 Maximiliano José Nigro1*
Maximiliano José Nigro1* Hinako Kirikae1,2
Hinako Kirikae1,2 Kasper Kjelsberg1
Kasper Kjelsberg1 Rajeevkumar Raveendran Nair1
Rajeevkumar Raveendran Nair1 Menno P. Witter1
Menno P. Witter1- 1Center for Neural Computation, Egil and Pauline Braathen and Fred Kavli Center for Cortical Microcircuits, Kavli Institute for Systems Neuroscience, Norwegian University of Science and Technology (NTNU), Trondheim, Norway
- 2Department of Organ Anatomy, Graduate School of Medicine, Tohoku University, Sendai, Japan
The wide diversity of cortical inhibitory neuron types populating the cortex allows the assembly of diverse microcircuits and endows these circuits with different computational properties. Thus, characterizing neuronal diversity is fundamental to describe the building blocks of cortical microcircuits and probe their function. To this purpose, the mouse has emerged as a powerful tool to genetically label and manipulate specific inhibitory cell-types in the mammalian brain. Among these cell-types, the parvalbumin-expressing interneuron type (PV-INs) is perhaps the most characterized. Several mouse lines have been generated to target PV-INs. Among these mouse lines, the PV-IRES-Cre lines is the most widely used and demonstrated a high specificity and efficiency in targeting PV-INs in different cortical areas. However, a characterization of the performance across cortical regions is still missing. Here we show that the PV-IRES-Cre mouse line labels only a fraction of PV immunoreactive neurons in perirhinal cortex and other association areas. Our results point to a yet uncharacterized diversity within the PV-INs and emphasize the need to characterize these tools in specific cortical areas.
Introduction
The cerebral cortex is populated by a wide diversity of glutamatergic and GABAergic neurons (Harris and Shepherd, 2015). GABAergic neurons represent 10–15% of the neuronal population, but their diversity allows them to participate in different microcircuits and shape information flow in the cortex (Kepecs and Fishell, 2014). The study of neuronal diversity was propelled by the discovery of molecular markers that are specifically expressed by neuronal classes. GABAergic neurons can be broadly divided in three major classes according to the non-overlapping expression of parvalbumin (PV), somatostatin (SST) and 5-hydroxytryptamine 3a receptor (5HT3aR) (Tremblay et al., 2016). The discovery of molecular markers has been exploited to produce transgenic mouse lines expressing fluorophores or recombinases to label or manipulate specific neuronal classes (Madisen et al., 2010; Taniguchi et al., 2011). Parvalbumin-expressing GABAergic neurons (PV-INs) are the major group of inhibitory neurons in the mammalian cortex, representing about 40% of the GABAergic population (Tremblay et al., 2016). PV-INs provide powerful perisomatic inhibition to excitatory neurons and are involved in feedforward and feedback circuits (Hu et al., 2014; Tremblay et al., 2016). PV-INs encompass two major morphological cell-types: fast-spiking basket cells that target the soma of postsynaptic neurons, and chandelier cells, targeting the axon initial segment. The development of Knock-in mice expressing Cre or FlpO recombinase under the control of the PV promoter (PV-IRES-Cre) has provided researchers with a powerful tool to explore the functional role of PV-INs in the cortical circuit (Hippenmeyer et al., 2005). The PV-IRES-Cre mouse line has been shown to efficiently and specifically target PV-INs in the visual cortex (Atallah et al., 2012), and in the medial entorhinal cortex (Martínez et al., 2017). Recently the PV-IRES-Cre mouse line has been used to quantify the density of PV-INs across the whole cortex (Whissell et al., 2015). These studies highlighted the low density of PV-INs in associative cortices, including the perirhinal cortex (PER). However, a thorough characterization of the specificity and efficiency of the PV-IRES-Cre line in PER is still missing. PER is part of the parahippocampal network and represents a gateway of sensory information entering the lateral entorhinal cortex (LEC), and the hippocampus (Witter et al., 2000). PER has been shown to provide an inhibitory control on the information entering the LEC (de Curtis and Paré, 2004). The mechanism underlying this inhibitory control has been hypothesized to be mediated by PV-INs in the deep layers of PER through feedforward inhibition (Willems et al., 2018). This proposed role of PV-INs in gating information flow to the LEC seems to contrast with the low density of labeled cells in PER of the PV-IRES-Cre mouse line. In the current study we quantified the specificity and efficiency of the PV-IRES-Cre mouse line in PER. We report that the PV-IRES-Cre targets only a fraction of PV immunoreactive neurons in PER and other association areas. We suggest that these results point to a yet uncharacterized diversity within the PV-INs.
Materials and Methods
Animal Models
The research described here was performed on commercially available transgenic mice. We used the following mouse lines: PV-IRES-Cre [B6;129P2-PvalbTM 1(cre)Arbr/J; stock 008069, The Jackson Laboratory], Ai9 [Gt(ROSA)26SorRCL-tdT; stock 007909, The Jackson Laboratory], R26R-EYFP [129 × 1-Gt(ROSA)26SorTM 1(EYFP)Cos/J, stock 006148, The Jackson Laboratory]. The PV-IRES-Cre and Ai9 mice were bred as homozygotes, whereas the R26R-EYFP mice were bred as heterozygotes. To express fluorescent proteins in PV-INs we crossed PV-IRES-Cre mice with either Ai9 [4 mice, 2 males (M) and 2 females (F)] or R26R-EYFP mice (3 mice, 2 M and 1 F). Mice were group housed, with water and food ad libitum and a reverse dark/light cycle of 12 h/12 h. We used adult mice (>2 months) of both sexes in the current study.
AAV2/1-FLEX-mCherry Purification
First, pAAV-CMV-βglobin-intron-FLEX-MCS-WPRE was created by cloning a FLEX cassette with multicloning site into Cla1 and HindIII sites in pAAV-CMV-βglobin-intron-MCS-WPRE (Agilent). Sequence of the FLEX cassette was obtained from Atasoy et al. (2008). Subsequently, mCherry sequence was synthesized and cloned in the inverted orientation into EcoR1 and BamH1 sites in pAAV-CMV-βglobin-intron-FLEX-MCS-WPRE to create pAAV CMV-βglobin-intron-FLEX-mCherry-WPRE. The positive clones were confirmed by restriction digestion analyses and subsequently by DNA sequencing. Endotoxin free plasmid maxipreps (Qiagen) were used for AAV preparations. Approximately 7 × 106 AAV 293 cells (Agilent) were seeded in DMEM containing 10% fetal bovine serum (Thermo Fisher Scientific) and penicillin/streptomycin antibiotics into 150 mm cell culture plates. Next day, calcium chloride mediated cotransfection was done with 22.5 μg pAAV-containing the transgene, 22.5 μg pHelper, 11.3 μg pRC, 11.3 μg pXR1 (NGVB, IU, United States) capsid plasmids. The medium was replaced by fresh 10% FBS containing DMEM after 7 h. The cells were scrapped out after 72 h, then centrifuged at 200g and cell pellet was subjected to lysis using 150 mm NaCl-20 mm Tris pH 8:0 buffer containing 10% sodium deoxy cholate. The lysate was then treated with Benzonase nuclease HC (Millipore) for 45 min at 37°C. Benzonase treated lysate was centrifuged at 3,000 × g for 15 min and the clear supernatant was then subjected to HiTrap® Heparin High Performance (GE) affinity column chromatography using a peristaltic pump. The elute from the Heparin column was then concentrated using Amicon Ultra centrifugal filters (Millipore). Titering of viral stock was determined as approximately 1011 infectious particles/ML.
Injections of Adeno-Associated Viruses
We injected AAVs carrying a cre dependent mCherry construct in one male PV-IRES-Cre/R26REYFP intraparenchymally. The mouse was anesthetized with isoflurane (4%, Nycomed, airflow 1 l/min) in an induction chamber. After induction, the mouse was moved to a stereotactic apparatus and placed on a heating pad (37°C) throughout the procedure. Eye ointment was applied to protect the cornea. The following analgesic was applied subcutaneously: buprenorphine hydrochloride (0.1 mg/Kg, Temgesic, Invidior), meloxicam (1 mg/Kg, Metacam Boerringer Ingelheim Vetmedica), bupivacaine hydrochloride (1 mg/Kg at the injection site, Marcain, Astra Zeneca). The head was fixed with ear bars before the surgery. The skin above the skull was shaved and disinfected with Pyrisept. An incision was made to expose the skull at the selected location. The skull was thinned with a drill and a small hole was made with a glass pipette at the appropriate coordinates to target the PER (from bregma: AP −3 mm, ML 4.4 mm, DV 1.3 and 2 mm). Pressure injections were performed with glass pipettes attached to an injector (Nanoliter 2010, World Precision Instruments) controlled by a microsyringe pump controller (Micro4 pump, World Precision Instruments). Two injections (200 nl each) along the dorsoventral axis were performed at 50 nl/min. After retracting the pipette, the wound was rinsed with saline and sutured. The mouse could recover in a heated chamber before being returned to the home cage. The mouse received post operational analgesic 24 h after the surgery (meloxicam 1 mg/Kg). The survival time for transduction and expression of the virus was 15 days.
Histology
Mice were anaesthetized with isoflurane and euthanized with an injection of pentobarbital (i.p. 100 mg/Kg, Apotekerforeninger). The mice were subsequently perfused transcardially with Ringer’s solution (0.85% NaCl, 0.025% KCl, 0.02% NaHCO3) before perfusion with 4% paraformaldehyde (PFA) in PB (pH 7.4). After removing the brains from the skull, they were stored at 4°C in PFA for 2–3 h and then moved in a solution containing sucrose 15% in PB and kept at 4°C overnight. The brains were moved to sucrose 30% in PB for 2 days before being sectioned at the freezing microtome. We cut sections of 50 μm, collected in six, equally spaced series in a solution containing: 30% glycerol, 30% ethylene glycol, 40% PBS. The series were stored at -20°C until used for immunostainings.
Slices were washed in PB (3 × 10 min) before being incubated in blocking solution (0.1% Triton-100X, 10% NGS in PB) for 1 h. After blocking, the slices were moved to a new well and incubated for 2–3 days at 4°C in a solution with the primary antibodies (0.1% Triton-100X, 1% NGS in PB). After washing in PB (3 × 1 h), the slices were incubated at 4°C overnight with the secondary antibodies. The slices were washed in PB (3 × 10 min) and then mounted on SuperFrost slides (Thermo Fisher Scientific) in PB, and left to dry overnight at RT. Once dried, the slides were washed in xylene before being coversliped with entellan in xylene (Merck Chemicals, Darmstadt, Germany). We used the following primary antibodies: Guinea Pig IgG anti-NeuN (1:1,000, Sigma Millipore, #ABN90P), Rabbit anti-parvalbumin (1:1,000, Swant, #PV-27), Mouse IgG1 anti-parvalbumin (1:1,000, Sigma, #P3088), Rat IgG2a anti-RFP (1:1,000, Proteintech, #5f8), Chicken IgY anti-GFP (1:1,000, Abcam, #ab13970). We used the following secondary antibodies: Goat anti-guinea pig (IgG H + L) A647 (1:500, Invitrogen, #A-21450), Goat anti-Rat (IgG H + L) A-546 (1:500, Invitrogen, #A11081), Goat anti-Rabbit (IgG H + L) A488 (1:500, Invitrogen, #A11008), Goat anti-chicken (IgY H + L) A488 (1:500, Invitrogen, #A11039), Goat anti-rabbit (IgG H + L) A546 (1:500, Invitrogen, #A11010), Goat anti-rabbit (IgG H + L) A635 (1:500, Invitrogen, A31576).
Image Acquisition and Analysis
We used a confocal microscope (Zeiss LSM 880 AxioImager Z2) to image regions of interest (ROIs) to count labeled neurons. Images of PER, wS1, PrL, IL and LEC were taken with a x20 air objective with 1 Airy unit pinhole size and saved as czi files. We selected 3–5 ROIs containing PER starting from the most rostral part of PER and proceeding caudally. We included 2 ROIs containing wS1 at bregma and caudally to bregma. We included 2 ROIs containing IL and PrL between + 1.94 and + 1.54 mm from bregma. We included 3 ROIs from LEC between −2.9 and 3.5 mm from Bregma. ROIs contained the whole cortical area of interest. Files containing the ROIs were uploaded in Neurolucida (Micro Bright Field Bioscience) for analysis. We created contours to delineate the layers of the ROIs using the NeuN signal. The PER was divided in area 35 (A35) and area 36 (A36), and delineated according to Beaudin et al. (2013), LEC was delineated according to Ohara et al. (2021) and Witter (2011), PrL and IL were delineated according to Van De Werd et al. (2010). Markers for each signal were used to count cells labeled by different markers. We counted all cells contained within the contours. Since we counted slices distanced 300 μm from each other, overcounting in the z axis is not relevant and correction was not applied. Quantifications were done in Neurolucida Explorer (Micro Bright Field Bioscience) and exported in excel files. The density of neurons labeled by a marker was measured as the number of labeled neurons in a contour divided by the area of the contour (cells/mm2). The specificity measures the percentage of cells labeled by the mouse line that co-express PV and was measured dividing the number of RFP-expressing neurons in Ai9 or GFP-expressing neurons in R26REYFP co-expressing PV by the number of RFP-expressing neurons or GFP-expressing in R26REYFP. The efficiency measures the percentage of PV-expressing neurons labeled by the mouse line and was calculated as the number of RFP-expressing neurons in Ai9 (or GFP-expressing in R26REYFP) co-expressing PV divided by the number of PV-expressing neurons. We quantified between 2 and 5 sections per cortical region. Values are reported as mean ± standard deviation). All graphs were created in excel, images of ROIs were created in Zen lite (Zeiss) and figures in illustrator.
Data for the comparison of the PV-IRES-Cre and PV-T2A-Cre were obtained from the Allen Institute website: Mouse Brain Connectivity Atlas, transgenic characterization. Samples for the PV-IRES-Cre: 100141217 Cre ISH, 81709692 tdTomato ISH, 81657984 FISH. Samples for the PV-T2A-Cre: 100141203 Cre ISH, 81658019 tdTomato ISH, 81811663 FISH.
Results
We used immunofluorescence for PV on tissue from mice expressing tdTomato in PV-INs to address the specificity and efficacy of the PV-IRES-Cre mouse line in the mouse PER. The distribution of tdTomato across the cortex recapitulates the pattern of PV expression: (i) higher expression in thalamo recipient layers (4 and 5B) in sensory cortex; (ii) a bias toward deep layers throughout the cortex; (iii) higher density in dorsal cortex than in ventral cortical areas (Figure 1; Tremblay et al., 2016).
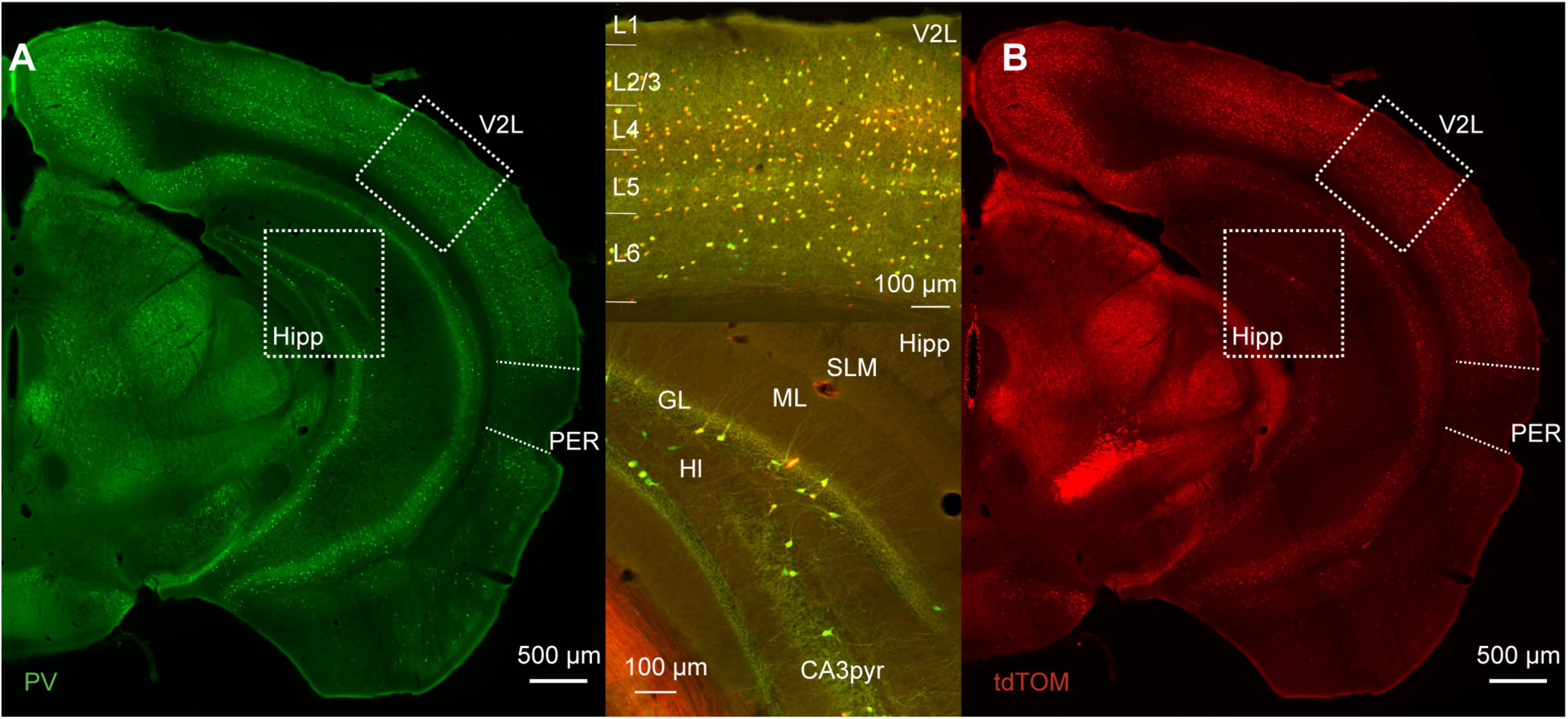
Figure 1. (A) Representative immunofluorescence staining for PV of a slice at about bregma −3 mm of a PVcre/Ai9 mouse. Dotted boxes highlight the lateral part of the secondary visual cortex (V2L) and the hippocampus (Hipp). The perirhinal cortex (PER) is delineated by the dotted line on the lateral side of the brain. (B) Representative image of the immunofluorescence for tdTomato in the same section as (A). Upper insert shows the merged signals in the cortex, and lower insert shows merged signals in the hippocampus (SLM, stratum lacunosum-moleculare; ML, molecular layer; GL, granule layer of the dentate gyrus; Hl, hilus; CA3pyr, pyramidal layer of the CA3 region).
We measured the density of PV-immunoreactive neurons (PV-IR) and tdTomato expressing neurons across the layers of PER. The pattern of tdTomato expression showed a bias toward the deep layers similarly to that of PV-IR neurons (Figures 2A,B,D,E). However, the density of tdTomato expressing neurons was lower than that of PV-IR neurons (tdTomato: 122.56 ± 31.3 cells/mm2; PV-IR: 274.68 ± 10.9 cells/mm2, n = 4 mice) (Figures 2A,B,D,E). Indeed, several PV-IR cells did not express tdTomato in the PV-IRES-Cre mouse line (Figure 2C). The density of PV-IR neurons not expressing tdTomato was higher than that of tdTomato expressing neurons (Figure 2F). This result suggests that the PV-IRES-Cre mouse line is very inefficient in labeling PV expressing GABAergic neurons in the mouse PER.
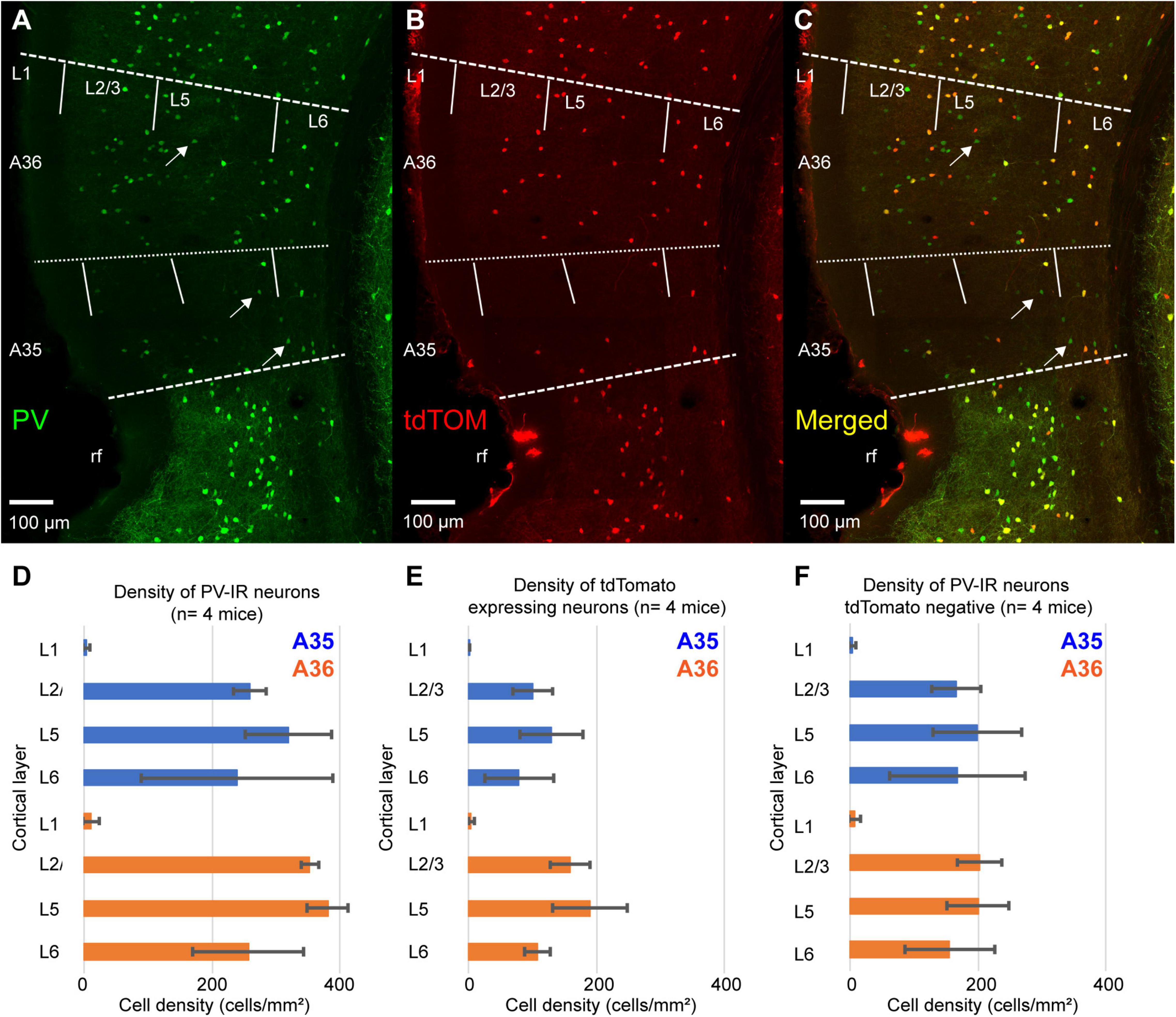
Figure 2. (A–C) Representative confocal stacks showing immunofluorescence for PV (A), tdTomato (B) and the overlap of the two signals (C) in PER of a PV-IRES-Cre mouse. Arrows indicate PV-IR neurons that do not express tdTomato. rf, rhinal fissure. (D) Bar graph showing the density of PV-IR neurons across the layers for A35 and A36. (E) Bar graph showing the density of tdTomato-expressing neurons across the layers of A35 and A36. (F) Bar graph showing the density of PV-IR that do not express tdTomato across the layers of A35 and A36.
We tested this hypothesis by quantifying the specificity (i.e., the percentage of tdTomato expressing neurons that also express PV) and the efficiency (i.e., the percent of PV-IR neurons labeled by tdTomato) of the PV-IRES-Cre mouse line in PER (Figure 3). For these measurements we used 4 PV-Cre/Ai9 (2 M, 2 F), and counted a total of 2985 PV-IR neurons, 1411 tdTomato expressing neurons and 1334 double labeled neurons. The majority of tdTomato expressing neurons was PV-IR, suggesting that the mouse line shows a high specificity for PV expressing neurons (n = 4 mice, specificity = 94.6 ± 2.7%) (Figures 3A,B). However, we found that less than 50% of PV-IR neurons throughout the layers of PER were labeled by tdTomato (n = 4 mice, efficiency = 44.1 ± 10%) (Figures 3A,C). These data demonstrate that the PV-IRES-Cre mouse line shows a low efficiency in labeling PV expressing neurons in PER. We confirmed our results using a different primary antibody against PV (mouse anti-PV, 2 mice, 1M and 1F) (Supplementary Figure 1). Moreover, the transgenic characterization of the Allen Institute for Brain Science demonstrates by fluorescent in situ hybridization (FISH) that many neurons expressing PV do not express tdTomato (Supplementary Figure 2).
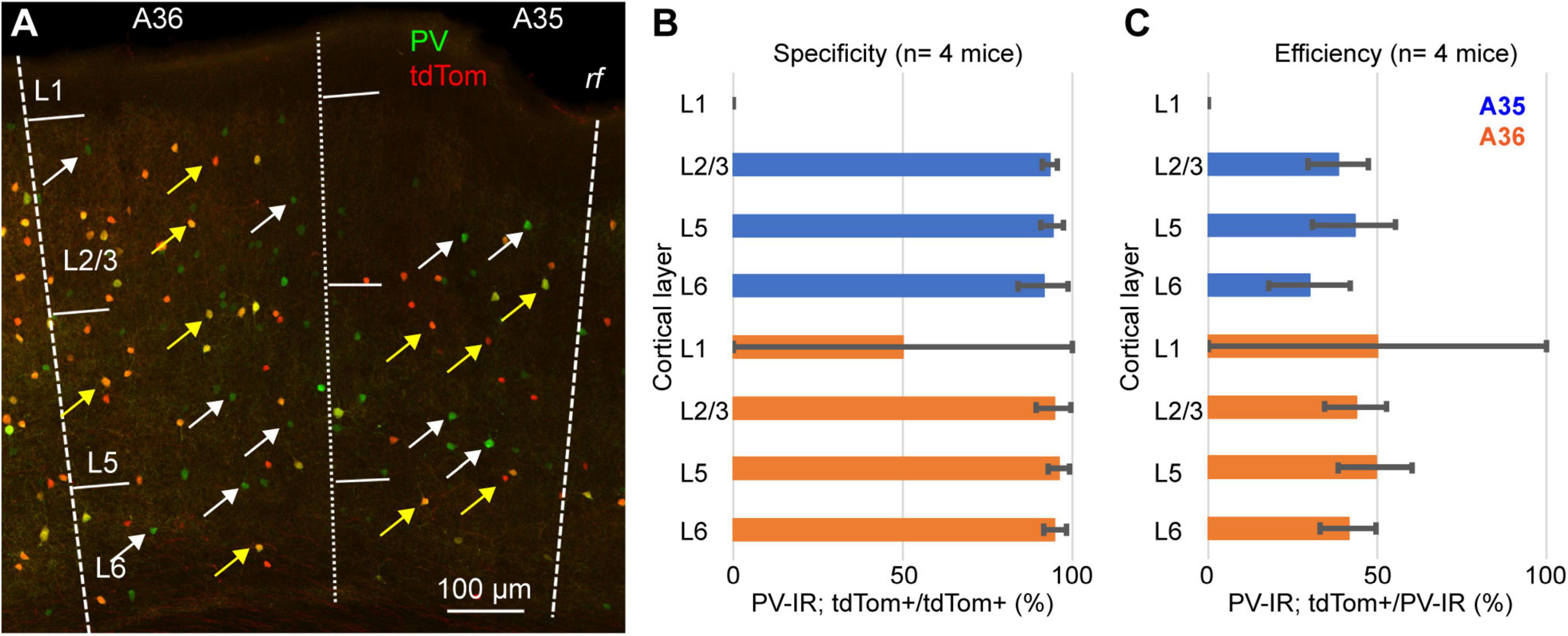
Figure 3. (A) Representative confocal stack showing the overlap of immunofluorescence for PV (green) and tdTomato (red) in PER; yellow arrows indicate double labeled cells and white arrows indicate PV-IR cell that do not express tdTomato. (B) Bar graph showing the specificity of the PV-IRES-Cre mouse line in PER in 4 animals. (C) Bar graph showing the efficacy of the PV-IRES-Cre mouse line in PER in 4 animals.
As an internal control, we quantified the specificity and efficiency of the PV-IRES-Cre line in the barrel cortex. For these quantifications we used the same tissue as for the measurements in PER (PV-Cre/Ai9, 3 mice, 2M, 1F). We counted a total of 1716 PV-IR neurons, 1805 tdTomato-expressing neurons, and 1613 double labeled neurons). As previously reported for the visual cortex (Atallah et al., 2012), the PV-IRES-Cre was very specific and efficient in labeling PV expressing neurons in the barrel cortex (Figure 4). We found that 89.64 ± 7.5% of tdTomato expressing neurons also expressed PV, and 93.97 ± 0.7% of PV expressing neurons was labeled by tdTomato. These results suggest that our measurements are not a consequence of our histological procedures and are consistent with results obtained by FISH.
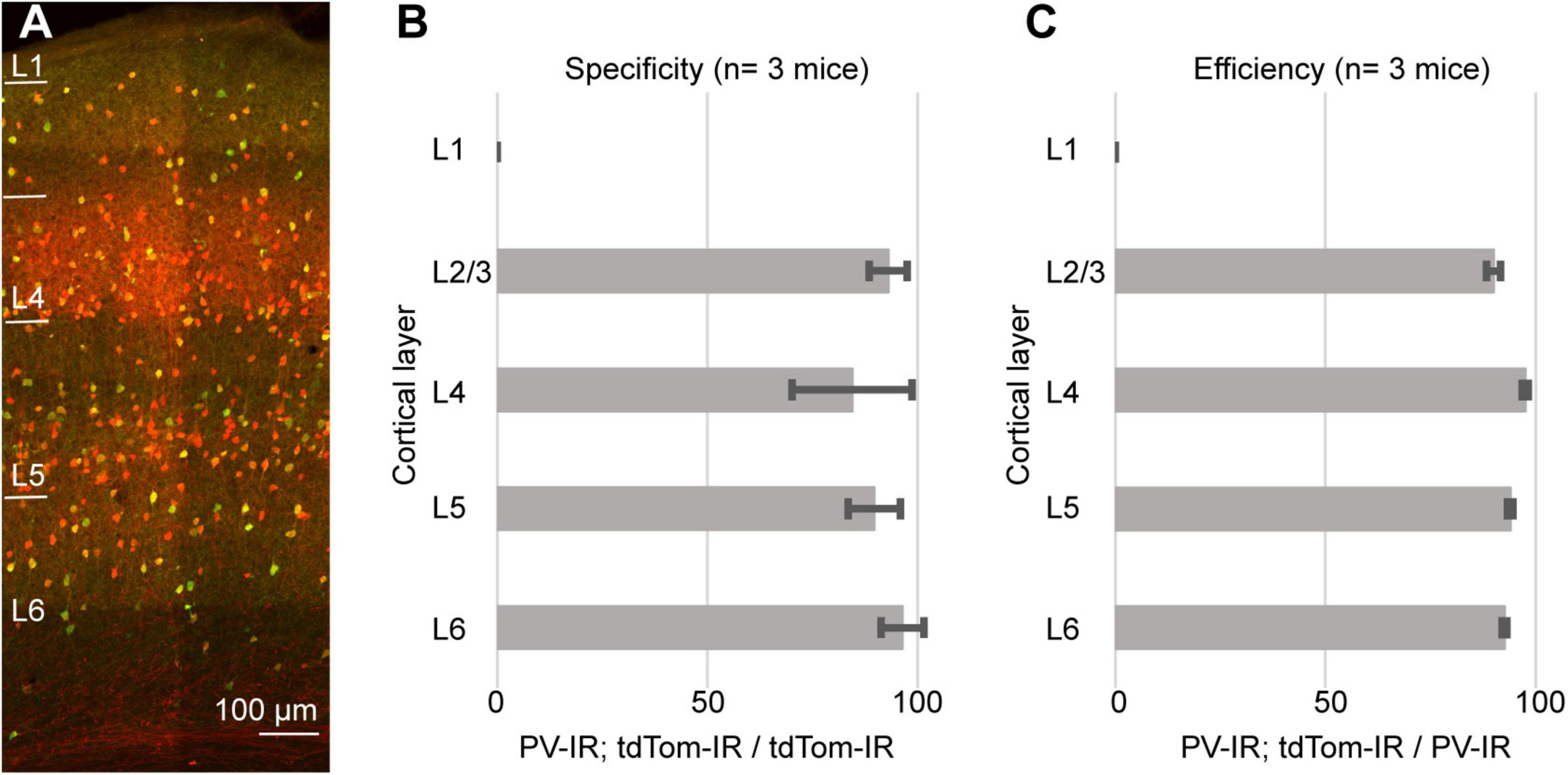
Figure 4. (A) Representative confocal stack showing the overlap of immunofluorescence for PV (green) and tdTomato (Red) in barrel cortex. (B) Bar graph showing the specificity of the PV-IRES-Cre mouse line in barrel cortex in 3 animals. (C) Bar graph showing the efficiency of the PV-IRES-Cre mouse line in barrel cortex in 3 animals.
Our results can be explained either assuming that a population of PV-expressing interneurons does not express Cre-recombinase, or that the Ai9 reporter line is not reliable in PER. To test the reliability of the reporter line, we crossed the PV-IRES-Cre line with the Rosa26-stop-EYFP and measured the specificity and efficiency (n = 2 mice, 2M). In these experiments we counted 1461 PV-IR neurons, 743 tdTomato-expressing neurons, and 719 double labeled neurons. Similar to our results with the Ai9 reporter line, the specificity was very high (96.85 ± 1%, n = 2 mice) but only 49.15 ± 6.35% of PV-IR neurons was labeled by YFP in PER (Figure 5). Since both reporter lines exploit the Rosa26 locus, we hypothesized that this locus might be selectively silenced in a subpopulation of PV-expressing neurons in PER. We tested this hypothesis by injecting a Cre dependent virus AAV-FLEX-mCherry in PER, reasoning that if the hypothesis was correct then we would find PV-expressing neurons that also expressed mCherry but not YFP. However, we found that almost all mCherry expressing neurons also expressed YFP (94.79%, n = 1 mouse, 1M, 96 mCherry cells), suggesting that the low efficiency resides in the PV-IRES-Cre mouse line rather than in the reporter line (Figure 6). The low efficiency of the PV-IRES-Cre was found in both sexes: 43.2 ± 7.2% in males (n = 2 mice), and 51.1 ± 9.6% in females (n = 2 mice).
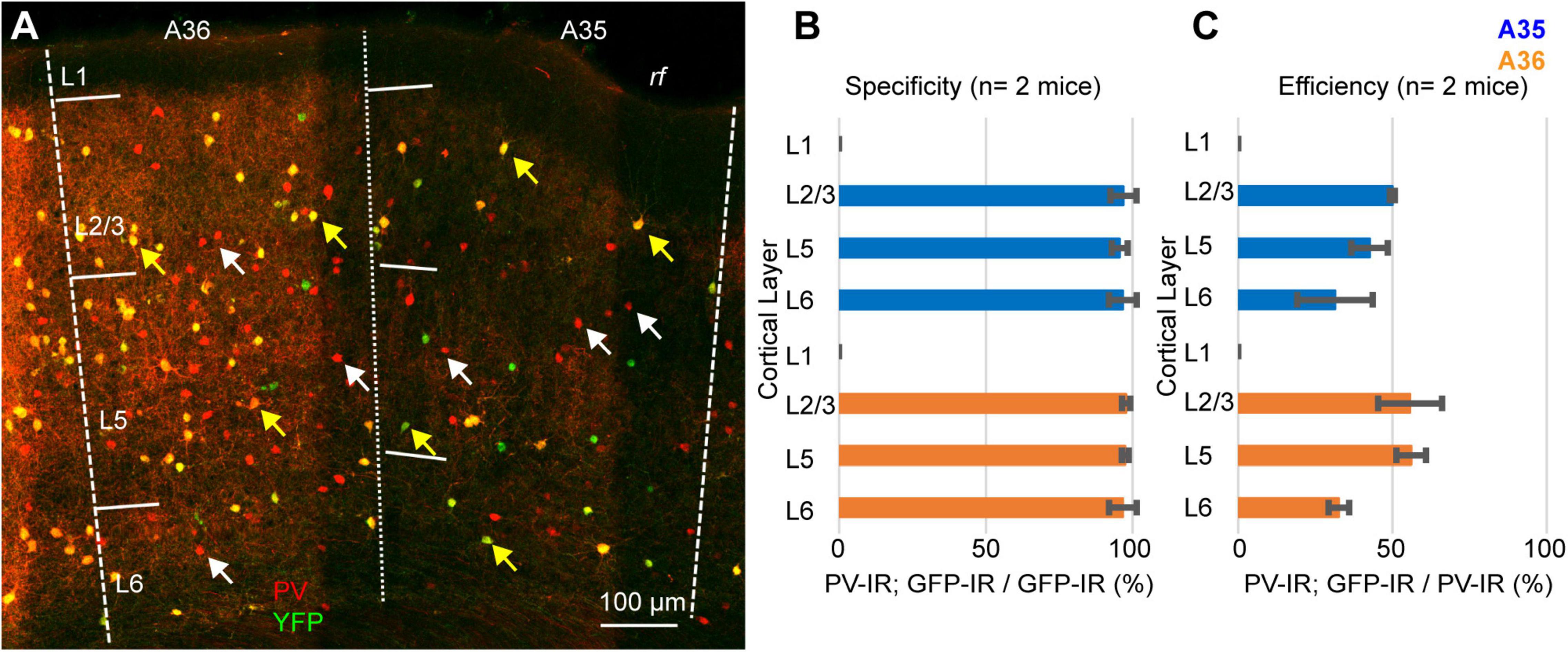
Figure 5. (A) Representative confocal stack showing the overlap of immunofluorescence for PV (red) and YFP (green) in PER. Yellow arrows indicate double-IR neurons, white arrows indicate PV-IR neurons not labeled with YFP. (B) Bar graph showing the specificity of the mouse line in 2 animals. (C) Bar graph showing the efficiency of the mouse line in 2 animals.
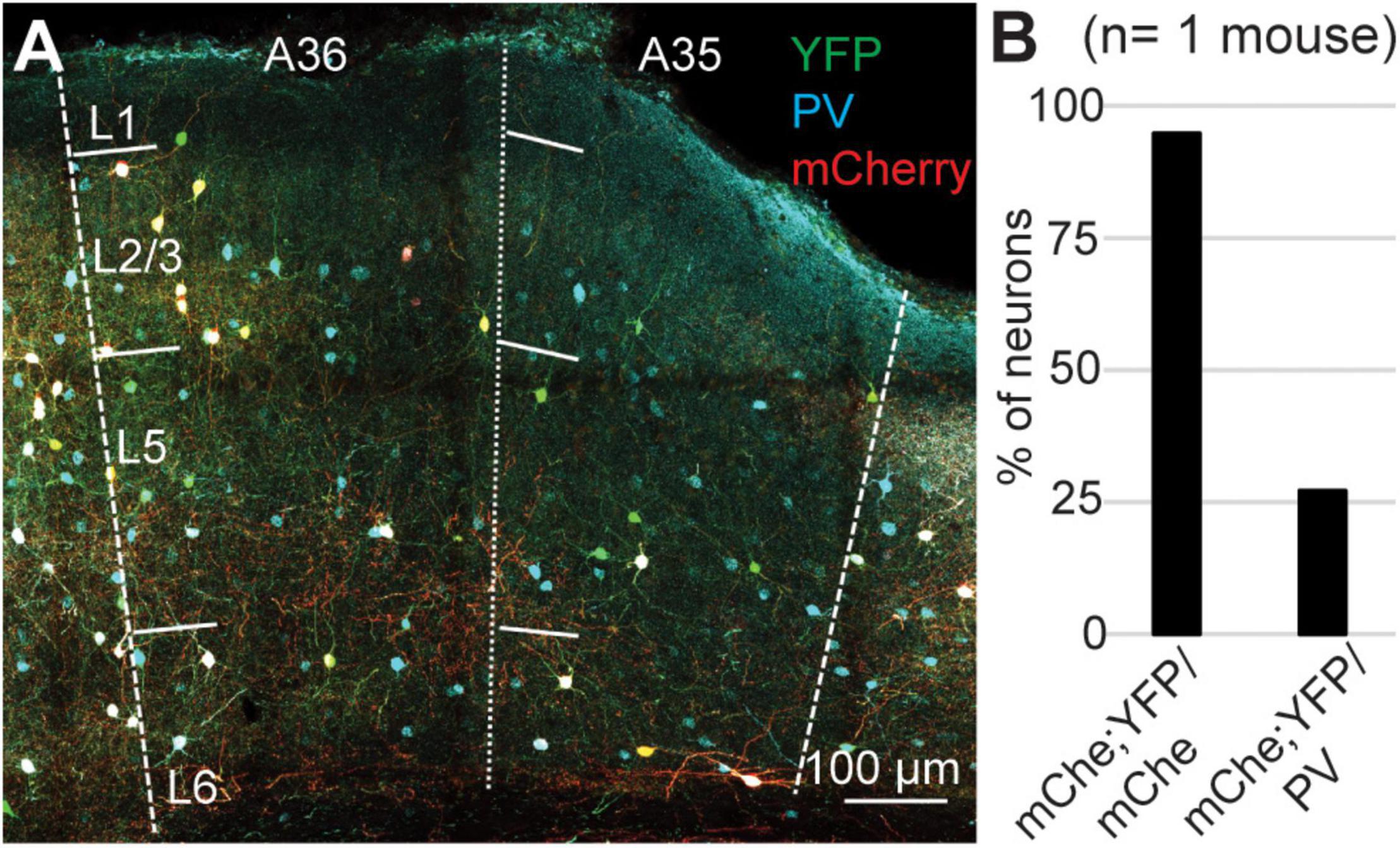
Figure 6. (A) Representative confocal stack showing the overlap of PV (turquoise), YFP (green) and mCherry (red) in PER. (B) Bar plot showing the percent of mCherry-expressing neurons that also expressed YFP, and the percent of PV-expressing neurons labeled by mCherry and YFP.
A recent report has demonstrated that the density of PV neurons is lower in associative areas of the cortex as compared to sensory-motor regions (Kim et al., 2017). These findings suggest that the low efficiency of the PV-IRES-Cre mouse line might be a shared feature of associative areas. To test this hypothesis, we measured the specificity and efficiency of the PV-IRES-Cre mouse line in three associative areas shown to have low density of PV neurons: prelimbic cortex (PrL), infralimbic cortex (IL) and LEC (Figure 7). As reported for other cortical regions, the specificity of the PV-IRES-Cre mouse line was very high in all three areas (Figures 7C,D), with the exception of L2 of IL where about 50% of tdTomato labeled neurons did not express PV (Figures 7A1–3,C). The efficiency of the PV-IRES-Cre mouse line was lower than in the barrel cortex and similar to that reported in the PER (Figures 3,7). These results suggest that the low efficiency of the PV-IRES-Cre mouse line is a phenomenon common to association cortices. The PV-IRES-Cre is the most widely used mouse line to label PV-INs, however, another mouse line has been generated by the Allen Institute, the PV-T2A-Cre (Madisen et al., 2010). The PV-T2A-Cre line has been used to quantify PV-INs in the whole brain (Kim et al., 2017). These authors reported a lower density of PV-INs in PER as compared to sensorimotor areas. A qualitative comparison of the two mouse lines using available in situ hybridization data from the Allen Institute revealed a low number of Cre-expressing neurons in both lines (Supplementary Figure 2). The expression of td-Tomato was higher in the PV-T2A-Cre than in the PV-IRES-Cre, and several td-Tomato cells do not seem to express PV (Supplementary Figure 2). The comparison of the two mouse lines suggests that the number of Cre expressing neurons is low in both, and further examination of the expression of PV, Cre, and td-Tomato is necessary in the PV-T2A-Cre.
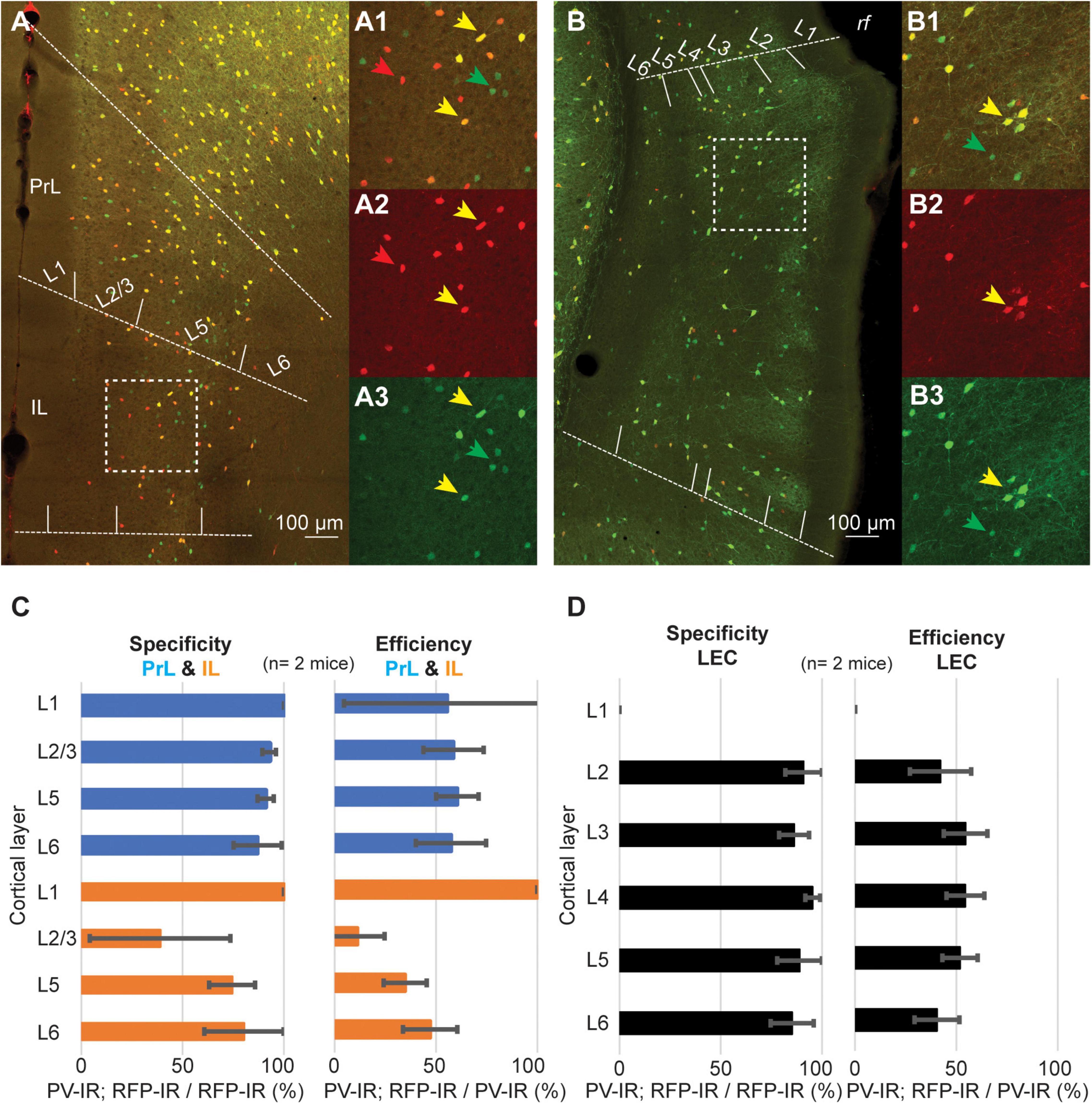
Figure 7. Representative confocal stack showing the overlap of PV (green) and tdTomato (red) in PrL and IL cortex (A), and in LEC (B). Inserts show magnification of the areas highlighted by dotted squares in (A,B) (A1–3, IL; B1–3, LEC). (C) Bar graphs showing the specificity and efficiency of the PV-IRES-Cre mouse line in PrL and IL. (D) Bar graphs showing the specificity and efficiency of the PV-IRES-Cre mouse line in LEC.
Discussion
We demonstrate here that the PV-IRES-Cre mouse line has a low efficiency in labeling PV-INs in PER and this phenomenon might be a common feature of associative areas of the cerebral cortex. The PV-IRES-Cre mouse line has become an invaluable tool for studying fast-spiking basket cells across the telencephalon (Atallah et al., 2012; Straub et al., 2016; Yu et al., 2019). This mouse line has recently been used to estimate the number of PV-expressing fast-spiking basket cells in the cortex and hippocampus (Whissell et al., 2015). Our findings suggest that the density of PV-INs obtained by counting fate-mapped neurons in the PV-IRES-Cre mouse line is dramatically underestimated.
A recent study quantified labeled neurons in the PV-2A-Cre mouse line and showed that association areas of the cortex, including PER, contain a low density of PV-INs (Kim et al., 2017). The PV-IRES-Cre and the PV-T2A-Cre differ in regulatory elements, including the elements for multicistronic expression: IRES vs. 2A (Madisen et al., 2010). A qualitative examination of the characterization performed by the Allen Institute showed that the size of the labeled population is similar in the two mouse lines, suggesting that the low efficiency might be due to a difference within the population of PV-INs in PER. Interestingly, similar results to ours have been shown in the retina of PV-IRES-Cre mice (Gábriel et al., 2016). The retina of PV-IRES-Cre/YFP mice showed labeling in glial cells, but not in cell-types expressing PV (Gábriel et al., 2016).
PV-expressing fast-spiking neurons have also been implicated in controlling information flow along the cortico-hippocampal network. These neurons strongly respond to cortical inputs and provide feedforward inhibition to excitatory neurons in PER, controlling their output to LEC (Willems et al., 2018). Because of the high specificity of the PV-IRES-Cre line in PER, we suggest that the PV-IRES-Cre mouse line can be used to study the cellular properties of fast-spiking basket cells in PER and their connectivity through paired recordings or circuits-mapping with soma-targeted optogenetics (Baker et al., 2016; Willems et al., 2018). However, the low efficiency of the PV-IRES-Cre mouse line might impair the optogenetic assessment of the circuits underlying the inhibitory control exerted by PER in the cortico-hippocampal network (e.g., using full-field light stimulation) (de Curtis and Paré, 2004).
Our work raises two questions: (i) what are the mechanisms by which cells expressing PV do not express Cre?; (ii) do labeled and unlabeled PV-INs belong to different subpopulations? The answer to the first question will require additional experiments to assess the activity of the locus containing Cre in labeled and unlabeled neurons in PER. One possible scenario is that in PER a population of PV-INs expresses very low levels of Cre recombinase. This might lead to non-parallel recombination of the lox-P sites and to absence of expression of the reporter (Liu et al., 2013). The second question concerns with the biological implications of our findings in the context of neuronal diversity. Patch clamp recordings of PV-INs in the PER of PV-IRES-Cre mice show that these neurons express a fast-spiking phenotype (Willems et al., 2018). However, there is currently no information of the firing pattern of PV-INs not labeled by the mouse line. Assessing this lack of information requires a way to label either all GABAergic neurons or the entire PV-expressing population. One possibility is a comprehensive analysis of the firing patterns and molecular identity of GABAergic neurons in the GAD1-Cre mouse or in the GAD67-EGFP mouse. A transcriptomic analysis of the same mouse lines might further dissect subpopulations of the PV-INs in the PER.
The current observations do emphasize the importance of careful characterization of the tools used to study cell-types across cortical regions. Cortical regions might contain specialized populations not targeted by current transgenic or viral strategies or these strategies might show region specific bias in the labeled population.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation. Data for Supplementary figure 1: ©2011 Allen Institute for Brain Science. Allen Mouse Brain Connectivity Atlas. Available from: https://connectivity.brain-map.org/.
Ethics Statement
The animal study was reviewed and approved by the Forsøksdyrforvaltningens tilsyns- og søknadssystem (ID 22312).
Author Contributions
MN, MW, and HK conceived the research. MN, HK, and KK performed the experiments and analyzed the data. RN contributed viral vectors. MN wrote the manuscript with feedback from all authors.
Funding
This study has been supported by the Center of Excellence scheme of the Research Council of Norway—Center for Neural Computation (Grant No. 223262), and the Kavli Foundation. This project has received funding from the European Union’s Horizon 2020 Research and Innovation Programme under the Marie Skłodowska-Curie (Grant No. 885955 to MN).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fncir.2021.781928/full#supplementary-material
Supplementary Figure 1 | (A–C) Representative immunostainings showing the expression of tdTomato (A, red), PV (B, green) and the merged image (C) in the perirhinal cortex of the PV-IRES-Cre line using a primary mouse anti-PV antibody. (D) Bar plot showing the densities of PV immunoreactive (PV-IR) neurons (green) and tdTomato expressing neurons (red). (E) Bar plot showing the specificity obtained with the mouse anti-PV. (F) Efficiency obtained with the mouse anti-PV. © 2011 Allen Institute for Brain Science. Allen Mouse Brain Connectivity Atlas. Available from: https://connectivity.brain-map.org/.
Supplementary Figure 2 | Comparison of the PV-IRES-Cre and PV-T2A-Cre mouse lines in the PER. (A) PER of a PV-IRES-Cre mouse showing ISH for Cre. (B) PER of a PV-T2A-Cre mouse showing ISH for Cre. (C) PER of a PV-IRES-Cre mouse showing ISH for td-Tomato. (D) PER of a PV-T2A-Cre mouse showing ISH for tdTomato. (E–J) FISH for PV and td-Tomato in PV-IRES-Cre (E, merged; G, PV; I, td-Tomato), and in PV-T2A-Cre (F, merged; H, PV; J, td-Tomato). Green arrows point to PV-expressing neurons that do not express tdTomato. Red arrows point to td-Tomato neurons that do not express PV. LEC, lateral entorhinal cortex.
References
Atallah, B. V., Bruns, W., Carandini, M., and Scanziani, M. (2012). Parvalbumin-expressing interneurons linearly transform cortical responses to visual stimuli. Neuron 73, 159–170. doi: 10.1016/j.neuron.2011.12.013
Atasoy, D., Aponte, Y., Su, H. H., and Sternson, S. M. (2008). A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping. J. Neurosci. 28, 7025–7030. doi: 10.1523/JNEUROSCI.1954-08.2008
Baker, C. A., Elyada, Y. M., Parra, A., and Bolton, M. M. (2016). Cellular resolution circuit mapping with temporal-focused excitation of soma-targeted channelrhodopsin. Elife 5:e14193. doi: 10.7554/eLife.14193
Beaudin, S. A., Singh, T., Agster, K. L., and Burwell, R. D. (2013). Borders and comparative cytoarchitecture of the perirhinal and postrhinal cortices in an F1 hybrid mouse. Cereb. Cortex 23, 460–476. doi: 10.1093/cercor/bhs038
de Curtis, M., and Paré, D. (2004). The rhinal cortices: a wall of inhibition between the neocortex and the hippocampus. Prog. Neurobiol. 74, 101–110. doi: 10.1016/j.pneurobio.2004.08.005
Gábriel, R., Erdélyi, F., Szabó, G., Lawrence, J. J., and Wilhelm, M. (2016). Ectopic transgene expression in the retina of four transgenic mouse lines. Brain Struct. Funct. 221, 3729–3741. doi: 10.1007/s00429-015-1128-2
Harris, K. D., and Shepherd, G. M. (2015). The neocortical circuit: themes and variations. Nat. Neurosci. 18, 170–181.
Hippenmeyer, S., Vrieseling, E., Sigrist, M., Portmann, T., Laengle, C., Ladle, D. R., et al. (2005). A developmental switch in the response of DRG neurons to ETS transcription factor signaling. PLoS Biol. 3:e159. doi: 10.1371/journal.pbio.0030159
Hu, H., Gan, J., and Jonas, P. (2014). Fast-spiking, parvalbumin+ GABAergic interneurons: from cellular design to microcircuit function. Science 345:1255263.
Kepecs, A., and Fishell, G. (2014). Interneuron cell types are fit to function. Nature 505, 318–326. doi: 10.1038/nature12983
Kim, Y., Yang, G. R., Pradhan, K., Venkataraju, K. U., Bota, M., Del Molino, L. C. G., et al. (2017). Brain-wide maps reveal stereotyped cell-type-based cortical architecture and subcortical sexual dimorphism. Cell 171, 456.–469. doi: 10.1016/j.cell.2017.09.020
Liu, J., Willet, S. G., Bankaitis, E. D., Xu, Y., Wright, C., and Gu, G. (2013). Non-parallel recombination limits Cre-LoxP-based reporters as precise indicators of conditional genetics manipulation. Genesis 51, 436–442. doi: 10.1002/dvg.22384
Madisen, L., Zwingman, T. A., Sunkin, S. M., Oh, S. W., Zariwala, H. A., Gu, H., et al. (2010). A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 13, 133–140. doi: 10.1038/nn.2467
Martínez, J. J., Rahsepar, B., and White, J. A. (2017). Anatomical and electrophysiological clustering of superficial medial entorhinal cortex interneurons. eNeuro 4:ENEURO.0263-16.2017. doi: 10.1523/ENEURO.0263-16.2017
Ohara, S., Blankvoort, S., Nair, R. R., Nigro, M. J., Nilssen, E. S., Kentros, C., et al. (2021). Local projections of layer Vb-to-Va are more prominent in lateral than in medial entorhinal cortex. Elife 10:e67262. doi: 10.7554/eLife.67262
Straub, C., Saulnier, J. L., Bègue, A., Feng, D. D., Huang, K. W., and Sabatini, B. L. (2016). Principles of synaptic organization of GABAergic interneurons in the striatum. Neuron 92, 84–92. doi: 10.1016/j.neuron.2016.09.007
Taniguchi, H., He, M., Wu, P., Kim, S., Paik, R., Sugino, K., et al. (2011). A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013. doi: 10.1016/j.neuron.2011.07.026
Tremblay, R., Lee, S., and Rudy, B. (2016). GABAergic interneurons in the neocortex: from cellular properties to circuits. Neuron 91, 260–292. doi: 10.1016/j.neuron.2016.06.033
Van De Werd, H., Rajkowska, G., Evers, P., and Uylings, H. B. (2010). Cytoarchitectonic and chemoarchitectonic characterization of the prefrontal cortical areas in the mouse. Brain Struct. Funct. 214, 339–353. doi: 10.1007/s00429-010-0247-z
Whissell, P. D., Cajanding, J. D., Fogel, N., and Kim, J. C. (2015). Comparative density of CCK-and PV-GABA cells within the cortex and hippocampus. Front. Neuroanat. 9:124. doi: 10.3389/fnana.2015.00124
Willems, J. G., Wadman, W. J., and Cappaert, N. L. (2018). Parvalbumin interneuron mediated feedforward inhibition controls signal output in the deep layers of the perirhinal-entorhinal cortex. Hippocampus 28, 281–296. doi: 10.1002/hipo.22830
Witter, M. (2011). “The hippocampus,” in The mouse Nervous System (First Edition), eds G. Paxinos, L. Puilles, and C. Watson (Cambridge, MA: Academic Press), 112–139.
Witter, M. P., Wouterlood, F. G., Naber, P. A., and Van Haeften, T. (2000). Anatomical organization of the parahippocampal-hippocampal network. Ann. NY Acad. Sci. 911, 1–24. doi: 10.1111/j.1749-6632.2000.tb06716.x
Keywords: parvalbumin interneurons, PV-IRES-Cre, interneuron diversity, perirhinal cortex, mouse line
Citation: Nigro MJ, Kirikae H, Kjelsberg K, Nair RR and Witter MP (2021) Not All That Is Gold Glitters: PV-IRES-Cre Mouse Line Shows Low Efficiency of Labeling of Parvalbumin Interneurons in the Perirhinal Cortex. Front. Neural Circuits 15:781928. doi: 10.3389/fncir.2021.781928
Received: 23 September 2021; Accepted: 18 October 2021;
Published: 08 November 2021.
Edited by:
Blake A. Richards, Montreal Institute for Learning Algorithm (MILA), CanadaReviewed by:
Michael M. Kohl, University of Glasgow, United KingdomHiroyuki Hioki, Juntendo University, Japan
Copyright © 2021 Nigro, Kirikae, Kjelsberg, Nair and Witter. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maximiliano José Nigro, TWF4aW1pbGlhbm8uai5uaWdyb0BudG51Lm5v