 Andrea Ravignani
Andrea Ravignani Bill Thompson
Bill Thompson Piera Filippi
Piera Filippi- 1Department of Language and Cognition, Max Planck Institute for Psycholinguistics, Nijmegen, Netherlands
- 2Artificial Intelligence Lab, Vrije Universiteit Brussel, Brussels, Belgium
- 3Research Department, Sealcentre Pieterburen, Pieterburen, Netherlands
- 4Institute of Language, Communication and the Brain, Aix-en-Provence, France
- 5Laboratoire Parole et Langage LPL UMR 7309, Centre National de la Recherche Scientifique, Aix-Marseille Université, Aix-en-Provence, France
- 6Laboratoire de Psychologie Cognitive LPC UMR7290, Centre National de la Recherche Scientifique, Aix-Marseille Université, Marseille, France
Language and music share many commonalities, both as natural phenomena and as subjects of intellectual inquiry. Rather than exhaustively reviewing these connections, we focus on potential cross-pollination of methodological inquiries and attitudes. We highlight areas in which scholarship on the evolution of language may inform the evolution of music. We focus on the value of coupled empirical and formal methodologies, and on the futility of mysterianism, the declining view that the nature, origins and evolution of language cannot be addressed empirically. We identify key areas in which the evolution of language as a discipline has flourished historically, and suggest ways in which these advances can be integrated into the study of the evolution of music.
Introduction
Language and music are typical human behaviors, absent in our closest living animal relatives. Since behavior and cognition do not fossilize, the earliest stages of language and music in our ancestors can only be reconstructed indirectly. Here, we argue for the value of taking into account the evolution of language when studying the evolution of music. There are multiple reasons for this. First, from a meta-scientific perspective, the empirical investigation of language evolution predates research on the evolution of music. Second, methodologically, the fields of music and language evolution show several commonalities (Table 1). For instance, in both fields, corpus-based research is complemented by laboratory-based psychological testing, electrophysiology and neuroimaging studies, and comparative experiments on animals. Third, many hypotheses concerning the origins of music also involve language, and vice-versa. Studying language and music within a common framework provides key insights and testable hypotheses in both disciplines. Here, we argue that anti-empiricist views on language have detrimental effects on understanding its origins and evolution. We identify equivalent anti-scientific tendencies in musicology. We suggest a few ways to counteract such effects that have been established for linguistics, proposing that similar approaches should be adopted in musicology.
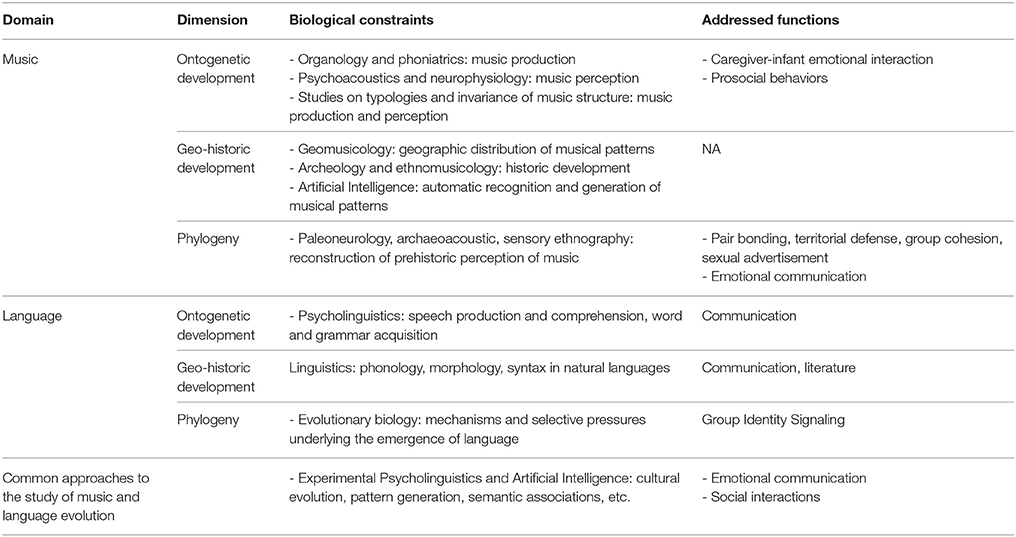
Table 1. Disciplines which can contribute to understanding the evolution of musicality, and their correspondence with affine disciplines in the evolution of language.
Language is defined here as the ability to produce and understand verbal units within interactional communication acts. A major issue in musicology is finding an operational definition of music. Cross (2003, p.79) defines music as “embodying, entraining and transposably intentionalising time in sound and action.” Following Honing et al. (2015), we distinguish the notions of “musicality”—a set of traits which evolve as constrained by our cognitive and biological system that shape musical behaviors across cultures - and “music”—a socio-cultural artifact building on the biological inclination for musicality.
Language and Music: Differences and Commonalities
In language, meaningless phonemes are concatenated into larger discrete units, such as morphemes and words, in accordance with phonological and morpho-syntactic rules. These units are arbitrarily linked to meanings and conceptual representations, which may be culturally transmitted (though sound-meaning mapping in language is not always arbitrary: Monaghan et al., 2012; Parise and Spence, 2012; Imai and Kita, 2014; Dingemanse et al., 2015, 2016). In contrast, while musical tones can be concatenated into phrases or melodies according to structural rules, they are not usually arbitrarily linked to external meanings. Furthermore, while musical melodies are typically made of discrete pitches at fixed interval scales and tonal centers (in case of tonal music), spoken language involves continuous pitch rise and falls (Jackendoff, 2009).
Despite these differences, language and music share several cognitive underpinnings. A number of studies have identified common cognitive mechanisms involved in the production and perception of structural relations in both instrumental music and propositional morpho-syntax (Fedorenko et al., 2009; Patel, 2010). For instance, timing principles are used in both language and music (Ravignani et al., 2017), where longer and louder units tend to be perceived as stressed, and changes in pitch modulation orient perception of boundaries between stressed and unstressed units (Cutler et al., 1997; Curtin et al., 2005). In addition, empirical evidence from brain imaging research indicates that amusic participants show deficits in fine-grained perception of pitch (Peretz and Hyde, 2003); patients fail to distinguish a question from a statement solely on the basis of changes in pitch direction (Patel et al., 2008; Liu et al., 2010). This observed difficulty in a sample of amusic patients supports the hypothesis that music and speech intonation share specific neural resources for processing pitch patterns (but see Ayotte et al., 2002). Further brain imaging studies report a considerable overlap in the brain areas involved in the perception of pitch and rhythm patterns in words and songs (Zatorre et al., 2002; Merill et al., 2012), and in sound pattern processing in melodies and linguistic phrases (Brown et al., 2006). In adults and children, musical training facilitates syllabic and pitch processing in language (Schön et al., 2004; Besson et al., 2007).
Similarly, in both music and verbal language, emotions are expressed through similar patterns of pitch, tempo and intensity (Scherer, 1995; Juslin and Laukka, 2003; Bowling et al., 2012). For instance, in both channels, happiness is expressed by fast speech rate/tempo, medium-high voice intensity/sound level, medium-high frequency energy, high fundamental frequency (F0)/pitch level, high F0/pitch variability, rising F0/pitch contour, fast voice onsets/tone attacks (Juslin and Laukka, 2003). Importantly, the use of voice modulation to express emotional information within interpersonal communication might have had adaptive value in the early species of our genus Homo, improving their ability to respond appropriately to survival opportunities (Mithen, 2005; Filippi, 2016; Frijda, 2016; Filippi et al., 2017a,b).
Finally, in addition to potentially shared cognitive foundations, music and language share the feature of being socially learned from the behavioral outputs of other individuals. It is well-established that the process of transmission from one-generation to another is an important part of the cultural evolutionary process that shapes languages (Kirby et al., 2014). It is becoming clear that cultural evolutionary processes may play a similar role in shaping music (Cross, 2001, 2009; Reybrouck, 2013; Ravignani et al., 2016a; Fitch, 2017a; Ravignani and Verhoef, 2018). Cultural evolution has emerged as a unifying framework in the language sciences (Christiansen and Chater, 2008), linking the cognitive bases for language with the diversity of languages observable throughout the world. A similar approach in the study of the evolution music is desirable (Trehub, 2015).
Research on the Evolution of Musicality: Pitfalls to Avoid
In the last decades, researchers from multiple disciplines have joined forces to unveil the origins and evolution of language. Despite patent progress in methodologies and applications (as attested by highly influential publications), a group of scientists, linguists and philosophers have strongly criticized the whole field of language evolution. This is particularly interesting because the “attack” has been led by one of the fathers of modern linguistics and cognitive science, Noam Chomsky.
A central theme of Chomsky's critiques to the empirical study of language is mysterianism: the idea that scientific knowledge is not always attainable (Chomsky, 2009, 2015; Piattelli-Palmarini et al., 2009). This would be due to the architecture of our minds imposing hard limits on what we can discover scientifically. In other words, Chomsky argues that the limits of human cognition hinder our capacity to unveil scientific mysteries through scientific investigation (Ravignani and Thompson, 2017). This perspective is not new, and had been already proposed in the cognitive sciences in general (McGinn, 1989).
Chomsky makes a distinction between I-language and E-language (Chomsky, 2015). I-language refers to internal linguistic representations, a universal language of thought also known as universal grammar, which, according to Chomsky, is innate. In his view, I-language has a dedicated brain area and is the only aspect of language worth studying, since it is not subject to variation through time and cultures. From his perspective, the ease with which infants acquire any of the hugely diverse set of grammatical structures observable across natural languages points to the existence of this innate universal grammar underlying language acquisition. Instead, E-language encompasses the multiple languages, i.e., the strings of sounds uttered in the outside world, which vary across individuals and cultures. Chomsky claims that processes underlying development and evolution of the wide variety of E-language instances cannot be investigated empirically. In fact, his ideas of mysterianism applied to language bring Chomsky to a simple conclusion: the most important questions about language (his I-language), including its cultural nature, its origins and development over time, and its acquisition are potentially unanswerable (Chomsky, 2015; Ravignani and Thompson, 2017). Notably, the I-language vs. E-languages distinction parallels that between musicality, the human cognitive-biological innate predispositions underlying music perception and production vs. music, intended as a cultural product (Honing et al., 2015; Honing, 2017).
Chomsky has a strong influence on debates about language evolution, and there are clear parallels between the fields of music and language evolution. Here we suggest in the strongest possible terms that research on the evolution of music should avoid Chomskyan mysterianism. By definition this perspective is scientifically stagnant, and the theoretical commitments driving Chomsky's mysterianism are widely rejected in the language sciences at large (Boeckx and Theofanopoulou, 2015; Corballis, 2017; Fitch, 2017b; Kirby, 2017; Ravignani and Thompson, 2017). Other schools of thought show some commonalities to mysterianism (McClelland, 2013; Coleman, 2016).
Another potential attack to the approach we propose comes from a field far away from Chomskyan thought. In particular, generations of ethnomusicologists and cultural anthropologists have strongly opposed the very idea of investigating evolutionary and cross-cultural features of music (Vandor, 1980; Nettl, 2005; Nattiez, 2012). These scholars often object that musical cultures from different parts of the world cannot be compared because comparison would occur through the eyes of a scholar bound to a specific culture. They argue that, across cultures, the word “music” maps to different meanings, and likewise what we call music in the Western world can be translated in many different ways across cultures and languages. According to most of these scholars, the concept of “universal”—corresponding to a feature which is found more often than not, above chance, across cultures, e.g., music often entailing percussion instruments—is pointless because it conflicts with cultural specificity. More generally, several scholars argue there is an irreconcilable divide between humanities and sciences (Gourlay, 1984; Cohen, 2001).
Both Chomskyan and anthropological schools of thought, while departing from opposite philosophical stances, reach the same theoretical conclusion: the nature of language and music is mostly unknown, and empirical efforts to unveil it are pointless. We strongly disagree with this conclusion. Even a cursory glance at the contemporary literature on language and cognition reveals astounding progress in what we know about these topics (Fitch, 2017c). Here we simply reiterate that this progress mostly results from broad contemporary adoption of the scientific method. Our view is that music-related disciplines can benefit equally from rejection of mysterianist skepticism and continued adoption of an integrated experimental approach, namely: (1) observing behaviors and the environment in which they occur, (2) formulating a hypothesis, (3) testing the hypothesis by performing an experiment or collecting new data, (4) using the results to build a model of the phenomenon of interest, (5) employing the model to generate a more refined hypothesis to be tested empirically.
Against the Chomskyan and anthropological perspectives above, we argue for an empirical approach to the origin and evolution of music and musicality. The new-born discipline of music evolution will benefit—and has already benefitted—by unifying the following approaches into one research framework: (1) empirical investigations, as opposed to armchair speculation, on ontogenetic and phylogenetic evolution of music; (2) comparative research, addressed by probing for presence of proto-musical behaviors in other animal species; (3) cross-cultural work, recognizing the diversity of world musical behaviors while comparing them to find common patterns; (4) proposing alternatives to the classical nature-nurture dichotomy. Below we discuss these four points succinctly.
Replacing Armchair Speculation with Models
First, centuries of scientific practice have shown that tightly integrating theory and empirics typically leads to scientific progress. The study of the evolution of music or language is not an exception to this. Theoretical frameworks should provide testable empirical questions (Iversen, 2016), insights for good experimental design and conceptual frameworks to interpret statistical results and generate new testable hypotheses. Ideally, theoretical contributions should be formulated as mathematical models and computer simulations. One advantage of modeling, as compared to constraint-free theorizing, is modeling's potential for falsifiability: Models rely on assumptions to make predictions, which can be empirically falsified. Another related advantage of models is their potential integration with experiments: model assumptions and predictions can be promptly translated into experimental constraints, in turn testable on humans or other living organisms. For these reasons, we strongly support the use of quantitative models of cognitive (Perfors et al., 2011; Fitch, 2014), cultural (Tamariz and Kirby, 2016), and evolutionary (Thompson et al., 2016) processes in music evolution research.
Comparative Cognition Can Inform Human Evolution
Second, the comparative approach to animal cognition can be useful in reconstructing the evolution of human behaviors. Proto-musical behaviors may emerge in other species by (1) homology, i.e. our last common ancestor with that species was endowed with a predisposition toward the behavior under scrutiny, or (2) convergent evolution, i.e., similar evolutionary pressures gave rise to similar genetic predisposition for proto-musical behaviors in humans and other species (Fuhrmann et al., 2014; Ravignani et al., 2014, 2016b; Wilson and Cook, 2016). For instance, recent studies found evidence for beat perception and production, relative pitch and tonal encoding (Hoeschele et al., 2015; Hoeschele and Bowling, 2016), octave generalization (Crickmore, 2003), and consonance (Cook and Fujisawa, 2006) in animals. Based on theoretically driven empirical research (Honing et al., 2015), we argue that, if musical tasks designed for humans are adapted - by modifying their form, not substance—to the specific species under inquiry, many “unthinkable tasks”—sensu Chomsky (2015)—may become manageable. In contrast, based on purely theoretical introspection, Chomsky argues that the cognitive differences between humans and other animals are a matter of quality, not quantity. For instance, he claims that “rats cannot deal with a prime number maze” (Chomsky, 2015; p. 105). To this, we respond that cicadas show behavioral patterns based on prime numbers (Grant, 2005; Tanaka et al., 2009); hence it is tenable that these insects could solve a cicada-adapted prime maze, say over an evolutionary timescale. Likewise, if precursors to music are studied across species with an open-minded attitude, suggestive parallels can be found across humans and other species.
Cross-Cultural Comparisons
Third, an equally important sort of comparative approach consists in cross-cultural research on music. Acceptance of cross-cultural work exhibits one of the starkest contrasts between the study of language and that of music. Cross-cultural comparison of languages has proceeded unhindered for centuries, with few roadblocks. The comparative study of music, instead, experienced a golden period before a crashing halt in the 1960s. Music research can learn from evolutionary linguistics, by performing more cross-cultural work. Its purpose is to account for the uniformity and diversity of musical forms across cultures, in turn to find patterns of music and musicality which are truly generalizable to mankind (the so called universals; Savage et al., 2015).
Beyond the Nature-Nurture Dichotomy
Fourth, language research has been historically characterized by an overreliance on the nature-nurture dichotomy. In our view, the evolution of language as a scientific endeavor has long been plagued by a spurious dichotomy that need not be imported into the evolution of music (Fitch, 2011). We see this opposition as reflecting unnecessarily strong but historically entrenched theoretical divisions. On the one hand, generative linguists in the Chomskyan school have traditionally sought an exclusively mind-internal explanation for the complex tapestry of structures and operations observable in languages (although Chomsky's contemporary mysterianism questions the future of this endeavor). On the other hand, empiricists have privileged the mechanisms of human interaction and cultural transmission as explanations for the diversity and complexity of languages. These opposing standpoints have fuelled much debate but little consensus (e.g., Carruthers et al., 2005).
Contemporary nativist-empiricist dialogues in the cognitive sciences at large now focus on developing paradigms that generalize this dichotomy into a continuous range of possibilities, with traditional nativism and empiricism at the extremes. One example of such a paradigm is Bayesian cognitive modeling, in which bio-cognitive constraints and empirical learning are integrated by a theory of subjective inference based on the principles of conditional probability. The Bayesian approach provides a framework in which both empirical evidence and innate (or earlier learned) biases can be expressed as influences on how individuals learn. While earlier approaches have treated these influences as distinct alternatives, the Bayesian approach allows the two to be balanced in a formally explicit way. Several scholars have argued that approaches like this, in which any flavor of theory can be formally instantiated, interrogated, and tested against empirical data, are the future of nativist-empiricist dialogues (e.g., Spelke and Kinzler, 2009). We hope this sort of integrative approach can be imported into the evolution of music (Trehub, 2015; Jacoby and McDermott, 2017; Ravignani et al., accepted). For example, the generative program applied to music (Lerdahl and Jackendoff, 1985; Rohrmeier, 2011) has generated a range of predictions which have been tested empirically (e.g., Koelsch et al., 2013).
In the language sciences, a new generation of researchers is pushing this approach, integrating models and empirics, even further toward larger-scale evolutionary questions (Blasi et al., 2017). This effort focuses on process-based explanations for behavior, allowing us to understand our species a unique interaction of bio-cognitive and cultural processes (Figure 1) via evolutionary modeling (Smith and Kirby, 2008). Modeling the evolution of minds as part of cultural systems enables us to formalize theories concerning optimal divisions of labor between specialized individual minds and the cultural processes that connect them (Thompson et al., 2016; de Boer and Thompson, 2018). Free from theoretical commitments to one evolutionary process dominating another, contemporary language scientists have at their explanatory disposal an evolutionary framework more powerful than exclusively biological or cultural explanations for behavior: co-evolution, in which evolving minds shape new behaviors, and evolving behaviors shape new minds. Like the Bayesian paradigm, co-evolutionary approaches to the origins of our abilities allow us to move past the idea that only biology or only culture is relevant to unique human behaviors, in a formally explicit way. Hence, modern approaches to the study of language evolution (e.g. Kirby et al., 2014; Thompson et al., 2016) show how biological and cognitive theory can be used to develop process-based, experimentally testable hypotheses about the emergence of behavior among culturally interacting individual minds (Trehub, 2015; Ravignani et al., accepted).
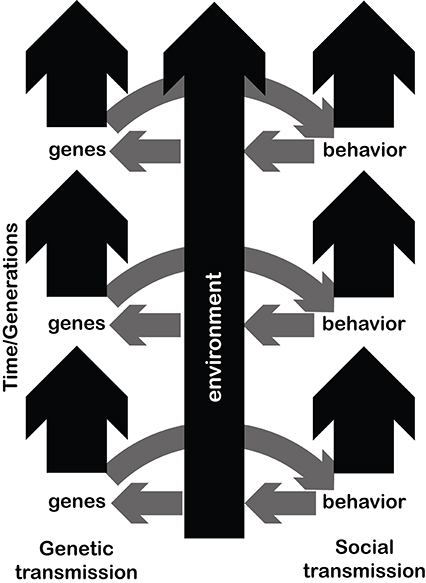
Figure 1. The arrows depict the potential three-way interaction between genes, environment and musical behaviors. Figure inspired by Deacon (1998). Over time, both genes (left) and behaviors (right) change and are transmitted. The environment might change stochastically, or might be directionally altered by behaviors of the species populating it. For a given generation or time period, genes affect behavioral patterns. Behavioral patterns also adapt to, and modify, their environmental medium. In turn, behaviorally-driven changes in the environment might affect the fitness landscape of a species, influencing in turn which genes will be passed on to the next generation.
Conclusions
We hope the evolution of music can hit the ground running under by adopting the inclusive approach described above (see Bowling et al., 2017). The main hypothesis under scrutiny is: does our species unique blend of biological and cultural features underpin remarkable human behaviors like language and music (Figure 1)?
The distinction between music and musicality provides a practical advantage in designing and interpreting experiments. Still, empirical research on the origins of music should adopt a hybrid approach, complementing experiments in tightly controlled settings—hence targeting only music or musicality—with research which integrates the two domains and explanatory levels.
Above we only describe the importance of one possible flow of ideas, from language to music. However, the inverse is also needed: More linguists would need to learn about music evolution and cognition research. For instance, phoneticians and phonologists should capitalize on music findings when investigating tone, prosody, rhythm, etc. (Brown, 2017).
In sum, recent debates provide a bird's-eye view of how the science of language has historically developed, and partly branched into the stormy study of language evolution. Scientists addressing music can benefit from historical breakthroughs and dead-ends in the study of language evolution, and use these insights to accelerate discovery in one of the most exciting topics in contemporary cognitive science, the evolution of music.
Author Contributions
All authors listed have made substantial, direct, and intellectual contributions to the work, and approved it for publication.
Funding
AR was supported by funding from the European Union's Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No. 665501 with the research Foundation Flanders (FWO) (Pegasus2 Marie Curie fellowship 12N5517N awarded to AR) and ERC grant 283435 ABACUS (awarded to Bart de Boer). PF was supported by grants ANR-16-CONV-0002 (ILCB), ANR-11-LABX-0036 (BLRI) and the Excellence Initiative of Aix-Marseille University (A*MIDEX). All authors were supported by a visiting fellowship in Language Evolution from the Max Planck Society.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ayotte, J., Peretz, I., and Hyde, K. (2002). Congenital amusia: a group study of adults afflicted with a music-specific disorder. Brain 125, 238–251. doi: 10.1093/brain/awf028
Besson, M., Schön, D., Moreno, S., Santos, A., and Magne, C. (2007). Influence of musical expertise and musical training on pitch processing in music and language. Restor. Neurol. Neurosci. 25, 399–410.
Blasi, D. E., Michaelis, S. M., and Haspelmath, M. (2017). Grammars are robustly transmitted even during the emergence of creole languages. Nat. Hum. Behav. 1, 723–729. doi: 10.1038/s41562-017-0192-4
Boeckx, C., and Theofanopoulou, C. (2015). Commentary on: Labels, cognomes, and cyclic computation: an ethological perspective. Front. Psychol. 6:784. doi: 10.3389/fpsyg.2015.00784
Bowling, D. L., Hoeschele, M., Gill, K. Z., and Fitch, W. T. (2017). The nature and nurture of musical consonance. Music Percept. 35, 118–121. doi: 10.1525/mp.2017.35.1.118
Bowling, D. L., Sundararajan, J., Han, S., and Purves, D. (2012). Expression of emotion in eastern and western music mirrors vocalization. PLoS ONE 7:e31942. doi: 10.1371/journal.pone.0031942
Brown, S., Martinez, M. J., and Parsons, L. M. (2006). Music and language side by side in the brain: a PET study of the generation of melodies and sentences. Eur. J. Neurosci. 23, 2791–2803. doi: 10.1111/j.1460-9568.2006.04785.x
Carruthers, P., Laurence, S., and Stich, S. (eds.). (2005). The Innate Mind: Structure and Contents, Vol. 1. New York, NY: Oxford University Press.
Chomsky, N. (2009). The mysteries of nature: how deeply hidden? J. Philos. 106, 167–200. doi: 10.5840/jphil2009106416
Christiansen, M., and Chater, N. (2008). Language as shaped by the brain. Behav. Brain Sci. 1, 489–508. doi: 10.1017/S0140525X08004998
Cohen, B. R. (2001). Science and humanities: across two cultures and into science studies. Endeavour 25, 8–12. doi: 10.1016/S0160-9327(00)01335-1
Coleman, S. (2016). “Panpsychism and neutral monism: how to make up one's mind,” in Panpsychism: Contemporary Perspectives. Philosophy of Mind Series, eds G. Bruntrup and L. Jaskolla (New York, NY: OUP), 249–282.
Cook, N. D., and Fujisawa, T. X. (2006). The psychophysics of harmony perception: harmony is a three-tone phenomenon. Empirical Musicol. Rev. 1, 106–126. doi: 10.18061/1811/24080
Corballis, M. C. (2017). Language evolution: a changing perspective. Trends Cogn. Sci. 21, 229–236. doi: 10.1016/j.tics.2017.01.013
Crickmore, L. (2003). A re-valuation of the ancient science of harmonics. Psychol. Music 31, 391–403. doi: 10.1177/03057356030314004
Cross, I. (2001). Music, cognition, culture, and evolution. Ann. N. Y. Acad. Sci. 930, 28–42. doi: 10.1111/j.1749-6632.2001.tb05723.x
Cross, I. (2003). Music and evolution: consequences and causes. Contemp. Music Rev. 22, 79–89. doi: 10.1080/0749446032000150906
Cross, I. (2009). The evolutionary nature of musical meaning. Music. Sci. 13, 179–200. doi: 10.1177/1029864909013002091
Curtin, S., Mintz, T. H., and Christiansen, M. H. (2005). Stress changes the representational landscape: evidence from word segmentation. Cognition 96, 233–262. doi: 10.1016/j.cognition.2004.08.005
Cutler, A., Oahan, D., and Van Donselaar, W. (1997). Prosody in the comprehension of spoken language: a literature review. Lang. Speech 40, 141–201. doi: 10.1177/002383099704000203
Deacon, T. W. (1998). The Symbolic Species: The Co-evolution of Language and the Brain. New York, NY: WW Norton & Company.
de Boer, B., and Thompson, B. (2018). Biology-culture co-evolution in finite populations. Sci. Rep. 8:1209. doi: 10.1038/s41598-017-18928-0
Dingemanse, M., Blasi, D. E., Lupyan, G., Christiansen, M. H., and Monaghan, P. (2015). Arbitrariness, iconicity, and systematicity in language. Trends Cogn. Sci. 19, 603–615. doi: 10.1016/j.tics.2015.07.013
Dingemanse, M., Schuerman, W., Reinisch, E., Tufvesson, S., Mitterer, H., Akita, K., Woodworth, N. L., et al. (2016). What sound symbolism can and cannot do: testing the iconicity of ideophones from five languages. Language 92, e117–e133. doi: 10.1353/lan.2016.0034
Fedorenko, E., Patel, A., Casasanto, D., Winawer, J., and Gibson, E. (2009). Structural integration in language and music: evidence for a shared system. Mem. Cogn. 37, 1–9. doi: 10.3758/MC.37.1.1
Filippi, P. (2016). Emotional and interactional prosody across animal communication systems: a comparative approach to the emergence of language. Front. Psychol. 7:1393. doi: 10.3389/fpsyg.2016.01393
Filippi, P., Congdon, J. V., Hoang, J., Bowling, D. L., Reber, S. A., Pašukonis, A., et al. (2017a). Humans recognize emotional arousal in vocalizations across all classes of terrestrial vertebrates: evidence for acoustic universals. Proc. R. Soc. B. 284:20170990. doi: 10.1098/rspb.2017.0990
Filippi, P., Ocklenburg, S., Bowling, D. L., Heege, L., Güntürkün, O., Newen, A., et al. (2017b). More than words (and faces): evidence for a Stroop effect of prosody in emotion word processing. Cogn. Emotion 31, 879–891. doi: 10.1080/02699931.2016.1177489
Fitch, W. T. (2011). Biological versus cultural evolution: beyond a false dichotomy: comment on “modeling the cultural evolution of language” by Luc Steels. Phys. Life Rev. 8, 357–358. doi: 10.1016/j.plrev.2011.10.020
Fitch, W. T. (2014). Toward a computational framework for cognitive biology: unifying approaches from cognitive neuroscience and comparative cognition. Phys. Life Rev. 11, 329–364. doi: 10.1016/j.plrev.2014.04.005
Fitch, W. T. (2017a). Cultural evolution: lab-cultured musical universals. Nat. Hum. Behav. 1, 1–2. doi: 10.1038/s41562-016-0018
Fitch, W. T. (2017b). On externalization and cognitive continuity in language evolution. Mind Lang. 32, 597–606. doi: 10.1111/mila.12162
Fitch, W. T. (2017c). Empirical approaches to the study of language evolution. Psychon. Bull. Rev. 24, 3–33. doi: 10.3758/s13423-017-1236-5
Frijda, N. H. (2016). The evolutionary emergence of what we call “emotions”. Cogn. Emot. 30, 609–620. doi: 10.1080/02699931.2016.1145106
Fuhrmann, D., Ravignani, A., Marshall-Pescini, S., and Whiten, A. (2014). Synchrony and motor mimicking in chimpanzee observational learning. Sci. Rep. 4:5283. doi: 10.1038/srep05283
Grant, P. R. (2005). The priming of periodical cicada life cycles. Trends Ecol. Evol. 20, 169–174. doi: 10.1016/j.tree.2005.01.016
Hoeschele, M., and Bowling, D. L. (2016). Sex differences in rhythmic preferences in the budgerigar (Melopsittacus undulatus): a comparative study with humans. Front. Psychol. 7:543. doi: 10.3389/fpsyg.2016.01543
Hoeschele, M., Merchant, H., Kikuchi, Y., Hattori, Y., and ten Cate, C. (2015). Searching for the origins of musicality across species. Phil. Trans. R. Soc. B 370:20140094. doi: 10.1098/rstb.2014.0094
Honing, H. (2017). “Musicality as an upbeat to music: introduction and research agenda,” in The Origins of Musicality, ed H. Honing (Cambridge, MA: MIT Press), 3–20.
Honing, H., ten Cate, C., Peretz, I., and Trehub, S. E. (2015). Without it no music: cognition, biology, and evolution of musicality. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:20140088. doi: 10.1098/rstb.2014.0088
Imai, M., and Kita, S. (2014). The sound symbolism bootstrapping hypothesis for language acquisition and language evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369:20130298. doi: 10.1098/rstb.2013.0298
Iversen, J. R. (2016). “In the beginning was the beat: evolutionary origins of musical rhythm in humans,” in The Cambridge Companion to Percussion, ed R. Hartenberger (Cambridge: Cambridge University Press), 281–295.
Jackendoff, R. (2009). Parallels and nonparallels between language and music. Music Percept. 26, 195–204. doi: 10.1525/mp.2009.26.3.195
Jacoby, N., and McDermott, J. H. (2017). Integer ratio priors on musical rhythm revealed cross-culturally by iterated reproduction. Curr. Biol. 27, 359–370. doi: 10.1016/j.cub.2016.12.031
Juslin, P. N., and Laukka, P. (2003). Communication of emotions in vocal expression and music performance: different channels, same code? Psychol. Bull. 129, 770–814. doi: 10.1037/0033-2909.129.5.770
Kirby, S. (2017). Culture and biology in the origins of linguistic structure. Psychon. Bull. Rev. 24, 118–137. doi: 10.3758/s13423-016-1166-7
Kirby, S., Griffiths, T., and Smith, K. (2014). Iterated learning and the evolution of language Introduction: can culture explain structure? Curr. Opin. Neurobiol. 28, 108–114. doi: 10.1016/j.conb.2014.07.014
Koelsch, S., Rohrmeier, M., Torrecuso, R., and Jentschke, S. (2013). Processing of hierarchical syntactic structure in music. Proc. Natl. Acad. Sci. U.S.A. 110, 15443–15448. doi: 10.1073/pnas.1300272110
Lerdahl, F., and Jackendoff, R. (1985). A Generative Theory of Tonal Music. Cambridge, MA: MIT Press.
Liu, F., Patel, A. D., Fourcin, A., and Stewart, L. (2010). Intonation processing in congenital amusia: discrimination, identification and imitation. Brain 133, 1682–1693. doi: 10.1093/brain/awq089
McClelland, T. (2013). The Neo-Russellian ignorance hypothesis. J. Conscious. Stud. 20, 4–125. Available online at: http://www.ingentaconnect.com/content/imp/jcs/2013/00000020/F0020003/art00006?crawler=true
McGinn, C. (1989). Can we solve the mind - body problem? Mind xcviii 391, 349–366. doi: 10.1093/mind/XCVIII.391.349
Merill, J., Sammler, D., Bangert, M., Goldhahn, D., Turner, R., and Friederici, A. D. (2012). Perception of words and pitch patterns in song and speech. Front. Psychol. 3:76. doi: 10.3389/fpsyg.2012.00076
Mithen, S. (2005). The Singing Neanderthals: The Origins of Music, Language, Mind, and Body. Cambridge, MA: Harvard University Press.
Monaghan, P., Mattock, K., and Walker, P. (2012). The role of sound symbolism in language learning. J. Exp. Psychol. Learn. Mem. Cogn. 38, 1152–1164. doi: 10.1037/a0027747
Nattiez, J.-J. (2012). Is the search for universals incompatible with the study of cultural specificity? Hum. Soc. Stud. 1, 67–94. doi: 10.2478/v10317-012-0005-2
Nettl, B. (2005). The Study of Ethnomusicology: Thirty-One Issues and Concepts. Champaign, IL: University of Illinois Press.
Parise, C. V., and Spence, C. (2012). Audiovisual crossmodal correspondences and sound symbolism: a study using the implicit association test. Exp. Brain Res. 220, 319–333. doi: 10.1007/s00221-012-3140-6
Patel, A. D., Wong, M., Foxton, J., Lochy, A., and Peretz, I. (2008). Speech intonation perception deficits in musical tone deafness (congenital amusia). Music Percept. 25, 357–368. doi: 10.1525/mp.2008.25.4.357
Peretz, I., and Hyde, K. L. (2003). What is specific to music processing? insights from congenital amusia. Trends Cogn. Sci. 7, 362–367. doi: 10.1016/S1364-6613(03)00150-5
Perfors, A., Tenenbaum, J. B., Griffiths, T. L., and Xu, F. (2011). A tutorial introduction to Bayesian models of cognitive development. Cognition 120, 302–321. doi: 10.1016/j.cognition.2010.11.015
Piattelli-Palmarini, M., Uriagereka, J., and Salaburu, P. (Eds.). (2009). Of Minds and Language: A Dialogue with Noam Chomsky in the Basque Country. OUP Oxford.
Ravignani, A., and Thompson, B. (2017). NOAM CHOMSKY, What kind of creatures are we? New York, NY: Columbia University Press, 2015. Pp. 200. Hb. $19.95. Lang. Soc. 46, 446–447. doi: 10.1017/S0047404517000288
Ravignani, A., and Verhoef, T. (2018). Which melodic universals emerge from repeated signaling games? Artif. Life 24. doi: 10.7287/peerj.preprints.3366v1
Ravignani, A., Bowling, D. L., and Fitch, W. T. (2014). Chorusing, synchrony and the evolutionary functions of rhythm. Front. Psychol. 5:1118. doi: 10.3389/fpsyg.2014.01118
Ravignani, A., Delgado, T., and Kirby, S. (2016a). Musical evolution in the lab exhibits rhythmic universals. Nat. Hum. Behav. 1:0007. doi: 10.1038/s41562-016-0007
Ravignani, A., Fitch, W. T., Hanke, F. D., Heinrich, T., Hurgitsch, B., Kotz, S. A., et al. (2016b). What pinnipeds have to say about human speech, music, and the evolution of rhythm. Front. Neurosci. 10:274. doi: 10.3389/fnins.2016.00274
Ravignani, A., Honing, H., and Kotz, S. A. (2017). The evolution of rhythm cognition: Timing in music and speech. Front. Hum. Neurosci. 11:308. doi: 10.3389/fnhum.2017.00303
Ravignani, A., Thompson, B., Grossi, T., Delgado, T., and Kirby, S. (accepted). Evolving building blocks of rhythm: How human cognition creates music via cultural transmission. Ann. N.Y. Acad. Sci. doi: 10.1111/nyas.13610
Reybrouck, M. (2013). From sound to music: an evolutionary approach to musical semantics. Biosemiotics 6, 585–606. doi: 10.1007/s12304-013-9192-6
Rohrmeier, M. (2011). Towards a generative syntax of tonal harmony. J. Math. Music 5, 35–53. doi: 10.1080/17459737.2011.573676
Savage, P. E., Brown, S., Sakai, E., and Currie, T. E. (2015). Statistical universals reveal the structures and functions of human music. Proc. Natl. Acad. Sci. U.S.A. 112, 8987–8992. doi: 10.1073/pnas.1414495112
Scherer, K. R. (1995). Expression of emotion in voice and music. J. Voice 9, 235–248. doi: 10.1016/S0892-1997(05)80231-0
Schön, D., Magne, C., and Besson, M. (2004). The music of speech: music training facilitates pitch processing in both music and language. Psychophysiology 41, 341–349. doi: 10.1111/1469-8986.00172.x
Smith, K., and Kirby, S. (2008). Cultural evolution: implications for understanding the human language faculty and its evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3591–3603. doi: 10.1098/rstb.2008.0145
Spelke, E. S., and Kinzler, K. D. (2009). Innateness, learning, and rationality. Child Dev. Perspect. 3, 96–98. doi: 10.1111/j.1750-8606.2009.00085.x
Tamariz, M., and Kirby, S. (2016). The cultural evolution of language. Curr. Opin. Psychol. 8, 37–43. doi: 10.1016/j.copsyc.2015.09.003
Tanaka, Y., Yoshimura, J., Simon, C., Cooley, J. R., and Tainaka, K.-I. (2009). Allele effect in the selection for prime-numbered cycles in periodical cicadas Proc. Natl. Acad. Sci. U.S.A. 106, 8975–8979. doi: 10.1073/pnas.0900215106
Thompson, B., Kirby, S., and Smith, K. (2016). Culture shapes the evolution of cognition. Proc. Natl. Acad. Sci. U.S.A. 113, 4530–4535. doi: 10.1073/pnas.1523631113
Trehub, S. E. (2015). Cross-cultural convergence of musical features. Proc. Natl. Acad. Sci. U.S.A. 112, 8809–8810. doi: 10.1073/pnas.1510724112
Wilson, M., and Cook, P. F. (2016). Rhythmic entrainment: why humans want to, fireflies can't help it, pet birds try, and sea lions have to be bribed. Psychon. Bull. Rev. 23, 1647. doi: 10.3758/s13423-016-1013-x
Keywords: evolution of music, evolution of language, cultural transmission, cultural evolution, universals, music cognition, comparative cognition, nature and nurture
Citation: Ravignani A, Thompson B and Filippi P (2018) The Evolution of Musicality: What Can Be Learned from Language Evolution Research? Front. Neurosci. 12:20. doi: 10.3389/fnins.2018.00020
Received: 15 September 2017; Accepted: 10 January 2018;
Published: 06 February 2018.
Edited by:
Aleksey Nikolsky, Independent Researcher, Los Angeles, CA, United StatesCopyright © 2018 Ravignani, Thompson and Filippi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea Ravignani, YW5kcmVhLnJhdmlnbmFuaUBnbWFpbC5jb20=