 Huimin Yan1Minglei Liu1
Huimin Yan1Minglei Liu1 Yuan Gao1,2,3,4
Yuan Gao1,2,3,4 Yanpeng Yuan1,2,3,4
Yanpeng Yuan1,2,3,4 Xiaojing Liu1,2,4
Xiaojing Liu1,2,4 Yangyang Wang1,2,4
Yangyang Wang1,2,4 Lanjun Li1,2Qingzhi Wang1
Lanjun Li1,2Qingzhi Wang1 Yanlin Wang1,2,3,4
Yanlin Wang1,2,3,4 Changhe Shi1,4
Changhe Shi1,4 Yuming Xu1,2,3,4*
Yuming Xu1,2,3,4* Jing Yang1,2,3*
Jing Yang1,2,3*- 1Department of Neurology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, China
- 2NHC Key Laboratory of Prevention and Treatment of Cerebrovascular Disease, Zhengzhou, Henan, China
- 3Henan Key Laboratory of Cerebrovascular Diseases, Zhengzhou University, Zhengzhou, Henan, China
- 4Institute of Neuroscience, Zhengzhou University, Zhengzhou, Henan, China
Background: Overwhelming evidence points to that genetic factors contributing to the development of Alzheimer’s disease (AD) and Parkinson’s disease (PD). Genome-Wide Association Study (GWAS) has come a long way in the last decade. So far, a large number of GWAS studies have been published on neurological diseases and many other diseases, providing us with a wealth of genetic information and unique biological insights.
Methods: Genomic DNA was extracted from both patients’ and controls’ peripheral blood samples utilizing the Blood Genome Extraction Kit. Single nucleotide polymorphisms (SNPs) were genotyped employing the enhanced multiple ligase detection reaction (iMLDR) technology.
Results: A case-control study was conducted, involving 211 AD patients, 508 PD patients (including 117 with dementia), and 412 healthy individuals. Age and sex stratification analysis revealed that rs871269/TNIP1 was associated with LOAD (p = 0.035), and rs5011436/TMEM106B was associated with AD in males (p = 0.044) in the genotype model. In the allele model, rs871269/TNIP1 was found to be associated with PD in the Chinese Han population (p = 0.0035, OR 0.741, 95% CI 0.559-0.983), and rs708382/GRN was identified as a risk factor for Parkinson’s disease dementia (PDD) in the Chinese Han population (p = 0.004, odds ratio (OR) 0.354, 95% confidence interval (CI) 0.171-0.733). However, no significant associations with AD or PD were observed for the remaining four loci (rs113020870/AGRN, rs6891966/HAVCR2, rs2452170/NTN5, rs1761461/LILRB2) in terms of allele or genotype frequencies.
Conclusion: This study identifies rs871269/TNIP1 as a potential risk factor for both LOAD and PD, rs708382/GRN as a risk factor for PDD, and rs5011436/TMEM106B as associated with AD in males when stratified by age.
1 Introduction
Alzheimer’s disease (AD) and Parkinson’s disease (PD) rank as the first and second most common neurodegenerative diseases, respectively (1). Globally, 44 million people are living with dementia, with AD accounting for approximately 50–75 percent. Remarkably, half of these cases are from China. This number is anticipated to more than triple by 2050 due to an aging population (2). The incidence of PD increases with age, the prevalence of PD in the elderly population over 65 years old is about 1.7%, and the prevalence of PD in the elderly population over 80 years old is as high as 4%, exerting immense pressure on society (3).
Most cases of AD and PD are sporadic with unknown etiology, with both environmental and genetic factors contributing to their development (4). Overwhelming evidence points to important genetic roles (5). AD and PD may share common pathological processes, such as abnormal protein deposition. The presence of Aβ, a signature pathological protein of AD, has been reported in PD patients (6), and lewy body deposits, characteristic of PD pathology, are also found in AD cases (7). Additionally, the APOE and MAPT genes, recognized risk genes for AD (8), have also been linked to increased PD risk, highlighting potential genetic and pathological overlaps between AD and PD (9).
Cognitive dysfunction, a prevalent non-motor symptom of PD, further complicates the disease landscape. Parkinson’s disease dementia (PDD) progresses at a rate of about 10% per year in PD patients, with a cumulative prevalence of 75% in patients with PD for more than 10 years (10). The Sydney multicenter study, the APOE longest cohort to date, reported an 83% incidence of dementia among patients surviving more than 20 years (11), underscoring the significant overlap between PD and dementia. Thus, it is of interest to explore whether risk loci for AD may be risk factors for PD-induced dementia.
Genome-Wide Association Study (GWAS) has come a long way in the last decade (12). Researchers can detect genetic markers associated with specific traits or diseases on a genome-wide scale (13). Traditional genetic link research methods have limitations for the study of this complex trait (14), but GWAS can identify risk variants on a genome-wide scale, thus providing a new perspective for the study of the mechanism of complex diseases (15). So far, a large number of GWAS studies have been published on neurological diseases and many other diseases, providing us with a wealth of genetic information and unique biological insights (16).
A recent large-scale meta-analysis pooling 13 AD Genome-Wide Association Study (GWAS) datasets from European ancestry samples identified 38 loci, including seven loci that have not been reported previously (17). Of the 31 loci that have been reported, some researchers have verified some of them in the Chinese population (18–20). However, the relevance of these new loci to AD in Asian populations remains unexplored. In light of these findings, this study replicates AD-related genetic variants in the AD and PD population to identify additional candidate sites that might alter their risk.
2 Materials and methods
2.1 Study population
A total of 1,131 subjects of Han ethnicity were enrolled in this study, encompassing 211 AD patients, 508 sporadic PD patients, and 412 control subjects. Blood samples were collected from all participants for SNP testing, and patient information, including information related to age, sex, and education (years of schooling), was collected at the time of sampling. The simple mental state examination (MMSE) was used to screen cognitive function, and the scores of controls were in the normal range. Among PD patients, 117 were diagnosed with PDD based on the criteria proposed by the Movement Disorder Society Task Force, and we classified those with a disease course of more than 10 years and no cognitive impairment into the group of PD without dementia (PDND). All recruited patients were sporadic cases, with young-onset patients excluded from the study. Case definitions adhered to the criteria set by the National Institute on Aging–Alzheimer’s Association (2018) and Parkinson’s United Kingdom Brain Bank (1997). All study participants provided written informed consent, and the study received approval from the Ethics Committee of the First Affiliated Hospital of Zhengzhou University.
2.2 Genotyping and data analysis
Genomic DNA was extracted from both patients’ and controls’ peripheral blood samples utilizing the Blood Genome Extraction Kit (BioTeke Co., Beijing, China). Single nucleotide polymorphisms (SNPs) were genotyped employing the enhanced multiple ligase detection reaction (iMLDR) technology (Geneskybiotech, Shanghai, China). All pertinent specific PCR primers and ligation primers are enumerated in Supplementary Table 1.
Statistical analyses were executed using IBM SPSS Statistics 26.0. The age disparity was evaluated employing the t-test. The Hardy–Weinberg equilibrium in genotype frequency among controls was assessed utilizing the χ2 test. Logistic regression analysis facilitated the computation of risk analysis for each SNP in both dominant and recessive models, post adjustment for age and gender. Chi-squared tests were employed to contrast differences in sex ratio, genotype frequency, and allele frequency post-age and gender-stratified analysis. A two-tailed p value less than 0.05 was deemed statistically significant.
3 Results
We enrolled 211 AD patients, 508 sporadic PD patients, and 412 control subjects. AD and control, PD and control, and PDD and PDND had no significant difference in age and sex (Table 1). The APOE ε4 allele is one of the major genetic risk factors for AD (21). Unlike AD, the association between the APOE gene and PD is less clear. Some studies have shown an association between the APOE ε4 allele and the risk of PD, but the association is not as strong as it is with AD (22). Therefore, we tried to explore the relationship between APOEε4(+) and PD and PDD. The results showed that APOEε4(+) had significant difference between AD and control, but no significant difference between PD and control, PDD and PDND. The association between the APOE gene and PD still needs more research to confirm and explain.

Table 1. Characteristics of the study groups.
The frequencies of all seven variants in both case and control groups were in accordance with the Hardy–Weinberg equilibrium (p > 0.05). In all sites, genotype and allele frequencies of AD and control were not statistically different (Table 2). Considering that our AD population encompasses people of all ages, but these seven loci were found in the LOAD population in the study by Wightman et al. (17). In addition, the proportion of female and male patients in AD is 3:1, and female gender as a high-risk factor for Alzheimer’s disease has also been the focus of academic research in recent years (2). Therefore, we conducted a stratified analysis by age and sex. We found that rs871269/TNIP1 was associated with LOAD (p = 0.035), and rs5011436/TMEM106B was associated with male AD (p = 0.044) in the genotype model (Table 3).
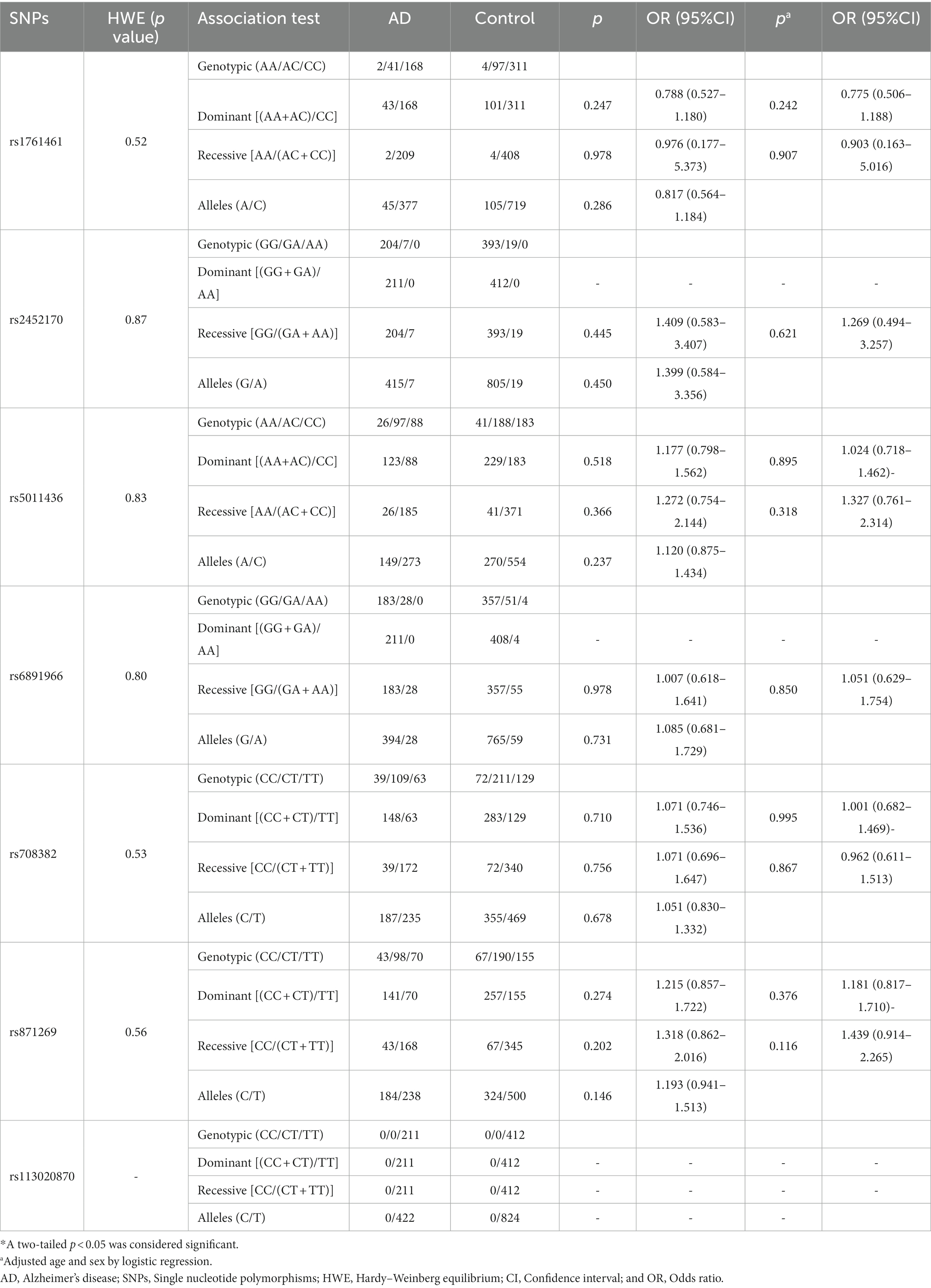
Table 2. Distribution of included SNPs in AD patients and control subjects.
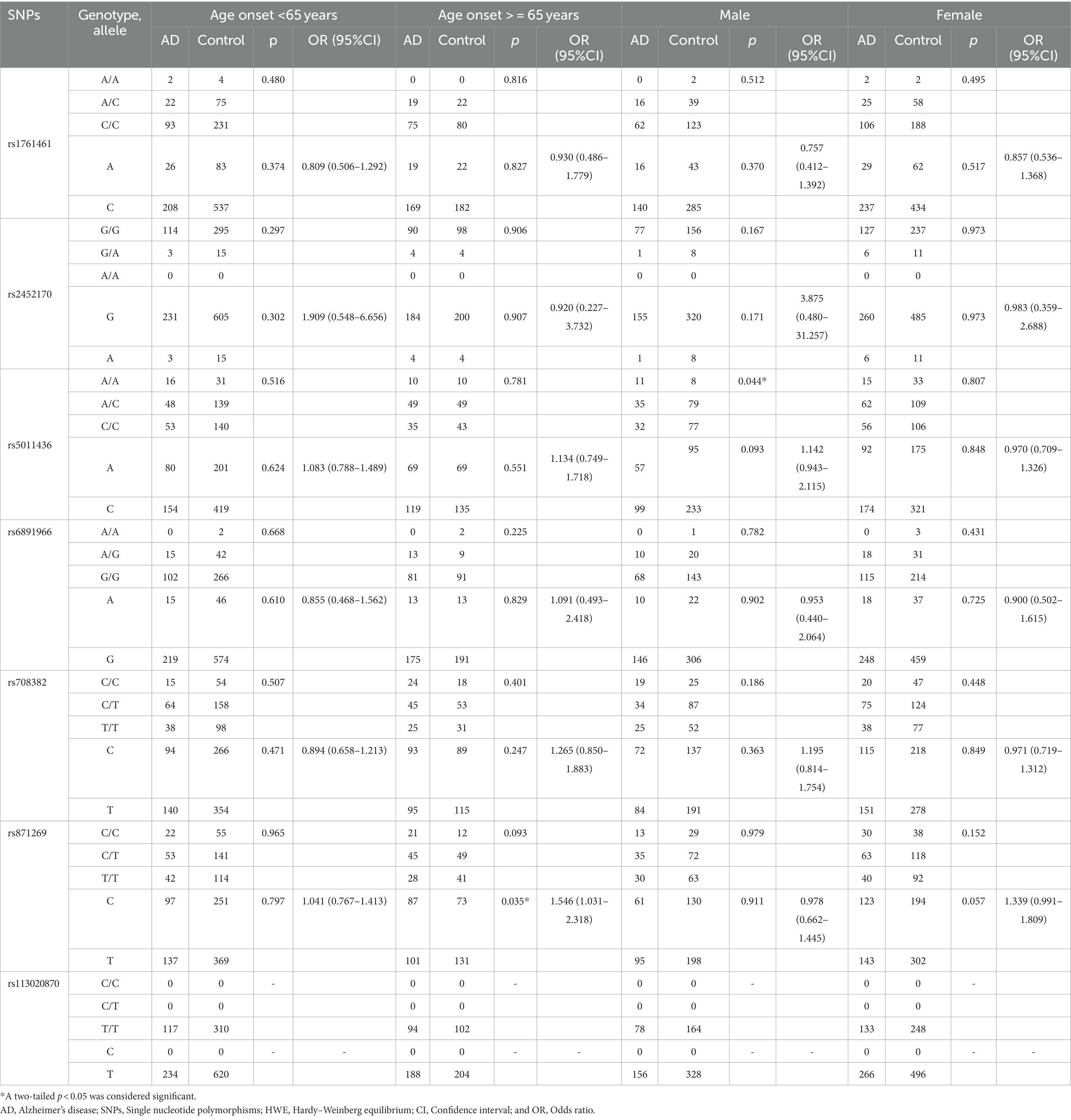
Table 3. Age-stratified and sex-stratified analysis of 7 loci.
In allelic models, rs871269/TNIP1 showed a significant difference between PD patients and controls (p = 0.0035, OR 0.741, 95% CI 0.559–0.983; Table 4). Similarly, a significant difference was observed in the variation of rs708382/GRN between PDD patients and PDND patients [p = 0.004, odds ratio (OR) 0.354, 95% confidence interval (CI) 0.171–0.733; Table 5], with the patient group exhibiting higher levels of the C allele compared to the control group. Because PD is divided into early-onset and late-onset according to the 40-year old boundary (23), we did not conduct a stratified analysis of PD by age and gender. Considering that we did many tests, we adopted Bonferroni correction method to adjust the statistical significance level, and the results showed that rs871269/TNIP1 was still statistically different between PD patients and controls, and rs708382/GRN was still statistically different between PDD patients and PDND controls. However, rs871269/TNIP1 and rs5011436/TMEM106B no longer showed statistical differences in AD sex and age stratification.
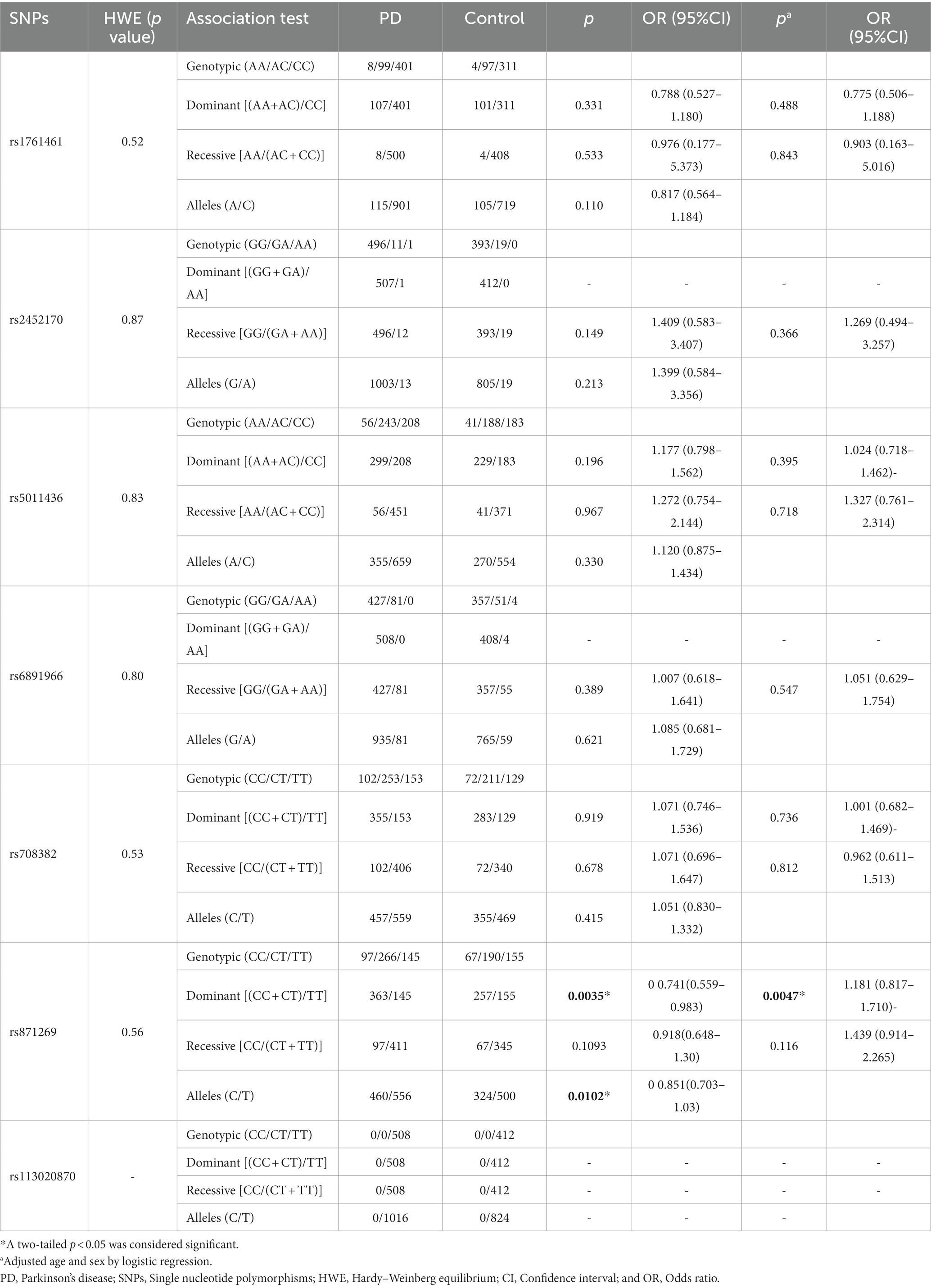
Table 4. Distribution of included SNPs in PD patients and control subjects.
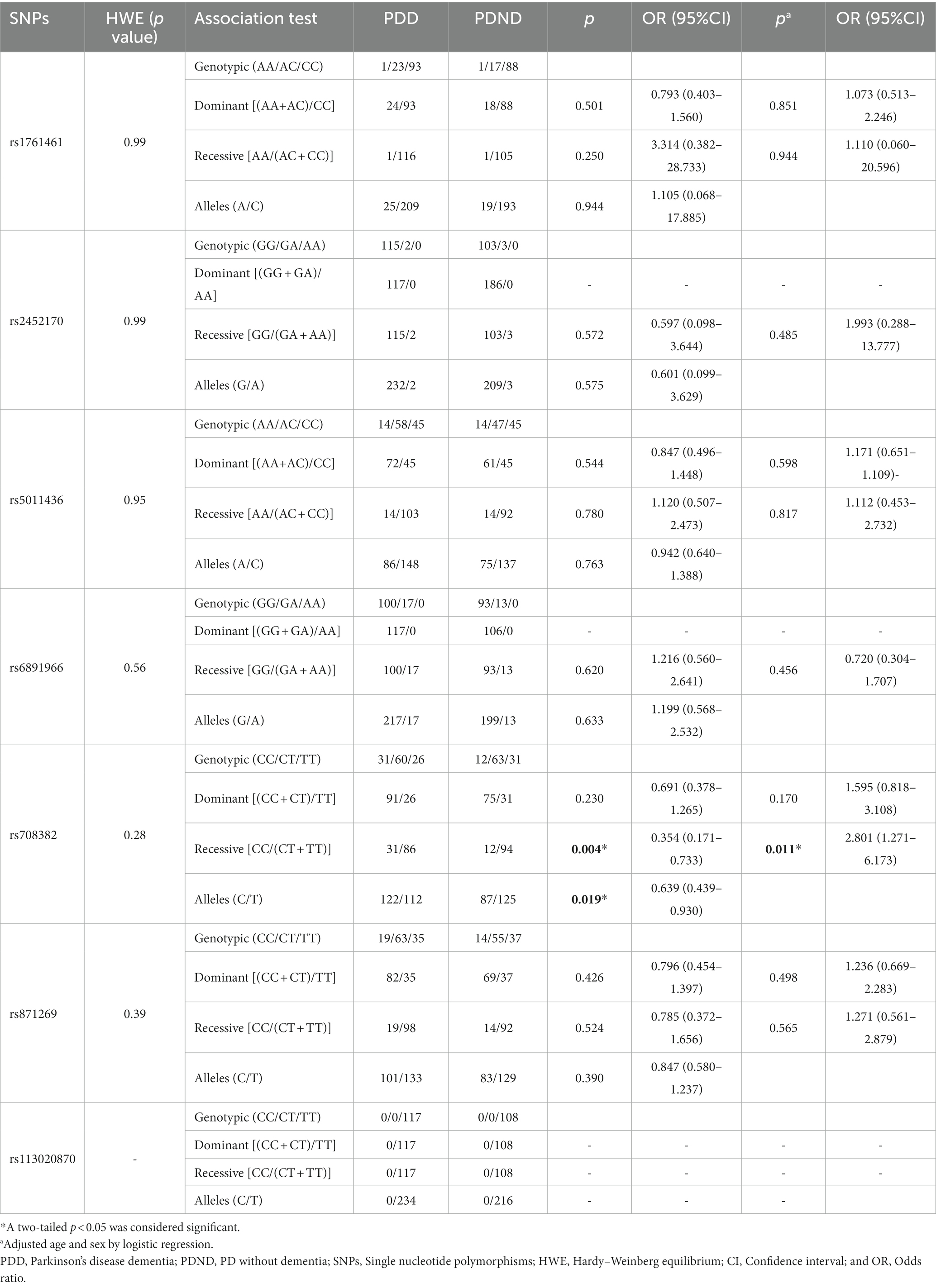
Table 5. Distribution of included SNPs in PDD patients and PDND patients.
In contrast, for the remaining four sites (rs113020870/AGRN, rs6891966/HAVCR2, rs2452170/NTN5, and rs1761461/LILRB2), no statistical difference in genotype or allele frequency was noted between AD, PD, and PDD patients, and the control group. Among these risk sites, no base mutation of rs113020870 was present in any of our patients or healthy controls. Comprehensive details regarding the relationship between the seven sites and AD, PD, and PDD levels are delineated in Tables 2–5.
4 Discussion
Considering the impact of ethnic heterogeneity, the current study explored seven newly identified AD-related variants within the Han Chinese population. The findings revealed distinct differences in allelic modes: rs871269/TNIP1 between LOAD patients and controls, and again rs5011436/TMEM106B between male AD patients and controls, rs871269/TNIP1 between PD patients and controls, and rs708382/GRN between PDD patients and PDND controls. To the best of our knowledge, this research is the inaugural study to demonstrate the correlation of SNPs in GRN, TNIP1, and TMEM106B genes in Chinese Han Cohort. The remaining four loci exhibited no significant disparities in genotype or allele frequency between AD, PDD, PD patients, and the control group. This partial observation may be attributed to genetic heterogeneity stemming from ethnic and geographical variations, as noted by Wightman et al. (17) (refer to Supplementary Table 2 for an exhaustive overview of the seven loci of GWAS results). Moreover, the interplay between environmental and genetic factors may potentially influence gene expression. In addition, our sample size is much smaller than that of Wightman et al. (17), so our results serve as a preliminary hint.
GRN, a recognized frontotemporal dementia (FTD) gene (24), holds associations not only with FTD but also with LOAD (25). Its biological significance may pertain more to protein clearance mechanisms rather than involvement in specific disease-related protein aggregation. A notable pathological characteristic of PD is the presence of Lewy bodies within the cytoplasm of neurons. These are primarily composed of α-synuclein, ubiquitin, and heat shock proteins (26). Such observations suggest a potential relation of GRN to PD or PDD.
TNIP1, known to induce excessive inflammation, has been previously identified in autoimmune GWAS (27). Within mouse microglia, TNIP1 is encompassed in a transcription module regulated by BCL3, correlating with prolonged microglial exposure to inflammation and aging (28). Various pieces of evidence have underscored that α-synuclein is the pivotal trigger of PD neuroinflammation (29). Some studies have discerned that α-synuclein can mediate the oxidative stress of microglial cells (30). However, the assertion that neuroinflammation is a fundamental process of PD remains a topic of uncertainty. The specific role of TNIP1 in the pathogenesis of PD necessitates further exploration.
TMEM106B has been previously linked with dementia, albeit not in preceding LOAD GWAS (31). The rs5011436 is an intronic variant of TMEM106B. The proximate exon variant, rs3173615, is postulated to be an associated signal-driven variant in FTD, leading to a diminished abundance of transmembrane protein 106B (the protein encoded by TMEM106B) through augmented protein degradation (32). Noteworthy differences in the expression of TMEM106B in extensive brain tissues of LOAD patients compared with the control group have been observed (33). The intricate relationship of TMEM106B with AD warrants further comprehensive research.
The present study harbors certain limitations, including a relatively modest sample size. After the Bonferroni correction, rs871269/TNIP1 and rs5011436/TMEM106B no longer showed statistical differences in AD sex and age stratification. Although Bonferroni correction is relatively strict, it still needs our further attention and verification. Interestingly, in Wightman et al. (17), rs871269 T allele is associated with lower AD risk, and our study indicates a protective effect in PD. However, the T allele is the dominant allele in our study population, while it is the minor allele in the European population, considering that it may be due to racial differences. It is imperative to note that the molecular mechanisms underlying the associations between rs708382/GRN and rs871269/TNIP1 and PD remain enigmatic. The design of additional functional experiments is essential to further elucidate the pathogenesis.
In conclusion, our research indicates that the variation of TNIP1 is significantly associated with LOAD and PD within the Chinese Han population, while TMEM106B with male AD and GRN is correlated with PDD. The involvement of geographic or environmental factors in the genetic outcomes at these sites remains an open question. Comprehensive further genetic analyses and functional studies are imperative to unravel the roles of these variants in the pathogenesis of AD and PD.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material; further inquiries can be directed to the corresponding authors.
Ethics statement
The study received approval from the Ethics Committee of the First Affiliated Hospital of Zhengzhou University. All study participants provided written informed consent.
Author contributions
HY: Conceptualization, Formal analysis, Methodology, Software, Writing – original draft, Validation, Writing – review & editing. ML: Formal analysis, Methodology, Writing – original draft. YG: Writing – original draft, Conceptualization, Data curation, Supervision. YY: Conceptualization, Supervision, Validation, Writing – original draft. XL: Conceptualization, Supervision, Validation, Writing – original draft. YangW: Writing – review & editing. LL: Writing – review & editing. QW: Writing – review & editing. YanlW: Writing – review & editing. CS: Writing – review & editing. YX: Funding acquisition, Writing – review & editing. JY: Formal analysis, Funding acquisition, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by grants from the National Natural Science Foundation of China to YX (U1904207), the Nonprofit Central Research Institute Fund of Chinese Academy of Medical Sciences to YX (2020-PT310-01), the National Natural Science Foundation of China to JY (82171434), the Excellent Youth Project of Henan Provincial Natural Science Foundation to JY (222300420070), the Henan Province Young and Middle-Aged Health Science and Technology Innovation Outstanding Youth Training Project to JY (YXKC2020031), and the Funding for Scientific Research and Innovation Team of The First Affiliated Hospital of Zhengzhou University (QNCXTD2023016).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fneur.2024.1326692/full#supplementary-material
References
1. Cui, L, Hou, NN, Wu, HM, Zuo, X, Lian, YZ, Zhang, CN, et al. 6 prevalence of Alzheimer’s disease and Parkinson’s disease in China: an updated systematical analysis. Front Aging Neurosci. (2020) 12:603854. doi: 10.3389/fnagi.2020.603854
2. Alzheimer’s Association. 2023 Alzheimer’s disease facts and figures. Alzheimers Dement. (2023) 19:1598–695. doi: 10.1002/alz.13016
3. Zheng, Z, Zhu, Z, Zhou, C, Cao, L, and Zhao, G. 8 burden of Parkinson disease in China, 1990–2019: findings from the 2019 global burden of disease study. Neuroepidemiology. (2023) 57:51–64. doi: 10.1159/000527372
4. Andrews, SJ, Renton, AE, Fulton-Howard, B, Podlesny-Drabiniok, A, Marcora, E, and Goate, AM. 9 the complex genetic architecture of Alzheimer’s disease: novel insights and future directions. EBioMedicine. (2023) 90:104511. doi: 10.1016/j.ebiom.2023.104511
5. Ye, H, Robak, LA, Yu, M, Cykowski, M, and Shulman, JM. 10 genetics and pathogenesis of Parkinson’s syndrome. Annu Rev Pathol Mech Dis. (2023) 18:95–121. doi: 10.1146/annurev-pathmechdis-031521-034145
6. Ta, M, Blauwendraat, C, Antar, T, Leonard, HL, Singleton, AB, Nalls, MA, et al. Genome-wide meta-analysis of CSF biomarkers in Alzheimer’s disease and Parkinson’s disease cohorts. MedRxiv [Preprint]. (2023). doi: 10.1101/2023.06.13.23291354
7. White, LR, Corrada, MM, Kawas, CH, Cholerton, BA, Edland, SE, Flanagan, ME, et al. 12 Neuropathologic changes of Alzheimer’s disease and related dementias: relevance to future prevention. J Alzheimers Dis. (2023) 95:307–16. doi: 10.3233/JAD-230331
8. Zenuni, H, Bovenzi, R, Bissacco, J, Grillo, P, Simonetta, C, Mascioli, D, et al. 13 clinical and neurochemical correlates of the APOE genotype in early-stage Parkinson’s disease. Neurobiol Aging. (2023) 131:24–8. doi: 10.1016/j.neurobiolaging.2023.07.011
9. Senkevich, K, Bandres-Ciga, S, Cisterna-García, A, Yu, E, Bustos, BI, Krohn, L, et al. Genome-wide association study stratified by MAPT haplotypes identifies potential novel loci in Parkinson’s disease. MedRxiv [Preprint]. (2023). doi: 10.1101/2023.04.14.23288478
10. Loureiro, D, Bilbao, R, Bordet, S, Grasso, L, Otero-Losada, M, Capani, F, et al. 15 a systematic review and meta-analysis on the association between orthostatic hypotension and mild cognitive impairment and dementia in Parkinson’s disease. Neurol Sci. (2023) 44:1211–22. doi: 10.1007/s10072-022-06537-3
11. Davis, AA, and Racette, B. Parkinson disease and cognitive impairment: Five new things. Neurol Clin Pract. (2016) 6:452–458. doi: 10.1212/CPJ.0000000000000285
12. He, J, and Gai, J. 1 genome-wide association studies (GWAS) In: Y Shavrukov, editor. Plant Genotyping: Methods and Protocols. United States: Springer US (2023). 123–46.
13. Tang, CS, Karim, A, Zhong, Y, Chung, PH, and Tam, PK. 2 genetics of Hirschsprung’s disease. Pediatr Surg Int. (2023) 39:104. doi: 10.1007/s00383-022-05358-x
14. Hautakangas, H, Winsvold, BS, Ruotsalainen, SE, Bjornsdottir, G, Harder, AVE, Kogelman, LJA, et al. 3 genome-wide analysis of 102,084 migraine cases identifies 123 risk loci and subtype-specific risk alleles. Nat Genet. (2022) 54:152–60. doi: 10.1038/s41588-021-00990-0
15. Qin, Y, Havulinna, AS, Liu, Y, Jousilahti, P, Ritchie, SC, Tokolyi, A, et al. 4 combined effects of host genetics and diet on human gut microbiota and incident disease in a single population cohort. Nat Genet. (2022) 54:134–42. doi: 10.1038/s41588-021-00991-z
16. Zhang, Q, Li, Q, Zhao, H, Shu, M, Luo, M, Li, Y, et al. 5 neurodegenerative disease and antioxidant biomarkers: a bidirectional Mendelian randomization study. Front Neurol. (2023) 14:1158366. doi: 10.3389/fneur.2023.1158366
17. Wightman, DP, Jansen, IE, Savage, JE, Shadrin, AA, Bahrami, S, Holland, D, et al. 17A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat Genet. 53:1276–82. doi: 10.1038/s41588-021-00921-z
18. Zhu, J, Liu, X, Yin, H, Gao, Y, and Yu, H. Convergent lines of evidence support BIN1 as a risk gene of Alzheimer’s disease. Hum Genom. (2021) 15:9. doi: 10.1186/s40246-021-00307-6
19. Tan, L, Wang, HF, Tan, MS, Tan, CC, Zhu, XC, Miao, D, et al. Effect of CLU genetic variants on cerebrospinal fluid and neuroimaging markers in healthy, mild cognitive impairment and Alzheimer’s disease cohorts. Sci Rep. (2016) 6:26027. doi: 10.1038/srep26027
20. Alzheimer’s Disease Neuroimaging InitiativeCao, H, Zhou, X, Chen, Y, Ip, FCF, Chen, Y, et al. Association of SPI1 haplotypes with altered SPI1 gene expression and Alzheimer’s disease risk. JAD. (2022) 86:1861–73. doi: 10.3233/JAD-215311
21. Uddin, MS, Kabir, MT, al Mamun, A, Abdel-Daim, MM, Barreto, GE, and Ashraf, GM. 18 APOE and Alzheimer’s disease: evidence mounts that targeting APOE4 may combat Alzheimer’s pathogenesis. Mol Neurobiol. (2019) 56:2450–65. doi: 10.1007/s12035-018-1237-z
22. Davis, AA, Inman, CE, Wargel, ZM, Dube, U, Freeberg, BM, Galluppi, A, et al. 19 APOE genotype regulates pathology and disease progression in synucleinopathy. Sci Transl Med. (2020) 12:eaay3069. doi: 10.1126/scitranslmed.aay3069
23. Camerucci, E, Stang, CD, Hajeb, M, Turcano, P, Mullan, AF, Martin, P, et al. 20 early-onset parkinsonism and early-onset Parkinson’s disease: a population-based study (2010-2015). J Parkinsons Dis. (2021) 11:1197–207. doi: 10.3233/JPD-202464
24. Logan, T, Simon, MJ, Rana, A, Cherf, GM, Srivastava, A, Davis, SS, et al. 21 rescue of a lysosomal storage disorder caused by Grn loss of function with a brain penetrant progranulin biologic. Cell. (2021) 184:4651–4668.e25. doi: 10.1016/j.cell.2021.08.002
25. Sassi, C, Guerreiro, R, Gibbs, R, Ding, J, Lupton, MK, Troakes, C, et al. 22 investigating the role of rare coding variability in Mendelian dementia genes (APP, PSEN1, PSEN2, GRN, MAPT, and PRNP) in late-onset Alzheimer’s disease. Neurobiol Aging. (2014) 35:2881.e1–6. doi: 10.1016/j.neurobiolaging.2014.06.002
26. Mehra, S, Sahay, S, and Maji, SK. 23 α-Synuclein misfolding and aggregation: implications in Parkinson’s disease pathogenesis. Biochim Biophys Acta Proteins Proteom. (2019) 1867:890–908. doi: 10.1016/j.bbapap.2019.03.001
27. Ren, P, Wang, JY, Chen, HL, Lin, XW, Zhao, YQ, Guo, WZ, et al. 24 diagnostic model constructed by nine inflammation-related genes for diagnosing ischemic stroke and reflecting the condition of immune-related cells. Front Immunol. (2022) 13:1046966. doi: 10.3389/fimmu.2022.1046966
28. Karaoz, E, Genç, ZS, Demircan, PÇ, Aksoy, A, and Duruksu, G. 25 protection of rat pancreatic islet function and viability by coculture with rat bone marrow-derived mesenchymal stem cells. Cell Death Dis. (2010) 1:e36. doi: 10.1038/cddis.2010.14
29. Marogianni, C, Sokratous, M, Dardiotis, E, Hadjigeorgiou, GM, Bogdanos, D, and Xiromerisiou, G. 26 neurodegeneration and inflammation—an interesting interplay in Parkinson’s disease. IJMS. (2020) 21:8421. doi: 10.3390/ijms21228421
30. Jinbo Cheng, BT, Liao, Y, Dong, Y, Han, H, Yang, N, Kong, X, et al. 27 microglial autophagy defect causes parkinson disease-like symptoms by accelerating inflammasome activation in mice. Autophagy. (2020) 16:2193–205. doi: 10.1080/15548627.2020.1719723
31. Perneel, J, Neumann, M, Heeman, B, Cheung, S, van den Broeck, M, Wynants, S, et al. 28 accumulation of TMEM106B C-terminal fragments in neurodegenerative disease and aging. Acta Neuropathol. (2023) 145:285–302. doi: 10.1007/s00401-022-02531-3
32. Cabron, A-S, Borgmeyer, U, Richter, J, Peisker, H, Gutbrod, K, Dörmann, P, et al. 29 lack of a protective effect of the Tmem106b “protective SNP” in the Grn knockout mouse model for frontotemporal lobar degeneration. Acta Neuropathol Commun. (2023) 11:21. doi: 10.1186/s40478-023-01510-3
Keywords: Alzheimer’s disease, Parkinson’s disease, Parkinson’s disease dementia, single nucleotide polymorphisms, Chinese Han cohort
Citation: Yan H, Liu M, Gao Y, Yuan Y, Liu X, Wang Y, Li L, Wang Q, Wang Y, Shi C, Xu Y and Yang J (2024) Assessing the impact of novel risk loci on Alzheimer’s and Parkinson’s diseases in a Chinese Han cohort. Front. Neurol. 15:1326692. doi: 10.3389/fneur.2024.1326692
Edited by:
Xueping Chen, Sichuan University, ChinaReviewed by:
Mariet Allen, Mayo Clinic Florida, United StatesDaniel Wickland, Mayo Clinic Florida, United States
Copyright © 2024 Yan, Liu, Gao, Yuan, Liu, Wang, Li, Wang, Wang, Shi, Xu and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuming Xu, eHV5dW1pbmdAenp1LmVkdS5jbg==; Jing Yang, eWFuZ2ppbmc5NTI3QDEyNi5jb20=