



- 1Institute of Cognitive Neuroscience and Institute of Neurology, University College London, London, UK
- 2School of Psychology, Cardiff University, Cardiff, UK
Although executive control and automatic behavior have often been considered separate and distinct processes, there is strong emerging and convergent evidence that they may in fact be intricately interlinked. In this review, we draw together evidence showing that visual stimuli cause automatic and unconscious motor activation, and how this in turn has implications for executive control. We discuss object affordances, alien limb syndrome, the visual grasp reflex, subliminal priming, and subliminal triggering of attentional orienting. Consideration of these findings suggests automatic motor activation might form an intrinsic part of all behavior, rather than being categorically different from voluntary actions.
It is widely believed that human cognition and behavior is governed by both voluntary and automatic processes. Voluntary “executive control” mechanisms are assumed to direct behavior in goal-directed ways through use of explicit knowledge and expectations. On the other hand, accumulating research has revealed that perceptual processing of visual stimuli can automatically and unconsciously modulate motor responses (see e.g., Eimer and Schlaghecken, 2003; Sumner, 2007). Traditionally, the processes underpinning automatic and unconscious triggering of actions have been considered separate from the processes underpinning voluntary action planning and control. Embedded in this concept of separate functional pathways is the idea that automatic processes are unconscious, fast, and rigid whereas voluntary action planning and control were considered to be conscious, and flexible (see e.g., Schneider and Shiffrin, 1977; Shiffrin and Schneider, 1977, 1984). However, several lines of evidence, briefly reviewed previously by Sumner and Husain (2008), challenge this traditional distinction.
In this review we consider recent empirical findings and discuss how they provide evidence that voluntary and automatic control of action might not in fact be so distinct. We suggest that many “automatic” mechanisms can in fact be surprisingly flexible, quite unlike the traditional, inflexible view of these processes. We begin by reviewing evidence that even simple, flashed visual stimuli can automatically modulate on-going motor responses. Then we discuss how automatically primed responses might affect interactions with real objects in the world around us, focusing on the subject of object affordance. Finally we turn to the issue of how such primed activity might be controlled, and to what extent such control could be automatic. Throughout the review, we draw on evidence from three converging approaches: using subliminal priming paradigms to show that unconscious motor activation can also be reversed unconsciously; using traditional “conscious inhibition” paradigms to show that such inhibition can also be triggered automatically; using the two types of paradigm together to see if they interact. The demonstration of flexible control over automatic processes suggests an intricate link between these historically distinct processes.
Evidence for Automatic Activation of Motor Responses
Perceptual processing of a visual stimulus can result in motor responses even when the observer does not intend to act. One of the most well-studied of these phenomena is the “visual grasp reflex”, where an observer makes a fast, reflexive eye movement (saccade) toward a suddenly appearing—and irrelevant—visual stimulus, despite their intention to look elsewhere (e.g., Theeuwes et al., 1998; Irwin et al., 2000). Even when irrelevant distractors do not fully succeed in capturing gaze they may nevertheless have a remarkable influence on on-going motor activity. For example, saccades can curve whilst in flight toward an irrelevant distractor on the way to correctly landing on the target (e.g., McPeek and Keller, 2001; Godijn and Theeuwes, 2002; McPeek et al., 2000, 2003). But as response latencies increase, saccades are more likely to curve away from a distractor (e.g., Walker et al., 2006), revealing an inhibitory mechanism acting to suppress unwanted motor activity toward the irrelevant stimulus (e.g., Sheliga et al., 1995).
Saccades toward targets can also be slowed when an irrelevant distractor is presented simultaneously—or nearly simultaneously—with the target (the saccade distractor effect; e.g., Walker et al., 1995, 2000). Furthermore, transient changes to the scene during saccade planning in simple tasks, reading, or visual search produce a characteristic “dip” in the frequency of saccades made around 90–100 ms after the change (saccadic inhibition effect; e.g., Reingold and Stampe, 1999, 2000, 2002, 2003; Buonocore and McIntosh, 2008; Edelman and Xu, 2009; Bompas and Sumner, 2011). These dips provide highly robust evidence for rapid modulation of on-going motor commands by visual information. Manual reaching responses too are affected by irrelevant non-target stimuli. Like saccades, reaches can be slowed (e.g., Tipper et al., 1997), curve toward (e.g., Tipper et al., 1997) or away from (e.g., Howard and Tipper, 1997) non-target stimuli in flight. These findings suggest that both manual and oculomotor responses can be automatically modulated by irrelevant visual inputs.
Although not often considered in this context, such effects of irrelevant stimuli might in fact be related to a long-established view that simply visually processing an object can automatically evoke action plans appropriate for interacting with it. Gibson (1979) described “affordances” as properties of the environment that automatically prime the observer to act in such a way. According to this view, seeing a coffee cup with its handle to the right affords—or facilitates—a reaching movement with the right hand to grasp the cup. Recently there has been renewed interest in affordances, and their effects have been examined using functional imaging as well as behavioral methods. For example, motor regions of the brain—such as those within the dorsal medial frontal cortex—are activated when observers merely look at a graspable object (e.g., Grèzes and Decety, 2002), even when they do not intend to act. In other tasks that require arbitrary responses to pictures of graspable objects (such as squeezing a trigger to indicate whether the object is man-made), the response is facilitated when it is congruent with the action afforded by the object (e.g., Tucker and Ellis, 1998; see Figures 1A and B for examples of typical affordance stimuli and their effects on response times). Findings such as these suggest that the brain has learnt to associate objects with actions appropriate to “capture” them, and these actions can be (partially) activated by visual processing of the object.
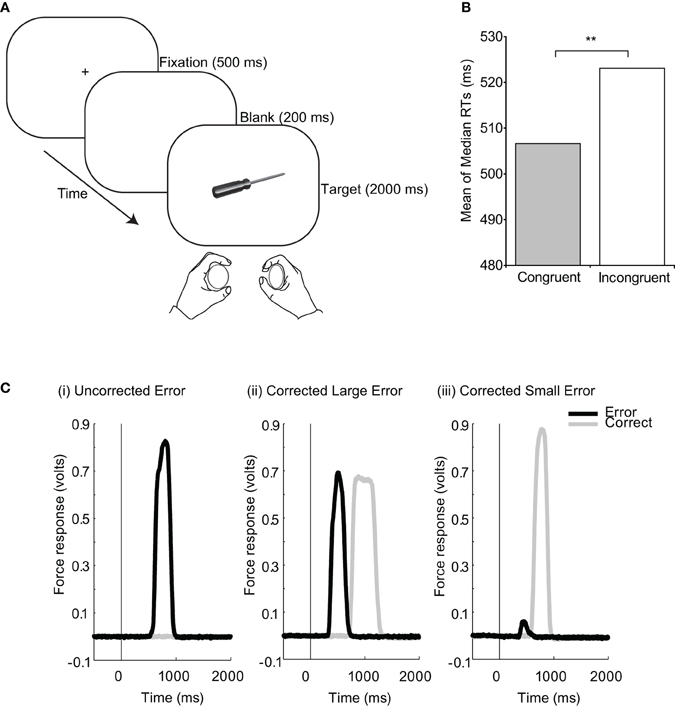
Figure 1. McBride et al. (2012) studied automatic priming of motor responses by visual objects in an object affordance task. Here, participants made speeded squeeze responses with either their left or right hand according to whether the object presented on each trial belonged in a kitchen or a toolbox (see A for task details). The objects could also afford a grasping action with either the left or right hand. Although object affordance was irrelevant to the task, it still modulated responses so that they were faster when the object afforded a congruent response than an incongruent one (B). Furthermore, partial squeeze responses (see C(ii) and C(iii) for examples of the responses recorded on individual trials) were made significantly more often on incongruent than on congruent trials—indicating that the afforded response had been activated and at least partially executed on incongruent trials. These figures are adapted with permission from Taylor and Francis and were originally published in McBride et al. (2012), Q. J. Exp. Psychol. 65, 13–24, www.tandfonline.com.
However, there has been some debate about whether object affordance effects genuinely arise from visual objects automatically eliciting motor plans. Anderson et al. (2002) observed that the most visually salient part of the objects used in many affordance studies were also often the graspable part of the object. They demonstrated that response times were faster whenever the side of the response corresponded with whichever side of the object was most visually salient, even when the object was not graspable (e.g., left hand responses to a picture of an analog clock showing a time of quarter to nine). Therefore, affordance-like behavioral effects do not necessarily arise from possibilities for action per se, but instead can stem from congruence between the required response and this shift of attention.
But this is not to say all affordance effects are perceptual. Object orientation was irrelevant in most other affordance experiments (e.g., Tucker and Ellis, 1998), but by contrast it was the response-defining property of the object in Anderson and colleagues' task. They also used line-drawings of common objects which may have evoked qualitatively different responses than those evoked by the photographs or images of 3D models used elsewhere (e.g., Tucker and Ellis, 1998; Phillips and Ward, 2002; McBride et al., 2012). Furthermore, perceptually lateralized stimuli that do not afford actions do not necessarily produce affordance effects (e.g., Buccino et al., 2009). Finally, object affordance effects have recently been shown with stimuli and responses that are not lateralized, and instead rely on compatibility between object size and response (pinch or power grasp) to produce affordance effects (e.g., Ellis and Tucker, 2000; Tucker and Ellis, 2001; Derbyshire et al., 2006; Ellis et al., 2007). These considerations suggest that there are good reasons to believe perceptual processing of graspable objects can automatically evoke motor responses associated with them.
Alien Hand Syndrome and Utilization Behavior
Some of the most striking evidence that visual objects can indeed automatically generate responses comes from studies of some unusual neurological cases. Patients with alien hand syndrome spontaneously and involuntarily grasp objects—even other people—in their environment as if magnetically drawn to them (for a review see Scepkowski and Cronin-Golumb, 2003). These individuals are aware of their hand making these movements, but report that they are not controlling them, and instead feel the movements are being controlled by an external agent. In a related syndrome, patients who display utilization behavior automatically grasp and use objects placed within their reach, even when the objects are not needed. For example, they may grasp and begin to peel an apple placed within their reach, even though they are not hungry, do not want to eat the apple, and know that it doesn't belong to them (e.g., Boccardi et al., 2002).
Alien limb and utilization behavior are rare neurological syndromes, and so case reports and experimental studies on patients with them have been correspondingly few. Some of the most detailed investigation comes from Riddoch and her colleagues (Riddoch et al., 1998; Humphreys and Riddoch, 2000). They showed that patients with an alien right hand can correctly pick up a cup with the left hand as long as the cup's handle is also on the left. However, when the handle is on the right the patients are more likely to grasp the cup with the right hand, despite instructions to the contrary. These “interference” errors were reduced when the task was to point to the object, rather than grasp, and also when the objects were inverted. Therefore, it seems that these patients responded according to well-learnt affordance-action associations rather than according to the instructions they were given. The action afforded by the object was disrupted when the object was inverted, or when the action required was not the one usually made to the object (pointing instead of grasping), so fewer interference errors were reported under these conditions.
Alien limb and utilization syndromes are most often associated with focal lesions to the medial frontal lobes (particularly the supplementary motor area; SMA e.g., Lhermitte, 1983; Boccardi et al., 2002), but have also been associated with damage to the corpus callosum (e.g., Biran and Chatterjee, 2004), as well as patients with parietal lesions following posterior cerebral artery stroke (e.g., Coulthard et al., 2007; Bartolo et al., 2011). Increasingly, they are recognized in patients with corticobasal degeneration (CBD), a slowly progressive neurodegenerative condition which affects cortical regions as well as the basal ganglia (e.g., Murray et al., 2007; Tiwari and Amar, 2008).
In a recent functional imaging study, Schaefer et al. (2010) examined the neural correlates of unwanted movements in a patient with alien limb syndrome with CBD. They reported that voluntary and alien movements activated similar brain regions, including motor and parietal cortices. However, the right inferior frontal gyrus (rIFG), which has been associated with inhibitory control of motor responses (e.g., Swann et al., 2009, 2012; Hampshire et al., 2010; Verbruggen et al., 2010; see also Aron, 2007), was activated only during alien movements. Such activation may reflect unsuccessful attempts to inhibit alien movements. Taken together, these studies highlight the impact of automatically afforded actions, and suggest that alien limb patients might find them particularly difficult to inhibit.
Evidence for Automatic Motor Activation from “Partial” Errors
Another line of evidence that potentially reveals the automatic effects of visual stimuli on actions comes from investigation of erroneous responses. Typically, most evidence for automatic motor priming by visual objects has been gleaned indirectly by measuring the eventual outcome of this process on reaction times, usually for manual button presses. Such responses are an all-or-nothing, binary measure: either the response made is of sufficient magnitude to trigger a button press, or it isn't. However, it is possible that small amounts of force applied (erroneously) to a button might be insufficient to trigger a measurable response and thereby escape detection.
With this in mind, there has recently been revived interest in employing continuous and sensitive measures of motor response to more directly investigate processes of automatic motor activation on a trial-by-trial basis. McBride et al. (2012) employed such a measure to investigate object affordances. They asked participants to classify object stimuli by squeezing one of two devices placed in their left and right hands (see Figure 1A) while measuring the force applied by either hand. Consistent with object affordance effects, responses were faster on trials where the object afforded an action with the same hand that was required to make the response (congruent trials), compared to the opposite hand (incongruent trials). But continuous, simultaneous force recordings also revealed that participants made small erroneous responses when there was conflict between the response afforded by the object and the response required by the task, i.e., when the stimulus afforded a response that was incongruent to the response required by the task. Such errors were later corrected (Figure 1C). These partial erroneous responses provide compelling evidence that viewing an object activates motor plans appropriate for interacting with that object, sometimes going far enough to produce a partial response.
Electromyography (EMG) has also been used to demonstrate “sub-threshold” erroneous responses on incompatible compared to compatible trials in a variety of paradigms including Eriksen flanker (e.g., Coles et al., 1985; Eriksen et al., 1985), and Simon (e.g., Burle et al., 2002) tasks. For example, continuous measurement of EMG from both arms of participants performing a flanker task has demonstrated that correct button-press responses on incompatible trials are frequently accompanied by some muscle activity in the opposite hand, i.e., for the response associated with the irrelevant flanker stimuli (Eriksen et al., 1985). Thus, response-related muscle activity measured by EMG could be measured in the absence of a “full” button-press response.
Such increased erroneous response activity on incongruent trials provides strong evidence that an irrelevant stimulus—or part of a stimulus—can automatically activate responses associated with it. These responses are not merely partially activated somewhere in the brain; the response can be measured in the muscles or in small hand movements with force transducers. These “partial” responses are not captured by current the models of decision-making, which instead assume that actions are either executed wholly once the threshold for accumulated evidence is reached, or not executed at all (see Smith and Ratcliff, 2004 for a review of commonly used models and their characteristics). These models assume that evidence in favor of particular action possibilities is accumulated until a certain threshold of evidence is reached. Models differ in how evidence accumulates. Some (e.g., random-walk) assume that evidence is accumulated as a single total so that evidence in favor of one response is necessarily evidence counter to alternative responses (e.g., Link and Heath, 1975), whereas others (e.g. accumulator models e.g., Usher and McClelland, 2001) assume that evidence in favor of competing responses is accumulated separately, often with mutual inhibition between the separate accumulators.
Importantly, all these decision models share the assumption that once the accumulated evidence reaches a “threshold,” the response is executed. This all-or-nothing property of decision models does not allow any gradation of the response. Either the evidence accumulating for a particular response reaches the decision threshold and the response is made, or it does not reach threshold and no response is made. We anticipate that investigations of partial responses evoked by automatic activation of motor responses will provide interesting constraints for future work in decision-making.
Invisible Influences
Thus far, in the evidence we have reviewed, the automatic nature of motor priming has been inferred from interference effects: if the participant or patient is engaged in a particular task and stimuli interfere with that task, we infer that responses to the task-irrelevant stimulus (or part of a stimulus) were not volitional. Another way to study automatic influences is to investigate the effects that invisible stimuli have on motor behavior. If an observer is unaware of a stimulus, then traditionally it is concluded that any response made to it cannot have been evoked voluntarily and must, therefore, have been made automatically.
One way to present a stimulus subliminally is to use the backwards masking technique (e.g., Ögmen and Breitmeyer, 2006). Using this technique, participants are usually required to make a manual button press as quickly as possible to a target stimulus (often a left or right pointing arrow). This target is preceded by a “prime” stimulus for a very short duration (say, 20 ms) which is followed by an overlapping (or surrounding in the case of meta-contrast masking) stimulus—or “mask.” This technique renders the prime stimulus imperceptible to the observer. Even when the participants cannot discriminate the identity of the prime under forced choice, responses are generally faster and more accurate when the prime stimulus was associated with the same response as the target stimulus (compatible trial) compared to when the prime was associated with the opposite response (incompatible trial; e.g., Leuthold and Kopp, 1998). Thus, manual responses can be partially activated automatically by visual stimuli even when they cannot be consciously discriminated.
Subliminal stimuli can also prime a shift of attention (e.g., McCormick, 1997; Ivanoff and Klein, 2003; for a review see Mulckhuyse and Theeuwes, 2010) so that observers respond more quickly and accurately to stimuli presented at the cued location relative to an un-cued location. For instance, McCormick (1997) manipulated the luminance of cues so that some were visible and others were not. The cues were mostly invalid so the target was most likely to appear at the opposite location to the cue. When participants perceived the cue they were faster to respond to a target presented at the location opposite the cue, suggesting that they had volitionally moved their attention to the most appropriate (i.e., statistically predicted) location. However, when participants were not aware of the cue they were faster to respond to targets at the cued, relative to un-cued, location. This provides evidence that the invisible cue produced an automatic and involuntary shift of attention to the cued location.
Cues do not have to be based on low-level differences in visual salience in order to produce reliable effects. Socially relevant eye-gaze can also direct attention exogenously. Responses are faster to target stimuli that have been preceded by a non-predictive face (or schematic drawing of a face) with the eyes gazing in the direction of the target (e.g., Friesen and Kingstone, 1998, 2003; Driver et al., 1999; but see Tipples, 2002). These so-called “gaze cueing effects” have been shown following cues that have been backwards-masked to render them invisible to the participant (Sato et al., 2007). In summary, shifts of attention and motor responses can be automatically and unconsciously triggered by visual stimuli. Effects of non-perceived stimuli such as these have provided key evidence that visual stimuli can automatically prime the observer to act.
Inhibition of Primed Actions
In the above section, we have reviewed evidence from parallel domains that visual stimuli can automatically generate actions. These automatically generated actions can interfere with the intended or task-relevant response, and can be triggered by stimuli that are not consciously perceived, potentially facilitating responses to them. But an important question that arises is how such automatically primed responses might be controlled to allow flexible, goal-directed, behavior. For it would not be useful to respond to every object that we see. Thus, it is necessary to consider how brain systems inhibit or override responses that have been triggered automatically by the environment and are not relevant to our current goals.
“Cognitive control” over simultaneously activated competing motor plans has been extensively studied using “conflict” tasks such as such as the Eriksen flanker task described above (see Eriksen and Eriksen, 1974), the Stroop color-word naming task (e.g., Stroop, 1935) and the Simon task (for a review see Lu and Proctor, 1995). In these tasks, multiple responses can be simultaneously activated and in conflict: one response according to the task instruction and one evoked automatically by the irrelevant stimulus (or irrelevant property of the target stimulus). Typical theoretical frameworks for the congruency effects shown in conflict tasks suggest that stimuli are simultaneously processed by two routes which converge at the level of response programming (e.g., dual route model by Kornblum et al., 1990; activation suppression model by Ridderinkhof, 2002; see Figure 2A).
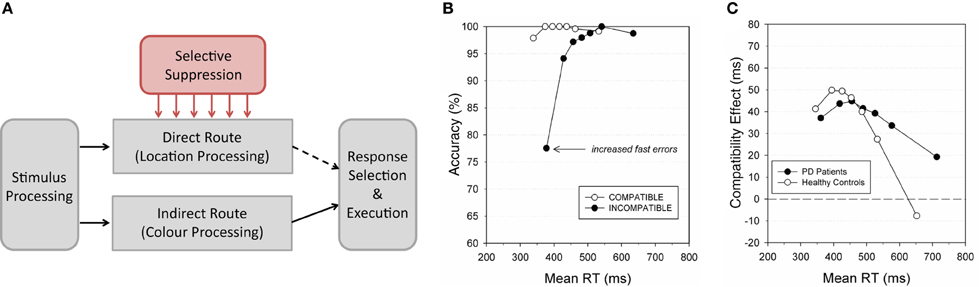
Figure 2. (A) Information processing during a Simon task as modelled by the Activation-Suppression model [e.g., Ridderinkhof (2002)]. The relevant stimulus dimension (color in the Simon task) is processed via the slow, indirect, processing route. Simultaneously, irrelevant stimulus location is processed via the fast, direct, processing route. Activation of the location-based response activation is inhibited by selective suppression (shown here in red), which facilitates execution of the correct, color-based, action. The selective suppression takes time to develop, which results in a relatively high proportion of fast erroneous (location-based) responses on incongruent trials, with near-perfect response accuracy when responses are slower as the inappropriate responses have been suppressed. These effects seen by plotting response accuracy as a function of response time in a Conditional Accuracy Function (CAF) as shown in (B). Furthermore, the interference on incongruent trials would be expected to reduce as unwanted responses are suppressed as RTs increase. This effect can be seen in the negative slope of a delta plot, as shown in (C). Delta plots can reveal individual and group differences in motor activation and suppression that cannot be seen in average measures of RT. Panels (B) and (C) were originally published in van den denWildenberg et al. (2010). Front. Hum. Neurosci. 4:222. doi: 10.3389/fnhum.2010.00222. Panel (B) was modified after Wylie et al. (2009), with permission from Elsevier. Panel (C) was modified after Wylie et al. (2010).
Processing by the fast, direct processing route is automatic, and occurs irrespective of task instructions. For example, the spatial location of a target stimulus in a Simon task would be processed quickly and automatically via the direct processing route. At the same time, processing of the task-relevant target attribute (e.g., target color in a Simon task) proceeds via a slower indirect processing route. On congruent trials, the same response is activated by both the direct and the indirect processing routes, producing fast, correct responses. On incongruent trials, however, the direct processing route and the indirect processing route activate different responses which results in increased error rates, and slower response times as the conflict between competing responses is resolved.
Importantly, models of information processing in conflict tasks often include an active inhibition mechanism which acts to selectively suppress inappropriate response activation resulting from the direct processing route. Evidence for such control over automatically activated responses can be gleaned by studying the temporal dynamics of interference effects—for example, by plotting the accuracy of responses as a function of response speed as a Conditional Accuracy Function (CAF, see van den denWildenberg et al., 2010 for a review; see Figure 2B). In conflict tasks, accuracy for compatible trials is near-perfect, while fast responses on incompatible trials are often near (e.g., Wylie et al., 2009) or below (e.g., Stins et al., 2007) chance level. This pattern of erroneous responses is unlikely to be purely a result of fast-guessing, which would be expected to result in similar accuracy levels for both incompatible and compatible trials. Instead, these findings are consistent with the suggestion that erroneous responses are activated quickly via the direct processing route, before being selectively suppressed by an inhibitory control mechanism.
How response activation and suppression change as a function of response speed can also be seen in a delta plot (e.g., Burle et al., 2002; Ridderinkhof, 2002; Ridderinkhof et al., 2005; Wylie et al., 2010). This shows the size of the compatibility effect (RT on compatible trials subtracted from the RT on incompatible trials) as a function of RT (see Figure 2C). The plots make use of the whole RT distribution, rather than relying on a single measure of the central tendency, and therefore, they can reveal differences in the dynamics of response activation and suppression across individuals or groups even when mean RTs are not reliably different. Delta plots in traditional conflict tasks typically show initial positive effects that level off, or even become negative, as response times increase (e.g., de Jong et al., 1994; see also van den denWildenberg et al., 2010 for a review). This leveling-off is consistent with suppression of the unwanted stimulus-triggered response activation. As this suppression takes time to develop, a pattern of facilitation followed by inhibition is shown in the RT delta plot. Note however, that it is not necessary to postulate the existence of an active inhibitory mechanism which acts to selectively suppress inappropriately activated responses to explain the temporal dynamics of interference effects: the patterns shown in CAFs and delta plots may instead be produced by early activation of the inappropriate response which spontaneously decays over time (but see Burle et al., 2002 for evidence against this suggestion).
Pre-Stimulus vs. Post-Stimulus Cognitive Control
Control mechanisms that can override inappropriate response plans which have been automatically evoked by the environment not only act to inhibit responses after they have been evoked by the stimulus. Pre-stimulus control mechanisms also seem to play a role. Thus, task set and previous experience can modulate conflicting response tendencies in a preparatory manner. Indeed, there is good evidence that pre-stimulus inhibitory mechanisms play a role in controlling responses in many contexts, including stop-signal (e.g., Verbruggen and Logan, 2009b), the anti-saccade (e.g., Everling and Munoz, 2000; Munoz and Everling, 2004), and reaction time tasks incorporating warning signals (e.g., Boulinguez et al., 2008).
The effects of pre-stimulus control have also been reported in traditional “conflict” tasks. Thus, the size of the congruency effect on the current trial in a conflict task can be modulated by (1) a pre-cue indicating whether the upcoming trial will be congruent or incongruent (e.g., Logan and Zbrodoff, 1982); (2) the ratio of congruent and incongruent trials in a block or experiment (e.g., Logan and Zbrodoff, 1979); and (3) whether the immediately preceding trial was congruent or incongruent (the so-called “Gratton effect” e.g., Gratton et al., 1988, 1992).
The Gratton effect has in particular been subject to much investigation. While some have suggested that it arises from repetition priming of exactly the same stimulus-response links from previous trials (e.g., Mayr et al., 2003), others have reported that it can occur without exact stimulus-response repetitions (e.g., Kerns et al., 2004; Akçay and Hazeltine, 2007). An influential conflict monitoring hypothesis proposed that following the response conflict experienced on an incongruent trial, cognitive control mechanisms which resolve this conflict are boosted for the next trial, in turn leading to reduced interference if the subsequent trial is also incongruent (e.g., Botvinick et al., 2001). This suggestions is supported by several observations that performance on incongruent trials is improved if they are preceded by another incongruent trial relative to a congruent trial (e.g., by Gratton et al., 1992 using Eriksen flankers; McBride et al., 2012 using object affordance; and by Stürmer et al., 2002 using the Simon task). However, this pattern has not consistently been reported and several researchers have documented selective benefits for compatible trials when the previous trial was also compatible (e.g., Kunde and Wühr, 2006; Akçay and Hazeltine, 2007; van Gaal et al., 2010a; Schlaghecken and Martini, 2011).
Schlaghecken and Martini (2011) recently accounted for these discrepant findings by suggesting that the effects of trial history on cognitive control were driven by a mechanism which responds to the previous experience of both the presence and the absence of conflict, arguing that the mechanism is one of context adaptation, rather than conflict adaptation. Whatever the mechanisms are that produce pre-stimulus control effects, it is clear that task set, instruction, and previous experience can modulate the apparently automatic priming of motor responses by visual objects.
Unconscious Control Over Unwanted Responses
Traditionally, cognitive control mechanisms resulting in response inhibition have been considered as tightly coupled to consciousness (e.g., Schneider and Shiffrin, 1977; Shiffrin and Schneider, 1977, 1984), just as for voluntary control over actions. The logic behind this view is that observers need to be aware of the interfering, control-evoking, stimulus in order for control mechanisms to be implemented and for unwanted motor responses to be suppressed. Such a suggestion is supported by evidence from studies showing that inhibition of primed responses only operates when stimuli are presented above—and not below—the threshold required for conscious awareness (e.g., Merikle et al., 1995 using the Stroop task).
However, there is now increasing evidence that some forms of cognitive control can be executed entirely automatically, without conscious awareness—or volition. Some of the most compelling evidence for the automatic inhibition of unconsciously triggered motor priming comes from several experiments by Eimer and Schlaghecken (for reviews, see Eimer and Schlaghecken, 2003; Sumner, 2007). In their paradigm, participants typically made a speeded button-press response according to the direction of a target arrow, which was preceded by a masked (subliminal) prime. When the interval between mask and target was short (e.g., 20–40 ms), participants showed the expected speeding of responses when prime and target were compatible relative to when they were incompatible (positive compatibility effect, PCE). However, when the interval between prime and mask was extended beyond around 100–150 ms, incompatible trials produced faster responses than compatible trials. In other words, the usual priming effect had reversed.
This negative compatibility effect (NCE) has now been widely reported with button-press responses, foot responses, and eye movements (e.g., Eimer and Schlaghecken, 1998, 2002, 2003; Schlaghecken and Eimer, 2000, 2002; Eimer et al., 2002; Aron et al., 2003; Schlaghecken et al., 2003; Seiss and Praamstra, 2004; Sumner et al., 2007; Boy et al., 2008; Hermens et al., 2010), as well as in electroencephalogram recordings (e.g., Eimer and Schlaghecken, 1998, 2003; Praamstra and Seiss, 2005). They can also bias free-choice responses (e.g., Klapp and Hinkley, 2002; Schlaghecken and Eimer, 2004; Klapp and Haas, 2005), and have been shown both with familiar stimulus response mappings—such as arrows—and also when stimuli have been arbitrarily mapped to responses (e.g., Boy et al., 2008; Sumner, 2008).
Many researchers have suggested that this reversed priming results from an inhibitory mechanism in the motor system which acts to suppress sub-threshold motor activation evoked by the prime (e.g., Eimer and Schlaghecken, 1998; Klapp and Hinkley, 2002; Schlaghecken and Eimer, 2002; Schlaghecken et al., 2006). The most recent evidence suggests that such inhibition is triggered by the arrival of new stimuli—that the appearance of a second stimulus after the prime (normally the mask) automatically elicits an “emergency brake” that cancels any motor activation initiated by the prime (Jaśkowski and Przekoracka-Krawczyk, 2005; Lleras and Enns, 2006; Jaśkowski, 2007, 2008; Boy et al., 2008).
However, there has been considerable debate over whether the NCE genuinely reflects an inhibitory mechanism rather than arising from a purely perceptual process, or alternatively, from positive priming (PCE) of a motor response associated with elements of the mask stimulus. Perceptual accounts suggest that the NCE occurs because perceptual processing of the target stimulus is slower following a compatible prime, due to habituation-like processes such as “repetition blindness” or an attentional refractory period (Bavelier et al., 2000; Huber, 2008; Sohrabi and West, 2008; see also van Leeuwen and Lachmann, 2004, also discussed in Lleras and Enns, 2005 alongside the object updating theory; Hochhaus and Johnston, 1996; Huber et al., 2001, 2002; Johnston et al., 2002; Sohrabi and West, 2009). Such perceptual processes may play a role in producing some reversed priming effects, but they cannot account for more recent findings. For example, Boy and Sumner (2010) found that when participants learned novel sensorimotor associations in a masked priming task, and those response mappings were suddenly switched, both positive and negative priming effects temporarily reversed (see Figure 3)—indicating that the old response mappings continue to be primed until the participants learn the new mappings sufficiently well. Perceptual accounts of inverse priming cannot explain this finding.
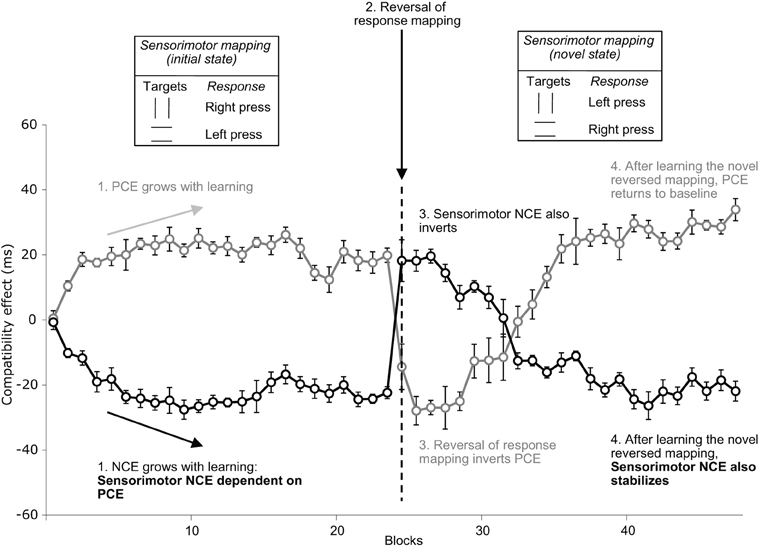
Figure 3. Evolution of Positive and Negative Compatibility Effects following the sudden reversal of the response mapping in the paradigm used by Boy and Sumner (2010, Experiment 4). Participants were trained to respond to targets in a masked prime paradigm alternating short and long mask-target intervals by blocks of five trials. Figure adapted with permission from Boy and Sumner (2010). J. Exp. Psychol. Hum. Percept. Perform. 36, 892–905, originally published by the American Psychological Association.
Alternatively, perceptual interactions between the prime and the mask could end up causing motor priming in the opposite direction to that expected from the prime. This idea has been variously termed “object-updating,” “active mask,” or “mask-induced priming” (Lleras and Enns, 2004; Verleger et al., 2004; see Sumner, 2007 for review). Many early experiments used masks that were constructed by superimposing features in the alternative primes. In this case the most visually salient features of the mask could be those that were new onsets in the stimulus sequence—i.e., those that were not in the prime. Thus, the prime-mask sequence could actually prime the response opposite the one associated with the prime. Object updating may play a strong role in producing the NCE when masks are constructed from prime features, but they cannot account for the NCE in other cases where masks do not contain elements of possible primes (e.g., Sumner, 2008).
Overall, therefore, reversed priming effects can be caused in several ways. For the purposes of this review, the most interesting one is a form of automatic motor inhibition, which can be revealed with appropriate stimuli. Finding the NCE with subliminally presented primes provides evidence that the mechanisms at its origin are deployed automatically. If the observer is not aware of the prime, then presumably they cannot volitionally suppress any motor response associated with it. However, note that the prime does not necessarily need to be subliminal for the NCE to occur (e.g., Klapp and Hinkley, 2002; Klapp, 2005; Lleras and Enns, 2005; Mattler, 2005; Sumner et al., 2006; Schlaghecken et al., 2008).
Automatic Inhibition in the Affordance Paradigm
If the NCE shown in masked priming genuinely reflects an automatic control mechanism to suppress sub-threshold activation of an unwanted motor response (see above), one might expect to see evidence of an NCE in other paradigms where visual stimuli automatically evoke motor responses. Vainio and colleagues have recently reported that the positive stimulus-response compatibility effects usually shown in object affordance tasks can become negative if the object stimulus is presented briefly and then removed (e.g., Vainio, 2009; Vainio et al., 2011; see also Vainio and Mustonen, 2011).
This NCE-like effect was reported even when the prime stimulus (e.g., a cup) was not relevant to the on-going task (respond to direction of a subsequently presented target arrow), quite unlike the masked prime paradigm where primes typically need to contain elements relevant to the task for NCEs to be observed (e.g., Eimer and Schlaghecken, 1998). To account for this discrepancy, Vainio and colleagues suggested that even though the cup primes in their experiments were irrelevant to the participants' task, a small degree of motor activation occurred due to the relatively long stimulus duration (compared to the primes in previous NCE studies), and the fact that the response association is highly over-learnt (compared to the semi-arbitrary correspondence between simple arrows or lines and a response). However, the associated motor activation was still sub-threshold, and thus able to be reversed by inhibition when perceptual support for that response was interrupted (producing the observed NCE). Overall, these studies suggest that actions which have been automatically primed by object affordances may also be subject to automatic control.
Automatic Triggering of “Endogenous” Control
In the masked prime paradigm, the participants are not actually instructed to employ response inhibition—it just appears to occur automatically following the prime and mask. In other paradigms, participants are specifically asked to endogenously inhibit their planned motor responses when cued to do so, for example in the go/no-go task and stop signal task (e.g., Logan, 1994). In both tasks, participants are instructed to respond as quickly as possible to go-signals, but to withhold their response when other (“stop”) stimuli occur.
Traditionally, such endogenous response inhibition has been thought to depend on the conscious detection of stop signals (e.g., Eimer and Schlaghecken, 2003). However, recent work suggests that endogenous suppression of pre-potent responses can also be primed or evoked unconsciously and automatically (e.g., Verbruggen and Logan, 2009a; van Gaal et al., 2008, 2009, 2010a,b). For example, van Gaal et al. (2009, 2010b) examined whether inhibition of a response could be triggered unconsciously by a masked stop-signal. They reported that although participants did not completely withhold their responses on unconscious (strongly masked) stop trials, they were significantly slowed relative to go trials, indicating there was some (incomplete) suppression of responses triggered by the imperceptible stop signal. Thus, the control processes involved in suppressing responses can be—at least partially—evoked by signals that are not consciously perceived.
Automatic Pre-Stimulus Control
In the case of both the NCE and the “endogenous inhibition” paradigms discussed above, inhibitory control processes appear to be evoked to deal with motor activation after it has been elicited. Is it also possible that pre-stimulus preparatory types of control can be elicited automatically? Many researchers have suggested that observers must consciously experience conflict in order for the pre-stimulus control mechanisms to be deployed (e.g., Kunde, 2003; Mayr, 2004; Ansorge et al., 2011). However, recent evidence from van Gaal et al. (2010a) suggests that some pre-stimulus control can be evoked automatically, without conscious awareness (see Figure 4). They used a meta-contrast masking paradigm to manipulate awareness of conflict-inducing stimuli. Conflict between co-activated responses was either conscious (weakly masked primes) or unconscious (strongly masked primes). The largest conflict adaptation effects occurred when both the current and the previous trial were weakly masked (visible). Importantly, a small but statistically significant conflict adaptation effect was evident when primes on trial n and trial n–1 were both presented below the threshold required for conscious awareness (strongly masked condition). This is consistent with the suggestion that unconsciously presented stimuli can automatically evoke these pre-stimulus conflict adaptation mechanisms, and can modulate the effects of subsequent conflicting stimuli. However, this result should be interpreted cautiously because responses are typically faster following fast responses (congruent trials), and slower following slow responses (incongruent trials; see e.g., Laming, 1979). If this effect were more apparent on fast (congruent) trials than on slow (incongruent) trials then it might entirely account for the small Gratton effect observed with non-conscious stimuli.
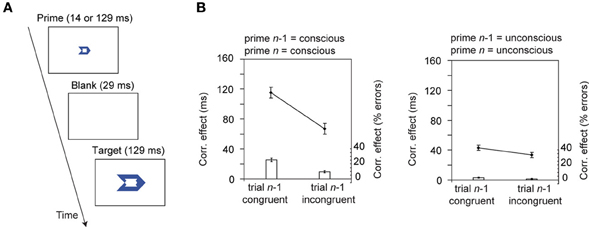
Figure 4. Automatic (unconscious) conflict adaptation effects as shown in van Gaal et al. (2010a). Observers responded to the direction of a target arrow stimulus that had been preceded by a backwards (meta-contrast) masked prime that either corresponded with the response required to the target (as shown in A), or was non-corresponding. The prime stimulus was either conscious (presented for 129 ms) or unconscious (presented for 14 ms). Correspondence effects (non-corresponding—corresponding) on trial n were modulated by whether trial n–1 was corresponding or not. These effects were largest when both trial n and trial n–1 had visible primes (B), but were still significant when both trial n and trial n–1 contained invisible primes. These figures were originally published in van Gaal et al. (2010a). PLoS One 5:e11508. doi: 10.1371/journal.pone.0011508.
Overlap between Brain Areas Responsible for Automatic and Voluntary Control
The traditional distinction drawn between automatic and voluntary cognitive control is not only being challenged by behavioral studies. Lesion and imaging studies have also revealed substantial overlap between brain regions traditionally associated with “voluntary” control and those active during automatic control. Brain areas in medial frontal cortex such as the SMA and anterior cingulate cortex (ACC) have traditionally been considered to be important for voluntary control (for a review, see Nachev et al., 2008). However, they also seem to be involved in mediating automatic motor activation and suppression of unwanted action plans (e.g., D'Ostilio and Garraux, 2011, 2012). For example, Sumner et al. (2007) used a masked prime paradigm to reveal that two extremely rare patients with microlesions of the SMA and/or the adjacent supplementary eye field (SEF) showed normal PCEs but failed to show NCEs, unlike healthy matched controls. These data are consistent with the view that the SMA and SEF may play causal roles in producing the automatic motor inhibition indexed behaviorally by the NCE. Thus, areas involved in the voluntary control of action might play a crucial role in automatic inhibition of unwanted actions (in this case, evoked by the subliminal prime).
There is also evidence from healthy observers that the SMA and nearby pre-SMA are involved in producing the unconscious NCE in healthy observers. Recent research from van Gaal et al. (2011b), found that individual differences in pre-SMA gray matter density were correlated with participants' ability to correctly respond to a target that had been preceded by a strongly masked (invisible) incompatible prime. Furthermore, Boy et al. (2010a) found that the fMRI signal was modulated by (invisible) prime compatibility in the SMA. Moreover, by studying normal participants' in vivo neurochemistry through magnetic resonance spectroscopy (MRS), Boy et al. (2010a) found that for a region including the SMA, the measured concentration of gamma-aminobutyric acid (GABA)—the main neurotransmitter responsible for neuronal inhibition—was correlated with the magnitude of the NCE.
There is also overlap between “voluntary” and “automatic” neural mechanisms involved in more conventional inhibition tasks such as go/no-go, and stop signal paradigms. van Gaal et al. (2010b) found that the amplitude of a fronto-central N2 event-related potential (ERP) component was reliably correlated with successful stopping on weakly masked (conscious) stop trials, and with the amount of slowing on strongly masked (unconscious) stop trials (as measured by the stop signal reaction time; SSRT). Thus, the size of this N2 component correlated with behavioral measures of both conscious and unconscious suppression of response. In addition, functional imaging has shown that strongly masked no-go signals activate much of the same brain areas that are activated by weakly masked no-go signals, particularly the pre-SMA and inferior frontal cortex (van Gaal et al., 2010b). The strength of activation in these areas was positively correlated with the amount of slow-down on strongly masked no-go trials—which supports the suggestion that this activity may be functional and have a direct effect on stopping behavior.
Taken together, these findings challenge the traditional assumption that voluntary control and involuntary mechanisms occur through distinct pathways in the brain. Rather, there is considerable overlap between the brain regions which are active during consciously and unconsciously triggered action control.
Dissociations in Automatic and Voluntary Control
Recent work from Boy et al. (2010b) suggests that the important distinction is not between control that is automatic compared to control that is voluntary, but rather between pre- and post-stimulus control (see discussion above on pre- vs. post-stimulus control). These investigators used a hybrid task which integrated masked priming into an Eriksen flanker paradigm (see Figure 5A). They showed an interaction between the post-stimulus inhibitory influences caused by prime-mask sequence (the NCE) and the post-stimulus control of flanker interference. Thus, these processes presumably share some common mechanisms (see Figure 5B). However, when examining the influence of the previous trials' flankers on performance in the current trial (the Gratton effect, see section on pre-stimulus control), no such interaction was found with the NCE. This suggests that pre-stimulus control mechanisms did not share processes with the NCE, and thus are distinct from post-stimulus mechanisms.
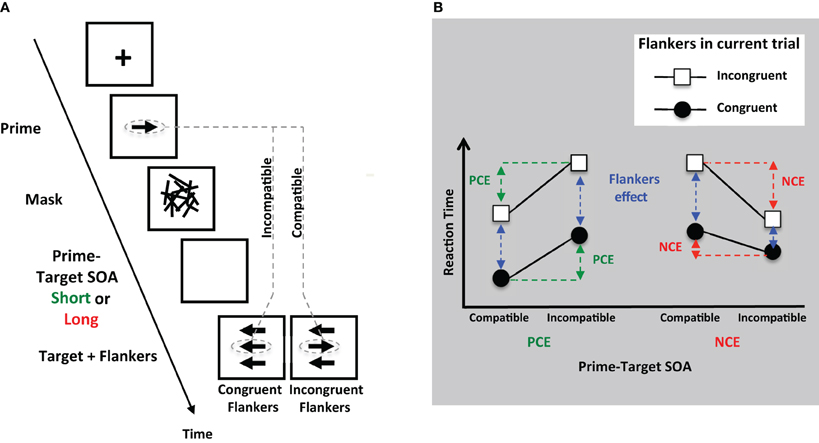
Figure 5. (A) Schematic illustration of the task in Boy et al. (2010b), which combines subliminal priming and flanker interference. If the stimulus onset asynchrony (SOA) between prime and target (response) cue is short (70 ms), a PCE in response times is found (green arrows in B). If the SOA is longer (180 ms), an NCE is found (red arrows in B). Compatibility refers to whether the direction of prime arrow and target (response) arrow is the same (compatible) or different (incompatible). Targets are flanked by arrows. If the flankers are congruent with the target, then response times are shorter than if flankers are incongruent (the flanker interference effect; blue arrows in B). 4-B. Results: for positive priming (PCE), the priming effect was additive with the effect of flanker interference. For the NCE, which measures subliminal inhibition, there was an interaction with flanker interference. This interaction did not occur for pre-stimulus control [data not shown here, see Boy et al. (2010b)].
Flexible Automaticty
Automatic and unconscious processes are traditionally regarded as inflexible (e.g., Schneider and Shiffrin, 1977; Shiffrin and Schneider, 1977, 1984), quite distinct in quality from the flexible nature of “voluntary” processes. However, there is increasing evidence that automatic and subliminal processes can in fact be modulated by “top-down” processes of attention, intention (“task set” or current goals) and expectation. For example, Kentridge and colleagues (e.g., Kentridge et al., 1999, 2004, 2008) have reported a series of studies in which attention modulates apparently subconscious processing, both in a “blindsight” patient, GY, and in normal participants. Focussing attention in time (e.g., Naccache et al., 2002) and in space (Lachter et al., 2004; Sumner et al., 2006; Marzouki et al., 2007) can also modulate the effects of masked primes on motor responses. For example, positive and negative compatibility effects in a masked prime task can be enhanced by exogenously pre-cueing prime location (Sumner et al., 2006), and in such a way that was not simply explained by an attentional boost to the perceptual strength of the prime. This suggests that attention does not only enhance perceptual processing, but can also modulate sensori-motor linkages.
Task set-up and instruction can also modulate many of the effects of visual stimuli on motor responses and control. As noted above, prime stimuli in masked priming tasks generally only affect responses when they share task-relevant elements with the target stimuli (e.g., Eimer and Schlaghecken, 1998; see also Huang et al., 2011). For example, Eimer and Schlaghecken (1998) found no NCE when participants responded to letter targets that had been preceded by masked arrow primes, even though arrow primes reliably prime responses when targets are also arrows. Moreover, recent evidence shows that object affordance effects are also dependent on the goals of the observer. Bub and Masson (2010) demonstrated that the handle of a mug only produced reliable affordance effects on reach and grasp responses, not when the response was made via a speeded button press.
Unconscious “endogenous” control over responses also depends on task instruction. Wokke et al. (2011) tested participants with a go/no-go task in which a masked (unconscious) prime stimulus preceded an unmasked (conscious) target. One of two possible targets was presented on each trial, but unlike other studies, stimuli were not consistently paired with either a “go” or a “no-go” response across a testing session. Instead, which of the possible targets required a “go” or a “no-go” response was cued on a trial-by-trial basis. Response inhibition rates were improved when a “no-go” target was preceded by an invisible “no-go” cue. As the instruction cue manipulation eliminated any long-term associations being built up between stimulus and response, it seems that control processes evoked by unconsciously presented stimuli can be triggered in a flexible manner according to task instruction.
Finally, pre-stimulus, conflict adaptation effects such as the Gratton effect can also be modulated, seemingly by reward (van Steenbergen et al., 2009). For example, van Steenbergen and colleagues (2009) showed that the conflict adaptation effect in a flanker task can be reduced by reward, even though rewards were given arbitrarily and were unrelated to the task. In fact Botvinick (2007) has recently suggested that conflict might be experienced as a negatively reinforcing event. As such, it is possible for the effects of conflict (a negative stimulus) to be counteracted by a positive stimulus (reward)—an example of possible flexible control over an automatic response.
Taken together, these findings indicate that seemingly “automatic” response activation and control can be implemented flexibly—quite unlike the traditionally inflexible view of automatic processes.
Conclusion
The evidence described in this review demonstrates that stimuli can automatically prime specific, purposeful actions. Simple stimuli can capture eye movements, produce activity in motor-related brain areas, and can trigger the actions afforded by an object. Because observers are constantly bombarded by a complex set of visual stimulations, such automatic activation of potential responses is likely to be important, either in facilitating responses or requiring inhibition so other responses can be made. While their effects might not be obvious in healthy adults, the effects of such automatic activation of motor programs can be dramatically revealed following brain damage (e.g., in alien limb syndrome or utilization behavior).
A necessary pre-requisite for flexible, goal-directed action is the ability to inhibit inappropriate, competing, responses even when those competing responses have been activated automatically. Cognitive control has traditionally been seen as tightly coupled to awareness, it has been suggested that an observer must be aware of a stimulus in order to inhibit motor activation evoked by that stimulus. Many of the findings reviewed here challenge this assumption and instead suggest there is substantial overlap between the mechanisms supporting conscious and unconscious control of responses. Thus, we suggest that while there may be differences between automatic and voluntary control, they may not be entirely distinct in the brain, that automatic processes may play a role in all behavior, and that we must revise traditional views that couple cognitive control to consciousness and automaticity to inflexibility.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by The Wellcome Trust and NIHR BRC at UCL/UCLH.
References
Akçay, C., and Hazeltine, E. (2007). Conflict monitoring and feature overlap: two sources of sequential modulations. Psychon. Bull. Rev. 14, 724–748.
Anderson, S. J., Yamagishi, N., and Karavia, V. (2002). Attentional processes link perception and action. Proc. R. Soc. B 269, 1225–1232.
Ansorge, U., Fuchs, I., Khalid, S., and Kunde, W. (2011). No conflict control in the absence of awareness. Psychol. Res. Psychol. Forsch. 75, 351–365.
Aron, A. R. (2007). The neural basis of inhibition in cognitive control. Neuroscientist 13, 214–228.
Aron, A. R., Schlaghecken, F., Fletcher, P. C., Bullmore, E. T., Eimer, M., Barker, R., Sahakian, B. J., and Robbins, T. W. (2003). Inhibition of subliminally primed responses is mediated by the caudate and thalamus: evidence from functional MRI and Huntington's disease. Brain 126, 713–723.
Bartolo, M., Zucchella, C., Pichiecchio, A., Pucci, E., Sandrini, G., and Sinforiani, E. (2011). Alien hand syndrome in left posterior stroke. Neurol. Sci. 32, 483–486.
Bavelier, D., Deruelle, C., and Proksch, J. (2000). Positive and negative compatibility effects. Percept. Psychophys. 62, 100–112.
Boccardi, E., Della Sala, S., Motto, C., and Spinnler, H. (2002). Utilisation behaviour consequent to bilateral SMA softening. Cortex 38, 239–308.
Bompas, A., and Sumner, P. (2011). Saccadic inhibition reveals the timing of automatic and voluntary signals in the human brain. J. Neurosci. 31, 12501–12512.
Botvinick, M. M. (2007). Conflict monitoring and decision making: reconciling two perspectives on anterior cingulate function. Cogn. Affect. Behav. Neurosci. 7, 356–366.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., and Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychol. Rev. 108, 624–652.
Boulinguez, P., Jaffard, M., Granjon, L., and Benraiss, A. (2008). Warning signals induce automatic EMG activations and proactive volitional inhibition: evidence from analysis of error distribution in simple RT. J. Neurophysiol. 99, 1572–1578.
Boy, F., Clarke, K., and Sumner, P. (2008). Mask stimulus triggers inhibition in subliminal visuomotor priming. Exp. Brain Res. 190, 111–116.
Boy, F., Evans, C. J., Edden, R. A. E., Singh, K. D., Husain, M., and Sumner, P. (2010a). Individual differences in subconscious motor control predicted by GABA concentration in SMA. Curr. Biol. 20, 1779–1785.
Boy, F., Husain, M., and Sumner, P. (2010b). Unconscious inhibition separates two forms of cognitive control. Proc. Natl. Acad. Sci. U.S.A. 107, 11134–11139.
Boy, F., and Sumner, P. (2010). Tight coupling between positive and reversed priming in the Masked Prime paradigm. J. Exp. Psychol. Hum. Percept. Perform. 36, 892–905.
Bub, D. N., and Masson, M. E. J. (2010). Grasping beer mugs: on the dynamics of alignment effects induced by handled objects. J. Exp. Psychol. Hum. Percept. Perform. 36, 341–358.
Buccino, G., Sato, M., Cattaneo, L., Rodà, F., and Riggio, L. (2009). Broken affordances, broken objects: a TMS study. Neuropsychologia 47, 3074–3078.
Buonocore, A., and McIntosh, R. D. (2008). Saccadic inhibition underlies the remote distractor effect. Exp. Brain Res. 191, 117–122.
Burle, B., Possamaï, C. -A., Vidal, F., Bonnet, M., and Hasbroucq, T. (2002). Executive control in the Simon effect: an electromyographic and distributional analysis. Psychol. Res. 66, 324–336.
Coles, M. G. H., Gratton, G., Bashore, T. R., Eriksen, C. W., and Donchin, E. (1985). A psychophysical investigation of the continuous flow model of human information processing. J. Exp. Psychol. Hum. Percept. Perform. 11, 529–553.
Coulthard, E., Rudd, A., Playford, E. D., and Husain, M. (2007). Alien Limb following posterior cerebral artery stroke: failure to recognize internally generated movements? Mov. Disord. 22, 1498–1502.
de Jong, R., Liang, C.-C., and Lauber, E. (1994). Conditional and unconditional automaticity: a dual-process model of effects of spatial stimulus-response correspondence. J. Exp. Psychol. Hum. Percept. Perform. 20, 731–750.
Derbyshire, N., Ellis, R., and Tucker, M. (2006). The potentiation of two components of the reach-to-grasp action during object categorisation in visual memory. Acta Psychol. 122, 74–98.
D'Ostilio, K., and Garraux, G. (2011). Automatic stimulus-induced medial premotor cortex activation without perception or action. PLoS One 6:e16613. doi: 10.1371/journal.pone.0016613
D'Ostilio, K., and Garraux, G. (2012). Dissociation between unconscious motor response facilitation and conflict in medial frontal areas. Eur. J. Cogn. Neurosci. 35, 332–340.
Driver, J., Davis, G., Picciardelli, P., Kidd, P., Maxwell, E., and Baron-Cohen, S. (1999). Gaze perception triggers reflexive visuospatial orienting. Vis. Cogn. 6, 509–540.
Edelman, J. A., and Xu, K. Z. (2009). Inhibition of voluntary saccadic eye movement commands by abrupt visual onsets. J. Neurophysiol. 101, 1222–1234.
Eimer, M., and Schlaghecken, F. (1998). Effects of masked stimuli on motor activation: behavioural and electrophysiological evidence. J. Exp. Psychol. Hum. Percept. Perform. 24, 1737–1747.
Eimer, M., and Schlaghecken, F. (2002). Links between conscious awareness and response inhibition: evidence from masked priming. Psychon. Bull. Rev. 9, 514–520.
Eimer, M., and Schlaghecken, F. (2003). Response facilitation and inhibition in subliminal priming. Biol. Psychol. 64, 7–26.
Eimer, M., Schubö, A., and Schlaghecken, F. (2002). Locus of inhibition in the masked priming of response alternatives. J. Mot. Behav. 34, 3–10.
Ellis, R., and Tucker, M. (2000). Micro-affordance: the potentiation of components of action by seen objects. Br. J. Psychol. 91, 451–471.
Ellis, R., Tucker, M., Symes, E., and Vainio, L. (2007). Does selecting one visual object from several require inhibition of actions associated with nonselected objects? J. Exp. Psychol. Hum. Percept. Perform. 33, 670–691.
Eriksen, B. A., and Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Percept. Psychophys. 16, 143–149.
Eriksen, C. W., Coles, M. G. H., Morris, L. R., and O'Hara, W. P. (1985). An electromyographic examination of response competition. Bull. Psychon. Soc. 23, 165–168.
Everling, S., and Munoz, D. P. (2000). Neuronal correlates for preparatory set associated with pro-saccades and anti-saccades in the primate frontal eye field. J. Neurosci. 20, 387–400.
Friesen, C. K., and Kingstone, A. (1998). The eyes have it! reflexive orienting is triggered by nonpredictive gaze. Psychon. Bull. Rev. 5, 490–495.
Friesen, C. K., and Kingstone, A. (2003). Abrupt onsets and gaze direction cues trigger independent reflexive attentional effects. Cognition 87, B1–B10.
Godijn, R., and Theeuwes, J. (2002). Programming of endogenous and exogenous saccades: evidence for a competitive integration model. J. Exp. Psychol. Hum. Percept. Perform. 28, 1039–1054.
Gratton, G., Coles, M. G. H., and Donchin, E. (1992). Optimizing the use of information: strategic control of activation of responses. J. Exp. Psychol. Gen. 121, 480–506.
Gratton, G., Coles, M. G. H., Sirevaag, E. J., Eriksen, C. W., and Donchin, E. (1988). Pre- and poststimulus activation of response channels: a psychophysiological analysis. J. Exp. Psychol. Hum. Percept. Perform. 14, 331–344.
Grèzes, J., and Decety, J. (2002). Does visual perception of object afford action? Evidence from a neuroimaging study. Neuropsychologia 40, 212–220.
Hampshire, A., Chamberlin, S. R., Monti, M. M., Duncan, J., and Owen, A. M. (2010). The role of the right inferior frontal gyrus: inhibition and attentional control. Neuroimage 50, 1313–1319.
Hermens, F., Sumner, P., and Walker, R. (2010). Inhibition of masked primes as revealed by saccade curvature. Vision Res. 50, 46–56.
Hochhaus, L., and Johnston, J. C. (1996). Pereceptual repetition blindness effects. J. Exp. Psychol. Hum. Percept. Perform. 22, 355–366.
Howard, L. A., and Tipper, S. P. (1997). Hand deviations away from visual cues: indirect evidence for inhibition. Exp. Brain Res. 113, 144–152.
Huang, Y., Zhou, T., and Chen, L. (2011). The precedence of topological change over top-down attention in masked priming. J. Vis. 11, 1–10.
Huber, D. E., Shiffrin, R. M., Lyle, K. B., and Ruys, K. I. (2001). Perception and preference in short-term word priming. Psychol. Rev. 108, 149–182.
Huber, D. E., Shiffrin, R. M., Quach, R., and Lyle, K. B. (2002). Mechanisms of source confusion and discounting in short-term priming: 1. Effects of prime duration and prime recognition. Mem. Cogn. 30, 745–757.
Humphreys, G. W., and Riddoch, M. J. (2000). One more cup of coffee for the road: object-action assemblies, response blocking and response capture after frontal lobe damage. Exp. Brain Res. 133, 81–93.
Irwin, D. E., Colcombe, A. M., Kramber, A. F., and Hahn, S. (2000). Attentional and oculomotor capture by onset, luminance and color singletons. Vision Res. 40, 1143–1458.
Ivanoff, J., and Klein, R. M. (2003). Orienting of attention without awareness is affected by measurement-induced attentional control settings. J. Vis. 3, 32–40.
Jaśkowski, P. (2007). The effect of nonmasking distractors in the priming of motor responses. J. Exp. Psychol. Hum. Percept. Perform. 32, 465–468.
Jaśkowski, P., and Przekoracka-Krawczyk, A. (2005). On the role of mask structure in subliminal priming. Acta Neurobiol. Exp. 65, 409–417.
Johnston, J. C., Hochhaus, L., and Ruthruff, E. (2002). Repetition blindness has a perceptual locus: evidence from online processing of targets in RSVP streams. J. Exp. Psychol. Hum. Percept. Perform. 28, 477–489.
Kentridge, R. W., Heywood, C. A., and Weiskrantz, L. (1999). Attention without awareness in blindsight. Proc. Biol. Sci. 266, 1805–1811.
Kentridge, R. W., Heywood, C. A., and Weiskrantz, L. (2004). Spatial attention speeds discrimination without awareness in blindsight. Neuropsychologia 42, 831–835.
Kentridge, R. W., Nijboer, T. C., and Heywood, C. A. (2008). Attended but unseen: visual attention is not sufficient for visual awareness. Neuropsychologia 46, 864–869.
Kerns, J. G., Cohen, J. D., MacDonald, A. W., Cho, R. Y., Stenger, V. A., and Carter, C. S. (2004). Anterior cingulate conflict monitoring and adjustments control. Science 303, 1023–1026.
Klapp, S. ((2005)). Two versions of the negative compatibility effect: comment on Lleras and Enns 2004. J. Exp. Psychol. Gen. 134, 431–435.
Klapp, S. T., and Haas, B. W. (2005). Nonconscious influence of masked stimuli on response selection is limited to concrete stimulus–response associations. J. Exp. Psychol. Hum. Percept. Perform. 31, 193–209.
Klapp, S. T., and Hinkley, L. B. (2002). The negative compatibility effect: unconscious inhibition influences reaction time and response selection. J. Exp. Psychol. Gen. 131, 255–269.
Kornblum, S., Hasbroucq, T., and Osman, A. (1990). Dimensional overlap: cognitive basis for stimulus-response compatibility – a model and taxonomy. Psychol. Rev. 97, 253–270.
Kunde, W. (2003). Sequential modulations of stimulus-response correspondence effects depend on awareness of response conflict. Psychon. Bull. Rev. 10, 198–205.
Kunde, W., and Wühr, P. (2006). Sequential modulations of correspondence effects across spatial dimensions and tasks. Mem. Cogn. 34, 356–367.
Lachter, J., Forster, K. I., and Ruthruff, E. (2004). Forty-five years after Broadbent (1958): still no identification without attention. Psychol. Rev. 111, 880–913.
Leuthold, H., and Kopp, B. (1998). Mechanisms of priming by masked stimuli: Inferences from event-related brain potentials. Psychol. Sci. 9, 263–269.
Lhermitte, F. (1983). “Utilization behaviour” and its relation to lesions of the frontal lobes. Brain 106, 237–255.
Link, S. W., and Heath, R. A. (1975). A sequential theory of psychological discrimination. Psychometrika 40, 77–105.
Lleras, A., and Enns, J. T. (2004). Negative compatibility of object updating? A cautionary tale of mask-dependent priming. J. Exp. Psychol. Gen. 133, 475–493.
Lleras, A., and Enns, J. T. ((2005)). Updating a cautionary tale of masked priming: a reply to Klapp 2005. J. Exp. Psychol. Hum. Percept. Perform. 134, 436–440.
Lleras, A., and Enns, J. T. ((2006)). How much like a target can a mask be? Geometric, spatial and temporal similarity in priming: a reply to Schlaghecken and Eimer 2006. J. Exp. Psychol. Gen. 135, 495–500.
Logan, G. D. (1994). “On the ability to inhibit through and action. A users' guide to the stop signal paradigm,” in Inhibitory Processes in Attention, Memory and Language, eds D. Dagenbach and T. H. Carr (San Diego, CA: Academic Press), 189–236.
Logan, G. D., and Zbrodoff, N. J. (1979). When it helps to be misled: facilitative effects of increasing the frequency of conflicting stimuli in a Stroop-like task. Mem. Cogn. 7, 166–174.
Logan, G. D., and Zbrodoff, N. J. (1982). Constraints on strategy construction in a speeded discrimination task. J. Exp. Psychol. Hum. Percept. Perform. 8, 502–520.
Lu, C-H., and Proctor, R. W. (1995). The influence of irrelevant location information on performance: a review of the Simon and spatial Stroop effects. Psychon. Bull. Rev. 2, 174–207.
Marzouki, Y., Grainger, J., and Theeuwes, J. (2007). Exogenous spatial cueing modulates subliminal masked priming. Acta Psychol. 126, 34–45.
Mattler, U. (2005). Inhibition and decay of motor and nonmotor priming. Percept. Psychophys. 67, 285–300.
Mayr, U., Awh, E., and Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nat. Neurosci. 6, 450–452.
McBride, J., Sumner, P., and Husain, M. (2012). Conflict in object affordance revealed by grip force. Q. J. Exp. Psychol. 65, 13–24.
McCormick, P. A. (1997). Orienting attention without awareness. J. Exp. Psychol. Hum. Percept. Perform. 23, 168–180.
McPeek, R. M., Han, J. H., and Keller, E. L. (2003). Competition between saccade goals in the superior colliculus produces saccade curvature. J. Neurophysiol. 89, 2577–2590.
McPeek, R. M., and Keller, E. L. (2001). Short-term priming, concurrent processing, and saccade curvature during a target selection task in the monkey. Vision Res. 41, 785–800.
McPeek, R. M., Skavenski, A. A., and Nakayama, K. (2000). Concurrent processing of saccades in visual search. Vision Res. 40, 2499–2516.
Merikle, P. M., Joordens, S., and Stolz, J. A. (1995). Measuring the relative magnitude of unconscious influences. Conscious. Cogn. 4, 422–439.
Mulckhuyse, M., and Theeuwes, J. (2010). Unconscious attentional orienting to exogenous cues: a review of the literature. Acta Psychol. 134, 299–309.
Munoz, D. P., and Everling, S. (2004). Look away: the anti-saccade task and the voluntary control of eye movement. Nat. Rev. Neurosci. 5, 218–228.
Murray, R., Neumann, M., Forman, M. S., Farmer, J., Massimo, L., Rice, A., Miller, B. L., Johnson, J. K., Clark, C. M., Hurtig, H. I., Gorno-Tempini, M. L., Lee, V. M.-Y., Trojanowski, J. Q., and Grossman, M. (2007). Cognitive and motor assessment in autopsy-proven corticobasal degeneration. Neurology 68, 1274–1283.
Naccache, L., Blandin, L., and Dehaene, S. (2002). Unconscious masked priming depends on temporal attention. Psychol. Sci. 13, 416–424.
Nachev, P., Kennard, C., and Husain, M. (2008). Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 9, 856–869.
Ögmen, H., and Breitmeyer, B. G. (2006). The First Half Second: Temporal Dynamics of Conscious and Unconscious Visual Processing. Cambridge, MA: MIT Press.
Phillips, J. C., and Ward, R. (2002). S-R correspondence effects of irrelevant visual affordance: time course and specificity of response activation. Vis. Cogn. 9, 540–558.
Praamstra, P., and Seiss, E. (2005). The neurophysiology of response competition: motor cortex activation and inhibition following subliminal response priming. J. Cogn. Neurosci. 17, 483–493.
Reingold, E. M., and Stampe, D. M. (1999). “Saccadic inhibition in complex visual tasks,” in Current Oculomotor Research: Physiological and Psychological Aspects, eds W. Becker, H. Deubel and T. Mergner (New York, NY: Plenum), 249–255.
Reingold, E. M., and Stampe, D. M. (2000). “Saccadic inhibition and gaze contingent research paradigms,” in Reading as A Perceptual Process, eds A. Kennedy, R. Radach, D. Heller and J. Pynte (Amsterdam: Elsevier), 119–145.
Reingold, E. M., and Stampe, D. M. (2002). Saccadic inhibition in voluntary and reflexive saccades. J. Cogn. Neurosci. 14, 371–388.
Reingold, E. M., and Stampe, D. M. (2003). “Using the saccadic inhibition paradigm to investigate saccadic control in reading,” in Mind's Eye: Cognitive and Applied Aspects of Eye Movement Research, eds R. Radach, J. Hyona and H. Deubel (Amsterdam: Elsevier Science), 347–360.
Ridderinkhof, K. R. (2002). “Activation and suppression in conflict tasks: empirical clarification through distributional analyses,” in Common Mechanisms in Perception and Action. Attention and Performance, Vol. XIX, eds W. Prinz and B. Hommel (Oxford: Oxford University Press), 494–519.
Ridderinkhof, K. R., Scheres, A., Oosterlaan, J., and Sergent, J. A. (2005). Delta plots in the study of individual differences: new tools reveal response inhibition deficits in AD/Hd that are eliminated by methylphenidate treatment. J. Abnorm. Psychol. 114, 197–215.
Riddoch, M. J., Edwards, M. G., Humphreys, G. W., West, R., and Heafield, T. (1998). Visual affordances direct action: neuropsychological evidence from manual interference. Cogn. Neuropsychol. 15, 645–683.
Sato, W., Okada, T., and Toichi, M. (2007). Attentional shift by gaze is triggered without awareness. Exp. Brain Res. 183, 87–94.
Scepkowski, L. A., and Cronin-Golumb, A. (2003). The alien hand: cases, categorizations and anatomical correlates. Behav. Cogn. Neurosci. Rev. 2, 261–277.
Schaefer, M., Heinze, H. J., and Galazky, I. (2010). Alien hand syndrome: neural correlates of movements without conscious will. PloS One 5:15010. doi: 10.1371/journal.pone.0015010
Schlaghecken, F., Blagrove, E., and Maylor, E. A. (2008). No difference between conscious and nonconscious visoumotor control: evidence from perceptual learning in the masked prime task. Conscious. Cogn. 17, 84–93.
Schlaghecken, F., Bowman, H., and Eimer, M. (2006). Dissociating local and global levels of perceputo-motor control in masked priming. J. Exp. Psychol. Hum. Percept. Perform. 32, 618–632.
Schlaghecken, F., and Eimer, M. (2000). A central-peripheral asymmetry in masked priming. Percept. Psychophys. 62, 1367–1382.
Schlaghecken, F., and Eimer, M. (2002). Motor activation with and without inhibition: evidence for a threshold mechanism in motor control. Percept. Psychophys. 64, 148–162.
Schlaghecken, F., and Eimer, M. (2004). Masked prime stimuli can bias “free” choices between response alternatives. Psychon. Bull. Rev. 11, 463–468.
Schlaghecken, F., and Martini, P. (2011). Context, not conflict, drives cognitive control. J. Exp. Psychol. Hum. Percept. Perform. doi: 10.1037/a0025791. [Epub ahead of print]
Schlaghecken, F., Münchau, A., Bloem, B. R., Rothwell, J., and Eimer, M. (2003). Slow frequency repetitive transcranial magnetic stimulation affects reaction times, but not priming effects, in a masked prime task. Clin. Neurophysiol. 114, 1272–1277.
Schneider, W., and Shiffrin, R. M. (1977). Controlled and automatic human information processing: i. detection, search, and attention. Psychol. Rev. 84, 1–66.
Seiss, E., and Praamstra, P. (2004). The basal ganglia and inhibitory mechanisms in response selection: evidence from subliminal priming of motor responses in Parkinson's disease. Brain 127, 330–339.
Sheliga, B. M., Riggio, L., and Rizzolatti, G. (1995). Spatial attention and eye movements. Exp. Brain Res. 105, 261–275.
Shiffrin, R. M., and Schneider, W. (1977). Controlled and automatic human information processing: I. Perceptual learning, automatic attending, and a general theory. Psychol. Rev. 84, 127–190.
Shiffrin, R. M., and Schneider, W. (1984). Automatic and controlled processing revisited. Psychol. Rev. 91, 269–276.
Smith, P. L., and Ratcliff, R. (2004). Psychology and neurobiology of simple decisions. Trends Neurosci. 27, 161–168.
Sohrabi, A., and West, R. L. (2008). “A biologicallly-plausible cognitive model (BPCM) of positive and negative congruency effects in masked priming,” in Poster Session Presented at the Annual Meeting of the Cognitive Science Society, Washington, DC.
Sohrabi, A., and West, R. L. (2009). Positive and negative congruency effects in masked priming: a neuro-computational model based on representation, attention, and conflict. Brain Res. 15, 126–132.
Stins, J. F., Polderman, J. C. T., Boomsma, D. I., and de Geus, E. J. C. (2007). Conditional accuracy in response interference tasks: evidence from the Eriksen flanker task and the spatial conflict task. Adv. Cogn. Psychol. 3, 389–396.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. J. Exp. Psychol. 18, 643–662.
Stürmer, B., Leuthold, H., Soetens, E., Schröter, H., and Sommwe, W. (2002). Control over location-based response activation in the Simon task: behavioral and electrophysiological evidence. J. Exp. Psychol. Hum. Percept. Perform. 28, 1345–1363.
Sumner, P. (2007). Negative and positive masked-priming - implications for motor inhibition. Adv. Cogn. Psychol. 3, 317–326.
Sumner, P. (2008). Mask-induced priming and the negative compatibility effect. Exp. Psychol. 55, 133–141.
Sumner, P., and Husain, M. (2008). At the edge of consciousness: automatic motor activation and voluntary control. Neuroscientist 14, 474–486.
Sumner, P., Nachev, P., Morris, P., Peters, A. M., Jackson, S., Kennard, C., and Husain, M. (2007). Human median frontal cortex mediates unconscious inhibition of voluntary action. Neuron 54, 697–711.
Sumner, P., Tsai, P. -C., Yu, K., and Nachev, P. (2006). Attentional modulation of sensorimotor processes in the absence of perceptual awareness. Proc. Natl. Acad. Sci. U.S.A. 103, 10520–10525.
Swann, N. C., Cai, W., Conner, C. R., Pieters, T. A., Claffey, M. P., George, J. S., Aron, A. R., and Tandon, N. (2012). Roles for the pre-supplementary motor area and the right inferior frontal gyrus in stopping action: electrophysiological responses and functional and structural connectivity. Neuroimage 59, 2860–2870.
Swann, N. C., Tandon, N., Canolty, R., Ellmore, T. M., McEvoy, L. K., Dreyer, S., DiSano, M., and Aron, A. (2009). Intracranial EEG reveals a time- and frequency-specific role for the right inferior frontal gyrus and primary motor cortex in stopping initiated responses. J. Neurosci. 29, 12675–12685.
Theeuwes, J., Kramer, A. F., Hahn, S., and Irwin, D. E. (1998). Our eyes do not always go where we want them to go: capture of the eyes by new objects. Psychol. Sci. 9, 379–385.
Tipper, S. P., Howard, L. A., and Jackson, S. R. (1997). Selective reaching to grasp: evidence for distractor interference effects. Vis. Cogn. 4, 1–38.
Tipples, J. (2002). Eye gaze is not unique: automatic orienting in response to uninformative arrows. Psychon. Bull. Rev. 9, 214–318.
Tiwari, D., and Amar, K. (2008). A case of corticobasal degeneration presenting with alien limb syndrome. Age Ageing 37, 600–601.
Tucker, M., and Ellis, R. (1998). On the relations between seen objects and components of potential actions. J. Exp. Psychol. Hum. Percept. Perform. 24, 830–846.
Tucker, M., and Ellis, R. (2001). The potentiation of grasp types during visual object categorization. Vis. Cogn. 8, 769–800.
Usher, M., and McClelland, J. (2001). The time course of perceptual choice: the leaky, competing accumulator model. Psychol. Rev. 108, 550–592.
Vainio, L. (2009). Interrupted object-based updating of reach program leads to a negative compatibility effect. J. Mot. Behav. 41, 305–315.
Vainio, L., Hammarèn, L., Hausen, M., Rekolainen, E., and Riskilä, S. (2011). Motor inhibition associated with the affordance of briefly displayed objects. Q. J. Exp. Psychol. 64, 1094–1110.
Vainio, L., and Mustonen, T. (2011). Mapping the Identity of a viewed hand in the motor system: evidence from stimulus-response compatibility. J. Exp. Psychol. Hum. Percept. Perform. 37, 207–221.
van den Wildenberg, W. P. M., Wylie, S. A., Forstmann, B. U., Burle, B., Hasbroucq, T., and Ridderinkhof, K. R. (2010). To head or to heed? Beyond the surface of selective action inhibition: a review. Front. Hum. Neurosci. 4:222. doi: 10.3389/fnhum.2010.00222
van Gaal, S., Lamme, V. A. F., Fahrenfort, J. J., and Ridderinkhof, K. R. (2011a). Dissociable brain mechanisms underlying the conscious and unconscious control of behavior. J. Cogn. Neurosci. 23, 91–105.
van Gaal, S., Scholte, H. S., Lamme, V. A. F., Fahrenfort, J. J., and Ridderinkhof, K. R. (2011b). Pre-SMA gray matter density predicts individual differences in action selection in the face of conscious and unconscious response conflict. J. Cogn. Neurosci. 23, 382–390.
van Gaal, S., Lamme, V. A. F., and Ridderinkhof, K. R. (2010a). Unconsciously triggered conflict adaptation. PloS One 5:222. doi: 10.1371/journal.pone.0011508
van Gaal, S., Ridderinkhof, K. R., Fahrenfort, J. J., Scholte, H. S., and Lamme, V. A. F. (2008). Frontal cortex mediates unconsciously triggered inhibitory control. J. Neurosci. 28, 8053–8062.
van Gaal, S., Ridderinkhof, K. R., Scholte, H. S., and Lamme, V. A. F. (2010b). Unconscious activation of the prefrontal no-go network. J. Neurosci. 30, 4143–4150.
van Gaal, S., Ridderinkhof, K. R., van den Wildenberg, W. P. M., and Lamme, V. A. F. (2009). Dissociating consciousness from inhibitory control: evidence for unconsciously triggered response inhibition in the Stop-Signal task. J. Exp. Psychol. Hum. Percept. Perform. 35, 1129–1139.
van Leeuwen, C., and Lachmann, T. (2004). Negative and positive congruence effects in letters and shapes. Percept. Psychophys. 66, 908–925.
van Steenbergen, H., Band, G. P. H., and Hommel, B. (2009). Reward counteracts conflict adaptation: evidence for a role of affect in executive control. Psychol. Sci. 20, 1473–1477.
Verbruggen, F., Aron, A. R., Stevens, M. A., and Chambers, C. D. (2010). Theta burst stimulation dissociates attention and action updating in human inferior frontal cortex. Proc. Natl. Acad. Sci. U.S.A. 107, 13966–13971.
Verbruggen, F., and Logan, G. D. (2009a). Automaticity of cognitive control: goal priming in response inhibition paradigms. J. Exp. Psychol. Learn. Mem. Cogn. 35, 1381–1388.
Verbruggen, F., and Logan, G. D. (2009b). Proactive adjustments of response strategies in the stop-signal paradigm. J. Exp. Psychol. Hum. Percept. Perform. 35, 835–854.
Verleger, R., and Jaśkowski, P. Aydemir, A. van der Lubbe, R. H., Groen, M. (2004). Qualitative differences between conscious and nonconscious processing? On inverse priming induced by masked arrows. J. Exp. Psychol. Gen. 133, 494–515.
Walker, R., Kentridge, R. W., and Findlay, J. M. (1995). Independent contributions of the orienting of attention, fixation offset and bilateral stimulation on human saccadic latencies. Exp. Brain Res. 103, 294–310.
Walker, R., Mannan, S., Maurer, D., Pambakian, A. L. M., and Kennard, C. (2000). The oculomotor distractor effect in normal and hemianopic vision. Proc. Biol. Sci. 267, 431–438.
Walker, R., McSorley, E., and Haggard, P. (2006). The control of saccade trajectories: direction of curvature depends on prior knowledge of target location and saccade latency. Percept. Psychophys. 68, 129–138.
Wokke, M. E., van Gaal, S., Scholte, H. S., Ridderinkhof, K. R., and Victor A. F., Lamme (2011). The flexible nature of unconscious cognition. PLoS One 6:e25729. doi: 10.1371/journal.pone.0025729
Wylie, S. A., Ridderinkhof, K. R., Bashore, T. R., and van den Wildenberg, W. P. M. (2010). The effect of Parkinson's disease on the dynamics of online and proactive cognitive control during action selection. J. Cogn. Neurosci. 22, 2058–2073.
Keywords: action, cognitive control, response inhibition, unconscious, volition
Citation: McBride J, Boy F, Husain M and Sumner P (2012) Automatic motor activation in the executive control of action. Front. Hum. Neurosci. 6:82. doi: 10.3389/fnhum.2012.00082
Received: 31 January 2012; Paper pending published: 25 February 2012;
Accepted: 22 March 2012; Published online: 24 April 2012.
Edited by:
Nicola de Pisapia, University of Trento, ItalyReviewed by:
William C. Gaetz, The Children's Hospital of Philadelphia, USAFriederike Schlaghecken, University of Warwick, UK
Copyright: © 2012 McBride, Boy, Husain and Sumner. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Jennifer McBride, Institute of Cognitive Neuroscience and Institute of Neurology, University College London, Alexandra House, 17 Queen Square, London, WC1N 3AR, UK. e-mail:ai5tY2JyaWRlQHVjbC5hYy51aw==