 Anagh Pathak
Anagh Pathak Dipanjan Roy
Dipanjan Roy Arpan Banerjee
Arpan Banerjee
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Comput. Neurosci., 26 May 2022
Volume 16 - 2022 | https://doi.org/10.3389/fncom.2022.866517
This article is part of the Research TopicInsights in Computational NeuroscienceView all 11 articles
Computational neuroscience has come a long way from its humble origins in the pioneering work of Hodgkin and Huxley. Contemporary computational models of the brain span multiple spatiotemporal scales, from single neuronal compartments to models of social cognition. Each spatial scale comes with its own unique set of promises and challenges. Here, we review models of large-scale neural communication facilitated by white matter tracts, also known as whole-brain models (WBMs). Whole-brain approaches employ inputs from neuroimaging data and insights from graph theory and non-linear systems theory to model brain-wide dynamics. Over the years, WBM models have shown promise in providing predictive insights into various facets of neuropathologies such as Alzheimer's disease, Schizophrenia, Epilepsy, Traumatic brain injury, while also offering mechanistic insights into large-scale cortical communication. First, we briefly trace the history of WBMs, leading up to the state-of-the-art. We discuss various methodological considerations for implementing a whole-brain modeling pipeline, such as choice of node dynamics, model fitting and appropriate parcellations. We then demonstrate the applicability of WBMs toward understanding various neuropathologies. We conclude by discussing ways of augmenting the biological and clinical validity of whole-brain models.
Billions of years of evolution have invested the nervous system with tremendous complexity. Modern neuroscience has sought to understand this complexity as a hierarchical ladder that spans multiple spatial and temporal scales, starting from the interaction of biomolecules through to more complex structures like neurons and neural networks. Building on the pioneering work of Hodgkin and Huxley, significant progress took place toward the elucidation of the function of the single neuron. However, in spite of all the remarkable achievements at the single neuron level, relatively little is known about how populations of neurons coordinate with one another to facilitate cognitive processes. While it is fair to characterize neurons, or even individual dendrites as the canonical units of computation in the brain, it is evident that complex cognitive processes rely on interactions between several neural ensembles (McIntosh, 2004; Bressler and Tognoli, 2006)1 that are distributed across the cortex (Deco et al., 2008). Gaining an understanding of neural circuitry assumes vital importance not only for explaining cognition, but also for the treatment of various neurological diseases.
Early efforts toward bridging the gap between single-neuron activity and circuit operation led to the formulation of neural mass models (Beurle, 1956; Wilson and Cowan, 1972) which conceptualized cortical activity as arising from the dynamic interplay of multiple neural populations (or masses) with excitatory-inhibitory feedback (Figure 1). Such models leverage the fact that while the spiking of individual neurons is highly irregular (even chaotic), the mean activity of neural ensembles obeys fairly low-dimensional dynamics (Deco et al., 2008) 2. Furthermore, mean-field descriptions of cortical tissue may be extended in space and endowed with spatial gradients in neural parameters that follow mathematically defined connectivity (Amari, 1977). Field theory, with its deep roots in physics, provides analytically tractable solutions and has been employed extensively in neuroscience to explain wide-ranging phenomena. A classic field-theoretic model is the one proposed by Amari, which considered lateral-inhibition to explain oscillatory waves and input-evoked transients (Amari, 1977). Two-dimensional field models support diverse phenomena such as spiral and target waves that are organized into complex checkerboard patterns, reminiscent of neural activity observed during different brain states (Ermentrout and Cowan, 1979; Jirsa and Haken, 1996).
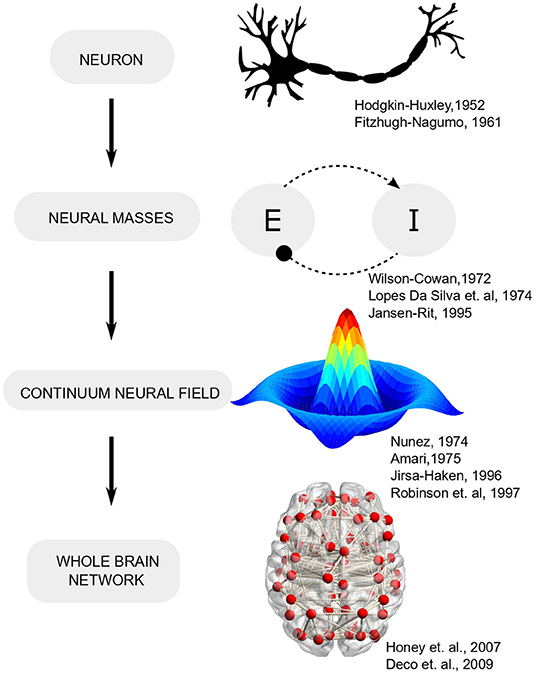
Figure 1. Evolution of computational neuroscience models from single neurons to network models.
By the turn of the century, neuroimaging modalities like PET and fMRI were being increasingly used to study cognition. The abstract nature of existing large-scale models made it difficult to exploit the rich datasets which were being churned in such experiments (Tagamets and Horwitz, 1998; Horwitz et al., 1999, 2000). Additionally, despite their success in providing theoretical accounts for neural phenomena such as traveling waves (Amari, 1977) or resting-state dynamics, Robinson et al. (2021) continuum field models had limited applicability in the clinical setting since crucial medical observables such as anatomical connectivity, functional correlations between brain areas or distribution of various cell types cannot be expressed in terms of mathematical expressions which can then be analytically solved within the field-theoretic framework. Therefore, it was deemed desirable to setup the neurodynamic model so that patient-specific neuroimaging data (e.g., DTI, fMRI connectivity) could be fused with simulations in order to facilitate precision medicine (Ritter et al., 2013; Deco and Kringelbach, 2014).
Within this framework, anatomical connectivity derived from diffusion MRI is used as a structural scaffold to simulate mesoscopic neural interactions (Horwitz et al., 2000; Honey et al., 2009). Nodes, representing mean-field activity of individual brain areas, evolve according to differential equations under the influence of coupling from other brain regions, external input and noise (Deco et al., 2009). Parameters representing biological or phenomenological properties of the nodes and edges are systematically varied, and time-series obtained for each run. For fMRI data, a further hemodynamic convolution is applied to the time-series and functional correlations (FC) are estimated from the resulting data (Deco and Kringelbach, 2014). Alternatively, for EEG/MEG studies, FC is estimated from amplitude envelopes that are extracted for each frequency band of interest and downsampled to correspond with BOLD time-scales (Hipp et al., 2012). Model fitting techniques are then utilized to obtain working points and regimes that best capture the corresponding empirical data. After model fitting, the researcher can ask how this system responds to various perturbations like external inputs (e.g., stimuli), noise or structural insults (e.g., lesions) (Figure 2).
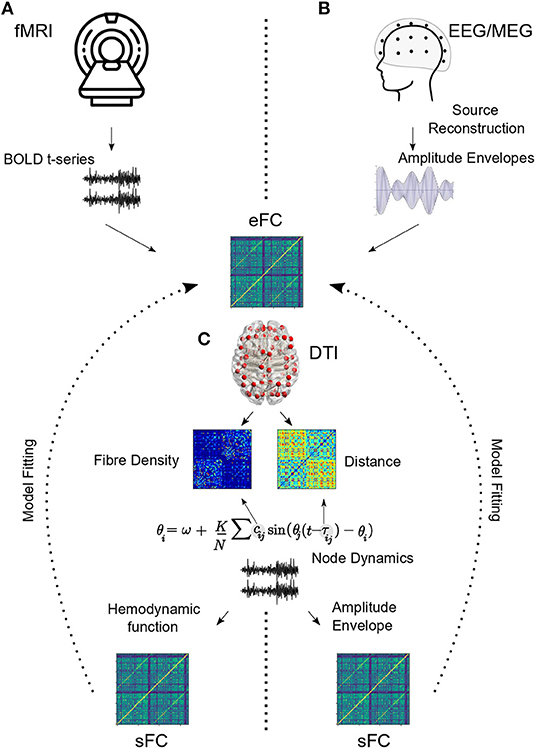
Figure 2. Model pipeline. (A) Inter-regional functional correlations (empirical functional connectivity or eFC) are extracted from resting state BOLD time-series. (B) For EEG/MEG, raw electrode/sensor time series are used to estimate source level activity, which is then bandpass filtered and hilbert transformed to obtain amplitude envelopes. Band specific functional connectivity is estimated from amplitude envelopes. DTI connectivity provides information about inter-regional white matter fiber density. Euclidean distances may be scaled to obtain temporal delay information. (C) Kuramoto model is used here to demonstrate how DTI information is incorporated into the node dynamics. For fMRI, an additional hemodynamic convolution is performed (Balloon-Windkessel) to project model output to BOLD time scales before estimating simulated FC (sFC). For EEG/MEG, amplitude envelopes are extracted and simulated FC is estimated. Model fitting procedures estimate model parameters based on eFC and sFC.
Whole-brain models provide actionable insights into various neurological deficits (e.g., identifying optimal resection zone in epilepsy), while also retaining a link to fundamental dynamical and graph theoretic concepts like attractors, metastability, stochastic dynamics, chaos and modularity (Popovych et al., 2019). In the following we outline the major variables that need to be considered before establishing a successful WBM pipeline.
Whole-brain modeling has tremendous clinical applicability as it provides prognostic tools and predictive insights for a host of neurological diseases (Deco and Kringelbach, 2014). However, since not all brain pathologies have the same origin or mechanism, the models seeking to understand them are also customized according to the specific etiology of the disease (Table 1). Following E.P Box's adage- “all models are wrong, but some are useful,” system equations are set up keeping in mind the specific properties of the underlying clinical context at the expense of biological realism. For example, it may be unnecessary to include conduction delays in models seeking to fit fMRI data due to the widely differing time-scales between BOLD activity (seconds) and axonal propagation (milliseconds). On the other hand, delays assume vital importance when the object of study involves electrophysiological spectral coherence between neural oscillators. Broadly, establishing an effective whole- brain modeling pipeline essentially comes down to the following choices- parcellation scheme, node dynamics, model fitting technique and type of perturbation applied.
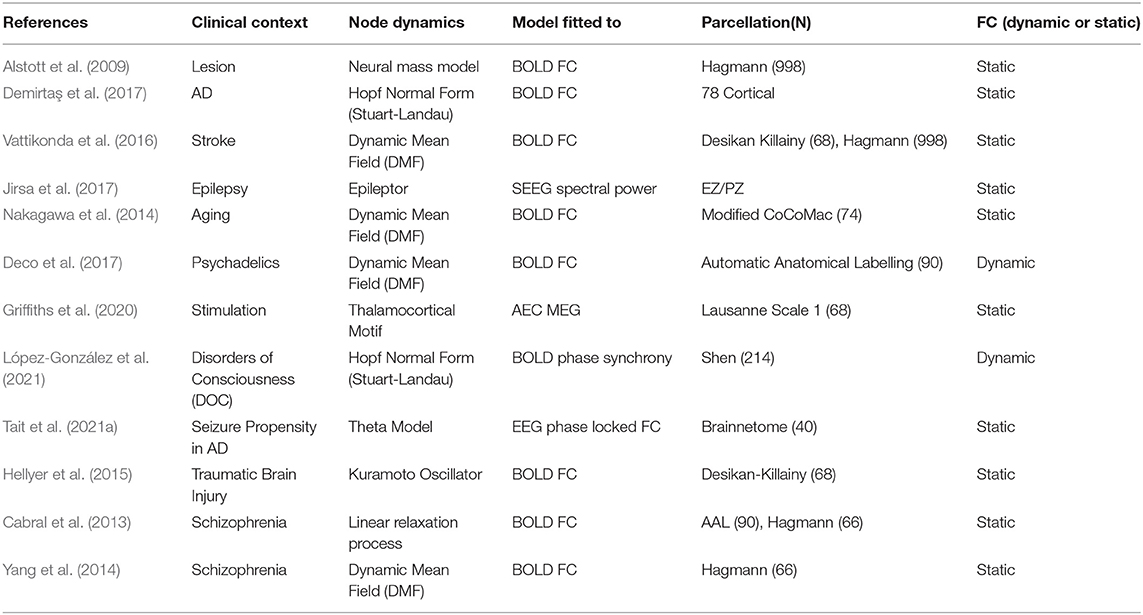
Table 1. List of studies employing WBMs to understand neuropathologies.
Firstly, thousands of voxels are reduced to only a few relevant areas of interest. Diffusion imaging is performed to extract anatomical connectivity matrices (Box 1). Connectivity matrices specify fiber density across various white matter tracts. A crucial decision at this stage is the choice of a parcellation scheme for obtaining an adjacency matrix. In the absence of a general consensus on what constitutes a “good” parcellation, one must consider carefully how the parcellation scheme may affect the WBM pipeline. Parcellation dictates the spatial resolution and topology of the model. Topological properties are known to be affected by the spatial scale of the parcellation used (Zalesky et al., 2010). Zalesky et al. (2010) demonstrate that while the basic properties of network topology such as scale-freeness or small-worldness remain invariant across spatial scales, the extent of these properties significantly varies between parcellations. Modeling has further corroborated that significant variability exists in graph-theoretical attributes of network dynamics as a function of the parcellation scheme (Domhof et al., 2021). Significant inter-parcellation variability also exists in resting-state dynamical models (Fornito et al., 2010). Additionally, since the computation time for whole-brain models scales with the number of coupled differential equations (same as the nodes in the network), finer-grained parcellations may be computationally cumbersome to solve. Further, diffusion MRI (dMRI) techniques are biased against short-range and intracortical connectivity, which may have significant ramifications for simulated dynamics (Proix et al., 2016). Highly granular parcellation schema may also lead to redundancy and rank-deficiency during source reconstruction (Tait et al., 2021b).
Box 1. Estimating anatomical connectivity
In-vivo estimation of white matter structural connectivity is enabled by diffusion magnetic resonance imaging (dMRI). Broadly, dMRI approaches measure the preferential direction of diffusion of water molecules in brain tissue. Computational algorithms estimate fiber orientations (streamlines) from dMRI data using a process known as tractography. Streamlines are counted and averaged according to pre-defined brain parcellations to yield adjacency graphs, which can then be submitted as input for whole-brain models. Computational libraries that perform tractography include FSL (Jenkinson et al., 2012), MRtrix (Tournier et al., 2012), BrainSUITE (Shattuck and Leahy, 2002), and DSI studio (Yeh et al., 2013). Considerable variability may exist in the output of different libraries due to differences in the choice of diffusion models, model parameters and tractographic algorithms. Thus, optimal algorithm selection remains an active area of research (Bastiani et al., 2012; Zhan et al., 2015; Petrov et al., 2017).
Broadly, atlases bin brain areas on the basis of either anatomical or functional similarities. Commonly used anatomical criteria for parcellating brain regions include gross anatomy, cytoarchitecture, myeloarchitecture, chemoarchitecture and gene expression profiles (Nowinski, 2021). By contrast, functional atlases utilize resting state or task-related functional correlations to allocate ROIs (Craddock et al., 2012; James et al., 2016). Anatomical atlases are known to fare poorly in comparison to functional atlases when it comes to reproducing FC patterns at the voxel scale (Craddock et al., 2012). Since functional homogeneity is a crucial precondition for modeling ROI dynamics, this would argue for the superiority of functional over anatomical parcellations for whole-brain modeling (Craddock et al., 2012). On the other hand, with anatomically defined ROIs it is easier to interpret results in the light of extant neuroscience literature. Therefore, multimodal atlases which integrate anatomical and functional criteria may offer a suitable tradeoff to ensure functional homogeneity while retaining anatomical specificity in whole-brain analysis (Glasser et al., 2016).
Ultimately, the scope of the study dictates the choice of parcellation. For example, it may be crucial to include sub-cortical nodes where the primary pathology may involve subcortical structures like the thalamus (Ji et al., 2016; Bazin et al., 2020).
Node dynamics consist of differential equations specifying the temporal evolution of the population activity of each region of interest (ROI). Each anatomically defined node may potentially consist of thousands of neurons and therefore, the dynamics of the ensemble is reduced to a low-dimensional description using mean-field formalisms. For example, Deco et al. reduce a spiking neuron model with synaptic conductance to yield a dynamic mean-field model that is subsequently used to specify node dynamics (Deco et al., 2013; Roy et al., 2014). Examples of node dynamics may range from the simple phenomenological ones, such as the Kuramoto model (Breakspear et al., 2010) or the normal form of the supercritical Hopf bifurcation (Lord et al., 2017) to the more biologically inspired ones such as the Wilson-Cowan model (Wilson and Cowan, 1972) or thalamocortical motifs (Griffiths et al., 2020) (see Figure 3 and Table 1). For example, the Kuramoto model reduces node dynamics to a phase variable, which evolves according to a natural frequency and a sinusoidal interaction term (Breakspear et al., 2010). On the other hand, both asynchronous and synchronous dynamics can be captured in the same set of equations in bifurcation models that can possess a relaxation solution (damped oscillations) or limit cycle solution (self-sustained oscillations) depending on the value of the bifurcation parameter (Figure 3) (Lord et al., 2017). Most computational studies model average functional connectivity, however, brain dynamics is also marked by transitions in the patterns of functional connectivity with time. Switching between FC configurations (FC state) requires node dynamics to possess multistable solutions which may be imparted through the addition of non-linear terms in model equations (Deco et al., 2013; Hansen et al., 2015).
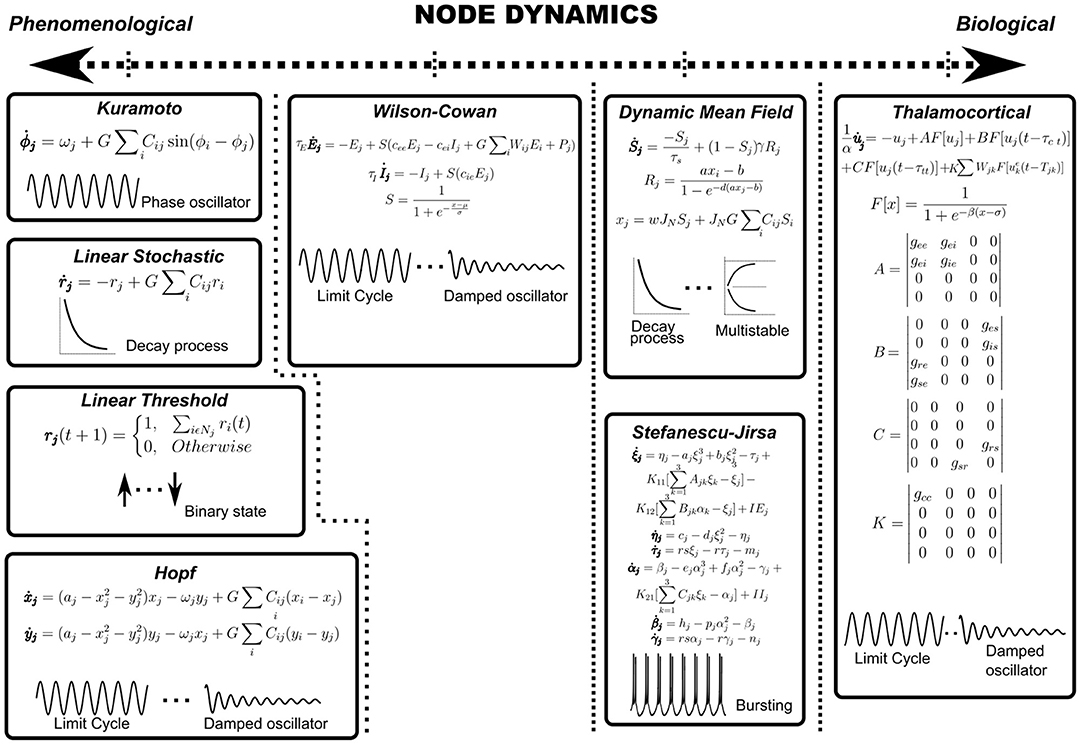
Figure 3. Node dynamics. Node dynamics may be categorized on the axis of biological validity. Purely phenomenological models such as Kuramoto, Linear Stochastic, Linear Threshold or the normal form of Hopf bifurcation may be regarded as toy models which seek to focus on a conceptual aspect of underlying phenomena at the expense of biological realism. On the other hand, it may be necessary to impart nodes with biologically detailed dynamics (SJ3D or Thalamocortical motifs) to explain a certain biological feature of the research problem. Further, nodes may be categorized on the type of underlying dynamics, such as damped oscillators, limit cycle oscillators, relaxation processes, multistable solutions or bursting patterns (represented schematically).
Heterogeneity in node dynamics may be introduced by assigning multiple oscillatory frequencies. For example, Deco et al. (2017) show that models utilizing multiple natural frequencies confirm better to empirical rsMEG networks. Similarly, Roberts et al. devise a principled approach for natural frequency allocation by scaling frequencies by the topological degree of each node (Gollo et al., 2017). For non-oscillatory node dynamics, temporal heterogeneity may be introduced by modulating exponential decay rates or synaptic time constants (Figure 3).
Another decision to be made at this step is the inclusion of transmission delays (Nakagawa et al., 2014). Computational models have demonstrated the value of including transmission delays, particularly in explaining oscillatory activity at electrophysiological time scales (Banerjee and Jirsa, 2007; Deco et al., 2009). Neural delays are known to play a crucial role in motor control, particularly in bimanual coordination (Banerjee and Jirsa, 2007) and in explaining perceptual variability in multi-sensory integration (Thakur et al., 2016). Conduction delays, on the order of a few milliseconds, can flip the phase relationship between two gamma oscillators from in-phase to out-of-phase (Pajevic et al., 2014). Network delays crucially dictate oscillation frequency (Niebur et al., 1991; Petkoski and Jirsa, 2019; Pathak et al., 2021) and propagation of cortical traveling waves (Ermentrout and Kleinfeld, 2001). Delays can even cause the complete cessation of self-sustained oscillations (amplitude death) in networks of coupled limit-cycle oscillators (Reddy et al., 1998). On the other hand, under certain conditions, time delays may also enhance neural synchrony (Dhamala et al., 2004).
Conduction delays may be estimated by scaling cortico-cortical tract lengths by conduction velocity, which is usually parametrically varied between 1-30 m/s, in accordance with experimental studies (Swadlow, 1982). However, given the millisecond scale of delays involved, it may be redundant to include delays in cases where the object of interest is fMRI BOLD time scales.
Typically, whole-brain models aim to explain data collected at fMRI BOLD or electrophysiological (EEG, MEG, sEEG) time scales. Functional time series collected from fMRI experiments are used to estimate inter-areal functional connectivity. For EEG or MEG, pre-processed signals are bandpass filtered in various frequency bands of interest and Hilbert-transformed to extract amplitude envelopes, which are then used to estimate functional connectivity (Hipp et al., 2012; Deco et al., 2017). For both fMRI and EEG/MEG, typically static correlations (presuming stationarity) are employed to estimate model fit. However, recent work has strongly argued that static measures fail to capture the rich, higher-order dynamics inherent in neuroimaging data, and therefore, have advocated the use of dynamic measures of functional connectivity (dFC) (Hutchison et al., 2013; Preti et al., 2017). dFC may be estimated through a windowed manner, or through techniques not requiring arbitrarily chosen temporal windows (Cabral et al., 2017). For dFC analysis, every time step (or time window) has a characteristic FC pattern associated with it. One way to perform model fitting for dFC is by collapsing this 3D data structure (ROI*ROI*time) to a 2D matrix (time*time) consisting of correlations between the leading eigenvectors at each time point or window; the resulting matrix may be considered as the object of model fitting (Cabral et al., 2017).
Model parameters are systematically varied and simulated FCs (static or dynamic) are estimated for each parametric set. Estimation of the optimal parameter set (often referred to as the dynamic working point of the system), offering closest concordance with empirical FC may be achieved by minimizing an error function or by maximizing correlation between empirical and simulated FCs (Deco and Kringelbach, 2014). Bayesian modeling is often employed to estimate parameters associated with the underlying generative models (Vattikonda et al., 2016; Hashemi et al., 2020). Here, models are initialized with a randomly chosen parameter set; stochastic gradient descent is then used to update model parameters.
Instead of FC, one could alternatively perform model fitting against other empirical features of the data. For example, Jirsa et al. (2017) develop a personalized epileptic brain model by estimating model parameters from the spectral distribution of stereotactic (SEEG) electrodes. Indeed, it is even possible to do away with model fitting altogether when the research question is of a qualitative nature. For example, Mejias and Wang (2022) simulate a large-scale model of primate neo-cortex to elucidate the emergence of distributed attractor states subserving various internal processes. In such studies, explaining the salient aspects of the underlying system takes precedence over precise model fitting (Mišić et al., 2015; Mejias and Wang, 2022).
After successfully fitting the model to relevant empirical data, it is desired to introduce various perturbations to the model in order to understand the fallout of various pathological scenarios (Deco et al., 2015). For example, in the case of stroke or TBI, one would like to induce partial or complete lesions at various network nodes and study the differential contribution of node topology in disease progression (Alstott et al., 2009; Vattikonda et al., 2016). Since the thrust here is to understand recoverability, individual node dynamics can be endowed with plasticity mechanisms that homeostatically regulate firing rates (Vattikonda et al., 2016; Abeysuriya et al., 2018; Páscoa Dos Santos and Verschure, 2021). Similarly, stimulation protocols require providing current input to specific nodes in the network to study network response (Griffiths et al., 2020). Epilepsy models require altering node dynamics such as channel properties or neurotransmitter concentrations to model seizure spread from the seizure onset zone (SOZ) (Jirsa et al., 2017). Levels of consciousness in whole-brain models can be manipulated by adjusting neural gain, say, mediated by subcortical structures (Shine, 2021).
Epilepsy is marked by the occurrence of frequent seizures, which often spread from an onset zone to other distal areas along white matter tracts. In some cases, this necessitates the surgical resection of epileptogenic tissue. Due to the obvious role of network dynamics and structural topology, epilepsy is particularly well-suited for whole-brain modeling, as described in previous sections (Engel Jr et al., 2013; Taylor et al., 2014; Jirsa et al., 2017). Since surgery carries obvious risks, it is desirable to minimize the extent of resected tissue. Jirsa et al. show that personalized whole-brain modeling can be used to aid medical decision making for optimal surgery (Vattikonda et al., 2016; Jirsa et al., 2017). Patient-specific brain connectivity is integrated to model empirical EEG data for the identification of the epileptogenic zone (EZ). This technique is particularly useful for instances where conventional methods for EZ identification provide sub-optimal results due to a lack of a clear MRI lesion (Hashemi et al., 2020). Recently, whole-brain modeling has also been used to explain seizures in non-epileptic conditions as well. For example, it is known that patients with Alzheimer's disease are about 6-10 times more likely to develop seizures as compared to the normal population (Pandis and Scarmeas, 2012). Tait et al. (2021a) using a whole-brain pipeline, find that functional connectomes of AD patients show a greater propensity to transition into seizure states as compared to healthy connectomes. Here individual nodes in the network are modeled as phase oscillators capable of producing neuronal spiking in response to inputs. By systematically varying the excitability parameter of individual nodes, the authors show that AD connectomes are more ictogenic as compared to control connectomes for a wide range of excitatory input (Tait et al., 2021a).
Neural tissue undergoes lesioning due to various factors like traumatic brain injury, stroke or neurodegenerative diseases (Alstott et al., 2009). Focal lesions can cause disruptions in large-scale functional connectivity, leading to severe cognitive and behavioral impairment. Alstott et al. (2009) demonstrate that the extent and severity of functional deterioration depends on the topological profile of the lesioned nodes, with nodes occupying the most central position causing the greatest network deficit upon lesioning. Vattikonda et al. (2016) extend this idea to gauge potential recoverability from stroke induced lesions by endowing node dynamics with an inhibitory plasticity mechanism that can rescue neural firing rates in response to structural insult (Figure 4). Recently, Good et al. used whole-brain modeling to predict the chronic outcomes following traumatic brain injury. Their approach, which utilizes the Virtual Brain simulation platform (Sanz Leon et al., 2013), is able to distinguish semiacute mild to moderate TBI patients from a control group (Good et al., 2022). The effect of lesions on segregative and integrative tendencies can be quantified using WBMs. For example, Hellyer et al. (2015) estimate metastability- a measure of segregation and integration, and find disrupted metastable dynamics in patients with traumatic brain injury (TBI). By simulating a network of phase oscillators on topology specified by connectomes obtained from TBI patients, the authors demonstrate how structural disconnection can lead to a reduction in metastable brain dynamics. These observations provide a mechanistic explanation for the significant reductions in cognitive flexibility and information processing, often seen in patients recovering from TBI lesions. Váša et al. (2015) highlight the usefulness of computational lesion studies by demonstrating how graph theoretic properties of network nodes such as modularity determine synchrony and metastability in response to virtual lesioning. The authors find that lesions to nodes with high eigenvector centrality or to nodes which connect segregated modules lead to a decrease in global synchrony along with an increase in global metastability (Váša et al., 2015).
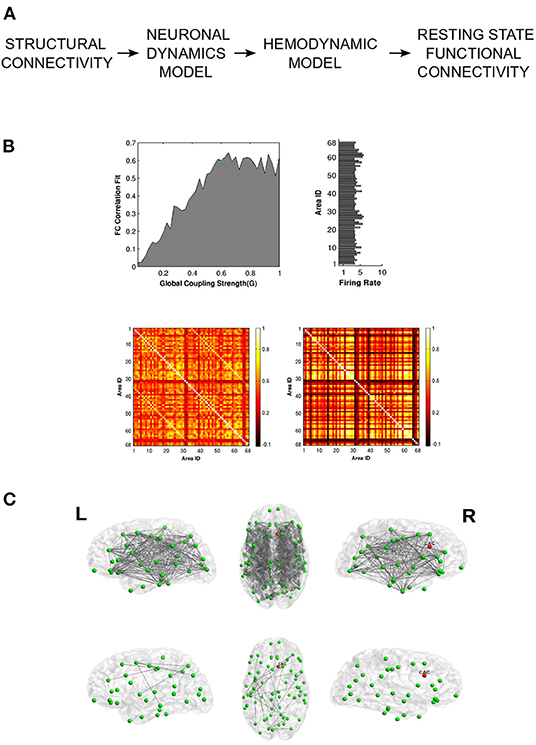
Figure 4. Vattikonda and colleagues use a whole-brain pipeline to elucidate network recovery after lesional insult. (A) DTI structural connectivity is used in conjunction with mean field node dynamics (with feedback inhibition) to generate ROI time series which are then convolved with hemodynamic function to obtain simulated BOLD time-series. Resting state FC is calculated using pearson correlation. (B) Global working point of the system is estimated by comparing simulated FC with empirical FC. Histogram shows the ROI-wise average firing rate for optimal global coupling. Heat Maps show empirical and optimal simulated FC (averaged across subjects). (C) Network recovery takes place due to local plasticity implemented through feedback inhibition. Here lesion was introduced in the right caudal anterior cingulate node. The top 3 figures display connections that have significantly changed before re-establishing local E-I balance. The bottom 3 figures represent the connections that have significantly changed after re-establishing local E-I balance. Adapted from Vattikonda et al. (2016), with permission from the authors.
Alzheimer's Disease (AD) has traditionally been regarded as a disease of the gray matter, however, recent neuroimaging studies have implicated white matter abnormalities in the pathogenesis of AD (Sachdev et al., 2013). This is reflected in aberrant functional connectivity patterns observed in preclinical populations. Demirtaş et al. (2017) fit a whole-brain model to healthy controls; the model parameters thus obtained are then systematically varied to generate FCs which match empirical FCs seen in preclinical AD, Mild Cognitive Impairment and AD. The authors find that simulated FCs mimic pathological FCs as the individual node dynamics is shifted toward damped oscillations by altering the bifurcation parameter (Demirtaş et al., 2017). Stefanovski et al. (2019) fuse PET-derived Amyloid beta levels with averaged healthy connectomes to shed light on possible pathogenetic mechanims of AD. In this model, Amyloid beta levels modulate the regional Excitation/Inhibition balance, providing a mechanistic explanation for EEG alterations in AD. Further, their whole-brain approach provides therapeutic insights by accounting for large-scale functional reversibility of EEG alterations by modeling the effect of memantine (NMDA receptor antagonist) on local neural populations (Stefanovski et al., 2019). Recently, whole-brain network models have also been utilized for virtual data completion to augment multimodal AD datasets such as the Alzheimer's Disease Neuroimaging Initiative (ADNI) dataset (Arbabyazd et al., 2021).
The dysconnection hypothesis posits that symptoms of Schizophrenia are best characterized as emerging from functional, rather than anatomical disconnection (Friston, 1998). In line with this assertion, several studies have observed extensive decrease in resting state functional connectivity of patients, pointing to disrupted integration between segregated brain areas (Lynall et al., 2010). Cabral et al. (2013) employ structural connectivity matrices obtained from adolescent patients with early onset schizophrenia and show that functional disruptions associated with Schizophrenia are better explained by reductions in global coupling rather than structural differences, in line with the dysconnection hypothesis. Yang et al. (2014) use whole-brain modeling to show that widely reported differences in global brain signal (GBS) in resting state fMRI of patients may be explained by changes in the net strength of overall brain connectivity in schizophrenia, further corroborating dysconnection. Anticevic et al. (2012) use whole-brain models to identify the role for glutamate in establishing large-scale functional patterns associated with Schizophrenia. Their whole-brain approach, which allows for the introduction of pharmacological manipulations, provides a framework for understanding the role of NMDA-mediated disruption of cortical excitation/inhibition balance and its role in producing the cognitive symptoms of schizophrenia.
Loss of consciousness is either temporary, like in deep sleep or anesthesia- or permanent- like in brain injury or other Disorders of Consciousness (DoC). Efficient classification of brain states as either reversibly or irreversibly unconscious is needed to advance therapeutics. One way to gauge whether a certain unconscious state is transient or permanent is from the response of that state to externally provided perturbation. Recently, whole-brain models have been used to characterize brain states in terms of their stability toward perturbation. Sanz Perl et al. (2021) demonstrate that perturbational analysis can complement machine-learning based algorithms which classify different states of consciousness. López-González et al. (2021) use structural connectivity from healthy and injured subjects to show that low-level states of consciousness are associated with decreased network interactions, leading to segregation of synchronization patterns in fMRI brain dynamics. Segregative tendencies are found to be associated with the global coupling parameter that scales the weights of the SC matrix.
Since whole-brain approaches leverage neuroimaging modalities for modeling neural dynamics, future improvement is contingent upon parallel advances in diffusion imaging, functional imaging and signal processing techniques. Here, we discuss a few directions that can significantly augment current neurocomputational models.
One potential avenue for enriching current whole-brain models is by improving the estimation of structural adjacency matrices. For example, DTI derived structural matrices are bidirectional, whereas actual white matter fibers have a well-defined point of origin and termination which imparts directionality and has obvious consequences for the emerging dynamics. Additionally, current protocols for structural estimation rely on the number of streamline (NOS) methods which reconstruct structure by counting the number of streamlines between ROI pairs. Although showing concordance with tract-tracing, the NOS method has inherent limitations as it does not consider other biologically crucial parameters like conduction speeds. Thus, estimation of structural connectivity matrices can be further improved by inclusion of myeloarchitecture, since myelin plays a crucial role in determining conduction speeds across axons (Boshkovski et al., 2021). One way to achieve this is by weighting the connectome with longitudinal relaxation rate (R1), which is sensitive to myelin. Boshkovski et al. (2021) show how including myelin weighted structural connectomes is successful at separating transmodal regions from unimodal regions . Inclusion of myelin in network simulations has particular application at electrophysiological time-scales where phase lags often arise due to finite conduction delays (Petkoski and Jirsa, 2019). g-ratio, which quantifies the ratio between axon diameter and myelin thickness, has recently been shown to be estimable through MRI protocols (Berman et al., 2019; Drakesmith et al., 2019). In vivo g-ratio mapping has the potential to provide novel insights into cortical conduction speeds (Berman et al., 2019; Drakesmith et al., 2019). Another method being currently explored for the estimation of cortical conduction velocity uses direct electrical stimulation to measure the propagation of electrophysiological responses across the cortex in patients implanted with intracranial electrodes for seizure monitoring (David, 2021). Harnessing signal propagation information has far-reaching applications, especially toward understanding various demyelinating disorders such as multiple sclerosis.
Further augmentation of whole-brain connectomes comes from incorporating neuromodulatory information (Deco et al., 2018; Kringelbach et al., 2020; Naskar et al., 2021). Multi-modal integration between diffusion imaging (structural connectivity) and PET (receptor density) allows for the infusion of dynamic information to static network models. Kringelbach et al. (2020) have employed a similar pipeline to model the bidirectional interaction of neuronal and neurotransmitter systems that sheds light on the action of psilocybin on human resting state activity. Understanding large-scale functional impact of neuromodulation is of primary importance to computational neuropsychiatry given the therapeutic potential of psychedelics in the treatment of anxiety and depression (Deco et al., 2018).
Another limitation of most current large-scale models is the absence of sub-cortical nodes in the network. This is partly due to inadequate resolution offered by most atlases at the sub-cortical level. Additionally, various sub-cortical structures (e.g., thalamus) possess unique network architecture, requiring the development of specialized node dynamics (see thalamocortical motifs, Figure 3). Here we direct the interested reader to some recent efforts toward addressing this lacuna (see Shine et al., 2018; Griffiths et al., 2020; Shine, 2021). Future developments in high field strength imaging, sub-cortical node dynamics and parcellations offer the possibility of having truly whole-brain models.
Despite substantial progress in the field, most successful whole-brain models are limited to either BOLD (fMRI) or BOLD time-scale (amplitude envelopes) functional correlations. Lacunae exist about the extent to which whole-brain models may explain phenomena at electrophysiological time-scales, especially since neural oscillations are so well-linked to the underlying white matter structure and are crucial to cognition (Chu et al., 2015; Hindriks et al., 2015). Signal processing techniques that circumvent or correct for volume/field spread effects which tend to contaminate electrophysiological data would go a long way toward informing whole-brain modeling (Hipp et al., 2012).
Similarly, the present thrust of whole-brain approaches is oriented toward modeling recordings while participants are not engaged in overt cognition, aka resting-state (Biswal et al., 1995; Deco et al., 2011; Popovych et al., 2019). Going forward, whole-brain models could also be explored for explaining various tasks and learning paradigms, requiring richer node dynamics with neuromodulatory and plasticity properties (Abel et al., 2013; Maniglia and Seitz, 2018; Zhang et al., 2021). Finally, foundational discoveries in graph theory and non-equilibrium physics will continue to offer new insights into the mechanistic underpinnings of large-scale brain dynamics.
Computational neuroscience aims to understand the biophysical principles underlying brain function. Many cognitive phenomena crucial for understanding the brain in health and disease evolve at the mesoscopic scale, where the firing patterns of individual neurons get averaged out, thereby offering an opportunity for radical dimensionality reduction. Whole-brain models leverage new advances in neuroimaging techniques to simulate white matter-mediated large-scale brain networks that underlie cognitive and behavioral processes in health and disease. In this article, we provided a brief outline of how coarsely grained models of brain dynamics may be employed to gain insights into the mechanistic underpinnings of brain dynamics, an endeavor central to the emerging field of computational psychiatry. We summarized the various choices at hand for the successful implementation of whole-brain pipelines and discussed those in the context of relevant case studies. Researchers must be mindful of how the choices of parcellation, node dynamics, model fitting procedure and perturbation impact the modeling pipeline and relate to the underlying scientific objective of the study.
We discussed how large-scale modeling has provided crucial insights into the biology of various neuropathologies like Epilepsy, Stroke, Traumatic Brain Injury, Alzheimer's Disease, Schizophrenia and Disorders of Consciousness. Like any emerging field, whole-brain modeling also requires further developments to tap into its full potential and we provided methodological and technical recommendations for the growth of large-scale modeling. Improvements in structural brain imaging and signal processing techniques can significantly enhance the accuracy of neurocomputational models. Similarly, the inclusion of sub-cortical, neurotransmitter and myelination information can lead the field toward truly whole-brain models. Going forward, the continued development of new computational platforms like the Virtual Brain simulator (Sanz Leon et al., 2013) is likely to bridge the gap between theory and implementation, making whole-brain modeling more accessible to medical professionals and biologists alike. In closing, whole-brain models are the newest addition to the rich arsenal of computational neuroscience techniques and promise to usher in a new era in personalized medicine.
AP and AB conceived the study. AP wrote the first draft. AP, DR, and AB wrote and revised manuscript. All authors contributed to the article and approved the submitted version.
DR, Ramalingaswami Fellowship, Department of Biotechnology, Government of India, Award ID: BT/RLF/Re-entry/07/2014. DR, Department of Science and Technology (DST), Ministry of Science and Technology, Government of India, Award ID: SR/CSRI/21/2016. AB, Ministry of Youth Affairs and Sports, Government of India, Award ID: F.NO.K-15015/42/2018/SP-V. AB, NBRC Flagship program, Department of Biotechnology, Government of India, Award ID: BT/MED-III/NBRC/Flagship/Flagship2019.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge the generous support of NBRC Core funds.
MRI, Magnetic resonance imaging; WBM, Whole-Brain Model; ROI, Region of Interest; BOLD, Blood oxygen level dependent; EEG/MEG, Electro/Magneto encephalography; FC, Functional connectivity.
1. ^Several thousand neurons exhibiting coordinated firing patterns.
2. ^In mean field models, coarse-grained variables representing ensemble activity such as the population firing rate are used to track the evolution of the system.
Abel, T., Havekes, R., Saletin, J. M., and Walker, M. P. (2013). Sleep, plasticity and memory from molecules to whole-brain networks. Curr. Biol. 23, R774–R788. doi: 10.1016/j.cub.2013.07.025
Abeysuriya, R. G., Hadida, J., Sotiropoulos, S. N., Jbabdi, S., Becker, R., Hunt, B. A., et al. (2018). A biophysical model of dynamic balancing of excitation and inhibition in fast oscillatory large-scale networks. PLoS Comput. Biol. 14, e1006007. doi: 10.1371/journal.pcbi.1006007
Alstott, J., Breakspear, M., Hagmann, P., Cammoun, L., and Sporns, O. (2009). Modeling the impact of lesions in the human brain. PLoS Comput. Biol. 5, e1000408. doi: 10.1371/journal.pcbi.1000408
Amari, S.-I. (1977). Dynamics of pattern formation in lateral-inhibition type neural fields. Biol. Cybern. 27, 77–87. doi: 10.1007/BF00337259
Anticevic, A., Gancsos, M., Murray, J. D., Repovs, G., Driesen, N. R., Ennis, D. J., et al. (2012). NMDA receptor function in large-scale anticorrelated neural systems with implications for cognition and schizophrenia. Proc. Natl. Acad. Sci. U.S.A. 109, 16720–16725. doi: 10.1073/pnas.1208494109
Arbabyazd, L., Shen, K., Wang, Z., Hofmann-Apitius, M., Ritter, P., McIntosh, A. R., et al. (2021). Virtual connectomic datasets in alzheimer's disease and aging using whole-brain network dynamics modelling. Eneuro 8, ENEURO.0475-20.2021. doi: 10.1523/ENEURO.0475-20.2021
Banerjee, A., and Jirsa, V. K. (2007). How do neural connectivity and time delays influence bimanual coordination? Biol. Cybern. 96, 265–278. doi: 10.1007/s00422-006-0114-4
Bastiani, M., Shah, N. J., Goebel, R., and Roebroeck, A. (2012). Human cortical connectome reconstruction from diffusion weighted mri: the effect of tractography algorithm. Neuroimage 62, 1732–1749. doi: 10.1016/j.neuroimage.2012.06.002
Bazin, P.-L., Alkemade, A., Mulder, M. J., Henry, A. G., and Forstmann, B. U. (2020). Multi-contrast anatomical subcortical structures parcellation. Elife 9, e59430. doi: 10.7554/eLife.59430
Berman, S., Filo, S., and Mezer, A. A. (2019). Modeling conduction delays in the corpus callosum using MRI-measured g-ratio. Neuroimage 195, 128–139. doi: 10.1016/j.neuroimage.2019.03.025
Beurle, R. L. (1956). Properties of a mass of cells capable of regenerating pulses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 240, 55–94. doi: 10.1098/rstb.1956.0012
Biswal, B., Zerrin Yetkin, F., Haughton, V. M., and Hyde, J. S. (1995). Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn. Reson. Med. 34, 537–541. doi: 10.1002/mrm.1910340409
Boshkovski, T., Kocarev, L., Cohen-Adad, J., Mišić, B., Lehéricy, S., Stikov, N., et al. (2021). The R1-weighted connectome: complementing brain networks with a myelin-sensitive measure. Netw. Neurosci. 5, 358–372. doi: 10.1162/netn_a_00179
Breakspear, M., Heitmann, S., and Daffertshofer, A. (2010). Generative models of cortical oscillations: neurobiological implications of the kuramoto model. Front. Hum. Neurosci. 4, 190. doi: 10.3389/fnhum.2010.00190
Bressler, S. L., and Tognoli, E. (2006). Operational principles of neurocognitive networks. Int. J. Psychophysiol. 60, 139–148. doi: 10.1016/j.ijpsycho.2005.12.008
Cabral, J., Fernandes, H. M., Van Hartevelt, T. J., James, A. C., Kringelbach, M. L., and Deco, G. (2013). Structural connectivity in schizophrenia and its impact on the dynamics of spontaneous functional networks. Chaos 23, 046111. doi: 10.1063/1.4851117
Cabral, J., Vidaurre, D., Marques, P., Magalh aes, R., Moreira, P. S., Soares, J. M., et al. (2017). Cognitive performance in healthy older adults relates to spontaneous switching between states of functional connectivity during rest. Sci. Rep. 7, 1–13. doi: 10.1038/s41598-017-05425-7
Chu, C. J., Tanaka, N., Diaz, J., Edlow, B. L., Wu, O., Hämäläinen, M., et al. (2015). EEG functional connectivity is partially predicted by underlying white matter connectivity. Neuroimage 108, 23–33. doi: 10.1016/j.neuroimage.2014.12.033
Craddock, R. C., James, G. A., Holtzheimer, III, P. E., Hu, X. P., and Mayberg, H. S. (2012). A whole brain fmri atlas generated via spatially constrained spectral clustering. Hum. Brain Mapp. 33, 1914–1928. doi: 10.1002/hbm.21333
David, O. (2021). Functional brain tractography. Brain Stimul. 14, 1729. doi: 10.1016/j.brs.2021.10.466
Deco, G., Cabral, J., Woolrich, M. W., Stevner, A. B., Van Hartevelt, T. J., and Kringelbach, M. L. (2017). Single or multiple frequency generators in on-going brain activity: a mechanistic whole-brain model of empirical MEG data. Neuroimage 152, 538–550. doi: 10.1016/j.neuroimage.2017.03.023
Deco, G., Cruzat, J., Cabral, J., Knudsen, G. M., Carhart-Harris, R. L., Whybrow, P. C., et al. (2018). Whole-brain multimodal neuroimaging model using serotonin receptor maps explains non-linear functional effects of LSD. Curr. Biol. 28, 3065–3074. doi: 10.1016/j.cub.2018.07.083
Deco, G., Jirsa, V., McIntosh, A. R., Sporns, O., and Kötter, R. (2009). Key role of coupling, delay, and noise in resting brain fluctuations. Proc. Natl. Acad. Sci. U.S.A. 106, 10302–10307. doi: 10.1073/pnas.0901831106
Deco, G., Jirsa, V. K., and McIntosh, A. R. (2011). Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 12, 43–56. doi: 10.1038/nrn2961
Deco, G., Jirsa, V. K., Robinson, P. A., Breakspear, M., and Friston, K. (2008). The dynamic brain: from spiking neurons to neural masses and cortical fields. PLoS Comput. Biol. 4, e1000092. doi: 10.1371/journal.pcbi.1000092
Deco, G., and Kringelbach, M. L. (2014). Great expectations: using whole-brain computational connectomics for understanding neuropsychiatric disorders. Neuron 84, 892–905. doi: 10.1016/j.neuron.2014.08.034
Deco, G., Ponce-Alvarez, A., Mantini, D., Romani, G. L., Hagmann, P., and Corbetta, M. (2013). Resting-state functional connectivity emerges from structurally and dynamically shaped slow linear fluctuations. J. Neurosci. 33, 11239–11252. doi: 10.1523/JNEUROSCI.1091-13.2013
Deco, G., Tononi, G., Boly, M., and Kringelbach, M. L. (2015). Rethinking segregation and integration: contributions of whole-brain modelling. Nat. Rev. Neurosci. 16, 430–439. doi: 10.1038/nrn3963
Demirtaş, M., Falcon, C., Tucholka, A., Gispert, J. D., Molinuevo, J. L., and Deco, G. (2017). A whole-brain computational modeling approach to explain the alterations in resting-state functional connectivity during progression of alzheimer's disease. Neuroimage 16, 343–354. doi: 10.1016/j.nicl.2017.08.006
Dhamala, M., Jirsa, V. K., and Ding, M. (2004). Enhancement of neural synchrony by time delay. Phys. Rev. Lett. 92, 074104. doi: 10.1103/PhysRevLett.92.074104
Domhof, J. W., Jung, K., Eickhoff, S. B., and Popovych, O. V. (2021). Parcellation-induced variation of empirical and simulated brain connectomes at group and subject levels. Netw. Neurosci. 5, 798–830. doi: 10.1162/netn_a_00202
Drakesmith, M., Harms, R., Rudrapatna, S. U., Parker, G. D., Evans, C. J., and Jones, D. K. (2019). Estimating axon conduction velocity in vivo from microstructural MRI. Neuroimage 203, 116186. doi: 10.1016/j.neuroimage.2019.116186
Engel Jr, J., Thompson, P. M., Stern, J. M., Staba, R. J., Bragin, A., and Mody, I. (2013). Connectomics and epilepsy. Curr. Opin. Neurol. 26, 186. doi: 10.1097/WCO.0b013e32835ee5b8
Ermentrout, G. B., and Cowan, J. D. (1979). A mathematical theory of visual hallucination patterns. Biol. Cybern. 34, 137–150. doi: 10.1007/BF00336965
Ermentrout, G. B., and Kleinfeld, D. (2001). Traveling electrical waves in cortex: insights from phase dynamics and speculation on a computational role. Neuron 29, 33–44. doi: 10.1016/S0896-6273(01)00178-7
Fornito, A., Zalesky, A., and Bullmore, E. T. (2010). Network scaling effects in graph analytic studies of human resting-state fMRI data. Front. Syst. Neurosci. 4, 22. doi: 10.3389/fnsys.2010.00022
Friston, K. J. (1998). The disconnection hypothesis. Schizophr. Res. 30, 115–125. doi: 10.1016/S0920-9964(97)00140-0
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell, J., Yacoub, E., et al. (2016). A multi-modal parcellation of human cerebral cortex. Nature 536, 171–178. doi: 10.1038/nature18933
Gollo, L. L., Roberts, J. A., and Cocchi, L. (2017). Mapping how local perturbations influence systems-level brain dynamics. Neuroimage 160, 97–112. doi: 10.1016/j.neuroimage.2017.01.057
Good, T., Schirner, M., Shen, K., Ritter, P., Mukherjee, P., Levine, B., et al. (2022). Personalized connectome-based modeling in patients with semi-acute phase tbi: Relationship to acute neuroimaging and 6 month follow-up. Eneuro 9, ENEURO.0075-21.2022. doi: 10.1523/ENEURO.0075-21.2022
Griffiths, J. D., McIntosh, A. R., and Lefebvre, J. (2020). A connectome-based, corticothalamic model of state-and stimulation-dependent modulation of rhythmic neural activity and connectivity. Front. Comput. Neurosci. 14, 575143. doi: 10.3389/fncom.2020.575143
Hansen, E. C., Battaglia, D., Spiegler, A., Deco, G., and Jirsa, V. K. (2015). Functional connectivity dynamics: modeling the switching behavior of the resting state. Neuroimage 105, 525–535. doi: 10.1016/j.neuroimage.2014.11.001
Hashemi, M., Vattikonda, A., Sip, V., Guye, M., Bartolomei, F., Woodman, M. M., et al. (2020). The bayesian virtual epileptic patient: a probabilistic framework designed to infer the spatial map of epileptogenicity in a personalized large-scale brain model of epilepsy spread. Neuroimage 217, 116839. doi: 10.1016/j.neuroimage.2020.116839
Hellyer, P. J., Scott, G., Shanahan, M., Sharp, D. J., and Leech, R. (2015). Cognitive flexibility through metastable neural dynamics is disrupted by damage to the structural connectome. J. Neurosci. 35, 9050–9063. doi: 10.1523/JNEUROSCI.4648-14.2015
Hindriks, R., Woolrich, M., Luckhoo, H., Joensson, M., Mohseni, H., Kringelbach, M. L., et al. (2015). Role of white-matter pathways in coordinating alpha oscillations in resting visual cortex. Neuroimage 106, 328–339. doi: 10.1016/j.neuroimage.2014.10.057
Hipp, J. F., Hawellek, D. J., Corbetta, M., Siegel, M., and Engel, A. K. (2012). Large-scale cortical correlation structure of spontaneous oscillatory activity. Nat. Neurosci. 15, 884–890. doi: 10.1038/nn.3101
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J.-P., Meuli, R., et al. (2009). Predicting human resting-state functional connectivity from structural connectivity. Proc. Natl. Acad. Sci. U.S.A. 106, 2035–2040. doi: 10.1073/pnas.0811168106
Horwitz, B., Friston, K. J., and Taylor, J. G. (2000). Neural modeling and functional brain imaging: an overview. Neural Netw. 13/, 829–846. doi: 10.1016/S0893-6080(00)00062-9
Horwitz, B., Tagamets, M., and McIntosh, A. R. (1999). Neural modeling, functional brain imaging, and cognition. Trends Cogn. Sci. 3, 91–98. doi: 10.1016/S1364-6613(99)01282-6
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A., Calhoun, V. D., Corbetta, M., et al. (2013). Dynamic functional connectivity: promise, issues, and interpretations. Neuroimage 80, 360–378. doi: 10.1016/j.neuroimage.2013.05.079
James, G. A., Hazaroglu, O., and Bush, K. A. (2016). A human brain atlas derived via n-cut parcellation of resting-state and task-based fmri data. Magn. Reson. Imaging 34, 209–218. doi: 10.1016/j.mri.2015.10.036
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W., and Smith, S. M. (2012). Fsl. Neuroimage 62, 782–790. doi: 10.1016/j.neuroimage.2011.09.015
Ji, B., Li, Z., Li, K., Li, L., Langley, J., Shen, H., et al. (2016). Dynamic thalamus parcellation from resting-state fMRI data. Hum. Brain Mapp. 37, 954–967. doi: 10.1002/hbm.23079
Jirsa, V. K., and Haken, H. (1996). Field theory of electromagnetic brain activity. Phys. Rev. Lett. 77, 960. doi: 10.1103/PhysRevLett.77.960
Jirsa, V. K., Proix, T., Perdikis, D., Woodman, M. M., Wang, H., Gonzalez-Martinez, J., et al. (2017). The virtual epileptic patient: individualized whole-brain models of epilepsy spread. Neuroimage 145, 377–388. doi: 10.1016/j.neuroimage.2016.04.049
Kringelbach, M. L., Cruzat, J., Cabral, J., Knudsen, G. M., Carhart-Harris, R., Whybrow, P. C., et al. (2020). Dynamic coupling of whole-brain neuronal and neurotransmitter systems. Proc. Natl. Acad. Sci. U.S.A. 117, 9566–9576. doi: 10.1073/pnas.1921475117
López-González, A., Panda, R., Ponce-Alvarez, A., Zamora-López, G., Escrichs, A., Martial, C., et al. (2021). Loss of consciousness reduces the stability of brain hubs and the heterogeneity of brain dynamics. Commun. Biol. 4, 1–15. doi: 10.1038/s42003-021-02537-9
Lord, L.-D., Stevner, A. B., Deco, G., and Kringelbach, M. L. (2017). Understanding principles of integration and segregation using whole-brain computational connectomics: implications for neuropsychiatric disorders. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 375, 20160283. doi: 10.1098/rsta.2016.0283
Lynall, M.-E., Bassett, D. S., Kerwin, R., McKenna, P. J., Kitzbichler, M., Muller, U., et al. (2010). Functional connectivity and brain networks in schizophrenia. J. Neurosci. 30, 9477–9487. doi: 10.1523/JNEUROSCI.0333-10.2010
Maniglia, M., and Seitz, A. R. (2018). Towards a whole brain model of perceptual learning. Curr. Opin. Behav. Sci. 20, 47–55. doi: 10.1016/j.cobeha.2017.10.004
McIntosh, A. R. (2004). Contexts and catalysts. Neuroinformatics 2, 175–181. doi: 10.1385/NI:2,2:175
Mejias, J. F., and Wang, X.-J. (2022). Mechanisms of distributed working memory in a large-scale network of macaque neocortex. Elife 11, e72136. doi: 10.7554/eLife.72136
Mišić, B., Betzel, R. F., Nematzadeh, A., Goni, J., Griffa, A., Hagmann, P., et al. (2015). Cooperative and competitive spreading dynamics on the human connectome. Neuron 86, 1518–1529. doi: 10.1016/j.neuron.2015.05.035
Nakagawa, T. T., Woolrich, M., Luckhoo, H., Joensson, M., Mohseni, H., Kringelbach, M. L., et al. (2014). How delays matter in an oscillatory whole-brain spiking-neuron network model for MEG alpha-rhythms at rest. Neuroimage 87, 383–394. doi: 10.1016/j.neuroimage.2013.11.009
Naskar, A., Vattikonda, A., Deco, G., Roy, D., and Banerjee, A. (2021). Multi-scale dynamic mean field model (MDMF) relates resting-state brain dynamics with local cortical excitatory-inhibitory neurotransmitter homeostasis. Netw. Neurosci. 5, 757–782. doi: 10.1162/netn_a_00197
Niebur, E., Schuster, H. G., and Kammen, D. M. (1991). Collective frequencies and metastability in networks of limit-cycle oscillators with time delay. Phys. Rev. Lett. 67, 2753. doi: 10.1103/PhysRevLett.67.2753
Nowinski, W. L. (2021). Evolution of human brain atlases in terms of content, applications, functionality, and availability. Neuroinformatics 19, 1–22. doi: 10.1007/s12021-020-09481-9
Pajevic, S., Basser, P. J., and Fields, R. D. (2014). Role of myelin plasticity in oscillations and synchrony of neuronal activity. Neuroscience 276, 135–147. doi: 10.1016/j.neuroscience.2013.11.007
Pandis, D., and Scarmeas, N. (2012). Seizures in alzheimer disease: clinical and epidemiological data: Seizures in alzheimer disease. Epilepsy Curr. 12, 184–187. doi: 10.5698/1535-7511-12.5.184
Páscoa Dos Santos, F., and Verschure, P. F. (2021). Excitatory-inhibitory homeostasis and diaschisis: tying the local and global scales in the post-stroke cortex. Front. Syst. Neurosci. 15, 806544doi: 10.3389/fnsys.2021.806544
Pathak, A., Sharma, V., Roy, D., and Banerjee, A. (2021). Preservation of neural synchrony at peak alpha frequency via global synaptic scaling compensates for white matter structural decline over adult lifespan. bioRxiv. doi: 10.1101/2021.10.24.465613
Petkoski, S., and Jirsa, V. K. (2019). Transmission time delays organize the brain network synchronization. Philos. Trans. R. Soc. A 377, 20180132. doi: 10.1098/rsta.2018.0132
Petrov, D., Ivanov, A., Faskowitz, J., Gutman, B., Moyer, D., Villalon, J., et al. (2017). “Evaluating 35 methods to generate structural connectomes using pairwise classification,” in International Conference on medical Image Computing and Computer-Assisted Intervention (Quebec: Springer), 515–522.
Popovych, O. V., Manos, T., Hoffstaedter, F., and Eickhoff, S. B. (2019). What can computational models contribute to neuroimaging data analytics? Front. Syst. Neurosci. 12, 68. doi: 10.3389/fnsys.2018.00068
Preti, M. G., Bolton, T. A., and Van De Ville, D. (2017). The dynamic functional connectome: state-of-the-art and perspectives. Neuroimage 160, 41–54. doi: 10.1016/j.neuroimage.2016.12.061
Proix, T., Spiegler, A., Schirner, M., Rothmeier, S., Ritter, P., and Jirsa, V. K. (2016). How do parcellation size and short-range connectivity affect dynamics in large-scale brain network models? Neuroimage 142, 135–149. doi: 10.1016/j.neuroimage.2016.06.016
Reddy, D. R., Sen, A., and Johnston, G. L. (1998). Time delay induced death in coupled limit cycle oscillators. Phys. Rev. Lett. 80, 5109. doi: 10.1103/PhysRevLett.80.5109
Ritter, P., Schirner, M., McIntosh, A. R., and Jirsa, V. K. (2013). The virtual brain integrates computational modeling and multimodal neuroimaging. Brain Connect 3, 121–145. doi: 10.1089/brain.2012.0120
Robinson, P. A., Henderson, J. A., Gabay, N. C., Aquino, K. M., Babaie-Janvier, T., and Gao, X. (2021). Determination of dynamic brain connectivity via spectral analysis. Front. Hum. Neurosci. 15, 655576. doi: 10.3389/fnhum.2021.655576
Roy, D., Sigala, R., Breakspear, M., McIntosh, A. R., Jirsa, V. K., Deco, G., et al. (2014). Using the virtual brain to reveal the role of oscillations and plasticity in shaping brain's dynamical landscape. Brain Connect. 4, 791–811. doi: 10.1089/brain.2014.0252
Sachdev, P. S., Zhuang, L., Braidy, N., and Wen, W. (2013). Is alzheimer's a disease of the white matter? Curr. Opin Psychiatry 26, 244–251. doi: 10.1097/YCO.0b013e32835ed6e8
Sanz Leon, P., Knock, S. A., Woodman, M. M., Domide, L., Mersmann, J., McIntosh, A. R., et al. (2013). The virtual brain: a simulator of primate brain network dynamics. Front. Neuroinform. 7, 10. doi: 10.3389/fninf.2013.00010
Sanz Perl, Y., Pallavicini, C., Pérez Ipi na, I., Demertzi, A., Bonhomme, V., Martial, C., et al. (2021). Perturbations in dynamical models of whole-brain activity dissociate between the level and stability of consciousness. PLoS Comput. Biol. 17, e1009139. doi: 10.1371/journal.pcbi.1009139
Shattuck, D. W., and Leahy, R. M. (2002). Brainsuite: an automated cortical surface identification tool. Med. Image Anal. 6, 129–142. doi: 10.1016/S1361-8415(02)00054-3
Shine, J. M. (2021). The thalamus integrates the macrosystems of the brain to facilitate complex, adaptive brain network dynamics. Progr. Neurobiol. 199, 101951. doi: 10.1016/j.pneurobio.2020.101951
Shine, J. M., Aburn, M. J., Breakspear, M., and Poldrack, R. A. (2018). The modulation of neural gain facilitates a transition between functional segregation and integration in the brain. Elife 7, e31130. doi: 10.7554/eLife.31130
Stefanovski, L., Triebkorn, P., Spiegler, A., Diaz-Cortes, M.-A., Solodkin, A., Jirsa, V., et al. (2019). Linking molecular pathways and large-scale computational modeling to assess candidate disease mechanisms and pharmacodynamics in Alzheimer's disease. Front. Comput. Neurosci. 13, 54. doi: 10.3389/fncom.2019.00054
Swadlow, H. A. (1982). Impulse conduction in the mammalian brain: physiological properties of individual axons monitored for several months. Science 218, 911–913. doi: 10.1126/science.7134984
Tagamets, M., and Horwitz, B. (1998). Integrating electrophysiological and anatomical experimental data to create a large-scale model that simulates a delayed match-to-sample human brain imaging study. Cereb. Cortex 8, 310–320. doi: 10.1093/cercor/8.4.310
Tait, L., Lopes, M. A., Stothart, G., Baker, J., Kazanina, N., Zhang, J., et al. (2021a). A large-scale brain network mechanism for increased seizure propensity in alzheimer's disease. PLoS Comput. Biol. 17, 1–21. doi: 10.1101/2021.01.19.427236
Tait, L., Özkan, A., Szul, M. J., and Zhang, J. (2021b). A systematic evaluation of source reconstruction of resting MEG of the human brain with a new high-resolution atlas: Performance, precision, and parcellation. Hum. Brain Mapp. 42, 4685–4707. doi: 10.1002/hbm.25578
Taylor, P. N., Kaiser, M., and Dauwels, J. (2014). Structural connectivity based whole brain modelling in epilepsy. J. Neurosci. Methods 236, 51–57. doi: 10.1016/j.jneumeth.2014.08.010
Thakur, B., Mukherjee, A., Sen, A., and Banerjee, A. (2016). A dynamical framework to relate perceptual variability with multisensory information processing. Sci. Rep. 6, 1–13. doi: 10.1038/srep31280
Tournier, J.-D., Calamante, F., and Connelly, A. (2012). Mrtrix: diffusion tractography in crossing fiber regions. Int. J. Imaging Syst. Technol. 22, 53–66. doi: 10.1002/ima.22005
Váša, F., Shanahan, M., Hellyer, P. J., Scott, G., Cabral, J., and Leech, R. (2015). Effects of lesions on synchrony and metastability in cortical networks. Neuroimage 118, 456–467. doi: 10.1016/j.neuroimage.2015.05.042
Vattikonda, A., Surampudi, B. R., Banerjee, A., Deco, G., and Roy, D. (2016). Does the regulation of local excitation-inhibition balance aid in recovery of functional connectivity? a computational account. Neuroimage 136, 57–67. doi: 10.1016/j.neuroimage.2016.05.002
Wilson, H. R., and Cowan, J. D. (1972). Excitatory and inhibitory interactions in localized populations of model neurons. Biophys. J. 12, 1–24. doi: 10.1016/S0006-3495(72)86068-5
Yang, G. J., Murray, J. D., Repovs, G., Cole, M. W., Savic, A., Glasser, M. F., et al. (2014). Altered global brain signal in schizophrenia. Proc. Natl. Acad. Sci. U.S.A. 111, 7438–7443. doi: 10.1073/pnas.1405289111
Yeh, F.-C., Verstynen, T. D., Wang, Y., Fernández-Miranda, J. C., and Tseng, W.-Y. I. (2013). Deterministic diffusion fiber tracking improved by quantitative anisotropy. PLoS ONE 8, e80713. doi: 10.1371/journal.pone.0080713
Zalesky, A., Fornito, A., Harding, I. H., Cocchi, L., Yücel, M., Pantelis, C., et al. (2010). Whole-brain anatomical networks: does the choice of nodes matter? Neuroimage 50, 970–983. doi: 10.1016/j.neuroimage.2009.12.027
Zhan, L., Zhou, J., Wang, Y., Jin, Y., Jahanshad, N., Prasad, G., et al. (2015). Comparison of nine tractography algorithms for detecting abnormal structural brain networks in alzheimer's disease. Front. Aging Neurosci. 7, 48. doi: 10.3389/fnagi.2015.00048
Keywords: whole brain model, neural mass, neural field, network, neuroimaging, DTI, connectome
Citation: Pathak A, Roy D and Banerjee A (2022) Whole-Brain Network Models: From Physics to Bedside. Front. Comput. Neurosci. 16:866517. doi: 10.3389/fncom.2022.866517
Received: 31 January 2022; Accepted: 02 May 2022;
Published: 26 May 2022.
Edited by:
Si Wu, Peking University, ChinaReviewed by:
Thomas Bolton, Advanced Telecommunications Research Institute International (ATR), JapanCopyright © 2022 Pathak, Roy and Banerjee. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Arpan Banerjee, YXJwYW5AbmJyYy5hYy5pbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.