 Chiara La Morgia
Chiara La Morgia Valerio Carelli
Valerio Carelli Michele Carbonelli
Michele Carbonelli- 1Unità Operativa Complessa Clinica Neurologica, IRCCS Istituto delle Scienze Neurologiche di Bologna, Ospedale Bellaria, Bologna, Italy
- 2Dipartimento di Scienze Biomediche e Neuromotorie, Università di Bologna, Bologna, Italy
Melanopsin retinal ganglion cells (mRGCs) are intrinsically photosensitive RGCs that mediate many relevant non-image forming functions of the eye, including the pupillary light reflex, through the projections to the olivary pretectal nucleus. In particular, the post-illumination pupil response (PIPR), as evaluated by chromatic pupillometry, can be used as a reliable marker of mRGC function in vivo. In the last years, pupillometry has become a promising tool to assess mRGC dysfunction in various neurological and neuro-ophthalmological conditions. In this review we will present the most relevant findings of pupillometric studies in glaucoma, hereditary optic neuropathies, ischemic optic neuropathies, idiopathic intracranial hypertension, multiple sclerosis, Parkinson's disease, and mood disorders. The use of PIPR as a marker for mRGC function is also proposed for other neurodegenerative disorders in which circadian dysfunction is documented.
Introduction
Melanopsin retinal ganglion cells (mRGCs) are intrinsically photosensitive RGCs expressing the photopigment melanopsin (1, 2). They constitute about 0.2–1% of total RGCs and contribute to the photoentrainment of circadian rhythms, through their projections to the suprachiasmatic nucleus (SCN), but also to other anatomical structures devoted to non-image forming functions of the eye. These include pupil regulation through their projections to the olivary pretectal nucleus (OPN) in the midbrain (3–5) and brain structures relevant for emotional processing (6). Recent data support the notion that distinct subpopulations of mRGCs mediate different functions in the central nervous system, including circadian rhythm regulation and pupil light reflex (PLR) through their projections to the OPN (5, 7).
The mRGC contribution to the pupil function has been extensively investigated over the recent years and it is now clear that rods mediate mainly the transient pupil contraction, whereas mRGCs contribute to the steady-state pupil constriction (8–11). In fact, mRGCs are characterized by a unique property, which is the capability of firing without fatigue in response to continuous stimulation, consistent with the intrinsic activation of these cells (12). In particular, post-illumination pupil response (PIPR), measured after 1.7 s from onset of the light stimulus, and its magnitude can be considered as specific measures of mRGC function (13). Different protocols, using different light paradigms and experimental setting of stimulation, have been tested and now established to assess in vivo PLR mediated by mRGCs (13, 14). Specifically, the contribution of mRGCs to pupil response has been evaluated using blue (470 nm) and red (640 nm) light, being the blue light able to maximally stimulating mRGCs (13–15).
The PLR mediated by mRGCs has been investigated in different ophthalmological conditions including glaucoma (16–18), retinitis pigmentosa (19), diabetes (20), Leber's congenital amaurosis (14), age-related macular degeneration (21), and ischemic optic neuropathies (22, 23). Moreover, various neurological and psychiatric disorders have been evaluated, including hereditary optic neuropathies (24–29), seasonal affective disorder (SAD) (30), idiopathic intracranial hypertension (IIH) (31, 32), multiple sclerosis (MS) (33), and Parkinson's disease (PD) (34).
In this review we will focus on pupillometry findings in neuro-ophthalmological disorders in which pupil and circadian functions have been investigated. In particular, we include disorders affecting the optic nerve such as glaucoma and hereditary optic neuropathies, neurodegenerative disorders with optic nerve involvement and circadian dysfunction and affective disorders for which a relevant role of mRGCs has been postulated. We will highlight the potential role of mRGC-mediated pupil function as an in vivo objective tool and possible biomarker for evaluating mRGC function in different neurodegenerative disorders.
Melanopsin RGCs and Pupil in Glaucoma and Anterior Ischemic Optic Neuropathy
Glaucoma is a chronic optic neuropathy characterized by loss of peripheral visual field secondary to a progressive and extensive loss of RGCs and their optic nerve fibers (35). The pathophysiology of glaucoma is not yet completely understood, even though two common and pivotal events are the increase in intraocular pressure and impaired microcirculation (vascular deregulation), both preceding the RGC death (36). Previous studies in monkey models of glaucoma reported that all classes of RGCs are susceptible to injury or damage since the early stages of the disease including the sub-population of mRGCs (37). Concordantly, recent clinical studies have shown high prevalence of sleep and circadian disorders, as well as depression in glaucoma patients, implying that the mRGC-driven photoentrainment of circadian rhythms may be affected in patients with glaucoma (38–41).
In the last years, several studies were published aimed at measuring in vivo the integrity of mRGC system in glaucoma by assessing the PLR (16–18). Overall, the results and the conclusions of these studies have been frequently inconsistent because of the different protocols and methodology adopted for chromatic pupillography. In fact, many variables may affect the results, such as time of dark adaptation, light stimulus (duration, intensity, and wavelength), time to measure the intrinsic melanopsin-mediated PIPR, direct or consensual pupil stimulation, and so on (see Table 1).
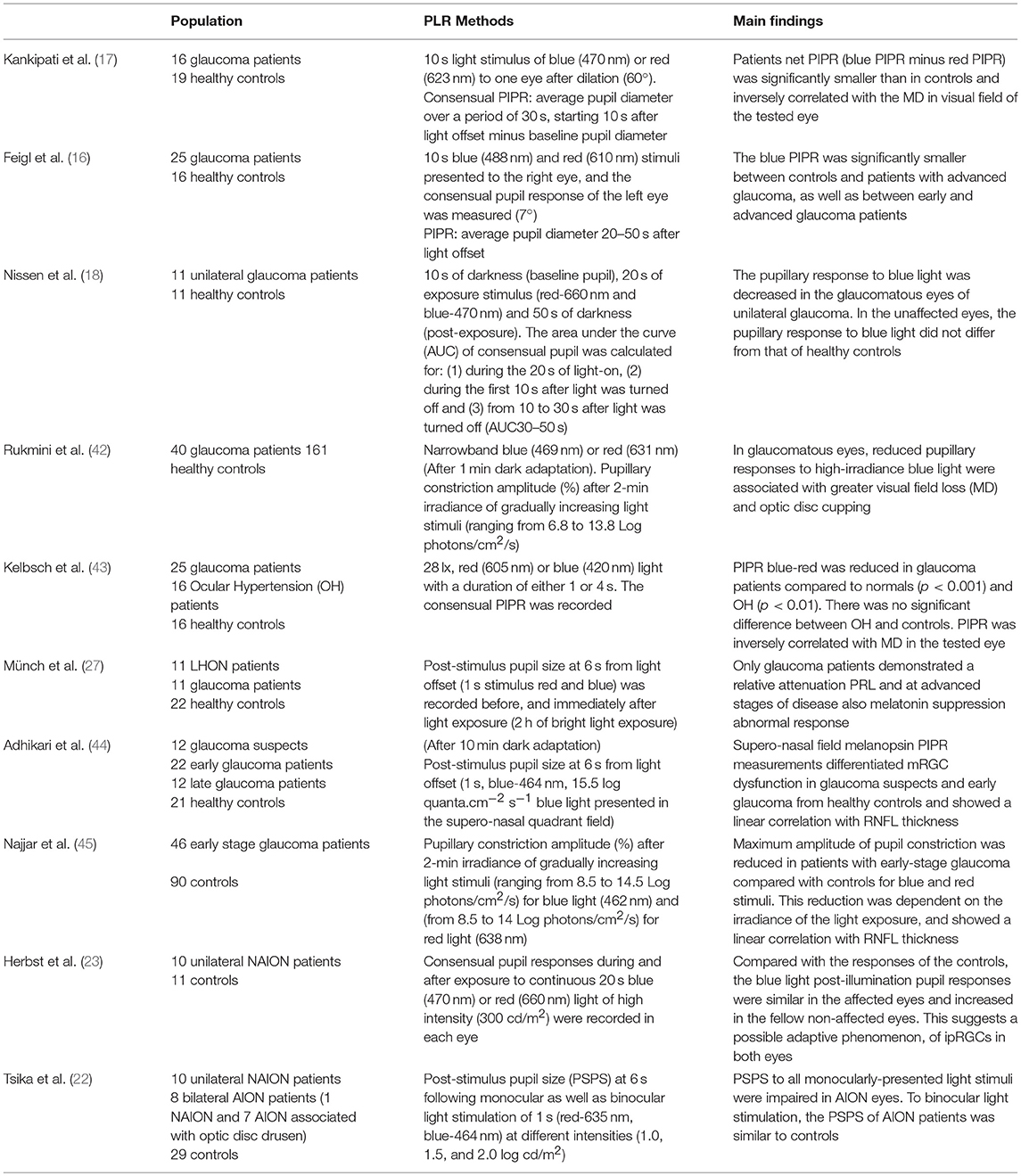
Table 1. Pupillometry findings in glaucoma and in anterior ischemic optic neuropathy.
Nonetheless, it is now clearly proven that the PLR, and particularly the PIPR, is altered in moderate and advanced stages of glaucoma, despite the use of different chromatic illumination paradigms (16–18, 27, 42, 43). These findings are also correlated with functional and structural features of the glaucomatous pathology, as demonstrated by the fact that PIPR is inversely correlated with the mean deviation in the visual fields (17, 42, 43). Moreover, an inverse correlation between PLR to high-irradiance blue light and optic disc cupping measured by Heidelberg Retinal Tomography was found (42), and the reduction of PLR to blue and red light correlates with retinal nerve fiber layer (RNFL) thinning (44, 45). These results are in line with a study demonstrating that there is a correlation between the severity of the glaucoma and the reduction of the PIPR (16). This is concordant with the knowledge that in glaucoma the central 10 degrees of the retina, where the mRGCs are more concentrated, are affected only in the last stages of the disease. However, in the last 2 years, a new method of light delivery (quadrant field pupillometry) (44), and a new light stimulation protocol (increasing light regimens) (45) were used to better investigate the pre-perimetric and early-stage glaucoma. By stimulating only the portion of the retina most precociously affected in glaucoma it was shown that the superonasal quadrant PIPR differentiates patients suspected of having glaucoma and with early glaucoma from healthy controls, and this finding correlated with RNFL thinning measured by OCT (44). Furthermore, by increasing logarithmically the light stimuli intensity, PLR is reduced in patients with early-stage glaucoma compared with controls at moderate to high irradiances with both blue and red light, and the maximal pupillary constriction amplitude is correlated to the RNFL thickness (45). To highlight the possible correlation of different measurements of mRGC functions, it is also worth mentioning that in advanced glaucoma, individuals with greater light-induced melatonin suppression (a measure of the retino-hypothalamic tract function) have also a smaller PIPR (27).
Finally, a functional damage of the mRGC-mediated PLR has been reported in the affected eyes of patients with unilateral or bilateral anterior ischemic optic neuropathy (AION), specifically 10 patients with unilateral non-arteritic ischemic optic neuropathy (NAION), 1 bilateral NAION, and 7 patients with bilateral AION associated with optic disc drusen, compared to the unaffected and control eyes (22). Differently, previous studies failed to demonstrate differences in the PLR between NAION and control eyes (23). Furthermore, if the bright blue stimuli were presented bilaterally and simultaneously to both eyes, bilateral AION patients showed, through binocular summation, the same post-stimulus pupil size of patients with unilateral AION and controls (22).
Melanopsin RGCs and Pupil in Hereditary Mitochondrial Optic Neuropathies
Mitochondrial optic neuropathies are inherited disorders of the optic nerve due to mitochondrial DNA (mtDNA) mutations affecting the mitochondrial-encoded subunits of complex I of the respiratory chain complex, pathogenic for Leber's hereditary Optic Neuropathy (LHON) or to mutations of the nuclear gene OPA1 causing Dominant Optic Atrophy (DOA) (46, 47). These inherited mitochondrial disorders are characterized by the selective loss of RGCs, in particular those originating the papillo-macular bundle, thus leading to optic atrophy secondary to mitochondrial dysfunctions with the invariable outcome of severe visual loss (46). In both disorders, previous data suggested the maintenance of the PLR even in the chronic stage of the disease, pointing to a pupil-visual dissociation (48, 49). In fact, in these disorders recent histological studies demonstrated a relative preservation of mRGCs compared to the massive loss of regular RGCs in both LHON and DOA, which supports the maintenance of the PLR in these patients (50). At this regard, interestingly, a previous post mortem study demonstrated the relative sparing of the retinofugal fibers to the pretectum in a LHON case, supporting the maintenance of the mRGC projections to the pretectum, which constitute the afferent pathway of the PLR (51). The reasons for the robustness of mRGCs in mitochondrial optic neuropathies are still unknown and under investigation, even though the possible role of peculiar metabolic properties, including the size of the soma, has been proposed (50, 52, 53). More recently, pupillometric studies showed a relative maintenance of the mRGC-mediated pupil response in LHON and DOA patients (24–29) (Table 2). Similarly to the maintenance of the PLR a preserved light-induced melatonin suppression has been demonstrated in LHON and DOA patients supporting a relative preservation of these cells in hereditary mitochondrial optic neuropathies (50). Interestingly, the preservation of mRGCs and PLR has also been demonstrated in an OPA1-mouse model (54).
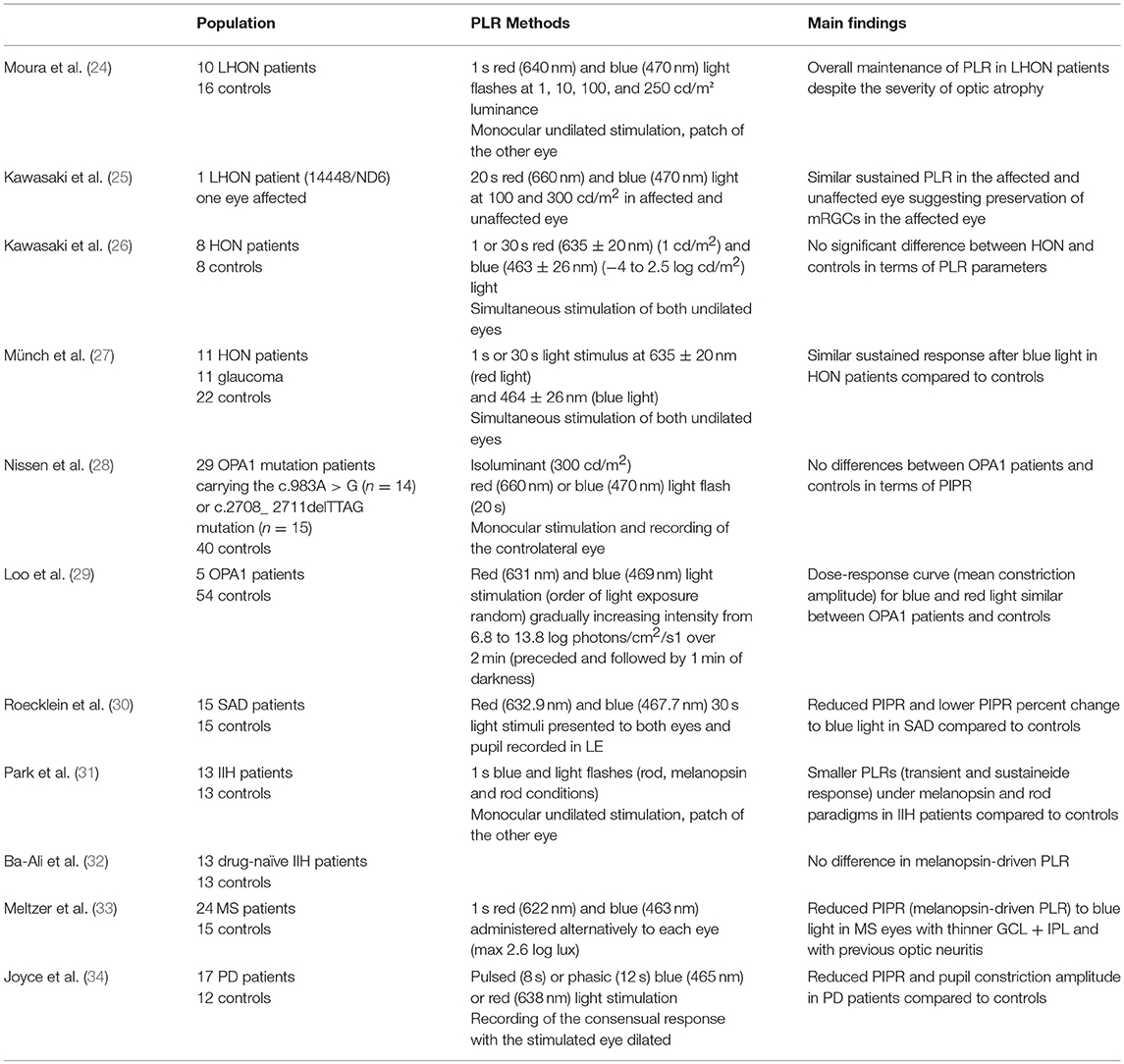
Table 2. Pupillometry findings in neurological disorders.
Melanopsin RGCs and Pupil in Other Neurological Disorders
In the last years the mRGC-mediated pupil light response has been investigated in various neurological disorders, including IIH, MS, and PD (31–34).
In a cohort of 13 IIH patients compared to 13 controls it was reported a significant reduction of PLR under melanopsin and rod paradigms in IIH subjects, suggesting the potential use of these parameters as an objective measure of RGC dysfunction in IIH (31). However, the abnormal mRGC-driven PLR has not been reported in a cohort of drug naïve IIH patients (32).
A significant reduction of the sustained pupil response to blue light in the eyes with thinner ganglion cell layer (GCL) + inner plexiform layer (IPL) was demonstrated in a group of 24 MS patients, in particular in those with a previous history of optic neuritis, compared to 15 controls (33). The authors proposed the use of the sustained pupil response to light mediated by mRGCs as a surrogate biomarker for neurodegeneration, including the retinohypothalamic tract, in MS patients (33). In consideration that mRGCs are a fundamental conduit for circadian photoentrainment, the sustained PLR to light may be used as a surrogate marker for RHT integrity and consequently for circadian measurements including melatonin rhythm. This may be relevant for potential light therapeutic interventions in these patients (33). Congruently, previous studies demonstrated an abnormal melatonin rhythm in MS patients (55).
An attenuated PIPR for short wavelength and reduced pupil constriction amplitude for long wavelength stimulation was described in a group of 17 early PD patients compared to a control group (34). Pupil metrics in this group were not influenced by disease severity, sleep quality, medications, or OCT measurements and were controlled for unrest pupil conditions. The authors proposed the pupil response mediated by mRGCs as potential biomarker for non-motor symptoms in PD, such as sleep and circadian dysfunction (34). In fact, there is a large body of evidence supporting the occurrence of circadian dysfunction in PD (56).
Finally, a recent study reported the occurrence of PLR dysfunction in R6/2 and Q175 Huntington's disease (HD) mouse models, with a prevalent contribution of cone dysfunction in young-middle-aged mice and of mRGCs in old mice (57). HD is a neurodegenerative disorder in which circadian dysfunction is a prominent and early disease trait pointing again to a possible mRGC dysfunction (56, 58).
Based on these recent findings, it seems reasonable that other neurological disorders, for which there is evidence of circadian dysfunction and mRGC pathology, such as Alzheimer's disease (AD) (59, 60), HD (56–58), and possibly others, may present an abnormal mRGC-driven PLR.
Melanopsin RGCs and Pupil in Affective Disorders
SAD is a psychiatric condition characterized by the recurrence of depression in winter, in relation to low levels of ambient light in this season (61). Even if the etiology of this disorder is still elusive, the possible role of individual seasonal variation in retinal sensitivity, and in particular of retinal subsensitivity in SAD has been proposed (62–64). Moreover, a polymorphism in the melanopsin (OPN4) gene (P10L) has been associated with SAD, suggesting that mRGCs and sensitivity to light may play a relevant role in the pathogenesis of SAD (65). Based on these premises, Roecklein and coauthors investigated the PIPR in 15 individuals with SAD compared to 15 controls. They found a reduced PIPR and a lower PIPR percent change in response to blue light in SAD subjects compared to controls, implying an abnormal mRGC-mediated response to light, as measured by PLR in SAD (30). Moreover, the PIPR response after blue light varied in relation to the OPN4 I394T genotype, another polymorphic variant, suggesting again a possible influence of genetic predisposition in modulating the sensitivity to light in SAD (30). Interestingly, this polymorphic variant has also been found to influence the steady-state pupil diameter in controls (66).
Differently, the melanopsin-mediated PIPR measurements were not significantly different between eight patients with non-seasonal depression and 13 age-matched healthy controls matched for day-light exposure (67). This finding possibly implies a different pathophysiological mechanism in SAD and non-seasonal depression. However, another study using a different light stimulation protocol, showed an abnormal PIPR in both seasonal-depressed and non-seasonal depressed patients (68).
Discussion
Intrinsically photosensitive retinal ganglion cells, the mRGCs, are unique photoreceptors located in the inner retina, which express the photopigment melanopsin (1, 2, 7). The presence of melanopsin makes these cells maximally sensitive to blue light at 470–480 nm and able to spontaneously spike for a long period, even when isolated from the surrounding retinal structures (7, 12). The mRGCs are crucial for non-image forming functions of the eye including circadian photoentrainment, sleep and melatonin synthesis, and PLR. Of particular importance, in this context, is the possibility of using some pupil metrics, such as the PIPR, as a specific signature of mRGC function in vivo (8, 13, 56) for ophthalmological and neurodegenerative disorders, which may present circadian dysfunction. In fact, mRGCs contribute mainly to the sustained component of the PLR and, using blue wavelength light, it is possible to isolate the melanopsin contribution to the PLR.
The availability of the mRGC-mediated PLR as a tool to indirectly test the circadian system status, as recently proposed (69), opens new avenues in the analysis of circadian, sleep, and non-motor features in many neurodegenerative disorders. Interestingly, it has been demonstrated in 15 healthy subjects, using combined evaluations including pupillometry, actigraphy, light sensors and body temperature, a close inverse relationship between pupil light response metrics and circadian status (70). In particular, for the pupil recordings it was used a protocol in which the right eye was dilated and different light stimuli including different light wavelength were tested (5 min stimuli) with 40 min of darkness between the light stimulations. Pupil parameters were analyzed using ad-hoc software. For the actigraphic recordings the subjects wore an actigraph with light sensor and non-parametric circadian measures, such as intradaily variability, interdaily stability, relative amplitude, L5 and M5, were obtained (70). The authors proposed the Circadian Status Index as an integrative measure to unify three aspects (robustness, timing, and level) of the three circadian rhythm measures (temperature, activity, and light), as well as a global parameter for pupil metrics (circadian photoreception PLR). However, the authors found an inverse relationship between the pupil and circadian metrics. These contradictory findings between circadian status robustness and the PLR might be referred to individual differences in the M1 cell population of mRGCs. Larger studies, more uniform light stimulation protocols and the inclusion of more circadian and pupil metrics are warranted to analyze the possible correlations between pupil metrics and circadian status. In fact, the current available pupillometric studies all suffer the limitation of great heterogeneity of stimulation protocols and consequent lack of reproducibility of their results. Similarly, all these studies are generally underpowered by the limited number of subjects analyzed.
Finally, since mRGCs are contributing to other non-visual functions of the eye, and different class of mRGCs have different projections to the CNS contributing to different functions (5), it must be emphasized that the finding of an abnormal mRGC-mediated PLR does not mean necessarily a global dysfunction of these cells. Overall, the availability of an easily accessible metric for mRGC function, in conjunction with other tests, such as melatonin suppression test, actigraphic recordings, and functional MRI, may represent a comprehensive strategy to further exploring the function of these cells in patients with different neuro-ophthalmological conditions.
Conclusions and Future Directions
In conclusion, the use of PLR mediated by mRGCs, as a measure of mRGC function, is of particular relevance for neurodegenerative disorders for which there is already evidence of circadian and sleep dysfunction, such as PD, AD, and HD. Similarly, it might be also relevant for other neurological disorders with evidence of circadian dysfunction such as frontotemporal dementia (71), Lewy-Body dementia (72), Progressive Supranuclear Palsy (73, 74), and possibly prion diseases, in particular fatal familial insomnia (75). Moreover, the study of PLR mediated by mRGCs might be particularly intriguing for conditions, in which light sensitivity is a predominant feature, such as photophobia (76, 77) and photosensitivity in epilepsy (78). At this regard, an abnormal PLR has been recently documented in migraineous photophobic subjects, even though it was not specifically assessed the mRGC contribution to PLR (79, 80).
Overall, after adequate standardization of light protocols, the availability of an easy accessible tool to assess mRGC function, as a surrogate marker for more general non-image forming functions of the eye, including circadian rhythms and sleep, is a particularly promising biomarker for neurodegenerative disorders.
Author Contributions
CLM and MC were responsible for conception, design, drafting, and revision of the manuscript. VC was responsible for conception and revision of the manuscript.
Funding
This work was supported by the Ministry of Health Young Researcher project (GR-2013-02358026) (to CLM) and by the Italian Ministry of Health and of Research and the Gino Galletti Foundation (to VC).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Berson DM, Dunn FA, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science (2002) 295:1070–3. doi: 10.1126/science.1067262
2. Hattar S, Liao HW, Takao M, Berson DM, Yau KW. Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science (2002) 295:1065–70. doi: 10.1126/science.1069609
3. Hannibal J, Hindersson P, Ostergaard J, Georg B, Heegaard S, Larsen PJ, et al. Melanopsin is expressed in PACAP-containing retinal ganglion cells of the human retinohypothalamic tract. Invest Ophthalmol Vis Sci. (2004) 45:4202–9. doi: 10.1167/iovs.04-0313
4. Hannibal J, Christiansen AT, Heegaard S, Fahrenkrug J, Kiilgaard JF. Melanopsin expressing human retinal ganglion cells: subtypes, distribution, and intraretinal connectivity. J Comp Neurol. (2017) 525:1934–61. doi: 10.1002/cne.24181
5. Chen SK, Badea TC, Hattar S. Photoentrainment and pupillary light reflex are mediated by distinct populations of ipRGCs. Nature (2011) 476:92–5. doi: 10.1038/nature10206
6. LeGates TA, Altimus CM, Wang H, Lee HK, Yang S, Zhao H, et al. Aberrant light directly impairs mood and learning through melanopsin-expressing neurons. Nature (2012) 491:594–8. doi: 10.1038/nature11673
7. Sand A, Schmidt TM, Kofuji P. Diverse types of ganglion cell photoreceptors in the mammalian retina. Prog Retin Eye Res. (2012) 31:287–302. doi: 10.1016/j.preteyeres.2012.03.003
8. Gamlin PD, McDougal DH, Pokorny J, Smith VC, Yau KW, Dacey DM. Human and macaque pupil responses driven by melanopsin-containing retinal ganglion cells. Vision Res. (2007) 47:946–54. doi: 10.1016/j.visres.2006.12.015
9. Kawasaki A, Kardon RH. Intrinsically photosensitive retinal ganglion cells. J Neuroophthalmol. (2007) 27:195–204. doi: 10.1097/WNO.0b013e31814b1df9
10. Young RS, Kimura E. Pupillary correlates of light-evoked melanopsin activity in humans. Vision Res. (2008) 48:862–71. doi: 10.1016/j.visres.2007.12.016
11. Keenan WT, Rupp AC, Ross RA, Somasundaram P, Hiriyanna S, Wu Z, et al. A visual circuit uses complementary mechanisms to support transient and sustained pupil constriction. Elife (2016) 5:e15392. doi: 10.7554/eLife.15392
12. Wong KY. A retinal ganglion cell that can signal irradiance continuously for 10 hours. J Neurosci. (2012) 32:11478–85. doi: 10.1523/JNEUROSCI.1423-12.2012
13. Adhikari P, Zele AJ, Feigl B. The post-illumination pupil response (PIPR). Invest Ophthalmol Vis Sci. (2015) 56:3838–49. doi: 10.1167/iovs.14-16233
14. Park JC, Moura AL, Raza AS, Rhee DW, Kardon RH, Hood DC. Toward a clinical protocol for assessing rod, cone, and melanopsin contributions to the human pupil response. Invest Ophthalmol Vis Sci. (2011) 52:6624–35. doi: 10.1167/iovs.11-7586
15. Kardon R, Anderson SC, Damarjian TG, Grace EM, Stone E, Kawasaki A. Chromatic pupil responses: preferential activation of the melanopsin-mediated versus outer photoreceptor-mediated pupil light reflex. Ophthalmology (2009) 116:1564–73. doi: 10.1016/j.ophtha.2009.02.007
16. Feigl B, Mattes D, Thomas R, Zele AJ. Intrinsically photosensitive (melanopsin) retinal ganglion cell function in glaucoma. Invest Ophthalmol Vis Sci. (2011) 52:4362–7. doi: 10.1167/iovs.10-7069
17. Kankipati L, Girkin CA, Gamlin PD. The post-illumination pupil response is reduced in glaucoma patients. Invest Ophthalmol Vis Sci. (2011) 52:2287–92. doi: 10.1167/iovs.10-6023
18. Nissen C, Sander B, Milea D, Kolko M, Herbst K, Hamard P, et al. Monochromatic pupillometry in unilateral glaucoma discloses no adaptive changes subserved by the ipRGCs. Front Neurol. (2014) 5:15. doi: 10.3389/fneur.2014.00015
19. Kardon R, Anderson SC, Damarjian TG, Grace EM, Stone E, Kawasaki A. Chromatic pupillometry in patients with retinitis pigmentosa. Ophthalmology (2011) 118:376–81. doi: 10.1016/j.ophtha.2010.06.033
20. Feigl B, Zele AJ, Fader SM, Howes AN, Hughes CE, Jones KA, et al. The post-illumination pupil response of melanopsin-expressing intrinsically photosensitive retinal ganglion cells in diabetes. Acta Ophthalmol. (2012) 90:e230–4. doi: 10.1111/j.1755-3768.2011.02226.x
21. Feigl B, Zele AJ. Melanopsin-expressing intrinsically photosensitive retinal ganglion cells in retinal disease. Optom Vis Sci. (2014) 91:894–903. doi: 10.1097/OPX.0000000000000284
22. Tsika C, Crippa SV, Kawasaki A. Differential monocular vs. binocular pupil responses from melanopsin-based photoreception in patients with anterior ischemic optic neuropathy. Sci Rep. (2015) 5:10780. doi: 10.1038/srep10780
23. Herbst K, Sander B, Lund-Andersen H, Wegener M, Hannibal J, Milea D. Unilateral anterior ischemic optic neuropathy: chromatic pupillometry in affected, fellow non-affected and healthy control eyes. Front Neurol. (2013) 4:52. doi: 10.3389/fneur.2013.00052
24. Moura AL, Nagy BV, La Morgia C, Barboni P, Oliveira AG, Salomão SR, et al. The pupil light reflex in Leber's hereditary optic neuropathy: evidence for preservation of melanopsin-expressing retinal ganglion cells. Invest Ophthalmol Vis Sci. (2013) 54:4471–7. doi: 10.1167/iovs.12-11137
25. Kawasaki A, Herbst K, Sander B, Milea D. Selective wavelength pupillometry in Leber hereditary optic neuropathy. Clin Exp Ophthalmol. (2010) 38:322–4. doi: 10.1111/j.1442-9071.2010.02212.x
26. Kawasaki A, Collomb S, Léon L, Münch M. Pupil responses derived from outer and inner retinal photoreception are normal in patients with hereditary optic neuropathy. Exp Eye Res. (2014) 120:161–6. doi: 10.1016/j.exer.2013.11.005
27. Münch M, Léon L, Collomb S, Kawasaki A. Comparison of acute non-visual bright light responses in patients with optic nerve disease, glaucoma and healthy controls. Sci Rep. (2015) 5:15185. doi: 10.1038/srep15185
28. Nissen C, Rönnbäck C, Sander B, Herbst K, Milea D, Larsen M, et al. Dissociation of pupillary post-illumination responses from visual function in confirmed OPA1 c.983A > G and c.2708_2711delTTAG autosomal dominant optic atrophy. Front Neurol. (2015) 6:5. doi: 10.3389/fneur.2015.00005
29. Loo JL, Singhal S, Rukmini AV, Tow S, Amati-Bonneau P, Procaccio V, et al. Multiethnic involvement in autosomal-dominant optic atrophy in Singapore. Eye (2017) 31:475–80. doi: 10.1038/eye.2016.255
30. Roecklein K, Wong P, Ernecoff N, Miller M, Donofry S, Kamarck M, et al. The post illumination pupil response is reduced in seasonal affective disorder. Psychiatr Res. (2013) 210:150–8. doi: 10.1016/j.psychres.2013.05.023
31. Park JC, Moss HE, McAnany JJ. The pupillary light reflex in idiopathic intracranial hypertension. Invest Ophthalmol Vis Sci. (2016) 57:23–9. doi: 10.1167/iovs.15-18181
32. Ba-Ali S, Jensen RH, Larsen LS, Lund-Andersen H, Hamann S. The melanopsin-mediated pupillary light response is not changed in patients with newly diagnosed idiopathic intracranial hypertension. Neuroophthalmology (2017) 42:65–72. doi: 10.1080/01658107.2017.1344251
33. Meltzer E, Sguigna PV, Subei A, Beh S, Kildebeck E, Conger D, et al. Retinal architecture and melanopsin-mediated pupillary response characteristics: a putative pathophysiologic signature for the retino-hypothalamic tract in multiple sclerosis. JAMA Neurol. (2017) 74:574–82. doi: 10.1001/jamaneurol.2016.5131
34. Joyce DS, Feigl B, Kerr G, Roeder L, Zele AJ. Melanopsin-mediated pupil function is impaired in Parkinson's disease. Sci Rep. (2018) 8:7796. doi: 10.1038/s41598-018-26078-0
35. Weinreb RN, Aung T, Medeiros FA. The pathophysiology and treatment of glaucoma: a review. JAMA (2014) 311:1901–11. doi: 10.1001/jama.2014.3192
36. Nickells RW, Howell GR, Soto I, John SW. Under pressure: cellular and molecular responses during glaucoma, a common neurodegeneration with axonopathy. Annu Rev Neurosci. (2012) 35:153–79. doi: 10.1146/annurev.neuro.051508.135728
37. Jakobs TC, Libby RT, Ben Y, John SW, Masland RH. Retinal ganglion cell degeneration is topological but not cell type specific in DBA/2J mice. J Cell Biol. (2005) 171:313–25. doi: 10.1083/jcb.200506099
38. Onen SH, Mouriaux F, Berramdane L, Dascotte JC, Kulik JF, Rouland JF. High prevalence of sleep-disordered breathing in patients with primary open-angle glaucoma. Acta Ophthalmol Scand. (2000) 78:638–41. doi: 10.1034/j.1600-0420.2000.078006638.x
39. Drouyer E, Dkhissi-Benyahya O, Chiquet C, WoldeMussie E, Ruiz G, Wheeler LA, et al. Glaucoma alters the circadian timing system. PLoS ONE (2008) 3:e3931. doi: 10.1371/journal.pone.0003931
40. Lanzani MF, de Zavalía N, Fontana H, Sarmiento MI, Golombek D, Rosenstein RE. Alterations of locomotor activity rhythm and sleep parameters in patients with advanced glaucoma. Chronobiol Int. (2012) 29:911–9. doi: 10.3109/07420528.2012.691146
41. Wang H, Zhang Y, Ding J, Wang N. Changes in the circadian rhythm in patients with primary glaucoma. PLoS ONE (2013) 29:e62841. doi: 10.1371/journal.pone.0062841
42. Rukmini AV, Milea D, Baskaran M, How AC, Perera SA, Aung T, et al. Pupillary responses to high-irradiance blue light correlate with glaucoma severity. Ophthalmology (2015) 122:1777–85. doi: 10.1016/j.ophtha.2015.06.002
43. Kelbsch C, Maeda F, Strasser T, Blumenstock G, Wilhelm B, Wilhelm H, et al. Pupillary responses driven by ipRGCs and classical photoreceptors are impaired in glaucoma. Graefes Arch Clin Exp Ophthalmol. (2016) 254: 1361–70. doi: 10.1007/s00417-016-3351-9
44. Adhikari P, Zele AJ, Thomas R, Feigl B. Quadrant field pupillometry detects melanopsin dysfunction in glaucoma suspects and early glaucoma. Sci Rep. (2016) 6:33373. doi: 10.1038/srep33373
45. Najjar RP, Sharma S, Atalay E, Rukmini AV, Sun C, Lock JZ, et al. Pupillary responses to full-field chromatic stimuli are reduced in patients with early-stage primary open-angle glaucoma. Ophthalmology (2018) 125:1362–71. doi: 10.1016/j.ophtha.2018.02.024
46. Carelli V, Ross-Cisneros FN, Sadun AA. Mitochondrial dysfunction as a cause of optic neuropathies. Prog Retin Eye Res. (2004) 23:53–89. doi: 10.1016/j.preteyeres.2003.10.003
47. Yu-Wai-Man P, Griffiths PG, Chinnery PF. Mitochondrial optic neuropathies-disease mechanisms and therapeutic strategies. Prog Retin Eye Res. (2011) 30:81–114. doi: 10.1016/j.preteyeres.2010.11.002
48. Bremner FD, Tomlin EA, Shallo-Hoffmann J, Votruba M, Smith SE. The pupil in dominant optic atrophy. Invest Ophthalmol Vis Sci. (2001) 42:675–8.
49. Wakakura M, Yokoe J. Evidence for preserved direct pupillary light response in Leber's hereditary optic neuropathy. Br J Ophthalmol. (1995) 79:442–6.
50. La Morgia C, Ross-Cisneros FN, Sadun AA, Hannibal J, Munarini A, Mantovani V, et al. Melanopsin retinal ganglion cells are resistant to neurodegeneration in mitochondrial optic neuropathies. Brain (2010) 133:2426–38. doi: 10.1093/brain/awq155
51. Bose S, Dhillon N, Ross-Cisneros FN, Carelli V. Relative post-mortem sparing of afferent pupil fibers in a patient with 3460 Leber's hereditary optic neuropathy. Graefes Arch Clin Exp Ophthalmol. (2005) 243:1175–9. doi: 10.1007/s00417-005-0023-6
52. La Morgia C, Ross-Cisneros FN, Hannibal J, Montagna P, Sadun AA, Carelli V. Melanopsin-expressing retinal ganglion cells: implications for human diseases. Vision Res. (2011) 51:296–302. doi: 10.1016/j.visres.2010.07.023
53. Georg B, Ghelli A, Giordano C, Ross-Cisneros FN, Sadun AA, Carelli V, et al. Melanopsin-expressing retinal ganglion cells are resistant to cell injury, but not always. Mitochondrion (2017) 36:77–84. doi: 10.1016/j.mito.2017.04.003
54. Perganta G, Barnard AR, Katti C, Vachtsevanos A, Douglas RH, MacLaren RE, et al. Non-image-forming light driven functions are preserved in a mouse model of autosomal dominant optic atrophy. PLoS ONE (2013) 8:e56350. doi: 10.1371/journal.pone.0056350
55. Damasceno A, Moraes AS, Farias A, Damasceno BP, dos Santos LM, Cendes F. Disruption of melatonin circadian rhythm production is related to multiple sclerosis severity: a preliminary study. J Neurol Sci. (2015) 353:166–8. doi: 10.1016/j.jns.2015.03.040
56. La Morgia C, Ross-Cisneros FN, Sadun AA, Carelli V. Retinal ganglion cells and circadian rhythms in Alzheimer's disease, Parkinson's disease, and beyond. Front Neurol. (2017) 8:162. doi: 10.3389/fneur.2017.00162
57. Ouk K, Hughes S, Pothecary CA, Peirson SN, Jennifer Morton A. Attenuated pupillary light responses and downregulation of opsin expression parallel decline in circadian disruption in two different mouse models of Huntington's disease. Hum Mol Genet. (2016) 25:5418–32. doi: 10.1093/hmg/ddw359
58. Morton AJ. Circadian and sleep disorder in Huntington's disease. Exp Neurol. (2013) 243:34–44. doi: 10.1016/j.expneurol.2012.10.014
59. La Morgia C, Ross-Cisneros FN, Koronyo Y, Hannibal J, Gallassi R, Cantalupo G, et al. Melanopsin retinal ganglion cell loss in Alzheimer disease. Ann Neurol. (2016) 79:90–109. doi: 10.1002/ana.24548
60. Feng R, Li L, Yu H, Liu M, Zhao W. Melanopsin retinal ganglion cell loss and circadian dysfunction in Alzheimer's disease. Mol Med Rep. (2016) 13:3397–400. doi: 10.3892/mmr.2016.4966
61. Magnusson A, Partonen T. The diagnosis, symptomatology, and epidemiology of seasonal affective disorder. CNS Spectr. (2005) 10:625–34; quiz 1–14. doi: 10.1017/s1092852900019593
62. Wehr TA, Duncan WC Jr, Sher L, Aeschbach D, Schwartz PJ, Turner EH, et al. A circadian signal of change of season in patients with seasonal affective disorder. Arch Gen Psychiatr. (2001) 58:1108–14. doi: 10.1001/archpsyc.58.12.1108
63. Hébert M, Dumont M, Lachapelle P. Electrophysiological evidence suggesting a seasonal modulation of retinal sensitivity in subsyndromal winter depression. J Affect Disord. (2002) 68:191–202. doi: 10.1016/s0165-0327(00)00192-0
64. Rohan KJ, Roecklein KA, Lacy TJ, Vacek PM. Winter depression recurrence one year after cognitive-behavioral therapy, light therapy, or combination treatment. Behav Ther. (2009) 40:225–38. doi: 10.1016/j.beth.2008.06.004
65. Roecklein KA, Rohan KJ, Duncan WC, Rollag MD, Rosenthal NE, Lipsky RH, et al. A missense variant (P10L) of the melanopsin (OPN4) gene in seasonal affective disorder. J Affect Disord. (2009) 114:279–85. doi: 10.1016/j.jad.2008.08.005
66. Higuchi S, Hida A, Tsujimura S, Mishima K, Yasukouchi A, Lee SI, et al. Melanopsin gene polymorphism I394T is associated with pupillary light responses in a dose-dependent manner. PLoS ONE (2013) 8:e60310. doi: 10.1371/journal.pone.0060310
67. Feigl B, Ojha G, Hides L, Zele AJ. Melanopsin-driven pupil response and light exposure in non-seasonal major depressive disorder. Front Neurol. (2018) 9:764. doi: 10.3389/fneur.2018.00764
68. Berman G, Muttuvelu D, Berman D, Larsen JI, Licht RW, Ledolter J, Kardon, et al. Decreased retinal sensitivity in depressive disorder: a controlled study. Acta Psychiatr Scand. (2018) 137:231–40. doi: 10.1111/acps.12851
69. Markwell EL, Feigl B, Zele AJ. Intrinsically photosensitive melanopsin retinal ganglion cell contributions to the pupillary light reflex and circadian rhythm. Clin Exp Optom. (2010) 93:137–49. doi: 10.1111/j.1444-0938.2010.00479
70. Bonmati-Carrion MA, Hild K, Isherwood C, Sweeney SJ, Revell VL, Skene DJ, et al. Relationship between human pupillary light reflex and circadian system status. PLoS ONE (2016) 11:e0162476. doi: 10.1371/journal.pone.0162476
71. Merrilees J, Hubbard E, Mastick J, Miller BL, Dowling GA. Rest-activity and behavioral disruption in a patient with frontotemporal dementia. Neurocase (2009) 15:515–26. doi: 10.1080/13554790903061371
72. Harper DG, Stopa EG, McKee AC, Satlin A, Fish D, Volicer L. Dementia severity and Lewy bodies affect circadian rhythms in Alzheimer disease. Neurobiol Aging (2004) 25:771–81. doi: 10.1016/j.neurobiolaging.2003.04.009
73. De Pablo-Fernández E, Courtney R, Warner TT, Holton JL. A histologic study of the circadian system in Parkinson disease, multiple system atrophy, and progressive supranuclear palsy. JAMA Neurol. (2018) 75:1008–12. doi: 10.1001/jamaneurol.2018.0640
74. Suzuki K, Miyamoto T, Miyamoto M, Hirata K. The core body temperature rhythm is altered in progressive supranuclear palsy. Clin Auton Res. (2009) 19:65–8. doi: 10.1007/s10286-009-0510-7
75. Roguski A, Gill AC. The role of the mammalian prion protein in the control of sleep. Pathogens (2017) 6:E58. doi: 10.3390/pathogens6040058
76. Noseda R, Copenhagen D, Burstein R. Current understanding of photophobia, visual networks and headaches. Cephalalgia (2018) 1:333102418784750. doi: 10.1177/0333102418784750
77. Noseda R, Kainz V, Jakubowski M, Gooley JJ, Saper CB, Digre K, Burstein R. A neural mechanism for exacerbation of headache by light. Nat Neurosci. (2010) 13:239–45. doi: 10.1038/nn.2475
78. Martins da Silva A, Leal B. Photosensitivity and epilepsy: current concepts and perspectives—A narrative review. Seizure (2017) 50:209–18. doi: 10.1016/j.seizure.2017.04.001
79. Cortez MM, Rea NA, Hunter LA, Digre KB, Brennan KC. Altered pupillary light response scales with disease severity in migrainous photophobia. Cephalalgia (2017) 37:801–11. doi: 10.1177/0333102416673205
Keywords: melanopsin retinal ganglion cells, light, pupil, neurodegeneration, optic nerve, optic neuropathies, Alzheimer, Parkinson
Citation: La Morgia C, Carelli V and Carbonelli M (2018) Melanopsin Retinal Ganglion Cells and Pupil: Clinical Implications for Neuro-Ophthalmology. Front. Neurol. 9:1047. doi: 10.3389/fneur.2018.01047
Received: 02 September 2018; Accepted: 19 November 2018;
Published: 07 December 2018.
Edited by:
Andrew J. Zele, Queensland University of Technology, AustraliaReviewed by:
Prakash Adhikari, Queensland University of Technology, AustraliaJason C. Park, University of Illinois at Chicago, United States
Birgit Sander, University of Copenhagen, Denmark
Copyright © 2018 La Morgia, Carelli and Carbonelli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chiara La Morgia, Y2hpYXJhLmxhbW9yZ2lhQHVuaWJvLml0