
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Evol. Neurosci. , 02 November 2011
Volume 3 - 2011 | https://doi.org/10.3389/fnevo.2011.00005
This article is part of the Research Topic NeuroNEEPS View all 6 articles
 William D. Lassek1,2* and Steven J. C. Gaulin2
William D. Lassek1,2* and Steven J. C. Gaulin2
Because the first neurons evolved in an environment high in the n−3 (omega-3) fatty acid docosahexaenoic acid (DHA), this fatty acid became a major component of neural structure and function and makes up 10% of the dry weight of the human brain. Since n−3 fatty acids must come from the diet, this suggests a possible positive role for dietary n−3 fatty acids in cognition and a possible negative role for n−6 fatty acids, which compete with n−3 for access to critical enzymes. Because human females must provide DHA for the growth of the unusually large brains of their offspring from maternal fat stored during childhood, their need for DHA is especially great. We used stepwise regression to determine whether particular dietary fatty acids and other nutrients were related to cognitive performance in over 4000 American children aged 6–16 from the Third National Health and Nutrition Examination Survey; a variety of possible biological, social, and environmental risk factors were statistically controlled. In this context the only dietary factors related to cognitive performance were n−3 and n−6 fatty acids. Dietary n−3 fatty acids were positively related to cognitive test scores in male and female children, while n−6 showed the reverse relationship, significantly so in females. In female children the positive effects of n−3 intake were twice as strong as in males and exceeded the negative effects of lead exposure. This suggests that increasing dietary intake of n−3 and decreasing n−6 fatty acids may have cognitive benefits in children, especially in females.
The n−3 (also called omega-3) long-chain fatty acid docosahexaenoic acid (DHA, 22:6n−3), with a 22-carbon chain and six double bonds, comprises about 10% of the dry weight of the human brain (Svennerholm, 1968; Rapoport, 2003). Animal studies have shown that DHA readily crosses the blood/brain barrier (Ouellet et al., 2009) and plays a critical positive role in all aspects of neuronal growth, synaptic connections, and functioning (Cockburn, 1994; Jamieson et al., 1999; Salem et al., 2001; Chang et al., 2009). This includes roles in regulating the activity of Na + K + ATPase in the neural membrane (Bourre et al., 1989; Turner et al., 2003; Kumosani et al., 2011), neuron size (Ahmad et al., 2002), neurogenesis (Auestad and Innis, 2000; Coti Bertrand et al., 2006; Beltz et al., 2007; Novak et al., 2008; Da Costa et al., 2009; Dagai et al., 2009; He et al., 2009), neurite growth (Calderon and Kim, 2004; Sakamoto et al., 2007; Liu et al., 2008; Novak et al., 2008; Cao et al., 2009), synapse formation and function (Yoshida et al., 1997; Cansev and Wurtman, 2007; Wu et al., 2008; Cao et al., 2009; Wurtman et al., 2009), neuronal integrity and vitality (Issa et al., 2006; Mukherjee et al., 2007; Niemoller et al., 2009), gene expression in the brain (Kitajka et al., 2002), brain glucose transport (Pifferi et al., 2007), cognitive development (Heinemann and Bauer, 2006; Bongiovanni et al., 2007; Coluccia et al., 2009), and learning ability (Bourre et al., 1989; Yoshida et al., 1997; Greiner et al., 1999; Salem et al., 2001; Takeuchi et al., 2002; Shirai and Suzuki, 2004; Garcia-Calatayud et al., 2005; Lim et al., 2005; Chung et al., 2008; Holguin et al., 2008; Fedorova et al., 2009; He et al., 2009; Hooijmans et al., 2009; Jiang et al., 2009).
Because animals lack the enzyme required to make an n−3 double bond, at least the basal n−3 fatty acid, alpha-linolenic acid (ALA, 18:3n−3), must be obtained from their diets. Mammals can convert ALA to eicosapentaenoic EPA, 20:5n−3), and can subsequently convert EPA to docosapentaenoic (DPA, 22:5n−3) and then to DHA, though conversion efficiency is quite low and capacity limited, especially for EPA to DHA (Pawlosky et al., 2001). As cited above, animal studies show that a deficiency of dietary n−3 fatty acids leads to a decrease in neuronal size and synapse number and impaired learning ability. Many studies in human infants have shown that levels of DHA in the maternal diet or blood during pregnancy and in maternal milk or formula are positively related to cognitive and visual development in infants, as reviewed by McCann and Ames (2005); Eilander et al. (2007); Innis (2009); Ryan et al. (2010); and Schuchardt et al. (2010). Studies involving the n−3 content of the maternal diet may underestimate the effect of DHA, because most DHA delivered by a mother to her fetus or nursing infant derives from her fat stores rather than current intake (Sauerwald et al., 2000).
Most studies relating the level of n−3 fatty acids in the diet or blood to cognitive measures in older children have also found a positive relationship, including studies in Italy (Agostoni et al., 1997), Scotland (Whalley et al., 2004), Maryland (Ryan and Nelson, 2008), Finland (Aberg et al., 2009), Alabama (Neggers et al., 2009), and Wales (Kirby et al., 2010), though a Dutch study did not (De Groot et al., 2007). Two studies in which children were provided with short-term supplemental dietary n−3 found a positive effect on cognitive measures (Richardson and Montgomery, 2005; Dalton et al., 2009) and two did not (Osendarp et al., 2007; Kennedy et al., 2009). None of these studies have considered sex differences which, for reasons we will outline below, are to be expected.
The reliance of human and other mammalian brains on the neuronal functions of DHA appears to be the result of a very ancient evolutionary contingency. Though DHA is now a relatively scarce and limiting resource for the development of large brains in terrestrial environments, neurons first evolved in an aquatic environment where high levels of DHA were readily available. The first links in this chain of contingency apparently reach back more than 3 billion years.
Ancient cyanobacteria evolved the ability to synthesize n−3 ALA for incorporation into the thylakoid membrane where it plays an essential role in photosynthesis – as it does today in the chloroplasts of all green plants. Dinoflagellates and certain cyanobacteria and algae subsequently evolved a metabolic pathway to efficiently convert ALA to DHA using the enzyme delta-4-desaturase, and these phytoplankton are still the source of DHA for all aquatic animal life. Fossilized acritarchs suggest that dinoflagellates may have evolved more than 3 billion years ago (Javaux et al., 2010).
The first neurons evolved in Precambrian cnidarians feeding on dinoflagellates and other phytoplankton rich in DHA (Nichols et al., 2003; Putnam et al., 2007); hence neurons could evolve a design that was dependent on substantial supplies of DHA. When larger and more elaborate brains evolved in marine vertebrates, their neurons could continue to rely on large amounts of DHA because the phytoplankton producing this long-chain polyunsaturated fatty acid also lay at the base of their food chain. (DHA’s function is not limited to the vertebrate nervous system; it also plays important roles in muscles, blood, and mitochondria.)
Marine arthropods also have a diet rich in DHA, but when exclusively terrestrial arthropods first colonized the land, they were cut off from the DHA supplied by phytoplankton and had minimal ability to synthesize longer-chain n−3. Although their bodies have significant amounts of ALA obtained from plants, their nervous systems contain little or no DHA (Jerde et al., 1975; Fontaneto et al., 2011); instead they use mainly ALA (Stark et al., 1993; Shanker et al., 2006). Their inability to convert alpha-linolenic to DHA may limit the complexity of their nervous systems. Reptiles subsequently evolved this conversion ability, although their synthetic pathway differs from that of phytoplankton (lacking delta-4 desaturase), and is very much less efficient. Although allometrically small reptilian brains have some DHA, the proportion of DHA is quite low compared with mammalian brains (Mitchell et al., 2007), and their limited n−3 supply may have similarly constrained the growth and hence the evolution of a more complex nervous system.
The evolution of endothermy in mammals greatly increased caloric requirements, and the concomitant 10-fold increase in consumption of plants and/or insects provided much larger amounts of ALA, permitting the synthesis of larger amounts of DHA despite the inefficiency of this process in terrestrial animals. These higher levels of dietary n−3 allowed for a considerable expansion of the mammalian brain, which is not only allometrically much larger than a reptile brain but also contains a much higher proportion of DHA (Mitchell et al., 2007). However, mammals still have a lower proportion than fish (Stoknes et al., 2004; USDA, 2011). Higher DHA levels in mammalian mitochondrial membranes also facilitated endothermic metabolism (Brand et al., 1991, 1994; Hulbert, 2007). Meanwhile, the evolution and diversification of flowering plants led to increases in the n−3 content of terrestrial plants, especially in their fruits, nuts, and seeds, and permitted the co-evolution of many new species of herbivorous insects with high levels of ALA.
The first primates were insectivores occupying a nocturnal, arboreal niche, and their enhanced feeding skills and diet permitted the further expansion of the primate brain. (Chimpanzee females continue to invest considerable amounts of time feeding on insects as shown by McGrew, 1979.) Frugivorous primates also obtain substantial amounts of insects in the fruit they eat (Redford et al., 1984) as well as higher concentrations of n−3 in nuts and seeds. Folivorous primates, like gorillas, have relatively smaller brains compared to frugivores (Clutton-Brock and Harvey, 1980; Harvey et al., 1980), and must still ingest a large volume of plants to provide the necessary n−3.
As the hominid brain expanded to a size seven times larger than expected from the brain:body size relationship in mammals, the need for n−3 and DHA increased proportionately. Because human synthesis of DHA from ALA remains very limited, like that of other terrestrial animals, sources of preformed DHA in the diet are important. Potential sources include the meat, organs, and eggs of herbivores and birds, and especially flesh from aquatic animals, which provides larger amounts of DHA. Some have argued that exploitation of aquatic nutritional resources was essential for the evolution of the large hominid brain (Broadhurst et al., 2002), and there is evidence for significant amounts of aquatic foods in hominid diets from two million years ago (Braun et al., 2010; Stewart, 2010).
In order to grow their very large brains, human fetuses, and nursing infants require much larger amounts of DHA than can be reliably obtained from maternal daily intake, and most of the DHA they receive comes from maternal fat stores. Studies using radioisotope-labeled fatty acids show that approximately 80% of the DHA and other essential long-chain fatty acids provided in human milk come from maternal fat rather than from the current diet (Sauerwald et al., 2000). These fatty acids are stored mainly in women’s gluteofemoral fat, and these depots are protected except during the third trimester and lactation when their fatty acids are mobilized (Lassek and Gaulin, 2006). Because men do not make these physiological investments in offspring, women’s need for these fatty acids greatly exceeds that of men, a fact that probably explains the unique human sex difference in body fat.
There is no facilitated transport of DHA into adipose, so the proportion of DHA in fat stores depends on the concentration of DHA in the blood. Because this concentration is relatively low compared with other fatty acids, the percentage in adipose is also relatively low (0.2–0.3%). Developing human females must therefore have substantial amounts of adipose tissue in order to store sufficient amounts of DHA to support the growth of large brains in their children. In a study of Dutch children, female fat increased from 14.8 to 25.5% of body weight during puberty, while male fat decreased from 10.5 to 9.3% (Boot et al., 1997). In a sample of young American women, there was a mean of 16.2 kg of adipose tissue at the end of puberty (Lassek and Gaulin, 2006). Assuming a DHA percentage of 0.2%, this amount of adipose would contain 32 g of DHA which would become available when adipose is mobilized during late pregnancy and lactation, when the fetal and infant DHA requirement is 100–200 mg/day (Clandinin et al., 1980a,b; Haggarty, 2004). A female child must store this DHA at the same time that she requires substantial amounts of DHA to support her own growth and development. Because she must allocate some of her limited dietary n−3 to storing DHA for her future children, there is a competition between her need for DHA for her own body and brain and her need to store DHA for future reproduction. Girls with proportionately larger amounts of gluteofemoral fat and lower waist-hip ratios have earlier menarche (Lassek and Gaulin, 2007). While human males usually have much less adipose than females, they have substantially more than typical primates (Pond and Mattacks, 1987).
The optimal amount of DHA per day in children has not been established, but participants in a 1999 workshop sponsored by National Institutes of Health recommended that at least 0.1% of calories should be DHA (Simopoulos and Leaf, 1999), which would be 220 mg for a 2000 calorie (8.4 MJ) diet (assuming 9 calories/g of fat). Similar amounts are recommended for pregnant and nursing women (Koletzko et al., 2007.) The per capita amount of DHA and ALA in the American food supply in 1990 was 70 mg and 2.4 g respectively (Gerrior et al., 2004).
While n−3 fatty acids are known to have positive effects on cognition, less is known about the effects of n−6 fatty acids, such as linoleic (LA, 18:2n−6) and arachidonic acid (AA, 20:4n−6). As with n−3, some form of n−6 must come from the diet, and there is no interconversion of n−3 and n−6 fatty acids. In the terrestrial synthetic pathway that evolved to elongate ALA, the LA-to-arachidonic conversion competes for the same enzymes used to synthesize DHA from ALA (Rubin and Laposata, 1992; Emken et al., 1994; Innis et al., 2004; Hibbeln et al., 2006; Harnack et al., 2009; Gibson et al., 2011). Because of this metabolic competition, higher n−6 fatty acid intake might be expected to have negative effects on cognition; and four studies have shown this (Agostoni et al., 1997; Whalley et al., 2004; Novak et al., 2008; Neggers et al., 2009).
Amounts of n−3 fatty acids have declined in the American diet during the twentieth century while n−6 have increased (Blasbalg et al., 2011). Reconstructions of the paleolithic diet suggest that over most of human evolution, there was more n−3 than n−6 in the diet (Kuipers et al., 2010). Per capita linoleic acid in the American food supply was 29.3 g in 1990 and the ratio of total n−6 to n−3 was 12.3 (Gerrior et al., 2004). Because of their metabolic competition for necessary enzymes, the very high amount of n−6 linoleic acid compared with n−3 in the diet of American children is likely to decrease the conversion rate of ALA to DHA.
Larger, population-based samples may help to clarify the relationships between fatty acid consumption and cognitive performance. Cognitive and dietary data collected in the Third National Health and Nutrition Examination Survey (NHANES III) conducted in 1988–1994 provides an opportunity to examine the relationship between dietary n−3 and n−6 fatty acid intake and cognitive outcomes in a large sample of American children, as well as the possible effects of other nutrients. We recently found a relationship between lower maternal waist–hip ratios and cognitive performance which may be mediated by n−3 fatty acids (Lassek and Gaulin, 2008). This leads us to predict that dietary n−3 will also be positively related to cognitive outcomes in this sample, whereas dietary n−6 will be inversely related to the same outcome measures. Because of the much greater requirement for n−3 fatty acids in human females, we predict that their cognitive performance will be more sensitive to the amount of n−3 in the diet and to the competing effects of dietary n−6.
Detailed dietary histories based on 24-h recall were obtained by skilled interviewers for 13,923 males and 15,182 females aged 0–90 in the NHANES III sample, 1988–1994. From this larger sample we restricted our focus to children 6–16 years old. The child sample included 26% non-Hispanic whites, 35% non-Hispanic blacks, 35% Mexican-Americans, and 5% other. Because of the oversampling of blacks and Hispanics, this sample is not representative of the American population.
Twenty-four hour dietary recall was used to estimate individual intake. Specific fatty acid content of the foods consumed was estimated using the food database of the University of Minnesota’s Nutrition Coordinating Center, and for other nutrients, the USDA National Nutrient Database for Standard Reference (USDA). Nutrients used in the analysis which were estimated by conjoining the dietary histories and food composition databases in this way included vitamins A, B6, B12, C, and E, iron, folate, riboflavin, niacin, and thiamine, serum electrolytes, specific sugars, saturated fats with 10, 12, 14, 17, 18, 20, and 22 carbons, monounsaturated fats with 14, 16, 18, 20, and 22 carbons, the n−6 fatty acids LA and AA (summed for total n−6), the n−3 fatty acids ALA, EPA, DPA, and DHA (added together for total n−3), and total saturated, monounsaturated, and polyunsaturated fats.
Four cognitive tests were administered to 2253 males and 2309 females aged 6–16 in the NHANES III sample, including the math and reading tests from the Wide Range Achievement Test-Revised and the digit span and block design tests from the Wechsler Intelligence Scale for Children-Revised. The mean scaled score across these four tests was used as a measure of cognitive performance for each of the 6–16 year-olds in this study; both cognitive scores and dietary data were available for 2103 females and 2051 males, and these subgroups thus comprise our primary sample.
Other (non-dietary) measures included in the analyses as possible confounding variables include race/ethnicity, family income, family size, and years of education of the householder parent. Serum lead was also included because it is known to have a significant negative relationship with cognitive performance in children (Wasserman et al., 1997; Needleman and Landrigan, 2004). Multiple linear regression was performed using SPSS-17. Sample weights and complex adjustments for the sampling methodology were not used; this is in accordance with the recommendations of Korn and Graubard (1991).
Table 1 provides descriptive statistics for the children in the study sample. Mean energy intake was 20% higher in males and dietary fatty acid intakes showed similar differences; the mean test score and ratio of n−6 to n−3 was higher in females. Dietary intakes of fatty acids show a high degree of individual variation.
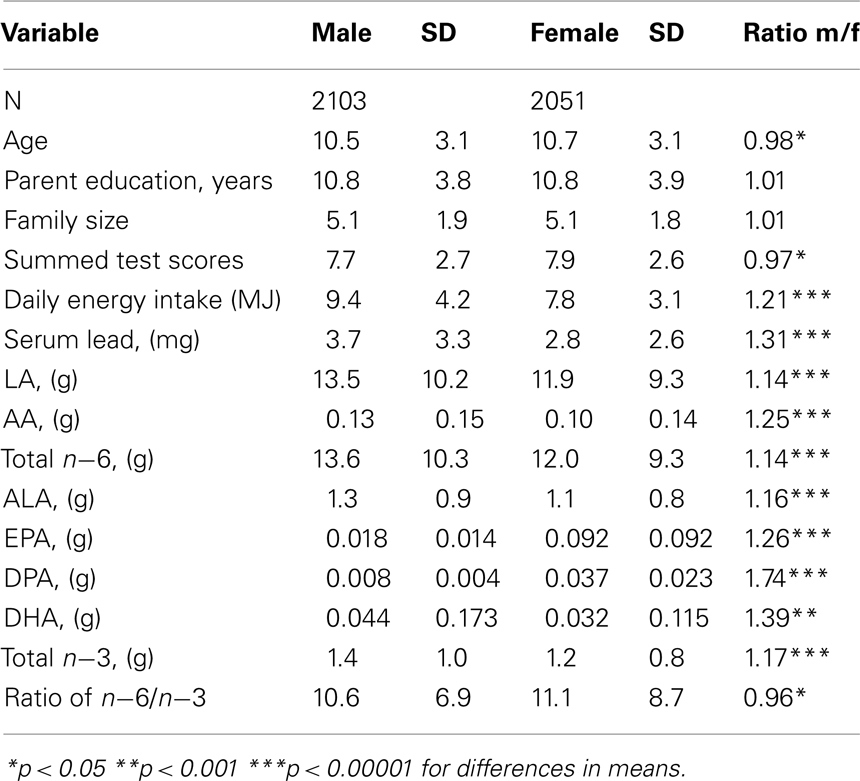
Table 1. Mean and SD by sex (data from NHANES III, ages 6–16).
Based on stepwise multiple regression, Table 2 shows the effect of significant nutrients on the cognitive performance of the children in the sample, controlling for race/ethnicity, family income and size, years of education of the householder parent, and serum lead. For both sexes, total dietary n−3 fatty acids are significantly positively related to test scores, while for females, total n−6 fatty acids are negatively related. None of the other 33 nutrients are significantly related, including iron and folate and total dietary saturated, monounsaturated, and polyunsaturated fatty acids. Based on its unstandardized regression coefficient, an increase in daily n−3 intake of 1 g increases the average test score by 0.38 points (±0.09 SD) in females and by 0.19 points (±0.10 SD) in males; this represents 0.14 and 0.07 of the SDs for the test scores. In females, but not males, when the ratio of n−6 to n−3 was used in place of the separate n−3 and n−6 intakes in the regression, it was significantly and negatively related to the cognitive score (beta = −0.071, p = 0.001). When the regression was run with both sexes, sex was not a significant variable.

Table 2. Standardized regression coefficients for the effect of dietary fatty acids on performance on four cognitive tests in youth 6–16, NHANES III.
National Health and Nutrition Examination Survey may sample more than one member in participating households; this creates a lack of independence among cases that is potentially problematic. To examine whether this lack of independence may have favored our hypothesis, we repeated the analysis using both sexes of children and limiting the sample to one child per household (1581 boys; 1682 girls), and found that there was no difference in the results of the regression.
Using a large sample drawn from NHANES III, dietary n−3 fatty acids are positively related to cognitive performance in children 6–16 years of age, while n−6 fatty acids are negatively related to cognitive performance in females in the same sample. As predicted, the contribution of dietary n−3 to cognitive performance is much greater (two-fold) in females, and females also show a significant negative effect for n−6 fatty acids which compete with n−3 for enzymes needed in the biosynthesis of DHA.
This result controls for other relevant variables known to affect cognitive outcomes. The special effects of n−3 and n−6 are apparent because dietary consumption of 33 other fatty acids and nutrients are not related to cognitive outcome measures based on the same dietary data set and the same children. The positive cognitive effect of dietary n−3 fatty acids, as measured by the imperfect method of 24-h recall, is of greater magnitude in girls than the negative effect of serum lead, a well known influence on cognition in children. Dietary iron and folate, which have also been found to relate to cognitive performance in some studies of children (Arija et al., 2006), were not significant when added to this regression.
Because of the complex sampling method used in the NHANES, these results should be viewed with caution and should not be considered representative of the American population. Also, the diet estimates used in this study were based on a single 24-h recall; and while this type of assessment is related to the long-term diet, it is not a highly accurate measure (Knutsen et al., 2003; Sekula et al., 2005; Slater et al., 2010). In addition, the children in the sample are past the ages of maximal brain growth, a period when the effects of dietary fatty acids would be expected to be greater. However, despite these limitations, their cognitive ability is still related to the amount of dietary n−3 fatty acids, and of the 40 nutritional variables used in the analysis, only n−3 and n−6 fatty acids were significantly related to cognitive ability.
The stronger effect of n−3 and significantly negative effect of n−6 in girls may reflect their greater need for n−3 fatty acids to sustain future pregnancy and lactation, as explained above. Because stored maternal fat is selectively used to support the development of the fetal and infant brain – via the placenta and breast milk – females must prepare for these demands by storing DHA in fat at a much higher rate than males during their childhood and adolescence, while their own brains and bodies are still growing. This competition between growth and reproductive goals, absent in boys, may make girls more subject to the antagonism between the n−6 and n−3 fatty acid families in commandeering necessary synthetic enzymes.
The effect of dietary fatty acids on cognition demonstrated here is relatively small – but of similar magnitude to the negative effect of lead. Both are environmental variables that can be altered, and clear steps have been taken in the case of lead. Since more than half of the variance in cognition is heritable (Plomin et al., 2000), and other environmental variables are often intractable (e.g., family income), dietary fatty acid intake may be of significant pragmatic importance.
These findings on the relationship between dietary fatty acids and cognitive performance are of particular interest in relation to current American food consumption patterns. The cognitive effects of dietary fatty acids in American children may be greater than in other populations because of the limited amount of n−3 fatty acids in the American diet combined with unusually high levels of n−6 (Blasbalg et al., 2011). Based on the energy intake of the sample children, the recommended amount of 0.1% of energy for DHA would be 250 in males and 210 mg in females per day. Assuming a very generous conversion rate of 2% for ALA to DHA, the mean total daily amount of DHA for the sample children would be just 70 mg for males and 54 mg for females, considerably short of the recommended amounts.
A coordinated increase in n−6 and decrease in n−3 supplies are the hallmarks of modern American industrial food production, and reflected in the high n−6/n−3 ratios in the NHANES III sample. Corn, which increased by 78% in the US diet from 1970 to 2000 (Putnam et al., 2002), has an n−6/n−3 ratio of 34 to 1 (USDA). Soybean oil, especially when partially hydrogenated or made from prevalent low-ALA varieties of soybeans, has a similarly high ratio. Moreover, n−3 in processed food is often removed because it is prone to spoil and thus reduces product shelf life (Holman, 1998).
An experimental study in rats using feeds corresponding to the Japanese diet (high n−3, low n−6) and American diet (high n−6, low n−3) found both better learning and many more synapses in the hippocampus in the rats fed the “Japanese” diet (Yoshida et al., 1997). It thus seems possible that the high n−6/n−3 ratio in the American diet might contribute to the relatively low ranking of American children in international testing (NCES, 2005) compared to children in countries with lower n−6/n−3 ratios, like Japan. Thus, evolutionary considerations may help lead to findings with considerable potential public health significance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Dr. Lassek was primarily responsible for the design of the analysis, data analysis, and the writing of the paper. Dr. Gaulin assisted in the design and data analysis and in the writing of the paper. We appreciate the helpful comments of the two reviewers.
Aberg, M. A. L., Aberg, N., Brisman, J., Sundberg, R., Winkvist, A., and Toren, K. (2009). Fish intake of Swedish male adolescents is a predictor of cognitive performance. Acta Paediatr. 98, 555–560.
Agostoni, C., Trojan, S., Bell, R., Riva, E., Bruzzese, M. G., and Giovannini, M. (1997). Developmental quotient at 24 months and fatty acid composition of diet in early infancy: a follow up study. Arch. Dis. Child. 76, 421–424.
Ahmad, A., Moriguchi, T., and Salem, N. (2002). Decrease in neuron size in docosahexaenoic acid-deficient brain. Pediatr. Neurol. 26, 210–218.
Arija, V., Esparo, G., Fernandez-Ballart, J., Murphy, M. M., Biarnes, E., and Canals, J. (2006). Nutritional status and performance in test of verbal and non-verbal intelligence in 6 year old children. Intelligence 34, 141–149.
Auestad, N., and Innis, S. M. (2000). Dietary n-3 fatty acid restriction during gestation in rats: neuronal cell body and growth-cone fatty acids. Am. J. Clin. Nutr. 71, 312S–314S.
Beltz, B. S., Tlusty, M. F., Benton, J. L., and Sandeman, D. C. (2007). Omega-3 fatty acids upregulate adult neurogenesis. Neurosci. Lett. 415, 154–158.
Blasbalg, T. L., Hibbeln, J. R., Ramsden, C. E., Majchrzak, S. F., and Rawlings, R. R. (2011). Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am. J. Clin. Nutr. 93, 950–962.
Bongiovanni, K. D., Depeters, E. J., and Van Eenennaam, A. L. (2007). Neonatal growth rate and development of mice raised on milk transgenically enriched with omega-3 fatty acids. Pediatr. Res. 62, 412–416.
Boot, A. M., Bouquet, J., Ridder, M. A. J. D., Krenning, E. P., and Keizer-Shrama, S. M. (1997). Determinants of body composition measured by dual-energy x-ray absorptiometry in Dutch children and adolescents. Am. J. Clin. Nutr. 66, 232–238.
Bourre, J. M., Francois, M., Youyou, A., Dumont, O., Piciotti, M., Pascal, G., and Durand, G. (1989). The effects of dietary alpha-linolenic acid on the composition of nerve membranes, enzymatic activity, amplitude of electrophysiological parameters, resistance to poisons and performance of learning tasks in rats. J. Nutr. 119, 1880–1892.
Brand, M. D., Couture, P., Else, P. L., Withers, K. W., and Hulbert, A. J. (1991). Evolution of energy metabolism. Proton permeability of the inner membrane of liver mitochondria is greater in a mammal than in a reptile. Biochem. J. 275, 81–86.
Brand, M. D., Couture, P., and Hulbert, A. J. (1994). Liposomes from mammalian liver mitochondria are more polyunsaturated and leakier to protons than those from reptiles. Comp. Biochem. Physiol. Biochem. Mol. Biol. 108, 181–188.
Braun, D. R., Harris, J. W. K., Levin, N. E., Mccoy, J. T., Herries, A. I. R., Bamford, M. K., Bishop, L. C., Richmond, B. G., and Kibunjia, M. (2010). Early hominin diet included diverse terrestrial and aquatic animals 1.95 Ma in East Turkana, Kenya. Proc. Natl. Acad. Sci. U.S.A. 107, 10002–10007.
Broadhurst, C. L., Wang, Y., Crawford, M. A., Cunnane, S. C., Parkington, J. E., and Schmidt, W. F. (2002). Brain specific lipids from marine, lacustrine, or terrestrial food resources: potential impact on early African Homo sapiens. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 131B, 653–673.
Calderon, F., and Kim, H.-Y. (2004). Docosahexaenoic acid promotes neurite growth in hippocampal neurons. J. Neurochem. 90, 979–988.
Cansev, M., and Wurtman, R. J. (2007). Chronic administration of docosahexaenoic acid or eicosapentaenoic acid, but not arachidonic acid, alone or in combination with uridine, increases brain phosphatide and synaptic protein levels in gerbils. Neuroscience 148, 421–431.
Cao, D., Kevala, K., Kim, J., Moon, H.-S., Jun, S. B., Lovinger, D., and Kim, H.-Y. (2009). Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J. Neurochem. 111, 510–521.
Chang, C.-Y., Ke, D.-S., and Chen, J.-Y. (2009). Essential fatty acids and human brain. Acta Neurol. Taiwan. 18, 231–241.
Chung, W. L., Chen, J. J., and Su, H. M. (2008). Fish oil supplementation of control and (n-3) fatty acid-deficient male rats enhances reference and working memory performance and increases brain regional docosahexaenoic acid levels. J. Nutr. 138, 1165–1171.
Clandinin, M. T., Chappell, J. E., Leong, S., Heim, T., Swyer, P. R., and Chance, G. W. (1980a). Extrauterine fatty acid accretion in infant brain: implications for fatty acid requirements. Early Hum. Dev. 4, 131–138.
Clandinin, M. T., Chappell, J. E., Leong, S., Heim, T., Swyer, R., and Chance, G. W. (1980b). Intrauterine fatty acid accretion rates in human brain: implications for fatty acid requirements. Early Hum. Dev. 4, 121–129.
Clutton-Brock, T. H., and Harvey, P. H. (1980). Primates, brains and ecology. J. Zool. (Lond.) 190, 309–323.
Coluccia, A., Borracci, P., Renna, G., Giustino, A., Latronico, T., Riccio, P., and Carratu, M. R. (2009). Developmental omega-3 supplementation improves motor skills in juvenile-adult rats. Int. J. Dev. Neurosci. 27, 599–605.
Coti Bertrand, P., O’usky, J. R., and Innis, S. M. (2006). Maternal dietary (n-3) fatty acid deficiency alters neurogenesis in the embryonic rat brain. J. Nutr. 136, 1570–1575.
Da Costa, K.-A., Rai, K. S., Craciunescu, C. N., Parikh, K., Mehedint, M. G., Sanders, L. M., Mclean-Pottinger, A., and Zeisel, S. H. (2009). Dietary docosahexaenoic acid supplementation modulates hippocampal development in the Pemt-/- mouse. J. Biol. Chem. 285, 1008–1015.
Dagai, L., Peri-Naor, R., and Birk, R. (2009). Docosahexaenoic acid significantly stimulates immediate early response genes and neurite outgrowth. Neurochem. Res. 34, 867–875.
Dalton, A., Wolmarans, P., Witthuhn, R. C., Van Stuijvenberg, M. E., Swanevelder, S. A., and Smuts, C. M. (2009). A randomised control trial in schoolchildren showed improvement in cognitive function after consuming a bread spread, containing fish flour from a marine source. Prostaglandins Leukot. Essent. Fatty Acids 80, 143–149.
De Groot, R. H., Hornstra, G., and Jolles, J. (2007). Exploratory study into the relation between plasma phospholipid fatty acid status and cognitive performance. Prostaglandins Leukot. Essent. Fatty Acids 76, 165–172.
Eilander, A., Hundscheid, D. C., Osendarp, S. J., Transler, C., and Zock, P. L. (2007). Effects of n-3 long chain polyunsaturated fatty acid supplementation on visual and cognitive development throughout childhood: a review of human studies. Prostaglandins Leukot. Essent. Fatty Acids 76, 189–203.
Emken, E. A., Adlof, R. O., and Gulley, R. M. (1994). Dietary linoleic acid influences desaturation and acylation of deuterium-labeled linoleic and linolenic acids in young males. Biochim. Biophys. Acta 1213, 277–288.
Fedorova, I., Hussein, N., Baumann, M. H., Di Martino, C., and Salem, N. Jr. (2009). An n-3 fatty acid deficiency impairs rat spatial learning in the Barnes maze. Behav. Neurosci. 123, 196–205.
Fontaneto, D., Tommaseo-Ponzetta, M., Galli, C., Risé, P., Glew, R. H., and Paoletti, M. G. (2011). Differences in fatty acid composition between aquatic and terrestrial insects used as food in human nutrition. Ecol. Food Nutr. 50, 351–367.
Garcia-Calatayud, S., Redondo, C., Martin, E., Ruiz, J. I., Garcia-Fuentes, M., and Sanjurjo, P. (2005). Brain docosahexaenoic acid status and learning in young rats submitted to dietary long-chain polyunsaturated fatty acid deficiency and supplementation limited to lactation. Pediatr. Res. 57, 719–723.
Gerrior, S., Bente, L., and Hiza, H. (2004). Nutrient content of the US food supply, 1909–2000 (Home Economics Res. Report Number 56). USDA Center for Nutrition Policy and Promotion, Alexandria.
Gibson, R. A., Muhlhausler, B., Makrides, M., Gibson, R. A., Muhlhausler, B., and Makrides, M. (2011). Conversion of linoleic acid and alpha-linolenic acid to long-chain polyunsaturated fatty acids (LCPUFAs), with a focus on pregnancy, lactation and the first 2 years of life. Matern. Child Nutr. 7(Suppl. 2), 17–26.
Greiner, R. S., Moriguchi, T., Hutton, A., Slotnick, B. M., and Salem, N. Jr. (1999). Rats with low levels of brain docosahexaenoic acid show impaired performance in olfactory-based and spatial learning tasks. Lipids 34(Suppl.), S239–S243.
Haggarty, P. (2004). Effect of placental function on fatty acid requirements during pregnancy. Eur. J. Clin. Nutr. 58, 1559–1570.
Harnack, K., Andersen, G., and Somoza1, V. (2009). Quantitation of alpha-linolenic acid elongation to eicosapentaenoic and docosahexaenoic acid as affected by the ratio of n6/n3 fatty acids. Nutr. Metab. (Lond.) 6, 8.
Harvey, P. H., Clutton-Brock, T. H., and Mace, G. M. (1980). Brain size and ecology in small mammals and primates. Proc. Natl. Acad. Sci. U.S.A. 77, 4387–4389.
He, C., Qu, X., Cui, L., Wang, J., and Kang, J. X. (2009). Improved spatial learning performance of fat-1 mice is associated with enhanced neurogenesis and neuritogenesis by docosahexaenoic acid. Proc. Natl. Acad. Sci. U.S.A. 106, 11370–11375.
Heinemann, K. M., and Bauer, J. E. (2006). Docosahexaenoic acid and neurologic development in animals. J. Am. Vet. Med. Assoc. 228, 700–705.
Hibbeln, J. R., Nieminen, L. R. G., Blasbalg, T. L., Riggs, J. A., and Lands, W. E. M. (2006). Healthy intakes of n-3 and n-6 fatty acids: estimations considering worldwide diversity. Am. J. Clin. Nutr. 83, 1483S–1493S.
Holguin, S., Huang, Y., Liu, J., and Wurtman, R. (2008). Chronic administration of DHA and UMP improves the impaired memory of environmentally impoverished rats. Behav. Brain Res. 191, 11–16.
Holman, R. T. (1998). The slow discovery of the importance of omega-3 essential fatty acids in human health. J. Nutr. 128, 427s–433s.
Hooijmans, C. R., Van Der Zee, C. E., Dederen, P. J., Brouwer, K. M., Reijmer, Y. D., Van Groen, T., Broersen, L. M., Lutjohann, D., Heerschap, A., Kiliaan, A. J., and Van Der Zee, C. E. E. M. (2009). DHA and cholesterol containing diets influence Alzheimer-like pathology, cognition and cerebral vasculature in APPswe/PS1dE9 mice. Neurobiol. Dis. 33, 482–498.
Innis, S. M. (2009). Omega-3 fatty acids and neural development to 2 years of age: do we know enough for dietary recommendations? J. Pediatr. Gastroenterol. Nutr. 48(Suppl. 1), S16–S24.
Innis, S. M., Vaghri, Z., and King, D. J. (2004). N-6 docosapentaenoic acid is not a predictor of low docosahexaenoic acid status in Canadian school children. Am. J. Clin. Nutr. 80, 768–773.
Issa, A. M., Moijica, W. A., Morton, S. C., Traina, S., Newberry, S. J., Hilton, L. G., Garland, R. H., and Maclean, C. H. (2006). The efficacy of omega-3 fatty acids on cognitive function in aging and dementia: a systematic review. Dement. Geriatr. Cogn. Disord. 21, 88–96.
Jamieson, E. C., Farquharson, J., Logan, R. W., Howatson, A. G., Patrick, W. J. A., Weaver, L. T., and Cockburn, F. (1999). Infant cerebellar gray and white matter fatty acids in relation to age and diet. Lipids 34, 1065–1071.
Javaux, E. J., Marshall, C. P., and Bekker, A. (2010). Organic-walled microfossils in 3.2-billion-year-old shallow-marine siliciclastic deposits. Nature 463, 934–938.
Jerde, R. S., Joel, P., Stroemer, J., Haight, R., and Joel, C. (1975). Comparison of nervous-tissue lipid fatty acid patterns of various animal species with particular reference to docosahexaenoic acid. Biochem. Soc. Trans. 3, 727–730.
Jiang, L.-H., Shi, Y., Wang, L.-S., and Yang, Z.-R. (2009). The influence of orally administered docosahexaenoic acid on cognitive ability in aged mice. J. Nutr. Biochem. 20, 735–741.
Kennedy, D. O., Jackson, P. A., Elliott, J. M., Scholey, A. B., Robertson, B. C., Greer, J., Tiplady, B., Buchanan, T., Haskell, C. F., Kennedy, D. O., Jackson, P. A., Elliott, J. M., Scholey, A. B., Robertson, B. C., Greer, J., Tiplady, B., Buchanan, T., and Haskell, C. F. (2009). Cognitive and mood effects of 8 weeks’ supplementation with 400 mg or 1000 mg of the omega-3 essential fatty acid docosahexaenoic acid (DHA) in healthy children aged 10–12 years. Nutr. Neurosci. 12, 48–56.
Kirby, A., Woodward, A., Jackson, S., Wang, Y., and Crawford, M. A. (2010). Childrens’ learning and behaviour and the association with cheek cell polyunsaturated fatty acid levels. Res. Dev. Disabil. 31, 731–742.
Kitajka, K., Puskas, L. G., Zvara, A., Hackler, L. H. Jr., Barcelo-Coblijn, G., Yoo, Y. K., and Farkas, T. (2002). The role of n-3 polyunsaturated fatty acids in brain: modulation of rat brain gene expression by dietary n-3 fatty acids. Proc. Natl. Acad. Sci. U.S.A. 99, 2619–2624.
Knutsen, S. F., Fraser, G. E., Beeson, W. L., Lindsted, K. D., and Shavlik, D. J. (2003). Comparison of adipose tissue fatty acids with dietary fatty acids as measured by 24-hour recall and food frequency questionnaire in black and white Adventists: the Adventist Health Study. Ann. Epidemiol. 13, 119–127.
Koletzko, B., Cetin, I., and Brenna, J. T. (2007). Dietary fat intakes for pregnant and lactating women. Br. J. Nutr. 98, 873–877.
Korn, E. L., and Graubard, B. I. (1991). Epidemiologic studies utilizing surveys: accounting for the sampling design. Am. J. Public Health 81, 1166–1173.
Kuipers, R. S., Luxwold, M. F., Dijck-Brouwer, D. A. J., Eaton, S. B., Crawford, M. A., Cordain, L., and Muskiet, F. A. J. (2010). Estimated macronutrient and fatty acid intakes from an East African Paleolithic diet. Br. J. Nutr. 104, 1666–1687.
Kumosani, T. A., Moselhy, S. S., Kumosani, T. A., and Moselhy, S. S. (2011). Modulatory effect of cod-liver oil on Na(+)-K(+) ATPase in rats’ brain. Hum. Exp. Toxicol. 30, 267–274.
Lassek, W. D., and Gaulin, S. J. C. (2006). Changes in body fat distribution in relation to parity in American women: a covert form of maternal depletion. Am. J. Phys. Anthropol. 131, 295–302.
Lassek, W. D., and Gaulin, S. J. C. (2007). Menarche is related to fat distribution. Am. J. Phys. Anthropol. 133, 1147–1151.
Lassek, W. D., and Gaulin, S. J. C. (2008). Waist-hip ratio and cognitive ability: is gluteofemoral fat a privileged store? Evol. Hum. Behav. 29, 26–34.
Lim, S.-Y., Hoshiba, J., Moriguchi, T., and Salem, N. Jr. (2005). N-3 fatty acid deficiency induced by a modified artificial rearing method leads to poorer performance in spatial learning tasks. Pediatr. Res. 58, 741–748.
Liu, J.-W., Almaguel, F. G., Bu, L., De Leon, D. D., and De Leon, M. (2008). Expression of E-FABP in PC12 cells increases neurite extension during differentiation: involvement of n-3 and n-6 fatty acids. J. Neurochem. 106, 2015–2029.
McCann, J. C., and Ames, B. N. (2005). Is docosahexaenoic acid, an n-3 long-chain polyunsaturated fatty acid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am. J. Clin. Nutr. 82, 281–295.
McGrew, W. C. (1979). “Evolutionary implications of sex differences in chimpanzee predation and tool use,” in The Great Apes, eds D. A. Hamburg, and E. R. McCown (London: Benjamin Cummings), 441–463.
Mitchell, T. W., Ekroos, K., Blanksby, S. J., Hulbert, A. J., and Else, P. L. (2007). Differences in membrane acyl phospholipid composition between an endothermic mammal and an ectothermic reptile are not limited to any phospholipid class. J. Exp. Biol. 210, 3440–3450.
Mukherjee, P. K., Chawla, A., Loayza, M. S., and Bazan, N. G. (2007). Docosanoids are multifunctional regulators of neural cell integrity and fate: significance in aging and disease. Prostaglandins Leukot. Essent. Fatty Acids 77, 233–238.
NCES. (2005). Highlights From the Trends in International Mathematics, and Science Study: TIMSS. (2003). Department of Education, National Center for Educational Statistics, 2005–2005, Washington, DC.
Needleman, H. L., and Landrigan, P. J. (2004). What level of lead in blood is toxic for a child? Am. J. Public Health 94, 8.
Neggers, Y. H., Kim, E.-K., Song, J.-M., Chung, E.-J., Um, Y.-S., and Park, T. (2009). Mental retardation is associated with plasma omega-3 fatty acid levels and the omega-3/omega-6 ratio in children. Asia Pac. J. Clin. Nutr. 18, 22–28.
Nichols, P. D., Danaher, K. T., and Koslow, J. A. (2003). Occurrence of high levels of tetracosahexaenoic acid in the jellyfish Aurelia sp. Lipids 11, 1207–1210.
Niemoller, T. D., Stark, D. T., and Bazan, N. G. (2009). Omega-3 fatty acid docosahexaenoic acid is the precursor of neuroprotectin D1 in the nervous system. World Rev. Nutr. Diet. 99, 46–54.
Novak, E. M., Dyer, R. A., and Innis, S. M. (2008). High dietary omega-6 fatty acids contribute to reduced docosahexaenoic acid in the developing brain and inhibit secondary neurite growth. Brain Res. 1237, 136–145.
Osendarp, S. J., Baghurst, K. I., Bryan, J., Calvaresi, E., Hughes, D., and Hussaini, M. (2007). Effect of a 12-mo micronutrient intervention on learning and memory in well-nourished and marginally nourished school-aged children: 2 parallel, randomized, placebo-controlled studies in Australia and Indonesia. Am. J. Clin. Nutr. 86, 1082–1093.
Ouellet, M., Emond, V., Chen, C., Julien, C., Bourasset, F., Oddo, S., Laferla, F., Bazinet, R. P., and Calon, F. (2009). Diffusion of docosahexaenoic and eicosapentaenoic acids through the blood-brain barrier: an in situ cerebral perfusion study. Neurochem. Int. 55, 476.
Pawlosky, R. J., Hibbeln, J. R., Novotny, J. A., and Salem, N. (2001). Physiological compartmental analysis of alpha-linolenic acid metabolism in adult humans. J. Lipid Res. 42, 1257–1265.
Pifferi, F., Jouin, M., Alessandri, J. M., Haedke, U., Roux, F., Perriere, N., Denis, I., Lavialle, M., and Guesnet, P. (2007). N-3 fatty acids modulate brain glucose transport in endothelial cells of the blood-brain barrier. Prostaglandins Leukot. Essent. Fatty Acids 77, 279–286.
Plomin, R., DeFries, J. C., McClearn, G. E., and McGuffin, P. (2000). Behavioral Genetics, 4th Edn. New York: Worth.
Pond, C. M., and Mattacks, C. A. (1987). The anatomy of adipose tissue in captive Macaca monkeys and its implications for human biology. Folia Primatol. (Basel) 48, 164–185.
Putnam, J., Allshouse, J., and Kantor, L. S. (2002). U.S. per capita food supply trends: more calories, refined carbohydrates, and fats. Food Rev. 25, 1–15.
Putnam, N. H., Srivastava, M., Hellsten, U., Dirks, B., Chapman, J., Salamov, A., Terry, A., Shapiro, H., Lindquist, E., Kapitonov, V. V., Jurka, J., Genikhovich, G., Grigoriev, I. V., Lucas, S. M., Steele, R. E., Finnerty, J. R., Technau, U., Martindale, M. Q., and Rokhsar, D. S. (2007). Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 317, 86–94.
Rapoport, S. I. (2003). In vivo approaches to quantifying and imaging brain arachidonic and docosahexaenoic acid metabolism. J. Pediatr. 143, S26–S34.
Redford, K. H., Bouchardet Da Fonseca, G. A., and Lacher, T. E. J. (1984). The relationship between frugivory and insectivory in primates. Primates 25, 433–440.
Richardson, A. J., and Montgomery, P. (2005). The Oxford-Durham Study: a randomized, controlled trial of dietary supplementation with fatty acids in children with developmental coordination disorder. Pediatrics 115, 1360–1366.
Rubin, D., and Laposata, M. (1992). Cellular interactions between n-6 and n-3 fatty acids: a mass analysis of fatty acid elongation/desaturation, distribution among complex lipids, and conversion to eicosanoids. J. Lipid Res. 33, 1431–1440.
Ryan, A. S., Astwood, J. D., Gautier, S., Kuratko, C. N., Nelson, E. B., and Salem, N. Jr. (2010). Effects of long-chain polyunsaturated fatty acid supplementation on neurodevelopment in childhood: a review of human studies. Prostaglandins Leukot. Essent. Fatty Acids 82, 305–314.
Ryan, A. S., and Nelson, E. B. (2008). Assessing the effect of docosahexaenoic acid on cognitive functions in healthy, preschool children: a randomized, placebo-controlled, double-blind study. Clin. Pediatr. (Phila) 47, 355–362.
Sakamoto, T., Cansev, M., and Wurtman, R. J. (2007). Oral supplementation with docosahexaenoic acid and uridine-5′-monophosphate increases dendritic spine density in adult gerbil hippocampus. Brain Res. 1182, 50–59.
Salem, N. Jr., Moriguchi, T., Greiner, R. S., Mcbride, K., Ahmad, A., Catalan, J. N., and Slotnick, B. (2001). Alterations in brain function after loss of docosahexaenoate due to dietary restriction of n-3 fatty acids. J. Mol. Neurosci. 16, 299–307.
Sauerwald, T. U., Demmelmair, H., Fidler, N., and Koletzko, B. (2000). Polyunsaturated fatty acid supply with human milk. Adv. Exp. Med. Biol. 478, 261–270.
Schuchardt, J. P., Huss, M., Stauss-Grabo, M., and Hahn, A. (2010). Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. Eur. J. Pediatr. 169, 149–164.
Sekula, W., Nelson, M., Figurska, K., Oltarzewski, M., Weisell, R., and Szponar, L. (2005). Comparison between household budget survey and 24-hour recall data in a nationally representative sample of Polish households. Public Health Nutr. 8, 430–439.
Shanker, K., Shireesha, K. K. S., Kumar, S. V., Srinivas, C., Rao, J. V., and Prasad, R. B. (2006). Isolation and characterization of neutral lipids of desilked eri silkworm pupae grown on castor and tapioca leaves. J. Agric. Food Chem. 2006, 3305–3309.
Shirai, N., and Suzuki, H. (2004). Effect of dietary docosahexaenoic acid and catechins on maze behavior in mice. Ann. Nutr. Metab. 48, 51–58.
Simopoulos, A. P., Leaf, A., and Salem, N. S. Jr. (1999). Workshop on the essentiality of and recommended dietary intakes for omega-6 and omega-3 fatty acids. J. Am. Coll. Nutr. 18, 487–489.
Slater, B., Enes, C. C., Lopez, R. V., Damasceno, N. R., Voci, S. M., Slater, B., Enes, C. C., Lopez, R. V. M., Damasceno, N. R. T., and Voci, S. M. (2010). Validation of a food frequency questionnaire to assess the consumption of carotenoids, fruits and vegetables among adolescents: the method of triads. Cad. Saude Publica 26, 2090–2100.
Stark, W. S., Lin, T. N., Brackhahn, D., Christianson, J. S., and Sun, G. Y. (1993). Fatty acids in the lipids of Drosophila heads: effects of visual mutants, carotenoid deprivation and dietary fatty acids. Lipids 28, 345–350.
Stewart, K. M. (2010). “The case for exploitation of wetlands by pre-sapiens hominins,” in Human Brain Evolution: The Influence of Freshwater and Marine Food Resources, eds S. C. Cunnane, and K. M. Stewart (Hoboken: John Wiley and Sons), 137–172.
Stoknes, I. S., Økland, H. M. W., Falch, E., and Synnes, M. (2004). Fatty acid and lipid class composition in eyes and brain from teleosts and elasmobranchs. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 138, 183–191.
Svennerholm, L. (1968). Distribution and fatty acid composition of phosphoglycerides in normal human brain. J. Lipid Res. 9, 570–579.
Takeuchi, T., Fukumoto, Y., and Harada, E. (2002). Influence of a dietary n-3 fatty acid deficiency on the cerebral catecholamine contents, EEG and learning ability in rat. Behav. Brain Res. 131, 193–203.
Turner, N., Else, P. L., and Hulbert, A. J. (2003). Docosahexaenoic acid (DHA) content of membranes determines molecular activity of the sodium pump: implications for disease states and metabolism. Naturwissenschaften 90, 521–523.
USDA. (2011). USDA National Nutrient Database for Standard Reference. Available at: http://www.ars.usda.gov/Services/docs.htm?docid=8964; see fatty acid composition of corn and pork, cow, and lamb brains
Wasserman, G. A., Liu, X., Lolacono, N. J., Factor-Litvak, P., Kline, J. K., Popovac, D., Morina, N., Musabegovic, A., Vrenezi, N., Capuni-Paracka, S., Lekic, V., Preteni-Redjepi, E., Hadzialjevic, S., Slavkovich, V., and Graziano, J. (1997). Lead exposure and intelligence in 7-year-old children: the Yugoslavia Prospective Study. Environ. Health Perspect. 105, 956–962.
Whalley, L. J., Fox, H. C., Wahle, K. W., Starr, J. M., and Deary, I. J. (2004). Cognitive aging, childhood intelligence, and the use of food supplements: possible involvement of n-3 fatty acids. Am. J. Clin. Nutr. 80, 1650–1657.
Wu, A., Ying, Z., and Gomez-Pinilla, F. (2008). Docosahexaenoic acid dietary supplementation enhances the effects of exercise on synaptic plasticity and cognition. Neuroscience 155, 751–759.
Wurtman, R. J., Cansev, M., and Ulus, I. H. (2009). Synapse formation is enhanced by oral administration of uridine and DHA, the circulating precursors of brain phosphatides. J. Nutr. Health Aging 13, 189–197.
Yoshida, S., Yasuda, A., Kawazato, H., Sakai, K., Shimada, T., Takeshita, M., Yuasa, S., Kobayashi, T., Watanabe, S., and Okuyama, H. (1997). Synaptic vesicle ultrastructural changes in the rat hippocampus induced by a combination of α-linolenate deficiency and a learning task. J. Neurochem. 68, 1261–1268.
Keywords: essential fatty acids, DHA, cognition, diet, brain, sex differences, evolution
Citation: Lassek WD and Gaulin SJC (2011) Sex differences in the relationship of dietary fatty acids to cognitive measures in American children. Front. Evol. Neurosci. 3:5. doi: 10.3389/fnevo.2011.00005
Received: 15 August 2011; Accepted: 14 October 2011;
Published online: 02 November 2011.
Edited by:
Melanie L. Shoup-Knox, University at Albany, USAReviewed by:
Paul M. Nealen, Indiana University of PA, USACopyright: © 2011 Lassek and Gaulin. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: William D. Lassek, Department of Epidemiology, Graduate School of Public Health, University of Pittsburgh, 527A Parran Hall, 130 DeSoto Street, Pittsburgh, PA 15261, USA. e-mail:d2lsbC5sYXNzZWtAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.