 Zhang Wanyi1†
Zhang Wanyi1† Jiang Lan
Jiang Lan- 1Department of Neurology, Chongqing Emergency Medical Center, Chongging University Central Hospital, Chongqing, China
- 2Department of Nursing, Chongqing Emergency Medical Center, Chongging University Central Hospital, Chongqing, China
- 3Outpatient Department, Chongqing Emergency Medical Center, Chongging University Central Hospital, Chongqing, China
- 4Department of Respiratory Medicine, Chongqing Emergency Medical Center, Chongging University Central Hospital, Chongqing, China
Parkinson’s disease (PD) and inflammatory bowel disease (IBD) are the two chronic inflammatory diseases that are increasingly affecting millions of people worldwide, posing a major challenge to public health. PD and IBD show similarities in epidemiology, genetics, immune response, and gut microbiota. Here, we review the pathophysiology of these two diseases, including genetic factors, immune system imbalance, changes in gut microbial composition, and the effects of microbial metabolites (especially short-chain fatty acids). We elaborate on the gut–brain axis, focusing on role of gut microbiota in the pathogenesis of PD and IBD. In addition, we discuss several therapeutic strategies, including drug therapy, fecal microbiota transplantation, and probiotic supplementation, and their potential benefits in regulating intestinal microecology and relieving disease symptoms. Our analysis will provide a new understanding and scientific basis for the development of more effective therapeutic strategies for these diseases.
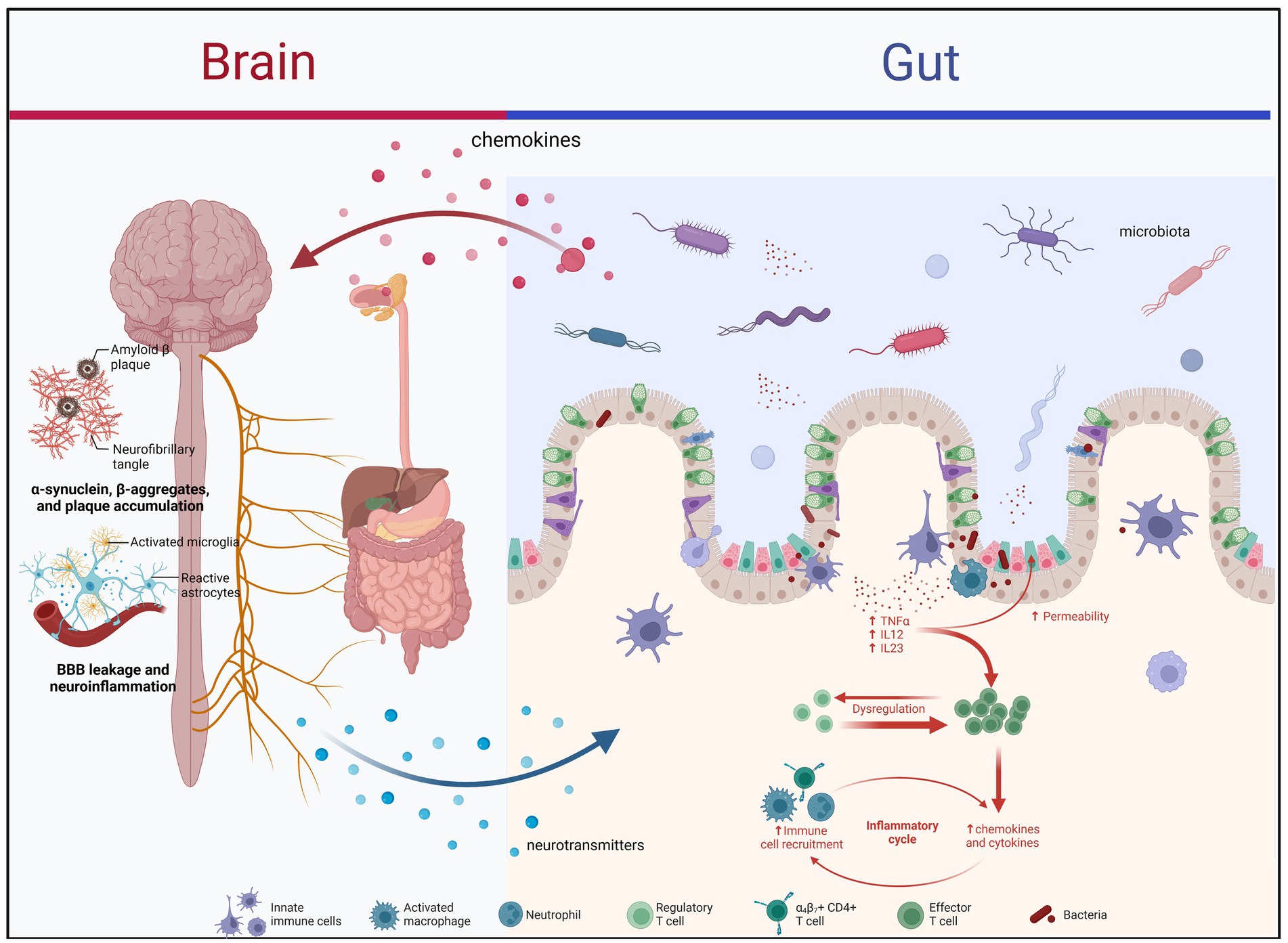
Graphical Abstract. PD and IBD mechanism of gut microbes.
1 Introduction
Parkinson’s disease (PD) and inflammatory bowel disease (IBD) are increasingly affecting the global population. PD is a neurodegenerative disease characterized by bradykinesia, resting tremor, and myotonia; its pathogenesis involves various factors, including genetics, environmental factors, and age (1, 2). IBD, including Crohn’s disease and ulcerative colitis, is a group of diseases characterized by chronic intestinal inflammation, leading to abdominal pain, diarrhea, and hematochezia. Its etiology is complex and diverse, involving immune system abnormalities, genetic susceptibility, and intestinal microbial imbalance (3–5). The occurrence of PD and IBD may be intricately linked to each other, and the interplay between the gut and the central nervous system—the “gut–brain axis”—provides a potential biological basis for this link (6–8). Meanwhile, some studies have shown that IBD is a candidate diagnostic marker for PD (9), and the incidence of PD is significantly increased in IBD patients (10). Here, we will review the risk factors, pathophysiological mechanisms, and treatment methods of PD and IBD, with special focus on the role of gut microbiota and short-chain fatty acids (SCFAs). We aim to provide comprehensive information on the relationship between these two seemingly different but closely related diseases for future research on diagnostic and treatment strategies.
2 Epidemiology of PD and IBD2.1 epidemiological relationship between PD and IBD
Parkinson’s disease is the second most common neurodegenerative disease worldwide, and its prevalence is gradually increasing with the aging of the global population. PD mainly affects adults over 50 years of age; however, cases of PD have also been reported in younger age groups. Males are slightly more likely to develop PD than females, and most cases are thought to be sporadic, although familial clusters have been reported in about 10% of patients (11, 12). The increase in the prevalence and incidence of IBD in industrialized and high-income countries can be attributed to changes in environmental factors, lifestyle, and genetic susceptibility. Although IBD can occur at any age, it is most common in adolescents and young adults, and its incidence is similar between men and women. However, family history is an important risk factor for IBD, and individuals with family history have a significantly increased risk of developing the disease (13–15).
2.1 Risk of PD in patients with IBD
A meta-analysis of nine studies involving 12,177,520 patients revealed that the incidence of PD in patients with IBD was higher than that in the general population (RR = 1.24; p < 0.001) (10). Further, the results of another meta-analysis confirmed that patients with IBD had a higher incidence of PD compared with the general population (OR = 1.30; p = 0.024) (16). However, it is unclear whether the risk of IBD in patients with PD is different from that in the general population. The findings of existing studies are inconsistent, and sufficient epidemiological studies are not available. A cohort study involving 1968 patients with PD and 6,792 controls was conducted in Taiwan in 2015. The authors found that the prevalence of IBD between the two groups was not significantly different (p = 0.561) (17). A Swedish case–control study involving 39,652 patients with IBD and 396,520 controls revealed that patients with IBD were more likely to have PD at the time of IBD diagnosis compared with the control population (OR = 1.4; 95% CI: 1.2–1.8). Similar results were observed in the subgroup analysis of Crohn’s disease (OR = 1.6; 95% CI: 1.1–2.3) and ulcerative colitis (OR = 1.4; 95% CI: 1.1–1.9) (18). The 2019 update of the World Movement Disorder Society diagnostic criteria for the prodromal stage of PD specifies that IBD is a candidate diagnostic marker for PD. This implies reliable and credible evidence; however, the corresponding prospective studies are lacking (9).
2.2 Risk of IBD in patients with PD
Freuer et al. analyzed 463,372 IBD-related datasets (7,045 cases and 456,327 controls) and 1,474,097 PD-related datasets (56,306 cases and 1,417,791 controls). Mendelian randomization (MR) analysis using inverse variance weighting (IVW) showed that IBD was not associated with the risk of PD (OR = 0.98, p = 0.48) (19). MR analysis was performed using the data obtained from 59,957 patients with IBD (25,042 cases and 34,915 controls) and 1,474,097 patients with PD-related using five statistical methods, including IVW and robust adjusted profile score (RAPS). The risk of IBD in patients with PD was higher than that in the control population (IVW and RAPS OR values were 1.062 and 1.063, respectively; both p < 0.05). The results of the remaining three statistical methods were negative (19). Two-way MR was used to analyze 214,053 patients with IBD (3,753 cases and 210,300 controls) and 482,730 patients with PD (33,674 cases and 449,056 controls). The authors observed the incidence of IBD in PD (OR = 1.014; 95% CI: 0.967–1.063; p = 0.573) and that of PD in IBD (OR = 0.978; 95% CI: 0.910–1.052; p = 0.549) (20). Currently, there is a paucity of definitive evidence to establish a direct association between PD and IBD; however, epidemiological correlations between the two disorders have been observed. Additional prospective and mechanistic studies are needed to clarify the possible causal relationship between these two diseases and the specific biological pathways involved in their interaction. This information may provide important clues for the development of new strategies for the prevention and treatment of both diseases.
3 Pathophysiology of PD and IBD
3.1 Genetic factors
Several authors have reported common genetic variants associated with the risk of PD and IBD (21). The NOD2/CARD15 gene may be a common risk gene for PD and IBD (22). This gene is located on chromosome 16 and encodes the NOD2 protein (23). Four single nucleotide polymorphisms (SNPs; R702W, G908R, L1007fs, and P268S) of the NOD2/CARD15 gene are highly expressed in patients with Crohn’s disease and PD (22, 24, 25). However, Appenzeller et al. suggested that the three SNPs (R702W, G908R, and L1007fs) are not associated with PD (26). The NOD2 protein encoded by this gene plays an important role in maintaining intestinal homeostasis. Any mutation in the NOD2 gene may increase the susceptibility to IBD in the corresponding population through nuclear factor-κb activation and cytokine response (27). The leucine-rich repeat kinase 2 (LRRK2) gene is located on chromosome 12 and encodes the LRRK2 protein (28). The LRRK2 gene is one of the important pathogenic genes in PD, and it is also related to IBD (29). The LRRK2 gene is highly expressed in peripheral blood mononuclear cells and may be involved in the inflammatory process. The expression of the LRRK2 gene in B cells, T cells, and CD16+ monocytes was higher in patients with PD than that in healthy controls. The interferon-γ stimulation can increase the expression of this gene in the immune cells of patients with Crohn’s disease (30, 31). Several SNP sites in the LRRK2 gene, including N1437H, R1441C/G/H, Y1699C, I2012T, G2019S, and I2020T, are the pathogenic mutation sites in PD, and M2397T is a risk site in Crohn’s disease. N2081D is the common risk locus of PD and Crohn’s disease, whereas N551K and R1398H are the common protective loci of PD and Crohn’s disease (29). N2081D is located in the kinase domain of the LRRK2 gene and is associated with increased kinase activity of the LRRK2 protein. R1398H is located in the Roc (Ras/GTPase-protein complex) domain of the LRRK2 gene, and mutations in this site can inactivate the LRRK2 protein by increasing GTPase activity. N551K is not in any domain of the LRRK2 gene, but N551K and R1398H show linkage disequilibrium (32, 33). Chronic neuroinflammation and intestinal inflammation are important pathophysiological processes in PD and IBD, respectively. Therefore, LRRK2 gene mutations may mediate inflammatory responses by affecting the kinase and GTPase activities of the LRRK2 protein, thereby participating in the pathogenesis of these diseases (29). In addition to the LRRK2 and NOD2 genes, other genes associated with autophagy, such as ATG16L1 (34, 35) and IRGM (36), are also associated with the pathology of IBD and may be involved in the pathogenesis of PD. These findings support the idea of abnormal autophagy as a shared pathophysiological feature of PD and IBD. Although these genetic findings provide valuable insights, the exact genetic link between PD and IBD remains a complex issue that requires further investigation. A better understanding of the genetic basis of these disorders may facilitate the development of therapeutic strategies targeting shared mechanisms, thereby providing patients with better treatment options and outcomes.
3.2 Immunomodulatory mechanisms
Chronic non-specific inflammation is often accompanied by structural and functional disorders of the gastrointestinal mucosal barrier. Crohn’s disease can affect any layers of mucosa from mouth to anus, whereas ulcerative colitis usually affects the lining of the colonic epithelium (37). C-reactive protein in the blood of patients with IBD is a reliable biomarker reflecting the severity of the disease. Atreya and Neurath observed an increase in the levels of tumor necrosis factor (TNF)-α and other cytokines in the gastrointestinal tract (38) and those of inflammation-related proteins, such as calprotectin, calgranulin C (also known as S100A12), and lactoferrin, in the feces of patients with IBD (39). Neuroinflammation is one of the important pathophysiological features of PD (40), and typical inflammation occurs in the gastrointestinal tract of patients with PD. The mRNA levels of TNF-α, interferons, interleukin (IL)-6, and IL-1β increase in the colon tissue of patients with PD. In addition, the levels of IL-1β, C-reactive protein, and calprotectin increase in the feces of these patients (41). α-Synuclein, an unfolded protein composed of 140 amino acid residues, is widely expressed in the human brain, especially in the synaptic terminals of neurons (42). This protein abnormally aggregates and forms fibrous structures called Lewy bodies in the brains of patients with PD; these structures are one of the most prominent pathological hallmarks of PD (43). Intestinal inflammation may cause brain inflammation through secondary systemic inflammatory response and eventually promote the abnormal accumulation of α-synuclein in the brain to induce PD (44). Immunohistochemical analysis of colon tissues from 8 patients with IBD (4 cases of Crohn’s disease and 4 cases of ulcerative colitis) and 4 controls showed that the level of α-synuclein in the non-inflammatory area of Crohn’s disease was 2.07 times higher that of the control group, and the level of α-synuclein in the inflammatory area was 2.35 times higher that of the control group (45). Kishimoto et al. fed drinking water containing 0.5% dextran sodium sulfate to A53T gene-mutant mice for inducing colitis. The results showed that the experimental group had earlier movement disorders, abnormal accumulation of α-synuclein and degeneration of dopaminergic neurons compared with the control mice (46). Abnormal accumulation of Lewy bodies in the enteric nervous system (ENS) has been detected in the early stage of PD, and ENS dysfunction may promote the development of gastrointestinal symptoms in PD patients (47). The ENS is the origin and entrance of pathological changes in PD, and spreads to the central nervous system through vagus nerve transmission, leading to further substantia nigra lesions. Enteric glial cells (EGCs), as the most abundant cells of ENS, are closely related to the intestinal microbiota (48) and respond to microbial invasion. Related studies have found that bacterial lipopolysaccharide (LPS) and IL-6 can activate EGC by binding to PRR on the membrane of EGC cells, trigger TLR4/NF-κB and other proinflammatory signaling pathways and the formation of NLRP3 inflammasome, and promote intestinal immune inflammation to clear pathogens (49, 50). EGC can exert immunosuppressive and anti-inflammatory effects by releasing GDNF and BDNF. After the release of GDNF, it can bind to RET on type 3 lymphocytes (ILC3), thereby activating ILC3 and promoting the release of anti-inflammatory factor IL-22 and the expression of repair genes in intestinal epithelial cells, thus protecting the inflammatory epithelium of colon (51). The released BDNF reduced the expression of nitric oxide synthase and pro-inflammatory factor IL-6 induced by LPS in mice by down-regulating the TLR4 receptor on EGC, and alleviated intestinal inflammation (52). Drokhlyansky et al. (53) applied single-cell sequencing to the analysis of human and mouse ENS and found that genes expressed in the intermuscular and mucosal EGCs were significantly different, and found that several PD risk genes were enriched in the ENS, among which NRXN1 and ANK2 were enriched in the EGC, indicating that the dysfunction of EGC may aggravate CNS disease. EGC reactive hyperplasia and its specific glial markers are found in the colon tissues of PD patients, and they appear in the early stage of PD (54). Therefore, EGC obtained by gastrointestinal mucosal biopsy and analyzed may be superior to α-syn in predicting early PD. In the latest study by Perez-Pardo et al. (55) immunofluorescence staining of fixed sections of the colon of dead mice also found that the expression of EGC-derived glial fibrillary acidic protein (GFAP) and α-syn was increased simultaneously, suggesting that EGC may also play a role in the pathological formation of intestinal α-syn. It has also been shown that α-syn can ascend to the central nervous system via the enteric glia Cx43 hemicchannel (through which glial-glial syncytial cells are connected as a pathway for intercellular communication between the gastrointestinal tract and the central nervous system) or the vagus nerve (56). It was further found that EGC also plays a role in the ascent of α-syn to the central nervous system. EGC in PD may be pathological activated, which may promote α-syn misfolding in ENS by participating in intestinal immune inflammation and help α-syn spread to the brain. In turn, α-syn may also act as an effector molecule to further promote the pathological activation of EGC. Therefore, chronic inflammation links these two diseases, providing potential targets for future therapeutic strategies. Future studies should focus on the specific mechanisms linking IBD and PD, especially the role of α-synuclein. In addition, strategies to intervene in intestinal inflammation should be explored to reduce the risk of PD or delay its progression.
3.3 Brain–gut axis: gut microbes and SCFAs
The “gut–brain axis” theory is based on experimental evidence indicating the link between the gut environment and the central nervous system. The theory proposes a connection between the emotional and cognitive centers of the brain with peripheral gut functions (57). The disruption of gut microbiota is closely related to autism, neurodegenerative diseases and emotional disorders (stress, depression, anxiety) (58). The composition of gut microbiota is affected by diet and environment, and the use of antibiotics is one of the important reasons for destroying the stability of gut microbiota (59). A rodent study showed that low-dose penicillin administered late during pregnancy and early after birth had long-term effects on mouse offspring, including altered gut microbiome composition, increased cytokine expression in the frontal cortex, altered blood–brain barrier integrity, and behavioral measures, with the mice showing anxitty-like behavior (60). Through metagenomic sequencing, Yang et al. found that a variety of phages and bacteria in the gut of patients with major depression were changed, among which the reduced abundance of Blautia and Eubacterium was significantly associated with depressive symptoms (61). In addition, a meta-analysis showed that Bacteroides, Paranobacillus, and Barnesiella were enriched in patients with depression, while Firmicutes, Spirospiraceae (UCG 003, UCG 002), and Bacteroides vulgaris were significantly depleted (62). More important, Kelly et al. (63) found that transplantation of “depressive microbiota” into germ-free mice induced depressive-like behaviors and features, including anhedonia and states of hopelessness. Therefore, by understanding the bidirectional communication system of the gut-brain axis, we can gain deeper insight into how changes in the gut microbiota affect brain function, which in turn affects individual emotional and behavioral performance. According to this theory, PD may be a consequence of intestinal dysbiosis or intestinal barrier dysfunction or both, which is caused by an unknown pathogen in the gastrointestinal tract. The main pathological manifestation of PD is Lewy body (LB), which is caused by the misfolding and aggregation of α-synuclein (α-syn) (64). Many studies have verified the “gut-brain axis” hypothesis that α-syn can spread from the gastrointestinal tract to the brain through the vagus nerve (65, 66). Kim and colleagues found that α-syn injected in the duodenum and pylorus of mice migrated through the vagus nerve to the substantia nigra, locus locus locus, olfactory bulb, cerebellum, and other brain regions to accumulate and precipitate, causing PD-related motor disorders and non-motor symptoms (65). A recent study (67) found that α-syn can promote the transmission of each other from the gut to the brain by interacting with Tau protein, triggering the loss of substantia nigra dopaminergic neurons. Furthermore, some researchers have used glucose probes to study the intestinal permeability of PD patients and found that the intestinal epithelial barrier in PD patients has similar dysfunction as that in patients with enteritis (68). A study using baboons as a model, published in Brain, found that α-syn not only travels from the gut to the brain, but also travels backwards (69). In recent years, studies have continuously revealed that psychological factors play an important role in the course of organic diseases (such as IBD) through the role of brain-gut axis (70). Psychological factors aggravate IBD by increasing intestinal permeability, changing intestinal flora and enhancing immune response mediated by brain-gut axis (71). Intestinal inflammation can cause psychological diseases. In recent years, animal studies have found that intestinal inflammation in mice with colitis can lead to increased serum C-reactive protein and cortisol levels, and lead to inflammation represented by increased cyclooxygenase-2 levels in the limbic system of the brain through the hypothalamic–pituitary–adrenal axis (HPA). Heightened reactivity and decreased brain-derived neurotrophic factor (BDNF), which is thought to be directly linked to psychological disorders such as anxiety (72). Therefore, pathological α-syn caused by intestinal barrier dysfunction moves from the intestine to the brain and induces PD, and PD patients are often accompanied by different degrees of gastrointestinal symptoms, which needs more basic and clinical research evidence to confirm.
PD and IBD are characterized by intestinal microbial dysbiosis (73, 74). The proportion of pro-inflammatory bacteria, such as Proteobacteria, increases, whereas the abundance of some beneficial bacteria (SCFA producers) decreases in the gut of patients with PD. This imbalance of microbiota may lead to impaired intestinal barrier function and increased intestinal permeability, allowing more pathogens and inflammatory molecules to enter the blood circulation. Ultimately, this affects brain function, promotes neuroinflammation, and induces abnormal aggregation of α-synuclein in PD (73). A decline in the production of SCFAs by bacteria, such as Faecalibacterium and Roseburia, in patients with IBD, which are essential for maintaining the health of the intestinal mucosa and suppressing inflammatory responses (75). The abundance of pro-inflammatory bacteria of the Enterobacteriaceae family is increased in patients with PD and IBD. The abundance of SCFA-producing bacteria, such as Prevotellaceae (Bacteroidota), Lachnospiraceae (including Roseburia; Firmicutes), and Faecalibacterium is decreased in these patients. While Verrucomicrobia, Verrucomicrobiaceae, the abundance of anti-inflammatory bacteria, including Akkermansia, Lactobacillaceae, and Actinobacteria (including Bifidobacterium) was heterogeneous. Anti-inflammatory bacteria, such as Akkermansia, Lactobacillaceae, and Bifidobacterium, can grow in an inflammatory environment, and their abundance increases later than the “intestinal inflammation” process of PD and IBD (76–78). Lactobacillus and Bifidobacterium can modulate the host’s immune response, enhancing gut health by improving mucosal barrier function and reducing inflammation. This is partly achieved through the production of short-chain fatty acids (SCFAs) like acetate, propionate, and butyrate, which have anti-inflammatory properties. These bacteria contribute to the strengthening of the gut barrier, preventing the translocation of harmful bacteria and endotoxins into the host’s circulatory system. This barrier function is crucial for preventing infections and maintaining immune homeostasis (79, 80).
Metabolites of gut microbiota, such as SCFAs, show similar changes in patients with PD and IBD. SCFAs are a group of saturated fatty acids with carbon atom number ≤ 6, including acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, isovaleric acid, caproic acid, and isocaproic acid. They are mainly produced by intestinal microorganisms in the colon by fermentation of the dietary fiber (81). Mechanistic studies in animal models have shown that butyrate has beneficial effects in maintaining the integrity of the gastrointestinal mucosal barrier, quenching oxygen at the epithelial interface and acting as an immunomodulatory agent. Propionate has been reported to induce satiation by regulating the production of anorexigenic hormones and intestinal gluconeogenesis, while also affecting glucose metabolism. Butyrate has been suggested to be associated with anti-cancer and anti-inflammatory effects, but direct evidence for this is lacking (82). In addition, it has been found that the use of butyrate in animal models of Parkinson’s disease can improve dyskinesia and dopamine deficiency (83), while propionate seems to be negatively correlated with the Unified Parkinson’s Disease Rating Scale III (84). Shin et al. detected SCFAs in PD patients and found that the concentrations of acetic acid, propionic acid and butyric acid in feces decreased, while the concentrations of acetic acid and propionic acid in plasma increased in PD patients. The severity of the disease was negatively correlated with the concentrations of SCFAs in feces (except propionic acid), and positively correlated with the concentrations of acetic acid, propionic acid and valeric acid in plasma (84). SCFAs are involved in the occurrence of PD by affecting the integrity of the blood–brain barrier, the function of microglia, neuronal autophagy and apoptosis, the integrity of the intestinal barrier, and intestinal inflammation (85). SCFAs play an immunomodulatory role in IBD by participating in regulating the differentiation of innate and adaptive immune cells and the function of related cells (86). Chen et al. reported that SCFA concentration decreases in the feces of patients with PD, whereas it increases in blood, urine, and saliva. This phenomenon may be related to the effect of SCFAs on intestinal mucosal permeability (87). A meta-analysis of 12 studies involving 572 patients with IBD and 282 healthy controls showed that fecal concentrations of acetic acid, propionic acid, butyric acid, and valeric acid decreased in patients with IBD. However, subgroup analysis showed that the changes in fecal SCFAs in patients with Crohn’s disease were different from those in patients with ulcerative colitis. Acetic acid, valeric acid, and total SCFAs showed a downward trend in patients with ulcerative colitis, whereas acetic acid, butyric acid, and valeric acid showed a downward trend in patients with Crohn’s disease. In addition, subgroup analysis found that the concentration of butyrate in patients with ulcerative colitis changed at different disease stages; it was lower than that in healthy controls in the active stage and higher in the remission stage (88). Future research should focus on the specific role of gut microbiota and their metabolites in the pathogenesis of PD and IBD. Therapeutic strategies should target regulating the composition of gut microbiota and increasing the production of beneficial SCFAs. The researchers should focus on the specific effects of SCFAs on intestinal mucosal permeability and central nervous system inflammation.
4 Treatment of PD and IBD
4.1 Medicine
Commonly used drugs for the treatment of IBD include non-biological and biological agents. Classical non-biological agents include aminosalicylic acid, thiopurines, and hormones. Biological agents include anti-TNF, interleukin, and other cytokines, and drugs acting on specific inflammation-related pathways (89–91). A 2023 meta-analysis of six studies with data on the use of medications for IBD showed a protective effect of medications for IBD on the onset of PD (RR = 0.88) (92). 5-aminosalicylic acid (5-ASA) and anti-TNF drugs are commonly used for the treatment of IBD. In a cross-sectional study of 144,018 patients with IBD, the risk of PD was lower in those who received anti-TNF drugs than in those who did not (IRR = 0.22; p = 0.03) (93). Ríos et al. conducted a study on 20,208,682 patients and found that people under 65 years of age were less likely to receive anti-PD medication while using 5-ASA than those not using 5-ASA (OR = 0.28; p = 0.0103) (94). The above findings support the idea that drugs for IBD may indirectly slow the course of PD or reduce the risk of its development by reducing the inflammatory response. The studies on the specific mechanisms underlying the protective effects of IBD drugs on PD, including clinical trials to validate the potential utility of these drugs in patients with PD, may lead to more promising treatment prospects for patients with IBD and PD. The literature suggests that treatment with L-dopa-carbidopa enteric-coated gel (LCIG), which is a commonly used treatment for advanced PD, may indirectly help improve the prevalent GI symptoms in PD patients. Continuous infusion of LCIG is designed to minimize fluctuations in plasma drug concentrations, which not only optimizes motor symptom control but may also help stabilize GI function by providing more stable dopamine stimulation (95). MR Analysis of PD and IBD suggested that the CXCR4 gene is a potential drug target. The gene encodes the chemokine receptor CXCR4, and flavonoids may become potential therapeutic drugs for PD and IBD by inhibiting the CXCR4 protein (96). Therefore, new therapies for PD and IBD can be developed by finding the gene targets of drugs.
4.2 Fecal microbiota transplantation and probiotic treatment
FMT and probiotic therapy are two novel approaches for the treatment of gut-related diseases. Both these approaches have been evaluated for the treatment of PD and IBD. These two approaches modulate the gut microbiome and show potential therapeutic value in regulating intestinal inflammation and gut–brain axis interactions. FMT can ameliorate the intestinal microbial imbalance in the mouse model of PD, increase the levels of striatal dopamine and 5-hydroxytryptamine, and play a neuroprotective role by inhibiting neuroinflammation (97). FMT can ameliorate motor symptoms (e.g., tremors and bradykinesia) and non-motor symptoms (such as constipation, anxiety, depression, and sleep disorders) of patients with PD to a certain extent (98, 99). However, FMT is considered a controversial treatment for patients with IBD. Although FMT helps relieve the symptoms of patients with IBD in some small clinical studies, patients become prone to adverse reactions, such as infection and fever (100, 101). In contrast, probiotics are more clinically useful in adjusting intestinal microecology.
According to the definition of the World Health Organization and the Food and Agriculture Organization of the United Nations, probiotics are a group of living microorganisms (102) that can provide health benefits to the host when ingested in appropriate amounts. Probiotics can live and reproduce in the intestinal tract, and have a variety of functions, including maintaining intestinal health, enhancing immunity, promoting nutrient absorption, and alleviating gastrointestinal symptoms. Among them, probiotics perform well in the maintenance of intestinal health. It maintains intestinal health by inhibiting the growth of harmful bacteria, increasing the stability of intestinal mucosal barrier, and promoting intestinal peristalsis (103). Probiotics have recognized antioxidant, anti-inflammatory, and neuroprotective effects, which can regulate central nervous system activity by targeting a variety of cellular and molecular processes, such as oxidative stress, inflammatory and anti-inflammatory pathways, and apoptosis (104). Sun et al. (105) showed that probiotic lactis Probio-M8 synergeted with traditional drug treatment regimens for Parkinson’s disease to enhance the clinical efficacy of PD treatment, while changing the host’s gut microbiome, gut microbial metabolic potential, and serum metabolites. Zhao et al. (106) showed that rotenone-induced PD mouse model damaged the intestinal barrier, leading to the leakage of pathogenic LPS and LBP, which activated the SN and TLR4 signaling pathway in the colon. Fecal microbiota transplantation intervention could protect rotenone-induced PD mouse model by improving the imbalance of intestinal microbiome. Inhibition of the LPS-TLR4 signaling pathway in the gut and brain may play an important role. One study found that long-term use of probiotics produced marked neuroprotective effects on dopaminergic neurons and improved motor deficits in a mouse model of genetic PD (107). The probiotic E. coli Nissle1917 is as effective as standard 5-ASA therapy in slowing the progression of ulcerative colitis (108). Daily supplementation of probiotics improved gait, balance function, and motor coordination in a mouse model of PD, and long-term administration of probiotics has a neuroprotective effect on dopamine neurons (109). The results of randomized double-blind controlled trials have indicated that probiotics can help relieve non-motor symptoms, such as constipation, abdominal pain, and bloating, as well as the total Unified Parkinson’s Disease Rating Scale score in patients with PD (110). Although FMT is controversial in the treatment of IBD and may be associated with some risks, it has shown the possibility of alleviating motor and non-motor symptoms in patients with PD. Probiotic therapy, a safer and more clinically accessible treatment, has shown promise in slowing the progression of ulcerative colitis and protecting dopamine neurons in PD models. Future studies are needed to explore the mechanisms of action of FMT and probiotic therapy in the treatment of PD and IBD, especially how they modulate the intestinal microbial community for optimal therapeutic effects. In addition, more clinical trials are necessary to evaluate the safety, efficacy, and long-term effects of these treatments, especially in patients with PD and IBD at different stages and types.
5 Conclusion and prospects
The seemingly different diseases, PD and IBD, have many similarities in epidemiological characteristics, pathophysiological mechanisms, and treatment approaches. Therapeutic strategies for IBD, such as anti-inflammatory and immunomodulatory approaches, are potentially protective against PD progression. In addition, modulation of gut microbiota, including interventions through diet, probiotics, or emerging drug targets such as CXCR4, can be used for the treatment of both diseases. The assessment of PD and IBD should adopt a holistic approach, investigating the interrelationship between these two conditions, and deciphering how their interactions influence disease onset and progression. Future research endeavors should focus on elucidating the precise mechanisms underpinning these associations and developing innovative therapeutic approaches to enhance the quality of life for individuals afflicted with these disorders.
Author contributions
ZW: Conceptualization, Data curation, Formal analysis, Investigation, Validation, Writing – original draft, Writing – review & editing. YJ: Conceptualization, Data curation, Formal analysis, Validation, Writing – original draft, Writing – review & editing. HW: Conceptualization, Formal analysis, Validation, Writing – review & editing. XB: Data curation, Validation, Writing – original draft. WX: Data curation, Validation, Writing – review & editing. JL: Conceptualization, Data curation, Formal analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing. ZL: Conceptualization, Data curation, Formal analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Balestrino, R, and Schapira, AHV. Parkinson disease. Eur J Neurol. (2020) 27:27–42. doi: 10.1111/ene.14108
2. Schapira, AHV, Chaudhuri, KR, and Jenner, P. Non-motor features of Parkinson disease. Nat Rev Neurosci. (2017) 18:435–50. doi: 10.1038/nrn.2017.62
3. Flynn, S, and Eisenstein, S. Inflammatory bowel disease presentation and diagnosis. Surg Clin North Am. (2019) 99:1051–62. doi: 10.1016/j.suc.2019.08.001
4. Rogler, G, Singh, A, Kavanaugh, A, and Rubin, DT. Extraintestinal manifestations of inflammatory bowel disease: current concepts, treatment, and implications for disease management. Gastroenterology. (2021) 161:1118–32. doi: 10.1053/j.gastro.2021.07.042
5. Ferro, JM, and Oliveira, SM. Neurology of inflammatory bowel disease. J Neurol Sci. (2021) 424:117426. doi: 10.1016/j.jns.2021.117426
6. Lee, HS, Lobbestael, E, Vermeire, S, Sabino, J, and Cleynen, I. Inflammatory bowel disease and Parkinson's disease: common pathophysiological links. Gut. (2021) 70:408–17. doi: 10.1136/gutjnl-2020-322429
7. Brudek, T . Inflammatory bowel diseases and Parkinson's disease. J Parkinsons Dis. (2019) 9:S331–44. doi: 10.3233/JPD-191729
8. Li, Y, Chen, Y, Jiang, L, Zhang, J, Tong, X, Chen, D, et al. Intestinal inflammation and Parkinson's disease. Aging Dis. (2021) 12:2052–68. doi: 10.14336/AD.2021.0418
9. Heinzel, S, Berg, D, Gasser, T, Chen, H, Yao, C, Postuma, RB, et al. Update of the Mds research criteria for prodromal Parkinson's disease. Mov Disord. (2019) 34:1464–70. doi: 10.1002/mds.27802
10. Zhu, Y, Yuan, M, Liu, Y, Yang, F, Chen, WZ, Xu, ZZ, et al. Association between inflammatory bowel diseases and Parkinson's disease: systematic review and meta-analysis. Neural Regen Res. (2022) 17:344–53. doi: 10.4103/1673-5374.317981
11. Global, Regional, and National Burden Of Parkinson's Disease . 1990-2016: a systematic analysis for the global burden of disease study 2016. Lancet Neurol. (2018) 17:939–53.
12. Ahmad, MH, Rizvi, MA, Ali, M, and Mondal, AC. Neurobiology of depression in Parkinson's disease: insights into epidemiology, molecular mechanisms and treatment strategies. Ageing Res Rev. (2023) 85:101840. doi: 10.1016/j.arr.2022.101840
13. Agrawal, M, and Jess, T. Implications of the changing epidemiology of inflammatory bowel disease in a changing world. United European Gastroenterol J. (2022) 10:1113–20. doi: 10.1002/ueg2.12317
14. Kuenzig, ME, Fung, SG, Marderfeld, L, Mak, JWY, Kaplan, GG, Ng, SC, et al. Twenty-first century trends in the global epidemiology of pediatric-onset inflammatory bowel disease: systematic review. Gastroenterology. (2022) 162:1147–1159.e4. doi: 10.1053/j.gastro.2021.12.282
15. Kaplan, GG, and Windsor, JW. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. (2021) 18:56–66. doi: 10.1038/s41575-020-00360-x
16. Li, M, Wan, J, Xu, Z, and Tang, B. The association between Parkinson's disease and autoimmune diseases: a systematic review and Meta-analysis. Front Immunol. (2023) 14:1103053. doi: 10.3389/fimmu.2023.1103053
17. Hsu, YT, Liao, CC, Chang, SN, Yang, YW, Tsai, CH, Chen, TL, et al. Increased risk of depression in patients with Parkinson disease: a Nationwide cohort study. Am J Geriatr Psychiatry. (2015) 23:934–40. doi: 10.1016/j.jagp.2014.10.011
18. Weimers, P, Halfvarson, J, Sachs, MC, Saunders-Pullman, R, Ludvigsson, JF, Peter, I, et al. Inflammatory bowel disease and Parkinson's disease: a Nationwide Swedish cohort study. Inflamm Bowel Dis. (2019) 25:111–23. doi: 10.1093/ibd/izy190
19. Freuer, D, and Meisinger, C. Association between inflammatory bowel disease and Parkinson's disease: a Mendelian randomization study. NPJ Parkinsons Dis. (2022) 8:55. doi: 10.1038/s41531-022-00318-7
20. Zeng, R, Wang, J, Zheng, C, Jiang, R, Tong, S, Wu, H, et al. Lack of causal associations of inflammatory bowel disease with Parkinson's disease and other neurodegenerative disorders. Mov Disord. (2023) 38:1082–8. doi: 10.1002/mds.29386
21. Gonzalez-Latapi, P, and Marras, C. Epidemiological evidence for An immune component of Parkinson's disease. J Parkinsons Dis. (2022) 12:S29–43. doi: 10.3233/JPD-223180
22. Bialecka, M, Kurzawski, M, Klodowska-Duda, G, Opala, G, Juzwiak, S, Kurzawski, G, et al. Card15 variants in patients with sporadic Parkinson's disease. Neurosci Res. (2007) 57:473–6. doi: 10.1016/j.neures.2006.11.012
23. McGovern, D, van Heel, D, Ahmad, T, and Jewell, DP. Nod2 (Card15), the first susceptibility gene for Crohn's disease. Gut. (2001) 49:752–4. doi: 10.1136/gut.49.6.752
24. Lv, C, Yang, X, Zhang, Y, Zhao, X, Chen, Z, Long, J, et al. Confirmation of three inflammatory bowel disease susceptibility loci in a Chinese cohort. Int J Color Dis. (2012) 27:1465–72. doi: 10.1007/s00384-012-1450-6
25. Ma, Q, An, X, Li, Z, Zhang, H, Huang, W, Cai, L, et al. P268s in Nod2 associates with susceptibility to Parkinson's disease in Chinese population. Behav Brain Funct. (2013) 9:19. doi: 10.1186/1744-9081-9-19
26. Appenzeller, S, Thier, S, Papengut, F, Klein, C, Hagenah, J, Kasten, M, et al. No association between Nod2 variants and Parkinson's disease. Mov Disord. (2012) 27:1191–2. doi: 10.1002/mds.25059
27. Trindade, BC, and Chen, GY. Nod1 and Nod2 in inflammatory and infectious diseases. Immunol Rev. (2020) 297:139–61. doi: 10.1111/imr.12902
28. Lewis, PA . Leucine rich repeat kinase 2: a paradigm for pleiotropy. J Physiol. (2019) 597:3511–21. doi: 10.1113/JP276163
29. Herrick, MK, and Tansey, MG. Is Lrrk2 the missing link between inflammatory bowel disease and Parkinson's disease? NPJ Parkinsons Dis. (2021) 7:26. doi: 10.1038/s41531-021-00170-1
30. Cook, DA, Kannarkat, GT, Cintron, AF, Butkovich, LM, Fraser, KB, Chang, J, et al. Lrrk2 levels in immune cells are increased in Parkinson's disease. NPJ Parkinsons Dis. (2017) 3:11. doi: 10.1038/s41531-017-0010-8
31. Gardet, A, Benita, Y, Li, C, Sands, BE, Ballester, I, Stevens, C, et al. Lrrk2 is involved in the Ifn-gamma response and host response to pathogens. J Immunol. (2010) 185:5577–85. doi: 10.4049/jimmunol.1000548
32. Hui, KY, Fernandez-Hernandez, H, Hu, J, Schaffner, A, Pankratz, N, Hsu, NY, et al. Functional variants in the Lrrk2 gene confer shared effects on risk for Crohn's disease and Parkinson's disease. Sci Transl Med. (2018) 10:eaai7795. doi: 10.1126/scitranslmed.aai7795
33. Gopalai, AA, Lim, JL, Li, HH, Zhao, Y, Lim, TT, Eow, GB, et al. Lrrk2 N551k and R1398h variants are protective in Malays and Chinese in Malaysia: a case-control association study for Parkinson's disease. Mol Genet Genomic Med. (2019) 7:E604. doi: 10.1002/mgg3.604
34. Don Wai Luu, L, Kaakoush, NO, and Castaño-Rodríguez, N. The role of Atg16l2 in autophagy and disease. Autophagy. (2022) 18:2537–46. doi: 10.1080/15548627.2022.2042783
35. Foerster, EG, Mukherjee, T, Cabral-Fernandes, L, Rocha, JDB, Girardin, SE, and Philpott, DJ. How autophagy controls the intestinal epithelial barrier. Autophagy. (2022) 18:86–103. doi: 10.1080/15548627.2021.1909406
36. Chen, SL, Li, CM, Li, W, Liu, QS, Hu, SY, Zhao, MY, et al. How autophagy, a potential therapeutic target, regulates intestinal inflammation. Front Immunol. (2023) 14:1087677. doi: 10.3389/fimmu.2023.1087677
37. Glassner, KL, Abraham, BP, and Quigley, EMM. The microbiome and inflammatory bowel disease. J Allergy Clin Immunol. (2020) 145:16–27. doi: 10.1016/j.jaci.2019.11.003
38. Atreya, R, and Neurath, MF. Il-23 blockade in anti-Tnf refractory Ibd: from mechanisms to clinical reality. J Crohns Colitis. (2022) 16:Ii54-Ii63. doi: 10.1093/ecco-jcc/jjac007
39. Liu, D, Saikam, V, Skrada, KA, Merlin, D, and Iyer, SS. Inflammatory bowel disease biomarkers. Med Res Rev. (2022) 42:1856–87. doi: 10.1002/med.21893
40. Tansey, MG, Wallings, RL, Houser, MC, Herrick, MK, Keating, CE, and Joers, V. Inflammation and immune dysfunction in Parkinson disease. Nat Rev Immunol. (2022) 22:657–73. doi: 10.1038/s41577-022-00684-6
41. Chapelet, G, Leclair-Visonneau, L, Clairembault, T, Neunlist, M, and Derkinderen, P. Can the gut be the missing piece in uncovering PD pathogenesis? Parkinsonism Relat Disord. (2019) 59:26–31. doi: 10.1016/j.parkreldis.2018.11.014
42. Praschberger, R, Kuenen, S, Schoovaerts, N, Kaempf, N, Singh, J, Janssens, J, et al. Neuronal identity defines Α-Synuclein and tau toxicity. Neuron. (2023) 111:1577–1590.e11. doi: 10.1016/j.neuron.2023.02.033
43. Blesa, J, Foffani, G, Dehay, B, Bezard, E, and Obeso, JA. Motor and non-motor circuit disturbances in early Parkinson disease: which happens first? Nat Rev Neurosci. (2022) 23:115–28. doi: 10.1038/s41583-021-00542-9
44. Rolli-Derkinderen, M, Leclair-Visonneau, L, Bourreille, A, Coron, E, Neunlist, M, and Derkinderen, P. Is Parkinson's disease a chronic low-grade inflammatory bowel disease? J Neurol. (2020) 267:2207–13. doi: 10.1007/s00415-019-09321-0
45. Prigent, A, Lionnet, A, Durieu, E, Chapelet, G, Bourreille, A, Neunlist, M, et al. Enteric alpha-Synuclein expression is increased in Crohn's disease. Acta Neuropathol. (2019) 137:359–61. doi: 10.1007/s00401-018-1943-7
46. Kishimoto, Y, Zhu, W, Hosoda, W, Sen, JM, and Mattson, MP. Chronic mild gut inflammation accelerates brain neuropathology and motor dysfunction in α-Synuclein mutant mice. NeuroMolecular Med. (2019) 21:239–49. doi: 10.1007/s12017-019-08539-5
47. Chen, M, and Mor, DE. Gut-to-brain α-Synuclein transmission in Parkinson's disease: evidence for prion-like mechanisms. Int J Mol Sci. (2023) 24:7205. doi: 10.3390/ijms24087205
48. Guyer, RA, Stavely, R, Robertson, K, Bhave, S, Mueller, JL, Picard, NM, et al. Single-cell Multiome sequencing clarifies enteric glial diversity and identifies An Intraganglionic population poised for neurogenesis. Cell Rep. (2023) 42:112194. doi: 10.1016/j.celrep.2023.112194
49. Barajon, I, Serrao, G, Arnaboldi, F, Opizzi, E, Ripamonti, G, Balsari, A, et al. Toll-like receptors 3, 4, and 7 are expressed in the enteric nervous system and dorsal root ganglia. J Histochem Cytochem. (2009) 57:1013–23. doi: 10.1369/jhc.2009.953539
50. Yang, PC, Li, XJ, Yang, YH, Qian, W, Li, SY, Yan, CH, et al. The influence of Bifidobacterium Bifidum and Bacteroides fragilis on enteric glial cell-derived neurotrophic factors and inflammasome. Inflammation. (2020) 43:2166–77. doi: 10.1007/s10753-020-01284-z
51. Ibiza, S, García-Cassani, B, Ribeiro, H, Carvalho, T, Almeida, L, Marques, R, et al. Glial-cell-derived neuroregulators control type 3 innate lymphoid cells and gut defence. Nature. (2016) 535:440–3. doi: 10.1038/nature18644
52. Kovler, ML, Gonzalez Salazar, AJ, Fulton, WB, Lu, P, Yamaguchi, Y, Zhou, Q, et al. Toll-like receptor 4-mediated enteric glia loss is critical for the development of necrotizing enterocolitis. Sci Transl Med. (2021) 13:Eabg3459. doi: 10.1126/scitranslmed.abg3459
53. Drokhlyansky, E, Smillie, CS, van Wittenberghe, N, Ericsson, M, Griffin, GK, Eraslan, G, et al. The human and mouse enteric nervous system at single-cell resolution. Cell. (2020) 182:1606–1622.e23. doi: 10.1016/j.cell.2020.08.003
54. Arizona Parkinson’s Disease ConsortiumBeach, TG, Adler, CH, Sue, LI, Vedders, L, Lue, LF, et al. Multi-organ distribution of phosphorylated alpha-Synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. (2010) 119:689–702. doi: 10.1007/s00401-010-0664-3
55. Perez-Pardo, P, Grobben, Y, Willemsen-Seegers, N, Hartog, M, Tutone, M, Muller, M, et al. Pharmacological validation of Tdo as a target for Parkinson's disease. FEBS J. (2021) 288:4311–31. doi: 10.1111/febs.15721
56. Esposito, G, Capoccia, E, Gigli, S, Pesce, M, Bruzzese, E, D’Alessandro, A, et al. Hiv-1 tat-induced diarrhea evokes An enteric glia-dependent Neuroinflammatory response in the central nervous system. Sci Rep. (2017) 7:7735. doi: 10.1038/s41598-017-05245-9
57. Carabotti, M, Scirocco, A, Maselli, MA, and Severi, C. The gut-brain Axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. (2015) 28:203–9.
58. Morais, LH, Schreiber, HLT, and Mazmanian, SK. The gut microbiota-brain Axis in behaviour and brain disorders. Nat Rev Microbiol. (2021) 19:241–55. doi: 10.1038/s41579-020-00460-0
59. Kwon, HJ, Mohammed, AE, Eltom, KH, Albrahim, JS, and Alburae, NA. Evaluation of antibiotic-induced behavioral changes in mice. Physiol Behav. (2020) 223:113015. doi: 10.1016/j.physbeh.2020.113015
60. Leclercq, S, Mian, FM, Stanisz, AM, Bindels, LB, Cambier, E, Ben-Amram, H, et al. Low-dose penicillin in early life induces Long-term changes in murine gut microbiota, brain cytokines and behavior. Nat Commun. (2017) 8:15062. doi: 10.1038/ncomms15062
61. Yang, J, Zheng, P, Li, Y, Wu, J, Tan, X, Zhou, J, et al. Landscapes of bacterial and metabolic signatures and their interaction in major depressive disorders. Sci Adv. (2020) 6:eaba8555. doi: 10.1126/sciadv.aba8555
62. Liang, S, Sin, ZY, Yu, J, Zhao, S, Xi, Z, Bruzzone, R, et al. Multi-cohort analysis of depression-associated gut Bacteria sheds insight on bacterial biomarkers across populations. Cell Mol Life Sci. (2022) 80:9. doi: 10.1007/s00018-022-04650-2
63. Kelly, JR, Borre, Y, O' Brien, C, Patterson, E, el Aidy, S, Deane, J, et al. Transferring the blues: depression-associated gut microbiota induces Neurobehavioural changes in the rat. J Psychiatr Res. (2016) 82:109–18. doi: 10.1016/j.jpsychires.2016.07.019
64. Morris, HR, Spillantini, MG, Sue, CM, and Williams-Gray, CH. The pathogenesis of Parkinson's disease. Lancet. (2024) 403:293–304. doi: 10.1016/S0140-6736(23)01478-2
65. Kim, S, Kwon, SH, Kam, TI, Panicker, N, Karuppagounder, SS, Lee, S, et al. Transneuronal propagation of pathologic α-Synuclein from the gut to the brain models Parkinson's disease. Neuron. (2019) 103:627–641.e7. doi: 10.1016/j.neuron.2019.05.035
66. Uemura, N, Yagi, H, Uemura, MT, Hatanaka, Y, Yamakado, H, and Takahashi, R. Inoculation of Α-Synuclein preformed fibrils into the mouse gastrointestinal tract induces Lewy body-like aggregates in the brainstem via the Vagus nerve. Mol Neurodegener. (2018) 13:21. doi: 10.1186/s13024-018-0257-5
67. Ahn, EH, Kang, SS, Liu, X, Chen, G, Zhang, Z, Chandrasekharan, B, et al. Initiation of Parkinson's disease from gut to brain by Δ-secretase. Cell Res. (2020) 30:70–87. doi: 10.1038/s41422-019-0241-9
68. Salat-Foix, D, Tran, K, Ranawaya, R, Meddings, J, and Suchowersky, O. Increased intestinal permeability and Parkinson disease patients: chicken or egg? Can J Neurol Sci. (2012) 39:185–8. doi: 10.1017/S0317167100013202
69. Arotcarena, ML, Dovero, S, Prigent, A, Bourdenx, M, Camus, S, Porras, G, et al. Bidirectional gut-to-brain and brain-to-gut propagation of Synucleinopathy in non-human primates. Brain. (2020) 143:1462–75. doi: 10.1093/brain/awaa096
70. Fairbrass, KM, Lovatt, J, Barberio, B, Yuan, Y, Gracie, DJ, and Ford, AC. Bidirectional brain-gut Axis effects influence mood and prognosis in IBD: a systematic review and meta-analysis. Gut. (2022) 71:1773–80. doi: 10.1136/gutjnl-2021-325985
71. Schneider, KM, Blank, N, Alvarez, Y, Thum, K, Lundgren, P, Litichevskiy, L, et al. The enteric nervous system relays psychological stress to intestinal inflammation. Cell. (2023) 186:2823–2838.e20. doi: 10.1016/j.cell.2023.05.001
72. Do, J, and Woo, J. From gut to brain: alteration in inflammation markers in the brain of dextran sodium sulfate-induced colitis model mice. Clin Psychopharmacol Neurosci. (2018) 16:422–33. doi: 10.9758/cpn.2018.16.4.422
73. Sun, MF, and Shen, YQ. Dysbiosis of gut microbiota and microbial metabolites in Parkinson's disease. Ageing Res Rev. (2018) 45:53–61. doi: 10.1016/j.arr.2018.04.004
74. Larabi, A, Barnich, N, and Nguyen, HTT. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in Ibd. Autophagy. (2020) 16:38–51. doi: 10.1080/15548627.2019.1635384
75. Mohebali, N, Weigel, M, Hain, T, Sütel, M, Bull, J, Kreikemeyer, B, et al. Faecalibacterium prausnitzii, Bacteroides faecis and Roseburia intestinalis attenuate clinical symptoms of experimental colitis by regulating Treg/Th17 cell balance and intestinal barrier integrity. Biomed Pharmacother. (2023) 167:115568. doi: 10.1016/j.biopha.2023.115568
76. Romano, S, Savva, GM, Bedarf, JR, Charles, IG, Hildebrand, F, and Narbad, A. Meta-analysis of the Parkinson's disease gut microbiome suggests alterations linked to intestinal inflammation. NPJ Parkinsons Dis. (2021) 7:27. doi: 10.1038/s41531-021-00156-z
77. Metwaly, A, Reitmeier, S, and Haller, D. Microbiome risk profiles as biomarkers for inflammatory and metabolic disorders. Nat Rev Gastroenterol Hepatol. (2022) 19:383–97. doi: 10.1038/s41575-022-00581-2
78. Oliveira, ECS, Quaglio, AEV, Magro, DO, di Stasi, LC, and Sassaki, LY. Intestinal microbiota and Mirna in Ibd: a narrative review about discoveries and perspectives for the future. Int J Mol Sci. (2023) 24:7176. doi: 10.3390/ijms24087176
79. Xiao, Y, Zhai, Q, Zhang, H, Chen, W, and Hill, C. Gut colonization mechanisms of Lactobacillus and Bifidobacterium: An argument for personalized designs. Annu Rev Food Sci Technol. (2021) 12:213–33. doi: 10.1146/annurev-food-061120-014739
80. Xiao, Y, Zhao, J, Zhang, H, Zhai, Q, and Chen, W. Mining Lactobacillus and Bifidobacterium for organisms with Long-term gut colonization potential. Clin Nutr. (2020) 39:1315–23. doi: 10.1016/j.clnu.2019.05.014
81. Dalile, B, van Oudenhove, L, Vervliet, B, and Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. (2019) 16:461–78. doi: 10.1038/s41575-019-0157-3
82. Chambers, ES, Byrne, CS, Morrison, DJ, Murphy, KG, Preston, T, Tedford, C, et al. Dietary supplementation with inulin-propionate Ester or inulin improves insulin sensitivity in adults with overweight and obesity with distinct effects on the gut microbiota, plasma metabolome and systemic inflammatory responses: a randomised cross-over trial. Gut. (2019) 68:1430–8. doi: 10.1136/gutjnl-2019-318424
83. Guo, TT, Zhang, Z, Sun, Y, Zhu, RY, Wang, FX, Ma, LJ, et al. Neuroprotective effects of sodium butyrate by restoring gut microbiota and inhibiting Tlr4 signaling in mice with Mptp-induced Parkinson's disease. Nutrients. (2023) 15:930. doi: 10.3390/nu15040930
84. Shin, C, Lim, Y, Lim, H, and Ahn, TB. Plasma short-chain fatty acids in patients with Parkinson's disease. Mov Disord. (2020) 35:1021–7. doi: 10.1002/mds.28016
85. Liu, X, du, ZR, Wang, X, Sun, XR, Zhao, Q, Zhao, F, et al. Polymannuronic acid prebiotic plus Lacticaseibacillus rhamnosus gg probiotic as a novel Synbiotic promoted their separate neuroprotection against Parkinson's disease. Food Res Int. (2022) 155:111067. doi: 10.1016/j.foodres.2022.111067
86. Gonçalves, P, Araújo, JR, and Di Santo, JP. A cross-talk between microbiota-derived short-chain fatty acids and the host mucosal immune system regulates intestinal homeostasis and inflammatory bowel disease. Inflamm Bowel Dis. (2018) 24:558–72. doi: 10.1093/ibd/izx029
87. Chen, SJ, and Lin, CH. Gut microenvironmental changes as a potential trigger in Parkinson's disease through the gut-brain Axis. J Biomed Sci. (2022) 29:54. doi: 10.1186/s12929-022-00839-6
88. Zhuang, X, Li, T, Li, M, Huang, S, Qiu, Y, Feng, R, et al. Systematic review and meta-analysis: short-chain fatty acid characterization in patients with inflammatory bowel disease. Inflamm Bowel Dis. (2019) 25:1751–63. doi: 10.1093/ibd/izz188
89. Magro, F, Cordeiro, G, Dias, AM, and Estevinho, MM. Inflammatory bowel disease-non-biological treatment. Pharmacol Res. (2020) 160:105075. doi: 10.1016/j.phrs.2020.105075
90. Jeong, DY, Kim, S, Son, MJ, Son, CY, Kim, JY, Kronbichler, A, et al. Induction and maintenance treatment of inflammatory bowel disease: a comprehensive review. Autoimmun Rev. (2019) 18:439–54. doi: 10.1016/j.autrev.2019.03.002
91. Vijiaratnam, N, Simuni, T, Bandmann, O, Morris, HR, and Foltynie, T. Progress towards therapies for disease modification in Parkinson's disease. Lancet Neurol. (2021) 20:559–72. doi: 10.1016/S1474-4422(21)00061-2
92. Li, HX, Zhang, C, Zhang, K, Liu, YZ, Peng, XX, and Zong, Q. Inflammatory bowel disease and risk of Parkinson's disease: evidence from a meta-analysis of 14 studies involving more than 13.4 million individuals. Front Med. (2023) 10:1137366. doi: 10.3389/fmed.2023.1137366
93. Peter, I, Dubinsky, M, Bressman, S, Park, A, Lu, C, Chen, N, et al. Anti-tumor necrosis Factor therapy and incidence of Parkinson disease among patients with inflammatory bowel disease. JAMA Neurol. (2018) 75:939–46. doi: 10.1001/jamaneurol.2018.0605
94. Pinel Ríos, J, Madrid Navarro, CJ, Pérez Navarro, MJ, Cabello Tapia, MJ, Piña Vera, MJ, Campos Arillo, V, et al. Association of Parkinson's disease and treatment with Aminosalicylates in inflammatory bowel disease: a cross-sectional study in a Spain drug dispensation records. BMJ Open. (2019) 9:E025574. doi: 10.1136/bmjopen-2018-025574
95. Kamel, WA, and Al-Hashel, JY. Lcig in treatment of non-motor symptoms in advanced Parkinson's disease: review of literature. Brain Behav. (2020) 10:E01757. doi: 10.1002/brb3.1757
96. Dogra, N, Jakhmola-Mani, R, Potshangbam, AM, Buch, S, and Pande Katare, D. Cxcr4 as possible Druggable target linking inflammatory bowel disease and Parkinson's disease. Metab Brain Dis. (2023) 38:1079–96. doi: 10.1007/s11011-022-01155-6
97. Sun, MF, Zhu, YL, Zhou, ZL, Jia, XB, Xu, YD, Yang, Q, et al. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson's disease mice: gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav Immun. (2018) 70:48–60. doi: 10.1016/j.bbi.2018.02.005
98. Huang, H, Xu, H, Luo, Q, He, J, Li, M, Chen, H, et al. Fecal microbiota transplantation to treat Parkinson's disease with constipation: a case report. Medicine. (2019) 98:E16163. doi: 10.1097/MD.0000000000016163
99. Xue, LJ, Yang, XZ, Tong, Q, Shen, P, Ma, SJ, Wu, SN, et al. Fecal microbiota transplantation therapy for Parkinson's disease: a preliminary study. Medicine. (2020) 99:E22035. doi: 10.1097/MD.0000000000022035
100. Li, M, Yang, L, Mu, C, Sun, Y, Gu, Y, Chen, D, et al. Gut microbial metabolome in inflammatory bowel disease: from association to therapeutic perspectives. Comput Struct Biotechnol J. (2022) 20:2402–14. doi: 10.1016/j.csbj.2022.03.038
101. Imdad, A, Pandit, NG, Zaman, M, Minkoff, NZ, Tanner-Smith, EE, Gomez-Duarte, OG, et al. Fecal transplantation for treatment of inflammatory bowel disease. Cochrane Database Syst Rev. (2023) 4:Cd012774. doi: 10.1002/14651858.CD012774.pub3
102. Ganguly, NK, Bhattacharya, SK, Sesikeran, B, Nair, GB, Ramakrishna, BS, Sachdev, HPS, et al. ICMR-DBT guidelines for evaluation of probiotics in food. Indian J Med Res. (2011) 134:22–5.
103. Suez, J, Zmora, N, Segal, E, and Elinav, E. The pros, cons, and many unknowns of probiotics. Nat Med. (2019) 25:716–29. doi: 10.1038/s41591-019-0439-x
104. Gao, J, Zhao, L, Cheng, Y, Lei, W, Wang, Y, Liu, X, et al. Probiotics for the treatment of depression and its comorbidities: a systemic review. Front Cell Infect Microbiol. (2023) 13:1167116. doi: 10.3389/fcimb.2023.1167116
105. Sun, H, Zhao, F, Liu, Y, Ma, T, Jin, H, Quan, K, et al. Probiotics synergized with conventional regimen in managing Parkinson's disease. Npj Parkinsons Dis. (2022) 8:62. doi: 10.1038/s41531-022-00327-6
106. Zhao, Z, Ning, J, Bao, XQ, Shang, M, Ma, J, Li, G, et al. Fecal microbiota transplantation protects rotenone-induced Parkinson's disease mice via suppressing inflammation mediated by the lipopolysaccharide-Tlr4 signaling pathway through the microbiota-gut-brain Axis. Microbiome. (2021) 9:226. doi: 10.1186/s40168-021-01107-9
107. Hsieh, TH, Kuo, CW, Hsieh, KH, Shieh, MJ, Peng, CW, Chen, YC, et al. Probiotics alleviate the progressive deterioration of motor functions in a mouse model of Parkinson's disease. Brain Sci. (2020) 10:206. doi: 10.3390/brainsci10040206
108. Kruis, W, Fric, P, Pokrotnieks, J, Lukás, M, Fixa, B, Kascák, M, et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard Mesalazine. Gut. (2004) 53:1617–23. doi: 10.1136/gut.2003.037747
109. Teran, MDM, Perez Visñuk, D, Savoy de Giori, G, de Moreno de LeBlanc, A, and LeBlanc, JG. Neuroprotective effect of thiamine-producing lactic acid bacteria in a murine parkinsonian model. Food Funct. (2022) 13:8056–67. doi: 10.1039/D2FO01195F
Keywords: inflammatory bowel disease, Parkinson’s disease, gut-brain axis, short-chain fatty acids, gut microbiota
Citation: Wanyi Z, Jiao Y, Wen H, Bin X, Xuefei W, Lan J and Liuyin Z (2024) Bidirectional communication of the gut-brain axis: new findings in Parkinson’s disease and inflammatory bowel disease. Front. Neurol. 15:1407241. doi: 10.3389/fneur.2024.1407241
Edited by:
Sheila Pirooznia, National Institutes of Health (NIH), United StatesReviewed by:
Christopher B. Forsyth, Rush University, United StatesAnastazja Gorecki, University of Notre Dame Australia, Australia
Panida Sittipo, Burapha University, Thailand
Copyright © 2024 Wanyi, Jiao, Wen, Bin, Xuefei, Lan and Liuyin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiang Lan, amxhbm9rQDE2My5jb20=; Zhou Liuyin, emhvdWxpdXlpbjYzNTNAMTYzLmNvbQ==
†These authors have contributed equally to this work