 Eleni Damianidou
Eleni Damianidou Lidia Mouratidou
Lidia Mouratidou Christina Kyrousi
Christina Kyrousi- 1University Mental Health, Neurosciences and Precision Medicine Research Institute “Costas Stefanis”, Athens, Greece
- 2First Department of Psychiatry, Medical School, Eginition Hospital, National and Kapodistrian University of Athens, Athens, Greece
Neurodevelopmental disorders (NDDs) are a heterogeneous group of impairments that affect the development of the central nervous system leading to abnormal brain function. NDDs affect a great percentage of the population worldwide, imposing a high societal and economic burden and thus, interest in this field has widely grown in recent years. Nevertheless, the complexity of human brain development and function as well as the limitations regarding human tissue usage make their modeling challenging. Animal models play a central role in the investigation of the implicated molecular and cellular mechanisms, however many of them display key differences regarding human phenotype and in many cases, they partially or completely fail to recapitulate them. Although in vitro two-dimensional (2D) human-specific models have been highly used to address some of these limitations, they lack crucial features such as complexity and heterogeneity. In this review, we will discuss the advantages, limitations and future applications of in vivo and in vitro models that are used today to model NDDs. Additionally, we will describe the recent development of 3-dimensional brain (3D) organoids which offer a promising approach as human-specific in vitro models to decipher these complex disorders.
Introduction
The cerebral cortex is responsible for many of the higher-level cognitive functions in humans, such as perception, decision making, and language. Disruption of the tightly coordinated processes regulating brain development provokes the so-called neurodevelopmental disorders (NDDs), characterized by cognitive deficits, developmental delay and intellectual disabilities (ID). Amongst the most common NDDs are autism spectrum disorder (ASD) and attention deficit and hyperactivity disorder (ADHD) (Doernberg and Hollander, 2016). A group of diseases caused by defective cortical formation, namely malformations of cortical development (MCDs), belong also to the complex collection of NDDs (Desikan and Barkovich, 2016), which are usually present in patients with ASD and ADHD concurrently (Ho et al., 2019). Defining the pathophysiological mechanisms which underlie brain developmental disorders will help to facilitate treatment in NDDs, while it will contribute to the limited knowledge we have on human brain development. For this reason, in recent years, scientists have made a significant effort to model such disorders.
Until recently, animal models played a central role in the investigation of NDDs, however, they cannot fully recapitulate the human clinical and/or molecular and cellular phenotypes (Zhao and Bhattacharyya, 2018). Considering that there is limited access to fetal human brain tissue due to ethical and practical constraints, the use of alternative human-specific model systems that are accessible and ethically justified is crucial. In this review, we will introduce the basic steps of brain development and the evolutionary differences among mammals. The advantages and limitations of the in vivo and in vitro models for NDDs will be also analyzed. Finally, we will review the key categories of NDDs and MCDs and the models used to investigate the implicated mechanisms (Figures 1A,B).
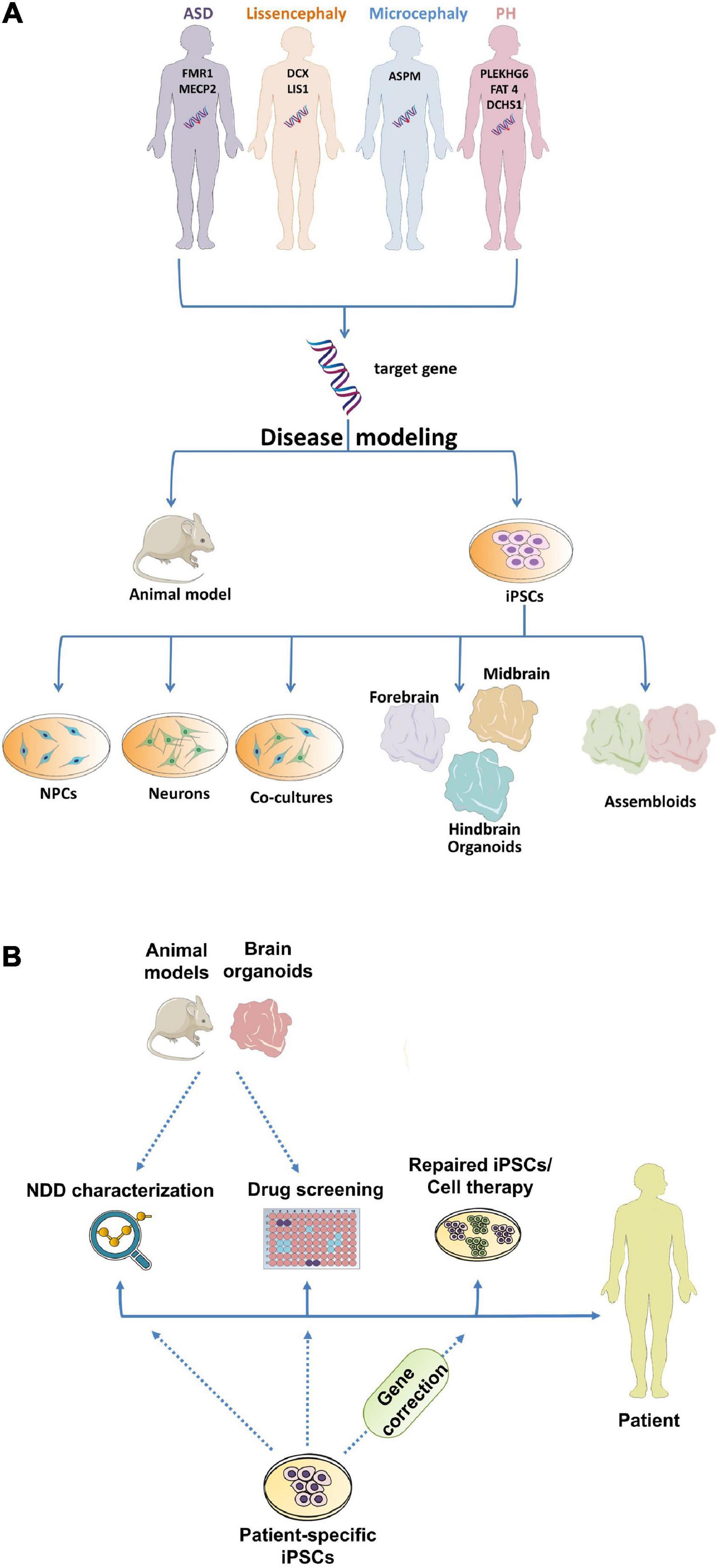
Figure 1. Utilization of different model systems and approaches in neurodevelopmental research. (A) Modeling of NDDs using in vivo and in vitro model systems. (B) Human-specific iPSC model systems are used for NDD characterization, drug screening and stem cell-based therapies, whereas animal models and human-specific COs can be used for NDD characterization and drug screening, aiming to improve patients’ quality of life. ASD, Autism Spectrum Disorder; PH, Periventricular Heterotopia; iPSCs, induced pluripotent stem cells; NPCs, neural progenitor cells; NDD, Neurodevelopmental disorder.
Evolutionary perspectives of mammalian neurogenesis
The development of the cerebral cortex begins early in the first trimester of gestation when neuroepithelial cells (NEs), the founder neural progenitor population located in the most rostral part of the neural tube, divide symmetrically to expand the neuroepithelial area (Subramanian et al., 2017). NEs, which form the ventricular zone (VZ) of the developing cortex, are polarized cells extending two thin processes: one contacting the apical side and the other contacting the basal side of the developing brain (pial) (Villalba et al., 2021). Shortly after the onset of corticogenesis, they transform into apical radial glia cells (aRGs). aRGs divide symmetrically, expanding the progenitor pool, and asymmetrically, giving rise to neurons directly or indirectly through generating other progenitor cells (Noctor et al., 2004). Amongst these progenitors are the intermediate progenitors (IPs) and the basal radial glia cells (bRGs) colonizing the subventricular zone (SVZ) and outer SVZ (oSVZ), respectively (Kalebic and Huttner, 2020). After their generation, newborn neurons assume a multipolar morphology and via the guidance of the RGs’ basal processes migrate away from the VZ and SVZ toward the developing cortical plate (CP) (Villalba et al., 2021). The continuous neuronal generation leads to the sequential formation of cortical layers (L1-L6) in an inside-out manner, where deep layer neurons form L5 and L6 and upper layer neurons form L2, L3, and L4 (Franchini, 2021).
Even though the main steps of neurogenesis are common among mammals (Franchini, 2021), the human cortex is greatly expanded, which is thought to be the foundation for the unique intellectual abilities of humans. A hallmark of humans’ but also of other primates’ and gyrencephalic cortices is the enlargement of supragranular layers, which originate from a unique progenitor pool, the oSVZ progenitors (Smart et al., 2002). In gyrencephalic species, bRGs are the most abundant neural progenitor (NPC) population in the SVZ (Hansen et al., 2010), and at the cellular level, they are highly heterogenous: there are bRGs with basal but no apical process, bRGs with only an apical process or bRGs with both basal and apical process (Betizeau et al., 2013). On the contrary, bRG-like cells in lissencephalic species like mice are few in number (5–10% of total NPCs between E12-E18) presenting only a basal process and located in the upper part of the SVZ (Wang et al., 2011). In gyrencephalic species, bRGs present a high proliferative capacity, whereas in mice they are mostly neurogenic (Smart et al., 2002; Betizeau et al., 2013). Thus, in gyrencephalic species and in particular, in humans, oSVZ is thicker with its progenitor cells playing a significant role in the formation of cortical folds (Borrell and Götz, 2014). Mechanical forces have highly contributed to the emergence of a folded cortex, with intercellular adhesion of migrating neurons being a crucial factor underpinning cortical folding (del Toro et al., 2017). Additionally, one of the main elements influencing cortical folding is the extracellular matrix (ECM) with human ECM components defining the mechanical properties of the developing cortex (Amin and Borrell, 2020). Transcriptional analysis in the mouse and primate brains has led to the identification of primate-specific and human-specific genes regulating the expansion of the neocortex (Boroviak et al., 2018). Characteristic examples of such human-specific genes are NOTCH2NL which contributes to the rapid evolution of the expanded human cortex (Fiddes et al., 2018) and ARHGAP11B which increases its size and folding (Florio et al., 2015). Primate-specific and bRG-specific genes were uncovered in fetal human brain samples, such as TMEM14B, DAG1, KCNK10, HP1BP3 (Liu et al., 2017). Also, the hominin-specific gene TBC1D3 was found to increase ERK signaling in humans’ bRGs and therefore, their proliferative capacity (Ju et al., 2016). A recent study by Pinson et al. revealed that a single lysine-to-arginine substitution in human TKTL1 leads to increased bRGs number and greater neurogenesis than in Neanderthals (Pinson et al., 2022). Nevertheless, the newly identified human-specific genes or human-specific isoforms cannot explain the phenotypic and morphological differences that are observed in human evolution as mentioned above, thus, attention has also been drawn to non-coding and regulatory sequences (Liu et al., 2017). Comparative genomic and epigenomic studies have identified: (a) regions in the human lineage named human accelerated regions (HARs), (b) non-coding region changes enriched in developmental enhancers (Bird et al., 2007) and (c) regions named human gained enhancers (HGEs) (Vermunt et al., 2016). These regions regulate gene expression of crucial key neurodevelopmental genes, some of which are associated with patterning in the frontal cortex, such as TBR1 (Won et al., 2019). Beyond these, other genes were proposed to be associated with the size of the frontal cortex such as FGF17 and EMX2 (Cholfin and Rubenstein, 2008). All the above, eventually lead to an increase in the neuronal number accompanied by a non-analog increase in the brain area size, which in turn leads to cortical gyrification. Interestingly, variants in human-specific genes, genes with human-specific expression pattern or HARs and HGEs have been found enriched in patients with NDDs, suggesting that human-specific changes are important not only for the evolution of the human brain but also for brain-related disorders.
Model systems: Advantages and disadvantages in studying brain development and disease
In vivo models
Animal models have been used for many years to dissect the mechanisms and disease-causing factors implicated in NDDs (Figures 1A,B), providing a huge advantage over human tissues whose use is limited. Among the animal models that have been highly used for studying brain development and brain-related diseases are zebrafish (Danio rerio) (Howe et al., 2013) and fruit fly (Drosophila melanogaster) (Gatto and Broadie, 2011). Their short life cycle and high genomic similarity with humans make them ideal to screen multiple candidate genes relatively fast. Drosophila contains almost 87% of genes involved in neurological and 75% of genes implicated in neurodevelopmental function. Similarly, zebrafish shares almost 70% homology with human genes with its neurodevelopmental processes being highly conserved through evolution (Howe et al., 2013). The chicken embryo (Gallus gallus) has also been a valuable tool in vertebrate neurodevelopment due to its large size, fast growth, easy accessibility for visualization and experimental manipulation and cost-effectiveness (Vergara and Canto-Soler, 2012; Zosen et al., 2021). Even though a huge breakthrough has been made in the research of brain-related disorders using these animal models, it is definite that they are distant from the human evolutionary perspective. Toward bridging this gap, scientists have turned to other model systems, such as rodents, to decipher the molecular and cellular mechanisms governing NDDs. The mouse model has been widely used because it presents a relatively fast reproduction time with several offspring. In addition, it presents more than 95% similarity with the human genome allowing accurate genomic manipulation of candidate genes. However, the use of mice also presents some limitations as it lacks several human-specific features, such as the gyrification of the cortex (Boroviak et al., 2018). Behavioral tests in rodents have been performed to model human behavioral deficits in individuals with NDDs (Kazdoba et al., 2016). Mice models exhibit several human NDD-related behavioral deficits including cognitive impairments, defects in learning and memory, hyperactivity and autism-like behaviors, but others cannot be precisely recapitulated, suggesting that assessment of higher brain functions in the lissencephalic brain is challenging. To overcome these limitations, non-human primates (NHPs) were proposed as potential models. The most used NHP is the monkey (Rhesus macaque), which presents a plethora of similarities with the human brain regarding cognitive function but also in terms of anatomical, structure, molecular and cellular features (Liu et al., 2020). Due to their long reproductive rate and demands in hosting they have only recently started to be used in NDDs research (Feng et al., 2020). Nevertheless, with the advantages of in vivo transient gene manipulation—i.e., in utero electroporation—and CRISPR/Cas9 technology precise nucleotides changes and transgene insertions/deletions—it has become economical and time effective to generate genetically manipulated NHPs (Chen et al., 2017; Zuo et al., 2017; Heide et al., 2020). These animal models serve as significant research tools to study genetic aspects of NDDs manifestation with great translational potentials.
Human-specific in vitro model systems
The striking differences between animal models and humans in terms of brain development raise questions about the value of information gained from animal studies. Considering that the use of human embryos or fetuses is ethically restricted, human-specific in vitro models could be a complementary approach to animal studies. A major breakthrough has been the differentiation of human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs) into all different cell lineages, possibly including neuronal lineage (Ardhanareeswaran et al., 2017). With the use of different culture media, hESCs and hiPSCs have been differentiated in two-dimensional (2D) neuronal cultures (Park et al., 2008), namely the monolayer cell cultures, including dopaminergic neurons (Sánchez-Danés et al., 2012), GABAergic neurons (Yang et al., 2017), serotonergic neurons (Vadodaria et al., 2016), hippocampal dentate granule neurons (Yu et al., 2014), motor neurons (Park et al., 2016), and hypothalamic neurons (Merkle et al., 2015). Besides neuronal cultures, astrocytes (Juopperi et al., 2012), oligodendrocytes, and microglia (Abud et al., 2017) have also been generated. These cultures have been used to study neuronal differentiation, neuronal activity, and neuronal morphology. The co-culture of neuronal and glial cells was also implemented to generate a more complex system and study cell-to-cell communication (Figure 1A). Considering that iPSCs enable the generation of scalable quantities of patient tissues, they facilitate the identification of therapeutic targets through drug screening (Elitt et al., 2018; Figure 1B). Moreover, aside from their contribution in NDD characterization and drug testing, iPSCs potential lies in their ability to serve as initial material for patient-specific stem cell-based cell therapies (Madrid et al., 2021; Figure 1B). Even though 2D human cultures transcended animal models concerning studying species-specific mechanisms, limitations arise from their use, especially regarding their inability to recapitulate cellular homeostasis and diversity. In particular, the lack of cellular diversity and cell interconnection among different brain regions, even in the co-culture system, may lead to the underestimation of the implicated cellular and molecular mechanisms.
Given these restrictions, the use of three-dimensional (3D) cell cultures, namely brain organoids which have been developed in the last decade, is pioneering in the study of brain development and disease. Brain organoids are stem cell-derived self-organized 3D tissues that mimic the cytoarchitecture, cellular composition and gene expression profile of the fetal human brain. The principle of their generation is based on cell sorting and spatially restricted lineage commitment (Benito-Kwiecinski and Lancaster, 2020). This model system is characterized by a high cellular diversity, where different NPCs, newborn and mature neurons as well as glial cells form a highly orchestrated and organized structure. Gene expression and electrophysiological studies have shown that brain organoids accurately resemble the first and second trimesters of the human fetal brain (Watanabe et al., 2017; Qian et al., 2018). In addition, single-cell RNA-sequencing data suggest that they model precisely cortical development on the molecular level (Camp et al., 2015; Amiri et al., 2018; Kanton et al., 2019; Velasco et al., 2019). Cerebral organoids (COs), the first brain organoid protocol (Lancaster et al., 2013; Lancaster and Knoblich, 2014), were generated from iPSCs based on their intrinsic properties to generate neuroectoderm. This protocol has been used since then to mimic early human brain development and evolution, as well as human brain diseases (Kyrousi and Cappello, 2020). Areas with specific regional characteristics like dorsal and ventral forebrain, choroid plexus, hippocampus and retina were found within these organoids (Lancaster et al., 2013). Interestingly, these regions can form connections with each other, making the modeling of interconnectivity possible. Newer protocols have attempted to direct regional identity in a more guided way by using patterning factors (Qian et al., 2019). Examples of single-region brain organoids include organoids of the cortex (Velasco et al., 2019), hypothalamus (Qian et al., 2016; Huang et al., 2021), cerebellum (Silva et al., 2021), and midbrain (Qian et al., 2016; Galet et al., 2020). In addition, protocols generating fused patterned organoids (or assembloids) have been developed to recapitulate the connections of different brain regions (Figure 1A). The fusion of dorsally and ventrally patterned organoids made possible to model the migration of inhibitory and excitatory neurons between and within these two areas, as well as the formation of a local inhibitory–excitatory neuronal circuit (Bagley et al., 2017; Birey et al., 2017). The generation of cortico-striatal assembloids (Miura et al., 2020) to model human cortico-striatal circuits of the forebrain has also been a big achievement, as dysfunctions in neural circuits of this pathway are thought to contribute to NDDs. The co-culture of medial ganglionic eminence organoids (MGEOs) and cortical organoids, generating human fused MGE-cortical organoids (Xiang et al., 2017) has been established to investigate complex interaction between specific brain regions. Following this approach, the fusion of human thalamic and COs to model axon projections (Xiang et al., 2019) was developed to study the molecular basis of human complex activities such as sensory-motor processing and attention. The different organoid protocols were shown to recapitulate closer the key aspects of human brain development than other model systems. Overall, the new methodologies in stem cell technology have expanded our toolbox for modeling NDDs. Choosing the appropriate model system depends on the focus of the investigation and the research question that needs to be answered (Figures 1A,B).
Using the correct system in modeling neurodevelopmental disorders
Neurodevelopmental disorders comprise a heterogeneous group of disorders characterized by a plethora of clinical phenotypes such as cognitive impairment, communication deficits, impaired psychomotor skills, and inability to reach developmental milestones (Parenti et al., 2020). NDDs include ASD, ADHD, ID, communication disorders, neurodevelopmental motor, learning and speech disorders. This group of disorders is characterized by high rates of comorbidity between several diseases within this diagnostic group. For example, individuals with ASD and ID exhibit often MCDs (Garcia-Forn et al., 2020), a wide spectrum of cortical abnormalities. MCDs are classified based on the perturbed developmental processes, thus, alterations in proliferation lead to microcephaly and macrocephaly, defects in neuronal migration result in periventricular/subcortical band heterotopia and Lissencephaly and deficits in the cortical organization lead to polymicrogyria (Barkovich et al., 2012; Severino et al., 2020). MCDs clinical manifestations include epilepsy, autistic features, ID and developmental delay, coupling MCDs with neuropsychiatric disorders. For example, individuals with Rett Syndrome present microcephaly and ASD at the same time, while in Seckel and Angelman syndrome patients exhibit microcephaly and ID (Sisodia et al., 2014). Patients with Prader-Willi (Bennett et al., 2015) and Timothy syndrome (Bauer et al., 2021) suffer from both ID and ASD, while in Fragile X syndrome characteristics of ASD, ID, and ADHD are present simultaneously. Interestingly, while monogenic causes predominate in MCDs (Table 1), NDDs usually exhibit polygenic pathophysiology (Li M. et al., 2018). Nevertheless, the etiology of these different syndromes is not well-understood up till now and thus, tremendous effort has been made to scrutinize their causality by using different model systems which will be presented in the following sections (Figures 1A,B). In addition, we will present MCDs based upon the observed neuropathological phenotype: (i) presence of ectopic neurons (periventricular heterotopia and subcortical band heterotopia), (ii) change of the gyrification index (Lissencephaly and polymicrogyria), (iii) abnormal brain size (microcephaly and macrocephaly) and how can we model them using the aforementioned model systems.
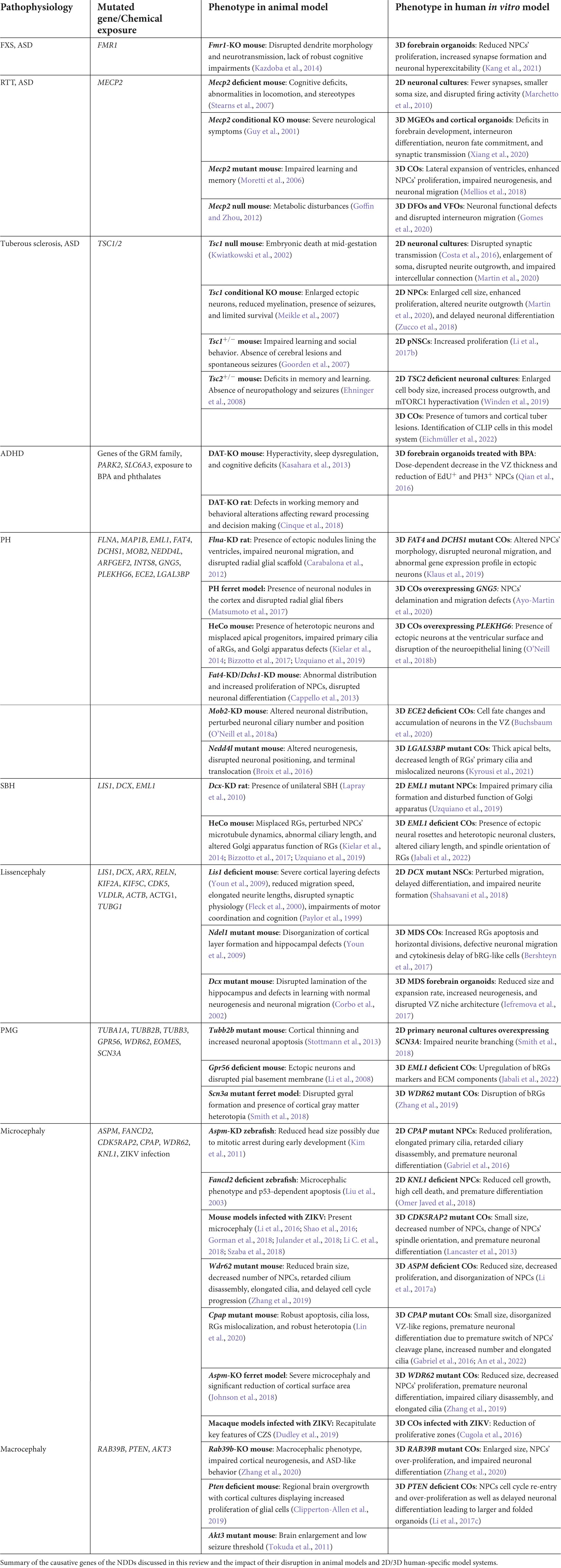
Table 1. Animal and human-specific model systems used in NDDs research.
Neurodevelopmental disorders
Autism spectrum disorders
Autism spectrum disorders involves deficits in social communication and repetitive behavior while it presents a complex mode of inheritance concerning genetic loci on several chromosomes (Muhle et al., 2004; Marco and Skuse, 2006; Rylaarsdam and Guemez-Gamboa, 2019). Candidate genes for ASD have key roles in neurodevelopment or/and neurotransmission (Rubenstein and Merzenich, 2003; McDougle et al., 2005). These genes encode regulatory proteins, such as transcription factors involved in neuronal synapses (Zoghbi, 2003) and networks relevant to neuroinflammation and neurotransmission (Voineagu et al., 2011). ASD remains one of the most heterogeneous NDD with more than 800 associated genes (Yin and Schaaf, 2017). Some of the most well-studied monogenic syndromes correlated with ASD in humans are: Fragile X syndrome (FXS) (mutation in FMR1) (Sitzmann et al., 2018), Rett syndrome (RTT) (mutation in MECP2) (Meloni et al., 2000), and tuberous sclerosis (mutations in TSC1 and TSC2) (Bjornsson et al., 1996), which will be analyzed in the next paragraphs.
The use of animal models has made and continues to make enormous progress in the understanding of the molecular mechanisms underlying FXS. The Fmr1 knockout (KO) mouse model exhibited synaptic deficits, abnormal dendritic spine morphology and neurotransmission defects, providing opportunities to evaluate novel drug targets (Kazdoba et al., 2014). Although, studies in FXS mouse models revealed deficient metabotropic pathway (Guo et al., 2016), its modulation led to unsuccessful clinical trials. The lack of effective clinical trials so far can be attributed to the fundamental developmental, biochemical, and physiological differences between animal models and humans. Indeed, transcriptome analysis of FXS forebrain organoids revealed a significant number of genes with altered expression, whereas only a few were differentially expressed in the FXS mouse brain (Kang et al., 2021). Moreover, this analysis showed a large number of overlapped genes differentially expressed in FXS forebrain organoids and fetal brain tissues (Kang et al., 2021). The differentially expressed genes in the FXS forebrain organoids were associated with neuronal migration, axonogenesis, neurogenesis, and neuronal differentiation (Kang et al., 2021). In addition, the rescue of the developmental deficits in FXS forebrain organoids was achieved by inhibiting the PI3K pathway, but not the metabotropic pathway which was disrupted in the FXS mouse models (Guo et al., 2016), suggesting that brain organoids mimic more accurately the FXS phenotype (Kang et al., 2021). Although dysregulation of PI3K pathway has been linked with FXS pathogenesis (Gross et al., 2015), modulation of this pathway has not been performed in clinical trials yet. Moreover, considering that pharmacological inhibition of PI3K pathway could rescue some of the developmental defects in FXS forebrain organoids (Kang et al., 2021), using PI3K pathway as a target in future therapeutic strategies might be promising. Overall, all the above indicate that human brain organoids could serve as preclinical models to identify human-specific therapeutic targets (Figure 1B).
To investigate RTT, transgenic mouse models and post-mortem human tissues have been widely used. Both Mecp2-mutant mice and human patients exhibit motor abnormalities (Stearns et al., 2007), robust neurological symptoms including uncoordinated gait and reduced spontaneous movement (Guy et al., 2001), learning and memory deficits (Moretti et al., 2006), metabolic disturbances (Goffin and Zhou, 2012), increased oxidative stress (Janc et al., 2016), seizures, and a shortened lifespan (Stearns et al., 2007). However, the neurological symptoms in mice are less severe and do not fully recapitulate the human phenotype. Two-dimensional neuronal cultures differentiated from patient-derived iPSCs harboring MeCP2 mutations showed impaired maturation, fewer synapses, excitatory/inhibitory (E/I) imbalance, smaller soma size and functional defects in firing activity (Marchetto et al., 2010). The recent innovations of brain organoids were used for RTT modeling, revealing impaired neurogenesis in human MGEOs and cortical organoids (Xiang et al., 2020) as well as deficits in neurogenesis and neuronal differentiation in COs (Mellios et al., 2018). Lastly, assembly of dorsal (DFOs) and ventral forebrain organoids (VFOs) from RTT patient-derived iPSCs revealed defects in interneuron migration (Gomes et al., 2020), suggesting a valuable model for understanding RTT during early stages of neural development.
Rodent studies have shown that loss of Tsc1/2 function in tuberous sclerosis impacts multiple processes at different developmental stages, including neuronal morphology and migration, synaptic plasticity and glial function (Tsai and Sahin, 2011). The germline loss of Tsc1 in mice resulted in embryonic lethality prior to brain development (Kwiatkowski et al., 2002), thus, spatial and temporal genetic manipulation at very early stages of cortical neurodevelopment is necessary. Loss of Tsc1 in cortical neurons during embryonic development in mice led to several neurological abnormalities, ectopic enlarged neurons and seizure activity (Meikle et al., 2007). Nevertheless, although the conditional KO mouse models have been and will continue to be a powerful research tool, this pathology may result from unique cellular and molecular aspects of human brain development. Moreover, although one mutant allele of TSC1/2 is sufficient to give rise to tuberous sclerosis in patients, heterozygous animal models demonstrate subtle or no symptoms (Goorden et al., 2007; Ehninger et al., 2008). Deficits in learning, memory and social behavior in Tsc1+/– and Tsc2+/– mice emerged in the absence of neuropathology and seizures (Goorden et al., 2007; Ehninger et al., 2008), indicating that animal models do not recapitulate faithfully the human phenotype. In contrast, the use of human stem cells enables to investigate the early stages of tuberous sclerosis neuropathology and identify the human-specific features of this disease (Figure 1B). It has been proposed that the haploinsufficiency of TSC1/2 could engage the pathomechanisms responsible for the formation of cortical tubers, the neuropathological hallmark of tuberous sclerosis (Winden et al., 2019). Neurons differentiated from hESCs in 2D displayed altered synaptic transmission, which was rescued by inhibition of mTORC1 pathway (Costa et al., 2016). Moreover, 2D NPCs differentiated from patient-specific iPSCs exhibited increased size and enhanced proliferation, consistent with the mTORC1 signaling activation (Martin et al., 2020) as well as disrupted neuronal differentiation (Zucco et al., 2018). Likewise, 2D patient-derived primitive neural stem cells (pNSCs) displayed increased proliferative capacity while differentiated neurons exhibited enlargement of the soma, perturbed neurite outgrowth and disrupted connections with other cells (Li et al., 2017b). Interestingly, although patient-specific iPSC-derived TSC2+/– neurons showed mTORC1 hyperactivation and associated increased cell body, only TSC2–/– neurons exhibited hyperactivity and transcriptional dysregulation observed in cortical tubers (Winden et al., 2019). A breakthrough in tuberous sclerosis research has been the development of patient-derived COs which recapitulated the emergence of brain tumors and dysplastic cortical regions (Eichmüller et al., 2022). More importantly, this model system revealed the presence of a novel interneuron progenitor population, namely CLIP cells, that give rise to both tumors and cortical tuber lesions (Eichmüller et al., 2022), highlighting the importance of using a proper model system in neurodevelopmental research.
Attention-deficit/hyperactivity disorder
Attention-deficit/hyperactivity disorder is a behavioral NDD characterized by ID, lack of attention, disorganization and difficulty in completing tasks (Ougrin et al., 2010). Although it is usually presented in children, symptoms might be maintained during adulthood. Findings from animal models suggest that ADHD is characterized by deficits in dopaminergic, noradrenergic and serotonergic systems, as well as more fundamental defects in neurotransmission (Russell, 2011). The etiology, although largely unknown, includes both genetic and environmental factors, while evidence shows that exposure to endocrine-disrupting chemicals, especially phthalates (Kim et al., 2009) and bisphenol A (BPA) (Rochester et al., 2018) might be causative. Genome-wide association studies have identified several candidate genes for ADHD including parkin 2 (PARK2) (Jarick et al., 2014), neuropeptide Y loci (Lesch et al., 2011) and genes in the metabotropic glutamate receptor family (GRM) (Elia et al., 2011). This polygenic etiology together with the variability in the clinical phenotype and the psychiatric root of ADHD renders its modeling extremely challenging.
Several animal models have been used to model ADHD including genetic, environmentally, and chemically induced models such as the spontaneously hypertensive rats (SHR) (Adriani et al., 2003), the Naples high-excitability rats (NHE) (Viggiano et al., 2002) and the neonatal 6-hydroxidopamine (6-OHDA) mice (Bouchatta et al., 2020). Since deregulation in the forebrain dopaminergic signaling and function has been associated with ADHD, dopamine transporter KO rodents (DAT-KO) have been generated by depleting SLC6A3 gene to study ADHD. Indeed, the DAT-KO mice presented hyperactivity, sleep dysregulation and cognitive deficits (Kasahara et al., 2013) while the DAT-KO rats showed a deficit in working memory and behavioral alterations affecting reward processing and decision making (Cinque et al., 2018). Furthermore, ADHD mouse models suggested that environmental factors in prenatal and perinatal stages, including epigenetic modifications, premature birth, and maternal smoking during pregnancy are major risk factors for ADHD (Halmøy et al., 2009; Zhu et al., 2014).
Nevertheless, the limited knowledge that we have of the developmental origin of ADHD and the implicated molecular and cellular mechanisms led to the development of rapid and low-cost human-specific in vitro models. The generation of ADHD patient-derived iPSCs has been reported (Sochacki et al., 2016; Jansch et al., 2018), offering the unique potential to bridge the gap between genetic and neural network associations in ADHD in a human-specific environment. Screening of drugs and chemicals was achieved by developing brain organoids providing a cheaper, faster, and more biologically relevant model (Schwartz et al., 2015) in understanding the environmental predisposition in ADHD (Figure 1B). For instance, the study of the effect of BPA, which is known to affect also rodent neurodevelopment (Kundakovic et al., 2013), demonstrated that the forebrain organoid system allowed quantitative investigation of the consequences of BPA exposure (Qian et al., 2016). In addition, with the help of robotic technology, an automated approach of large-scale human midbrain organoid cultures was generated for screening the general and dopaminergic-specific toxic effects of several compounds (Renner et al., 2020). Nonetheless, the lack of the blood-brain barrier, from which many chemicals cross, may have an important effect on understanding the underlying mechanisms considering toxicity. Moreover, the lack of any behavioral readout in 2D/3D model systems is not negligible and should be taken into consideration. It is thus, commonly accepted that a single model cannot capture all the traits of a complex brain disorder and that a combination of different approaches is needed to fulfill this request (Figures 1A,B).
Other neurodevelopmental disorders
While the focus has been drawn on ASD and ADHD, especially in recent years, other NDDs such as ID, motor disorders, learning and speech disabilities are highly uncharacterized. Our knowledge regarding these NDDs has mainly derived from comparative studies in human subjects, as research in animal models is limited and 2D/3D in vitro studies have not been conducted yet. Considering that molecular genetic testing coupled with genome-wide association studies (GWAS) have identified causative genes in individuals with these NDDs (Niemi et al., 2018; Gialluisi et al., 2021; Mountford et al., 2021) it is of high importance to delve into the etiopathogenic mechanisms using both in vivo and in vitro research tools (Figures 1A,B).
Malformations of cortical development characterized by the presence of ectopic neurons
Periventricular heterotopia
Individuals with periventricular heterotopia (PH) present a normal cortex with no major disruptions or changes. However, at the cellular level, patients exhibit clusters of gray matter organized either as laminar or as nodules close to the lateral ventricles. These areas of gray matter represent a subpopulation of neurons that fail to migrate toward the cortical plate (Watrin et al., 2015). Patients with PH suffer from various types of seizures and epilepsy, possibly due to the formation of an aberrant local neuronal network and synaptic connectivity. It has been proposed that the major etiopathogenic mechanisms of PH can be genetic mutations or environmental factors such as radiation, injury and infection (Watrin et al., 2015). While originally, PH was considered merely a neuronal migration disorder, recently focus has been drawn on the importance of NPCs’ proliferation and differentiation in the disease manifestation.
The X-linked FLNA which stabilizes the cytoskeleton was one of the first genes that have been associated with PH using animal models. The Flna-knockdown (KD) mouse model showed the importance of radial glia organization and the function of progenitor cells during neurogenesis and migration (Carabalona et al., 2012). Likewise, a PH ferret model exhibited neuronal nodules located in the cortex and disorganized radial glial fibers (Matsumoto et al., 2017). The significance of microtubule and actin dynamics in PH pathogenesis was also highlighted with the identification of MAP1B, encoding the microtubule-associated protein 1B, as a risk factor for PH (Heinzen et al., 2018). Interestingly, while pathogenic mutations in the microtubule-associated protein EML1, leads to subcortical band heterotopia in humans (Shaheen et al., 2017) (Section “Subcortical band heterotopia”), disruption of the Eml1 resulted in the heterotopic cortex (HeCo) in mouse mutants (Kielar et al., 2014). HeCo mice exhibited a population of aRGs located away from the VZ, which depicted perturbed primary cilia and disrupted Golgi apparatus function (Bizzotto et al., 2017; Uzquiano et al., 2019). In addition, Cappello et al., using also the mouse model demonstrated that Fat4-Dchs1 and Yap are key regulators in mammalian neurogenesis, suggesting that changes in the number of progenitor cells and their differentiation ability led to heterotopic neurons in the subcortex (Cappello et al., 2013). Notably, a Hippo signaling factor serving as a molecular modulator of Fat4 and Dchs1, namely Mob2, has also been implicated in PH pathogenesis (O’Neill et al., 2018a). Mob2 KD in the developing mouse cortex altered neuronal distribution and led to impaired ciliary numbers and position within migrating neurons (O’Neill et al., 2018a). Mechanistically, it has been also proposed that vesicle and/or membrane trafficking from the trans-Golgi network changes the transport of polarized molecules to the radial glial cells’ surface, thereby disrupting their proliferation and migration during cortical development (Sheen, 2012). For instance, mutations in the ARFGEF2 gene encoding the vesicle trafficking Brefeldin A-associated guanine exchange factor 2 (BIG2), are implicated in the manifestation of PH in humans (Sheen, 2014). Moreover, disruption of the E3 ubiquitin ligase NEDD4L leads to PH in humans, with mutants showing sensitivity to proteasome degradation in mouse models (Broix et al., 2016). Mutations in INTS8 have been also linked with severe NDDs characterized by PH, profound cognitive impairment, limb and facial dysmorphism and epilepsy, with patients exhibiting global transcriptome perturbations (Oegema et al., 2017).
Even though extensive research has been done in the past years using animal models, these systems failed to faithfully recapitulate the human phenotype. More recent work in 2D and 3D human-specific model systems highlighted the importance of the morphology and function of NPCs in the manifestation of PH (Klaus et al., 2019). Specifically, patient-derived DCHS1 and FAT4 deficient COs presented numerous neuronal nodules at ventricular positions, poorly organized germinal zones and morphologically disrupted NPCs. Additionally, KD of these genes in control COs led to an aberrant migration pattern in a subset of neurons (Klaus et al., 2019). Interestingly, single-cell RNA-sequencing analysis revealed a subpopulation of neurons with dysregulated genes involved in axon guidance, neuronal migration and patterning, elucidating the phenotype observed in patients, where only a subgroup of neurons fail to migrate, generating ectopic nodules (Klaus et al., 2019). Among the differentially expressed genes in the patient-specific ectopic cluster of neurons, GNG5, which is naturally expressed in NPCs, was the most upregulated (Klaus et al., 2019), indicating that its downregulation is essential for proper neuronal migration. Overexpression of GNG5 in control COs provoked premature NPCs’ delamination, induced alterations in NPCs’ morphology and led to neuronal migration defects (Ayo-Martin et al., 2020). Hence, it is now believed that the cross-talk between NPCs’ morphology and altered migration behavior of newly born neurons is a human-specific molecular and cellular explanation for PH manifestation (Klaus et al., 2019; Ayo-Martin et al., 2020). The importance of a human-specific model to study PH was also highlighted in a publication describing a novel de novo mutation in the human-specific isoform of the PLEKHG6, which is responsible for altering progenitors’ differentiation and neuronal migration downstream of RhoA (O’Neill et al., 2018b) highlighting again the necessity for the use of human-specific model systems. Besides, by using brain organoids, cell non-autonomous mechanisms have also been implicated in the etiology of PH by modeling the role of ECE2 (Buchsbaum et al., 2020), demonstrating the importance of the extracellular organization in the manifestation of PH. Likewise, mutations in LGALS3BP, encoding a secreted protein that interacts with the ECM, have been associated with PH, autism and seizures (O’Neill et al., 2018b). LGALS3BP deficiency led to increased apical belt thickness, prevention of NPCs’ delamination, ectopically located neurons and decreased RGs’ ciliary length in COs, indicating that its expression affects corticogenesis (Kyrousi et al., 2021). This study suggested that ECM, with LGALS3BP being a mediator of ECM signals, is crucial for human cortical development at the cellular level. Interestingly, patients with LGALS3BP mutations exhibit also changes in the local gyrification index, characteristics of other MCDs (Section “Malformations of cortical development characterized by altered gyrification index”), reinforcing the comorbidity between different MCDs.
Subcortical band heterotopia
Subcortical band heterotopia (SBH) is characterized by a smooth band of gray matter in the superficial and middle portions of the white matter. It emerges when newly generated neurons fail to migrate to the correct location and accumulate in the white matter. In humans, SBH is associated with ID, drug-resistant seizures and epilepsy (Sahu et al., 2019). The majority of patients are females carrying mutations in the X-linked DCX gene, exhibiting diffuse thick bands of ectopic neurons (Dobyns, 2010). Due to the stochastic nature of X-inactivation females with DCX mutations present a mosaic state with two populations of neurons: one that expresses DCX and migrates properly and the deficient one that gives rise to the heterotopic band. In males with DCX deficiency neurons lack completely the protein, giving rise to the more severe lissencephalic “smooth” cortex (Section “Lissencephaly”) (Matsumoto et al., 2001). Another locus that has been linked with SBH contains the PAFAH1B1 gene, which encodes the LIS1 protein. LIS1-associated SBH exhibits thin or intermediate posterior bands, with posterior heterotopia being more severe than anterior heterotopia (Mineyko et al., 2010). Additionally, individuals with mutations in EML1, exhibit SBH, megalencephaly, polymicrogyria (section “Polymicrogyria”), and agenesis of corpus callosum (Shaheen et al., 2017; Oegema et al., 2019). Overall, MCDs result from single point mutations in critical developmental genes or from variations in several genetic loci, increasing disease probability. Interestingly, disruption of common genes might lead to divergent disease phenotypes (Table 1), with the variable penetrance of genetic variants complexifying, even more, the overall clinical manifestation (Klingler et al., 2021).
Although KD of Dcx in rats has provided valuable insight into the SBH epileptogenicity, they only harbor a unilateral SBH, not mimicking accurately the human phenotype (Lapray et al., 2010). Disruption of Eml1 leads to HeCo mutant mice, a unique model that has been widely used to decipher the molecular basis of SBH. HeCo mice exhibit progenitor anomalies with some RGs leaving the VZ and diving in basal locations (Kielar et al., 2014). HeCo mice RGs presented disrupted primary cilia and perturbed Golgi apparatus mechanisms that probably contributed to the abnormal RGs’ delamination (Bizzotto et al., 2017; Uzquiano et al., 2019). The contribution of primary cilia in SBH pathophysiology is not surprising considering their role in the regulation of RG behavior. Nonetheless, although HeCo mouse models exhibit heterotopia they do not present the whole spectrum of pathologies found in humans, such as polymicrogyria and megalencephaly, indicating differential disease manifestation in humans and non-human models.
The emergence of cutting-edge stem cell technology has been extremely valuable in deciphering the etiopathogenesis of SBH. Two-dimensional cortical progenitors differentiated from EML1 mutant iPSCs exhibited perturbed primary cilia formation and disrupted function of Golgi apparatus (Uzquiano et al., 2019). Moreover, EML1-deficient forebrain organoids presented ectopic neural rosettes accumulated at the basal side of VZ and heterotopic neuronal clusters (Jabali et al., 2022). RGs in the VZ of mutant organoids exhibited altered spindle orientation and shorter primary cilia which might directly impact RG delamination. Additionally, single-cell RNA-sequencing in the ectopic progenitor populations showed upregulation of bRGs markers and human-specific ECM components (Jabali et al., 2022). Although this model system revealed the implicated pathomechanisms in EML1 deficiency, the developmental stage of organoids does not allow us to study the full scope of MCDs caused by EML1 impairment.
Malformations of cortical development characterized by altered gyrification index
Lissencephaly
Lissencephaly is characterized by defective neuronal migration and results in the lack of development of sulci and gyri which leads to a smooth brain surface. It encompasses a broad group of brain malformations with abnormalities in the formation of gyri ranging from agyria (absence of gyri) to oligogyria (reduced gyrification) (Severino et al., 2020). Besides those, pachygyria (broad gyri) and SBH (double cortex) are also included in the spectrum of Lissencephaly (Kattuoa and Das, 2022). Depending on the degree of malformation, patients might present developmental delay and mental disabilities. Among the key mechanisms leading to Lissencephaly is the disruption of the proper development of gyri and sulci. It is caused due to genetic and non-genetic factors such as lack of oxygenated blood to the brain during fetal development or viral infections of the mother of the fetus (Leruez-Ville and Ville, 2017). Interestingly, many associated genes are causative for the manifestation of SBH as well (section “Subcortical band heterotopia”), supporting the notion that impairments in common genes result in divergent clinical pictures. Miller Dicker Syndrome (MDS) is a cortical malformation characterized by Lissencephaly often associated with microcephaly (section “Microcephaly”) leading to ID and mortality (Herman and Siegel, 2008). This syndrome is caused by deletions in the human band 17p13.3, which includes PAFAH1B1 (LIS1 protein) (Herman and Siegel, 2008). Genetic causes of Lissencephaly also involve the DCX gene which is essential for proper neuronal migration (Corbo et al., 2002). Other genetic causes of Lissencephaly include mutations in ARX playing an important role in forebrain development (Gécz et al., 2006), RELN which encodes the protein reelin (Hong et al., 2000) and other genes like KIF2A, KIF5C, CDK5, VLDLR, ACTB, ACTG1, and TUBG1 (Parrini et al., 2016) which, however, have not been properly modeled so far.
The function of Lis1 and the pathways associated with it have been studied in mouse models for Lissencephaly (Youn et al., 2009). Mice with a graded reduction in Lis1 dosage in disorganized cortical layers, cerebellum, hippocampus and olfactory bulb have been suitable models to study MDS. These models have aided in demonstrating impairments of motor coordination and cognition, severe disruptions of hippocampal cellular and synaptic physiology as well as in vivo migration defects (Paylor et al., 1999; Fleck et al., 2000). Additionally, NDEL1 which interacts with the LIS1/dynein complex has also been implicated in Lissencephaly pathogenesis. Specifically, Ndel1 null mouse brains exhibited severe cortical layering defects and hippocampal deficits (Youn et al., 2009). Another genetic cause associated with classic Lissencephaly, and SBH is mutations in the DCX gene. Dcx mutant mice displayed disrupted lamination in the hippocampus, leading to deficits in learning (Corbo et al., 2002). However, the small and unfolded smooth brain of mice makes it difficult to model the wide spectrum of phenotypes in a precise way. For example, despite the disruption of the hippocampus, Dcx mutant mice presented a normal cortex with regular patterns of neocortical neurogenesis and neuronal migration (Corbo et al., 2002). In contrast, small interfering RNA (siRNA) inhibition of DCX in rat embryonic cortical slices led to severe defects in neuronal migration (Bai et al., 2003). These opposing results suggest that there is a more complex role for DCX, highlighting the necessity of human-specific models.
Differentiation of iPSCs into 2D neural cells provides a model system to further scrutinize the molecular and cellular regulation of Lissencephaly and in particular, the function of DCX in neurodevelopment. Neural stem cells (NCSs) with absent or reduced DCX protein expression showed delayed differentiation, impaired migration and neurite formation, recapitulating the lissencephalic phenotype (Shahsavani et al., 2018). As previously mentioned, in the developing human cortex the oSVZ, in which IP cells and bRGs are localized, is thicker compared to other mammals (Hansen et al., 2010). While IPs are conserved between human and mouse, bRGs are fewer in the developing cortex of lissencephalic mice. Moreover, human bRGs have been associated with the evolutionary increase in the cortical size of the human brain (Nonaka-Kinoshita et al., 2013). All the above may explain why Lis1 deficient mice presented a milder phenotype compared to human patients with heterozygous PAFAH1B1 mutations. The implementation of brain organoid cultures has demonstrated the effect of MDS Lissencephaly mutations on distinct biological processes and cell types. Specifically, there has been a recapitulation of developmental lineage from NEs to RGs, IPs and bRGs with spatial and temporal resolution, which could not have been achieved with the previous methods (Kadoshima et al., 2013). This has highlighted the vulnerability of NEs and human-specific bRGs in MDS, whereas aRGs and IPs were less affected (Bershteyn et al., 2017). MDS-forebrain organoids exhibited small size, reduced expansion rate, changes in RGs’ division plane, increased neurogenesis and altered cortical niche architecture (Iefremova et al., 2017). Finally, the involvement of the ECM in cortical gyrification and thereby, in the manifestation of MCDs characterized by altered gyrification index is not insignificant considering that patients harboring LGALS3BP mutations, exhibit altered gyrification, while its overexpression in the mouse brain promoted cortical folding, indicating that LGALS3BP regulates cortical expansion and gyrification (Kyrousi et al., 2021). Even though bRGs seem to have a robust effect on gyrification and hence, the emergence of MCDs with altered gyrification index, due to the almost total absence of bRGs in mice, it is challenging to study bRG behavior in this model. In contrast, in brain organoids, there are bRG-like cells with matching characteristics in vivo such as morphology, molecular identity and mitotic behavior. Nevertheless, how the in vitro-derived bRG cells recapitulate the functional properties and molecular identity of primary bRG cells has not yet been determined, making the synergy of in vivo and in vitro models essential (Figures 1A,B).
Polymicrogyria
Polymicrogyria (PMG) is a MCD characterized by the presence of multiple abnormally small gyri which leads to an irregularly folded cortical surface with an increased gyrification index. Individuals with PMG present cognitive impairments, epilepsy and developmental delay. It has been associated with the malfunction of several genes including TUBA1A (Cushion et al., 2013), TUBB2B (Guerrini et al., 2012), TUBB3 (Poirier et al., 2010), GPR56 (Piao et al., 2004), WDR62 (Yu et al., 2010), EOMES (Baala et al., 2007), SCN3A (Smith et al., 2018), as well as environmental causes. Considering that bRGs are responsible for cortical folding in gyrencephalic species, PMG might be an outcome of increased migration and/or excess of bRGs (Klingler et al., 2021). It is a highly heterogeneous disorder in terms of topographic distribution, clinical, and imaging features, as well as severity levels, varying from focal forms to bilateral involvement (Leventer et al., 2010). PMG often occurs concurrently with other MCDs, such as Lissencephaly, macrocephaly and heterotopia (Stutterd et al., 2020), but it is probably the least characterized among the more common MCDs (Stutterd and Leventer, 2014), thus more effort has to be made to identify the implicated mechanisms.
Only a few animal models harboring mutations in PMG-associated genes, that are mentioned here, have been reported (Table 1). Nonetheless, they fail to recapitulate accurately the human phenotype. For instance, Tubb2-deficient mouse model exhibited cortical thinning and increased apoptosis of cortical neurons which led to perinatal lethality (Stottmann et al., 2013). Likewise, while individuals carrying EML1 mutations exhibit a wide spectrum of impairments including a PMG-like cortex, HeCo mouse models failed to mimic this phenotype (Kielar et al., 2014) (section “Subcortical band heterotopia”). Additionally, although loss of GPR56 in humans leads to bilateral frontoparietal PMG (Piao et al., 2004), in mice it led to ectopic cortical neurons due to neuronal over-migration, structural aberrations in the RG endfeet and breaches in the pial basement membrane (Li et al., 2008). Overall, Gpr56 mutant mice exhibited disorganized cortical lamination and a cobblestone-like malformation (Li et al., 2008). Although mouse models have been a valuable tool in MCDs research, using complex gyrencephalic cortices is essential to decipher the PMG-implicated pathomechanisms. Interestingly, the PMG-associated GPR56 e1m promoter preferentially drove gene expression in GABAergic neurons of the developing cortex in common marmosets, indicating a possible link with GPR56-mutation associated epilepsy (Murayama et al., 2020). Moreover, altered expression of SCN3A, which encodes the brain-enriched voltage-gated sodium channel NaV1.3, disrupted cortical gyral formation and led to cortical gray matter heterotopia in a ferret animal model, highlighting the necessity of ion channels function at early stages of cortical development (Smith et al., 2018).
Even though initial steps have been taken to model PMG, animal models cannot mimic properly the human clinical phenotype, therefore, the need for human-specific model systems is urgent. Additionally, the lack of human-specific models for several PMG-linked genes, such as GPR56, impedes the disease characterization and modeling, hence steps need to be taken toward this direction. Overexpression of SCN3A mutant variants in 2D primary human neuronal cultures attenuated neurite branching, supporting the notion that SCN3A regulates neuronal development (Smith et al., 2018). Transcriptome analysis in EML1-deficient COs revealed an upregulation of bRGs markers and human-specific ECM components in the ectopic neural rosette population, supporting the involvement of bRGs in PMG pathophysiology (Jabali et al., 2022). Considering that the upregulated ECM genes COL1A2, COL3A1, and LUM have been linked with cortical expansion and folding (Long et al., 2018), it is tempting to speculate that the increase in their activity plays a role in the development of PMG-like phenotype in patients harboring EML1 mutations (Oegema et al., 2019). Moreover, modeling the WDR62-associated microcephaly gene (section “Microcephaly”) using COs depicted disruption of bRGs (Zhang et al., 2019), indicating that this model system could be a valuable tool to delve into PMG implicated pathways. Nonetheless, the absence of folding in brain organoids constrains the modeling of PMG-associated phenotypes. Deletion of PTEN in human COs led to the expansion of the NPC population and substantial surface folding (Li et al., 2017c), indicating that we are one step closer to engineering folded brain organoids that can be used for PMG modeling. While actions have been taken toward the generation of folded organoids, the existing protocols are preliminary. In contrast to cortical gyrification which takes place around gestational week 30, when the fetal brain is largely composed of neurons, folded organoids contain mainly NPCs, thus they mimic an earlier developmental stage (Karzbrun et al., 2018). Overall, while the organoid wrinkling is equivalent to cortical gyrification there are crucial biological differences, indicating that this field is still developing.
Malformations of cortical development characterized by abnormal brain size
Microcephaly
Microcephaly is characterized by reduced brain growth and is defined as a head circumference with ≤−3 SD. It results from an imbalance between progenitors’ production and cell death and leads to a reduced number of neuronal and glial cells (Ashwal et al., 2009; Álamo-Junquera et al., 2015). It is one of the most studied MCDs, with extensive research showing that some of the causes of primary microcephaly (present at birth) are impairments in DNA repair pathways (Zhou et al., 2013), mutations in genes encoding centrosomal proteins (Guernsey et al., 2010), double-stranded breaks (Yue et al., 2013), toxic exposures (Parens et al., 2017), in utero infection (Antoniou et al., 2020) and metabolic conditions (Ashwal et al., 2009; Table 1). Secondary microcephaly (postnatal manifestation) can be associated with migration defects (Juric-Sekhar et al., 2011), increased cell death (Simmons et al., 2019), and metabolic disorders (Ashwal et al., 2009). Besides, microcephaly can also be a clinical feature in other NDDs and syndromes like RTT (Banerjee et al., 2012), Angelman syndrome (Sun et al., 2019) and Seckel syndrome (Al-Dosari et al., 2010), while, maternal infections from Zika virus (ZIKV) lead to congenital zika syndrome (CZS) with infants exhibiting severe microcephaly (Moore et al., 2017; Antoniou et al., 2020).
Zebrafish has been a useful animal model to decipher the genetic implications and molecular mechanisms of microcephaly. Knocking down of the abnormal spindle-like microcephaly associated gene (Aspm), which is associated with the manifestation of autosomal recessive primary microcephaly (MCPH), resulted in reduced head size in zebrafish (Kim et al., 2011). Zebrafish Fnacd2 mutants also presented microcephaly due to increased p53-dependent apoptosis (Liu et al., 2003). Moreover, immunocompromised (AG129, C57BL/6 models) (Shao et al., 2016; Julander et al., 2018; Li C. et al., 2018) and immunocompetent [Swiss Jim Lambert (SJL), ICR, STAT2 KO models] (Li et al., 2016; Gorman et al., 2018; Szaba et al., 2018) mice have been a useful tool to investigate the effects of ZIKV infection. In addition, mouse models have been widely used to investigate the effects of disruption of MCPH-associated genes in the developing brain (McIntyre et al., 2012; Insolera et al., 2014; Zhang et al., 2019). Loss of CPAP, a protein crucial for centriole biogenesis, induced p53-dependent cell death which severely disrupted embryonic mouse brains (Lin et al., 2020). However, patient mutations in mice led to mild microcephalic phenotypes and did not give insights into cellular and molecular mechanisms. Additionally, there is no clear evidence that a disease-causing mutation in the mouse brain is analogous to human microcephaly. Interestingly, Aspm KO in the ferret, a species with a larger gyrencephalic cortex and greater NPC diversity than mice, led to severe microcephaly with significantly reduced cortical surface area (Johnson et al., 2018). To recapitulate more accurately the pathogenesis, neurophysiology and therapeutic development of CZS, NHPs have also been used as they present similarities to humans in gestational biology (Nguyen et al., 2017). Specifically, macaques recapitulated key features of human microcephaly via ZIKV infection (Dudley et al., 2019). Nevertheless, the demands of NHS breeding and management as well as the generation of transgenic animals are challenging, highlighting the need for model systems that are not only physiologically relevant to the human brain but also easy to manipulate.
Human-specific 2D and 3D in vitro models have contributed to decoding the complexity of microcephaly, which was challenging to be deciphered in the existing in vivo models. Differentiation of Seckel patient-derived iPSCs in 2D, revealed reduced proliferation and elongated cilia in CPAP mutant NPCs (Gabriel et al., 2016). Interestingly, Seckel NPCs exhibited retarded cilium disassembly and delayed G1-S transition, which led to NPC premature differentiation (Gabriel et al., 2016). In the same line, 2D differentiation of hESCs into NPCs harboring mutations in the microcephaly associated gene KNL1, which is essential for spindle assembly checkpoint signaling, revealed reduced cell growth, high cell death, and premature differentiation at the expense of NPC proliferation (Omer Javed et al., 2018). CDK5RAP2, which encodes the CDK5 regulatory subunit-associated protein 2, was the first gene used to study microcephaly in brain organoids. The generation of patient-derived COs has shown that changes in the mitotic spindle orientation affect progenitors’ proliferation and lead to premature neuronal differentiation (Lancaster et al., 2013). Moreover, modeling ASPM mutations demonstrated the ability of human COs to recapitulate microcephaly (Li et al., 2017a). In turn, CPAP mutant COs displayed reduced size—a key feature of microcephaly—and increased number of cilia which affected progenitors’ mode of division and led to premature neurogenesis (Gabriel et al., 2016). Likewise, a CPAP mutation identified in MCPH patients induced cell death in COs and led to the generation of smaller organoids (An et al., 2022). Additionally, the CPAP mutation provoked NPC spindle misorientation which resulted in premature differentiation (An et al., 2022). Finally, WDR62 deletion, which encodes a centrosomal protein, in COs revealed impaired ciliary length, decreased NPC proliferation and premature neuronal differentiation which eventually led to a reduction in the size of mutant organoids (Zhang et al., 2019). The perturbed ciliary structure and function in the 2D/3D models of microcephaly highlight the necessity of primary cilia in regulating crucial developmental processes including NPCs’ proliferation and differentiation. Last but not least, brain organoids have successfully recapitulated the effects of ZIKV infection in cortical development, revealing a RG-specific infection, especially in the VZ, resulting in a reduction of the proliferative zones (Cugola et al., 2016).
Macrocephaly
Macrocephaly is defined as head circumference with ≥2 SD for a given age and gender (Accogli et al., 2022), originating from a higher proliferation rate of progenitors as well as defects in growth factor signaling (Sun and Hevner, 2014). Heterozygous loss-of-function mutations in PTEN were identified in patients with macrocephaly and associated with ASD, concurrently (Butler et al., 2005). Individuals with PTEN mutations exhibit brain enlargement and white matter abnormalities (Vanderver et al., 2014). Nonetheless, PTEN mutations do not have the same effects throughout different brain areas. Abnormal scaling of different brain regions could indicate disrupted connectivity, the presence of ectopic tissue, or abnormal innervation of crucial brain areas, which may contribute to behavioral phenotypes. Interestingly, loss of PTEN leads to an increase in the PI3K-AKT-mTOR signaling, which in turn has been suggested to have important roles in macrocephaly (Yeung et al., 2017). For instance, mutations in the small GTPase RAB39B which interacts with the PI3K components are associated with X-linked macrocephaly, ASD and ID (Woodbury-Smith et al., 2017). Moreover, AKT3 activating mutations have been linked with macrocephaly and polymicrogyria (Lee et al., 2012).
Prenatal neurodevelopmental abnormalities in macrocephaly are difficult to be understood because of the limited access to human brain tissues. Thus, genetically modified animal models remain a necessary strategy to scrutinize the pathogenesis of macrocephaly on a molecular and behavioral basis. Rab39b KO mice exhibited macrocephaly and autism-like behaviors (Zhang et al., 2020). In particular, they presented an increased proliferative rate and decreased cell cycle exit of NPCs which led to the expansion of the progenitors’ pool (Zhang et al., 2020). Pten+/– mouse models presented regional brain overgrowth, which is similar to brain enlargement in adult PTEN-ASD patients (Clipperton-Allen et al., 2019). This phenotype may be associated with increased gliogenesis due to reduced suppression of the PI3K pathway (Clipperton-Allen et al., 2019). In addition, Akt3 mutant mice harboring a dominant missense mutation showed enhanced signaling activity and increased brain size (Tokuda et al., 2011).
In humans, the majority of upper layer neurons derive from the oSVZ, where bRGs are abundant. On the contrary, in rodents, bRG-like cells are fewer in number. Thus, human iPSCs and iPSC-derived brain organoids are a valuable platform for studying macrocephaly, as the human brain exhibit increased cortical size and complexity. Indeed, RAB39b mutant COs exhibited a more severe phenotype (Zhang et al., 2020) compared to KO mouse models, highlighting the valuable role of brain organoids in modeling human early development. These mutant brain organoids had a more prominent increase in the neural progenitor pool and upper layer neurons compared to mutant mice, explaining the severe phenotype that appeared in mutant organoids (Zhang et al., 2020). Likewise, the deletion of PTEN resulted in enlarged and folded brain organoids. PTEN homozygous mutations in organoids increased PI3K pathway signaling, promoted cell cycle re-entry of NPCs and transiently delayed neuronal differentiation, resulting in expansion of the progenitor population (Li et al., 2017c). However, the fact that the developmental timing between organoids and mutant mice is not compatible, shows the importance of making side-by-side comparisons between mouse models and human brain organoids.
Brain organoids as tool to model neurodegenerative and neuropsychiatric disorders
Besides the applications of brain organoids in NDDs research, they can be also used to elucidate impaired processes in neurodegenerative and neuropsychiatric diseases (Sun et al., 2021). The poor physiological relevance of animal models’ brain, the limited number of therapies as well as the inefficient treatment effects in neurodegenerative disorders dictate the need for revolutionary tools. Moreover, the requirement for a non-invasive analysis of patient-derived tissue and an effective drug-screening system has contributed to the emergence of brain organoid cultures. Even though brain organoids do not recapitulate the aging brain, they have been used to model several neurodegenerative disorders. Since these disorders are associated with extended periods of prodromal disease, where cellular abnormalities are accumulating without manifesting as clinical symptoms, organoids allow us to investigate these early cellular changes that drive neurodegeneration (Wray, 2021). Moreover, advances in organoid technology have enabled the protracted culture of brain organoids, leading to increased cellular diversity and neuronal maturation (Quadrato et al., 2017). These matured organoids developed spontaneously active neuronal networks and acquired traits of mature neurons, suggesting that, beyond modeling early neurodevelopmental processes, they have the potential to model higher-order functions of the human brain (Quadrato et al., 2017). Organoids have been used to model Alzheimer’s disease (AD) (Gonzalez et al., 2018; Ghatak et al., 2019), Parkinson’s disease (PD) (Kim et al., 2019; Smits et al., 2019), amyotrophic lateral sclerosis (ALS) (Osaki et al., 2018), frontotemporal dementia (Seo et al., 2017), Huntington’s disease (Conforti et al., 2018), and spinal muscular atrophy (Hor et al., 2018). Considering that 2D model systems lack the complex extracellular environment necessary to recapitulate protein aggregation, such as the extracellular deposition of misfolded amyloid-β (Aβ) plaques, the molecular hallmark of AD, 3D culture conditions provide a powerful tool to investigate Aβ pathology (Lee et al., 2016; Bubnys and Tsai, 2022). Additionally, the emergence of human midbrain organoids has been pioneering in PD research, which is characterized by the loss of dopaminergic neurons in the substantia nigra pars compacta and the presence of Lewy body inclusions (Galet et al., 2020). Likewise, the complexity of the human brain concerning physiological, functional and structural aspects of brain development as well as the increase in cognitive abilities have pointed out the need for human-specific model systems to study neuropsychiatric diseases. Toward this direction, organoids have provided valuable information regarding implicated pathomechanisms in schizophrenia (Kathuria et al., 2020), bipolar disorder (Mertens et al., 2015), psychosis (Sawada et al., 2020), and depression (Vadodaria et al., 2019). In particular, modeling the effect of schizophrenia-linked genes using brain organoids revealed disrupted neural rosette structures and altered proliferation of NPCs which was rescued by WNT antagonism, indicating that organoids can be a valuable tool for drug testing (Figure 1B) (Srikanth et al., 2018). Taking into account, that patients with NDDs can exhibit simultaneously neurodegenerative or neuropsychiatric disorders, the use of a model system that can recapitulate them faithfully is of great potential.
Conclusion and future perspectives
Creating a functional cerebral cortex requires the coordination of crucial developmental processes such as progenitors’ proliferation and differentiation, neuronal migration and eventually their maturation and integration into the existing neuronal network. Disruptions of these complex and delicate processes lead to a plethora of brain impairments, the NDDs. Their clinical manifestation encompasses developmental delay, epilepsy, ID but also various structural abnormalities that affect the size, thickness and folding of the cortex. Understanding the molecular etiology of NDDs is crucial for patient management. Nonetheless, their multifactorial and polygenic nature makes their modeling difficult. Although rodent and other animal models have been the foundation for deciphering the implicated pathways in NDDs, the use of human-specific models is essential. Toward this direction, iPSC-derived 2D and 3D in vitro cultures show great potential for modeling human neurodevelopmental pathologies. In this review, we elaborated on both the in vivo and in vitro models used to study NDDs (Figures 1A,B).
Although the tremendous progress in stem cell and organoid development has decoded complex pathogenic pathways, brain organoids present numerous limitations as a model system (Paşca, 2018). In particular, they do not develop past the stage of a prenatal brain and cannot recapitulate the six-layered spatial organization of the cortical plate (Lancaster et al., 2013). Nevertheless, recent advances in the patterning of brain organoids have led to prolonged in vitro development, enabling significant levels of maturity (Quadrato et al., 2017; Velasco et al., 2019). Additionally, despite the existence of bRGs, COs have not presented folding on the pial surface and gyrification (Qian et al., 2019). Moreover, the E/I neurons although functional, are less mature compared to adult neurons. Thus, cell diversity in organoids remains to be further enriched, also given that they lack non-neuronal cell types such as microglia (Dutta et al., 2017) which could be achieved with new pattering protocols (Ormel et al., 2018) or with co-cultures with non-neuronal lineage cell types in the future. The lack of vascularization, which restricts the delivery of nutrients and oxygen has been another significant obstacle to the long survival of these cultures, as a necrotic core is built up inside the organoids (Qian et al., 2019). Toward addressing the lack of vascularization, the grafting of human brain organoids into adult mouse brains has been performed (Mansour et al., 2018). Interestingly, functional intra-graft neuronal networks, as well as graft-to-host synaptic connectivity were observed in these mice. Another promising tool for overcoming such problems is organoids-on-a-chip. A recent report on the human kidney has examined the fluid flow on vascularization and maturation of hiPSC-derived kidney organoids. They engineered them in a 3D printed chamber which led to the expansion of endothelial progenitors within the organoids and eventually the formation of blood vessels in a flow-enhanced way (Homan et al., 2019). Another limitation is the variability of the different organoid cultures, which depends on the quality of the founder cell line (i.e., the line and passage of the iPSCs) (Hernández et al., 2022). Lastly, one major obstacle to the use of brain organoids to model NDDs is the lack of any behavioral readout of the in vitro cultures. Summarizing, focusing on six-layer human architecture, in vitro vascularization, patterning and further refinement in tissue identity can offer great promise, toward the development of a more robust human-specific model system (Chiaradia and Lancaster, 2020). Nonetheless, the advanced development and engraftment of organoids in animal brains constitute an ethical gray zone, making ethicists wonder if these experiments are essential to answer a scientific question or whether boundaries are pushed beyond limits (Reardon, 2020; Powell, 2022).
Author contributions
ED and LM wrote the manuscript. CK conceived, wrote, and revised the presented work. All authors read and approved the final manuscript.
Funding
This work was supported by the Hellenic Foundation for Research & Innovation (2881) and the International Brain Research Organization (IBRO).
Acknowledgments
We thank Panagiota Nti Kostantzo for the critical reading of the manuscript. Our group apologizes that, because of space limitations, some primary publications could not be cited.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abud, E. M., Ramirez, R. N., Martinez, E. S., Healy, L. M., Nguyen, C. H. H., Newman, S. A., et al. (2017). IPSC-derived human microglia-like cells to study neurological diseases. Neuron 94, 278–293.e9. doi: 10.1016/j.neuron.2017.03.042
Accogli, A., Geraldo, A. F., Piccolo, G., Riva, A., Scala, M., Balagura, G., et al. (2022). Diagnostic Approach to Macrocephaly in Children. Front. Pediatr. 9:794069. doi: 10.3389/fped.2021.794069
Adriani, W., Caprioli, A., Granstrem, O., Carli, M., and Laviola, G. (2003). The spontaneously hypertensive-rat as an animal model of ADHD: Evidence for impulsive and non-impulsive subpopulations. Neurosci. Biobehav. Rev. 27, 639–651. doi: 10.1016/j.neubiorev.2003.08.007
Álamo-Junquera, D., Sunyer, J., Iñiguez, C., Ballester, F., Garcia-Esteban, R., Forns, J., et al. (2015). Prenatal head growth and child neuropsychological development at age 14 months. Am. J. Obstet. Gynecol. 212, 661.e1–11. doi: 10.1016/j.ajog.2014.12.001
Al-Dosari, M. S., Shaheen, R., Colak, D., and Alkuraya, F. S. (2010). Novel CENPJ mutation causes Seckel syndrome. J. Med. Genet. 47, 411–414. doi: 10.1136/jmg.2009.076646
Amin, S., and Borrell, V. (2020). The Extracellular Matrix in the Evolution of Cortical Development and Folding. Front. Cell Dev. Biol. 8:604448. doi: 10.3389/fcell.2020.604448
Amiri, A., Coppola, G., Scuderi, S., Wu, F., Roychowdhury, T., Liu, F., et al. (2018). Transcriptome and epigenome landscape of human cortical development modeled in organoids. Science 362:eaat6720 doi: 10.1126/science.aat6720
An, H. L., Kuo, H. C., and Tang, T. K. (2022). Modeling Human Primary Microcephaly With hiPSC-Derived Brain Organoids Carrying CPAP-E1235V Disease-Associated Mutant Protein. Front. Cell Dev. Biol. 10:830432. doi: 10.3389/fcell.2022.830432
Antoniou, E., Orovou, E., Sarella, A., Iliadou, M., Rigas, N., Palaska, E., et al. (2020). Zika Virus and the Risk of Developing Microcephaly in Infants: A Systematic Review. Int. J. Environ. Res. Public Health 17:3806. doi: 10.3390/ijerph17113806
Ardhanareeswaran, K., Mariani, J., Coppola, G., Abyzov, A., and Vaccarino, F. M. (2017). Human induced pluripotent stem cells for modelling neurodevelopmental disorders. Nat. Rev. Neurol. 13, 265–278. doi: 10.1038/nrneurol.2017.45
Ashwal, S., Michelson, D., Plawner, L., and Dobyns, W. B. (2009). Practice parameter: Evaluation of the child with microcephaly (an evidence-based review): Report of the Quality Standards Subcommittee of the American Academy of Neurology and the Practice Committee of the Child Neurology Society. Neurology 73, 887–897. doi: 10.1212/WNL.0b013e3181b783f7
Ayo-Martin, A. C., Kyrousi, C., Di Giaimo, R., and Cappello, S. (2020). GNG5 Controls the Number of Apical and Basal Progenitors and Alters Neuronal Migration During Cortical Development. Front. Mol. Biosci. 7:578137. doi: 10.3389/fmolb.2020.578137
Baala, L., Briault, S., Etchevers, H. C., Laumonnier, F., Natiq, A., Amiel, J., et al. (2007). Homozygous silencing of T-box transcription factor EOMES leads to microcephaly with polymicrogyria and corpus callosum agenesis. Nat. Genet. 39, 454–456. doi: 10.1038/ng1993
Bagley, J. A., Reumann, D., Bian, S., Lévi-Strauss, J., and Knoblich, J. A. (2017). Fused cerebral organoids model interactions between brain regions. Nat. Methods 14, 743–751. doi: 10.1038/nmeth.4304
Bai, J., Ramos, R. L., Ackman, J. B., Thomas, A. M., Lee, R. V., and LoTurco, J. J. (2003). RNAi reveals doublecortin is required for radial migration in rat neocortex. Nat. Neurosci. 6, 1277–1283. doi: 10.1038/nn1153
Banerjee, A., Castro, J., and Sur, M. (2012). Rett syndrome: Genes, synapses, circuits, and therapeutics. Front. Psychiatry 3:34. doi: 10.3389/fpsyt.2012.00034
Barkovich, A. J., Guerrini, R., Kuzniecky, R. I., Jackson, G. D., and Dobyns, W. B. (2012). A developmental and genetic classification for malformations of cortical development: Update 2012. Brain 135, 1348–1369. doi: 10.1093/brain/aws019
Bauer, R., Timothy, K. W., and Golden, A. (2021). Update on the Molecular Genetics of Timothy Syndrome. Front. Pediatr. 9:668546. doi: 10.3389/fped.2021.668546
Benito-Kwiecinski, S., and Lancaster, M. A. (2020). Brain Organoids: Human Neurodevelopment in a Dish. Cold Spring Harb. Perspect. Biol. 12:a035709. doi: 10.1101/cshperspect.a035709
Bennett, J. A., Germani, T., Haqq, A. M., and Zwaigenbaum, L. (2015). Autism spectrum disorder in Prader-Willi syndrome: A systematic review. Am. J. Med. Genet. A. 167A, 2936–2944. doi: 10.1002/ajmg.a.37286
Bershteyn, M., Nowakowski, T. J., Pollen, A. A., Di Lullo, E., Nene, A., Wynshaw-Boris, A., et al. (2017). Human iPSC-derived cerebral organoids model cellular features of lissencephaly and reveal prolonged mitosis of outer radial glia. Cell Stem Cell 20, 435–449.e4. doi: 10.1016/j.stem.2016.12.007
Betizeau, M., Cortay, V., Patti, D., Pfister, S., Gautier, E., Bellemin-Ménard, A., et al. (2013). Precursor diversity and complexity of lineage relationships in the outer subventricular zone of the primate. Neuron 80, 442–457. doi: 10.1016/j.neuron.2013.09.032
Bird, C. P., Stranger, B. E., Liu, M., Thomas, D. J., Ingle, C. E., Beazley, C., et al. (2007). Fast-evolving noncoding sequences in the human genome. Genome Biol. 8:R118. doi: 10.1186/gb-2007-8-6-r118
Birey, F., Andersen, J., Makinson, C. D., Islam, S., Wei, W., Huber, N., et al. (2017). Assembly of functionally integrated human forebrain spheroids. Nature 545, 54–59. doi: 10.1038/nature22330
Bizzotto, S., Uzquiano, A., Dingli, F., Ershov, D., Houllier, A., Arras, G., et al. (2017). Eml1 loss impairs apical progenitor spindle length and soma shape in the developing cerebral cortex. Sci. Rep. 7:17308. doi: 10.1038/s41598-017-15253-4
Bjornsson, J., Short, M. P., Kwiatkowski, D. J., and Henske, E. P. (1996). Tuberous sclerosis-associated renal cell carcinoma. Clinical, pathological, and genetic features. Am. J. Pathol. 149, 1201–1208.
Boroviak, T., Stirparo, G. G., Dietmann, S., Hernando-Herraez, I., Mohammed, H., Reik, W., et al. (2018). Single cell transcriptome analysis of human, marmoset and mouse embryos reveals common and divergent features of preimplantation development. Development 145:dev167833. doi: 10.1242/dev.167833
Borrell, V., and Götz, M. (2014). Role of radial glial cells in cerebral cortex folding. Curr. Opin. Neurobiol. 27, 39–46. doi: 10.1016/j.conb.2014.02.007
Bouchatta, O., Manouze, H., Ba-M’Hamed, S., Landry, M., and Bennis, M. (2020). Neonatal 6-OHDA lesion model in mouse induces cognitive dysfunctions of attention-deficit/hyperactivity disorder (ADHD) during young age. Front. Behav. Neurosci. 14:27. doi: 10.3389/fnbeh.2020.00027
Broix, L., Jagline, H., Ivanova, E., Schmucker, S., Drouot, N., Clayton-Smith, J., et al. (2016). Mutations in the HECT domain of NEDD4L lead to AKT-mTOR pathway deregulation and cause periventricular nodular heterotopia. Nat. Genet. 48, 1349–1358. doi: 10.1038/ng.3676
Bubnys, A., and Tsai, L.-H. (2022). Harnessing cerebral organoids for Alzheimer’s disease research. Curr. Opin. Neurobiol. 72, 120–130. doi: 10.1016/j.conb.2021.10.003
Buchsbaum, I. Y., Kielkowski, P., Giorgio, G., O’Neill, A. C., Di Giaimo, R., Kyrousi, C., et al. (2020). ECE2 regulates neurogenesis and neuronal migration during human cortical development. EMBO Rep. 21:e48204. doi: 10.15252/embr.201948204
Butler, M. G., Dasouki, M. J., Zhou, X.-P., Talebizadeh, Z., Brown, M., Takahashi, T. N., et al. (2005). Subset of individuals with autism spectrum disorders and extreme macrocephaly associated with germline PTEN tumour suppressor gene mutations. J. Med. Genet. 42, 318–321. doi: 10.1136/jmg.2004.024646
Camp, J. G., Badsha, F., Florio, M., Kanton, S., Gerber, T., Wilsch-Bräuninger, M., et al. (2015). Human cerebral organoids recapitulate gene expression programs of fetal neocortex development. Proc. Natl. Acad. Sci. U.S.A. 112, 15672–15677. doi: 10.1073/pnas.1520760112
Cappello, S., Gray, M. J., Badouel, C., Lange, S., Einsiedler, M., Srour, M., et al. (2013). Mutations in genes encoding the cadherin receptor-ligand pair DCHS1 and FAT4 disrupt cerebral cortical development. Nat. Genet. 45, 1300–1308. doi: 10.1038/ng.2765
Carabalona, A., Beguin, S., Pallesi-Pocachard, E., Buhler, E., Pellegrino, C., Arnaud, K., et al. (2012). A glial origin for periventricular nodular heterotopia caused by impaired expression of Filamin-A. Hum. Mol. Genet. 21, 1004–1017. doi: 10.1093/hmg/ddr531
Chen, Y., Yu, J., Niu, Y., Qin, D., Liu, H., Li, G., et al. (2017). Modeling Rett Syndrome Using TALEN-Edited MECP2 Mutant Cynomolgus Monkeys. Cell 169, 945–955.e10.. doi: 10.1016/j.cell.2017.04.035
Chiaradia, I., and Lancaster, M. A. (2020). Brain organoids for the study of human neurobiology at the interface of in vitro and in vivo. Nat. Neurosci. 23, 1496–1508. doi: 10.1038/s41593-020-00730-3
Cholfin, J. A., and Rubenstein, J. L. R. (2008). Frontal cortex subdivision patterning is coordinately regulated by Fgf8. Fgf17, and Emx2. J. Comp. Neurol. 509, 144–155. doi: 10.1002/cne.21709
Cinque, S., Zoratto, F., Poleggi, A., Leo, D., Cerniglia, L., Cimino, S., et al. (2018). Behavioral Phenotyping of Dopamine Transporter Knockout Rats: Compulsive Traits, Motor Stereotypies, and Anhedonia. Front. Psychiatry 9:43. doi: 10.3389/fpsyt.2018.00043
Clipperton-Allen, A. E., Cohen, O. S., Aceti, M., Zucca, A., Levy, J., Ellegood, J., et al. (2019). Pten haploinsufficiency disrupts scaling across brain areas during development in mice. Transl. Psychiatry 9:329. doi: 10.1038/s41398-019-0656-6
Conforti, P., Besusso, D., Bocchi, V. D., Faedo, A., Cesana, E., Rossetti, G., et al. (2018). Faulty neuronal determination and cell polarization are reverted by modulating HD early phenotypes. Proc. Natl. Acad. Sci. 115:E762–E771. doi: 10.1073/pnas.1715865115
Corbo, J. C., Deuel, T. A., Long, J. M., LaPorte, P., Tsai, E., Wynshaw-Boris, A., et al. (2002). Doublecortin is required in mice for lamination of the hippocampus but not the neocortex. J. Neurosci. Off. J. Soc. Neurosci. 22, 7548–7557. doi: 10.1523/JNEUROSCI.22-17-07548.2002
Costa, V., Aigner, S., Vukcevic, M., Sauter, E., Behr, K., Ebeling, M., et al. (2016). mTORC1 inhibition corrects neurodevelopmental and synaptic alterations in a human stem cell model of tuberous sclerosis. Cell Rep. 15, 86–95. doi: 10.1016/j.celrep.2016.02.090
Cugola, F. R., Fernandes, I. R., Russo, F. B., Freitas, B. C., Dias, J. L. M., Guimarães, K. P., et al. (2016). The Brazilian Zika virus strain causes birth defects in experimental models. Nature 534, 267–271. doi: 10.1038/nature18296
Cushion, T. D., Dobyns, W. B., Mullins, J. G. L., Stoodley, N., Chung, S.-K., Fry, A. E., et al. (2013). Overlapping cortical malformations and mutations in TUBB2B and TUBA1A. Brain 136, 536–548. doi: 10.1093/brain/aws338
del Toro, D., Ruff, T., Cederfjäll, E., Villalba, A., Seyit-Bremer, G., Borrell, V., et al. (2017). Regulation of cerebral cortex folding by controlling neuronal migration via FLRT adhesion molecules. Cell 169, 621–635.e16. doi: 10.1016/j.cell.2017.04.012
Desikan, R. S., and Barkovich, A. J. (2016). Malformations of cortical development. Ann. Neurol. 80, 797–810. doi: 10.1002/ana.24793
Dobyns, W. B. (2010). The clinical patterns and molecular genetics of lissencephaly and subcortical band heterotopia. Epilepsia 51, 5–9. doi: 10.1111/j.1528-1167.2009.02433.x
Doernberg, E., and Hollander, E. (2016). Neurodevelopmental disorders (ASD and ADHD): DSM-5, ICD-10, and ICD-11. CNS Spectr. 21, 295–299. doi: 10.1017/S1092852916000262
Dudley, D. M., Aliota, M. T., Mohr, E. L., Newman, C. M., Golos, T. G., Friedrich, T. C., et al. (2019). Using macaques to address critical questions in zika virus research. Annu. Rev. Virol. 6, 481–500. doi: 10.1146/annurev-virology-092818-015732
Dutta, D., Heo, I., and Clevers, H. (2017). Disease Modeling in Stem Cell-Derived 3D Organoid Systems. Trends Mol. Med. 23, 393–410. doi: 10.1016/j.molmed.2017.02.007
Ehninger, D., Han, S., Shilyansky, C., Zhou, Y., Li, W., Kwiatkowski, D. J., et al. (2008). Reversal of learning deficits in a Tsc2+/- mouse model of tuberous sclerosis. Nat. Med. 14, 843–848. doi: 10.1038/nm1788
Eichmüller, O. L., Corsini, N. S., Vértesy, Á, Morassut, I., Scholl, T., Gruber, V.-E., et al. (2022). Amplification of human interneuron progenitors promotes brain tumors and neurological defects. Science 375:eabf5546. doi: 10.1126/science.abf5546
Elia, J., Glessner, J. T., Wang, K., Takahashi, N., Shtir, C. J., Hadley, D., et al. (2011). Genome-wide copy number variation study associates metabotropic glutamate receptor gene networks with attention deficit hyperactivity disorder. Nat. Genet. 44, 78–84. doi: 10.1038/ng.1013
Elitt, M. S., Barbar, L., and Tesar, P. J. (2018). Drug screening for human genetic diseases using iPSC models. Hum. Mol. Genet. 27, R89–R98. doi: 10.1093/hmg/ddy186
Feng, G., Jensen, F. E., Greely, H. T., Okano, H., Treue, S., Roberts, A. C., et al. (2020). Opportunities and limitations of genetically modified nonhuman primate models for neuroscience research. Proc. Natl. Acad. Sci. U.S.A. 117, 24022–24031. doi: 10.1073/pnas.2006515117
Fiddes, I. T., Lodewijk, G. A., Mooring, M., Bosworth, C. M., Ewing, A. D., Mantalas, G. L., et al. (2018). Human-specific NOTCH2NL genes affect notch signaling and cortical neurogenesis. Cell 173, 1356–1369.e22. doi: 10.1016/j.cell.2018.03.051
Fleck, M. W., Hirotsune, S., Gambello, M. J., Phillips-Tansey, E., Suares, G., Mervis, R. F., et al. (2000). Hippocampal abnormalities and enhanced excitability in a murine model of human lissencephaly. J. Neurosci. Off. J. Soc. Neurosci. 20, 2439–2450. doi: 10.1523/JNEUROSCI.20-07-02439.2000
Florio, M., Albert, M., Taverna, E., Namba, T., Brandl, H., Lewitus, E., et al. (2015). Human-specific gene ARHGAP11B promotes basal progenitor amplification and neocortex expansion. Science 347, 1465–1470. doi: 10.1126/science.aaa1975
Franchini, L. F. (2021). Genetic Mechanisms Underlying Cortical Evolution in Mammals. Front. Cell Dev. Biol. 9:591017. doi: 10.3389/fcell.2021.591017
Gabriel, E., Wason, A., Ramani, A., Gooi, L. M., Keller, P., Pozniakovsky, A., et al. (2016). CPAP promotes timely cilium disassembly to maintain neural progenitor pool. EMBO J. 35, 803–819. doi: 10.15252/embj.201593679
Galet, B., Cheval, H., and Ravassard, P. (2020). Patient-Derived Midbrain Organoids to Explore the Molecular Basis of Parkinson’s Disease. Front. Neurol. 11:1005. doi: 10.3389/fneur.2020.01005
Garcia-Forn, M., Boitnott, A., Akpinar, Z., and De Rubeis, S. (2020). Linking autism risk genes to disruption of cortical development. Cells 9:2500. doi: 10.3390/cells9112500
Gatto, C. L., and Broadie, K. (2011). Drosophila modeling of heritable neurodevelopmental disorders. Curr. Opin. Neurobiol. 21, 834–841. doi: 10.1016/j.conb.2011.04.009
Gécz, J., Cloosterman, D., and Partington, M. (2006). ARX: A gene for all seasons. Curr. Opin. Genet. Dev. 16, 308–316. doi: 10.1016/j.gde.2006.04.003
Ghatak, S., Dolatabadi, N., Trudler, D., Zhang, X., Wu, Y., Mohata, M., et al. (2019). Mechanisms of hyperexcitability in Alzheimer’s disease hiPSC-derived neurons and cerebral organoids vs isogenic controls. eLife 8:e50333. doi: 10.7554/eLife.50333
Gialluisi, A., Andlauer, T. F. M., Mirza-Schreiber, N., Moll, K., Becker, J., Hoffmann, P., et al. (2021). Genome-wide association study reveals new insights into the heritability and genetic correlates of developmental dyslexia. Mol. Psychiatry 26, 3004–3017. doi: 10.1038/s41380-020-00898-x
Goffin, D., and Zhou, Z. J. (2012). The neural circuit basis of Rett syndrome. Front. Biol. 7:428–435. doi: 10.1007/s11515-012-1248-5
Gomes, A. R., Fernandes, T. G., Vaz, S. H., Silva, T. P., Bekman, E. P., Xapelli, S., et al. (2020). Modeling Rett Syndrome With Human Patient-Specific Forebrain Organoids. Front. Cell Dev. Biol. 8:610427. doi: 10.3389/fcell.2020.610427
Gonzalez, C., Armijo, E., Bravo-Alegria, J., Becerra-Calixto, A., Mays, C. E., and Soto, C. (2018). Modeling amyloid beta and tau pathology in human cerebral organoids. Mol. Psychiatry 23, 2363–2374. doi: 10.1038/s41380-018-0229-8
Goorden, S. M. I., van Woerden, G. M., van der Weerd, L., Cheadle, J. P., and Elgersma, Y. (2007). Cognitive deficits in Tsc1+/- mice in the absence of cerebral lesions and seizures. Ann. Neurol. 62, 648–655. doi: 10.1002/ana.21317
Gorman, M. J., Caine, E. A., Zaitsev, K., Begley, M. C., Weger-Lucarelli, J., Uccellini, M. B., et al. (2018). An Immunocompetent mouse model of zika virus infection. Cell Host. Microbe 23, 672–685.e6. doi: 10.1016/j.chom.2018.04.003
Gross, C., Chang, C.-W., Kelly, S. M., Bhattacharya, A., McBride, S. M. J., Danielson, S. W., et al. (2015). Increased expression of the PI3K enhancer PIKE mediates deficits in synaptic plasticity and behavior in fragile X syndrome. Cell Rep. 11, 727–736. doi: 10.1016/j.celrep.2015.03.060
Guernsey, D. L., Jiang, H., Hussin, J., Arnold, M., Bouyakdan, K., Perry, S., et al. (2010). Mutations in centrosomal protein CEP152 in primary microcephaly families linked to MCPH4. Am. J. Hum. Genet. 87, 40–51. doi: 10.1016/j.ajhg.2010.06.003
Guerrini, R., Mei, D., Cordelli, D. M., Pucatti, D., Franzoni, E., and Parrini, E. (2012). Symmetric polymicrogyria and pachygyria associated with TUBB2B gene mutations. Eur. J. Hum. Genet. 20, 995–998. doi: 10.1038/ejhg.2012.21
Guo, W., Molinaro, G., Collins, K. A., Hays, S. A., Paylor, R., Worley, P. F., et al. (2016). Selective disruption of metabotropic glutamate receptor 5-homer interactions mimics phenotypes of fragile x syndrome in mice. J. Neurosci. Off. J. Soc. Neurosci. 36, 2131–2147. doi: 10.1523/JNEUROSCI.2921-15.2016
Guy, J., Hendrich, B., Holmes, M., Martin, J. E., and Bird, A. (2001). A Mouse Mecp2-null mutation causes neurological symptoms that mimic Rett syndrome. Nat. Genet. 27, 322–326. doi: 10.1038/85899
Halmøy, A., Fasmer, O. B., Gillberg, C., and Haavik, J. (2009). Occupational outcome in adult ADHD: Impact of symptom profile, comorbid psychiatric problems, and treatment: A cross-sectional study of 414 clinically diagnosed adult ADHD patients. J. Atten. Disord. 13, 175–187. doi: 10.1177/1087054708329777
Hansen, D. V., Lui, J. H., Parker, P. R. L., and Kriegstein, A. R. (2010). Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature 464, 554–561. doi: 10.1038/nature08845
Heide, M., Haffner, C., Murayama, A., Kurotaki, Y., Shinohara, H., Okano, H., et al. (2020). Human-specific ARHGAP11B increases size and folding of primate neocortex in the fetal marmoset. Science 369, 546–550. doi: 10.1126/science.abb2401
Heinzen, E. L., O’Neill, A. C., Zhu, X., Allen, A. S., Bahlo, M., Chelly, J., et al. (2018). De novo and inherited private variants in MAP1B in periventricular nodular heterotopia. PLoS Genet. 14:e1007281. doi: 10.1371/journal.pgen.1007281
Herman, T. E., and Siegel, M. J. (2008). Miller–Dieker syndrome, type 1 lissencephaly. J. Perinatol. 28, 313–315. doi: 10.1038/sj.jp.7211920
Hernández, D., Rooney, L. A., Daniszewski, M., Gulluyan, L., Liang, H. H., Cook, A. L., et al. (2022). Culture Variabilities of Human iPSC-Derived Cerebral Organoids Are a Major Issue for the Modelling of Phenotypes Observed in Alzheimer’s Disease. Stem Cell Rev. Rep. 18, 718–731. doi: 10.1007/s12015-021-10147-5
Ho, C. S. H., Dubeau, F., Séguin, R., and Ducharme, S. (2019). Prevalence of neuropsychiatric symptoms associated with malformations of cortical development. Epilepsy Behav. 92, 306–310. doi: 10.1016/j.yebeh.2019.01.011
Homan, K. A., Gupta, N., Kroll, K. T., Kolesky, D. B., Skylar-Scott, M., Miyoshi, T., et al. (2019). Flow-enhanced vascularization and maturation of kidney organoids in vitro. Nat. Methods 16, 255–262. doi: 10.1038/s41592-019-0325-y
Hong, S. E., Shugart, Y. Y., Huang, D. T., Shahwan, S. A., Grant, P. E., Hourihane, J. O., et al. (2000). Autosomal recessive lissencephaly with cerebellar hypoplasia is associated with human RELN mutations. Nat. Genet. 26, 93–96. doi: 10.1038/79246
Hor, J. H., Soh, E. S.-Y., Tan, L. Y., Lim, V. J. W., Santosa, M. M., and Winanto, et al. (2018). Cell cycle inhibitors protect motor neurons in an organoid model of Spinal Muscular Atrophy. Cell Death Dis. 9:1100. doi: 10.1038/s41419-018-1081-0
Howe, K., Clark, M. D., Torroja, C. F., Torrance, J., Berthelot, C., Muffato, M., et al. (2013). The zebrafish reference genome sequence and its relationship to the human genome. Nature 496, 498–503. doi: 10.1038/nature12111
Huang, W.-K., Wong, S. Z. H., Pather, S. R., Nguyen, P. T. T., Zhang, F., Zhang, D. Y., et al. (2021). Generation of hypothalamic arcuate organoids from human induced pluripotent stem cells. Cell Stem Cell 28, 1657–1670.e10. doi: 10.1016/j.stem.2021.04.006
Iefremova, V., Manikakis, G., Krefft, O., Jabali, A., Weynans, K., Wilkens, R., et al. (2017). An Organoid-Based Model of Cortical Development Identifies Non-Cell-Autonomous Defects in Wnt Signaling Contributing to Miller-Dieker Syndrome. Cell Rep. 19, 50–59. doi: 10.1016/j.celrep.2017.03.047
Insolera, R., Bazzi, H., Shao, W., Anderson, K. V., and Shi, S.-H. (2014). Cortical neurogenesis in the absence of centrioles. Nat. Neurosci. 17, 1528–1535. doi: 10.1038/nn.3831
Jabali, A., Hoffrichter, A., Uzquiano, A., Marsoner, F., Wilkens, R., Siekmann, M., et al. (2022). Human cerebral organoids reveal progenitor pathology in EML1-linked cortical malformation. EMBO Rep. 23:e54027. doi: 10.15252/embr.202154027
Janc, O. A., Hüser, M. A., Dietrich, K., Kempkes, B., Menzfeld, C., Hülsmann, S., et al. (2016). Systemic Radical Scavenger Treatment of a Mouse Model of Rett Syndrome: Merits and Limitations of the Vitamin E Derivative Trolox. Front. Cell Neurosci. 10:266. doi: 10.3389/fncel.2016.00266
Jansch, C., Günther, K., Waider, J., Ziegler, G. C., Forero, A., Kollert, S., et al. (2018). Generation of a human induced pluripotent stem cell (iPSC) line from a 51-year-old female with attention-deficit/hyperactivity disorder (ADHD) carrying a duplication of SLC2A3. Stem Cell Res. 28, 136–140. doi: 10.1016/j.scr.2018.02.005
Jarick, I., Volckmar, A.-L., Pütter, C., Pechlivanis, S., Nguyen, T. T., Dauvermann, M. R., et al. (2014). Genome-wide analysis of rare copy number variations reveals PARK2 as a candidate gene for attention-deficit/hyperactivity disorder. Mol. Psychiatry 19, 115–121. doi: 10.1038/mp.2012.161
Johnson, M. B., Sun, X., Kodani, A., Borges-Monroy, R., Girskis, K. M., Ryu, S. C., et al. (2018). Aspm knockout ferret reveals an evolutionary mechanism governing cerebral cortical size. Nature 556, 370–375. doi: 10.1038/s41586-018-0035-0
Ju, X.-C., Hou, Q.-Q., Sheng, A.-L., Wu, K.-Y., Zhou, Y., Jin, Y., et al. (2016). The hominoid-specific gene TBC1D3 promotes generation of basal neural progenitors and induces cortical folding in mice. eLife 5:e18197. doi: 10.7554/eLife.18197
Julander, J. G., Siddharthan, V., Park, A. H., Preston, E., Mathur, P., Bertolio, M., et al. (2018). Consequences of in utero exposure to Zika virus in offspring of AG129 mice. Sci. Rep. 8:9384. doi: 10.1038/s41598-018-27611-x
Juopperi, T. A., Kim, W. R., Chiang, C.-H., Yu, H., Margolis, R. L., Ross, C. A., et al. (2012). Astrocytes generated from patient induced pluripotent stem cells recapitulate features of Huntington’s disease patient cells. Mol. Brain 5:17. doi: 10.1186/1756-6606-5-17
Juric-Sekhar, G., Kapur, R. P., Glass, I. A., Murray, M. L., Parnell, S. E., and Hevner, R. F. (2011). Neuronal migration disorders in microcephalic osteodysplastic primordial dwarfism type I/III. Acta Neuropathol. 121, 545–554. doi: 10.1007/s00401-010-0748-0
Kadoshima, T., Sakaguchi, H., Nakano, T., Soen, M., Ando, S., Eiraku, M., et al. (2013). Self-organization of axial polarity, inside-out layer pattern, and species-specific progenitor dynamics in human ES cell-derived neocortex. Proc. Natl. Acad. Sci. U.S.A. 110, 20284–20289. doi: 10.1073/pnas.1315710110
Kalebic, N., and Huttner, W. B. (2020). Basal progenitor morphology and neocortex evolution. Trends Neurosci. 43, 843–853. doi: 10.1016/j.tins.2020.07.009
Kang, Y., Zhou, Y., Li, Y., Han, Y., Xu, J., Niu, W., et al. (2021). A human forebrain organoid model of fragile X syndrome exhibits altered neurogenesis and highlights new treatment strategies. Nat. Neurosci. 24, 1377–1391. doi: 10.1038/s41593-021-00913-6
Kanton, S., Boyle, M. J., He, Z., Santel, M., Weigert, A., Sanchís-Calleja, F., et al. (2019). Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 574, 418–422. doi: 10.1038/s41586-019-1654-9
Karzbrun, E., Kshirsagar, A., Cohen, S. R., Hanna, J. H., and Reiner, O. (2018). Human Brain Organoids on a Chip Reveal the Physics of Folding. Nat. Phys. 14, 515–522. doi: 10.1038/s41567-018-0046-7
Kasahara, Y., Kubo, Y., and Sora, I. (2013). [Analysis of dopamine transporter knockout mice as an animal model of AD/HD]. Nihon Shinkei Seishin Yakurigaku Zasshi 33, 185–189.
Kathuria, A., Lopez-Lengowski, K., Jagtap, S. S., McPhie, D., Perlis, R. H., Cohen, B. M., et al. (2020). Transcriptomic Landscape and Functional Characterization of Induced Pluripotent Stem Cell-Derived Cerebral Organoids in Schizophrenia. JAMA Psychiatry 77, 745–754. doi: 10.1001/jamapsychiatry.2020.0196
Kattuoa, M. L., and Das, J. M. (2022). “Lissencephaly,” in StatPearls, (Treasure Island, FL: StatPearls Publishing).
Kazdoba, T. M., Leach, P. T., and Crawley, J. N. (2016). Behavioral phenotypes of genetic mouse models of autism. Genes Brain Behav. 15, 7–26. doi: 10.1111/gbb.12256
Kazdoba, T. M., Leach, P. T., Silverman, J. L., and Crawley, J. N. (2014). Modeling fragile X syndrome in the Fmr1 knockout mouse. Intractable Rare Dis. Res. 3, 118–133. doi: 10.5582/irdr.2014.01024
Kielar, M., Tuy, F. P. D., Bizzotto, S., Lebrand, C., de Juan Romero, C., Poirier, K., et al. (2014). Mutations in Eml1 lead to ectopic progenitors and neuronal heterotopia in mouse and human. Nat. Neurosci. 17, 923–933. doi: 10.1038/nn.3729
Kim, B.-N., Cho, S.-C., Kim, Y., Shin, M.-S., Yoo, H.-J., Kim, J.-W., et al. (2009). Phthalates exposure and attention-deficit/hyperactivity disorder in school-age children. Biol. Psychiatry 66, 958–963. doi: 10.1016/j.biopsych.2009.07.034
Kim, H., Park, H. J., Choi, H., Chang, Y., Park, H., Shin, J., et al. (2019). Modeling G2019S-LRRK2 Sporadic Parkinson’s Disease in 3D Midbrain Organoids. Stem Cell Rep. 12, 518–531. doi: 10.1016/j.stemcr.2019.01.020
Kim, H.-T., Lee, M.-S., Choi, J.-H., Jung, J.-Y., Ahn, D.-G., Yeo, S.-Y., et al. (2011). The microcephaly gene aspm is involved in brain development in zebrafish. Biochem. Biophys. Res. Commun. 409, 640–644. doi: 10.1016/j.bbrc.2011.05.056
Klaus, J., Kanton, S., Kyrousi, C., Ayo-Martin, A. C., Di Giaimo, R., Riesenberg, S., et al. (2019). Altered neuronal migratory trajectories in human cerebral organoids derived from individuals with neuronal heterotopia. Nat. Med. 25, 561–568. doi: 10.1038/s41591-019-0371-0
Klingler, E., Francis, F., Jabaudon, D., and Cappello, S. (2021). Mapping the molecular and cellular complexity of cortical malformations. Science 371:eaba4517. doi: 10.1126/science.aba4517
Kundakovic, M., Gudsnuk, K., Franks, B., Madrid, J., Miller, R. L., Perera, F. P., et al. (2013). Sex-specific epigenetic disruption and behavioral changes following low-dose in utero bisphenol A exposure. Proc. Natl. Acad. Sci. U.S.A. 110, 9956–9961. doi: 10.1073/pnas.1214056110
Kwiatkowski, D. J., Zhang, H., Bandura, J. L., Heiberger, K. M., Glogauer, M., el-Hashemite, N., et al. (2002). A mouse model of TSC1 reveals sex-dependent lethality from liver hemangiomas, and up-regulation of p70S6 kinase activity in Tsc1 null cells. Hum. Mol. Genet. 11, 525–534. doi: 10.1093/hmg/11.5.525
Kyrousi, C., and Cappello, S. (2020). Using brain organoids to study human neurodevelopment, evolution and disease. WIREs Dev. Biol. 9:e347. doi: 10.1002/wdev.347
Kyrousi, C., O’Neill, A. C., Brazovskaja, A., He, Z., Kielkowski, P., Coquand, L., et al. (2021). Extracellular LGALS3BP regulates neural progenitor position and relates to human cortical complexity. Nat. Commun. 12:6298. doi: 10.1038/s41467-021-26447-w
Lancaster, M. A., and Knoblich, J. A. (2014). Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 9, 2329–2340. doi: 10.1038/nprot.2014.158
Lancaster, M. A., Renner, M., Martin, C. A., Wenzel, D., Bicknell, L. S., Hurles, M. E., et al. (2013). Cerebral organoids model human brain development and microcephaly. Nature 501, 373–379. doi: 10.1038/nature12517
Lapray, D., Popova, I. Y., Kindler, J., Jorquera, I., Becq, H., Manent, J.-B., et al. (2010). Spontaneous epileptic manifestations in a DCX knockdown model of human double cortex. Cereb. Cortex 20, 2694–2701. doi: 10.1093/cercor/bhq014
Lee, H.-K., Velazquez Sanchez, C., Chen, M., Morin, P. J., Wells, J. M., Hanlon, E. B., et al. (2016). Three Dimensional Human Neuro-Spheroid Model of Alzheimer’s Disease Based on Differentiated Induced Pluripotent Stem Cells. PLoS One 11:e0163072. doi: 10.1371/journal.pone.0163072
Lee, J. H., Huynh, M., Silhavy, J. L., Kim, S., Dixon-Salazar, T., Heiberg, A., et al. (2012). De novo somatic mutations in components of the PI3K-AKT3-mTOR pathway cause hemimegalencephaly. Nat. Genet. 44, 941–945. doi: 10.1038/ng.2329
Leruez-Ville, M., and Ville, Y. (2017). Fetal cytomegalovirus infection. Best Pract. Res. Clin. Obstet. Gynaecol. 38, 97–107. doi: 10.1016/j.bpobgyn.2016.10.005
Lesch, K.-P., Selch, S., Renner, T. J., Jacob, C., Nguyen, T. T., Hahn, T., et al. (2011). Genome-wide copy number variation analysis in attention-deficit/hyperactivity disorder: Association with neuropeptide Y gene dosage in an extended pedigree. Mol. Psychiatry 16, 491–503. doi: 10.1038/mp.2010.29
Leventer, R. J., Jansen, A., Pilz, D. T., Stoodley, N., Marini, C., Dubeau, F., et al. (2010). Clinical and imaging heterogeneity of polymicrogyria: A study of 328 patients. Brain 133, 1415–1427. doi: 10.1093/brain/awq078
Li, C., Wang, Q., Jiang, Y., Ye, Q., Xu, D., Gao, F., et al. (2018). Disruption of glial cell development by Zika virus contributes to severe microcephalic newborn mice. Cell Discov. 4:43. doi: 10.1038/s41421-018-0042-1
Li, C., Xu, D., Ye, Q., Hong, S., Jiang, Y., Liu, X., et al. (2016). Zika Virus Disrupts Neural Progenitor Development and Leads to Microcephaly in Mice. Cell Stem Cell 19, 120–126. doi: 10.1016/j.stem.2016.04.017
Li, M., Santpere, G., Imamura Kawasawa, Y., Evgrafov, O. V., Gulden, F. O., Pochareddy, S., et al. (2018). Integrative functional genomic analysis of human brain development and neuropsychiatric risks. Science 362:eaat7615.
Li, R., Sun, L., Fang, A., Li, P., Wu, Q., and Wang, X. (2017a). Recapitulating cortical development with organoid culture in vitro and modeling abnormal spindle-like (ASPM related primary) microcephaly disease. Protein Cell 8, 823–833. doi: 10.1007/s13238-017-0479-2
Li, Y., Cao, J., Chen, M., Li, J., Sun, Y., Zhang, Y., et al. (2017b). Abnormal Neural Progenitor Cells Differentiated from Induced Pluripotent Stem Cells Partially Mimicked Development of TSC2 Neurological Abnormalities. Stem Cell Rep. 8, 883–893. doi: 10.1016/j.stemcr.2017.02.020
Li, Y., Muffat, J., Omer, A., Bosch, I., Lancaster, M. A., Sur, M., et al. (2017c). Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell 20, 385–396.e3. doi: 10.1016/j.stem.2016.11.017
Li, S., Jin, Z., Koirala, S., Bu, L., Xu, L., Hynes, R. O., et al. (2008). GPR56 regulates pial basement membrane integrity and cortical lamination. J. Neurosci. 28, 5817–5826. doi: 10.1523/JNEUROSCI.0853-08.2008
Lin, Y., Lee, Y., Li, S., and Tang, T. K. (2020). Loss of CPAP in developing mouse brain and its functional implication for human primary microcephaly. J. Cell Sci. 133:jcs243592 doi: 10.1242/jcs.243592
Liu, J., Liu, W., Yang, L., Wu, Q., Zhang, H., Fang, A., et al. (2017). The primate-specific gene TMEM14B marks outer radial glia cells and promotes cortical expansion and folding. Cell Stem Cell 21, 635–649.e8. doi: 10.1016/j.stem.2017.08.013
Liu, T. X., Howlett, N. G., Deng, M., Langenau, D. M., Hsu, K., Rhodes, J., et al. (2003). Knockdown of zebrafish Fancd2 causes developmental abnormalities via p53-dependent apoptosis. Dev. Cell. 5, 903–914. doi: 10.1016/S1534-5807(03)00339-3
Liu, Z., Wang, X., Newman, N., Grant, K. A., Studholme, C., and Kroenke, C. D. (2020). Anatomical and diffusion MRI brain atlases of the fetal rhesus macaque brain at 85, 110 and 135 days gestation. Neuroimage 206:116310. doi: 10.1016/j.neuroimage.2019.116310
Long, K. R., Newland, B., Florio, M., Kalebic, N., Langen, B., Kolterer, A., et al. (2018). Extracellular matrix components HAPLN1, lumican, and collagen i cause hyaluronic acid-dependent folding of the developing human neocortex. Neuron 99, 702–719.e6. doi: 10.1016/j.neuron.2018.07.013
Madrid, M., Sumen, C., Aivio, S., and Saklayen, N. (2021). Autologous induced pluripotent stem cell–based cell therapies: Promise, progress, and challenges. Curr. Protoc. 1:e88. doi: 10.1002/cpz1.88
Mansour, A. A., Gonçalves, J. T., Bloyd, C. W., Li, H., Fernandes, S., Quang, D., et al. (2018). An in vivo model of functional and vascularized human brain organoids. Nat. Biotechnol. 36, 432–441. doi: 10.1038/nbt.4127
Marchetto, M. C. N., Carromeu, C., Acab, A., Yu, D., Yeo, G. W., Mu, Y., et al. (2010). A model for neural development and treatment of Rett syndrome using human induced pluripotent stem cells. Cell 143, 527–539. doi: 10.1016/j.cell.2010.10.016
Marco, E. J., and Skuse, D. H. (2006). Autism-lessons from the X chromosome. Soc. Cogn. Affect. Neurosci. 1, 183–193. doi: 10.1093/scan/nsl028
Martin, P., Wagh, V., Reis, S. A., Erdin, S., Beauchamp, R. L., Shaikh, G., et al. (2020). TSC patient-derived isogenic neural progenitor cells reveal altered early neurodevelopmental phenotypes and rapamycin-induced MNK-eIF4E signaling. Mol. Autism 11:2. doi: 10.1186/s13229-019-0311-3
Matsumoto, N., Hoshiba, Y., Morita, K., Uda, N., Hirota, M., Minamikawa, M., et al. (2017). Pathophysiological analyses of periventricular nodular heterotopia using gyrencephalic mammals. Hum. Mol. Genet. 26, 1173–1181. doi: 10.1093/hmg/ddx038
Matsumoto, N., Leventer, R. J., Kuc, J. A., Mewborn, S. K., Dudlicek, L. L., Ramocki, M. B., et al. (2001). Mutation analysis of the DCX gene and genotype/phenotype correlation in subcortical band heterotopia. Eur. J. Hum. Genet. 9, 5–12. doi: 10.1038/sj.ejhg.5200548
McDougle, C. J., Erickson, C. A., Stigler, K. A., and Posey, D. J. (2005). Neurochemistry in the pathophysiology of autism. J. Clin. Psychiatry 66, 9–18.
McIntyre, R. E., Lakshminarasimhan Chavali, P., Ismail, O., Carragher, D. M., Sanchez-Andrade, G., Forment, J. V., et al. (2012). Disruption of mouse Cenpj, a regulator of centriole biogenesis, phenocopies Seckel syndrome. PLoS Genet. 8:e1003022. doi: 10.1371/journal.pgen.1003022
Meikle, L., Talos, D. M., Onda, H., Pollizzi, K., Rotenberg, A., Sahin, M., et al. (2007). A mouse model of tuberous sclerosis: Neuronal loss of Tsc1 causes dysplastic and ectopic neurons, reduced myelination, seizure activity, and limited survival. J. Neurosci. 27, 5546–5558. doi: 10.1523/JNEUROSCI.5540-06.2007
Mellios, N., Feldman, D. A., Sheridan, S. D., Ip, J. P. K., Kwok, S., Amoah, S. K., et al. (2018). MeCP2-regulated miRNAs control early human neurogenesis through differential effects on ERK and AKT signaling. Mol. Psychiatry 23, 1051–1065. doi: 10.1038/mp.2017.86
Meloni, I., Bruttini, M., Longo, I., Mari, F., Rizzolio, F., D’Adamo, P., et al. (2000). A mutation in the rett syndrome gene, MECP2, causes X-linked mental retardation and progressive spasticity in males. Am. J. Hum. Genet. 67, 982–985. doi: 10.1086/303078
Merkle, F. T., Maroof, A., Wataya, T., Sasai, Y., Studer, L., Eggan, K., et al. (2015). Generation of neuropeptidergic hypothalamic neurons from human pluripotent stem cells. Development 142, 633–643. doi: 10.1242/dev.117978
Mertens, J., Wang, Q.-W., Kim, Y., Yu, D. X., Pham, S., Yang, B., et al. (2015). Differential responses to lithium in hyperexcitable neurons from patients with bipolar disorder. Nature 527, 95–99. doi: 10.1038/nature15526
Mineyko, A., Doja, A., Hurteau, J., Dobyns, W. B., Das, S., and Boycott, K. M. (2010). A novel missense mutation in LIS1 in a child with subcortical band heterotopia and pachygyria inherited from his mildly affected mother with somatic mosaicism. J. Child Neurol. 25, 738–741. doi: 10.1177/0883073809343312
Miura, Y., Li, M.-Y., Birey, F., Ikeda, K., Revah, O., Thete, M. V., et al. (2020). Generation of human striatal organoids and cortico-striatal assembloids from human pluripotent stem cells. Nat. Biotechnol. 38, 1421–1430. doi: 10.1038/s41587-020-00763-w
Moore, C. A., Staples, J. E., Dobyns, W. B., Pessoa, A., Ventura, C. V., Fonseca, E. B., et al. (2017). Characterizing the pattern of anomalies in congenital zika syndrome for pediatric clinicians. JAMA Pediatr. 171, 288–295. doi: 10.1001/jamapediatrics.2016.3982
Moretti, P., Levenson, J. M., Battaglia, F., Atkinson, R., Teague, R., Antalffy, B., et al. (2006). Learning and memory and synaptic plasticity are impaired in a mouse model of Rett syndrome. J. Neurosci. 26, 319–327. doi: 10.1523/JNEUROSCI.2623-05.2006
Mountford, H. S., Hill, A., Barnett, A. L., and Newbury, D. F. (2021). Genome-Wide Association Study of Motor Coordination [Internet]. Front. Hum. Neurosci. 15:669902. doi: 10.3389/fnhum.2021.669902
Muhle, R., Trentacoste, S. V., and Rapin, I. (2004). The genetics of autism. Pediatrics 113:e472–e486. doi: 10.1542/peds.113.5.e472
Murayama, A. Y., Kuwako, K., Okahara, J., Bae, B.-I., Okuno, M., Mashiko, H., et al. (2020). The polymicrogyria-associated GPR56 promoter preferentially drives gene expression in developing GABAergic neurons in common marmosets. Sci. Rep. 10:21516. doi: 10.1038/s41598-020-78608-4
Nguyen, S. M., Antony, K. M., Dudley, D. M., Kohn, S., Simmons, H. A., Wolfe, B., et al. (2017). Highly efficient maternal-fetal Zika virus transmission in pregnant rhesus macaques. PLoS Pathog. 13:e1006378. doi: 10.1371/journal.ppat.1006378
Niemi, M. E. K., Martin, H. C., Rice, D. L., Gallone, G., Gordon, S., Kelemen, M., et al. (2018). Common genetic variants contribute to risk of rare severe neurodevelopmental disorders. Nature 562, 268–271. doi: 10.1038/s41586-018-0566-4
Noctor, S. C., Martínez-Cerdeño, V., Ivic, L., and Kriegstein, A. R. (2004). Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat. Neurosci. 7, 136–144. doi: 10.1038/nn1172
Nonaka-Kinoshita, M., Reillo, I., Artegiani, B., Martínez-Martínez, M. Á, Nelson, M., Borrell, V., et al. (2013). Regulation of cerebral cortex size and folding by expansion of basal progenitors. EMBO J. 32, 1817–1828. doi: 10.1038/emboj.2013.96
Oegema, R., Baillat, D., Schot, R., van Unen, L. M., Brooks, A., Kia, S. K., et al. (2017). Human mutations in integrator complex subunits link transcriptome integrity to brain development. PLoS Genet. 13:e1006809. doi: 10.1371/journal.pgen.1006809
Oegema, R., McGillivray, G., Leventer, R., Le Moing, A.-G., Bahi-Buisson, N., Barnicoat, A., et al. (2019). EML1-associated brain overgrowth syndrome with ribbon-like heterotopia. Am. J. Med. Genet. C Semin Med. Genet. 181, 627–637. doi: 10.1002/ajmg.c.31751
Omer Javed, A., Li, Y., Muffat, J., Su, K.-C., Cohen, M. A., Lungjangwa, T., et al. (2018). Microcephaly modeling of kinetochore mutation reveals a brain-specific phenotype. Cell Rep. 25, 319–27. doi: 10.1016/j.celrep.2018.09.032
O’Neill, A. C., Kyrousi, C., Einsiedler, M., Burtscher, I., Drukker, M., Markie, D. M., et al. (2018a). Mob2 Insufficiency Disrupts Neuronal Migration in the Developing Cortex. Front. Cell Neurosci. 12:57. doi: 10.3389/fncel.2018.00057
O’Neill, A. C., Kyrousi, C., Klaus, J., Leventer, R. J., Kirk, E. P., Fry, A., et al. (2018b). A primate-specific isoform of PLEKHG6 regulates neurogenesis and neuronal migration. Cell Rep. 25, 2729–2741.e6. doi: 10.1016/j.celrep.2018.11.029
Ormel, P. R., Vieira de Sá, R., van Bodegraven, E. J., Karst, H., Harschnitz, O., Sneeboer, M. A. M., et al. (2018). Microglia innately develop within cerebral organoids. Nat. Commun. 9:4167. doi: 10.1038/s41467-018-06684-2
Osaki, T., Uzel, S. G. M., and Kamm, R. D. (2018). Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human iPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 4:eaat5847. doi: 10.1126/sciadv.aat5847
Ougrin, D., Chatterton, S., and Banarsee, R. (2010). Attention deficit hyperactivity disorder (ADHD): Review for primary care clinicians. London J. Prim. Care 3, 45–51. doi: 10.1080/17571472.2010.11493296
Parens, R., Nijhout, H. F., Morales, A., Xavier Costa, F., and Bar-Yam, Y. (2017). A possible link between pyriproxyfen and microcephaly. PLoS Curr. 9:ecurrents.outbreaks.5afb0bfb8cf31d9a4baba7b19b4edbac. doi: 10.1371/currents.outbreaks.5afb0bfb8cf31d9a4baba7b19b4edbac
Parenti, I., Rabaneda, L. G., Schoen, H., and Novarino, G. (2020). Neurodevelopmental disorders: From genetics to functional pathways. Trends Neurosci. 43, 608–621. doi: 10.1016/j.tins.2020.05.004
Park, I.-H., Arora, N., Huo, H., Maherali, N., Ahfeldt, T., Shimamura, A., et al. (2008). Disease-specific induced pluripotent stem cells. Cell 134, 877–886. doi: 10.1016/j.cell.2008.07.041
Park, J.-H., Park, H.-S., Hong, S., and Kang, S. (2016). Motor neurons derived from ALS-related mouse iPS cells recapitulate pathological features of ALS. Exp. Mol. Med. 48:e276. doi: 10.1038/emm.2016.113
Parrini, E., Conti, V., Dobyns, W. B., and Guerrini, R. (2016). Genetic basis of brain malformations. Mol. Syndromol. 7, 220–233. doi: 10.1159/000448639
Paylor, R., Hirotsune, S., Gambello, M. J., Yuva-Paylor, L., Crawley, J. N., and Wynshaw-Boris, A. (1999). Impaired learning and motor behavior in heterozygous Pafah1b1 (Lis1) mutant mice. Learn. Mem. 6, 521–537. doi: 10.1101/lm.6.5.521
Paşca, S. P. (2018). The rise of three-dimensional human brain cultures. Nature 553, 437–445. doi: 10.1038/nature25032
Piao, X., Hill, R. S., Bodell, A., Chang, B. S., Basel-Vanagaite, L., Straussberg, R., et al. (2004). G protein-coupled receptor-dependent development of human frontal cortex. Science 303, 2033–2036. doi: 10.1126/science.1092780
Pinson, A., Xing, L., Namba, T., Kalebic, N., Peters, J., Oegema, C. E., et al. (2022). Human TKTL1 implies greater neurogenesis in frontal neocortex of modern humans than Neanderthals. Science 377:eabl6422. doi: 10.1126/science.abl6422
Poirier, K., Saillour, Y., Bahi-Buisson, N., Jaglin, X. H., Fallet-Bianco, C., Nabbout, R., et al. (2010). Mutations in the neuronal ß-tubulin subunit TUBB3 result in malformation of cortical development and neuronal migration defects. Hum. Mol. Genet. 19, 4462–4473. doi: 10.1093/hmg/ddq377
Powell, K. (2022). Hybrid brains: The ethics of transplanting human neurons into animals. Nature 608, 22–25. doi: 10.1038/d41586-022-02073-4
Qian, X., Jacob, F., Song, M. M., Nguyen, H. N., Song, H., and Ming, G.-L. (2018). Generation of human brain region-specific organoids using a miniaturized spinning bioreactor. Nat. Protoc. 13, 565–580. doi: 10.1038/nprot.2017.152
Qian, X., Nguyen, H. N., Song, M. M., Hadiono, C., Ogden, S. C., Hammack, C., et al. (2016). Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell 165, 1238–1254. doi: 10.1016/j.cell.2016.04.032
Qian, X., Song, H., and Ming, G.-L. (2019). Brain organoids: Advances, applications and challenges. Development 146:dev166074. doi: 10.1242/dev.166074
Quadrato, G., Nguyen, T., Macosko, E. Z., Sherwood, J. L., Min Yang, S., Berger, D. R., et al. (2017). Cell diversity and network dynamics in photosensitive human brain organoids. Nature 545, 48–53. doi: 10.1038/nature22047
Reardon, S. (2020). Can lab-grown brains become conscious? Nature 586, 658–661. doi: 10.1038/d41586-020-02986-y
Renner, H., Grabos, M., Becker, K. J., Kagermeier, T. E., Wu, J., Otto, M., et al. (2020). A fully automated high-throughput workflow for 3D-based chemical screening in human midbrain organoids. eLife 9:e52904. doi: 10.7554/eLife.52904
Rochester, J. R., Bolden, A. L., and Kwiatkowski, C. F. (2018). Prenatal exposure to bisphenol A and hyperactivity in children: A systematic review and meta-analysis. Environ. Int. 114, 343–356. doi: 10.1016/j.envint.2017.12.028
Rubenstein, J. L. R., and Merzenich, M. M. (2003). Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2, 255–267. doi: 10.1034/j.1601-183X.2003.00037.x
Russell, V. A. (2011). Overview of animal models of attention deficit hyperactivity disorder (ADHD). Curr. Protoc. Neurosci. Chapter 9:Unit9.35. doi: 10.1002/0471142301.ns0935s54
Rylaarsdam, L., and Guemez-Gamboa, A. (2019). Genetic Causes and Modifiers of Autism Spectrum Disorder. Front. Cell Neurosci. 13:385. doi: 10.3389/fncel.2019.00385
Sahu, S., Buhler, E., Vermoyal, J.-C., Watrin, F., Represa, A., and Manent, J.-B. (2019). Spontaneous epileptiform activity in a Rat model of bilateral subcortical band heterotopia. Epilepsia 60, 337–348. doi: 10.1111/epi.14633
Sánchez-Danés, A., Richaud-Patin, Y., Carballo-Carbajal, I., Jiménez-Delgado, S., Caig, C., Mora, S., et al. (2012). Disease-specific phenotypes in dopamine neurons from human iPS-based models of genetic and sporadic Parkinson’s disease. EMBO Mol. Med. 4, 380–395. doi: 10.1002/emmm.201200215
Sawada, T., Chater, T. E., Sasagawa, Y., Yoshimura, M., Fujimori-Tonou, N., Tanaka, K., et al. (2020). Developmental excitation-inhibition imbalance underlying psychoses revealed by single-cell analyses of discordant twins-derived cerebral organoids. Mol. Psychiatry 25, 2695–2711. doi: 10.1038/s41380-020-0844-z
Schwartz, M. P., Hou, Z., Propson, N. E., Zhang, J., Engstrom, C. J., Santos Costa, V., et al. (2015). Human pluripotent stem cell-derived neural constructs for predicting neural toxicity. Proc. Natl. Acad. Sci. U.S.A. 112, 12516–12521. doi: 10.1073/pnas.1516645112
Seo, J., Kritskiy, O., Watson, L. A., Barker, S. J., Dey, D., Raja, W. K., et al. (2017). Inhibition of p25/Cdk5 Attenuates Tauopathy in Mouse and iPSC Models of Frontotemporal Dementia. J. Neurosci. 37, 9917–9924. doi: 10.1523/JNEUROSCI.0621-17.2017
Severino, M., Geraldo, A. F., Utz, N., Tortora, D., Pogledic, I., Klonowski, W., et al. (2020). Definitions and classification of malformations of cortical development: Practical guidelines. Brain 143, 2874–2894. doi: 10.1093/brain/awaa174
Shaheen, R., Sebai, M. A., Patel, N., Ewida, N., Kurdi, W., Altweijri, I., et al. (2017). The genetic landscape of familial congenital hydrocephalus. Ann. Neurol. 81, 890–897. doi: 10.1002/ana.24964
Shahsavani, M., Pronk, R. J., Falk, R., Lam, M., Moslem, M., Linker, S. B., et al. (2018). An in vitro model of lissencephaly: Expanding the role of DCX during neurogenesis. Mol. Psychiatry 23, 1674–1684. doi: 10.1038/mp.2017.175
Shao, Q., Herrlinger, S., Yang, S.-L., Lai, F., Moore, J. M., Brindley, M. A., et al. (2016). Zika virus infection disrupts neurovascular development and results in postnatal microcephaly with brain damage. Development 143, 4127–4136. doi: 10.1242/dev.143768
Sheen, V. L. (2012). Periventricular heterotopia: Shuttling of proteins through vesicles and actin in cortical development and disease. Scientifica 2012:480129. doi: 10.6064/2012/480129
Sheen, V. L. (2014). Filamin A mediated Big2 dependent endocytosis: From apical abscission to periventricular heterotopia. Tissue Barriers 2:e29431. doi: 10.4161/tisb.29431
Silva, T. P., Sousa-Luís, R., Fernandes, T. G., Bekman, E. P., Rodrigues, C. A. V., Vaz, S. H., et al. (2021). Transcriptome profiling of human pluripotent stem cell-derived cerebellar organoids reveals faster commitment under dynamic conditions. Biotechnol. Bioeng. 118, 2781–2803. doi: 10.1002/bit.27797
Simmons, A. J., Park, R., Sterling, N. A., Jang, M.-H., van Deursen, J. M. A., Yen, T. J., et al. (2019). Nearly complete deletion of BubR1 causes microcephaly through shortened mitosis and massive cell death. Hum. Mol. Genet. 28, 1822–1836. doi: 10.1093/hmg/ddz022
Sisodia, R., Raj, R. K. S., and Goel, V. (2014). Seckel syndrome: A rare case report. J. Indian Soc. Pedod. Prev. Dent. 32, 160–163. doi: 10.4103/0970-4388.130983
Sitzmann, A. F., Hagelstrom, R. T., Tassone, F., Hagerman, R. J., and Butler, M. G. (2018). Rare FMR1 gene mutations causing fragile X syndrome: A review. Am. J. Med. Genet. A 176, 11–18. doi: 10.1002/ajmg.a.38504
Smart, I. H. M., Dehay, C., Giroud, P., Berland, M., and Kennedy, H. (2002). Unique morphological features of the proliferative zones and postmitotic compartments of the neural epithelium giving rise to striate and extrastriate cortex in the monkey. Cereb. Cortex 12, 37–53. doi: 10.1093/cercor/12.1.37
Smith, R. S., Kenny, C. J., Ganesh, V., Jang, A., Borges-Monroy, R., Partlow, J. N., et al. (2018). Sodium Channel SCN3A (Na(V)1.3) Regulation of Human Cerebral Cortical Folding and Oral Motor Development. Neuron 99, 905–913.e7. doi: 10.1016/j.neuron.2018.07.052
Smits, L. M., Reinhardt, L., Reinhardt, P., Glatza, M., Monzel, A. S., Stanslowsky, N., et al. (2019). Modeling Parkinson’s disease in midbrain-like organoids. NPJ Park. Dis. 5:5. doi: 10.1038/s41531-019-0078-4
Sochacki, J., Devalle, S., Reis, M., Mattos, P., and Rehen, S. (2016). Generation of urine iPS cell lines from patients with Attention Deficit Hyperactivity Disorder (ADHD) using a non-integrative method. Stem Cell Res. 17, 102–106. doi: 10.1016/j.scr.2016.05.015
Srikanth, P., Lagomarsino, V. N., Muratore, C. R., Ryu, S. C., He, A., Taylor, W. M., et al. (2018). Shared effects of DISC1 disruption and elevated WNT signaling in human cerebral organoids. Transl. Psychiatry 8:77. doi: 10.1038/s41398-018-0122-x
Stearns, N. A., Schaevitz, L. R., Bowling, H., Nag, N., Berger, U. V., and Berger-Sweeney, J. (2007). Behavioral and anatomical abnormalities in Mecp2 mutant mice: A model for Rett syndrome. Neuroscience 146, 907–921. doi: 10.1016/j.neuroscience.2007.02.009
Stottmann, R. W., Donlin, M., Hafner, A., Bernard, A., Sinclair, D. A., and Beier, D. R. (2013). A mutation in Tubb2b, a human polymicrogyria gene, leads to lethality and abnormal cortical development in the mouse. Hum. Mol. Genet. 22, 4053–4063. doi: 10.1093/hmg/ddt255
Stutterd, C. A., Dobyns, W. B., Jansen, A., Mirzaa, G., and Leventer, R. J. (2020). Polymicrogyria Overview 1. Definition and Clinical Characteristics of Polymicrogyria. Seattle, WA: University of Washington.
Stutterd, C. A., and Leventer, R. J. (2014). Polymicrogyria: A common and heterogeneous malformation of cortical development. Am. J. Med. Genet. C Semin Med. Genet. 166C, 227–239. doi: 10.1002/ajmg.c.31399
Subramanian, L., Bershteyn, M., Paredes, M. F., and Kriegstein, A. R. (2017). Dynamic behaviour of human neuroepithelial cells in the developing forebrain. Nat. Commun. 8:14167. doi: 10.1038/ncomms14167
Sun, A. X., Yuan, Q., Fukuda, M., Yu, W., Yan, H., Lim, G. G. Y., et al. (2019). Potassium channel dysfunction in human neuronal models of Angelman syndrome. Science 366, 1486–1492. doi: 10.1126/science.aav5386
Sun, N., Meng, X., Liu, Y., Song, D., Jiang, C., and Cai, J. (2021). Applications of brain organoids in neurodevelopment and neurological diseases. J. Biomed. Sci. 28:30. doi: 10.1186/s12929-021-00728-4
Sun, T., and Hevner, R. F. (2014). Growth and folding of the mammalian cerebral cortex: From molecules to malformations. Nat. Rev. Neurosci. 15, 217–232. doi: 10.1038/nrn3707
Szaba, F. M., Tighe, M., Kummer, L. W., Lanzer, K. G., Ward, J. M., Lanthier, P., et al. (2018). Zika virus infection in immunocompetent pregnant mice causes fetal damage and placental pathology in the absence of fetal infection. PLoS Pathog. 14:e1006994. doi: 10.1371/journal.ppat.1006994
Tokuda, S., Mahaffey, C. L., Monks, B., Faulkner, C. R., Birnbaum, M. J., Danzer, S. C., et al. (2011). A novel Akt3 mutation associated with enhanced kinase activity and seizure susceptibility in mice. Hum. Mol. Genet. 20, 988–999. doi: 10.1093/hmg/ddq544
Tsai, P., and Sahin, M. (2011). Mechanisms of neurocognitive dysfunction and therapeutic considerations in tuberous sclerosis complex. Curr. Opin. Neurol. 24, 106–113. doi: 10.1097/WCO.0b013e32834451c4
Uzquiano, A., Cifuentes-Diaz, C., Jabali, A., Romero, D. M., Houllier, A., Dingli, F., et al. (2019). Mutations in the Heterotopia Gene Eml1/EML1 Severely Disrupt the Formation of Primary Cilia. Cell Rep. 28, 1596.e–1611.e. doi: 10.1016/j.celrep.2019.06.096
Vadodaria, K. C., Ji, Y., Skime, M., Paquola, A. C., Nelson, T., Hall-Flavin, D., et al. (2019). Altered serotonergic circuitry in SSRI-resistant major depressive disorder patient-derived neurons. Mol. Psychiatry 24, 808–818. doi: 10.1038/s41380-019-0377-5
Vadodaria, K. C., Mertens, J., Paquola, A., Bardy, C., Li, X., Jappelli, R., et al. (2016). Generation of functional human serotonergic neurons from fibroblasts. Mol. Psychiatry 21, 49–61. doi: 10.1038/mp.2015.161
Vanderver, A., Tonduti, D., Kahn, I., Schmidt, J., Medne, L., Vento, J., et al. (2014). Characteristic brain magnetic resonance imaging pattern in patients with macrocephaly and PTEN mutations. Am. J. Med. Genet. A 164A, 627–633. doi: 10.1002/ajmg.a.36309
Velasco, S., Kedaigle, A. J., Simmons, S. K., Nash, A., Rocha, M., Quadrato, G., et al. (2019). Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 570, 523–527. doi: 10.1038/s41586-019-1289-x
Vergara, M. N., and Canto-Soler, M. V. (2012). Rediscovering the chick embryo as a model to study retinal development. Neural Dev. 7:22. doi: 10.1186/1749-8104-7-22
Vermunt, M. W., Tan, S. C., Castelijns, B., Geeven, G., Reinink, P., de Bruijn, E., et al. (2016). Epigenomic annotation of gene regulatory alterations during evolution of the primate brain. Nat. Neurosci. 19, 494–503. doi: 10.1038/nn.4229
Viggiano, D., Vallone, D., Welzl, H., and Sadile, A. G. (2002). The Naples High- and Low-Excitability rats: Selective breeding, behavioral profile, morphometry, and molecular biology of the mesocortical dopamine system. Behav. Genet. 32, 315–333. doi: 10.1023/A:1020210221156
Villalba, A., Götz, M., and Borrell, V. (2021). The regulation of cortical neurogenesis. Curr. Top Dev. Biol. 142, 1–66. doi: 10.1016/bs.ctdb.2020.10.003
Voineagu, I., Wang, X., Johnston, P., Lowe, J. K., Tian, Y., Horvath, S., et al. (2011). Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature 474, 380–384. doi: 10.1038/nature10110
Wang, X., Tsai, J.-W., LaMonica, B., and Kriegstein, A. R. (2011). A new subtype of progenitor cell in the mouse embryonic neocortex. Nat. Neurosci. 14, 555–561. doi: 10.1038/nn.2807
Watanabe, M., Buth, J. E., Vishlaghi, N., de la Torre-Ubieta, L., Taxidis, J., Khakh, B. S., et al. (2017). Self-Organized Cerebral Organoids with Human-Specific Features Predict Effective Drugs to Combat Zika Virus Infection. Cell Rep. 21, 517–532. doi: 10.1016/j.celrep.2017.09.047
Watrin, F., Manent, J.-B., Cardoso, C., and Represa, A. (2015). Causes and consequences of gray matter heterotopia. CNS Neurosci. Ther. 21, 112–122. doi: 10.1111/cns.12322
Winden, K. D., Sundberg, M., Yang, C., Wafa, S. M. A., Dwyer, S., Chen, P.-F., et al. (2019). Biallelic Mutations in TSC2 Lead to Abnormalities Associated with Cortical Tubers in Human iPSC-Derived Neurons. J. Neurosci. 39, 9294–9305. doi: 10.1523/JNEUROSCI.0642-19.2019
Won, H., Huang, J., Opland, C. K., Hartl, C. L., and Geschwind, D. H. (2019). Human evolved regulatory elements modulate genes involved in cortical expansion and neurodevelopmental disease susceptibility. Nat. Commun. 10:2396. doi: 10.1038/s41467-019-10248-3
Woodbury-Smith, M., Deneault, E., Yuen, R. K. C., Walker, S., Zarrei, M., Pellecchia, G., et al. (2017). Mutations in RAB39B in individuals with intellectual disability, autism spectrum disorder, and macrocephaly. Mol. Autism 8:59. doi: 10.1186/s13229-017-0175-3
Wray, S. (2021). Modelling neurodegenerative disease using brain organoids. Semin. Cell Dev. Biol. 111, 60–66. doi: 10.1016/j.semcdb.2020.05.012
Xiang, Y., Tanaka, Y., Cakir, B., Patterson, B., Kim, K.-Y., Sun, P., et al. (2019). hESC-Derived Thalamic Organoids Form Reciprocal Projections When Fused with Cortical Organoids. Cell Stem Cell 24, 487–497.e7. doi: 10.1016/j.stem.2018.12.015
Xiang, Y., Tanaka, Y., Patterson, B., Hwang, S.-M., Hysolli, E., Cakir, B., et al. (2020). Dysregulation of BRD4 Function Underlies the Functional Abnormalities of MeCP2 Mutant Neurons. Mol. Cell 79, 84–98.e9. doi: 10.1016/j.molcel.2020.05.016
Xiang, Y., Tanaka, Y., Patterson, B., Kang, Y.-J., Govindaiah, G., Roselaar, N., et al. (2017). Fusion of Regionally Specified hPSC-Derived Organoids Models Human Brain Development and Interneuron Migration. Cell Stem Cell 21, 383–398.e7. doi: 10.1016/j.stem.2017.07.007
Yang, N., Chanda, S., Marro, S., Ng, Y.-H., Janas, J. A., Haag, D., et al. (2017). Generation of pure GABAergic neurons by transcription factor programming. Nat. Methods 14, 621–628. doi: 10.1038/nmeth.4291
Yeung, K. S., Tso, W. W. Y., Ip, J. J. K., Mak, C. C. Y., Leung, G. K. C., Tsang, M. H. Y., et al. (2017). Identification of mutations in the PI3K-AKT-mTOR signalling pathway in patients with macrocephaly and developmental delay and/or autism. Mol. Autism 8:66. doi: 10.1186/s13229-017-0182-4
Yin, J., and Schaaf, C. P. (2017). Autism genetics – an overview. Prenat. Diagn. 37, 14–30. doi: 10.1002/pd.4942
Youn, Y. H., Pramparo, T., Hirotsune, S., and Wynshaw-Boris, A. (2009). Distinct dose-dependent cortical neuronal migration and neurite extension defects in Lis1 and Ndel1 mutant mice. J. Neurosci. 29, 15520–15530. doi: 10.1523/JNEUROSCI.4630-09.2009
Yu, D. X., Di Giorgio, F. P., Yao, J., Marchetto, M. C., Brennand, K., Wright, R., et al. (2014). Modeling hippocampal neurogenesis using human pluripotent stem cells. Stem Cell Rep. 2, 295–310. doi: 10.1016/j.stemcr.2014.01.009
Yu, T. W., Mochida, G. H., Tischfield, D. J., Sgaier, S. K., Flores-Sarnat, L., Sergi, C. M., et al. (2010). Mutations in WDR62, encoding a centrosome-associated protein, cause microcephaly with simplified gyri and abnormal cortical architecture. Nat. Genet. 42, 1015–1020. doi: 10.1038/ng.683
Yue, J., Lu, H., Lan, S., Liu, J., Stein, M. N., Haffty, B. G., et al. (2013). Identification of the DNA repair defects in a case of Dubowitz syndrome. PLoS One 8:e54389. doi: 10.1371/journal.pone.0054389
Zhang, W., Ma, L., Yang, M., Shao, Q., Xu, J., Lu, Z., et al. (2020). Cerebral organoid and mouse models reveal a RAB39b-PI3K-mTOR pathway-dependent dysregulation of cortical development leading to macrocephaly/autism phenotypes. Genes Dev. 34, 580–597. doi: 10.1101/gad.332494.119
Zhang, W., Yang, S.-L., Yang, M., Herrlinger, S., Shao, Q., Collar, J. L., et al. (2019). Modeling microcephaly with cerebral organoids reveals a WDR62–CEP170–KIF2A pathway promoting cilium disassembly in neural progenitors. Nat. Commun. 10:2612. doi: 10.1038/s41467-019-10497-2
Zhao, X., and Bhattacharyya, A. (2018). Human Models Are Needed for Studying Human Neurodevelopmental Disorders. Am. J. Hum. Genet. 103, 829–857. doi: 10.1016/j.ajhg.2018.10.009
Zhou, Z.-W., Tapias, A., Bruhn, C., Gruber, R., Sukchev, M., and Wang, Z.-Q. (2013). DNA damage response in microcephaly development of MCPH1 mouse model. DNA Repair 12, 645–655. doi: 10.1016/j.dnarep.2013.04.017
Zhu, J., Lee, K. P., Spencer, T. J., Biederman, J., and Bhide, P. G. (2014). Transgenerational transmission of hyperactivity in a mouse model of ADHD. J. Neurosci. 34, 2768–2773. doi: 10.1523/JNEUROSCI.4402-13.2014
Zoghbi, H. Y. (2003). Postnatal neurodevelopmental disorders: Meeting at the synapse? Science 302, 826–830. doi: 10.1126/science.1089071
Zosen, D., Hadera, M. G., Lumor, J. S., Andersen, J. M., and Paulsen, R. E. (2021). Chicken embryo as animal model to study drug distribution to the developing brain. J. Pharmacol. Toxicol. Methods 112:107105. doi: 10.1016/j.vascn.2021.107105
Zucco, A. J., Pozzo, V. D., Afinogenova, A., Hart, R. P., Devinsky, O., and D’Arcangelo, G. (2018). Neural progenitors derived from Tuberous Sclerosis Complex patients exhibit attenuated PI3K/AKT signaling and delayed neuronal differentiation. Mol. Cell Neurosci. 92, 149–163. doi: 10.1016/j.mcn.2018.08.004
Keywords: neurodevelopmental disorders, malformations of cortical development, disease modeling, animal models, two-dimensional (2D) human-specific cultures, brain organoids
Citation: Damianidou E, Mouratidou L and Kyrousi C (2022) Research models of neurodevelopmental disorders: The right model in the right place. Front. Neurosci. 16:1031075. doi: 10.3389/fnins.2022.1031075
Received: 29 August 2022; Accepted: 07 October 2022;
Published: 20 October 2022.
Edited by:
Myrto Denaxa, Alexander Fleming Biomedical Sciences Research Center, GreeceReviewed by:
Panagiotis Politis, Biomedical Research Foundation of the Academy of Athens (BRFAA), GreeceWai Kit Chan, University of Edinburgh, United Kingdom
Copyright © 2022 Damianidou, Mouratidou and Kyrousi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christina Kyrousi, Y2t5cm91c2lAbWVkLnVvYS5ncg==
†These authors have contributed equally to this work