
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Neurol. , 11 January 2023
Sec. Neuromuscular Disorders and Peripheral Neuropathies
Volume 13 - 2022 | https://doi.org/10.3389/fneur.2022.1098857
This article is part of the Research Topic Surgical Treatment of Peripheral Neuropathic Pain, Peripheral Nerve Tumors, and Peripheral Nerve Injury View all 25 articles
 Jing Fu1Xigong Li2Feilu Jin3
Jing Fu1Xigong Li2Feilu Jin3 Yanzhao Dong2
Yanzhao Dong2 Haiying Zhou2
Haiying Zhou2 Ahmad Alhaskawi2
Ahmad Alhaskawi2 Zewei Wang4
Zewei Wang4 Jingtian Lai4
Jingtian Lai4 Chengjun Yao4
Chengjun Yao4 Sohaib Hasan Abdullah Ezzi5
Sohaib Hasan Abdullah Ezzi5 Vishnu Goutham Kota2
Vishnu Goutham Kota2 Mohamed Hasan Abdulla Hasan Abdulla2Bin Chen2
Mohamed Hasan Abdulla Hasan Abdulla2Bin Chen2 Hui Lu2,6*
Hui Lu2,6*Peripheral nerve diseases are significantly correlated with severe fractures or trauma and surgeries, leading to poor life quality and impairment of physical and mental health. Human dental pulp stem cells (DPSCs) are neural crest stem cells with a strong multi-directional differentiation potential and proliferation capacity that provide a novel cell source for nerve regeneration. DPSCs are easily extracted from dental pulp tissue of human permanent or deciduous teeth. DPSCs can express neurotrophic and immunomodulatory factors and, subsequently, induce blood vessel formation and nerve regeneration. Therefore, DPSCs yield valuable therapeutic potential in the management of peripheral neuropathies. With the purpose of summarizing the advances in DPSCs and their potential applications in peripheral neuropathies, this article reviews the biological characteristics of DPSCs in association with the mechanisms of peripheral nerve regeneration.
Peripheral neuron degeneration, inflammation, and necroptosis caused by trauma, diabetes, and neurodegenerative disorders may cause motor-sensory dysfunctions (1–3). Therefore, the current therapeutic regimen mainly focuses on neuron regeneration and function restoration in post-traumatic events.
Conventional therapies have limited efficacy in restoring nerve function since the regeneration of neurons, and glial cells require sufficient neuronal precursor cells, which are absent or lacking in the mature nervous system (4). Stem cell-based therapies bring new insight into the biotherapy of peripheral neuropathies, providing adequate cell sources capable of self-renewal and multi-directional differentiation (4–8). However, challenges remain in the mass production of autografts or autologous cells for sufficient nerve regeneration (5–8).
Derived from the neural crest, DPSCs yield great potential in differentiation into neurons, expression of various neurotrophic factors for axonal regeneration, and functions of immunomodulation, indicating that DPSCs are an ideal cell source for peripheral nerve regeneration (5–7). In this review, we will summarize the biological characteristics of DPSCs and their respective application in animal models of peripheral neuropathies, with a focus on their regenerative mechanisms for future application.
Dental pulp, identified as a typical soft tissue, is rich in blood vessels, nerves, and mesenchymal tissue. Dental pulp has a central role in the development of primary and secondary teeth and further maintenance throughout life (7, 9, 10). Gronthos et al. (11) first described that DPSCs were initially discovered from the third molar dental pulp, which is later found in other dental pulps including deciduous teeth, permanent teeth, and supernumerary teeth. DPSCs display fibroblast-like morphology with higher proliferation capacity but lack specific surface biomarkers (Table 1) (12–20). While highly express MSC-like phenotypic biomarkers including CD29, CD90, and CD73 (12, 13), DPSCs are also found to express stemness-related markers such as Oct-4, Nanog, and Sox-2 (14, 15), and cytoskeleton-related markers such as Nestin and Vimentin (15). Moreover, several studies have demonstrated the expression of cranial neural crest cell-related neural markers by DPSCs, including glial fibrillary acidic protein (GFAP), β-III tubulin, and microtubule-associated protein-2 (MAP-2) (15, 16, 21). Recently, several special markers are proposed to distinguish DPSCs from gingiva-derived mesenchymal stem cells (GMSCs) including Calreticulin, Annexin A5, and Rho GDP dissociation inhibitor alpha (17). Furthermore, a recent study by Lei et al. (18) demonstrated that the CD271 is the most effective stem cell surface marker for dental mesenchymal stem cells (DMSCs), which display high odontogenic potential.
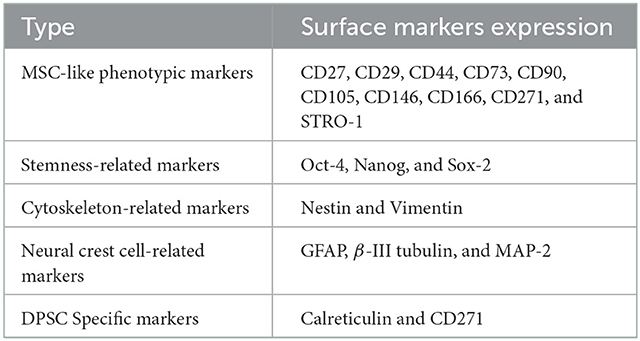
Table 1. The features of DPSCs.
DPSCs have shown the potential of multiple differentiations, with promising therapeutic value as bioengineered autografts for different tissue repair (Figure 1). Therefore, DPSCs can be applied in the biotherapy for a variety of peripheral neuropathies (5–7, 19, 20, 22–26). Studies show that DPSCs could induce restoration in peripheral nerves via three mechanisms, which include neuronal differentiation, paracrine and immunomodulatory effects (Figure 2) (5–7, 19, 20, 22–26).
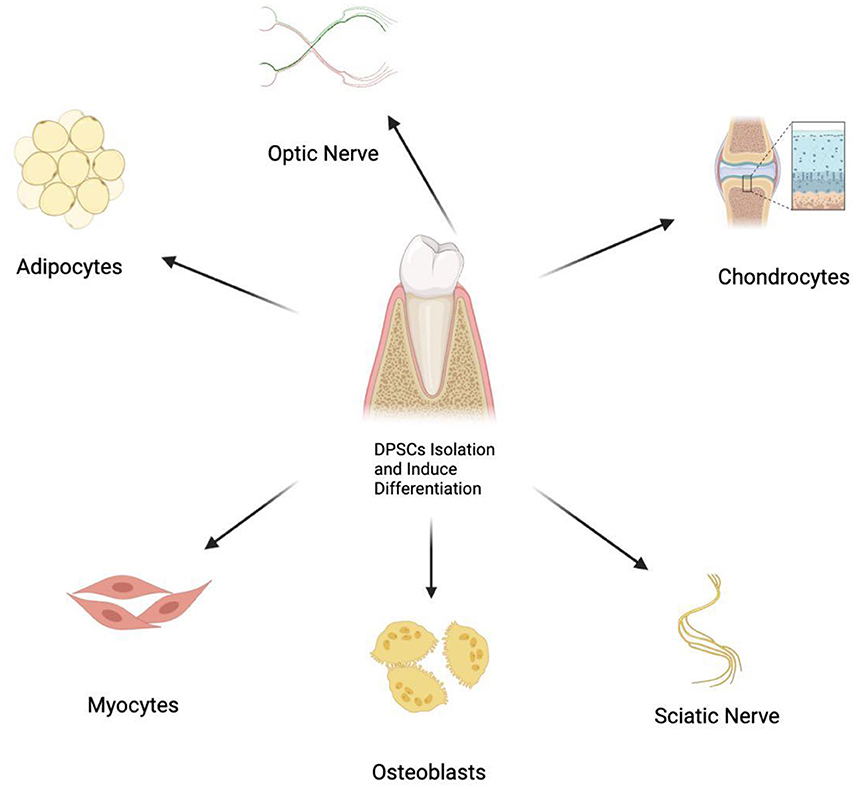
Figure 1. Multiple differentiation potential of DPSCs into various cell types.
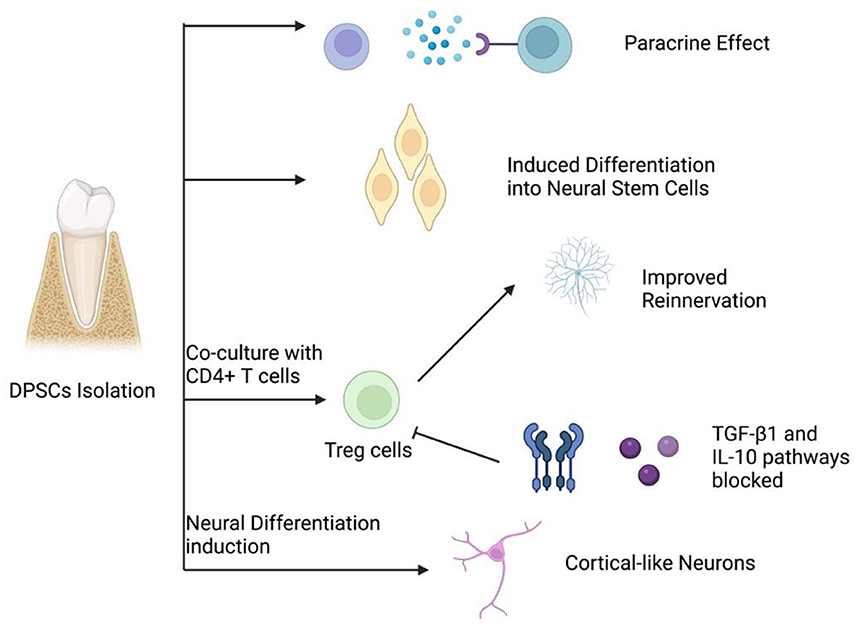
Figure 2. Four main mechanisms of peripheral nerve regeneration by DPSCs.
DPSCs can directly differentiate into neuron-like cells and express early neural markers (such as Nestin) (15). These neuron-like cells could migrate to the lesion sites and subsequently participate in the nerve regeneration process (5–7). Some studies have also shown that during migration, the transplanted DPSCs can recruit endogenous neural stem cells for tissue reconstruction (20). In addition, Kiraly et al. (15) revealed that the induction and differentiation of DPSCs are promoted by activating the intracellular cyclic adenosine phosphate signaling pathway. Chen et al. (27) proposed that the elevated intracellular cyclic adenosine phosphate can activate protein kinase A (PKA), which then up-regulate regeneration-related genes, such as arginase I, and promotes peptide synthesis. On the other hand, PKA also inhibits the Rho protein activation induced by myelin and induce nerve regeneration. Heng et al. (28) suggest that EphrinB2 signaling can modulate the neural differentiation of DPSCs, while EphB4 -inhibition in DPSCs could significantly up-regulate expression of the neural markers microtubule-associated protein 2, Musashi1, NGN2, and neuron-specific enolase. In 2018 Urraca et al. (29) described DPSC-derived neurons expressing GABAA and MAP2 genes, which is previously absent in undifferentiated DPSC and may provide future neurogenetic research with a useful tool.
Several studies have indicated that DPSCs could participate in the process of nerve repair in a paracrine manner (10, 20, 30). DPSCs can express brain-derived neurotrophic factor (BDNF), glia cell-line derived neurotrophic factor (GDNF), NGF, and neurotrophin-3 (NT3) at a substantial level, and exert neuroprotective functions in the process of peripheral nerve regeneration (10). Some researchers propose that the effects of these neurotrophic factors on neural cells are achieved via the intervention of PI3K/AKT signaling pathway. Even in the presence of neurotrophic factors such as NGF and GDNF, inhibition of PI3K or AKT activation can result in neuron apoptosis or necroptosis (31). The Erk signaling pathway plays a essential role in axonal growth stimulation by modulating neurotrophic factors (32). Additionally, the neurotrophin expression in DPSCs isenhanced in certain neural inductive conditions (33). The conditioned medium of DPSCs has been shown to increase Schwann cell proliferation rate while inducing neurite growth in vitro (31, 32). Previous studies revealed that the DPSC-derived secretome or DPSC conditional medium, which includes soluble factors and extracellular vesicles, proved to be therapeutically relevant in the management of neurodegenerative disorders and nerve injuries via the regulation of several processes, including neuroprotection, anti-inflammation, anti-apoptosis, and angiogenesis (34).
The immunomodulatory properties of DPSCs may also exert crucial functions during the neural repair mechanism (35–38). Several studies have shown not only CD4+T cells co-cultured with DPSCs can highly express regulatory T cells (Treg), but DPSCs implanted in vivo could reverse the decrease in Treg expression induced by transforming growth factor β1 (TGFβ1) and interleukin-10 signaling pathways inhibition, suggesting that DPSCs could interfere with immunoregulation during nerve regeneration (35, 36). On the other hand, it has been discovered that DPSCs may inhibit TNF-α, and thus up-regulate anti-inflammation cytokines and promote nerve regeneration (37). furthermore, DPSCs can induce inhibition in cytotoxic T cell proliferation and activation via CD73, a central enzyme in the crosstalk of immunosuppressive adenosine and extracellular pro-inflammatory ATP, which is highly expressed in DPSCs (38).
Peripheral nerve injuries lead to notable functional impairments and decreased quality of life (39). Despite advances in microscopic techniques for neurosurgery, clinicians always desire to improve postoperative nerve regeneration and rehabilitation for better functional restoration via various methods including biotherapy (40, 41). Currently, the transplanted DPSCs alone or in combination with several novel nerve conduits are promising therapy for patients suffering from peripheral nerve injury (42, 43). Takaoka et al. (44) transplanted DPSCs into a rat model with a 10-mm sciatic nerve defect and found improvement in axon growth, remyelination, electrophysiological activities, and alleviated muscle atrophy at 12 weeks post-transplantation. DPSC-embedded polymeric biomaterial based on ethyl acrylate and hydroxy ethyl acrylate copolymer shows sufficient bioactivity to promote regeneration of the injured sciatic nerve (45). A similar study has shown that preloading collagen conduits with Schwann cell-like cells (SCLCs) induced from DPSCs could enhance sciatic nerve repair (46). In addition, collagen scaffolds preloaded with DPSCs post-differentiation could exhibit certain traits of SCLC that promote the outgrowth of axons and myelination in 2-dimensional or 3-dimensional culture conditions (47). These results demonstrate that DPSCs are excellent stem cell sources for peripheral nerve regeneration (Figure 3).
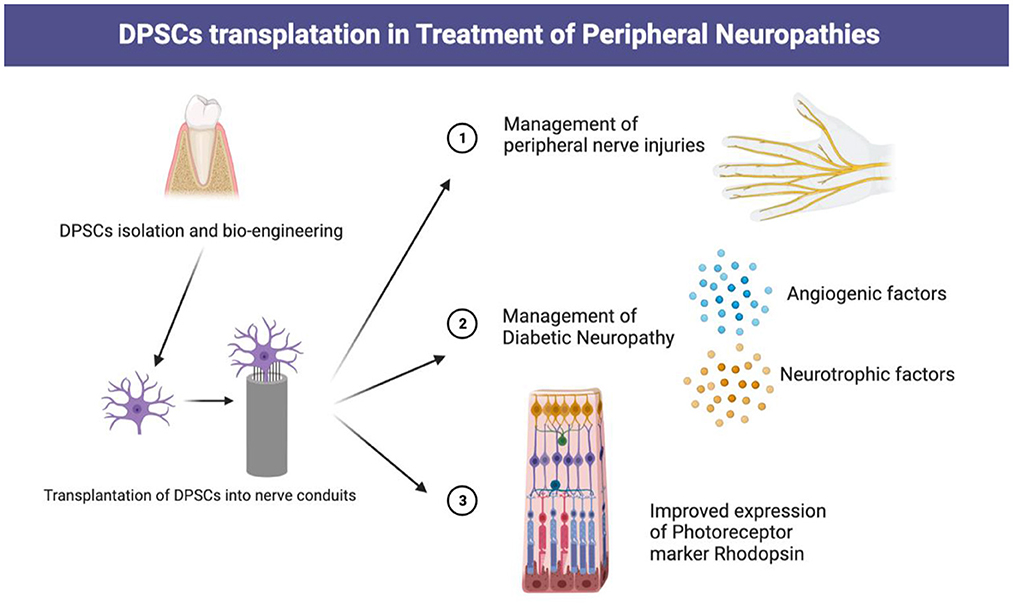
Figure 3. Transplantation of DPSCs in management of various peripheral neuropathies.
Diabetic neuropathy, the most common complication of type 1 and type 2 Diabetes Mellitus (DM), and has become a substantial health concern worldwide, especially for the elderly. It has been estimated that over 50% of long-term DM patients will eventually develop neuropathy, which could lead to diabetic foot ulcers associated with serious disabilities (48). The COVID milieu since 2019 has driven healthcare professionals to emphasize the matter of management and nursing of patients suffering from diabetic neuropathy since DM patients are prone to have compromised immunity and disturbed microenvironment (49). Typically, patients suffering from diabetic neuropathy show decreased peripheral nerve vascularity and a deficiency of angiogenic and neurotrophic factors, which may account for the pathogenesis of neuropathies (50, 51). Recently, the therapeutic effects of DPSCs in diabetic neuropathy have become recognized by many researchers, which raised controversies regarding the optimal application method of DPSCs. Makino et al. suggested that DPSCs transplantation can significantly improve the blood flow, nerve conduction velocity, capillary density, and intra-epidermal nerve fiber density of the damaged nerves while up-regulating the expression levels of angiogenic and neurotrophic factor genes (52). Another study proved that transplanted-DPSCs can significantly reduce the number of macrophages in the diabetic peripheral nerve microenvironment and specifically inhibit M1 macrophage expression while up-regulating M2 macrophage expression, eventually decreasing the M1/M2 macrophage ratio (19). Apart from regulating macrophage expression, DPSCs could also exert anti-inflammatory effects via inhibiting tumor necrosis factor α (TNFα) and interleukin-6 (IL-6) expression while up-regulating TGF-β expression. These findings provide some perspectives on possible future applications of DPSCs in diabetic neuropathy management.
Neurons in the retina and optic nerve share a mutual origin from the embryonic diencephalon. After neuroepithelium formation of the retina, neurons lose the ability to divide, rendering neuronal renewal in the retina impossible (53). Therefore, blindness caused by retinal injuries remains a major cause of disability worldwide. Retinal ganglion cells express a large number of neurotrophic factor receptors, which may enhance retinal ganglion cell survival and axonal regeneration (54). Mead et al. found that DPSCs secreted a large number of neurofibrillary tangles (NTF), which enhanced neural βIII-tubulin+ retinal cell proliferation and lengthened the neuritis (55). In addition, transplantation of DPSCs into the vitreous humor of mice after optic nerve injury promoted Brn-3a+ retinal ganglion cell survival and axonal regeneration (55). It has been reported that 44% of DPSCs expressed a photoreceptor marker rhodopsin in a conditioned medium from the damaged retina (56). This promising novel mechanism should be further explored for clinical applications.
Dental pulp tissue yields great reproductive ability and is rich in varying categories of stem cells with unique differentiation potentials. It has been concluded that DPSCs may be isolated from both postnatal teeth and extremely rare natal teeth (11). Immortalized DPSCs are also an excellent source of pluripotent stem cells with high molecular, morphological and genetic resemblance with non-immortalized DPSCs, which introduces the possibility of building a reservoir with immortalized DPSCs from patients suffering from a wide spectrum of neurogenetic disorders (57). On the other hand, Wilson et al. (58) have assessed the tumorigenic potential of immortalized DPSC in vitro and in mice and observed no tumor formation, indicating the probable safety of immortalized DPSC in future clinical applications.
As presented in Table 2, current research on the application of DPSC in the management of central and peripheral neuropathies is majorly concentrated on differentiation induced in vitro and implantation either via nerve conduit scaffold or direct injections. However, for such a therapeutic regimen to be carried out clinically, further clinical and lab research are required to achieve large-scale DPSC manufacture, storage, and transportation with minimum possibility of contamination. Concerns have been raised about the compromised quality of DPSC culture possibly in association with poor oral hygiene and long-distance transportation (57).
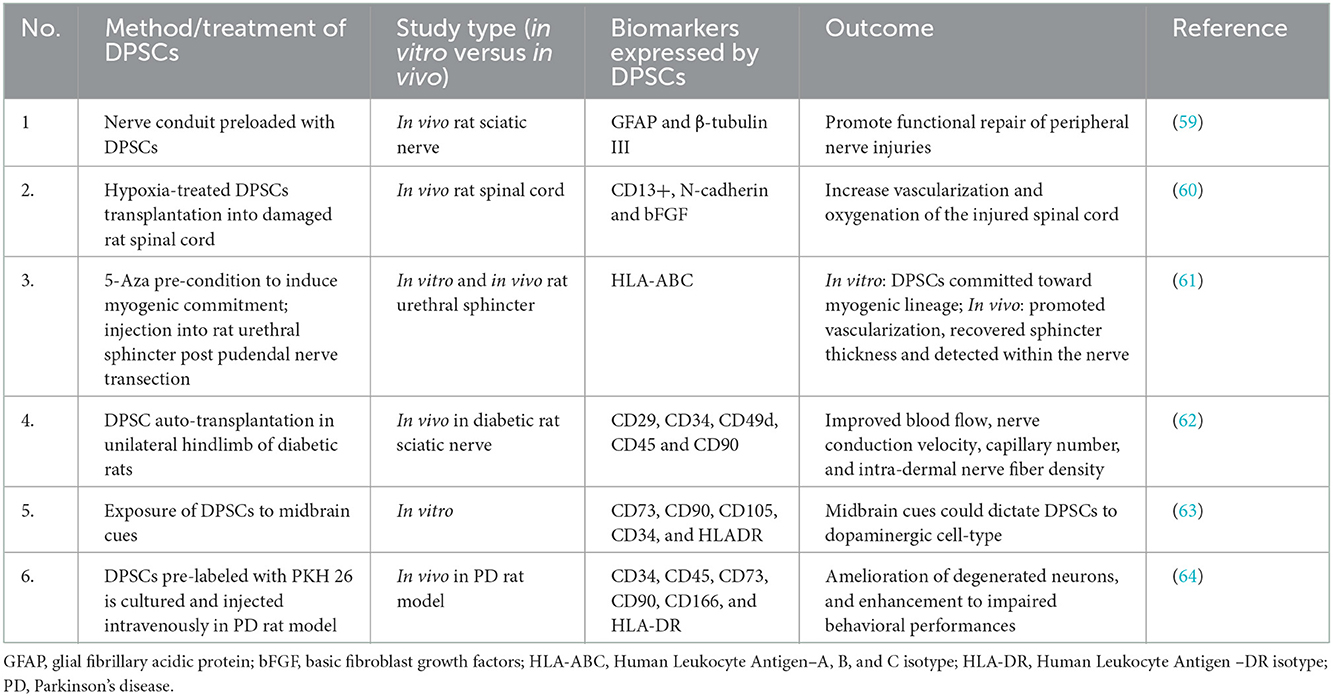
Table 2. Several current studies involving the application of DPSCs in nerve regeneration.
Stem cell-based therapies shed light on the biotherapies of peripheral nerve disease. DPSCs may enhance peripheral nerve regeneration via the induction of neuronal differentiation and the up-regulation of various neurotrophic factors. DPSCs in combination with biomaterials could be the prospect of neural tissue repair. Furthermore, DPSCs have a wide range of application prospects in peripheral nerve diseases, such as peripheral nerve injury, diabetic neuropathy, and retina injury. While DPSCs transplantation shows promising therapeutic potential in the management of peripheral nerve diseases, further research is required to establish a therapeutic approach and a regimen of dosage, efficacy, and safety. In conclusion, DPSCs yield great potential in peripheral neural tissue regeneration and repair, yet various issues remain to be solved through further assessment and experimentation.
HL and JF designed the study. FJ, XL, and YD performed data collection. HZ, AA, ZW, and JL analyzed the results. CY, SE, VK, BC, and MH drafted the manuscript. All authors have read and approved the final manuscript.
This study was funded by Zhejiang Provincial Natural Science Foundation Academic Exchange Program (Grant Numbers: LY20H060007 and LY20H090018). The funding bodies had no role in the design of the study; in the collection, analysis, and interpretation of data; in drafting the manuscript.
We thank Zhenyu Sun for comments on the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Donzelli R, Capone C, Sgulò FG, Mariniello G, Donzelli O, Maiuri F. Microsurgical repair by autografting in traumatic injuries of peripheral nerves. J Neurosurg Sci. (2022) 66:208–14. doi: 10.23736/S0390-5616.19.04572-7
2. Baumard J, Lesourd M, Remigereau C, Jarry C, Etcharry-Bouyx F, Chauviré V, et al. Tool use in neurodegenerative diseases: planning or technical reasoning? J Neuropsychol. (2018) 12:409–26. doi: 10.1111/jnp.12121
3. Farrell MC, Weiss SJ, Goodrich C, Lehmann MP, Delarato N. Food aversion leading to nutritional optic neuropathy in a child with severe vitamin a deficiency. J Neuroophthalmol. (2021) 41:e718–9. doi: 10.1097/WNO.0000000000001142
4. Klimovich P, Rubina K, Sysoeva V, Semina E. New Frontiers in Peripheral Nerve Regeneration: Concerns and Remedies. Int J Mol Sci. (2021) 22(24). doi: 10.3390/ijms222413380
5. Zainal Ariffin SH, Kermani S, Zainol Abidin IZ, Megat Abdul Wahab R, Yamamoto Z, et al. Differentiation of dental pulp stem cells into neuron-like cells in serum-free medium. Stem Cells Int. (2013) 2013:250740. doi: 10.1155/2013/250740
6. Luo L, He Y, Wang X, Key B, Lee BH, Li H, et al. Potential roles of dental pulp stem cells in neural regeneration and repair. Stem Cells Int. (2018) 2018:1731289. doi: 10.1155/2018/1731289
7. Young F, Sloan A, Song B. Dental pulp stem cells and their potential roles in central nervous system regeneration and repair. J Neurosci Res. (2013) 91:1383–93. doi: 10.1002/jnr.23250
8. de Freria CM, Van Niekerk E, Blesch A, Lu P. Neural stem cells: promoting axonal regeneration and spinal cord connectivity. Cells. (2021) 10:3296. doi: 10.3390/cells10123296
9. Karaöz E, Demircan PC, Saglam O, Aksoy A, Kaymaz F, Duruksu G. Human dental pulp stem cells demonstrate better neural and epithelial stem cell properties than bone marrow-derived mesenchymal stem cells. Histochem Cell Biol. (2011) 136:455–73. doi: 10.1007/s00418-011-0858-3
10. Martens W, Bronckaers A, Politis C, Jacobs R, Lambrichts I. Dental stem cells and their promising role in neural regeneration: an update. Clin Oral Investig. (2013) 17:1969–83. doi: 10.1007/s00784-013-1030-3
11. Gronthos S, Mankani M, Brahim J, Robey PG, Shi S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci U S A. (2000) 97:13625–30. doi: 10.1073/pnas.240309797
12. Gronthos S, Brahim J, Li W, Fisher LW, Cherman N, Boyde A, et al. Stem cell properties of human dental pulp stem cells. J Dent Res. (2002) 81:531–5. doi: 10.1177/154405910208100806
13. Kawashima N. Characterisation of dental pulp stem cells: a new horizon for tissue regeneration? Arch Oral Biol. (2012) 57:1439–58. doi: 10.1016/j.archoralbio.2012.08.010
14. Cheng PH, Snyder B, Fillos D, Ibegbu CC, Huang AHC, Chan AWS. Postnatal stem/progenitor cells derived from the dental pulp of adult chimpanzee. BMC Cell Biol. (2008) 9:20. doi: 10.1186/1471-2121-9-20
15. Király M, Porcsalmy B, Pataki A, Kádár K, Jelitai M, Molnár B, et al. Simultaneous PKC and cAMP activation induces differentiation of human dental pulp stem cells into functionally active neurons. Neurochem Int. (2009) 55:323–32. doi: 10.1016/j.neuint.2009.03.017
16. Feng X, Xing J, Feng G, Sang A, Shen B, Xu Y, et al. Age-dependent impaired neurogenic differentiation capacity of dental stem cell is associated with Wnt/beta-catenin signaling. Cell Mol Neurobiol. (2013) 33:1023–31. doi: 10.1007/s10571-013-9965-0
17. Alvarez R, Lee HL, Hong C, Wang CY. Single CD271 marker isolates mesenchymal stem cells from human dental pulp. Int J Oral Sci. (2015) 7:205–12. doi: 10.1038/ijos.2015.29
18. Lei T, Wang J, Liu Y, Chen P, Zhang Z, Zhang X, et al. Calreticulin as a special marker to distinguish dental pulp stem cells from gingival mesenchymal stem cells. Int J Biol Macromol. (2021) 178:229–39. doi: 10.1016/j.ijbiomac.2021.02.126
19. Omi M, Hata M, Nakamura N, Miyabe M, Ozawa S, Nukada H, et al. Transplantation of dental pulp stem cells improves long-term diabetic polyneuropathy together with improvement of nerve morphometrical evaluation. Stem Cell Res Ther. (2017) 8:279. doi: 10.1186/s13287-017-0729-5
20. Sugimura-Wakayama Y, Katagiri W, Osugi M, Kawai T, Ogata K, Sakaguchi K, et al. Peripheral nerve regeneration by secretomes of stem cells from human exfoliated deciduous teeth. Stem Cells Dev. (2015) 24:2687–99. doi: 10.1089/scd.2015.0104
21. Bhandi S, Alkahtani A, Reda R, Mashyakhy M, Boreak N, Maganur PC, et al. Parathyroid hormone secretion and receptor expression determine the age-related degree of osteogenic differentiation in dental pulp stem cells. J Pers Med. (2021) 11:349. doi: 10.3390/jpm11050349
22. Sharma. Changes in neurotrophic factors of adult rat laryngeal muscles during nerve regeneration. Neuroscience. (2016) 333:44–53. doi: 10.1016/j.neuroscience.2016.07.004
23. Zheng K, Feng G, Zhang J, Xing J, Huang D, Lian M, et al. Basic fibroblast growth factor promotes human dental pulp stem cells cultured in 3D porous chitosan scaffolds to neural differentiation. Int J Neurosci. (2021) 131:625–33. doi: 10.1080/00207454.2020.1744592
24. Tsutsui TW. Dental pulp stem cells: advances to applications. Stem Cells Cloning. (2020) 13:33–42. doi: 10.2147/SCCAA.S166759
25. Li Z, Li K, Zhu L, Kan Q, Yan Y, Kumar P, et al. Inhibitory effect of IL-17 on neural stem cell proliferation and neural cell differentiation. BMC Immunol. (2013) 14:20. doi: 10.1186/1471-2172-14-20
26. Kumar A, Kumar V, Rattan V, Jha V, Bhattacharyya S. Secretome cues modulate the neurogenic potential of bone marrow and dental stem cells. Mol Neurobiol. (2017) 54:4672–82. doi: 10.1007/s12035-016-0011-3
27. Chen ZL, Yu WM, Strickland S. Peripheral regeneration. Annu Rev Neurosci. (2007) 30:209–33. doi: 10.1146/annurev.neuro.30.051606.094337
28. Heng BC, Gong T, Xu J, Lim LW, Zhang C. EphrinB2 signalling modulates the neural differentiation of human dental pulp stem cells. Biomed Rep. (2018) 9:161–8. doi: 10.3892/br.2018.1108
29. Urraca N, Hope K, Victor AK, Belgard TG, Memon R, Goorha S, et al. Significant transcriptional changes in 15q duplication but not Angelman syndrome deletion stem cell-derived neurons. Mol Autism. (2018) 9:6. doi: 10.1186/s13229-018-0191-y
30. Kolar MK, Itte VN, Kingham PJ, Novikov LN, Wiberg M, Kelk P. The neurotrophic effects of different human dental mesenchymal stem cells. Sci Rep. (2017) 7:12605. doi: 10.1038/s41598-017-12969-1
31. Yao R Cooper GM. Requirement for phosphatidylinositol-3 kinase in the prevention of apoptosis by nerve growth factor. Science. (1995) 267:2003–6. doi: 10.1126/science.7701324
32. Traverse S, Gomez N, Paterson H, Marshall C, Cohen P. Sustained activation of the mitogen-activated protein (MAP) kinase cascade may be required for differentiation of PC12 cells. Comparison of the effects of nerve growth factor and epidermal growth factor. Biochem J. (1992) 288(Pt 2):351–5. doi: 10.1042/bj2880351
33. Sun ZG, Ma JT, Liu HW, Hu M, Huang HT. ERK/MAPK and PI3K/AKT signal channels simultaneously activated in nerve cell and axon after facial nerve injury. Saudi J Biol Sci. (2017) 24:1853–8. doi: 10.1016/j.sjbs.2017.11.027
34. Kichenbrand C, Velot E, Menu P, Moby V. Dental pulp stem cell-derived conditioned medium: an attractive alternative for regenerative therapy. Tissue Eng Part B Rev. (2019) 25:78–88. doi: 10.1089/ten.teb.2018.0168
35. Raza SS, Wagner AP, Hussain YS, Khan MA. Mechanisms underlying dental-derived stem cell-mediated neurorestoration in neurodegenerative disorders. Stem Cell Res Ther. (2018) 9:245. doi: 10.1186/s13287-018-1005-z
36. Hong JW, Lim JH, Chung CJ, Kang TJ, Kim TY, Kim YS, et al. Immune tolerance of human dental pulp-derived mesenchymal stem cells mediated by CD4(+)CD25(+)FoxP3(+) regulatory T-cells and induced by TGF-beta1 and IL-10. Yonsei Med J. (2017) 58:1031–9. doi: 10.3349/ymj.2017.58.5.1031
37. Yang C, Li X, Sun L, Guo W, Tian W. Potential of human dental stem cells in repairing the complete transection of rat spinal cord. J Neural Eng. (2017) 14:026005. doi: 10.1088/1741-2552/aa596b
38. Ahmadi P, Yan M, Bauche A, Smeets R, Müller CE, Koch-Nolte F, et al. Human dental pulp cells modulate CD8(+) T cell proliferation and efficiently degrade extracellular ATP to adenosine in vitro. Cell Immunol. (2022) 380:104589. doi: 10.1016/j.cellimm.2022.104589
39. Lopes B, Sousa P, Alvites R, Branquinho M, Sousa AC, Mendonça C, et al. Peripheral nerve injury treatments and advances: one health perspective. Int J Mol Sci. (2022) 23:918. doi: 10.3390/ijms23020918
40. Zhou HY, Jiang S, Ma FX, Lu H. Peripheral nerve tumors of the hand: clinical features diagnosis and treatment. World J Clin Cases. (2020) 8:5086–98. doi: 10.12998/wjcc.v8.i21.5086
41. Lu H, Chen L, Jiang S, Shen H. A rapidly progressive foot drop caused by the posttraumatic Intraneural ganglion cyst of the deep peroneal nerve. BMC Musculoskelet Disord. (2018) 19:298. doi: 10.1186/s12891-018-2229-x
42. Pfister BJ, Gordon T, Loverde JR, Kochar AS, Mackinnon SE, Cullen DK. Biomedical engineering strategies for peripheral nerve repair: surgical applications state of the art and future challenges. Crit Rev Biomed Eng. (2011) 39:81–124. doi: 10.1615/CritRevBiomedEng.v39.i2.20
43. Luzuriaga J, Polo Y, Pastor-Alonso O, Pardo-Rodríguez B, Larrañaga A, Unda F, et al. Advances and perspectives in dental pulp stem cell based neuroregeneration therapies. Int J Mol Sci. (2021) 22:3546. doi: 10.3390/ijms22073546
44. Takaoka S, Uchida F, Ishikawa H, Toyomura J, Ohyama A, Watanabe M, et al. Transplanted neural lineage cells derived from dental pulp stem cells promote peripheral nerve regeneration. Hum Cell. (2022) 35:462–71. doi: 10.1007/s13577-021-00634-9
45. Soria JM, Sancho-Tello M, Esparza MA, Mirabet V, Bagan JV, Monleón M, et al. Biomaterials coated by dental pulp cells as substrate for neural stem cell differentiation. J Biomed Mater Res A. (2011) 97:85–92. doi: 10.1002/jbm.a.33032
46. Sanen K, Martens W, Georgiou M, Ameloot M, Lambrichts I, Phillips J. Engineered neural tissue with Schwann cell differentiated human dental pulp stem cells: potential for peripheral nerve repair? J Tissue Eng Regen Med. (2017) 11:3362–72. doi: 10.1002/term.2249
47. Martens W, Sanen K, Georgiou M, Struys T, Bronckaers A, Ameloot M, et al. Human dental pulp stem cells can differentiate into Schwann cells and promote and guide neurite outgrowth in an aligned tissue-engineered collagen construct in vitro. FASEB J. (2014) 28:1634–43. doi: 10.1096/fj.13-243980
48. Reardon R, Simring D, Kim B, Mortensen J, Williams D, Leslie A. The diabetic foot ulcer. Aust J Gen Pract. (2020) 49:250–5. doi: 10.31128/AJGP-11-19-5161
49. Zhou H, Jin Q, Lu H. Exposure risk of patients with chronic infectious wounds during the COVID-19 outbreak and its countermeasures. J Orthop Surg Res. (2020) 15:452. doi: 10.1186/s13018-020-01976-0
50. Feldman EL, Nave KA, Jensen TS, Bennett DLH. New horizons in diabetic neuropathy: mechanisms, bioenergetics, and pain. Neuron. (2017) 93:1296–313. doi: 10.1016/j.neuron.2017.02.005
51. Zhang. Mesenchymal stem cells to treat diabetic neuropathy: a long and strenuous way from bench to the clinic. Cell Death Discov. (2016) 2:16055. doi: 10.1038/cddiscovery.2016.55
52. Makino E, Nakamura N, Miyabe M, Ito M, Kanada S, Hata M, et al. Conditioned media from dental pulp stem cells improved diabetic polyneuropathy through anti-inflammatory neuroprotective and angiogenic actions: Cell-free regenerative medicine for diabetic polyneuropathy. J Diabetes Investig. (2019) 10:1199–208. doi: 10.1111/jdi.13045
53. Stafford. Regenerating optic pathways from the eye to the brain. Science. (2017) 356:1031–4. doi: 10.1126/science.aal5060
54. Berry M, Ahmed Z, Lorber B, Douglas M, Logan A. Regeneration of axons in the visual system. Restor Neurol Neurosci. (2008) 26:147–74. doi: 10.2174/1570159X14666161227150250
55. Mead B, Logan A, Berry M, Leadbeater W, Scheven BA. Intravitreally transplanted dental pulp stem cells promote neuroprotection and axon regeneration of retinal ganglion cells after optic nerve injury. Invest Ophthalmol Vis Sci. (2013) 54:7544–56. doi: 10.1167/iovs.13-13045
56. Bray AF, Cevallos RR, Gazarian K, Lamas M. Human dental pulp stem cells respond to cues from the rat retina and differentiate to express the retinal neuronal marker rhodopsin. Neuroscience. (2014) 280:142–55. doi: 10.1016/j.neuroscience.2014.09.023
57. Urraca N, Memon R, El-Iyachi I, Goorha S, Valdez C, Tran QT, et al. Characterization of neurons from immortalized dental pulp stem cells for the study of neurogenetic disorders. Stem Cell Res. (2015) 15:722–30. doi: 10.1016/j.scr.2015.11.004
58. Wilson R, Urraca N, Skobowiat C, Hope KA, Miravalle L, Chamberlin R, et al. Assessment of the tumorigenic potential of spontaneously immortalized and hTERT-immortalized cultured dental pulp stem cells. Stem Cells Transl Med. (2015) 4:905–12. doi: 10.5966/sctm.2014-0196
59. Luo L, He Y, Jin L, Zhang Y, Guastaldi FP, Albashari AA, et al. Application of bioactive hydrogels combined with dental pulp stem cells for the repair of large gap peripheral nerve injuries. Bioact Mater. (2021) 6:638–54. doi: 10.1016/j.bioactmat.2020.08.028
60. Zhu S, Ying Y, He Y, Zhong X, Ye J, Huang Z, et al. Hypoxia response element-directed expression of bFGF in dental pulp stem cells improve the hypoxic environment by targeting pericytes in SCI rats. Bioact Mater. (2021) 6:2452–66. doi: 10.1016/j.bioactmat.2021.01.024
61. Zordani A, Pisciotta A, Bertoni L, Bertani G, Vallarola A, Giuliani D, et al. Regenerative potential of human dental pulp stem cells in the treatment of stress urinary incontinence: in vitro and in vivo study. Cell Prolif . (2019) 52(6):e12675. doi: 10.1111/cpr.12675
62. Hata M, Omi M, Kobayashi Y, Nakamura N, Tosaki T, Miyabe M, et al. Transplantation of cultured dental pulp stem cells into the skeletal muscles ameliorated diabetic polyneuropathy: therapeutic plausibility of freshly isolated and cryopreserved dental pulp stem cells. Stem Cell Res Ther. (2015) 6:162. doi: 10.1186/s13287-015-0156-4
63. Kanafi M, Majumdar D, Bhonde R, Gupta P, Datta I. Midbrain cues dictate differentiation of human dental pulp stem cells towards functional dopaminergic neurons. J Cell Physiol. (2014) 229:1369–77. doi: 10.1002/jcp.24570
Keywords: dental pulp stem cells, neuron, Schwann cells, peripheral nerve diseases, neurotrophic factors
Citation: Fu J, Li X, Jin F, Dong Y, Zhou H, Alhaskawi A, Wang Z, Lai J, Yao C, Ezzi SHA, Kota VG, Hasan Abdulla Hasan Abdulla M, Chen B and Lu H (2023) The potential roles of dental pulp stem cells in peripheral nerve regeneration. Front. Neurol. 13:1098857. doi: 10.3389/fneur.2022.1098857
Received: 15 November 2022; Accepted: 28 December 2022;
Published: 11 January 2023.
Edited by:
Bruce Douglas Carter, Vanderbilt University, United StatesReviewed by:
Nareshwaran Gnanasegaran, Singapore-MIT Alliance for Research and Technology (SMART), SingaporeCopyright © 2023 Fu, Li, Jin, Dong, Zhou, Alhaskawi, Wang, Lai, Yao, Ezzi, Kota, Hasan Abdulla Hasan Abdulla, Chen and Lu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hui Lu,  aHVpbHVAemp1LmVkdS5jbg==
aHVpbHVAemp1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.