
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Neurol. , 02 October 2019
Sec. Headache and Neurogenic Pain
Volume 10 - 2019 | https://doi.org/10.3389/fneur.2019.00982
This article is part of the Research Topic Functional and Structural Brain Alterations in Headache: A Trait or a State? View all 14 articles
 Zsigmond Tamás Kincses1,2*†
Zsigmond Tamás Kincses1,2*† Dániel Veréb1†
Dániel Veréb1† Péter Faragó1
Péter Faragó1 Eszter Tóth1
Eszter Tóth1 Krisztián Kocsis1
Krisztián Kocsis1 Bálint Kincses1
Bálint Kincses1 András Király1,3
András Király1,3 Bence Bozsik1Árpád Párdutz1Délia Szok1
Bence Bozsik1Árpád Párdutz1Délia Szok1 János Tajti1
János Tajti1 László Vécsei1,4
László Vécsei1,4 Bernadett Tuka1,4
Bernadett Tuka1,4 Nikoletta Szabó1,3
Nikoletta Szabó1,3Background: Migraine research is booming with the rapidly developing neuroimaging tools. Structural and functional alterations of the migrainous brain were detected with MRI. The outcome of a research study largely depends on the working hypothesis, on the chosen measurement approach and also on the subject selection. Against all evidence from the literature that migraine subtypes are different, most of the studies handle migraine with and without aura as one disease.
Methods: Publications from PubMed database were searched for terms of “migraine with aura,” “migraine without aura,” “interictal,” “MRI,” “diffusion weighted MRI,” “functional MRI,” “compared to,” “atrophy” alone and in combination.
Conclusion: Only a few imaging studies compared the two subforms of the disease, migraine with aura, and without aura, directly. Functional imaging investigations largely agree that there is an increased activity/activation of the brain in migraine with aura as compared to migraine without aura. We propose that this might be the signature of cortical hyperexcitability. However, structural investigations are not equivocal. We propose that variable contribution of parallel, competing mechanisms of maladaptive plasticity and neurodegeneration might be the reason behind the variable results.
Migraine is a heterogeneous disease affecting cca 10–20% of the population worldwide. It is associated with significant disability, reduced quality of life, and consequently poses an enormous financial burden to society (1). Since migraine ranks among the top disorders causing disability (2). It is in the focus of neuroimaging, molecular, and pharmaceutical research. In 20% of the cases, migraine headache is preceded or accompanied by reversible focal neurological symptoms, such as visual, motor, sensory, or speech disturbances (3). The ICHD-3 classification (4) bases the diagnosis of migraine on the patient's medical history and physical examination. Accordingly, migraine can be categorized into migraine with aura (MWA) and without aura (MWoA) as subtypes of the disease (among other categories). Besides the similarities in the epidemiology, clinical presentation, and the genetic evidence that MWA and MWoA largely overlap (5), the question has been raised a few years ago: are MWA and MWoA separate entities (6–8) or rather the two ends of a spectrum? Nevertheless, studying mixed groups of migraine patients should be avoided in further investigations if critical differences exist. Nevertheless, studying mixed groups of migraine patients should be avoided in further investigations if critical differences exist.
Since magnetic resonance imaging (MRI) makes it possible to investigate the structure and function of the brain in vivo, hundreds of papers were published in the last 20 years that describe the migrainous brain using neuroimaging methodology. The majority of these studies examined mixed patient groups or compared only one subtype to healthy individuals.
This review article summarizes the most significant neuroimaging results from studies comparing MWA patients to MWoA. To identify relevant articles, we searched the PubMed database for terms of “migraine with aura,” “migraine without aura,” “interictal,” “MRI,” “diffusion weighted MRI” (DWI), “functional MRI” (fMRI), “compared to,” “atrophy” alone and in combination up to June 2019.
Brain activity during rest and task performance can be described non-invasively by measuring the blood oxygen level dependent (BOLD) signal with functional MRI. Traditional fMRI studies compare signal differences in various phases of a task, but recently, there has also been a growing interest in studying brain activity patterns during rest. Interestingly, remote areas show synchronous activity, which renders resting state activity into functional networks (9, 10). Although fMRI parameters remain basically the same, we are witnessing a rapid development in the statistical analysis of fMRI scans.
Two publications confirmed brain activation differences between MWA and MWoA in the interictal phase (11, 12). Datta et al. (11) described higher BOLD response in migraine in response to visual stimuli in a BOLD fMRI study. This higher BOLD response was more robust in MWA than in MWoA patients. Interestingly the resting perfusion parameters of the two groups was not different, hence the authors discussed their finding in the light of the existing evidences that it relates to hyperresponsiveness of the visual cortex in MWA. On the contrary, resting brain perfusion did not differ between patients and controls or between MWA and MWoA patients (11). In a considerably larger cohort Cucchiara et al. found similarly greater BOLD amplitude in the visual cortex in MWA that positively correlated with visual discomfort score. No such correlation was found in MWoA (12).
Limited data are available on resting brain activity. Increased expression of the visual resting functional network was found in MWA compared to MWoA and controls (13). In our earlier investigation we found higher amplitude of resting state activity fluctuation in all identified resting state networks in the 0.08–0.04 Hz frequency range in MWA as compared to MWoA (14). On the contrary, lower amplitudes were found in the default mode network in MWoA compared to controls.
Reduced connectivity between the occipital lobe and anterior insula was found in MWA but not in MWoA, and the connectivity strength correlated with migraine severity in MWA (15). Increased connectivity was found in the default mode network in the pre-central gyrus, post-central gyrus, insular cortex, angular gyrus, supramarginal gyrus in MWA compared to MWoA (16).
Several investigations have shown that there are gray matter alterations in migraine: the gray matter density of several pain related cortical regions is reduced compared to healthy individuals (17). It should be noted that similar brain structural alterations were found in other chronic pain conditions. Importantly, only a few investigations concentrated on comparing the two subgroups of migraine.
Granziera et al. found increased cortical thickness and altered microstructure in migraineurs in the white matter beneath motion processing areas, namely motion processing visual areas and V3A area, but there were no differences between MWA and MWoA patients (18). In a similar cohort, voxel-based morphometry (VBM) did not detect any differences between the two patient groups (13). In a multicentre study involving a considerably larger migraine population, MWoA patients exhibited thinner cortex in the left central sulcus, in the left occipito-temporal gyrus, in the right cuneus and the superior parietal gyrus bilaterally. In some of these regions, the cortical thickness correlated with the frequency of migraine attacks and disease duration (19). Interestingly, a few of these regions were not only thinner in MWoA as compared to controls, but also when compared to MWA.
Among the structural abnormalities, white matter microstructure changes, as described by DWI, are receiving more and more attention. DWI is sensitive to the diffusion of water molecules, which in the brain is largely restricted by the membranes of cellular and sub-cellular elements. By fitting a diffusion tensor model it is possible to estimate diffusion parameters that reflect the microscopic organization of the measured volume (20).
White matter microstructural changes in MWA were reported, but studies are not congruent in calculated diffusion parameters and results. DaSilva et al. presented lower fractional anisotropy (FA) in the ventral trigemino-thalamic pathway in MWA and lower FA was detected in the ventrolateral periaqueductal gray matter (PAG) in MWoA (21). No correlation was found with clinical parameters. While migraineurs showed reduced FA subjacent to visual motion processing areas, no differences in diffusion parameters were found between MWA and MWoA (18). Similarly, tract-based-spatial statistics (TBSS) and a pre-defined region-of-interest analysis from fMRI results did not reveal microstructural white matter alterations between the two subtypes (13, 22). On the other hand, we found extensive white matter regions showing higher FA in MWA in a whole brain TBSS analysis (8). Also, we found that clinical parameters, such as disease duration and estimated lifetime attack number were associated with lower axial diffusivity (AD) in the left superior longitudinal fascicle, the left corticospinal tract and with the right superior longitudinal fascicle of MWA patients (8).
There are only few neurological disorders that were investigated so extensively and the hypotheses for its pathophysiology went through such evolution like migraine. In spite of this enormous body of research, the “migraine puzzle” is still incomplete. While migraine was thought to be a functional disease of the brain, recent studies have shown that brain structure and microstructure also exhibit profound alterations. Independent MRI studies observed functional and structural differences between MWA and MWoA in the interictal period. In summary, it can be pointed out that studies concur in finding higher brain activity/activation in MWA, but structural differences between the two subtypes of the disease are not so well-established, and results are ambiguous in the literature (Table 1).
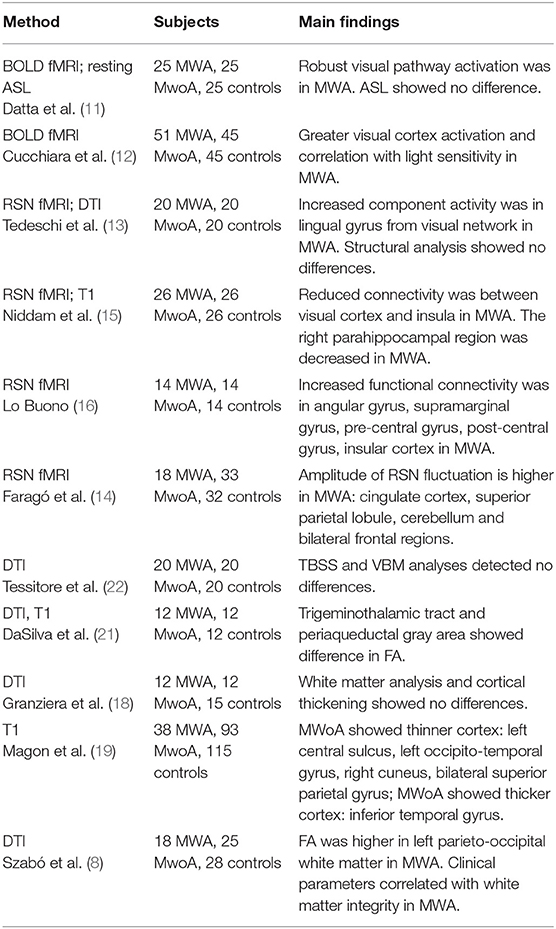
Table 1. Structural and functional MRI studies comparing migraine without to migraine with aura.
In MWA, during the transient visual, sensory or language symptoms a slow depolarization wave called cortical spreading depression (CSD) spreads through the cortex (23). During visual aura, BOLD signal changes develop in the occipital cortex and progress slowly, reflecting underlying depolarization waves (24). Apart from being the putative cause of the aura symptoms, CSD has been associated with neuroinflammation, possibly contributing further to the headache by activating the meningeal nociceptors and the neurons in the spinal trigeminal nucleus and trigeminal nucleus caudalis (25, 26). Neurophysiological investigations showed that the two subtypes of the disease differ considerably. The amplitudes of visual evoked potentials (VEP) were higher in migraineurs (27–30). Recent reports showed that hyperexcitability, as measured via VEP is predominantly true for MWA (31, 32). The threshold of transcranial magnetic stimulation (TMS) evoked phosphenes is also lower in migraineurs and the prevalence of phosphenes is higher (33). Interestingly, a recent metaanalysis pointed out that, similarly to the VEP results, this kind of TMS measured hyperexcitability is only true for patients experiencing aura (34). Moreover, the perception of cross-modal interaction that depends highly on cortical hyperexcitability differs between healthy volunteers and migraineurs (35). The effect is more pronounced in MWA. A possible background mechanism behind this hyperexcitability might be the altered neurochemical milieu, the imbalance of the excitatory and inhibitory neurotransmitter levels in migraineurs as detected by MR spectroscopy or other neurochemical approaches [e.g., glutamate and GABA, see (36, 37) for a review]. However, no study investigated the differences between MWA and MWoA. The above-mentioned functional imaging studies also demonstrated higher activation/activity in MWA. Considering that hyperexcitability comes along with increased firing frequency (38) that has a higher energy demand, it easily follows that BOLD fMRI studies find increased amplitude of response or resting activity fluctuation. In consequence to this increased activity, especially if it is regionally specific, interregional connections might strengthen, that could be measured as increased functional connectivity.
Nevertheless, one should not forget that fMRI is measuring the indirect vascular response to neuronal activity/activation. Since migraine is a neurovascular disease the identified differences in any fMRI study might be due to the filtering effect of the altered hemodynamic response function. In fact, altered vasomotor reactivity was identified in MWA (39).
The results of structural investigations are far less equivocal about the differences between the two subtypes of the disease.
A prominent reason behind the differences in the outcome of the structural studies might be the pathomechanism itself. Structural alterations could either be a (1) consequence or the (2) cause of the disease. In case of the former, one might consider two alternatives:
(a) The recurring painful attacks and hyperexcitability could lead to maladaptive plasticity. Use-dependent plasticity induced morphological changes are well-known in the gray and white matter (40–42). Repeated pain stimuli can also increase gray matter density in pain processing regions including the cingulate and the contralateral somatosensory cortex (43). One might hypothesize that the increased firing frequency due to hyperexcitability may induce similar use-dependent plastic changes. And finally, plastic changes were reported in animals after induction of CSD (44, 45). These processes presumably appear in the form of increased FA and thickened cortex (40–42).
(b) The underlying pathology might also cause degenerative processes in migraine. CSD might well-contribute to the noxious process (46), as it induces neuroinflammation and cellular damage (47–49). The recurring painful attacks and the cortical hyperexcitability might lead to excessive glutamate release (50), which is also known to induce excitotoxicity and cell death (51). CSD causes upregulation of matrix-metalloproteases (MMP) (52) and increased MMP activity was described in human migraineurs (53). This can lead to the leakage of the blood-brain barrier and inflammatory response and neuronal damage (54). In line with these hypotheses, increased ictal levels of S100B (a marker of glial damage) and neuron specific enolase (a marker of neuronal damage) were detected in migraineurs (55). These degenerative processes could presumably appear in the form of white matter disintegration (reduction of FA) and cortical atrophy.
We propose that these parallel, competing mechanisms coexist, but their relative contribution is different in MWA and MWoA. However, one might see the two sub-forms of the disease a spectrum, rather than two distinct entities and hence homogenous patient groups cannot be reproducibly formed.
An alternative explanation for the structural alterations might well-be that they are not consequential but rather causal factors of the disease. Accordingly, the genetic background is different between the two subtypes. Pisanu et al. demonstrated that genetic risk factors calculated on migraine-associated single nucleotide polymorphism differ between subgroups suggesting MWA and MWoA have different genetic backgrounds that contribute to the pathogenesis (56). Even so, we cannot exclude the possibility that co-morbidities and epigenetics have an influence on migraine pathogenesis (57, 58).
A number of findings showed that the clinical expression of migraine is consistent with perivascular trigeminal activation and release of neuropeptides [calcitonin gene-related peptide (CGRP), substance P and pituitary adenylate cyclase-activating polypeptide-38 (PACAP-38)] (59–66). It was also shown that CSD is tightly connected to CGRP release (67). Several aspects of the CGRP-related trigemino-vascular functions are also abnormal in FHM1-mutant mice showing an overall hyperexcitability phenotype (68). We showed that interictal PACAP-38 concentrations were lower in migraineurs, which approached normal levels during headache (69) and this altered interictal PACAP-38 serum level correlated with the microstructural integrity of pain related brain structures (70). Whether neuropeptide concentrations are different in MWA and MWoA is still to be investigated.
Whether CSD is the initiator of all the events of migraine attack (activation of distinct brain stem nuclei, neuropeptide release at the periphery, activation, and sensitization at the level of trigeminal nociceptors) remains controversial: although that there is evidence that CSD can induce activation trigeminal nociception in animals (71), but migrainous aura can occur without headache and the pain can start during the aura onset, moreover most of the migraineurs do not experience aura phenomenon at all, which suggests that CSD alone is insufficient and non-essential for the attack. If the latter is true CSD is not the cause, but the consequence or a part of the disease (72).
Importantly, several other reasons could be pointed out behind the variable results of structural studies. For example, the headache frequency is different in MWA and MWoA (73), which means the studies should be strictly matched for clinical parameters. The time since the last and until the next headache should also be strictly monitored. Unfortunately, none of the above mentioned studies are controlled for these factors.
Importantly, we have not considered white matter hyperintensities in our review in details, but it has to be pointed out that the prevalence of these lesions are also different in the two subtypes of the diseases (74, 75). The etiology of these lesions is not entirely clear yet, but thought to be microinfarction with a numerous factors contributing to it. Neurogenic inflammation, endothelial changes, thrombocyte aggregation may play a role, and the induced oligaemia might be deepened by the CSD (76).
Among primary headache disorders, migraine is a heterogeneous disease with two major subtypes. Functional imaging studies repeatedly confirmed various metrics of hyperexcitability. The results of structural imaging studies are far from being equivocal. We propose that variable contribution of parallel, competing mechanisms of maladaptive plasticity and neurodegeneration might be the reason behind the variable results. Therefore, in further research projects MWA and MWoA should be handled separately and groups should be strictly matched for clinical parameters if the two subtypes are directly compared.
ZK, DV, LV, and NS formulated the review hypothesis. NS, ZK, KK, ET, BB, BK, PF, DS, ÁP, BT, AK, JT, and DV went through on literature, collected the articles. NS, DV, and ZK wrote the manuscript.
This study was supported by Neuroscience Research Group of the Hungarian Academy of Sciences and the University of Szeged, GINOP-2.3.2-15-2016-00034 grant, EFOP-3.6.1-16-2016-00008, NAP 2.0 (2017-1.2.1-NKP-2017-00002), National Brain Research Program (KTIA_13_NAP-A-II/20), UNKP-17-3 New National Excellence Program of the Ministry of Human Capacities, also University of Szeged, Ministry of Human Capacities, Hungary grant (20391-3/2018/FEKUSSTRAT). BK was supported by the UNKP-18-3 New National Excellence Program of the Ministry of Human Capacities. Support was provided by the project CEITEC 2020 (LQ1601) from the Ministry of Education, Youth and Sports of the Czech Republic and from the National Center for Advancing Translational Sciences of the National Institutes of Health and European Union's Horizon 2020 research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Lipton RB, Bigal ME. Migraine: epidemiology, impact, and risk factors for progression. Headache. (2005) 45(Suppl. 1):S3–13. doi: 10.1111/j.1526-4610.2005.4501001.x
2. Disease GBD, Injury I, Prevalence C. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet. (2016) 388:1545–602. doi: 10.1016/S0140-6736(16)31678-6
3. Lauritzen M. Pathophysiology of the migraine aura. The spreading depression theory. Brain. (1994) 117:199–210. doi: 10.1093/brain/117.1.199
4. Headache classification committee of the International Headache Society (IHS) the International Classification of Headache Disorders 3rd edition. Cephalalgia. (2018) 38:1–211. doi: 10.1177/0333102417738202
5. Zhao H, Eising E, de Vries B, Vijfhuizen LS, International Headache Genetics C, Anttila V, et al. Gene-based pleiotropy across migraine with aura and migraine without aura patient groups. Cephalalgia. (2016) 36:648–57. doi: 10.1177/0333102415591497
6. Ranson R, Igarashi H, MacGregor EA, Wilkinson M. The similarities and differences of migraine with aura and migraine without aura: a preliminary study. Cephalalgia. (1991) 11:189–92. doi: 10.1046/j.1468-2982.1991.1104189.x
7. Manzoni GC, Torelli P. Migraine with and without aura: a single entity? Neurol Sci. (2008) 29(Suppl 1):S40–3. doi: 10.1007/s10072-008-0884-7
8. Szabó N, Farago P, Kiraly A, Vereb D, Csete G, Toth E, et al. Evidence for plastic processes in migraine with aura: a diffusion weighted MRI study. Front Neuroanat. (2017) 11:138. doi: 10.3389/fnana.2017.00138
9. Beckmann CF, DeLuca M, Devlin JT, Smith SM. Investigations into resting-state connectivity using independent component analysis. Philos Trans R Soc Lond B Biol Sci. (2005) 360:1001–13. doi: 10.1098/rstb.2005.1634
10. Mantini D, Perrucci MG, Del Gratta C, Romani GL, Corbetta M. Electrophysiological signatures of resting state networks in the human brain. Proc Natl Acad Sci USA. (2007) 104:13170–5. doi: 10.1073/pnas.0700668104
11. Datta R, Aguirre GK, Hu S, Detre JA, Cucchiara B. Interictal cortical hyperresponsiveness in migraine is directly related to the presence of aura. Cephalalgia. (2013) 33:365–74. doi: 10.1177/0333102412474503
12. Cucchiara B, Datta R, Aguirre GK, Idoko KE, Detre J. Measurement of visual sensitivity in migraine: validation of two scales and correlation with visual cortex activation. Cephalalgia. (2015) 35:585–92. doi: 10.1177/0333102414547782
13. Tedeschi G, Russo A, Conte F, Corbo D, Caiazzo G, Giordano A, et al. Increased interictal visual network connectivity in patients with migraine with aura. Cephalalgia. (2016) 36:139–47. doi: 10.1177/0333102415584360
14. Farago P, Tuka B, Toth E, Szabo N, Kiraly A, Csete G, et al. Interictal brain activity differs in migraine with and without aura: resting state fMRI study. J Headache Pain. (2017) 18:8. doi: 10.1186/s10194-016-0716-8
15. Niddam DM, Lai KL, Fuh JL, Chuang CY, Chen WT, Wang SJ. Reduced functional connectivity between salience and visual networks in migraine with aura. Cephalalgia. (2016) 36:53–66. doi: 10.1177/0333102415583144
16. Lo Buono V, Bonanno L, Corallo F, Pisani LR, Lo Presti R, Grugno R, et al. Functional connectivity and cognitive impairment in migraine with and without aura. J Headache Pain. (2017) 18:72. doi: 10.1186/s10194-017-0782-6
17. May A. Pearls and pitfalls: neuroimaging in headache. Cephalalgia. (2013) 33:554–65. doi: 10.1177/0333102412467513
18. Granziera C, DaSilva AF, Snyder J, Tuch DS, Hadjikhani N. Anatomical alterations of the visual motion processing network in migraine with and without aura. PLoS Med. (2006) 3:e402. doi: 10.1371/journal.pmed.0030402
19. Magon S, May A, Stankewitz A, Goadsby PJ, Schankin C, Ashina M, et al. Cortical abnormalities in episodic migraine: a multi-center 3T MRI study. Cephalalgia. (2019) 39:665–73. doi: 10.1177/0333102418795163
20. Beaulieu C. The basis of anisotropic water diffusion in the nervous system–a technical review. NMR Biomed. (2002) 15:435–55. doi: 10.1002/nbm.782
21. DaSilva AF, Granziera C, Tuch DS, Snyder J, Vincent M, Hadjikhani N. Interictal alterations of the trigeminal somatosensory pathway and periaqueductal gray matter in migraine. Neuroreport. (2007) 18:301–5. doi: 10.1097/WNR.0b013e32801776bb
22. Tessitore A, Russo A, Conte F, Giordano A, De Stefano M, Lavorgna L, et al. Abnormal connectivity within executive resting-state network in migraine with aura. Headache. (2015). 55:794–805. doi: 10.1111/head.12587
23. Leao A. Spreading depression of activity in the cerebral cortex. J Neurophysiol. (1944) 7:359–90. doi: 10.1152/jn.1944.7.6.359
24. Hadjikhani N, Sanchez Del Rio M, Wu O, Schwartz D, Bakker D, Fischl B, et al. Mechanisms of migraine aura revealed by functional MRI in human visual cortex. Proc Natl Acad Sci USA. (2001) 98:4687–92. doi: 10.1073/pnas.071582498
25. Moskowitz MA, Macfarlane R. Neurovascular and molecular mechanisms in migraine headaches. Cerebrovasc Brain Metab Rev. (1993) 5:159–77.
26. Moskowitz MA, Nozaki K, Kraig RP. Neocortical spreading depression provokes the expression of c-fos protein-like immunoreactivity within trigeminal nucleus caudalis via trigeminovascular mechanisms. J Neurosci. (1993) 13:1167–77. doi: 10.1523/JNEUROSCI.13-03-01167.1993
27. Connolly JF, Gawel M, Rose FC. Migraine patients exhibit abnormalities in the visual evoked potential. J Neurol Neurosurg Psychiatry. (1982) 45:464–7. doi: 10.1136/jnnp.45.5.464
28. Gawel M, Connolly JF, Rose FC. Migraine patients exhibit abnormalities in the visual evoked potential. Headache. (1983) 23:49–52. doi: 10.1111/j.1526-4610.1983.hed2302049.x
29. Diener HC, Scholz E, Dichgans J, Gerber WD, Jack A, Bille A, et al. Central effects of drugs used in migraine prophylaxis evaluated by visual evoked potentials. Ann Neurol. (1989) 25:125–30. doi: 10.1002/ana.410250204
30. Oelkers R, Grosser K, Lang E, Geisslinger G, Kobal G, Brune K, et al. Visual evoked potentials in migraine patients: alterations depend on pattern spatial frequency. Brain. (1999) 122:1147–55. doi: 10.1093/brain/122.6.1147
31. Sand T, Zhitniy N, White LR, Stovner LJ. Visual evoked potential latency, amplitude and habituation in migraine: a longitudinal study. Clin Neurophysiol. (2008) 119:1020–7. doi: 10.1016/j.clinph.2008.01.009
32. Coppola G, Bracaglia M, Di Lenola D, Di Lorenzo C, Serrao M, Parisi V, et al. Visual evoked potentials in subgroups of migraine with aura patients. J Headache Pain. (2015) 16:92. doi: 10.1186/s10194-015-0577-6
33. Aurora SK, Ahmad BK, Welch KM, Bhardhwaj P, Ramadan NM. Transcranial magnetic stimulation confirms hyperexcitability of occipital cortex in migraine. Neurology. (1998) 50:1111–4. doi: 10.1212/WNL.50.4.1111
34. Brigo F, Storti M, Tezzon F, Manganotti P, Nardone R. Primary visual cortex excitability in migraine: a systematic review with meta-analysis. Neurol Sci. (2013) 34:819–30. doi: 10.1007/s10072-012-1274-8
35. Brighina F, Bolognini N, Cosentino G, Maccora S, Paladino P, Baschi R, et al. Visual cortex hyperexcitability in migraine in response to sound-induced flash illusions. Neurology. (2015) 84:2057–61. doi: 10.1212/WNL.0000000000001584
36. Younis S, Hougaard A, Vestergaard MB, Larsson HBW, Ashina M. Migraine and magnetic resonance spectroscopy: a systematic review. Curr Opin Neurol. (2017) 30:246–62. doi: 10.1097/WCO.0000000000000436
37. Tajti J, Szok D, Nagy-Grocz G, Tuka B, Petrovics-Balog A, Toldi J, et al. Kynurenines and PACAP in migraine: medicinal chemistry and pathogenetic aspects. Curr Med Chem. (2017) 24:1332–49. doi: 10.2174/0929867324666170227115019
38. Bindman LJ, Lippold OC, Redfearn JW. The action of brief polarizing currents on the cerebral cortex of the rat (1) during current flow and (2) in the production of long-lasting after-effects. J Physiol. (1964) 172:369–82. doi: 10.1113/jphysiol.1964.sp007425
39. Wolf ME, Jäger T, Bäzner H, Hennerici M. Changes in functional vasomotor reactivity in migraine with aura. Cephalalgia. (2009) 29:1156–64. doi: 10.1111/j.1468-2982.2009.01843.x
40. Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U, May A. Neuroplasticity: changes in grey matter induced by training. Nature. (2004) 427:311–2. doi: 10.1038/427311a
41. Boyke J, Driemeyer J, Gaser C, Buchel C, May A. Training-induced brain structure changes in the elderly. J Neurosci. (2008) 28:7031–5. doi: 10.1523/JNEUROSCI.0742-08.2008
42. Scholz J, Klein MC, Behrens TE, Johansen-Berg H. Training induces changes in white-matter architecture. Nat Neurosci. (2009) 12:1370–1. doi: 10.1038/nn.2412
43. Teutsch S, Herken W, Bingel U, Schoell E, May A. Changes in brain gray matter due to repetitive painful stimulation. Neuroimage. (2008) 42:845–9. doi: 10.1016/j.neuroimage.2008.05.044
44. Dehbandi S, Speckmann EJ, Pape HC, Gorji A. Cortical spreading depression modulates synaptic transmission of the rat lateral amygdala. Eur J Neurosci. (2008) 27:2057–65. doi: 10.1111/j.1460-9568.2008.06188.x
45. Haghir H, Kovac S, Speckmann EJ, Zilles K, Gorji A. Patterns of neurotransmitter receptor distributions following cortical spreading depression. Neuroscience. (2009) 163:1340–52. doi: 10.1016/j.neuroscience.2009.07.067
46. Moskowitz MA. Pathophysiology of headache–past and present. Headache. (2007) 47(Suppl 1):S58–63. doi: 10.1111/j.1526-4610.2007.00678.x
47. Arnold G, Reuter U, Kinze S, Wolf T, Einhaupl KM. Migraine with aura shows gadolinium enhancement which is reversed following prophylactic treatment. Cephalalgia. (1998) 18:644–6. doi: 10.1046/j.1468-2982.1998.1809644.x
48. Knotkova H, Pappagallo M. Imaging intracranial plasma extravasation in a migraine patient: a case report. Pain Med. (2007) 8:383–7. doi: 10.1111/j.1526-4637.2006.00197.x
49. Cui Y, Takashima T, Takashima-Hirano M, Wada Y, Shukuri M, Tamura Y, et al. 11C-PK11195 PET for the in vivo evaluation of neuroinflammation in the rat brain after cortical spreading depression. J Nucl Med. (2009) 50:1904–11. doi: 10.2967/jnumed.109.066498
50. D'Andrea G, Cananzi AR, Joseph R, Morra M, Zamberlan F, Ferro Milone F, et al. Platelet glycine, glutamate and aspartate in primary headache. Cephalalgia. (1991) 11:197–200. doi: 10.1046/j.1468-2982.1991.1104197.x
51. Longoni M, Ferrarese C. Inflammation and excitotoxicity: role in migraine pathogenesis. Neurol Sci. (2006) 27(Suppl 2):S107–10. doi: 10.1007/s10072-006-0582-2
52. Gursoy-Ozdemir Y, Qiu J, Matsuoka N, Bolay H, Bermpohl D, Jin H, et al. Cortical spreading depression activates and upregulates MMP-9. J Clin Invest. (2004) 113:1447–55. doi: 10.1172/JCI200421227
53. Bernecker C, Pailer S, Kieslinger P, Horejsi R, Moller R, Lechner A, et al. Increased matrix metalloproteinase activity is associated with migraine and migraine-related metabolic dysfunctions. Eur J Neurol. (2010) 18:571–6. doi: 10.1111/j.1468-1331.2010.03205.x
54. Gupta VK. CSD, BBB and MMP-9 elevations: animal experiments versus clinical phenomena in migraine. Expert Rev Neurother. (2009) 9:1595–614. doi: 10.1586/ern.09.103
55. Yilmaz N, Karaali K, Ozdem S, Turkay M, Unal A, Dora B. Elevated S100B and neuron specific enolase levels in patients with migraine-without aura: evidence for neurodegeneration? Cell Mol Neurobiol. (2011) 31:579–85. doi: 10.1007/s10571-011-9651-z
56. Pisanu C, Preisig M, Castelao E, Glaus J, Pistis G, Squassina A, et al. A genetic risk score is differentially associated with migraine with and without aura. Hum Genet. (2017) 136:999–1008. doi: 10.1007/s00439-017-1816-5
57. Yang MH, Yang FY, Oyang YJ. Application of density estimation algorithms in analyzing co-morbidities of migraine. Netw Model Anal Health Inform Bioinform. (2013) 2:95–107. doi: 10.1007/s13721-013-0028-8
58. Roos-Araujo D, Stuart S, Lea RA, Haupt LM, Griffiths LR. Epigenetics and migraine; complex mitochondrial interactions contributing to disease susceptibility. Gene. (2014) 543:1–7. doi: 10.1016/j.gene.2014.04.001
59. Knyihar-Csillik E, Tajti J, Mohtasham S, Sari G, Vecsei L. Electrical stimulation of the Gasserian ganglion induces structural alterations of calcitonin gene-related peptide-immunoreactive perivascular sensory nerve terminals in the rat cerebral dura mater: a possible model of migraine headache. Neurosci Lett. (1995) 184:189–92. doi: 10.1016/0304-3940(94)11203-U
60. Knyihar-Csillik E, Tajti J, Chadaide Z, Csillik B, Vecsei L. Functional immunohistochemistry of neuropeptides and nitric oxide synthase in the nerve fibers of the supratentorial dura mater in an experimental migraine model. Microsc Res Tech. (2001) 53:193–211. doi: 10.1002/jemt.1084
61. Knyihar-Csillik E, Chadaide Z, Okuno E, Krisztin-Peva B, Toldi J, Varga C, et al. Kynurenine aminotransferase in the supratentorial dura mater of the rat: effect of stimulation of the trigeminal ganglion. Exp Neurol. (2004) 186:242–7. doi: 10.1016/j.expneurol.2003.12.001
62. Villalon CM, Olesen J. The role of CGRP in the pathophysiology of migraine and efficacy of CGRP receptor antagonists as acute antimigraine drugs. Pharmacol Ther. (2009) 124:309–23. doi: 10.1016/j.pharmthera.2009.09.003
63. Olesen J. Calcitonin gene-related peptide. (CGRP) in migraine. Cephalalgia. (2010). doi: 10.1177/0333102410388438
64. Tajti J, Pardutz A, Vamos E, Tuka B, Kuris A, Bohar Z, et al. Migraine is a neuronal disease. J Neural Transm. (2010) 118:511–24. doi: 10.1007/s00702-010-0515-3
65. Tuka B, Helyes Z, Markovics A, Bagoly T, Nemeth J, Mark L, et al. Peripheral and central alterations of pituitary adenylate cyclase activating polypeptide-like immunoreactivity in the rat in response to activation of the trigeminovascular system. Peptides. (2012) 33:307–16. doi: 10.1016/j.peptides.2011.12.019
66. Akerman S, Goadsby PJ. Neuronal PAC1 receptors mediate delayed activation and sensitization of trigeminocervical neurons: Relevance to migraine. Sci Transl Med. (2015) 7:308ra157. doi: 10.1126/scitranslmed.aaa7557
67. Tozzi A, de Iure A, Di Filippo M, Costa C, Caproni S, Pisani A, et al. Critical role of calcitonin gene-related peptide receptors in cortical spreading depression. Proc Natl Acad Sci USA. (2012) 109:18985–90. doi: 10.1073/pnas.1215435109
68. Chan KY, Labastida-Ramirez A, Ramirez-Rosas MB, Labruijere S, Garrelds IM, Danser AH, et al. Trigeminovascular calcitonin gene-related peptide function in Cacna1a R192Q-mutated knock-in mice. J Cereb Blood Flow Metab. (2017) 39:718–29. doi: 10.1177/0271678X17725673
69. Tuka B, Helyes Z, Markovics A, Bagoly T, Szolcsanyi J, Szabo N, et al. Alterations in PACAP-38-like immunoreactivity in the plasma during ictal and interictal periods of migraine patients. Cephalalgia. (2013) 33:1085–95. doi: 10.1177/0333102413483931
70. Vereb D, Szabo N, Tuka B, Tajti J, Kiraly A, Farago P, et al. Correlation of neurochemical and imaging markers in migraine: PACAP38 and DTI measures. Neurology. (2018) 91:e1166–74. doi: 10.1212/WNL.0000000000006201
71. Dalkara T, Zervas NT, Moskowitz MA. From spreading depression to the trigeminovascular system. Neurol Sci. (2006) 27(Suppl 2):S86–90. doi: 10.1007/s10072-006-0577-z
72. Charles A. Migraine: a brain state. Curr Opin Neurol. (2013) 26:235–9. doi: 10.1097/WCO.0b013e32836085f4
73. Jürgens TP, Schulte LH, May A. Migraine trait symptoms in migraine with and without aura. Neurology. (2014) 82:1416–24. doi: 10.1212/WNL.0000000000000337
74. Fazekas F, Koch M, Schmidt R, Offenbacher H, Payer F, Freidl W, et al. The prevalence of cerebral damage varies with migraine type: a MRI study. Headache. (1992) 32:287–91. doi: 10.1111/j.1526-4610.1992.hed3206287.x
75. Pavese N, Canapicchi R, Nuti A, Bibbiani F, Lucetti C, Collavoli P, et al. White matter MRI hyperintensities in a hundred and twenty-nine consecutive migraine patients. Cephalalgia. (1994) 14:342–5. doi: 10.1046/j.1468-2982.1994.1405342.x
Keywords: DTI, functional MRI, microstructure, migraine with and without aura, pathomechanism
Citation: Kincses ZT, Veréb D, Faragó P, Tóth E, Kocsis K, Kincses B, Király A, Bozsik B, Párdutz Á, Szok D, Tajti J, Vécsei L, Tuka B and Szabó N (2019) Are Migraine With and Without Aura Really Different Entities? Front. Neurol. 10:982. doi: 10.3389/fneur.2019.00982
Received: 30 June 2019; Accepted: 28 August 2019;
Published: 02 October 2019.
Edited by:
Massimo Filippi, Vita-Salute San Raffaele University, ItalyReviewed by:
Claudia Altamura, Campus Bio-Medico University, ItalyCopyright © 2019 Kincses, Veréb, Faragó, Tóth, Kocsis, Kincses, Király, Bozsik, Párdutz, Szok, Tajti, Vécsei, Tuka and Szabó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zsigmond Tamás Kincses, a2luY3Nlcy56c2lnbW9uZC50YW1hc0BtZWQudS1zemVnZWQuaHU=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.