
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Neurol. , 22 August 2022
Sec. Dementia and Neurodegenerative Diseases
Volume 13 - 2022 | https://doi.org/10.3389/fneur.2022.675301
This article is part of the Research Topic Frontotemporal Dementia and its Spectrum in Latin America and the Caribbean: a Multidisciplinary Perspective View all 21 articles
 Andrea López-Cáceres1,2*
Andrea López-Cáceres1,2* Francy Cruz-Sanabria3,4
Francy Cruz-Sanabria3,4 Pilar Mayorga5
Pilar Mayorga5 Ana Isabel Sanchez6,7
Ana Isabel Sanchez6,7 Silvia Gonzalez-Nieves8
Silvia Gonzalez-Nieves8 Paola Ayala-Ramírez1
Paola Ayala-Ramírez1 Ignacio Zarante1
Ignacio Zarante1 Diana Matallana5,9
Diana Matallana5,9Frontotemporal dementia (FTD) is a heterogeneous neurodegenerative disease of presenile onset. A better characterization of neurodegenerative disorders has been sought by using tools such as genome-wide association studies (GWAS), where associations between single nucleotide polymorphisms (SNPs) and cognitive profiles could constitute predictive biomarkers for these diseases. However, in FTD, associations between genotypes and cognitive phenotypes are yet to be explored. Here, we evaluate a possible relationship between genetic variants and some cognitive functions in an FTD population.
Methodology: A total of 47 SNPs in genes associated with neurodegenerative diseases were evaluated using the Sequenom MassARRAY platform along with their possible relationship with performance in neuropsychological tests in 105 Colombian patients diagnosed with FTD.
Results and discussion: The SNPs rs429358 (APOE), rs1768208 (MOBP), and rs1411478 (STX6), were identified as risk factors for having a low cognitive performance in inhibitory control and phonological verbal fluency. Although the significance level was not enough to reach the corrected alpha for multiple comparison correction, our exploratory data may constitute a starting point for future studies of these SNPs and their relationship with cognitive performance in patients with a probable diagnosis of FTD. Further studies with an expansion of the sample size and a long-term design could help to explore the predictive nature of the potential associations we identified.
Frontotemporal dementia (FTD) is an early-onset, heterogeneous neurodegenerative disorder with a strong genetic component (1). Positive family history has been reported in FTD in up to 40% of cases (2, 3), with the most frequent mutations found in the following genes: microtubule-associated protein tau (MAPT), granulin (GRN), and C9orf72 (2, 4). According to clinical involvement, FTD is classified into behavioral and language variants (semantic dementia, primary progressive aphasia) (5–7). It also coexists with motor neuron disease (FTD-MND) and atypical parkinsonian disorders (1, 8, 9).
The clinical and molecular heterogeneity of FTD, as well as the overlapping of symptoms with other neurodegenerative diseases (1, 2), have led to it being characterized through genome-wide association studies (GWAS) (10, 11). These typically involve the use of single nucleotide polymorphisms (SNPs) that are common in a given population and can establish risk by association with different phenotypes related to the onset, development, and progression of the disease (10, 12, 13). More than 40 risk loci have been identified for dementia within the genome (10), reporting the APOE ε4 allele with the strongest risk factor for late-onset alzheimer's disease (AD), and as a modulator of the expression of other degenerative dementias (14, 15). Specifically for FTD, three significant SNPs (rs6966915, rs1020004, and rs1990622) have been reported in the transmembrane protein 106B (TMEM106B) gene (7p21.3), a protein involved in endolysosomal transport and in the modulation of GRN protein levels (10, 16). Besides, some other loci such as 6p21.3, encompassing HLA locus, and 11q14 encompassing RAB38/CTSC were statistically significant in GWAS for FTD (10, 11).
An association between risk polymorphisms and cognitive profiles in mild cognitive impairment (MCI), AD, FTD, and amyotrophic lateral sclerosis (ALS) have recently been explored to evaluate disease development and progression (9, 17). The association between the studied loci and deficits in cognitive processes such as executive functions, language, visuospatial skills, and memory have been found in the four diseases (12). The association between polymorphic variants and cognitive performance suggests that exploring this may be a useful measure to detect risk variants that could eventually be considered predictive biomarkers for neurodegenerative diseases. It also makes it possible to evaluate disease development and progression (16, 18, 19). Thus, our study's main focus is to evaluate a relationship between cognitive performance and SNPs associated with neurodegenerative diseases in a sample of Colombian patients with a probable diagnosis of FTD.
An analytical, observational, non-probabilistic convenience study was conducted between January 2012 and December 2014 in 105 patients with a probable diagnosis of FTD, determined through consensus by a multidisciplinary group of specialists (Neurologist, Geriatrician, Psychiatrist, and Neuropsychologist) at the Memory Clinic at Hospital Universitario San Ignacio (Bogotá, Colombia). FTD patients were diagnosed according to established criteria in the behavioral variant of FTD (bv-FTD), non-fluent/agrammatic-variant primary progressive aphasia (nfvPPA), and semantic-variant primary progressive aphasia (svPPA) (6, 7), following the guidelines developed by an International Consortium for the Diagnosis of Frontotemporal Dementia (5, 7). Exclusion criteria include visual and hearing impairments, severe alteration of mobility, delirium, absence of caregiver or informant, significant cerebrovascular disease, and other previously recognized neurological diseases. This study was approved by the ethics committee at Hospital Universitario San Ignacio and Pontificia Universidad Javeriana. All participants received and signed informed consent.
A total of seven validated neuropsychological tests (see Table 1) on memory, praxis, verbal fluency, attention, and executive function were used to assess the cognitive profile of each patient (20–22, 28, 29). Taking into account as a reference the normative data for the tests of the Neuronorma Colombia neuropsychological evaluation battery (22, 29). The values obtained were converted to scale scores and subsequently dichotomized into 1 and 0. Performances that were lower than expected, considering age and education with respect to population parameters (percentile ≤ 6), were coded as 1, while performances above said percentile were coded as 0 (22, 29).
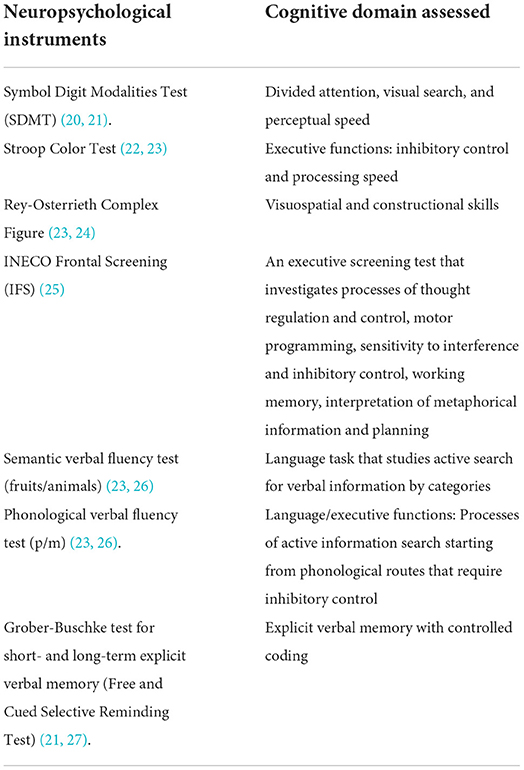
Table 1. Neuropsychological tests.
All evaluated patients had a 3-cc blood sample taken in EDTA tubes from which the genomic DNA was extracted using the Salting Out protocol. The DNA was then quantified using a NanoDrop® ND-1000 spectrophotometer. SNP genotyping was performed using a custom-designed panel on the Sequenom MassARRAY platform, developed at the University of Pennsylvania, in which 47 SNP-type genetic variants were evaluated in genes associated with neurodegenerative diseases (16, 23, 24), FTD, Alzheimer's disease (AD), amyotrophic lateral sclerosis (ALS), Parkinson's disease, and progressive supranuclear palsy (PSP) (see Annex 1) (16, 23). The assay consisted of an initial locus-specific PCR reaction, followed by single-base extension using mass-modified dideoxynucleotide terminators of an oligonucleotide primer which anneals immediately upstream of the polymorphic site of interest (25). Although not all included SNPs are relevant for FTD, it is more cost and time-efficient to use a single panel that can be applied generally since there is a significant overlap in neurodegenerative disease phenotypes (13, 26, 27, 30).
The clinical and sociodemographic characteristics were analyzed by calculating frequencies and central tendency measures (median-range). ANOVA and chi-squared tests were used to determine group differences in sociodemographic variables.
The allelic and genotypic frequencies were calculated by the counting method, and the Hardy-Weinberg (HW) equilibrium was determined for each SNP with the Arlequin v.3.5 software. The allelic frequencies obtained in the study were compared with the allelic frequencies reported in the 1,000 Genomes Global and in 1,000 Genomes Colombia samples by the χ2 association test, reporting their respective p-value (Stata/MP 14.0).
In order to identify a possible relationship between performance in cognitive tests and the SNPs assessed, a logistic regression model was calculated for each test and for each genetic variant in the R software. For all statistical tests, an alpha value of 0.05 was established. Based on these models, those SNPs that could significantly predict performance in each neuropsychological test were identified. To reduce Type I error for multiple comparisons, p-values were subjected to the Bonferroni correction with n = 47.
Similarly, the odds ratio (OR) of the allele related to these results was reported for each of the identified SNPs. Alleles with an OR < 1 were interpreted as being associated with adequate performance in the test, while ORs > 1 was associated with the risk of poor performance in neuropsychological tests.
Of the 105 patients with FTD, 61 patients met the criteria for bv-FTD, 28 met the criteria for PPA, and 16 patients met the criteria for SD. The median age of patients at the time of diagnosis was 61 years (range 40–86 years). No sex differences were found in the total sample or inside each clinical variant (see Table 2). As for the patient's education level, only 8.5% (9) had primary education, 19.2% (28) completed secondary education, and 35.6% (31) had undertaken university studies. It was not possible to determine the education level of 38 patients.

Table 2. Sociodemographic characteristics.
We found four SNPs that were not in HW equilibrium: rs7412 in APOE (p-value = 0.029), rs6656401 in CR1 (p-value = 0.024), rs983392 in MS4A6A (p-value = 0.009), and rs1411478 in STX6 (p-value = 0.014). The first three SNPs are associated with AD and the last one with progressive supranuclear palsy (PSP). We also determined the frequency of the minor or risk allele in the SNPs associated with FTD and AD. By comparing them with the 1,000 Genomes Colombia and 1,000 Genomes Global samples, we reported a statistically significant difference in rs12546767 in KIAA0196 gen (p-value < 0.001, p-value < 0.00001, respectively) (see Appendix 2).
Sixteen polymorphisms were significantly correlated with performance in one (or more than one) neuropsychological test (p-value < 0.05). Three of the alleles that were identified with a risk of poor performance in these tests correspond to the minor allele. A higher risk of poor performance in the phonological verbal fluency task was found for the STX6 rs1411478 A allele. Similarly, MOBP rs1768208 T allele and APOE rs429358 C allele were identified as risk factors for poor performance in the Stroop Color Test (see Table 3). However, these findings did not survive the Bonferroni correction.
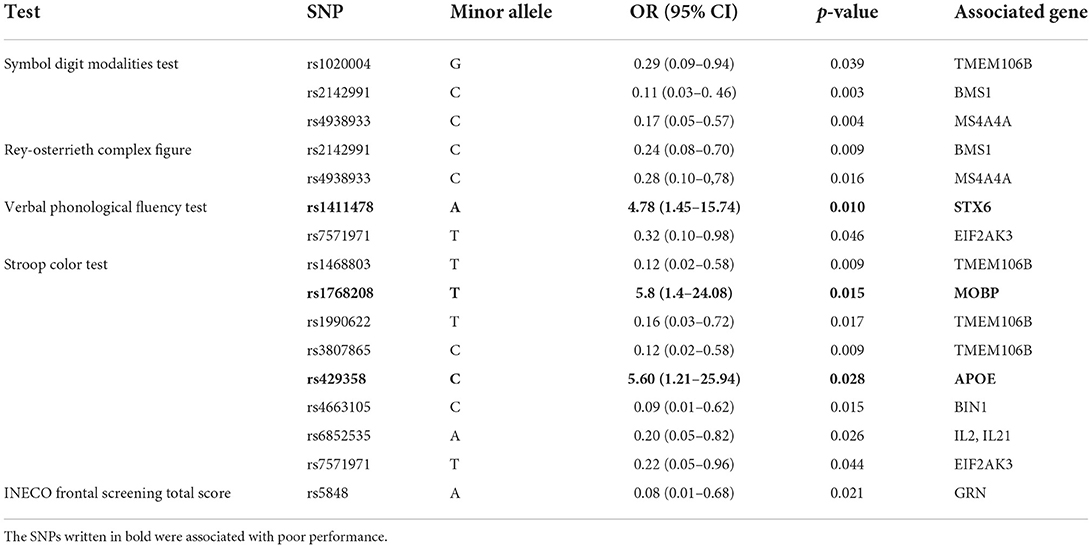
Table 3. OR values for SNPs with significant associations with neuropsychological test results.
Frontotemporal dementia is a heterogeneous disease in both its clinical and genetic components (32–34). We find that the sociodemographic characteristics of this cohort of patients were consistent with what was reported in the literature. BvFTD was the most common clinical variant followed by language variants (35, 36). Regarding distribution by sex and the incidence of disease, there was no significant difference between the clinical groups (11, 26, 31). All the SNPs associated with FTD reached HW equilibrium in our population. The SNPs that did not reach HW equilibrium in our cohort of patients with a clinical diagnosis of FTD were located in genes associated with AD and PSP (APOE, CR1, MS4A6A, and STX6 genes), which can be explained due to the sample size, or because these genes are subject to selection with each other between FTD and other neurodegenerative diseases (12, 37).
In addition, we compared the allele frequencies in our FTD sample with 1,000 Genomes and 1,000 Genomes Colombia populations, and we found that KIAA0196 rs12546767 showed higher frequency in our sample, supporting the findings reported in previous studies in which this SNP has an increased disease association signal in the combined ALS and FTD (38).
Regarding correlations between cognitive performance and the SNP array panel, carriers of the APOE rs429358C allele and MOBP rs1768208T allele showed deficits in inhibitory control. Furthermore, carriers of STX6 rs1411478A allele performed poorly in phonological verbal fluency. Although the significant level of the identified risk between these alleles and the cognitive performance was not enough to reach the corrected alpha for multiple comparison correction, this information should not underrate because in studies with a larger sample with longitudinal data associations, associations with different SNPs and cognitive performance have been found, as in studies with a cumulative score, combining more than one allele (16, 38, 39).
For our three alleles, few studies are found related to cognitive performance; To MOBP rs1768208T allele, Massimo and col. found that this allele is associated with a disruption of white matter networks in frontal regions, whereby MOBP rs1768208T + individuals demonstrated faster rates of decline in executive function through time (16). Moreover, MOBP rs1768208 has been independently identified as a risk factor in confirmed cases of corticobasal degeneration (DCB) and in cases of PSP (40). Literature on the APOE rs429358 and cognitive processes yields variable results: some studies conducted in healthy adults have found associations with deficits in naming and orientation skills (41), while others have described better cognitive performances measured by the Mini-Mental State Examination (MMSE) (42). Specifically for APOE rs429358C, there are no association studies with neurocognitive tests, but Xue-Bin Li et al. suggest that the APOE rs429358C allele genotype is associated with an increased risk of developing post-stroke depression, and may be detrimental to the recovery of nerve function after stroke (43). For STX6, there are no studies of its association with cognitive performance in this SNP to date. However, Ferrari and col. demonstrated that the rs1411478A allele has a significantly lower expression of STX6 in white matter but not in any other brain region in PSP (23). As mentioned previously, MOBP rs1768208T and STX6 rs1411478A alleles have been associated with disrupting white matter. This has revealed that cognitive performances are related to cortical thickness in frontotemporal regions and degradation in white matter integrity (35, 44, 45).
In conclusion, as no preliminary studies have been performed regarding the associations between cognitive performance and these SNPs in FTD, these results highlight the value of incorporating multiple biomarkers to help disentangle the mechanistic heterogeneity of cognitive decline (46). Our results may constitute a starting point for future studies involving these SNPs and their relationship with cognitive performance in patients with a probable diagnosis of FTD.
The original contributions presented in the study are included in the article. Requests to access the datasets should be directed to http://hdl.handle.net/10554/61347 or to the corresponding authors.
The studies involving human participants were reviewed and approved by Ethics Committee of the Pontificia Universidad Javeriana in Bogotá, Colombia approved the study. The patients/participants provided their written informed consent to participate in this study.
AL-C, FC-S, PM, and PA-R developed the study concept and the study design. AL-C, FC-S, and PM performed testing and data collection. AL-C, FC-S, PM, AS, SG-N, and PA-R data analysis and interpretation under the supervision of IZ and DM. AL-C, FC-S, PM, AS, SG-N, PA-R, IZ, and DM drafted the manuscript. DM and IZ provided critical revisions. All authors contributed to the article and approved the submitted version.
US-South American Initiative for Genetic-Neural-Behavioral Interactions in Human Neurodegenerative Research (R01AG057234-01A1), The Institute of Memory and Cognition (INTELLECTUS) the National Program of Science, Technology and Innovation in Health APS. This work was supported by, Colciencias 371-2011, 370-201 code: 120354531693.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fneur.2022.675301/full#supplementary-material
1. Seltman RE, Matthews BR. Frontotemporal lobar degeneration: epidemiology, pathology, diagnosis and management. CNS Drugs. (2012) 26:841–70. doi: 10.2165/11640070-000000000-00000
2. Takada LT. The genetics of monogenic frontotemporal dementia. Dement Neuropsychol. (2015) 9:219–29. doi: 10.1590/1980-57642015dn93000003
3. Onyike CU, Diehl-Schmid J. The epidemiology of frontotemporal dementia. Int Rev Psychiatry. (2013) 25:130–7. doi: 10.3109/09540261.2013.776523
4. Rademakers R, Neumann M, Mackenzie IR. Advances in understanding the molecular basis of frontotemporal dementia. Nat Rev Neurol. (2012) 8:423–34. doi: 10.1038/nrneurol.2012.117
5. Rascovsky K, Hodges JR, Knopman D, Mendez MF, Kramer JH, Neuhaus J, et al. Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain. (2011) 134:2456–77. doi: 10.1093/brain/awr179
6. Ghosh S, Lippa CF. Clinical subtypes of frontotemporal dementia. Am J Alzheimer's Dis Other Dementiasr. (2015) 30:653–61. doi: 10.1177/1533317513494442
7. Gorno-Tempini ML, Hillis AE, Weintraub S, Kertesz A, Mendez M, Cappa SF, et al. Classification of primary progressive aphasia and its variants. Neurology. (2011) 76:1006–14. doi: 10.1212/WNL.0b013e31821103e6
8. Boeve BF, Boylan KB, Graff-Radford NR, Dejesus-Hernandez M, Knopman DS, Pedraza O, et al. Characterization of frontotemporal dementia and/or amyotrophic lateral sclerosis associated with the GGGGCC repeat expansion in C9ORF72. Brain. (2012) 135:765–83. doi: 10.1016/j.jalz.2012.05.2129
9. Edwards TL, Scott WK, Almonte C, Burt A, Powell EH, Beecham GW, et al. Genome-Wide association study confirms SNPs in SNCA and the MAPT region as common risk factors for parkinson disease. Ann Hum Genet. (2010) 74:97–109. doi: 10.1111/j.1469-1809.2009.00560.x
10. Ibanez L, Farias FHG, Dube U, Mihindukulasuriya KA, Harari O. Polygenic risk scores in neurodegenerative diseases: a review. Curr Genet Med Rep. (2019) 7:22–9. doi: 10.1007/s40142-019-0158-0
11. Ferrari R, Hernandez DG, Nalls MA, Rohrer JD, Ramasamy A KJ, Dobson-Stone C, et al. Frontotemporal dementia and its subtypes: a genome-wide association study. Lancet Neurol. (2014) 13:686–99. doi: 10.1016/S1474-4422(14)70065-1
12. Hagenaars SP, Radaković R, Crockford C, Fawns-Ritchie C, Harris SE, Gale CR, et al. Genetic risk for neurodegenerative disorders, and its overlap with cognitive ability and physical function Chloe Fawns-Ritchie 1,2, International FTD-Genomics Consortium (IFGC). PLoS ONE. (2018) 13:e0198187. doi: 10.1371/journal.pone.0198187
13. Raffaele F, Claudia M, John H. Genetics and molecular mechanisms of frontotemporal lobar degeneration: an update and future avenues. Neurobiol Aging. (2019) 78:98–110. doi: 10.1016/j.neurobiolaging.2019.02.006
14. Boccardi M, Sabattoli F, Testa C, Beltramello A, Soininen H, Frisoni GB. APOE and modulation of Alzheimer's and frontotemporal dementia. Neurosci Lett. (2004) 356:167–70. doi: 10.1016/j.neulet.2003.11.042
15. Pastor P, Roe CM, Villegas A, Bedoya G, Chakraverty S, Garcia G, et al. Apolipoprotein Eepsilon4 modifies Alzheimer's disease onset in an E280A PS1 kindred. Ann Neurol. (2003) 54:163–9. doi: 10.1002/ana.10636
16. Massimo L, Rennert L, Xie SX, Olm C, Bove J, Van Deerlin V, et al. Common genetic variation is associated with longitudinal decline and network features in behavioral variant frontotemporal degeneration. Neurobiol Aging. (2021) 108:16–23. doi: 10.1016/j.neurobiolaging.2021.07.018
17. Cruz-Sanabria F, Bonilla-Vargas K, Estrada K, Mancera O, Vega E, Guerrero E, et al. Analysis of cognitive performance and polymorphisms of SORL1, PVRL2, CR1, TOMM40, APOE, PICALM, GWAS_14q, CLU, and BIN1 in patients with mild cognitive impairment and cognitively healthy controls. Neurologia. (2018) 36:1–10. doi: 10.1016/j.nrleng.2018.07.012
18. Karch CM, Goate AM. Alzheimer's disease risk genes and mechanisms of disease pathogenesis. Biol Psychiatry. (2014) 77:1–9. doi: 10.1016/j.biopsych.2014.05.006
19. Pedraza O, Allen M, Jennette K, Carrasquillo M, Crook J, Serie D, et al. Evaluation of memory endophenotypes for association with CLU,CR1 and PICALM variants in African-American and caucasian subjects. Alzheimers Dement. (2014) 10:205–13. doi: 10.1016/j.jalz.2013.01.016
20. Duarte, Pedroza L. Lenguaje, memoria y habilidades visuoconstructivas en el envejecimiento normal: Datos normativos con la Batería Neuronorma Colombia. Doctoral dissertation, Universidad Nacional de Colombia-Bogotá. Available online at: https://repositorio.unal.edu.co/handle/unal/59359
21. Torralva T, Roca M, Gleichgerrcht E, Lopez P, Manes F. INECO Frontal Screening (IFS): a brief, sensitive, and specific tool to assess executive functions in dementia. J Int Neuropsychol Soc. (2009) 15:777–86. doi: 10.1017/S1355617709990415
22. Fiorentino N, Gleichgerrcht E, Roca M, Cetkovich M, Manes F, Torralva T. The INECO Frontal Screening tool differentiates behavioral variant - frontotemporal dementia (bv-FTD) from major depression. Dement Neuropsychol. (2013) 7:33–9. doi: 10.1590/S1980-57642013DN70100006
23. McMillan CT, Toledo JB, Avants BB, Cook PA, Wood EM, Suh E, et al. Genetic and neuroanatomic associations in sporadic frontotemporal lobar degeneration. Neurobiol Aging. (2014) 35:1473–82. doi: 10.1016/j.neurobiolaging.2013.11.029
24. Suh ER, Lee EB, Neal D, Wood EM, Toledo JB, Rennert L, et al. Semi-automated quantification of C9orf72 expansion size reveals inverse correlation between hexanucleotide repeat number and disease duration in frontotemporal degeneration. Acta Neuropathol. (2015) 130:363–72. doi: 10.1007/s00401-015-1445-9
25. Gabriel S, Ziaugra L, Tabbaa D. SNP Genotyping using the sequenom massARRAY iPLEX Platform. Curr Protoc Hum Genet. (2009) 60:2–12. doi: 10.1002/0471142905.hg0212s60
26. Ramos EM, Koros C, Dokuru DR, Van Berlo V, Kroupis C, Wojta K, et al. Frontotemporal dementia spectrum: first genetic screen in a Greek cohort. Neurobiol Aging. (2019) 75:224.e1–224.e8. doi: 10.1016/j.neurobiolaging.2018.10.029
27. Hoglinger GU, Melhem NM, Dickson DW, Sleiman PMA, Wang L-S, Klei L, et al. Identification of common variants influencing risk of the tauopathy progressive supranuclear palsy. Nat Genet. (2011) 43:699–705. doi: 10.1038/ng.859
28. Hernández L, Montañés P, Gamez A, Cano C, Nuñez E. Neuropsicología del envejecimiento normal. Rev la Asoc Colomb Gerontol y Geriatría. (2007) 21:992–1004.
29. Espitia, Mendieta A. Funciones ejecutivas en el envejecimiento normal: datos normativos con la batería Neuronorma. Doctoral dissertation, Universidad Nacional de Colombia-Bogotá. Available online at: https://repositorio.unal.edu.co/handle/unal/59390
30. Deerlin VM, Van Sleiman PMA, Martinez-lage M, Chen- A, Wang L, Graff-radford NR, et al. Common variants at 7p21 are associated with with TDP-43 inclusions. Nat Genet. (2010) 42:234–9. doi: 10.1038/ng.536
31. Shpilyukova YA, Fedotova EY, Illarioshkin SN. Genetic diversity in frontotemporal dementia. Mol Biol (Mosk). (2020) 54:13–23. doi: 10.1134/S0026893320010136
32. López-Cáceres A, Velasco-Rueda M, Garcia-Cifuentes E, Zarante I, Matallana D. Analysis of heritability across the clinical phenotypes of frontotemporal dementia and the frequency of the C9ORF72 in a Colombian population. Front Neurol. (2021) 12:1–6. doi: 10.3389/fneur.2021.681595
33. Kelley RE, El-Khoury R. Frontotemporal dementia. Neurol Clin. (2016) 34:171–81. doi: 10.1016/j.ncl.2015.08.007
34. Benussi A, Padovani A, Borroni B. Phenotypic heterogeneity of monogenic frontotemporal dementia. Front Aging Neurosci. (2015) 7:1–19. doi: 10.3389/fnagi.2015.00171
35. Greaves C V, Rohrer JD. An update on genetic frontotemporal dementia. J Neurol. (2019) 266:2075–86. doi: 10.1007/s00415-019-09363-4
36. Waldö ML. The frontotemporal dementias. Psychiatr Clin North Am. (2015) 38:193–209. doi: 10.1016/j.psc.2015.02.001
37. Li Y, Graubard BI. Testing hardy-weinberg equilibrium and homogeneity of hardy-weinberg disequilibrium using complex survey data. Biometrics. (2009) 65:1096–104. doi: 10.1111/j.1541-0420.2009.01199.x
38. Diekstra FP, Van Deerlin VM, Van Swieten JC, Al-Chalabi A, Ludolph AC, Weishaupt JH, et al. C9orf72 and UNC13A are shared risk loci for amyotrophic lateral sclerosis and frontotemporal dementia: a genome-wide meta-analysis. Ann Neurol. (2014) 76:120–33. doi: 10.1002/ana.24198
39. Laukka EJ, Köhncke Y, Papenberg G, Fratiglioni L, Bäckman L. Combined genetic influences on episodic memory decline in older adults without dementia. Neuropsychology. (2020) 34:654–66. doi: 10.1037/neu0000637
40. Yokoyama JS, Karch CM, Fan CC, Bonham LW, Kouri N, Ross OA, et al. Shared genetic risk between corticobasal degeneration, progressive supranuclear palsy, and frontotemporal dementia HHS public access. Acta Neuropathol. (2017) 133:825–37. doi: 10.1007/s00401-017-1693-y
41. Zhen J, Huang X, Van Halm-Lutterodt N, Dong S, Ma W, Xiao R, et al. ApoE rs429358 and rs7412 Polymorphism and gender differences of serum lipid profile and cognition in aging chinese population. Front Aging Neurosci. (2017) 9:248. doi: 10.3389/fnagi.2017.00248
42. Prada D, Colicino E, Power MC, Cox DG, Weisskopf MG, Hou L, et al. Influence of multiple APOE genetic variants on cognitive function in a cohort of older men - results from the Normative Aging Study. BMC Psychiatry. (2014) 14:1–9. doi: 10.1186/s12888-014-0223-x
43. Li X, Bin Wang J, Xu AD, Huang JM, Meng LQ, Huang RY, et al. Apolipoprotein E polymorphisms increase the risk of post-stroke depression. Neural Regen Res. (2016) 11:1790–6. doi: 10.4103/1673-5374.194748
44. Rowley J, Fonov V, Wu O, Eskildsen SF, Schoemaker D, Wu L, et al. White matter abnormalities and structural hippocampal disconnections in amnestic mild cognitive impairment and Alzheimer's disease. PLoS ONE. (2013) 8:e74776. doi: 10.1371/journal.pone.0074776
45. Bouchard RW. Diagnostic criteria of dementia. Can J Neurol Sci. (2007) 34:S11–8. doi: 10.1017/S0317167100005497
Keywords: frontotemporal dementia, neuropsychological tests, cognition, SNP array, neurodegenerative disease
Citation: López-Cáceres A, Cruz-Sanabria F, Mayorga P, Sanchez AI, Gonzalez-Nieves S, Ayala-Ramírez P, Zarante I and Matallana D (2022) Association between risk polymorphisms for neurodegenerative diseases and cognition in colombian patients with frontotemporal dementia. Front. Neurol. 13:675301. doi: 10.3389/fneur.2022.675301
Received: 02 March 2021; Accepted: 18 July 2022;
Published: 22 August 2022.
Edited by:
Sonia Maria Dozzi Brucki, University of São Paulo, BrazilReviewed by:
Benedetta Nacmias, University of Florence, ItalyCopyright © 2022 López-Cáceres, Cruz-Sanabria, Mayorga, Sanchez, Gonzalez-Nieves, Ayala-Ramírez, Zarante and Matallana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea López-Cáceres, YWxvcGV6Y2FjZXJlc0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.