 Laurent M. A. Favié1,2*
Laurent M. A. Favié1,2* Arlette R. Cox3Agnes van den Hoogen2Cora H. A. Nijboer4Cacha M. P. C. D. Peeters-Scholte5Frank van Bel2,6Toine C. G. Egberts1,7Carin M. A. Rademaker1
Arlette R. Cox3Agnes van den Hoogen2Cora H. A. Nijboer4Cacha M. P. C. D. Peeters-Scholte5Frank van Bel2,6Toine C. G. Egberts1,7Carin M. A. Rademaker1 Floris Groenendaal2,6
Floris Groenendaal2,6
- 1Department of Clinical Pharmacy, University Medical Center Utrecht, Utrecht, Netherlands
- 2Department of Neonatology, Wilhelmina Children’s Hospital, University Medical Center Utrecht, Utrecht, Netherlands
- 3Department of Pharmacy, Academic Medical Center, Amsterdam, Netherlands
- 4Laboratory of NeuroImmunology and Developmental Origins of Disease (NIDOD), University Medical Center Utrecht, Utrecht, Netherlands
- 5Department of Neurology, Leiden University Medical Center, Leiden, Netherlands
- 6Brain Center Rudolf Magnus, University Medical Center Utrecht, Utrecht, Netherlands
- 7Department of Pharmacoepidemiology and Clinical Pharmacology, Faculty of Science, Utrecht University, Utrecht, Netherlands
Background: Hypoxic–ischemic encephalopathy following perinatal asphyxia is a leading cause of neonatal death and disability worldwide. Treatment with therapeutic hypothermia reduced adverse outcomes from 60 to 45%. Additional strategies are urgently needed to further improve the outcome for these neonates. Inhibition of nitric oxide synthase (NOS) is a potential neuroprotective target. This article reviews the evidence of neuroprotection by nitric oxide (NO) synthesis inhibition in animal models.
Methods: Literature search using the EMBASE, Medline, Cochrane, and PubMed databases. Studies comparing NOS inhibition to placebo, with neuroprotective outcome measures, in relevant animal models were included. Methodologic quality of the included studies was assessed.
Results: 26 studies were included using non-selective or selective NOS inhibition in rat, piglet, sheep, or rabbit animal models. A large variety in outcome measures was reported. Outcome measures were grouped as histological, biological, or neurobehavioral. Both non-selective and selective inhibitors show neuroprotective properties in one or more outcome measures. Methodologic quality was either low or moderate for all studies.
Conclusion: Inhibition of NO synthesis is a promising strategy for additional neuroprotection. In humans, intervention can only take place after the onset of the hypoxic–ischemic event. Therefore, combined inhibition of neuronal and inducible NOS seems the most likely candidate for human clinical trials. Future studies should determine its safety and effectiveness in neonates, as well as a potential sex-specific neuroprotective effect. Researchers should strive to improve methodologic quality of animal intervention studies by using a systematic approach in conducting and reporting of these studies.
Introduction
Hypoxic–ischemic encephalopathy (HIE) following perinatal asphyxia (i.e., severe oxygen deprivation at birth) is one of the leading causes of neonatal death and adverse neuromotor outcome in term and near-term infants worldwide. In high-income countries, the incidence of HIE has been estimated between 0.5 and 1.0 for every thousand live births, although some sources have reported an incidence as high as 8 per 1,000 live births (1, 2). In low- and middle-income countries, the incidence of HIE is higher, affecting more than 1.1 million babies annually (3–5).
The overall burden of HIE is high, in terms of quality-adjusted life years, years of life lost, and years lived with disability, not to mention a great financial cost for both society and the families involved (6, 7). With an estimated annual one million deaths worldwide, HIE is accountable for roughly 25% of all deaths in the neonatal period (3, 8).
Hypoxic–ischemic brain injury is not a single event, evoked by the actual asphyxia, but rather an ongoing process that leads to significant neuronal cell death over hours to days after the initial insult (9, 10). Several distinct phases have been identified in this process. The primary energy failure takes place during the hypoxic–ischemic event, resulting in failure of oxidative metabolism, cytotoxic edema, and accumulation of excitotoxins (11). After resuscitation and restoration of cerebral circulation, a latent phase, lasting approximately 6 h, commences (12, 13). Subsequently, starting between 6 and 15 h after asphyxia, the brain experiences a secondary energy failure that can last for days. This phase is marked by seizures, renewed cytotoxic edema, release of excitotoxins, impaired cerebral oxidative energy metabolism, and finally, neuronal cell death (14).
Currently, the only treatment that has proven to effectively reduce hypoxic–ischemic brain injury following perinatal asphyxia is the application of therapeutic hypothermia (TH). During TH the brain temperature is lowered to 33–34°C which is maintained for 72 h (1). Since the introduction of TH, the combined adverse outcome of death and disability, such as hearing loss, cerebral palsy, and other neuromotor disorders, has been reduced from approximately 60–45% (15–17). TH has widely been implemented as the standard of care treatment for moderate to severe HIE in high-income countries. However, TH needs to be started within 6 h after birth, leaving clinicians with a narrow window for establishing the diagnosis and severity of HIE as well as transportation to a medical facility equipped for TH (18). Additional neuroprotective strategies for HIE are urgently needed to augment TH, but when hypothermia is not yet feasible, act as a first line treatment option (3, 4, 19).
A potential target for (additional) neuroprotection in patients with HIE is the inhibition of nitric oxide synthase (NOS, enzyme commission number 1.14.13.39). NOS is an enzyme catalyzing production of nitric oxide (NO) from l-arginine. After perinatal asphyxia, NO can react with the superoxide free radical to form toxic peroxynitrite, setting a pre-apoptotic pathway in motion, resulting in neuronal loss (10, 20). Nitrotyrosine, an end product of this process, has been demonstrated post mortem in neonatal brain and spinal cord tissue after severe HIE (21, 22).
Three isoforms of NOS have been identified: endothelial (eNOS), neuronal (nNOS), and inducible NOS (iNOS) (23). All isoforms are upregulated after asphyxia; both nNOS and eNOS immediately after reperfusion and iNOS from several hours onward (24). While eNOS is regarded to be critical in maintaining pulmonary blood flow, preventing pulmonary hypertension and thereby maintaining adequate oxygenation of tissues throughout the body, excessive activation of nNOS and iNOS is associated with deleterious effects on the brain (24, 25). To illustrate, in mice genetically deficient of eNOS, infarct size after middle cerebral artery occlusion is larger compared with wild-type animals, due to a reduction in regional cerebral blood flow (26). By contrast, nNOS knockout mice are protected against hypoxic–ischemic brain injury, while mice lacking iNOS showed a delayed reduction in brain injury (27–32).
The aim of this study is to review the available evidence on NOS inhibition as a potential neuroprotective strategy in animal models translational for neonatal HIE and to identify one or more NOS inhibiting compounds that could evolve from preclinical to clinical studies in the near future.
Methods
Search Strategy
Studies assessing the neuroprotective effects of NOS inhibitors in HIE models were identified. A literature search using the EMBASE, Medline, Cochrane, and PubMed databases was performed. The primary keywords were Animals (newborn), Hypoxia, and Nitric Oxide Synthesis; the searches were limited to the English language. The complete search string is included in Supplementary Material. After the exclusion of duplicates, the titles and abstracts were independently screened by two researchers (Laurent M. A. Favié and Arlette R. Cox). A final selection was made after full text evaluation. Any discrepancies were resolved by a third researcher (Floris Groenendaal). In addition, the reference lists of the retrieved articles were searched for additional studies.
Selection Criteria
Studies were included based on the following inclusion criteria: animal models of a postnatal age in which brain development corresponds to near term or term brain development in humans, transient hypoxia or hypoxia–ischemia (HI), neuroprotection as outcome defined by histological, biochemical, and/or neurobehavioral parameters and inclusion of both a treatment group administering at least one NOS inhibitor and a control group that received sham treatment or consisted of untreated animals.
Data Synthesis
The year of publication, name of first author, the class and type of NOS inhibitor, the animal model, the method used to achieve HI, the dose and number of animals in each treatment group, the type of control group and number of control animals, the timing of administration with regards to the HI insult, and the results on the reported outcome parameters were recorded for each study. Each outcome parameter was categorized as histological, biochemical, or neurobehavioral.
Quality Assessment
The methodological quality of the included articles was assessed using the SYRCLE’s risk of bias (RoB) tool for animal intervention studies (33). This tool is based on the Cochrane RoB tool and consists of 10 items on which an article can be scored. Each item was scored 0, 1, or 2 points by two researchers (Laurent M. A. Favié and Arlette R. Cox) independently. If no evidence for adherence or evidence for non-adherence was found, a score of 0 was awarded. When evidence for adherence was present but inconclusive, one point was scored. If the item was fully adhered to, two points were scored. Any discrepancies were resolved after consultation with a third researcher (Agnes van den Hoogen). Because of the nature of the included studies and the timing of the interventions, “allocation concealment” was deemed unfeasible and was not rated for any of the articles. Articles scoring 1–6 points were considered low quality, 7–12 points moderate quality, and 13–18 points high quality. An example of the tool is included in Supplementary Material.
Results
Eligible Studies
The search yielded a total of 348 studies; 280 studies after removal of duplicates. After screening of title and abstract, 238 articles were excluded. Screening of the reference lists identified one additional article. 43 articles were thus assessed in full detail. Of these, 26 were deemed eligible for inclusion (Figure 1); the data were extracted from these studies, and these studies were assessed for methodological quality. Performing a meta-analysis was considered impossible because of the heterogeneity of the studies in outcome, administered NOS inhibitor, and animal models.
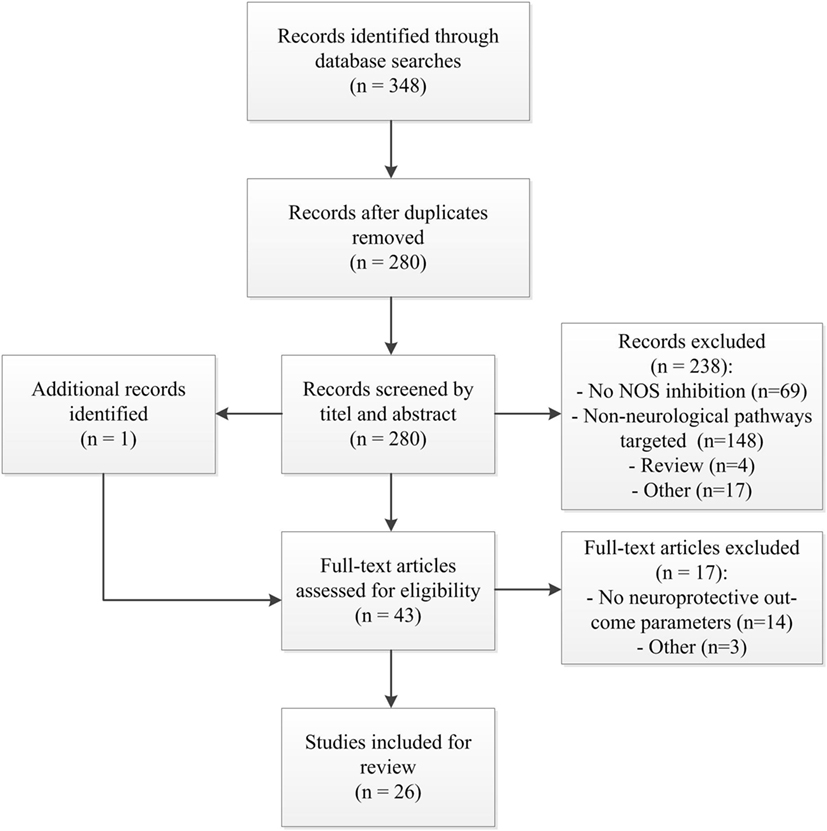
Figure 1. Study flow diagram. Abbreviations; n, number of studies; NOS, nitric oxide synthase.
Study Characteristics
The included studies and their descriptive characteristics are summarized in Table 1. Eight studies (31%) tested a non-specific NOS inhibitor (34–41), another eight (31%) applied an nNOS-specific inhibitor (42–49); three studies (12%) used an iNOS-specific inhibitor (50–52); and six (23%) used an inhibitor of both nNOS and iNOS (53–58). One study (3%) used separate groups for nNOS and iNOS inhibition (59). Four different species of animals were used: rat (n = 11, 42%), piglet (n = 10, 38%), sheep (n = 3, 12%), and rabbit (n = 2, 8%).

Table 1. Study characteristics including RoB score.
Different models for HI were used, mostly dependent on the animal species. All rat studies applied the Vannucci–Rice model in P7–P14 pups. All newborn (P1–P5) piglet studies induced brain injury by hypoxia for 30–60 min, in 30% of studies combined with transient bilateral artery occlusion. In sheep aged 2–11 days (one study), hypoxia for 30 min was combined with hypotension for 5 min. Also, two studies using sheep at 103–104 days gestation (term = 147 days) were included, in which brain injury was induced by hypoxia due to occlusion of the umbilical cord for 25 min. In rabbits, fetuses (embryonic day 22, 70% gestation) were subjected to an HI event by uterine ischemia for 40 min.
The dosing regimen of the included studies is summarized in Table 2. Seventeen studies (65%, all non-specific or nNOS-specific inhibitors) describe only a single administration, and nine studies (35%, all iNOS of combined nNOS and iNOS inhibitors) described repeated dosing. With regards to timing of the intervention, 12 studies (46%) administered the (first) dose before the onset of the HI event; 9 (35%) after the event; and the remaining 5 (19%) incorporated groups with administration both before and after the event.
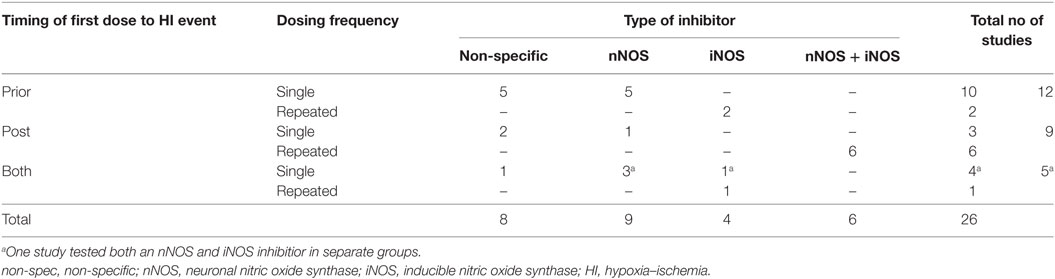
Table 2. Dosing frequency and timing of intervention for the included studies.
Outcome
The results of the reported outcome parameters for each study are presented in Table 1. A wide variety of histological, biochemical, and neurobehavioral outcome parameters were reported. Histological parameters included ipsilateral/contralateral weight ratio disparity and analysis of cortical and striatal lesions. Biochemical parameters included free radical formation and other biomarkers for neurological damage, but also cerebral energy status and electrocortical brain activity. Neurobehavioral parameters included overall survival, survival with normal EEG and results of neurobehavioral tests.
In the group of non-specific NOS inhibitors, administration before onset of the insult proved neuroprotective in 7/8 settings (88%), while administration directly after the insult was partially beneficial in 2/3 settings (67%).
For nNOS inhibitors, administration before the insult showed neuroprotective properties in 9/10 settings (90%) and when administered directly after the insult (1/1). When administration was delayed by 15 min or more, neuroprotective properties were lost (4/4).
When treatment with an iNOS inhibitor was started before the insult, neuroprotection was achieved (4/4). Administering the first dose after induction of HI showed neuroprotection in 33% of the settings (1/3). Hsu et al. administered the iNOS inhibitor aminoguanidine (AG) 30 min before and 3 h after the insult as a single dose. Both were neuroprotective compared with the control group, although less parameters were tested in the post insult treatment group.
All studies testing combined inhibition of nNOS and iNOS reported (partially) neuroprotective outcome. van den Tweel et al. (53) showed that 2-iminobiotin (2-IB) is neuroprotective in rats in a dose-dependent matter.
A direct comparison between two different inhibitors was made in two studies. Yu et al. reported superior neuroprotection of the novel nNOS inhibitor JI-8 compared with 7-nitro indazole (7-NI) when administered before the insult in equimolar doses. Hsu et al. observed that both 7-NI and AG are neuroprotective when administered 30 min before HI and that 7-NI is superior to AG in this setting. When the compounds were administered 3 h after HI, 7-NI lost its neuroprotective effect while AG remained neuroprotective compared with both vehicle and 7-NI.
Methodological Quality
Eleven studies (42%) were ranked low quality, 15 (58%) were considered moderate quality; none of the studies were ranked in the high quality group. On average, RoB score was 7 (3–12). Overall, animal baseline characteristics, randomization for treatment allocation, blinding of investigators and/or outcome assessors, and random selection for outcome assessment were often not mentioned and therefore scored 0.
Discussion
This systematic review shows that both selective and non-selective NOS inhibitors have neuroprotective qualities in various animal models of HI brain damage using histological, biochemical, and neurobehavioral outcome parameters. In animal studies, induction of the insult and administration of the potentially neuroprotective agent (before and/or after the insult) can be timed precisely. By contrast, this is not the case in clinical practice. The onset of perinatal asphyxia is often sudden and unpredictable. Therefore, administration of any drug before the onset of the insult is impossible, and administration directly after the insult (i.e., directly after birth) is highly improbable. All non-selective NOS inhibitors reviewed in this study were administered before insult or directly after; there are no data on delayed administration. Furthermore, non-selective inhibitors will also target eNOS, which could counteract the potential neuroprotective effects of nNOS and iNOS inhibition. For selective nNOS inhibitors, neuroprotection was lost when administration was withheld by as little as 15 min. For selective iNOS inhibitors, administration before the insult shows greater neuroprotective potential than post-insult treatment. The combination of nNOS and iNOS inhibition shows neuroprotective properties on histological, biochemical, and neurobehavioral outcome parameters when administered after the insult in a repeated dosing regimen. Thus, combined nNOS/iNOS inhibition with a repeated dosing regimen seems the most promising strategy to advance into human clinical trials. In fact, several phase II studies with 2-IB are currently underway, in addition to TH (NTR5221) as well as without TH in low-income countries (NCT01626924, EudraCT2015-003063-12).
Because of the wide variety in reported outcome measures, a clear-cut comparison between inhibitors based on outcome was difficult to make. Twelve studies report no neuroprotection on one or more outcome parameters after NOS inhibition. Potentially, this can be attributed to timing of the intervention or suboptimal dosing. When a NOS inhibitor is administered before the insult, the compound will be present in the tissues and circulation at the time of the actual insult, increasing the compound’s potential to exhibit neuroprotective effects. Most studies have tested one NOS inhibitor in a single dose. In studies testing different dosages, a higher dose often shows a better neuroprotective outcome, although some studies indicate a U-shaped effect. For 2-IB, the optimal dose in rats appears to be 30 mg/kg intraperitoneal (53). In piglets, increasing the dose by five times to 1.0 mg/kg intravenous does not provide greater neuroprotective properties compared with 0.2 mg/kg (57). Although most studies measured histological and biochemical outcome parameters associated with neuroprotection, the clinically most relevant parameter of improved neurobehavioral outcome was reported in four studies only. Yu et al. (45) and Ji et al. (49) showed that nNOS inhibition administered before the insult resulted in less deaths, and less neurobehavioral abnormalities in rabbits. Nijboer et al. (54) and Bjorkman et al. (57) report a (partial) neuroprotective effect for 2-IB on neurobehavioral outcome parameters in rats and piglets, respectively. Assessing neurobehavioral outcome requires a longer follow up period, which often involves intensive hands-on trained personnel especially in larger animal models, as well as validated tools to score the desired outcome parameter, making it very expensive. Using histological and biochemical markers provides researchers with a more time- and cost-effective alternative. Although data are limited, results on neurobehavioral outcome parameters, combined with results from histological and biochemical parameters, identify NOS inhibition as a potential neuroprotective strategy in humans.
Important differences exist between the adult and the neonatal brain with regard to susceptibility to injury, plasticity and cell death pathways. Therefore, adult animal models are not suitable to examine neuroprotective interventions for HIE. Across species, key brain maturation events regarding susceptibility and regenerative capacities have been identified at different moments before and after birth and are related to the developmental stage of the human neonatal brain (60–62). It is generally accepted that rats, at postnatal days 7–14 (P7–14), are comparable to near term/term human neonates with regards to cerebral cortex development (63, 64). The Vannucci–Rice model of unilateral common carotid artery ligation followed by a period of systemic hypoxia results in apoptotic-necrotic cell degeneration in P7–14 rats, similar to HIE (64–67). In term piglets aged 1–5 days, hypoxia leads to basal ganglia and somatosensory cortical injury, largely comparable to damage seen in human neonates after perinatal asphyxia (64, 68, 69). Introducing HI in utero to fetal rabbits provides animals with a motor phenotype similar to human cerebral palsy (64, 70). In term and preterm sheep models, hypoxia and asphyxia cause abnormalities in cerebral oxygen metabolism and hemodynamics as well as electrocortical brain activity comparable to human neonates after HI and basal ganglia injury representative for cerebral palsy (71–73).
Of interest is the potential role of sex-specific cell death pathways involved in HIE and possible sex-specific neuroprotective therapies. In general, females seem to be less susceptible to brain injury. This effect is seen across species, age groups, and origin of injury (74). In adult animal models, reduction in ischemic injury in females has been attributed to estradiol levels (74). Although estradiol will not be as predominant in prepubertal animal models, there is evidence of sexual dimorphism regarding sex steroids in central nervous system development in mice and rats (75, 76). Other studies show sex-specific cell death pathways leading to brain injury after HI both in vitro and in vivo. For instance, there is evidence that brain injury after HI in males is evoked by caspase-independent pathways whereas in females, caspase-dependent pathways are responsible (77–82). Therefore, neuroprotective agents such as NOS inhibitors that interact, either upstream or downstream, with the caspase-dependent pathway may be effective in females only.
The role of sex was only sparsely investigated in the studies included. For the majority of the studies (65%), the sex of the animals used was not reported. Six studies (23%) used rats of both sexes but have not reported sex-specific outcome. Yu et al. reported no outcome differences between sex for 7-NI and JI-9 but this statement was not supported by statistical analysis, possibly due to the small sample size in each of the groups (45). Nijboer et al. showed a statistically significant difference in histological and biochemical outcome parameters between sexes in rats, concluding that 2-IB was neuroprotective in female rats only (54). Other studies with different neuroprotective agents in both animals and humans also indicate a (potential) neuroprotective effect in females only (81–84).
Methodological quality assessment using the SYRCLE’s RoB tool resulted in only low and moderate scores for the publications used in this study. In all of the studies, at least on one or more items no information was available, forcing a score of 0 in that area. It is unknown whether these items were not adhered to during the experiment, or simply not included in the final manuscript due to regulations imposed by the editorial guidelines of the publishing journal. Unfortunately, it is not yet common practice to be as complete and precise in reporting data for animal studies as it is for human studies (33, 85). However, since this problem was addressed in a commentary published in the Lancet in 2002, awareness has been steadily increasing (86, 87). Fourteen of the studies included in this review were published in or before 2002; seven (50%) scoring low and an equal number scoring moderate. For the 12 included studies published in 2003 or later, 8 (67%) were awarded a moderate score. The SYRCLE’s RoB tool proved to be an adequate tool to consistently score the methodological quality of the included studies. However, this tool was developed recently and experience is still sparse. We would like to encourage future researchers to adhere to the items listed in this tool when conducting and reporting animal intervention studies to improve the methodological quality of studies as well as to use this tool when attempting a systematic review of animal literature. To illustrate the need for improvement in methodological quality and because of the possibility that low scores reflect lack of reporting and not lack of quality in the design of the study, we decided not to omit low quality studies nor did we emphasize the RoB scores when comparing the NOS inhibitors discussed in this study.
An important limitation of this study is that no independent statistics could be applied due to the large heterogeneity in study designs. Ideally, all NOS inhibitors should be tested in identical animal models with identical outcome measures. In reality, researchers over the past decades have used various animal models, dose and timing of NOS inhibitors, and reported outcome parameters. For the purpose of this review, we choose to report all of these and base our conclusions on the best available evidence. Based on this heterogeneity, these conclusions should be interpreted with caution.
Despite the low to moderate methodological quality according to the RoB tool, presented in Supplementary Material, and the lack of independent statistics, the evidence presented in this systematic review still indicates NOS inhibition as a promising strategy for (additional) neuroprotection in human neonates after perinatal asphyxia. Combined inhibition of nNOS and iNOS started as soon as possible after birth and in a repeated dosing regimen seems to have the best potential based on the combined outcome parameters, translation to clinical practice and methodological quality. Human studies (phase 2, open-label) with 2-IB, an inhibitor of both nNOS and iNOS, are currently taking place. Future clinical studies should make clear whether the sex-specific neuroprotective effect of drugs such as 2-IB observed in rats is present in humans as well. Furthermore, well designed placebo-controlled studies are needed to determine the safety of 2-IB in neonates and its effectiveness both with and without TH.
Author Contributions
LF, AC, and FG were involved in study selection; LF, AC, and AH conducted the methodological quality assessment. All the authors discussed the results and read and approved the final version of the manuscript. LF drafted the manuscript; AC, AH, CN, CP-S, FB, TE, CR, and FG provided critical feedback to each draft.
Conflict of Interest Statement
FB, FG, and CP-S are inventors of 2-iminobiotin as neuroprotective agent for neonates with HIE. CP-S is consultant for and shareholder of Neurophyxia BV’s-Hertogenbosch, The Netherlands. The other authors report no potential conflict of interest.
Acknowledgments
The authors would like to thank Justin van der Swaluw for his preliminary work in defining the search strategy.
Supplementary Material
The Supplementary Material for this article can be found online at https://www.frontiersin.org/articles/10.3389/fneur.2018.00258/full#supplementary-material.
References
1. Jacobs SE, Berg M, Hunt R, Tarnow-Mordi WO, Inder TE, Davis PG. Cooling for newborns with hypoxic ischaemic encephalopathy. In: Jacobs SE, editor. Cochrane Database of Systematic Reviews. Chichester, UK: John Wiley & Sons Ltd (2013). 385 p.
2. Arnaez J, García-Alix A, Arca G, Caserío S, Valverde E, Moral MT, et al. Population-based study of the national implementation of therapeutic hypothermia in infants with hypoxic-ischemic encephalopathy. Ther Hypothermia Temp Manag (2017) 8(1):24–9. doi:10.1089/ther.2017.0024
3. Pauliah SS, Shankaran S, Wade A, Cady EB, Thayyil S. Therapeutic hypothermia for neonatal encephalopathy in low- and middle-income countries: a systematic review and meta-analysis. PLoS One (2013) 8:e58834. doi:10.1371/journal.pone.0058834
4. Montaldo P, Pauliah SS, Lally PJ, Olson L, Thayyil S. Cooling in a low-resource environment: lost in translation. Semin Fetal Neonatal Med (2015) 20:72–9. doi:10.1016/j.siny.2014.10.004
5. Lee ACC, Kozuki N, Blencowe H, Vos T, Bahalim A, Darmstadt GL, et al. Intrapartum-related neonatal encephalopathy incidence and impairment at regional and global levels for 2010 with trends from 1990. Pediatr Res (2013) 74:50–72. doi:10.1038/pr.2013.206
6. Eunson P. The long-term health, social, and financial burden of hypoxic-ischaemic encephalopathy. Dev Med Child Neurol (2015) 57:48–50. doi:10.1111/dmcn.12727
7. Blencowe H, Vos T, Lee AC, Philips R, Lozano R, Alvarado MR, et al. Estimates of neonatal morbidities and disabilities at regional and global levels for 2010: introduction, methods overview, and relevant findings from the Global Burden of Disease study. Pediatr Res (2013) 74:4–16. doi:10.1038/pr.2013.203
8. Lawn JE, Cousens S, Zupan J. Neonatal survival 1 4 million neonatal deaths: when? Where? Why? Lancet (2005) 365(9462):891–900. doi:10.1016/S0140-6736(05)71048-5
9. Gunn AJ, Thoresen M. Hypothermic neuroprotection. NeuroRx (2006) 3:154–69. doi:10.1016/j.nurx.2006.01.007
10. van Bel FV, Groenendaal F. Drugs for neuroprotection after birth asphyxia: pharmacologic adjuncts to hypothermia. Semin Perinatol (2016) 40:152–9. doi:10.1053/j.semperi.2015.12.003
11. Wassink G, Gunn ER, Drury PP, Bennet L, Gunn AJ. The mechanisms and treatment of asphyxial encephalopathy. Front Neurosci (2014) 8:40. doi:10.3389/fnins.2014.00040
12. Iwata O, Iwata S, Bainbridge A, De Vita E, Matsuishi T, Cady EB, et al. Supra- and sub-baseline phosphocreatine recovery in developing brain after transient hypoxia-ischaemia: relation to baseline energetics, insult severity and outcome. Brain (2008) 131:2220–6. doi:10.1093/brain/awn150
13. Azzopardi D, Wyatt JS, Cady EB, Delpy DT, Baudin J, Stewart AL, et al. Prognosis of newborn infants with hypoxic-ischemic brain injury assessed by phosphorus magnetic resonance spectroscopy. Pediatr Res (1989) 25:445–51. doi:10.1203/00006450-198905000-00004
14. Gunn AJ, Laptook AR, Robertson NJ, Barks JD, Thoresen M, Wassink G, et al. Therapeutic hypothermia translates from ancient history in to practice. Pediatr Res (2017) 81:202–9. doi:10.1038/pr.2016.198
15. Azzopardi D, Brocklehurst P, Edwards D, Halliday H, Levene M, Thoresen M, et al. The TOBY Study. Whole body hypothermia for the treatment of perinatal asphyxial encephalopathy: a randomised controlled trial. BMC Pediatr (2008) 8:17. doi:10.1186/1471-2431-8-17
16. Azzopardi DV, Strohm B, Edwards AD, Dyet L, Halliday HL, Juszczak E, et al. Moderate hypothermia to treat perinatal asphyxial encephalopathy. N Engl J Med (2009) 361:1349–58. doi:10.1056/NEJMoa0900854
17. Groenendaal F, Casaer A, Dijkman KP, Gavilanes AWD, De Haan TR, Ter Horst HJ, et al. Introduction of hypothermia for neonates with perinatal asphyxia in the Netherlands and Flanders and the Dutch-Flemish working group on neonatal neurology. Neonatology (2013) 104:15–21. doi:10.1159/000348823
18. Olsen SL, DeJonge M, Kline A, Liptsen E, Song D, Anderson B, et al. Optimizing therapeutic hypothermia for neonatal encephalopathy. Pediatrics (2013) 131:e591–603. doi:10.1542/peds.2012-0891
19. Robertson N, Nakakeeto M, Hagmann C, Cowan F, Acolet D, Iwata O, et al. Hypothermia for birth asphyxia in low-resource settings: a pilot randomised controlled trial. Lancet (2008) 372:801–3. doi:10.1016/S0140-6736(08)61329-X
20. Beckman J, Koppenol W. Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am J Physiol (1996) 271:C1424–37. doi:10.1146/annurev.arplant.50.1.277
21. Groenendaal F, Lammers H, Smit D, Nikkels PGJ. Nitrotyrosine in brain tissue of neonates after perinatal asphyxia. Arch Dis Child Fetal Neonatal Ed (2006) 91:F429–33. doi:10.1136/adc.2005.092114
22. Groenendaal F, Vles J, Lammers H, De Vente J, Smit D, Nikkels PGJ. Nitrotyrosine in human neonatal spinal cord after perinatal asphyxia. Neonatology (2007) 93:1–6. doi:10.1159/000106432
23. Förstermann U, Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J (2012) 33:829–37. doi:10.1093/eurheartj/ehr304
24. Liu H, Li J, Zhao F, Wang H, Qu Y, Mu D. Nitric oxide synthase in hypoxic or ischemic brain injury. Rev Neurosci (2015) 26:105–17. doi:10.1515/revneuro-2014-0041
25. Fan X, Kavelaars A, Heijnen CJ, Groenendaal F, van Bel F. Pharmacological neuroprotection after perinatal hypoxic-ischemic brain injury. Curr Neuropharmacol (2010) 8:324–34. doi:10.2174/157015910793358150
26. Huang Z, Huang PL, Ma J, Meng W, Ayata C, Fishman MC, et al. Enlarged infarcts in endothelial nitric oxide synthase knockout mice are attenuated by nitro-L-arginine. J Cereb Blood Flow Metab (1996) 16:981–7. doi:10.1097/00004647-199609000-00023
27. Dawson VL, Kizushi VM, Huang PL, Snyder SH, Dawson TM. Resistance to neurotoxicity in cortical cultures from neuronal nitric oxide synthase-deficient mice. J Neurosci (1996) 16:2479–87.
28. Ferriero DM, Holtzman DM, Black SM, Sheldon RA. Neonatal mice lacking neuronal nitric oxide synthase are less vulnerable to hypoxic-ischemic injury. Neurobiol Dis (1996) 3:64–71. doi:10.1006/nbdi.1996.0006
29. Ferriero DM, Sheldon RA, Black SM, Chuai J. Selective destruction of nitric oxide synthase neurons with quisqualate reduces damage after hypoxia-ischemia in the neonatal rat. Pediatr Res (1995) 38:912–8. doi:10.1203/00006450-199512000-00014
30. Hara H, Huang PL, Panahian N, Fishman MC, Moskowitz MA. Reduced brain edema and infarction volume in mice lacking the neuronal isoform of nitric oxide synthase after transient MCA occlusion. J Cereb Blood Flow Metab (1996) 16:605–11. doi:10.1097/00004647-199607000-00010
31. Huang Z, Huang PL, Panahian N, Dalkara T, Fishman MC, Moskowitz MA. Effects of cerebral ischemia in mice deficient in neuronal nitric oxide synthase. Science (1994) 265:1883–5. doi:10.1126/science.7522345
32. Panahian N, Yoshida T, Huang PL, Hedley-Whyte ET, Dalkara T, Fishman MC, et al. Attenuated hippocampal damage after global cerebral ischemia in mice mutant in neuronal nitric oxide synthase. Neuroscience (1996) 72:343–54. doi:10.1016/0306-4522(95)00563-3
33. Hooijmans CR, Rovers MM, De Vries RBM, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE’s risk of bias tool for animal studies. BMC Med Res Methodol (2014) 14:43. doi:10.1186/1471-2288-14-43
34. Trifiletti R. Neuroprotective effects of N(G)-nitro-L-arginine in focal stroke in the 7-day old rat. Eur J Pharmacol (1992) 218:197–8. doi:10.1016/0014-2999(92)90168-4
35. Hamada Y, Hayakawa T, Hattori H, Mikawa H. Inhibitor of nitric oxide synthesis reduces hypoxic-ischemic brain damage in the neonatal rat. Pediatr Res (1994) 35:10–4. doi:10.1203/00006450-199401000-00003
36. Nunagami Y, Zubrow AB, Mishra OP, Delivoria-Papadopoulos M. Lipid free radical generation and brain cell membrane alteration following nitric oxide synthase inhibition during cerebral hypoxia in the newborn piglet. J Neurochem (1997) 69:1542–7. doi:10.1046/j.1471-4159.1997.69041542.x
37. Groenendaal F, de Graaf RA, van Vliet G, Nicolay K. Effects of hypoxia-ischemia and inhibition of nitric oxide synthase on cerebral energy metabolism in newborn piglets. Pediatr Res (1999) 45:827–33. doi:10.1203/00006450-199906000-00008
38. Ashraf QM, Zubrow AB, Mishra OP, Delivoria-Papadopoulos M. Nitration of Bax and Bcl-2 proteins during hypoxia in cerebral cortex of newborn piglets and the effect of nitric oxide synthase inhibition. Biol Neonate (2002) 81:65–72. doi:10.1159/000047186
39. Zubrow AB, Delivoria-Papadopoulos M, Ashraf QM, Fritz KI, Mishra OP. Nitric oxide-mediated Ca2+/calmodulin-dependent protein kinase IV activity during hypoxia in neuronal nuclei from newborn piglets. Neurosci Lett (2002) 335:5–8. doi:10.1016/S0304-3940(02)01138-2
40. Dorrepaal CA, van Bel F, Moison RM, Shadid M, van de Bor M, Steendijk P, et al. Oxidative stress during post-hypoxic-ischemic reperfusion in the newborn lamb: the effect of nitric oxide synthesis inhibition. Pediatr Res (1997) 41:321–6. doi:10.1203/00006450-199703000-00003
41. Blumberg RM, Taylor DL, Yue X, Aguan K, Mckenzie J, Cady EB, et al. Increased nitric oxide synthesis is not involved in delayed cerebral energy failure following focal hypoxic-ischemic injury to the developing brain. Pediatr Res (1999) 46:224–31. doi:10.1203/00006450-199908000-00016
42. Ishida A, Trescher WH, Lange MS, Johnston MV. Prolonged suppression of brain nitric oxide synthase activity by 7-nitroindazole protects against cerebral hypoxic-ischemic injury in neonatal rat. Brain Dev (2001) 23:349–54. doi:10.1016/S0387-7604(01)00237-6
43. Parikh NA, Katsetos CD, Ashraf QM, Haider SH, Legido A, Delivoria-Papadopoulos M, et al. Hypoxia-induced caspase-3 activation and DNA fragmentation in cortical neurons of newborn piglets: role of nitric oxide. Neurochem Res (2003) 28:1351–7. doi:10.1023/A:1024992214886
44. Ashraf QM, Haider SH, Katsetos CD, Delivoria-Papadopoulos M, Mishra O. Nitric oxide-mediated alterations of protein tyrosine phosphatase activity and expression during hypoxia in the cerebral cortex of newborn piglets. Neurosci Lett (2004) 362:108–12. doi:10.1016/j.neulet.2004.02.069
45. Yu L, Derrick M, Ji H, Silverman RB, Whitsett J, Vasquez-Vivar J, et al. Neuronal nitric oxide synthase inhibition prevents cerebral palsy following hypoxia-ischemia in fetal rabbits: comparison between JI-8 and 7-nitroindazole. Dev Neurosci (2011) 33:312–9. doi:10.1159/000327244
46. Mishra OP, Zubrow AB, Ashraf QM, Delivoria-Papadopoulos M. Effect of nitric oxide synthase inhibition during post-hypoxic reoxygenation on Bax and Bcl-2 protein expression and DNA fragmentation in neuronal nuclei of newborn piglets. Brain Res (2006) 1101:20–8. doi:10.1016/j.brainres.2006.05.021
47. Drury PP, Davidson JO, van den Heuij LG, Tan S, Silverman RB, Ji H, et al. Partial neuroprotection by nNOS inhibition during profound asphyxia in preterm fetal sheep. Exp Neurol (2013) 250:282–92. doi:10.1016/j.expneurol.2013.10.003
48. Drury PP, Davidson JO, Mathai S, van den Heuij LG, Ji H, Bennet L, et al. NNOS inhibition during profound asphyxia reduces seizure burden and improves survival of striatal phenotypic neurons in preterm fetal sheep. Neuropharmacology (2014) 83:62–70. doi:10.1016/j.neuropharm.2014.03.017
49. Ji H, Tan S, Igarashi J, Li H, Derrick M, Martasek P, et al. Selective neuronal nitric oxide synthase inhibitors and the prevention of cerebral palsy. Ann Neurol (2009) 65:209–17. doi:10.1002/ana.21555
50. Ikeno S, Nagata N, Yoshida S, Takahashi H, Kigawa J, Terakawa N. Immature brain injury via peroxynitrite production induced by inducible nitric oxide synthase after hypoxia-ischemia in rats. J Obstet Gynaecol Res (2000) 26:227–34. doi:10.1111/j.1447-0756.2000.tb01316.x
51. Tsuji M, Higuchi Y, Shiraishi K, Kume T, Akaike A, Hattori H. Protective effect of aminoguanidine on hypoxic-ischemic brain damage and temporal profile of brain nitric oxide in neonatal rat. Pediatr Res (2000) 47:79. doi:10.1203/00006450-200001000-00015
52. Tutak E, Satar M, Zorludemir S, Erdogan S, Yapicioglu H, Narli N, et al. Neuroprotective effects of indomethacin and aminoguanidine in the newborn rats with hypoxic-ischemic cerebral injury. Neurochem Res (2005) 30:937–42. doi:10.1007/s11064-005-5978-5
53. van den Tweel ER, van Bel F, Kavelaars A, Peeters-Scholte CM, Haumann J, Nijboer CH, et al. Long-term neuroprotection with 2-iminobiotin, an inhibitor of neuronal and inducible nitric oxide synthase, after cerebral hypoxia-ischemia in neonatal rats. J Cereb Blood Flow Metab (2005) 25:67–74. doi:10.1038/sj.jcbfm.9600007
54. Nijboer CHA, Groenendaal F, Kavelaars A, Hagberg HH, van Bel F, Heijnen CJ. Gender-specific neuroprotection by 2-iminobiotin after hypoxia-ischemia in the neonatal rat via a nitric oxide independent pathway. J Cereb Blood Flow Metab (2007) 27:282–92. doi:10.1038/sj.jcbfm.9600342
55. Peeters-Scholte C, Koster J, Veldhuis W, van den Tweel E, Zhu C, Kops N, et al. Neuroprotection by selective nitric oxide synthase inhibition at 24 hours after perinatal hypoxia-ischemia. Stroke (2002) 33:2304–10. doi:10.1161/01.STR.0000028343.25901.09
56. Peeters-Scholte C, Koster J, van den Tweel E, Blomgren K, Hamers N, Zhu C, et al. Effects of selective nitric oxide synthase inhibition on IGF-1, caspases and cytokines in a newborn piglet model of perinatal hypoxia-ischaemia. Dev Neurosci (2002) 24:396–404. doi:10.1159/000069045
57. Bjorkman ST, Ireland Z, Fan X, van der Wal WM, Roes KCB, Colditz PB, et al. Short-term dose-response characteristics of 2-iminobiotin immediately postinsult in the neonatal piglet after hypoxia-ischemia. Stroke (2013) 44:809–11. doi:10.1161/STROKEAHA.112.677922
58. Van den Tweel ERW, Peeters-Scholte CMPCD, van Bel F, Heijnen CJ, Groenendaal F. Inhibition of nNOS and iNOS following hypoxia-ischaemia improves long-term outcome but does not influence the inflammatory response in the neonatal rat brain. Dev Neurosci (2002) 24:389–95. doi:10.1159/000069044
59. Hsu Y-C, Chang Y-C, Lin Y-C, Sze C-I, Huang C-C, Ho C-J. Cerebral microvascular damage occurs early after hypoxia-ischemia via nNOS activation in the neonatal brain. J Cereb Blood Flow Metab (2014) 34:668–76. doi:10.1038/jcbfm.2013.244
60. Clancy B, Finlay BL, Darlington RB, Anand KJS. Extrapolating brain development from experimental species to humans. Neurotoxicology (2007) 28:931–7. doi:10.1016/j.neuro.2007.01.014
61. Dobbing J, Sands J. Developmental events. Growth (Lakeland) (1979) 2:503–5. doi:10.1016/0378-3782(79)90022-7
62. Pressler R, Auvin S. Comparison of brain maturation among species: an example in translational research suggesting the possible use of bumetanide in newborn. Front Neurol (2013) 4:36. doi:10.3389/fneur.2013.00036
63. Romijn HJ, Hofman MJ, Gransbergen A. At what age is the developing cortex of the rat comparable to that of the full term newborn baby? Early HumDev (1991) 26:61–7. doi:10.1016/0378-3782(91)90044-4
64. Northington FJ, Graham EM, Martin LJ. Apoptosis in perinatal hypoxic–ischemic brain injury: how important is it and should it be inhibited? Brain Res Rev (2005) 50:244–57. doi:10.1016/j.brainresrev.2005.07.003
65. Rice JE, Vannucci RC, Brierley JB. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann Neurol (1981) 9:131–41. doi:10.1002/ana.410090206
66. Chavez-Valdez R, Martin LJ, Northington FJ. Programmed necrosis: a prominent mechanism of cell death following neonatal brain injury. Neurol Res Int (2012) 2012:257563. doi:10.1155/2012/257563
67. Northington FJ, Chavez-Valdez R, Martin LJ. Neuronal cell death in neonatal hypoxia-ischemia. Ann Neurol (2011) 69:743–58. doi:10.1002/ana.22419
68. Traystman RJ. Primary sensory and forebrain motor systems in the newborn brain are hypoxia-ischemia. J Comp Neurol (1997) 285:262–85.
69. Roland EH, Poskitt K, Rodriguez E, Lupton BA, Hill A. Perinatal hypoxic-ischemic thalamic injury: clinical features and neuroimaging. Ann Neurol (1998) 44:161–6. doi:10.1002/ana.410440205
70. Derrick M, Luo NL, Bregman JC, Jilling T, Ji X, Fisher K, et al. Preterm fetal hypoxia – ischemia causes hypertonia and motor deficits in the neonatal rabbit : a model for human cerebral palsy? J Neurosci (2004) 24:24–34. doi:10.1523/JNEUROSCI.2816-03.2004
71. Bax M, Tydeman C, Hons BA. Clinical and MRI correlates of cerebral palsy the European Cerebral Palsy Study. J Am Med Assoc (2006) 296:1602–8. doi:10.1001/jama.296.13.1602
72. Rosenberg AA. Cerebral blood flow and O2 metabolism after asphyxia in neonatal lambs. Pediatr Res (1986) 20:778–82. doi:10.1203/00006450-198608000-00016
73. Williams CE, Gunn AJ, Mallard C, Gluckman PD. Outcome after ischemia in the developing sheep brain: an electroencephalographic and histological study. Ann Neurol (1992) 31:14–21. doi:10.1002/ana.410310104
74. Hurn PD, Vannucci SJ, Hagberg H. Adult or perinatal brain injury: does sex matter? Stroke (2005) 36:193–5. doi:10.1161/01.STR.0000153064.41332.f6
75. Dewing P, Shi T, Horvath S, Vilain E. Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Brain Res Mol Brain Res (2003) 118:82–90. doi:10.1016/S0169-328X(03)00339-5
76. Becu-Villalobos D, Gonzalez Iglesias A, Diaz-Torga G, Hockl P, Libertun C. Brain sexual differentiation and gonadotropins secretion in the rat. Cell Mol Neurobiol (1997) 17:699–715. doi:10.1023/A:1022542221535
77. Du L, Bayir H, Lai Y, Zhang X, Kochanek PM, Watkins SC, et al. Innate gender-based proclivity in response to cytotoxicity and programmed cell death pathway. J Biol Chem (2004) 279:38563–70. doi:10.1074/jbc.M405461200
78. Hagberg H, Wilson MA, Matsushita H, Zhu C, Lange M, Gustavsson M, et al. PARP-1 gene disruption in mice preferentially protects males from perinatal brain injury. J Neurochem (2004) 90:1068–75. doi:10.1111/j.1471-4159.2004.02547.x
79. Zhu C, Xu F, Wang X, Shibata M, Uchiyama Y, Blomgren K, et al. Different apoptotic mechanisms are activated in male and female brains after neonatal hypoxia-ischaemia. J Neurochem (2006) 96:1016–27. doi:10.1111/j.1471-4159.2005.03639.x
80. Hill CA, Fitch RH. Sex differences in mechanisms and outcome of neonatal hypoxia-ischemia in rodent models: implications for sex-specific neuroprotection in clinical neonatal practice. Neurol Res Int (2012) 2012:867531. doi:10.1155/2012/867531
81. Joly L-M, Mucignat V, Mariani J, Plotkine M, Charriaut-Marlangue C. Caspase inhibition after neonatal ischemia in the rat brain. J Cereb Blood Flow Metab (2004) 24:124–31. doi:10.1097/01.WCB.0000100061.36077.5F
82. Renolleau S, Fau S, Goyenvalle C, Joly LM, Chauvier D, Jacotot E, et al. Specific caspase inhibitor Q-VD-OPh prevents neonatal stroke in P7 rat: a role for gender. J Neurochem (2007) 100:1062–71. doi:10.1111/j.1471-4159.2006.04269.x
83. Renolleau S, Fau S, Goyenvalle C, Charriaut-Marlangue C. Sex, neuroprotection, and neonatal ischemia. Dev Med Child Neurol (2007) 49:477. doi:10.1111/j.1469-8749.2007.00477.x
84. Kaandorp JJ, Benders MJNL, Schuit E, Rademaker CMA, Oudijk MA, Porath MM, et al. Maternal allopurinol administration during suspected fetal hypoxia: a novel neuroprotective intervention? A multicentre randomised placebo controlled trial. Arch Dis Child Fetal Neonatal Ed (2015) 100:F216–23. doi:10.1136/archdischild-2014-306769
85. Kilkenny C, Parsons N, Kadyszewski E, Festing MFW, Cuthill IC, Fry D, et al. Survey of the quality of experimental design, statistical analysis and reporting of research using animals. PLoS One (2009) 4:e7824. doi:10.1371/journal.pone.0007824
86. Sandercock P, Roberts I. Systematic reviews of animal experiments. Lancet (2002) 360:586. doi:10.1016/S0140-6736(02)09812-4
87. Hooijmans CR, Rovers M, de Vries RB, Leenaars M, Ritskes-Hoitinga M. An initiative to facilitate well-informed decision-making in laboratory animal research: report of the First International Symposium on Systematic Reviews in Laboratory Animal Science. Lab Anim (2012) 46:356–7. doi:10.1258/la.2012.012052
Keywords: nitric oxide synthase inhibition, neuroprotection, animal models, hypoxic–ischemic encephalopathy, 2-iminobiotin, review
Citation: Favié LMA, Cox AR, van den Hoogen A, Nijboer CHA, Peeters-Scholte CMPCD, van Bel F, Egberts TCG, Rademaker CMA and Groenendaal F (2018) Nitric Oxide Synthase Inhibition as a Neuroprotective Strategy Following Hypoxic–Ischemic Encephalopathy: Evidence From Animal Studies. Front. Neurol. 9:258. doi: 10.3389/fneur.2018.00258
Received: 27 December 2017; Accepted: 03 April 2018;
Published: 19 April 2018
Edited by:
Stephane Vladimir Sizonenko, Geneva University Hospitals (HUG), SwitzerlandReviewed by:
Akira Yoshii, University of Illinois at Chicago, United StatesSheffali Gulati, All India Institute of Medical Sciences, India
Copyright: © 2018 Favié, Cox, van den Hoogen, Nijboer, Peeters-Scholte, van Bel, Egberts, Rademaker and Groenendaal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laurent M. A. Favié, bC5tLmEuZmF2aWVAdW1jdXRyZWNodC5ubA==