
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Neural Circuits , 21 December 2018
Volume 12 - 2018 | https://doi.org/10.3389/fncir.2018.00106
This article is part of the Research Topic Neuronal Co-transmission View all 11 articles
 Tomas Hökfelt1*
Tomas Hökfelt1* Swapnali Barde1
Swapnali Barde1 Zhi-Qing David Xu1,2
Zhi-Qing David Xu1,2 Eugenia Kuteeva1†
Eugenia Kuteeva1† Joelle Rüegg3,4,5
Joelle Rüegg3,4,5 Erwan Le Maitre1†
Erwan Le Maitre1† Mårten Risling1Jan Kehr6,7
Mårten Risling1Jan Kehr6,7 Robert Ihnatko8,9
Robert Ihnatko8,9 Elvar Theodorsson8,9
Elvar Theodorsson8,9 Miklos Palkovits10
Miklos Palkovits10 William Deakin11
William Deakin11 Gyorgy Bagdy12,13,14
Gyorgy Bagdy12,13,14 Gabriella Juhasz11,12,15
Gabriella Juhasz11,12,15 H. Josée Prud’homme16
H. Josée Prud’homme16 Naguib Mechawar16,17
Naguib Mechawar16,17 Rochellys Diaz-Heijtz1
Rochellys Diaz-Heijtz1 Sven Ove Ögren1
Sven Ove Ögren1Neuropeptides are auxiliary messenger molecules that always co-exist in nerve cells with one or more small molecule (classic) neurotransmitters. Neuropeptides act both as transmitters and trophic factors, and play a role particularly when the nervous system is challenged, as by injury, pain or stress. Here neuropeptides and coexistence in mammals are reviewed, but with special focus on the 29/30 amino acid galanin and its three receptors GalR1, -R2 and -R3. In particular, galanin’s role as a co-transmitter in both rodent and human noradrenergic locus coeruleus (LC) neurons is addressed. Extensive experimental animal data strongly suggest a role for the galanin system in depression–like behavior. The translational potential of these results was tested by studying the galanin system in postmortem human brains, first in normal brains, and then in a comparison of five regions of brains obtained from depressed people who committed suicide, and from matched controls. The distribution of galanin and the four galanin system transcripts in the normal human brain was determined, and selective and parallel changes in levels of transcripts and DNA methylation for galanin and its three receptors were assessed in depressed patients who committed suicide: upregulation of transcripts, e.g., for galanin and GalR3 in LC, paralleled by a decrease in DNA methylation, suggesting involvement of epigenetic mechanisms. It is hypothesized that, when exposed to severe stress, the noradrenergic LC neurons fire in bursts and release galanin from their soma/dendrites. Galanin then acts on somato-dendritic, inhibitory galanin autoreceptors, opening potassium channels and inhibiting firing. The purpose of these autoreceptors is to act as a ‘brake’ to prevent overexcitation, a brake that is also part of resilience to stress that protects against depression. Depression then arises when the inhibition is too strong and long lasting – a maladaption, allostatic load, leading to depletion of NA levels in the forebrain. It is suggested that disinhibition by a galanin antagonist may have antidepressant activity by restoring forebrain NA levels. A role of galanin in depression is also supported by a recent candidate gene study, showing that variants in genes for galanin and its three receptors confer increased risk of depression and anxiety in people who experienced childhood adversity or recent negative life events. In summary, galanin, a neuropeptide coexisting in LC neurons, may participate in the mechanism underlying resilience against a serious and common disorder, MDD. Existing and further results may lead to an increased understanding of how this illness develops, which in turn could provide a basis for its treatment.
The first evidence for chemical signaling in the central nervous system was reported by Eccles et al. (1954), when they demonstrated that acetylcholine is the transmitter released from motor neuron collaterals onto Renshaw cells in the spinal cord. Some 10 years later the Canadian electrophysiologist Hugh McLennan in his monograph “Synaptic transmission” (McLennan, 1963) reviewed in some detail the evidence for a number of molecules being transmitters: “Acetylcholine,” “Catecholamines,” “5-Hydroxytryptamine,” “Substance P,” “Factor I and the Inhibitory Transmitter,” “GABA and Glutamic Acid,” and “Cerebellar Excitatory Factor” were the chapter sub-headings. Some further compounds were mentioned, like other amino acids. A detailed table of the regional distribution of these molecules was included. In the “Conclusions” McLennan stated “With the exception of a number of cholinergic and rather fewer adrenergic systems, the data supporting a certain type of chemical mediation in any given situation are quite inadequate, and in spite of the inherent difficulties the number of problems to be solved are of great interest.” Indeed, many efforts in the following years rapidly expanded the number of candidates and ‘certified’ their transmitter status – work still ongoing. However, to identify a molecule as a transmitter was at that time often a difficult process with strong pro and contra arguments. More recently completely different molecules have appeared on the scene, not stored in vesicles and thus not exocytosed, like nitric oxide (NO) and hydrogen sulfide (H2S), sometimes called “gasotransmitters” (Paul and Snyder, 2015). Subsequently, substance P, mentioned already by McLennan, was identified as a member of the by far most diverse group of signaling molecules (>100) in the nervous system, the neuropeptides (Burbach, 2010).
The purpose of the present article is to review data on one of these peptides, galanin, which was discovered by Tatemoto et al. (1983) at Karolinska Institutet, a peptide that is a co-transmitter in many systems. In particular, focus is on recent results describing the distribution of galanin and it three receptors GalR1-3 in the ‘normal’ human brain by studying post mortem tissue samples (Le Maitre et al., 2013). More importantly, results are discussed showing significant changes in expression of the galanin family ‘members’ in post mortem brains from depressed patients having committed suicide, as compared to controls (Barde et al., 2016). A hypothesis is presented on a possible role of galanin, coexisting in noradrenergic neurons in the locus coeruleus (LC), in the development of depression and in resilience. This hypothesis is based on results from extensive animal experiments, so discussion of the human studies is preceded by an overview of “neuropeptides” with some comments on “methodological approaches,” of “neuropeptide – small transmitter molecule coexistence,” of the neuropeptide “galanin,” followed by a summary of the critical and relevant animal experiments.
The concept of neuropeptide transmitters was introduced by the late Dutch scientist David de Wied and colls. (see De Wied and De Kloet, 1987). Neuropeptides are different from classic transmitters in several ways (Strand, 1991). In brief, neuropeptides are ribosomally synthesized as large precursor molecules in cell soma and dendrites (Noda et al., 1982; Mains et al., 1987), and the bioactive peptide(s) is excised from prepropeptide precursors by convertase enzymes (Seidah and Chretien, 1999). Packed in storage vesicles the peptides are axonally transported and released by exocytosis from nerve terminals, and also from dendrites and soma.
Neuropeptides in the nervous system encompass > 100 members (Burbach, 2010), almost always acting via one or more of a correspondingly large number of 7-transmembrane, G protein-coupled receptors (GPCRs) (>200). Much research is ongoing in the neuropeptide field. A search on PubMed with the terms “neuropeptides, review” (August 1, 2018) generated 35.579 hits. However, work on neuropeptides has not been without controversies. Already in the 1990’ies doubts were expressed with regard to functional significance [see for example the article entitled “Superfluous neurotransmitters” (i.e., neuropeptides) by Bowers (1994)]. The recent statement by Sudhof (2017) still reflects a cautious attitude: “At the forefront of early molecular neuroscience was the identification of neuropeptide precursors and neuropeptide receptors (Noda et al., 1982), but since then the question of neuropeptide signaling has largely faded from view with a few exceptions.”
However, peptides have an important and well accepted physiological function, when they are expressed in neurosecretory systems (Scharrer and Scharrer, 1937; Bargmann, 1949; Bargmann and Scharrer, 1951; Swaab et al., 1975; Vandesande and Dierickx, 1975; Brownstein and Mezey, 1986; Swanson et al., 1986; Ceccatelli et al., 1989; Meister, 1993; Morris et al., 1998; Gainer et al., 2002; Landgraf and Neumann, 2004; Jurek and Neumann, 2018), releasing their peptides into the general circulation (e.g., vasopressin, oxytocin) (Acher and Chauvet, 1954; Du Vigneaud, 1954), or into the hypothalamic portal circulation [thyrotropin releasing hormone (TRH), luteinizing releasing hormone (LHRH), somatostatin (a.k.a. growth hormone release-inhibiting hormone, GHR-IH), corticotropin releasing factor/hormone (CRF/CRH), and growth hormone releasing hormone (GHRH)] (Guillemin, 1978; Schally et al., 1978; Spiess et al., 1981, 1983; Vale et al., 1981; Brazeau et al., 1982; Rivier et al., 1982).
It is fair to say that many of the initial, high expectations of neuropeptides were not met. Examples are: (i) the discovery of the first endogenous ligands met- and leu-enkephalin for the morphine receptor (Hughes et al., 1975), present in dorsal horn interneurons (Hokfelt et al., 1977b), was expected to lead to new efficacious medicines for fighting pain, without the serious side effects of morphine; and (ii) antagonists to substance P, present in sensory neurons and the spinal dorsal horn (Lembeck, 1957; Hokfelt et al., 1975b; Takahashi and Otsuka, 1975) and acting as a transmitter (Otsuka et al., 1975; Henry, 1976) via NK1 receptors (Mantyh et al., 1995), were anticipated to represent a new type of painkiller.
These ‘failures’ have occurred in spite of considerable efforts from academia and pharmaceutical companies. For example, a substance P (neurokinin 1, NK1) antagonist was tested some 25 years later in the clinic but did not induce analgesia (Hill, 2000; Herbert and Holzer, 2002). However, and interestingly, it was also reported in a placebo-controlled trial in patients with moderate to severe major depression that the substance P (NK1) antagonist MK-869 (Aprepitant, EMEND), has robust antidepressant activity (Kramer et al., 1998). Moreover, the improvement was similar to that observed (in the same study) with the widely used antidepressant serotonin reuptake inhibitor (SSRI) paroxetine (Paxil, Seroxat) and essentially without (the common sexual) side effects seen with SSRIs (Kramer et al., 1998). However, a phase 3 trial failed to reproduce the antidepressant effects of MK-869 (Keller et al., 2006). Reasons for the failure in the treatment of depression have recently been analyzed (Rupniak and Kramer, 2017), and psychiatric studies of NK1 are still ongoing (e.g., Frick et al., 2016; Schank and Heilig, 2017). Neuropeptides and pharmacotherapy for depression will be discussed further below.
There is, however, one ‘sphere’ where neuropeptides have achieved a significant ‘status,’ and that is as markers for specific neuron populations, in particular in cortex and hippocampus1, without defining their functional role. This said, there are interesting examples, where a neuropeptide is essential for particular mouse behaviors. For example, in the lateral amygdaloid nucleus gastrin releasing peptide (GRP) regulates fear via the GRP receptor (Shumyatsky et al., 2002), and the same peptide and receptor modulate sighing in the preBötzinger complex in the ventrolateral medulla oblongata (Li et al., 2016). Arcuate AgRP neurons projecting to i.a. the parabrachial nucleus (Broberger et al., 1998) represent another example. These neurons are GABAergic and also express and release NPY, thus a good example of peptide and small molecule co-transmission. Alhadeff et al. (2018) have now shown that, of these three molecules, NPY via its NPY Y1 receptor is selectively responsible for a pain-inhibiting effect. Finally, based on a Drosophila study (Asahina et al., 2014), Zelikowsky et al. (2018) use a battery of the most recent methodologies to conduct a landmark study that demonstrates a key role for the neuropeptide tachykinin 2/neurokinin B and its receptor NK3 in chronic isolation stress, opening up for a new treatment strategy of this serious mood disorder.
The therapeutic potential of neuropeptide signaling has been extensively discussed based on animal experiments. These experiments also consider a possible role of neuropeptides in behaviors related to stress and mood regulation, and explore their receptors as possible targets for antidepressant drug development, a main theme of this review (Herbert, 1993; Maubach et al., 1999; Hokfelt et al., 2003; Holmes et al., 2003; Sajdyk et al., 2004; Nemeroff and Vale, 2005; Millan, 2006; Steckler, 2008; Wu et al., 2011; Griebel and Holsboer, 2012; Griebel and Holmes, 2013).
Four methods are of crucial importance for the exploration of neuropeptides and their coexistence with small molecule transmitters: Immunohistochemistry (IHC), radioimmunoassay (RIA), in situ hybridization (ISH) and real-time (quantitative) polymerase chain reaction (qPCR).2 These methods allow not only studies of the localization and levels of various neuropeptides but also give a hint toward functionality.
Neuropeptides released from nerve endings have to be replaced by ribosomal synthesis in cell soma followed by axonal transport. Thus, replacement can take a considerable time, of course especially in neurons with long projections, and especially in large brains like the human brain. However, here dendritic release is special as the distance between site of release and site of synthesis is short and allows for rapid replacement. In fact, dendritic release is associated with distinct features: peptide release (see below) via exocytosis is stimulated by depolarization-induced Ca2+ entry through voltage-gated calcium channels, whereby the SNARE proteins in the dendrites may partly differ from those in nerve endings (Ludwig and Leng, 2006; Kennedy and Ehlers, 2011; Ovsepian and Dolly, 2011; van den Pol, 2012; Ludwig et al., 2016).
Neuropeptide dynamics distinctly contrast those of classic transmitters: the latter are enzymatically produced also at release sites (in the nerve endings), and they have a membrane reuptake mechanism (transporters) at both the cell and storage vesicle membrane (Kanner, 1994; Liu and Edwards, 1997; Chen et al., 2004; Eiden et al., 2004; Hahn and Blakely, 2007; Torres and Amara, 2007). These transporters allow rapid replacement at the site of release, i.e., no axonal transport is needed. Such transporters have not been demonstrated for neuropeptides. This said, there is evidence that galanin after intraventricular injection can accumulate in a small number of neurons, e.g., in the hippocampus (Jansson et al., 2000).
Monitoring peptide mRNA levels with ISH provides a measure of activity of specific neurons. If analyzed in an experimental paradigm, one may even associate involvement of a peptide with a certain function. For example, an increase in galanin transcripts in dorsal root ganglion (DRG) neurons, after peripheral nerve injury, has been interpreted as a defense against pain (Xu et al., 2008) and as a signal for repair (Hobson et al., 2010).
However, reporting of mRNA levels alone always raises the issue of translation: Can the presence of transcript really equal the presence of protein (peptide)? Many studies suggest this to be the case in DRGs, for example. Also, the experiments on human postmortem brains, where transcript (qPCR) and peptide (RIA) were analyzed in the same samples (Barde et al., 2016) support this view (see below). Ideally this issue can be solved by double-labeling of individual cells: ISH for transcript and IHC for neuropeptide (Grabinski et al., 2015). Contrasting ISH it is, however, difficult to quantify peptide levels at the microscopic level with IHC. Also, IHC requires fixed tissues, whereas snap-frozen fresh tissue is used for ISH. Nevertheless, these histochemical/biochemical approaches have been applied in countless animal experimental studies to explore a possible functional role of neuropeptides in specific neuronal populations.
In the 1970’s several groups reported that a neuron may release more than one transmitter. These findings were often considered to violate “Dale’s principle,” a rule generally thought to state that a neuron only produces and releases one neurotransmitter. This was subsequently clarified as a misunderstanding (e.g., Eccles, 1986). Several of the early studies on transmitter co-existence focused on invertebrates, and only on classic transmitters and not neuropeptides (Kerkut et al., 1967; Brownstein et al., 1974; Hanley et al., 1974; Cottrell, 1976). Since then the analysis of co-transmission in this class of animals has been extremely informative. Thanks to in-depth analyses of the comparatively easily accessible and well-characterized systems in invertebrates using front-line methods, detailed knowledge of the mechanisms underlying co-transmission, and of its functional consequences has been generated (as reviewed in, e.g., Kupfermann, 1991; Bargmann, 1993; Nusbaum et al., 2017; Nassel, 2018). In the present article, the focus is on transmitter coexistence in mammalian systems.
In mammals, co-existence of noradrenaline (NA) and serotonin (5-hydroxytryptamine, 5-HT) in the same synaptic vesicle of sympathetic nerves in the pineal gland was reported (Jaim-Etcheverry and Zieher, 1973); but, serotonin presumably originated from pinealocytes and had been translocated into the storage sites with the help of cell and vesicular membrane transporter molecules. At that time, evidence was also presented for a developmental transmitter “switch” from a cholinergic to a noradrenergic transmitter phenotype in sympathetic neurons in vitro, with some neurons temporarily expressing both acetylcholine and noradrenaline (Furshpan et al., 1976); later work revealed that this also occurred in vivo (Landis and Keefe, 1983). Furthermore, several groups, in particular Burnstock and coworkers, provided evidence that ATP is a transmitter and co-transmitter (Burnstock, 1972), at that time a controversial view (Burnstock, 2012).
This was also the period when attention started to focus on peptides/neuropeptides in the brain. David de Wied and colleagues in the Netherlands studied the effects of pituitary hormones on behavior (de Wied and Bohus, 1966). Guillemin and Schally’s groups discovered that the hypothalamic thyrotropin-releasing hormone is a tripeptide (Boler et al., 1969; Burgus et al., 1970), and several new peptides were isolated from the intestine and brain (Tatemoto and Mutt, 1980; Mutt, 1989). Also substance P was isolated from the intestine (von Euler and Gaddum, 1931), but only after 40 years (!) was it chemically identified as an undecapeptide (Chang and Leeman, 1970; Chang et al., 1971). Last but not least, a very large number of important peptides were isolated from the skin of various frog species by Erspamer et al. (1978). In a visionary review, Burnstock raised the question “Do some nerve cells release more than one transmitter?” with focus on ATP and also mentioning neuropeptides (Burnstock, 1976).
At that time the neuropeptide somatostatin was, surprisingly, localized to peripheral sympathetic neurons (Hokfelt et al., 1977a) already known to signal via NA, the transmitter of sympathetic neurons (von Euler, 1948; Hamberger and Norberg, 1963) (Figures 1A,B). Somatostatin had been discovered as an inhibitor of growth hormone release from the anterior pituitary (Brazeau et al., 1973; Vale et al., 1975; Guillemin, 2008). However, it turned out that somatostatin was not only present, as expected, in neurosecretory nerve endings in the hypothalamic median eminence (Dubois et al., 1974; Hokfelt et al., 1974a; Pelletier et al., 1975), but also in many other brain nuclei (Hokfelt et al., 1974a, 1975a; Brownstein et al., 1975; Dubé et al., 1975; Elde and Parsons, 1975). This indicated roles far beyond that of a hypothalamic hormone controlling pituitary growth hormone release. Then somatostatin was shown to have a depressant action on cortical neurons (Renaud et al., 1975). So somatostatin in noradrenergic neurons was the first example of coexistence of a neuropeptide transmitter with a classic neurotransmitter in mammals (Hokfelt et al., 1977a).
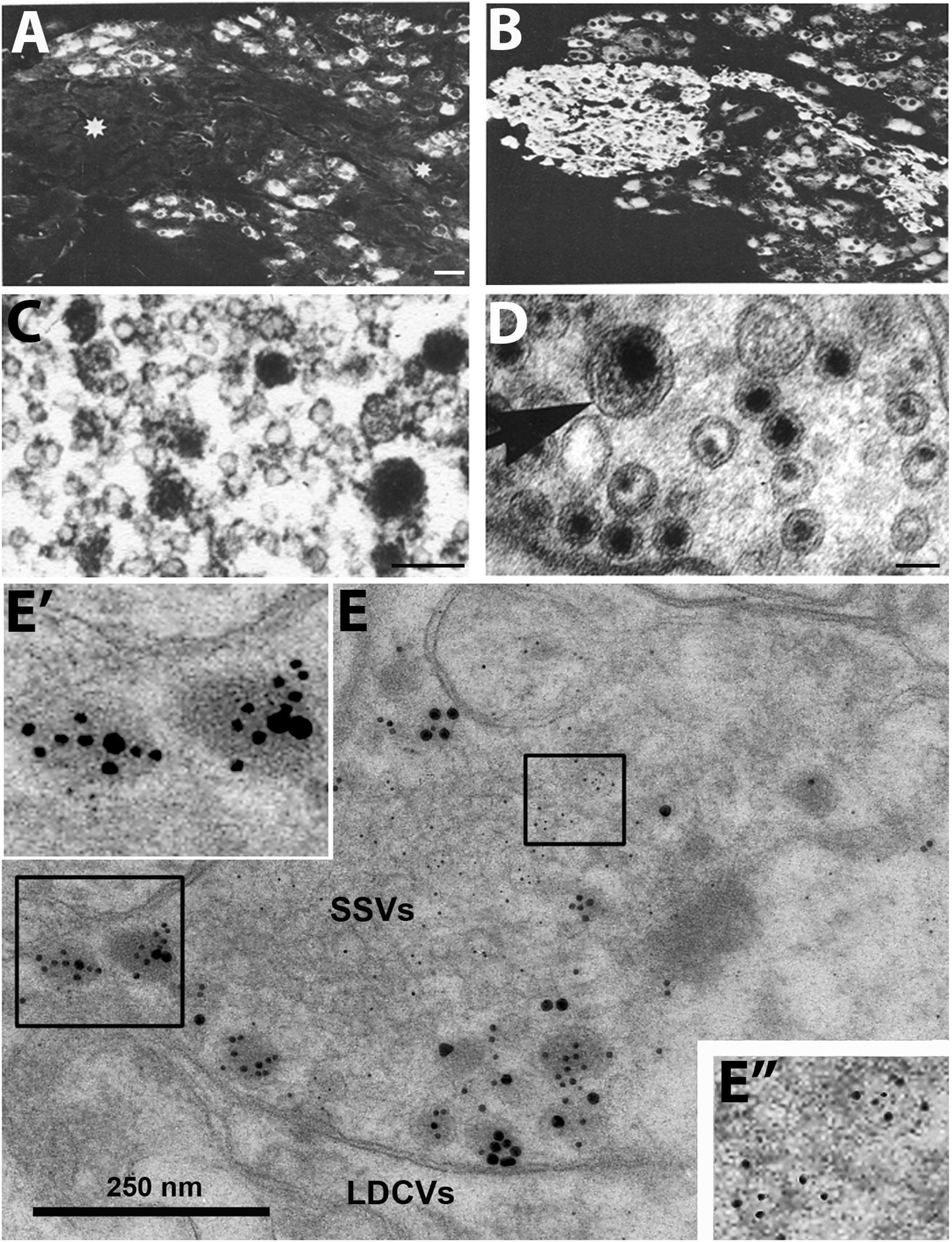
Figure 1. Immunofluorescence micrographs of the guinea-pig inferior mesenteric ganglion (A,B) and electron micrographs from different types of nerve endings (C–E). (A,B) Two adjacent sections incubated with antibodies to somatostatin (A) and the noradrenaline (NA) synthesizing enzyme dopamine ß-hydroxylase (DBH) (B). The majority of the principal ganglion cells are somatostatin-positive, whereas the small intensely fluorescent (SIF) cells (asterisk) lack the peptide. Virtually all ganglion cells and the SIF cells are DBH-positive, i.e., are noradrenergic. (C–E) Examples of transmitter storage in nerve endings based on or immunohistochemistry (C,E) or potassium permanganate fixation (D). (D) In sympathetic nerve endings NA (black precipitate) is stored in both (small) synaptic vesicles and large dense core vesicle (LDCVs) (arrow). Note that content varies between vesicles, both in the synaptic and LDCVs. (C) Substance P, a neuropeptide (black precipitate), in a sensory nerve ending in the monkey dorsal horn, is stored exclusively in LDCVs, all synaptic vesicles are empty. (E) Peptide and glutamate co-storage and coexistence in the dorsal horn of the rat spinal cord based on immunogold immunohistochemistry. Substance P/CGRP is detected with 10/20 nm gold particles and glutamate with 5 nm gold particles. Note that substance P and CGRP can be stored within the same LDCV (left box, magnified in E’). Staining for glutamate is restricted to synaptic vesicles (right box, magnified in E”). The results suggest that glutamate, a small molecule transmitter, is not stored in LDCVs in sensory nerve endings, and release of peptide and amino acid may be separate events. This contrasts NA (see D). Bars: 40 μm, for (A,B); 100 nm for (C,D); 250 nm for (E). (A,B) From Hokfelt et al. (1977a). (C) From DiFiglia et al. (1982), with permission. (D,E) Courtesy of Dr. A. Merighi (cf., Merighi, 2002).
Other early examples of this type of coexistence were vasoactive intestinal polypeptide with acetylcholine (Lundberg et al., 1979), and the neuropeptide Y (NPY) with NA (Lundberg et al., 1982). In the brain substance P was found in 5-HT (serotonin) neurons (Chan-Palay et al., 1978; Hokfelt et al., 1978), and cholecystokinin (CCK) in dopamine neurons (Hokfelt et al., 1980b), followed by many more combinations.
Regarding function, it could be shown, for example, that VIP contributes to the atropine-resistant vasodilation in exocrine glands (Lundberg et al., 1980), that NPY interacts with NA in sympathetic functions (Allen et al., 1982; Lundberg et al., 1982; Ekblad et al., 1984), and that CCK affects dopamine release (Kovacs et al., 1981; Starr, 1982), binding (Fuxe et al., 1981; Murphy and Schuster, 1982) and behavior (Crawley et al., 1984). In an elegant landmark study on a frog sympathetic ganglion Jan and Jan demonstrated that cholinergic presynaptic fibers express and release an LHRH-like peptide that is responsible for the late, slow excitatory post-synaptic potential via ‘volume transmission’ (Jan and Jan, 1982).
Taken together, these findings suggested a new principle: co-transmission - the release of a neuropeptide and a classic (small molecule) transmitter from the same neuron. In fact, the view emerged that neuropeptides always ‘co-exist’ with small molecule transmitters. Moreover, many groups, using IHC at the ultrastructural level, found that peptides are stored in large dense core vesicles (LDCVs) (diameter ∼1,000 Å) (Goldsmith and Ganong, 1975; Swaab et al., 1975; Vandesande and Dierickx, 1975; Castel and Hochman, 1976; Dube et al., 1976; Krisch, 1976; Pelletier et al., 1981; Merighi, 2002) (Figures 1C,E), whereas monoamines like NA are present both in synaptic vesicles (diameter ∼500 Å) and LDCVs as shown with potassium permanganate fixation (KMnO4) (Figure 1D) (Richardson, 1966; Hokfelt and Jonsson, 1968). The number of LDCVs in a nerve ending is mostly low compared to synaptic vesicles, indicating a lower content of peptide molecules vs. classic transmitters. However, the affinity at peptide receptors is in the low nanomolar range, allowing efficacious signaling even by low numbers of peptide molecules in the extracellular space.
It was not clear, whether IHC could exclude that peptides are stored in synaptic vesicles. Pelletier et al. (1981) incubated adjacent, ultrathin sections with antibodies against substance P and 5-HT, respectively, but in both cases only LDCVs were stained, not synaptic vesicles. This in spite of the fact that monoamines are (mainly) stored in synaptic vesicles (Figure 1D). Thus, it did not seem possible to visualize the main transmitter (5-HT) in the synaptic vesicles with IHC, contrasting, e.g., the KMnO4 method for NA (Figure 1D). So perhaps IHC also failed to demonstrate neuropeptides in synaptic vesicles? Therefore, subcellular fractionation studies were carried out, strongly suggesting lack of peptide in the synaptic vesicle pool but presence of NPY in the fraction with many LDCVs (Figures 2A–E) (Lundberg et al., 1981; Fried et al., 1985)3. In contrast to monoaminergic neurons, in sensory glutamatergic neurons the amino acid appears to be exclusively stored in synaptic vesicles (Merighi, 2002) (Figures 1C,E).
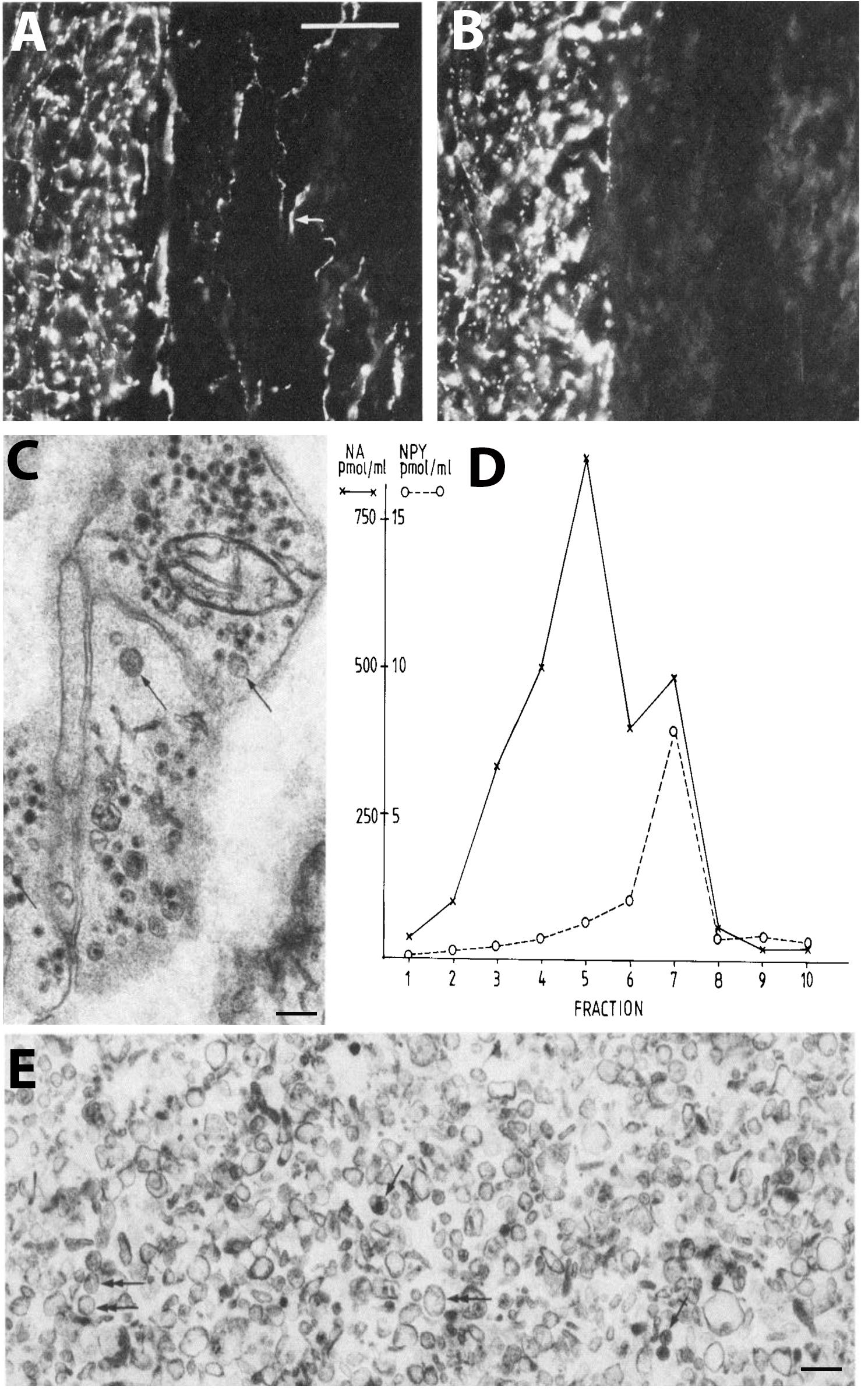
Figure 2. Coexistence and subcellular distribution of neuropeptide Y (NPY) and noradrenaline (NA) in the rat vas deferens. (A,B) Immunohistochemical visualization of NPY- (A) and tyrosine hydroxylase (TH)-(B) positive nerve terminals in adjacent sections. Overlapping, dense NPY and noradrenergic networks are seen in the muscle layer. Note sparse NPY-only positive nerves (arrow) in the subepithelial region, possibly cholinergic nerves. (C) Electron microscopic micrograph of several nerve terminal profiles in the muscle layer after potassium permanganate (KMnO4) fixation, showing small synaptic vesicles with a dense core and LDCVs. The dense core indicates presence of NA both in the synaptic and LDCVs (cf. Figure 1D). No profiles without small vesicle with a dense core are seen, suggesting a pure adrenergic innervation of the muscle layer. (D,E) Subcellular distribution of NA (x) and NPY (o) in a density gradient of rat vas deferens. There is only one peak for NPY (fraction 7; E), whereas there are two peaks for NA (fraction 5 and 7), tentatively representing synaptic vesicles and LDCVs, respectively. Note many LDCVs (arrows), as well as many vesicles of the same size but without dense core (double-headed arrow). The peptide is only present in the heavy fraction (in agreement with Figures 1C,E), whereas NA is present also in the light one (in agreement with Figure 1D). On the abscissa, totally recovered sedimentable substance is given as picomoles per milliliter after centrifugation at 145,000 × gmax for 45 min. On the ordinate, density gradient fractions 1–10 are given, corresponding to the following sucrose molarities: 1 (0.26 M), 2 (0.32 M), 3 (0.47 M), 4 (0.56 M), 5 (0.69 M), 6 (0.74 M), 7 (0.84 M), 8 (0.91 M), 9 (0.98 M), 10 (1.2 M). Recoveries of NA = 70%, of NPY = 65%, and of protein = 87%. Reprinted from Fried et al. (1985), with permission.
Furthermore, peptides are in general released when neurons fire at high frequency or in bursts (Lundberg et al., 1980; Andersson et al., 1982; Bondy et al., 1987; Bartfai et al., 1988; De Camilli and Jahn, 1990; Verhage et al., 1991; Consolo et al., 1994; Xia et al., 2009), and often extrasynaptically (Zhu et al., 1986) (Figure 3). The latter was already indicated in a pioneering study on the presynaptic structure of the synapse, showing docking sites for the synaptic vesicles which, however, are not spacious enough to leave room for LDCVs which are twice-the-size (1,000 Å) (Pfenninger et al., 1969) (Figure 3)4. This is of course not valid for somato-dendritic release and where true synapses do not exist, nor for the peripheral autonomic nervous system, where there is a considerable distance between the nerve ground plexus (Hillarp, 1949; Falck, 1962) and the smooth muscle cells, as shown with electron microscopy combined with electrophysiology (Merrillees et al., 1963). Furthermore, in the brain, extrasynaptically released neuropeptides may diffuse over long distances, so called volume transmission (Fuxe et al., 2010).
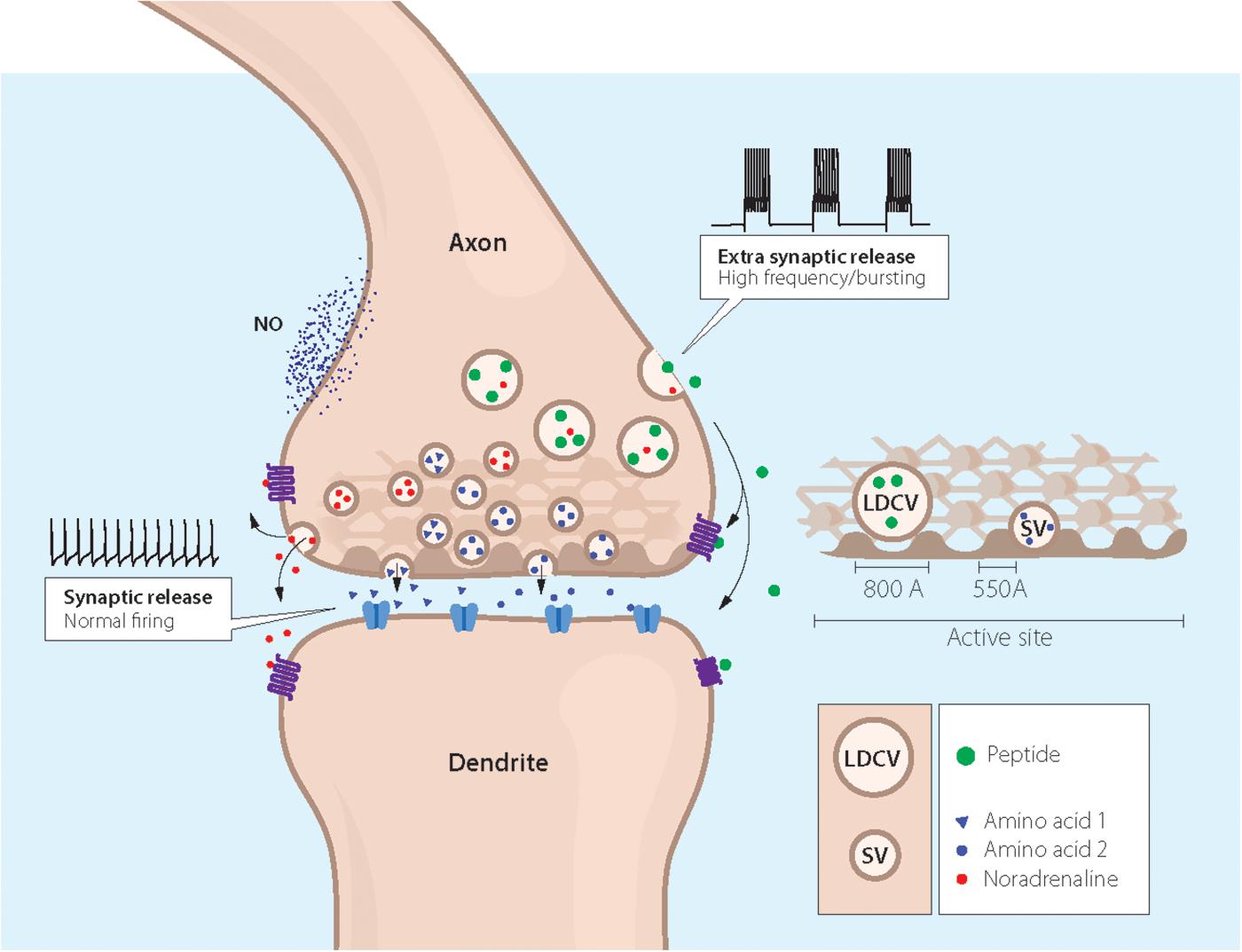
Figure 3. Cartoon showing coexistence of a neuropeptide with classic and ‘unconventional’ neurotransmitters in a nerve ending synapsing on a dendrite. Two types of storage vesicles are shown: synaptic vesicles (diameter 500 Å) storing classic transmitters (e.g., 5-HT, NA, GABA or glutamate), mainly released at synapses; large dense core vesicles (LDCVs) storing neuropeptides and, in amine neurons NA or 5-HT. The peptides are in general released extrasynaptically (“volume transmission”), when neurons fire with high frequency or in bursts. Peptide receptors are essentially extrasynaptic or presynaptic, whereas ligand-gated receptors are mostly localized in the postsynaptic membrane. ‘Gaseous’ (e.g., nitric oxide, NO) and other non-conventional transmitters are not stored in vesicles, but are generated upon demand (Snyder and Ferris, 2000). The presynaptic grid, an egg basket-like structure, originally described by Pfenninger et al. (1969), is indicated in the nerve ending and high-lighted to the right. Note that the LDCV does not fit into the grid and thus cannot attach to the presynaptic membrane for release. In contrast, there is room for the synaptic vesicle. This supports the concept that peptides are mostly not released into the synaptic cleft. Drawing by Mattias Karlen. Modified from Pfenninger et al. (1969),Lundberg and Hokfelt (1983), and Lang et al. (2015).
The exocytotic machinery underlying neurotransmitter release has been thoroughly characterized with regard to release of small molecule transmitters stored in synaptic vesicles (De Camilli and Jahn, 1990; Sudhof, 2014). However, the exocytotic neuropeptide release from LDCVs is less well defined. In early studies on synaptosomes it was shown that CCK release from LDCVs is triggered by small elevation of Ca2+ concentration in the bulk cytoplasm, whereas glutamate release from the synaptic vesicles requires the higher concentrations produced close to Ca2+ channels in the active zone (Verhage et al., 1991). This is in agreement with the localization of the two types of vesicles consistently observed in electron microscopic micrographs of the nerve endings: many synaptic vesicles with some close to the presynaptic membrane, versus a few LDCVs virtually always distant from the synapse (Figure 3).
There is evidence for involvement of SNAREs [soluble N-ethyl maleimide (NEM)-sensitive factor attachment protein receptor protein family] (Sudhof, 2014) also in dendritic release from magnocellular dendrites (Schwab et al., 2001; de Kock et al., 2003; Ovsepian and Dolly, 2011). The calcium-dependent activator protein for secretion (CAPS) (Walent et al., 1992) has been identified as a priming factor for exocytosis of LDCVs (Stevens and Rettig, 2009; James and Martin, 2013). Thus CAPS2, but not CAPS1, is required for LDCV exocytosis as shown in cerebellar granule cells and hippocampal interneurons (Sadakata et al., 2004; Shinoda et al., 2011).
Taken together, these early findings suggested that neuropeptides were not the main neuronal messengers. Moreover, when neuropeptides are released, the fast small molecule transmitters are already active in the synaptic cleft – i.e., no peptide release without release of classic transmitters. The discovery of coexistence and co-transmission was summarized in several books/reviews (Burnstock, 1978; Hokfelt et al., 1980a, 1986, 1987a; Cuello, 1982; Chan-Palay and Palay, 1984; Jaim-Etcheverry, 1994; Merighi, 2002), and since then further efforts have been made to understand co-signaling involving neuropeptides, including co-release of both an excitatory and an inhibitory neuropeptide. For an up-to-date overview of many aspects on neuropeptide signaling (see e.g., Salio et al., 2006; van den Pol, 2012; Ludwig et al., 2016).
More recently it has become clear that coexistence of small molecule transmitters, encompassing various combinations of GABA, glycine, glutamate, dopamine and acetylcholine (e.g., Guiterrez, 2009; Hnasko and Edwards, 2012; Trudeau et al., 2014) (Figure 3). For example, coexistence of GABA and glycine was first reported in the cerebellum (Ottersen et al., 1988), and then in the spinal cord (Todd and Sullivan, 1990; Ornung et al., 1994), where evidence for GABA-glycine co-transmission was obtained in the dorsal horn, and possible co-release from the same synaptic vesicles (Jonas et al., 1998) (Figure 3). Moreover, mesencephalic dopamine neurons can also release glutamate (Hnasko et al., 2010) and GABA (Tritsch et al., 2012), whereby GABA is not synthesized via the classic enzyme glutamate decarboxylase (GAD) but via aldehyde dehydrogenase 1a1 (Kim et al., 2015).
Thus, the number and combinations of transmitters present in a nerve ending (and/or dendrites) virtually seem endless, and it is difficult to define rules according to which neurotransmitters co-exist and are involved in co-transmission, as is discussed further in this Frontiers special topic. Furthermore, neurotransmitter switching, the gain of one and loss of another transmitter in the same, mammalian neuron, can occur not only during development but also in adult animals (Spitzer, 2017).
There is an increasing interest in neuropeptide/neurotrans-mitter coexistence and a need to explore transcriptional changes in defined healthy and diseased brain circuitries (Akil et al., 2010). In fact, there are many interesting results from animal disease models, suggesting involvement of neuropeptides and neuropeptide coexistence in patho-physiological processes with potential therapeutic implications. However, information on the significance of transmitter and neuropeptide coexistence in the normal and diseased human nervous system is limited. In this article, the focus is on galanin co-existing in noradrenergic neurons in the LC, and on galanin receptor expression in postmortem brains from normal subjects and depressed patients who committed suicide (Le Maitre et al., 2013; Barde et al., 2016). This is in line with previous extensive work carried out on postmortem brains from depressed humans, showing changes in transcripts related to neurotransmitters/neuropeptides and their receptors and to transporters, growth factors in nerve cells, and in glia, in cortical, limbic, hypothalamic and lower brain stem regions (Evans et al., 2004; Iwamoto et al., 2004; Aston et al., 2005; Choudary et al., 2005; Kang et al., 2007; Anisman et al., 2008; Kozicz et al., 2008; Tochigi et al., 2008; Klempan et al., 2009; Sequeira et al., 2009, 2012; Sibille et al., 2009; Poulter et al., 2010; Bernard et al., 2011; Bloem et al., 2012; Kerman et al., 2012; Zhurov et al., 2012; Labonte et al., 2013, 2017; Li et al., 2013; Du et al., 2014; Lopez et al., 2014a,b; Hayley et al., 2015; Maheu et al., 2015; Torres-Platas et al., 2016; Roy et al., 2017).
Galanin was originally isolated from porcine intestine as a 29-amino acid (30 in humans) neuropeptide (Tatemoto et al., 1983; Schmidt et al., 1991) (Figure 4A) with a wide distribution in the rat brain as shown with RIA (Skofitsch and Jacobowitz, 1986), IHC (Rokaeus et al., 1984; Melander et al., 1985, 1986b,c,d; Skofitsch and Jacobowitz, 1985; Merchenthaler et al., 1993), and ISH (Gundlach et al., 1990b; Jacobowitz and Skofitsch, 1991; Jacobowitz et al., 2004). The distribution of galanin in the mouse brain is similar to that in rat, both with regard to galanin peptide (Perez et al., 2001) and to its mRNA (Cheung et al., 2001; Lein et al., 2007). The galanin system has also been characterized in the monkey brain (Melander and Staines, 1986; Kordower and Mufson, 1990; Walker et al., 1991) (for human brain, see below).
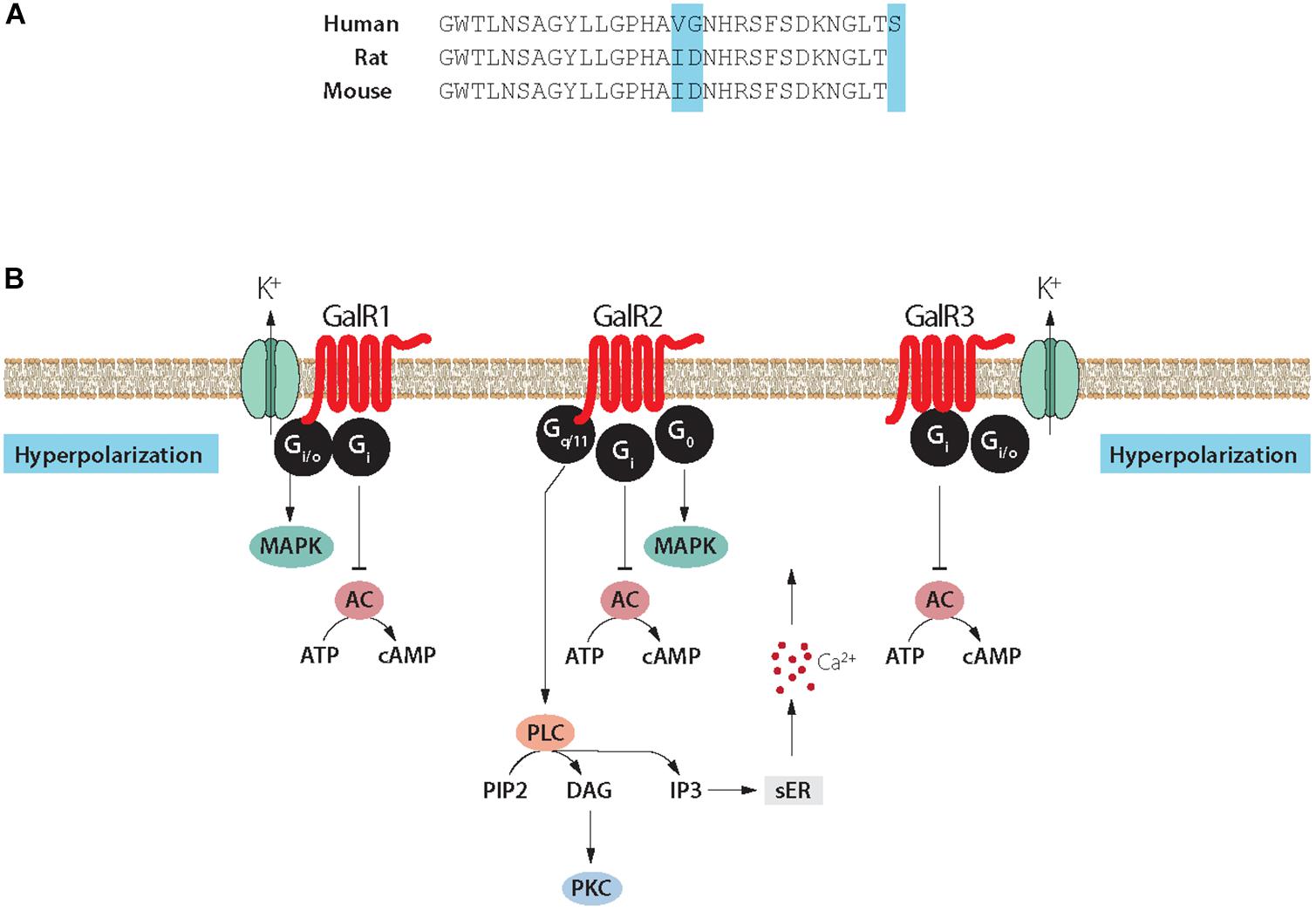
Figure 4. (A) Structure of galanin in three species. Galanin is composed of 29 amino acids in most species, except humans (30 amino acids). Note conservation of N-terminal portion. (B) Signaling pathways of galanin receptor subtypes. Galanin, via GalR1 and GalR3, opens potassium channels leading to membrane hyperpolarization. Galanin can via GalR2 activate PLC resulting in generation of IP3, release of Ca2+ from the smooth endoplasmic reticulum, opening of Ca2+ channels and eventually transmitter release. AC, adenylate cyclase; cAMP, 3′, 5′-cyclic adenosine monophosphate; DAG, diacylglycerol; K+, G-protein-regulated inwardly rectifying potassium channel; sER, smooth endoplasmic reticulum; IP3, inositol triphosphate; PIP2, phosphatidylinositol bisphosphate; PKC, protein kinase C; PLC, phospholipase C. Modified from Iismaa and Shine (1999) and Lang et al. (2015). Drawing by Mattias Karlén.
For many years galanin was considered as the sole endogenous ligand for GalR1-3 but more recently additional ligands were described (Lang et al., 2015)5. Currently, three galanin receptors, GalR1-3, have been cloned, all three belonging to the family of seven transmembrane-spanning GPCRs, with different transduction mechanisms, with GalR1 and -R3 having distinct similarities (Habert-Ortoli et al., 1994; Fathi et al., 1997; Howard et al., 1997; Wang et al., 1997; Ahmad et al., 1998; Smith et al., 1998; Iismaa and Shine, 1999; Branchek et al., 2000; Lang et al., 2007, 2015) (Figure 4B). The three galanin receptors are present in most parts of the rat brain, but could not be detected e.g., in dorsal cortical areas and the hippocampal formation (HiFo) in early autoradiographic ligand binding studies (Skofitsch et al., 1986; Melander et al., 1986a, 1988).
Galanin receptors have also been mapped in the mouse brain using 125I-galanin binding autoradiography (Jungnickel and Gundlach, 2005). A direct comparison with results in rat in the study by, e.g., O’Donnell et al. (2003) reveals an overall similar distribution but with some remarkable, apparently qualitative species differences. Thus, mouse shows, i.a., a strong signal in two important regions, the striatum and the cerebellum (Jungnickel and Gundlach, 2005) which both lack binding in the rat (Skofitsch et al., 1986; Melander et al., 1988; O’Donnell et al., 2003). To our knowledge, no attempts have been made to identify the cellular localization and origin of, e.g., the structures binding galanin in the mouse striatum.
The cloning of the receptors allowed localization with ISH and qPCR, which revealed that the transcripts for GalR1 and GalR2 are widely distributed in the rat brain, primarily in the brain stem and in ventral cortical areas (Landry et al., 1998; Mitchell et al., 1999; O’Donnell et al., 1999, 2003; Burazin et al., 2000; Waters and Krause, 2000; Mennicken et al., 2002). However, the GalR2 transcript is transiently highly expressed in neocortex during the first week after birth (Burazin et al., 2000). The distribution of GalR3 is limited (Mennicken et al., 2002). Only the GalR1 transcript has been mapped with ISH in the mouse brain (Hohmann et al., 2003; Lein et al., 2007). Thus, The Allen Brain Atlas (Lein et al., 2007) lacks results on GalR2 or GalR3, suggesting that they are expressed at low levels. This is also supported by the demonstration that the 125I-galanin binding sites are absent in a GalR1 knock-out mouse (Jungnickel and Gundlach, 2005). Taken together, these results suggest that GalR1 is the predominant receptor in the mouse brain, and that distinct species differences exist between mouse and rat.
GalR3 has emerged as a complex receptor (Lang et al., 2015), not present in all mammals (Liu et al., 2010). Its signaling properties are still not well defined, even though GalR3-transfected cell lines have now been generated (Lu et al., 2005b; Robinson et al., 2013). However, these cells could so far not be used for stable signaling experiments (see Lang et al., 2015). Still, GalR3 presumably acts via a PTX sensitive Gi/o-type G protein which in turn regulates inwardly rectifying K+ channels (Smith et al., 1998), as do GalR1 receptors (Smith et al., 1998) (Figure 4B). This lack of knowledge contrasts the substantial information about GalR1 and GalR2 (see Lang et al., 2015). The cloning of the receptors was useful, also because it has been difficult to raise specific antibodies to GalR1-3 (Lu and Bartfai, 2009; Brunner et al., 2018). A similar situation exists for other GPCRs (Michel et al., 2009). Detailed tables on the distribution of galanin and GalR1-3 in rodent brain are found in O’Donnell et al. (1999, 2003), Burazin et al. (2000), Hohmann et al. (2003), and Jungnickel and Gundlach (2005).
Early research on galanin was initiated because of its strong reaction to nerve injury. Transection of the sciatic nerve in rat causes an >100-fold increase in galanin synthesis (mRNA and peptide levels) in the corresponding somata of DRG somata (Hokfelt et al., 1987b). Upregulation could also be detected in the brain after various types of injury/manipulations (Cortes et al., 1990a,b; Villar et al., 1990; Agoston et al., 1994; Palkovits, 1995). In fact, galanin meets the criteria of a neurotransmitter/-modulator, but also has trophic functions, as shown both in brain and the peripheral nervous system (Hobson et al., 2010). Galanin has, in fact, many characteristics similar to the brain-derived neurotrophic factor (BDNF), including storage in, and exocytotic release from LDCVs and both transmitter and trophic functions (Barde, 1994). For example, galanin affects spine density (Sciolino et al., 2015), and it is well-known that BDNF influences dendritic morphology (Bennett and Lagopoulos, 2014). Thus, trophic functions of galanin are potentially interesting but will not be discussed here.
A further early finding in the rat was the coexistence (Figures 5A,B”) of galanin (Figure 5B) in both noradrenergic neurons in the LC (Figure 5B’) (Rokaeus et al., 1984; Skofitsch and Jacobowitz, 1985; Melander et al., 1986b,c; Holets et al., 1988; Moore and Gustafson, 1989) and in serotonergic neurons in the dorsal raphe nucleus (DRN) (Melander et al., 1986c; Fuxe et al., 1990; Priestley et al., 1993; Xu and Hokfelt, 1997), two systems associated with mood-related behavior. The LC neurons also express transcripts for both GalR1 and -R2 (O’Donnell et al., 1999; Burazin et al., 2000).
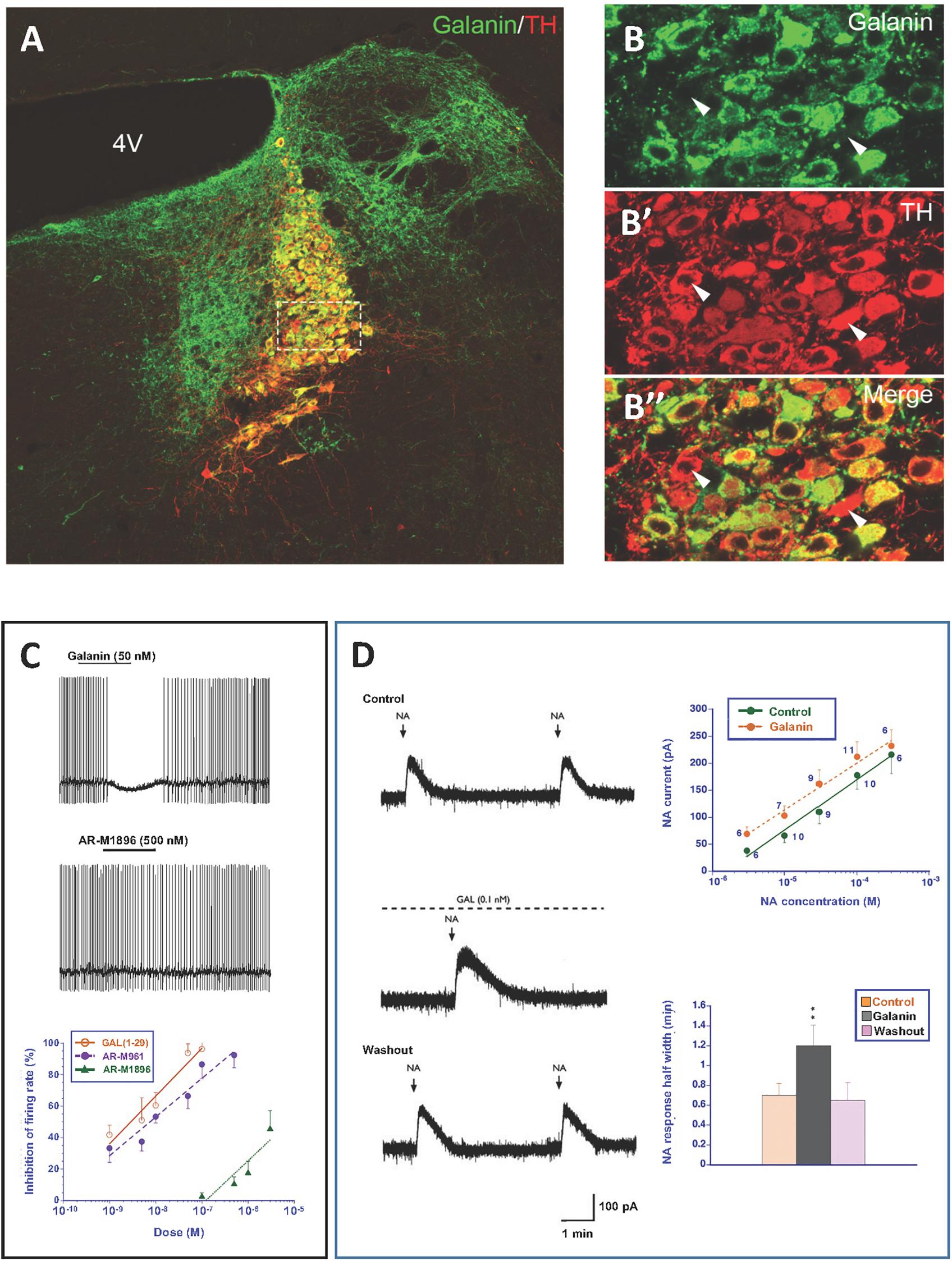
Figure 5. (A–B”) Immunofluorescence micrographs of the dorsal pontine periventricular region of mouse after double-staining of a section with antibodies to galanin (green) and tyrosine hydroxylase (TH) (red), the rate-limiting enzyme for catecholamine synthesis and thus a marker for NA neurons. Note that both antibodies stain neurons in the locus coerulus (LC) (B,B’), whereby many (yellow, B”), but not all TH-positive neurons express galanin [arrowheads point to TH-only neurons (red), apparently lacking galanin] (B’). Galanin is also present in many structures outside the LC. Colchicine treated animal. Courtesy Joanne Bakker and Mingdong Zhang. Bar for (A) 200 μm, for (B–B”) 20 μm. (C) Effect of galanin and the GalR2 agonist AR-M1896 on LC neurons (upper two traces), and the dose–response curves of galanin (red), the AR-M1896 (green) and the mixed GalR1-GalR2 M961 agonist (magenta) (lower trace). Note strong hyperpolarization of galanin and a less strong effect of M961, whereas that AR-M1896 hardly causes any effect at all. From Ma et al. (2001). (D, left panel) Effect of galanin on the response of LC neurons to NA. NA (applied from a pipette at the arrowhead) induces a persistent outward current (upper trace). When galanin (0.1 nM) is present, the NA-induced outward current is enhanced, and the duration is prolonged (middle trace). After wash out of galanin, the amplitude and duration of the NA response was similar to that seen before galanin administration (lower trace). (D, right panel) Effect of galanin on dose-response (upper figure) and duration (lower figure) of NA. The NA dose-response curve is shifted to the left, when galanin (0.1 nM) is present (upper figure). The duration of the NA-induced current is increased in the presence of galanin (lower figure). ∗∗P < 0.01. From Xu et al. (2001) with permission.
Thereafter galanin biology has since the early 1990’s been regularly summarized in books/journal from meetings (Hökfelt et al., 1991, 1998; Hökfelt and Crawley, 2005; Hokfelt, 2010; Hokfelt and Tatemoto, 2010); and in peer-reviewed articles focusing on the nervous system (only such published after 2004, and not included in the books/journals cited above, are listed here) (Lundstrom et al., 2005; Holmes and Picciotto, 2006; Karlsson and Holmes, 2006; Ogren et al., 2006, 2007, 2010; Robinson et al., 2006; Walton et al., 2006; Wrenn and Holmes, 2006; Lu et al., 2007; Tortorella et al., 2007; Picciotto, 2008; Robinson and Brewer, 2008; Butzkueven and Gundlach, 2010; Picciotto et al., 2010; Webling et al., 2012; Diaz-Cabiale et al., 2014; Freimann et al., 2015; Weinshenker and Holmes, 2016; Millon et al., 2017a; Genders et al., 2018a); and in some major comprehensive reviews (Lang et al., 2007, 2015).
Locus coeruleus is a small, compact bilateral nucleus in the pons located in the gray matter close to the lateral aspect of the 4th ventricle (Maeda, 2000). Dahlstrom and Fuxe first reported that NA is a transmitter in the rat LC, a.k.a. the A6 group (Dahlstrom and Fuxe, 1964). They used the formaldehyde, or Falck-Hillarp, fluorescence method that allows microscopic visualization of catecholamines and serotonin in tissue sections (Carlsson et al., 1962; Falck, 1962; Falck et al., 1962).
In the rat, the LC contains 2,800–3,600 neurons (both sides) (with an additional 260 neurons in the subcoeruleus area, the vast majority of which are noradrenergic with wide projections to virtually all parts of the central nervous system (Ungerstedt, 1971; Descarries and Saucier, 1972; Swanson and Hartman, 1975; Swanson, 1976; Morrison et al., 1978; Moore and Bloom, 1979; Goldman and Coleman, 1981; Foote et al., 1983; Aston-Jones et al., 1995). NA nerve terminals are also extensively present in primate cortex (Lewis et al., 1986).
When explored with electrophysiological methods galanin has effects on the membrane potential of several neuron systems (see Xu et al., 2005). Galanin hyperpolarizes noradrenergic LC neurons in a slice preparation (Seutin et al., 1989; Sevcik et al., 1993; Pieribone et al., 1995), mediated via GalR1 (Ma et al., 2001) (Figure 5C). However, the GalR2 (R3) agonist ARM-1986 (Liu et al., 2001; Lu et al., 2005b) does not cause any effect on the membrane potential (Ma et al., 2001) (Figure 5C). GalR2 may instead have a presynaptic role in the projection areas of LC neurons, perhaps mainly acting as an autoreceptor (Ma et al., 2001). In agreement, galanin is present in noradrenergic [dopamine ß-hydroxylase (DBH)]-positive nerve terminals in cortex and the hippocampus (Melander et al., 1986d; Xu et al., 1998). Galanin activation of GalR1, but not -R2 or R3, has been shown also in other studies on the rat and mouse LC (Hawes et al., 2005; Mitsukawa et al., 2009). In addition to this direct effect, galanin at low concentrations (10-9M) enhances the autoinhibitory effect of NA on LC neurons via alpha-2A receptors (Xu et al., 2001) (Figure 5D). This may in fact be the primary action of galanin in controlling the firing of LC neurons. Thus, galanin can via different autoinhibitory mechanisms exert a two-step inhibition on LC neurons, at low concentrations enhancing the inhibitory alpha-2A receptor effect.
Autoinhibition of LC neurons, mediated by NA via alpha-2A receptors, was early discovered by Svensson et al. (1975) and Aghajanian et al. (1977). It is assumed that autoinhibition, both at NA and serotonin neurons, at least in part, is responsible for the delayed onset of the clinical effect of monoamine reuptake inhibitors (Artigas et al., 1996; Mongeau et al., 1997; Millan, 2006). Autoinhibition via NA in LC was originally suggested to be a consequence of the release from collaterals (Aghajanian et al., 1977). There is, however, evidence that NA can be released from soma/dendrites (Pudovkina et al., 2001; Pudovkina and Westerink, 2005), and more recently release was shown to occur from individual vesicles by combined measurements using amperometry and patch clamp methodologies (Huang et al., 2007). This is in agreement with electron microscopic analysis, showing synaptic vesicles with a dense core in LC dendrites (Shimizu et al., 1979). Thus, collaterals are not necessarily the only structure involved in the autoinhibition.
There is another source of catecholamine input to the LC neurons originating from one of the three C neuron groups in the medulla oblongata: adrenaline (epinephrine) containing afferents (Figure 6) (Hokfelt et al., 1974b, 1984; Howe et al., 1980; Armstrong et al., 1982), which synapse on LC dendrites (Milner et al., 1989). This was supported by early tracing experiments, although at that time no transmitter histochemical identification was performed (Cedarbaum and Aghajanian, 1978). One likely origin is C1 neurons, since they display a high degree of collateralization, including inputs to the LC (Figure 6) (Haselton and Guyenet, 1990).
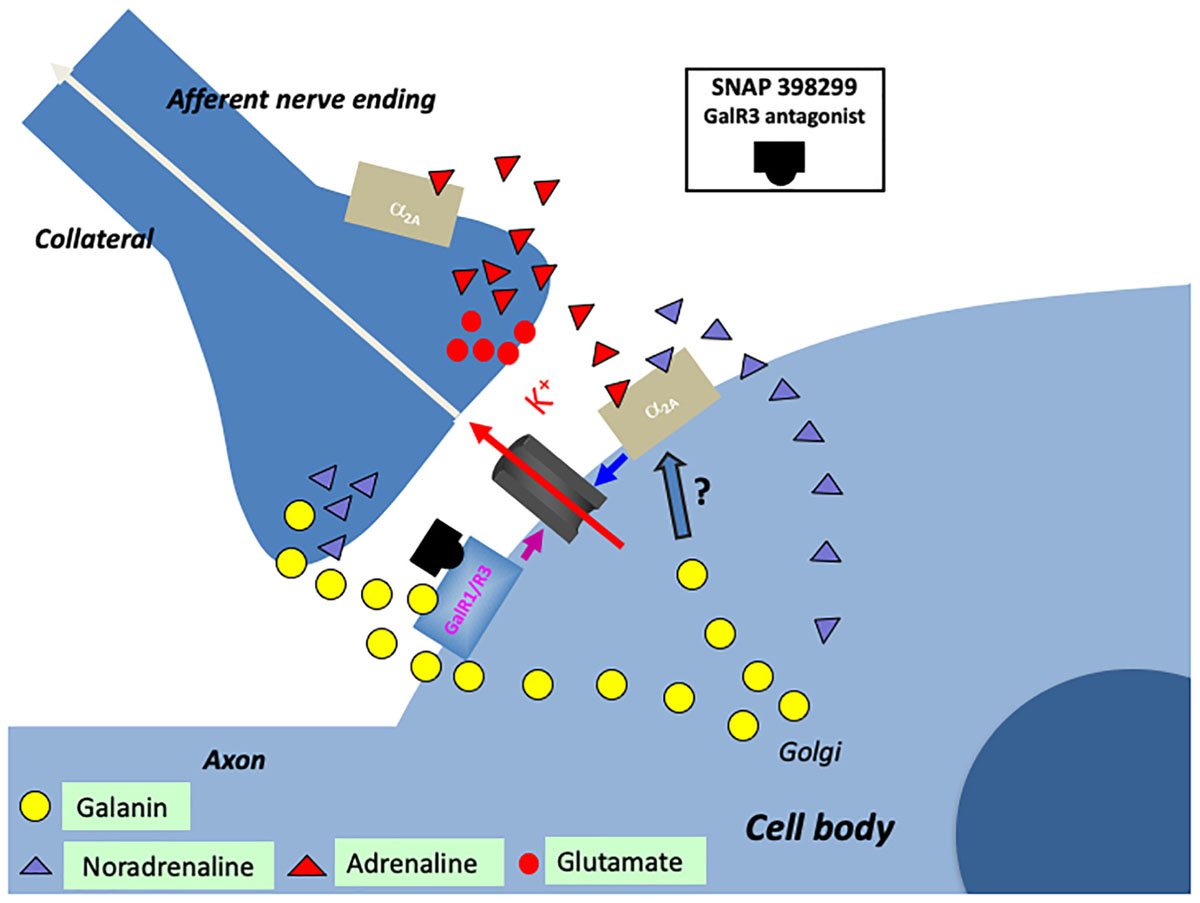
Figure 6. Cartoon showing several transmitters and signaling pathways in the locus coeruleus (LC) (part of a cell body with initial axon and an afferent nerve ending and a possible axon collateral). A noradrenergic LC neuron co-expresses galanin (yellow LDCVs) originating in the Golgi complex. The peptide in the LDCVs is, after transport to the somatic and dendritic cell membrane, released by exocytosis. Galanin then acts on inhibitory autoreceptors (GalR1/R3), opening potassium channels, in this way attenuating noradrenaline (NA) release in the forebrain. Galanin at low concentrations enhance the alpha2A mediated inhibition of the LC neuron (by an unknown mechanism). Galanin could also be released from collaterals. The GalR3 antagonist (SNAP 398299) may exert an antidepressive action by disinhibiting the LC neuron and restituting forebrain NA levels. With regard to small transmitters, NA (purple triangles) can be released from soma-dendrites and collaterals, acting on somato-dendritic, postsynaptic and presynaptic alpha2A receptors. The afferent nerve ending originates from C1 neurons which are glutamatergic (red dots) and co-release adrenaline (red triangles). Also adrenaline can act on the alpha2A receptors. The basis for this cartoon is animal experiments, and in the case of the galanin system, results from human postmortem brains are also incorporated.
Early studies suggested that the adrenaline (Cedarbaum and Aghajanian, 1976) and the C1 neurons (Aston-Jones et al., 1991) inhibit LC neurons. However, the more recent discovery that the C1 neurons are glutamatergic together with optogentic analysis demonstrated excitation as the primary effect (Figure 6) (Abbott et al., 2012). Released adrenaline may act as a modulator not only on postsynaptic but also presynaptic (Li et al., 1995) alpha-2A receptors, which will, respectively, directly and indirectly, dampen LC neuron activity (Figure 6) (Guyenet et al., 2013).
Taken together, galanin prevents overexcitation of LC, but is only one of several molecules performing this task (Aston-Jones et al., 1991; Singewald and Philippu, 1998; Van Bockstaele, 1998; Berridge and Waterhouse, 2003; Van Bockstaele and Valentino, 2013). This comprehensive network is perhaps a sign of how important it is to balance the activity of the noradrenergic LC neurons, which are involved in the control of many bodily functions (see below).
Kehr and colleagues have analyzed the effect of intracerebroventricularly administered galanin in freely moving rats and mice, monitoring several neurotransmitters using in vivo microdialysis (Ungerstedt, 1984) and a sensitive HPLC method. Their studies indicate that galanin reduces basal and desipramine-induced extracellular NA levels (Yoshitake et al., 2003, 2004). This effect is assumed to be exerted via GalR1 at the noradrenergic cell bodies/dendrites in the LC.
Studies on the hypothalamic magnocellular hormones vasopressin and oxytocin have provided compelling evidence that these two peptides are not only released from nerve endings in the posterior pituitary but also, independently, from dendrites in the paraventricular and supraoptic nuclei (Morris et al., 1998; Landgraf and Neumann, 2004; Ludwig and Leng, 2006; Kennedy and Ehlers, 2011; Ovsepian and Dolly, 2011; Ludwig et al., 2016). There is evidence for involvement of SNAREs [soluble N-ethyl maleimide (NEM)-sensitive factor attachment protein receptor protein family] (Sudhof, 2014) in release from magnocellular dendrites (Schwab et al., 2001; de Kock et al., 2003; Ovsepian and Dolly, 2011). Results from studies on CAPS2-dependant neuropeptide release from soma of dorsal root ganglion neurons (Bost et al., 2017; Shaib et al., 2018) may also be relevant for dendritic/somatic release in the brain. Galanin may be released from soma and dendrites in the LC (Pieribone et al., 1995; Vila-Porcile et al., 2009) (Figure 6). Therefore, it has been hypothesized that stress-induced firing increases galanin release from nerve terminals in the forebrain and dendrites-soma of LC neurons. This could lead to activation of GalR1 autoreceptors and inhibition of firing of LC neurons, a possible mechanism involved in resilience and development of depression-like behavior in animals (Sciolino et al., 2015) (see below).
Neuropeptide Y is expressed in LC neurons in rat (Everitt et al., 1984; Chronwall et al., 1985; Yamazoe et al., 1985; Holets et al., 1988) and human (Chan-Palay et al., 1990). Recently it has been shown in mice that dopamine is co-released with NA in the hippocampus (Kempadoo et al., 2016; Takeuchi et al., 2016) and the paraventriculer thalamic nucleus (Beas et al., 2018) and is involved in memory consolidation and control of stress responsitivity, respectively.
Galanin influences mood-related behavior in a region-specific way (Bing et al., 1993; Moller et al., 1999). Moreover, results from a number of rat experimental models suggest that galanin can be both prodepressive/anxiogenic and antidepressive (Fuxe et al., 1990, 1991, 1998; Weiss et al., 1998, 2005; Bellido et al., 2002; Khoshbouei et al., 2002; Barrera et al., 2005; Sergeyev et al., 2005; Lu et al., 2005a, 2007, 2008; Holmes and Picciotto, 2006; Karlsson and Holmes, 2006; Ogren et al., 2006; Kuteeva et al., 2008, 2010; Kozlovsky et al., 2009; Picciotto et al., 2010; Le Maitre et al., 2011; Sciolino et al., 2012, 2015; Weinshenker and Holmes, 2016).
In many of the early studies listed above on depressive-like behavior the receptor involved was not identified, or the site of action was not defined experimentally, but there was a general consensus that it is GalR1 that mediates the depressive behavior and that GalR2 may be prodepressive (summarized in Mitsukawa et al., 2009; Kuteeva et al., 2010; Hoyer and Bartfai, 2012; Webling et al., 2012; Freimann et al., 2015).
Two recent studies support involvement of GalR1, and suggest the ventral periaqueductal gray as one likely site of action. Using a rat model of depression based on chronic mild stresses (CMS) (Willner et al., 1987; Moreau et al., 1992), behavior was evaluated in the open field test, the forced swim test (FST), and by monitoring sucrose consumption (Wang et al., 2016). Transcript levels of galanin and GalR1-3 in various, laser-dissected brain regions, including the hippocampal formation (HiFo), vPAG, the DRN and the LC were analyzed with quantitative real time PCR (qPCR) (Wang et al., 2016). Only GalR1 mRNA levels were significantly changed (increased), in a single region, the vPAG. Moreover, after knocking down GalR1 in the vPAG using siRNA, the depressive behavioral phenotypic parameters were similar to unstressed controls. This result suggested that the depression-like behavior in rats exposed to CMS is likely related to an elevated expression of GalR1 in the vPAG. The phenotype of the GalR1-positive neurons was not identified, despite comparing their distribution with serotonin, glutamate (vesicular glutamate transporter type 2, VGLUT2) and GABA (glutamic acid decarboxylase, GAD) neurons (Wang et al., 2016).
In another study, the galanin system was monitored with qPCR, ISH and RIA methodologies following mild blast-induced traumatic brain injury (mbTBI) (Kawa et al., 2016). Significant increases in galanin peptide and transcript were observed in the LC, at 1 day with qPCR, at 3 days with RIA and from 2 h to 7 days with ISH. The increases thus remained for 7 days (ISH) (the longest period studied). With regard to galanin receptors, GalR1 mRNA was significantly increased in vPAG at 1 and 7 days, likely in the same neuron population as seen in the CMS model (Wang et al., 2016). These findings suggest a long-lasting role for the galanin system in the endogenous response to mbTBI. Again, the phenotype of these GalR1-positive neurons was not identified. Nevertheless, in both cases stress, and possibly depression-like behavior, are associated with increased levels of GalR1 transcript in the vPAG. Interestingly, the galanin system has also been shown to modulate stress-related responses related to mild TBI in a model of postraumatic stress disorder (PTSD) (Kozlovsky et al., 2009).
The robust and lasting effect of mbTBI on the expression of galanin (at least 7 days) not only in LC but also in 5-HT neurons (Kawa et al., 2016) is more sustained than the transient increase in tyrosine hydroxylase (TH) (3 days) and tryptophan hydroxylase 2 (1 day) seen in the same mbTBI model (Kawa et al., 2015). Thus, in mbTBI the coexisting peptide may have a more long-lasting and important effect than the small molecule transmitter.
In another study, i.p. injection (a stress by itself) and swim stress increased both galanin and TH mRNA levels in the LC, but not TPH2 or galanin transcripts in the DRN (Kuteeva et al., 2008), indicating that the serotonergic system is less sensitive to stress than the noradrenergic system. This has also been shown in other studies employing different types of stress (e.g., Wilkinson and Jacobs, 1988; Jordan et al., 1994; Kuteeva et al., 2008, 2010). The long lasting effects of stress can also be gauged against the fact that peptides can exert effects over long periods of time (Herbert, 1993; van den Pol, 2012). One example is a study on the lamprey locomotor network that revealed that a 10-min administration of substance P causes a long-lasting (>24 h) modulation of the frequency and regularity of NMDA-evoked locomotor bursts (Parker and Grillner, 1999).
GalR2 may also be involved, but here an opposite effect has been recorded, i.e., galanin actions via this receptor are antidepressive (Gottsch et al., 2005; Lu et al., 2005a, 2007, 2008; Kuteeva et al., 2008, 2010; Kinney et al., 2009; Le Maitre et al., 2011; Saar et al., 2013a,b; Kawa et al., 2016), in some cases associated with the vPAG. For example, in a neuropharmacological study (Kuteeva et al., 2008) the time of immobility (Figure 7A) and climbing (Figure 7B) were recorded in the FST. Galanin, the GalR1 receptor agonist M617, the GalR2(R3) agonist AR-M1896, the GalR2 antagonist M871 or aCSF were infused intracerebroventricularly. Galanin significantly increased immobility time, as did the GalR1 receptor agonist M617 (Figure 7A). In contrast, the GalR2(R3) agonist AR-M1896 decreased immobility, similar to fluoxetine, whereas the GalR2 antagonist M871 increased the time of immobility. Together these results support the view of GalR1 being pro- and GalR2 antidepressive. Moreover, the antidepressive effect of the GalR2 antagonist suggests there is an in vivo, tonic activation of this receptor under forced swimming (stress). This provides further evidence that galanin is released in vivo under stressful conditions. More recently it has been shown that an anxiolytic-/antidepressive effect of galanin injected directly into the DRN is mediated via GalR2 (Silote et al., 2013; de Souza et al., 2018).
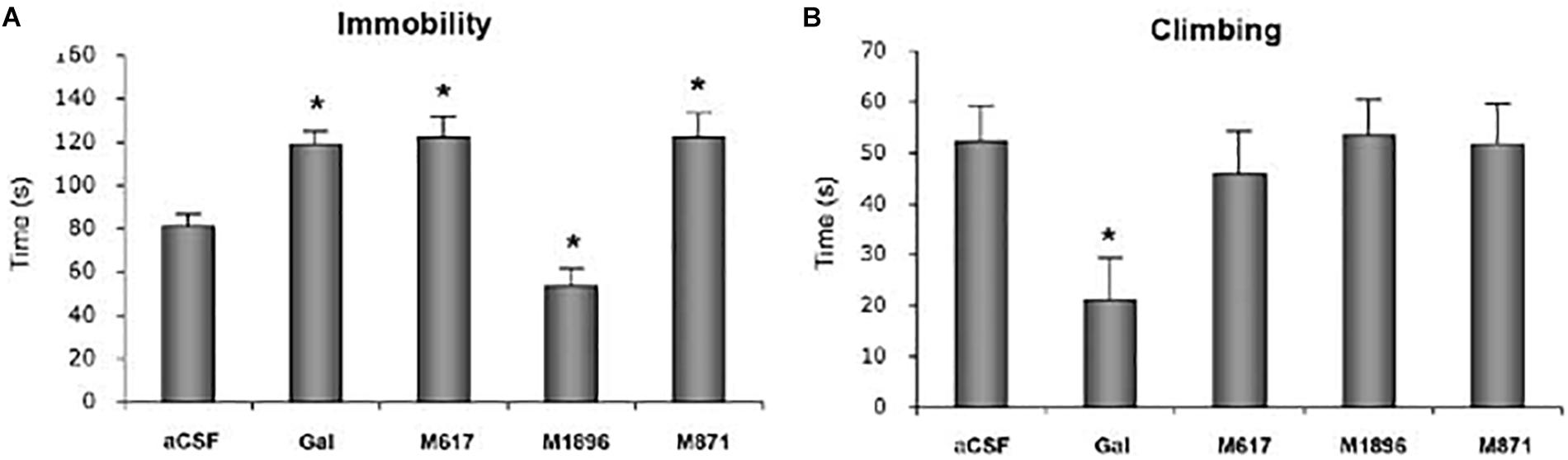
Figure 7. Time of immobility (A) and climbing (B) in the forced swim test (FST). Rats received i.c.v. infusion of aCSF, galanin (Gal), the GalR1 receptor agonist M617, the GalR2(R3) agonist AR-M1896 or the GalR2 antagonist M871 (M871) 20 min prior to a 5 min test. Data presented as mean ±SEM. significant difference from the control swim group; ∗one-way ANOVA, Fisher’s PLSD. Galanin, the GalR1 agonist and the GalR2 antagonist increase the immobility time versus a decrease after the GalR2(3) agonist. From Kuteeva et al. (2008), with permission.
Taken together, galanin receptors GalR1 and GalR2 play a differential role in regulation of depression-like behavior. Thus, galanin exerts a prodepressive effect, presumably via GalR1, while stimulation of GalR2 has an antidepressant-like effect.
While little interest has been paid to GalR3 in relation to mood, possibly due to its low expression in the rat (Mennicken et al., 2002) and mouse (Lein et al., 2007) brain, a GalR3 knockout mouse exhibits an anxiety-like phenotype (Brunner et al., 2014).
Following early studies on intra-membrane receptor-receptor interactions (Fuxe and Agnati, 1985), receptor di- and heteromerization have become a recognized mechanism for signaling through GPCRs (Bouvier, 2001; Devi, 2001; Agnati et al., 2003). Recent studies reveal that galanin receptor heteromers exist, introducing a further degree of complexity in interpreting galaninergic signaling in the brain (Fuxe et al., 2012), and in relation to mood control. Thus, in addition to GalR1 and 5-HT1A receptor heterodimers (Borroto-Escuela et al., 2010), the galanin (1–15) fragment alone induces strong depression- and anxiogenic-related effects and may regulate mood via binding to GalR1 and GalR2 heterocomplexes (Millon et al., 2014, 2017a,b). Interestingly, galanin (1-15) causes a dose-dependent hyperpolarization of a population of hippocampal CA3 neurons (Xu et al., 1999), and after iodination it binds to other regions including the dorsal hippocampus, as shown in autoradiographic studies (Hedlund et al., 1992). Taken together these results provide evidence for a functional role of galanin (1–15), perhaps unexpected in view of results showing a high affinity of the N-terminal galanin (1-16) fragment to galanin binding sites in the brain (Fisone et al., 1989).
The LC and NA have since the 1960’s been a focus of clinical and preclinical monoamine research, because of their involvement in stress, mood control and treatment of mood disorders (Bunney and Davis, 1965; Schildkraut, 1965; Weiss et al., 1981, 1994; Svensson, 1987; Simson and Weiss, 1988; Page and Valentino, 1994; Schatzberg and Schildkraut, 1995; Aston-Jones et al., 1996; Bremner et al., 1996a,b; Harro and Oreland, 2001; Charney, 2004; Millan, 2006; Samuels and Szabadi, 2008; Seki et al., 2018).
There is a strong relationship between stress and the LC: stress increases NA turnover, as well as tyrosine hydroxylase activity and transcription in the LC (Korf et al., 1973; Zigmond et al., 1974; Abercrombie and Jacobs, 1987; Komori et al., 1990; Smith et al., 1991; Melia et al., 1992; Aston-Jones et al., 1996; Rusnak et al., 1998; Chang et al., 2000; McDevitt et al., 2009; Ong et al., 2014; Kawa et al., 2015). Moreover, stress activation of LC neurons results in release of NA in the forebrain (Abercrombie et al., 1988; Jordan et al., 1994; Vahabzadeh and Fillenz, 1994; Ihalainen et al., 1999; Yoshitake et al., 2004) and cortical EEG activation, i.e., arousal (Page et al., 1993). Here CRF (Vale et al., 1981) is an important mediator of the stress-induced LC activation (Valentino and Van Bockstaele, 2015).
Also galanin expression is upregulated in LC neurons in response to stress/exercise (Holmes et al., 1995; Sweerts et al., 1999; O’Neal et al., 2001; Sciolino et al., 2012; Weinshenker and Holmes, 2016), establishing a relation between stress, NA and galanin in LC. Similarly, a single dose of the monoamine-depleting drug reserpine (Pletscher et al., 1955; Carlsson, 1975) causes an increase in galanin mRNA levels in LC neurons (Austin et al., 1990; Gundlach et al., 1990a). The same treatment results in a complete depletion of galanin in the noradrenergic cortical/hippocampal nerve terminals (Xu et al., 1998), that are the projections of the LC neurons: evidence for the view that release of a neuropeptide leads to increased synthesis of transcript and peptide. NPY expression in LC has, contrasting galanin, not been reported to be regulated by stress, but NPY mRNA is increased after reserpine administration (Gundlach et al., 1990a).
An involvement of LC in depression-like behavior has been studied by Weiss and colleagues focused on a link with the ascending mesencephalic dopamine system (Weiss et al., 1981, 1996, 1998, 2005). They based their experiments on the study by Grenhoff et al. (1993) showing that burst stimulation of LC inhibits DA neurons in the ventral tegmental area (VTA) (a.k.a the A10 group) (Dahlstrom and Fuxe, 1964). Weiss and colleagues have found that infusion of galanin into the VTA reduced exploratory behavior and increased immobility in the Porsolt test (a.k.a. Forced Swim Test, FST), an increase that was blocked by the galanin antagonist galantide. These findings link the LC-galanin system to studies showing involvement of the VTA and the reward system in stress and depression (Everitt and Robbins, 2005; Nestler and Carlezon, 2006; Thomas et al., 2008; Nestler, 2015; Pena et al., 2017).
The LC is involved in other mood-related behaviors such as addiction and reward (Maldonado and Koob, 1993), and galanin plays a role also in this context (Picciotto, 2008; Genders et al., 2018a). Thus, galanin binding and levels of GalR1 mRNA are increased in the LC during opiate withdrawal (Zachariou et al., 2003). Moreover, galanin-knockout mice exhibit more pronounced signs of opiate withdrawal, and galanin and the galanin ligand galnon both attenuate opiate reward and signs of withdrawal (Zachariou et al., 2003).
The distribution of galanin in the ‘normal’ human brain has been studied with RIA (Bennet et al., 1991; Barde et al., 2016), IHC (Chan-Palay, 1988a,b, 1990; Gentleman et al., 1989; Kowall and Beal, 1989; Beal et al., 1990; Kordower and Mufson, 1990; Kordower et al., 1992; Gabriel et al., 1994) and ISH (Miller et al., 1999; Le Maitre et al., 2013). In addition, the receptor distribution was analyzed with autoradiographic ligand binding methodology (Kohler et al., 1989; Kohler and Chan-Palay, 1990). Here, recent results obtained with ISH, qPCR and RIA on the galanin system in the LC and some other regions are summarized (Le Maitre et al., 2013; Barde et al., 2016). These studies were based on the identified gene sequences of the human galanin peptide and receptors (Evans and Shine, 1991; Jacoby et al., 1997; Lorimer et al., 1997; Fathi et al., 1998; Kolakowski et al., 1998; Smith et al., 1998).
The ISH analysis of the human LC (Le Maitre et al., 2013) revealed expression of TH, the rate-limiting enzyme for catecholamine synthesis in presumably all noradrenergic neurons (Figures 8A,B), galanin mRNA in many LC neurons (Figures 8C,D) and GalR3 mRNA in many, perhaps all neurons (Figures 8E,F), the latter two overlapping with the TH distribution. However, whereas the levels of TH and GalR3 mRNA are relatively similar in all cells, there was a large variation in the intensity of the galanin mRNA signal (c.f. Figures 8A,B,E,F with Figures 8C,D). This likely reflects the fact that galanin is a releasable molecule and that individual neurons are in different activity states. Note that the exposure time of the emulsion dipped slides is very different for the three markers (10 days for TH versus several months for GalR3), reflecting differences in mRNA levels (Figures 8A–F). Thus, GalR3 mRNA levels are very low, in agreement with low levels in rat (Mennicken et al., 2002) and potentially undetectable levels in mouse (Lein et al., 2007). In fact, GalR3 transcripts could only be visualized in human brains with very short post mortem delays prior to freezing (2–4 h).
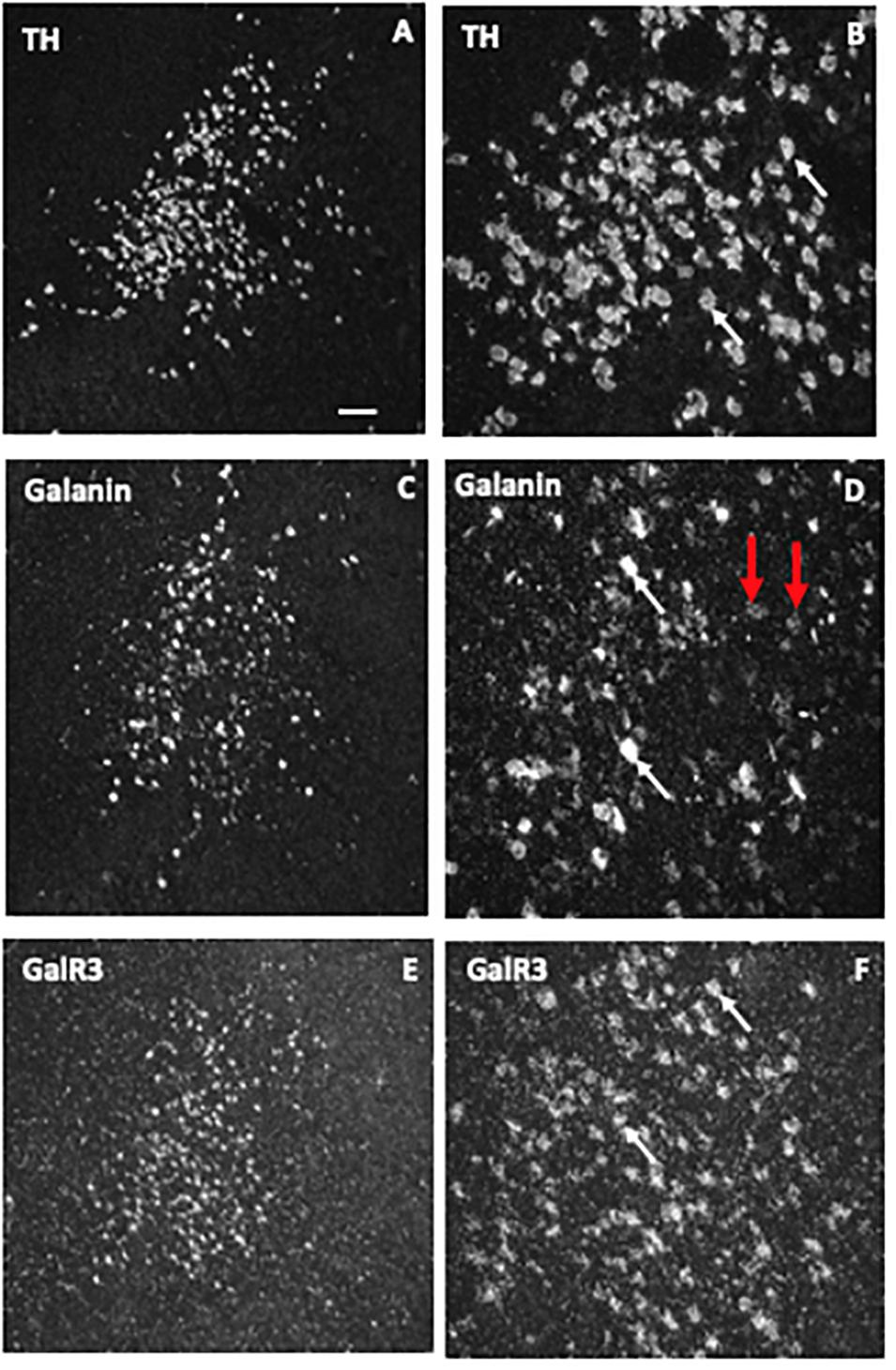
Figure 8. Dark-field ISH photomicrographs showing the distribution of transcripts for tyrosine hydroxylase (TH), galanin, and GalR3 in the locus coeruleus. The three markers TH (A,B), galanin (C,D), and GalR3 (E,F) show overlapping distribution patterns. TH and GalR3 transcript levels seem approximately similar in all cells. In contrast, there is variability in the strength of the signal for galanin mRNA (white arrow points to neurons with a strong signal, red ones to such with a weak signal). Exposure time: TH, 10 days; galanin, 4 weeks; GalR3, 8 weeks. This difference in exposure time transcript reflects difference in transcript levels, that is GalR3 mRNA levels are very low. Reprinted from Le Maitre et al. (2013) [Scale bars: 200 μM for (A,C,E); 50 μM for (B,D,F)].
Barde and colleagues analyzed > 200 postmortem brain samples from ‘normal’ (and depressed, see below) female and male subjects, including the following regions: in addition to LC, dorsolateral prefrontal cortex (DLPFC), anterior cingulate cortex (ACC), DRN and the medullary raphe nuclei (MRN)6 (Barde et al., 2016). Three methods were applied: RIA to monitor peptide levels, qPCR for transcript levels, and pyrosequencing to analyze DNA methylation. Comparable information is presented for ligand (galanin) (Table 1A) and transcripts (Table 1B) of galanin and GalR1-3 in the normal (control) and (‘depressed’) male and female brain (Barde et al., 2016).
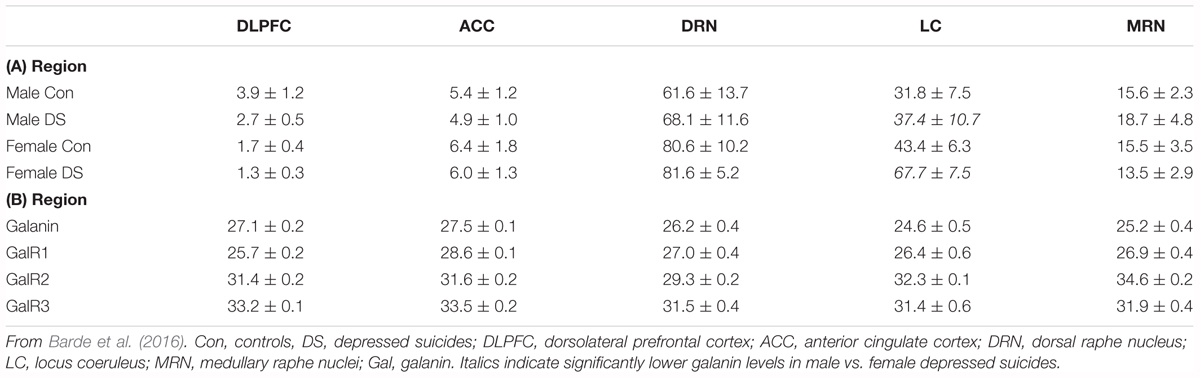
Table 1. (A,B) Concentration of galanin peptide (pmol/mg ± SEM) analyzed by RIA (A) and raw Ct values ± SEM monitored by qPCR (B) for male and female control and suicide samples from five brain regions.
When evaluating the results it should be noted that peptide levels in normal brains mostly reflect peptide present in nerve terminals and less so in cell bodies. Animal studies have shown that galanin peptide (like many other peptides) can best be detected in cell bodies after inhibition of axonal transport by colchicine (Rokaeus et al., 1984; Skofitsch and Jacobowitz, 1985; Melander et al., 1986c). In contrast, peptide transcripts are easily seen in cell bodies/dendrites, and often confined to these neuronal compartments.
With RIA, marked regional differences in galanin levels were observed, being highest in DRN (> LC > MRN > ACC = DLPFC), whereby DRN levels were 2 times higher than in LC and ∼12 times higher than in ACC/DLPFC. The qPCR analysis revealed the highest galanin mRNA levels in LC (reflecting the many positive cell bodies), about 4-fold higher than in DRN, and 6-fold higher than DLFPC, in agreement with the ISH results (Le Maitre et al., 2013) (Table 1A). Thus, the results from the LC suggest that there is a good translation from mRNA to peptide. The RIA results are generally in agreement with IHC studies on the primate brain (Kordower et al., 1992), and on the rat brain, when analyzed with RIA (Skofitsch and Jacobowitz, 1986) and IHC (Skofitsch and Jacobowitz, 1985; Melander et al., 1986b; Merchenthaler et al., 1993).
The results are also in line with the cited immunohistochemical results in the rat with a high density of galanin-positive nerve terminals in the DRN, and fewer in the LC (Skofitsch and Jacobowitz, 1985; Melander et al., 1986b; Merchenthaler et al., 1993). It is likely that galanin in cortical areas is present in thin and rather sparse afferents to the cortex, possibly originating in LC, as is the case in rat (Xu et al., 1998) and also in local neurons (see below).
With regard to receptors only transcripts and methylation were studied. This is partly due to a lack of specific antibodies for the galanin receptors, as discussed (Lu and Bartfai, 2009; Brunner et al., 2018), and there was no attempt to use Western blotting, Elisa or IHC for receptors. The strongest signal by far was noted for GalR1 mRNA, with the highest levels seen in DLPFC (> LC > MRN = DRN > ACC) (Table 1B). GalR1 levels in DLFPC were 2 times higher than in LC, and the GalR1 mRNA levels in DLFPC were 8-fold higher compared to the ‘adjacent’ ACC. GalR2 mRNA levels were in general considerably lower in LC (64-fold lower than GalR1). The GalR3 mRNA levels were low, although four times higher in the lower brain stem (LC, DRN) than in cortical regions, in agreement with the ISH results showing that GalR3 mRNA is present in NA neurons in LC, and possibly in 5-HT neurons in the DRN (Le Maitre et al., 2013).
In summary, GalR1 mRNA is the most prominent galanin receptor transcript in the human brain, including cortical regions. This is in agreement with early studies on human postmortem brain with iodinated galanin and autoradiography that revealed a distinct cortical signal (Kohler et al., 1989; Kohler and Chan-Palay, 1990), thus likely representing GalR1. The results differ from rat, since the early ligand binding studies in adult rat lacked binding in dorsal cortical areas (Skofitsch et al., 1986; Melander et al., 1988), and since GalR1 and -R2 mRNA levels are low (O’Donnell et al., 1999; Burazin et al., 2000). Thus, cortical receptor levels may represent another species difference not only between rat and human, but also between rat and mouse (Jungnickel and Gundlach, 2005). However, overall, GalR1 is also the most prominent galanin receptor in the rat brain (O’Donnell et al., 1999; Burazin et al., 2000) and likely also in mouse brain (Hohmann et al., 2003; Jungnickel and Gundlach, 2005; Lein et al., 2007). Waters and Krause (2000) monitored the levels of transcript for all three galanin receptors in the rat brain: GalR1 is highest in amygdala and spinal cord, whereas in cortex GalR2 > GalR1 > GalR3, and in hippocampus GalR2 > GalR1 = GalR3. In that study values were expressed as mean pg/25 μg total RNA.
The interpretation of the human qPCR results is not straight forward. For example, there is a lack of knowledge of the cellular localization of the transcripts in the prefrontal cortex regions, i.e., these areas have not been studied with ISH. It is likely that the transcripts are present in neurons, but a glial localization cannot be excluded (Butzkueven and Gundlach, 2010). Under certain circumstances galanin is expressed in specialized glial cells, e.g., after colchicine treatment alone or after spreading depression (Xu et al., 1992; Shen et al., 2003, 2005). The colchicine-induced signal was abolished by thyroidectomy (Calza et al., 1998). However, the results strongly suggest that galanin, and possibly all three galanin receptors, or at least GalR1, are expressed locally in cells, likely in cortical neurons. In contrast, ISH results are available for DRN and LC and reveal neuronal localization (Le Maitre et al., 2013). In particular, the results on the LC offer a possibility to form a hypothesis about the galanin system in this nucleus being involved in stress and genesis of depression, as discussed below.
Overall major differences exist between species, both with regard to galanin and galanin receptor expression, whereby galanin signaling seems to be more important for dorsal cortical functions in the human brain than in rodents. In contrast, in the rat ventral cortical areas, like entorhinal and piriform cortices, have abundant galanin receptor expression (Skofitsch et al., 1986; Melander et al., 1988; O’Donnell et al., 1999; Burazin et al., 2000), suggesting involvement in limbic processes.
Major depressive disorder (MDD) is a common and serious disease afflicting up to 2–5% (12-month prevalence; lifetime prevalence 10–15%) of the population worldwide, and women being more susceptible than men. Thus, MDD is a leading cause of disability worldwide associated with much suffering and major costs for society (Murray and Lopez, 1997; Kessler et al., 2003; Wittchen et al., 2011; Ferrari et al., 2013; World Health Organization [WHO], 2013). Adverse life events usually precede depression episodes, and experiences of physical and emotional abuse during early childhood and parental neglect are important predisposing vulnerability factors, strongly indicating that environmental psychosocial stressors are essential in pathogenesis (Kendler, 2012, 2013; Lutz et al., 2017; Tanti et al., 2017). The heritability is significant, about 35% (Sullivan et al., 2000). Interaction of genetic and environmental factors including stressful life events plays a major role in the development of MDD (Nestler et al., 2002; Akil, 2005; de Kloet et al., 2005; McEwen, 2007; McEwen et al., 2015). Epigenetic mechanisms through altered DNA methylation (Meaney and Ferguson-Smith, 2010; Zhang and Meaney, 2010) are probably involved, leading to stable changes in brain function that may underlie the psychopathology (Labonte et al., 2013; Vialou et al., 2013).
Over the last several decades two major hypotheses of the cause of unipolar depression have dominated, clinically associated with catecholamines (Bunney and Davis, 1965; Schildkraut, 1965; Schatzberg and Schildkraut, 1995) and with serotonin (Coppen, 1968; Maes and Meltzer, 1995). Pharmacological management of depression therefore often involves drugs that target the monoamine transporters, which include SSRIs, the transporter for noradrenaline (NA) (NRIs) or a combination of both (SNRIs) (Gardier et al., 1996; Mongeau et al., 1997; Millan, 2006), as well as a number of other medications (Berton and Nestler, 2006; Millan et al., 2015b). However, the therapeutic efficacy of these antidepressants is hampered by a slow onset of action, a limited response rate and considerable side effects (Montgomery, 2006; Trivedi et al., 2006). Of particular importance is the treatment resistant depression which affects some 20% of afflicted subjects (Akil et al., 2018). These issues have led to an intensive search for novel therapeutic approaches for MDD (Berton and Nestler, 2006; Artigas, 2015; Akil et al., 2018) (and see below), including targeting receptors for neuropeptides (Maubach et al., 1999; Hokfelt et al., 2003; Holmes et al., 2003; Nemeroff and Vale, 2005; Griebel and Holsboer, 2012), the most diverse family of brain messenger molecules (Burbach, 2010).
The evidence from animal experiments described led us to explore to what extent galanin may be involved in MDD and other mood disorders, and whether results from the analysis of human brain can guide the search for new antidepressants.
A candidate gene study of a cohort of European White ethnic origin totaling 2,361 from Manchester, United Kingdom and Budapest, Hungary was carried out (Juhasz et al., 2014) and revealed that variants in genes for galanin and its three receptors confer increased risk of depression and anxiety in people who experienced childhood adversity and/or recent negative life events (Figure 9). Genetic factors were only relevant in the moderate or high stress exposure groups when applying Bayesian multivariate analysis (Juhasz et al., 2014; Gonda et al., 2018). The rank order of the relevance of gene polymorphisms was GalR2 > GalR3 > GalR1 > galanin, with strong relevance for the first three in the moderately or highly exposed persons by recent negative life events in the last 12 months. All four were more relevant than the serotonin transporter gene-linked polymorphic region (5-HTTLPR) of the serotonin transporter gene. The effects were seen in the Manchester and the Budapest population, and in both males and females. This impact was seen only if taking stress into account, after medium and strong stress (the GalR2 gene) or strong stress (the GalR3, GalR2 and galanin genes), underlining the importance of environmental factors. In addition, the GalR2 gene polymorphism was more relevant than candidate gene polymorphisms of the genes for BDNF, the serotonin 1A receptor (HTR1A), the cannabinoid 1 receptor (CB1) and the serotonin 2A receptor (HTR2A) in the moderately stress exposed subjects (Gonda et al., 2018). The traditional analysis based on general linear models confirmed the gene-environment interaction; namely, no main effect of genes, but a significant modulatory effect of environment-induced development of depression were found.
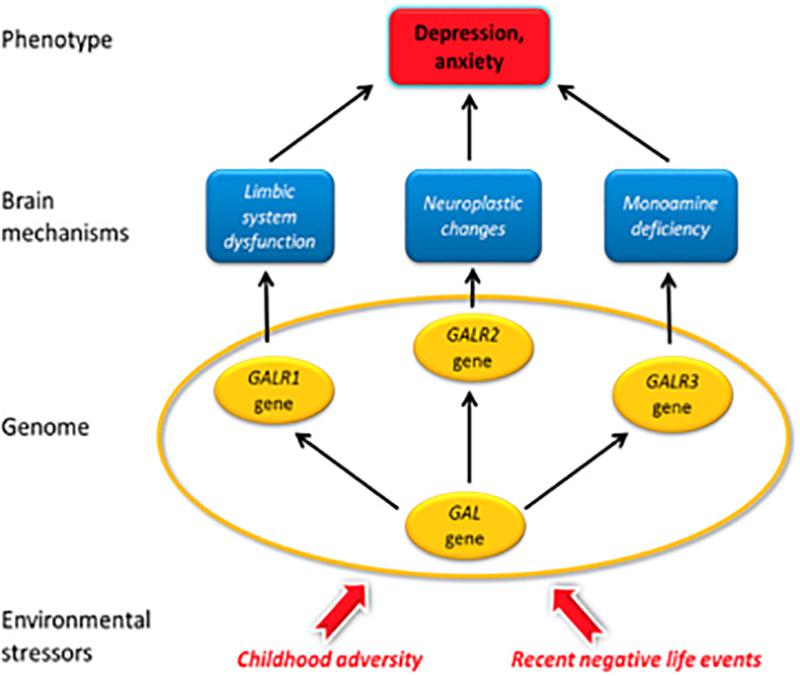
Figure 9. Galanin mechanisms hypothetically involved in MDD in humans. Galanin and its receptors are colocalized in some monoaminergic neurons in the brain. The galanin system is highly sensitive to experimental and naturalistic stressors. Stress-induced activation of the galanin system represents the first phase in the development of depression. Recent analysis of human brain has shown that the Gi protein-coupled GalR3 (and not GalR1 as in rodents) is the main galanin receptor in noradrenergic neurons in the locus coeruleus and probably the dorsal raphe nucleus and that the Gi protein-coupled GalR1 is the main receptor in the forebrain. Antidepressive effects may be achieved by (i) GalR3 antagonists, by reinstating normal monoamine turnover in LC neurons in the lower brainstem projecting to the forebrain, and by (ii) GalR1 antagonists in the forebrain by normalization of limbic system activity, or by (iii) agonists at GalR2, a Gq protein-coupled receptor, promoting neuroprotection. A candidate gene analysis suggests that GalR1 risk variants may compromise galanin signaling during childhood, whereas GalR2 signaling may be influenced by recent negative life events. In addition, all four galanin system genes have relevant roles in the development of depression-related phenotypes in those persons who were highly exposed to life stressors. From Juhasz et al. (2014).
Evidence for collaboration between small neurotransmitter and neuropeptide in the development of depression was also identified in this study, namely a gene–gene–environment interaction between the GalR2 and 5-HTTLPR genes in strongly exposed persons (Gonda et al., 2018). This could be of interest regarding antidepressant drug targets. The expression of the GALR2 polymorphism is about 2.5 times higher compared to 5-HTTLPR. Currently, the most frequently used antidepressants are the SSRIs, NRIs and SNRIs. Preliminary pre-clinical results suggest that an SNRI (venlafaxine) does not alter the transcript levels of galanin and its receptors (Petschner et al., 2016). Other studies show that chronic treatment with SSRIs increases galanin mRNA levels in various brain regions (Christiansen et al., 2011; Rovin et al., 2012; Yamada et al., 2013). The higher relevance of the GalR2, GalR3 and GalR1 gene polymorphisms in stress-induced depression and the galanin system-independent effects of the currently used antidepressants suggest that novel antidepressants acting on GalR1-3 could be developed. Such compounds could perhaps be more effective in SSRI/SNRI non-responders.
For a long time, it has been assumed that there is an interaction between stressful life events and a polymorphism in the promoter region of the 5-HT transporter (5-HTT) gene (SLCA4) (Lesch et al., 1996; Caspi et al., 2003). In the Newmood cohort this effect was weak and not significant in most comparisons, when corrections for multiple testing were applied (Juhasz et al., 2015). Furthermore, Bayesian relevance analyses consistently failed to show relevance for 5-HTTLPR (Juhasz et al., 2014; Gonda et al., 2018). Parallel to these findings, a recent large meta-analysis could not confirm an interaction (Culverhouse et al., 2018). The findings on the galanin system provide evidence for a more robust and relevant effect of galanin system genes compared to 5-HTTLPR of the serotonin transporter gene (Juhasz et al., 2014; Gonda et al., 2018). The 5-HTT and GalR2 receptor act jointly in the development of depression (Gonda et al., 2018).
Involvement of galanin in depression is further supported by a gender-specific association of galanin polymorphisms with antidepressant treatment response (Unschuld et al., 2010) and by a study reporting an association of galanin and MDD in the Chinese Han population (Wang et al., 2013). In addition, the first large genome-wide association study (GWAS) obtained a suggestive association of GAL with MDD using a gene based test, which retained low association p-values in two additional independent cohorts (Wray et al., 2012). A very large GWAS failed to identify risk genes (Major Depressive Disorder Working Group of the Psychiatric Gwas Consortium, 2013), but recent advances in large MDD GWAS studies resulted in several SNPs being associated with MDD (e.g., Hyde et al., 2016; Okbay et al., 2016; Xiao et al., 2017; Wray et al., 2018), and provided further evidence that genetic risk for depression is a continuous measure that translate environmental adversities into depressive symptoms. Taken together, the genetic analysis of the four members of the galanin system genes are complemented by a study on postmortem brains from depressed suicides (Barde et al., 2016), strengthening an involvement of galaninergic mechanisms in depression, as discussed below.
Differences in levels of galanin peptide, and of transcripts for, and DNA methylation of, galanin and GalR1-3 between MDD patients and matched controls were observed in an analysis of > 200 postmortem samples from five male and female brain regions (DLPFC, ACC, DRN, LC, and MRN). The significant and selective differences and changes in the galanin system in depressed versus control brains are summarized in Table 2. The most pronounced changes were observed for galanin and GalR3 in the DLPFC, and for galanin and GalR3 in the DRN and LC, in males and females (examples of results in Figures 10A,B). In DRN and LC there was an upregulation of the transcripts, paralleled by a decrease in DNA methylation. The decrease in methylation in galanin and GalR3 was most pronounced in female DRN and in male and female LC. In DLPFC, galanin mRNA levels were decreased in males and increased in females, the only distinct sex difference observed in the study. The changes in GalR1 were also increased and confined to three regions, DLPFC (male and female), DRN (male) and MRN (male), versus no change in LC. No differences were seen with regard to GalR2, except a decrease in MRN, included as a control region. The complete lack of changes in ACC contrasts the dramatic alterations in DLPFC, both regions belonging to the prefrontal cortex complex.
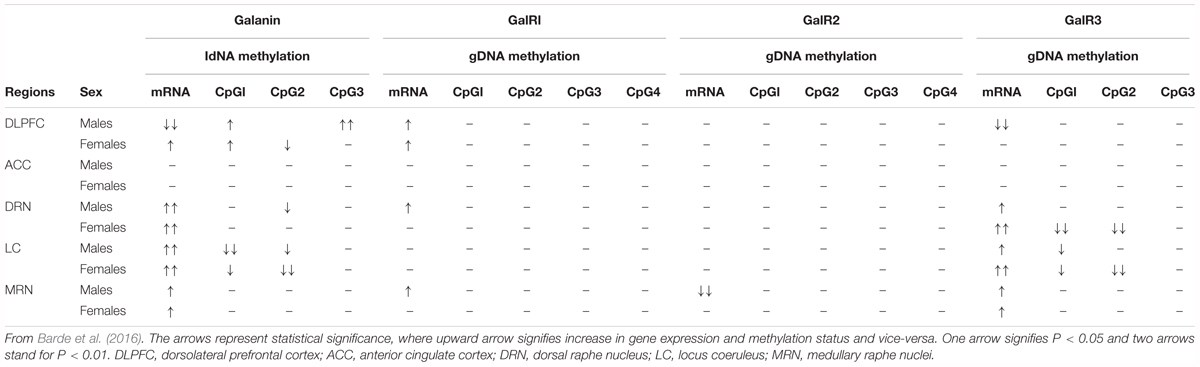
Table 2. Overview of mRNA and DNA methylation changes.
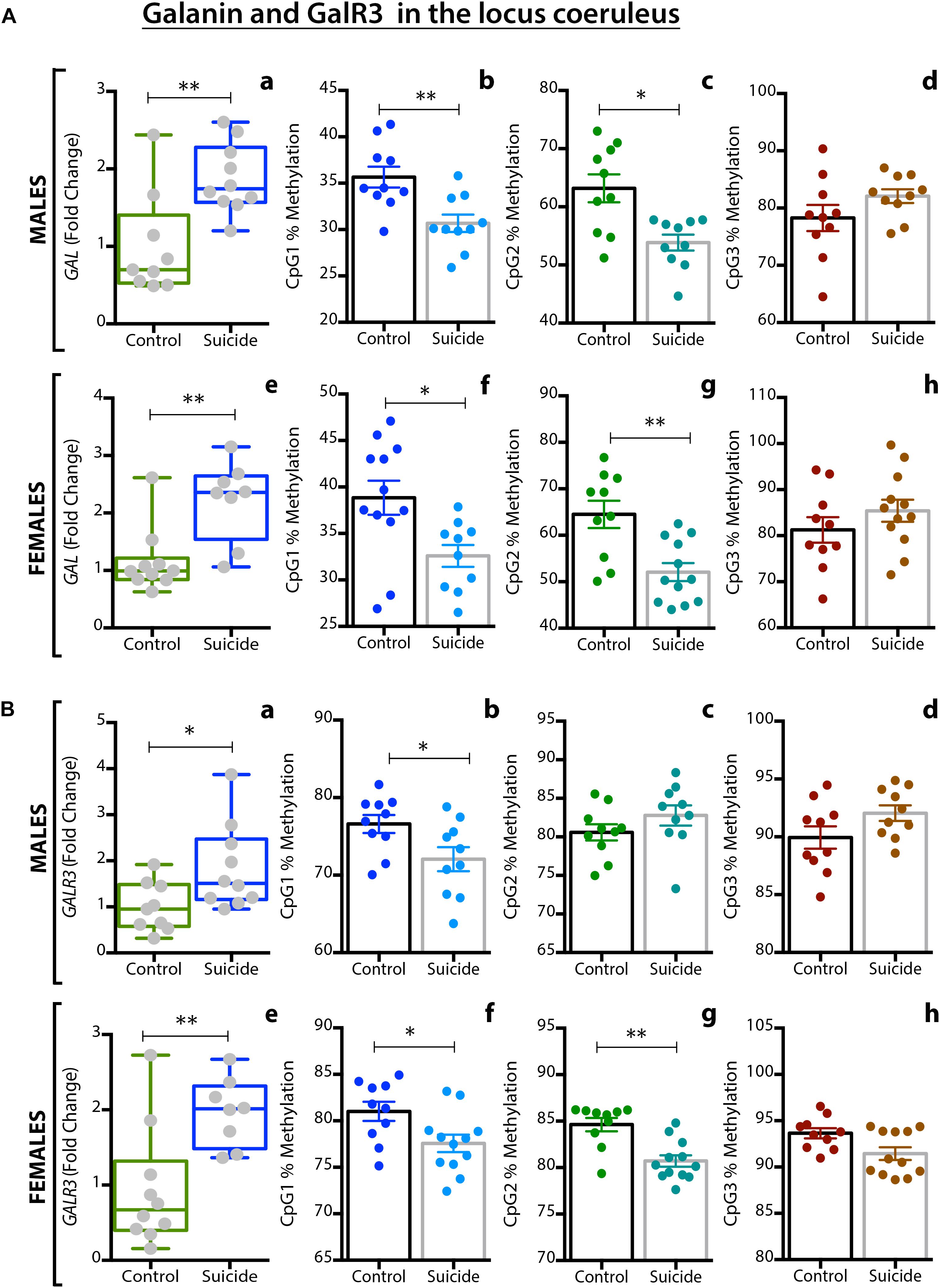
Figure 10. (A,B) Alterations in galanin (A) and GalR3 (B) gene expression and DNA methylation in the locus coeruleus (LC) of male and female depressed subjects who committed suicide, as compared to matched controls. (a,e) Expression levels of the two genes in the LC of male (a) and female (e) controls and depressed suicide (DS) subjects. (b–d,f–h) Percentage of DNA methylation levels at individual CpG sites of the two genes in male (b–d) and female (f–h) controls and DS subjects. All data are presented as mean ± SEM; males: n = 10 controls, 10 DS subjects; females: n = 12 controls, 10 DS subjects. Significant differences between DS subjects and controls are indicated: ∗P < 0.05, ∗∗P < 0.01. CON, controls. From Barde et al. (2016).
With regard to methylation, changes were always opposite to those in transcript levels. This is in agreement with the general view that methylation suppresses transcript synthesis (Moore et al., 2013). The results lend further support for an involvement of epigenetic mechanisms in MDD (Mill and Petronis, 2007; Machado-Vieira et al., 2011; Vialou et al., 2013; Lolak et al., 2014; Lopizzo et al., 2015; Saavedra et al., 2016; Hoffmann et al., 2017; Nagy et al., 2018).
Taken together, the results suggest that galaninergic mechanisms, in several brain regions, are involved in MDD, and that epigenetic changes mediated by DNA methylation play an important role, in agreement with a candidate gene study (Juhasz et al., 2014).
The LC in humans is a compact (but less so than in rodents), ‘blue’ (pigmented) nucleus consisting of a total (both sides) of around 50,000 neurons (German et al., 1988; Baker et al., 1989; Chan-Palay and Asan, 1989; Miller et al., 1999; Szot et al., 2000). As in other mammalian species studied, galanin is expressed in a large proportion of the human noradrenergic neurons (Chan-Palay et al., 1990; Kordower et al., 1992; Miller et al., 1999; Le Maitre et al., 2013), suggesting conservation during evolution. However, there are differences with regard to receptors. Thus the GalR3 receptor seems to be the most prominent receptor in the LC, contrasting the robust expression of GalR1 and GalR2 in the rat LC (O’Donnell et al., 1999; Burazin et al., 2000). Neither GalR1 nor -R2 mRNA was, surprisingly, detected with ISH in human NA neurons, although a GalR1 signal was seen in the LC region and in other regions, suggesting that the probe was functional. However, it cannot be excluded the NA LC neurons contain lower levels of GalR1 than other types of neurons and thus escaped detection. Thus, a distinct species difference seems to exist. Another apparent species difference was the possible lack of galanin expression in the human 5-HT neurons (Le Maitre et al., 2013), as is the case also in the mouse (Larm et al., 2003; Kuteeva et al., 2004; Lein et al., 2007; Fu et al., 2010), versus a robust expression of galanin in rat 5-HT neurons (Melander et al., 1986c; Fuxe et al., 1990; Priestley et al., 1993; Xu and Hokfelt, 1997).
How and when neuropeptides and classic transmitters are released has been explored (see Lundberg and Hokfelt, 1983; Hokfelt, 1991; Lundberg, 1996). Neuropeptides, stored in LDCVs, are mainly released when neurons are firing at a high rate or in bursts, e.g., during stress. This release occurs extrasynaptically, and not only from nerve endings but also from soma/dendrites. Merging this information and the results from MDD patients (Barde et al., 2016) it is possible to generate a hypothesis how depression in humans, likely a (stress-related) subtype of MDD, may develop (Figure 11): Under normal circumstances LC neurons fire at low frequencies, releasing NA in cortical regions, acting on post- and pre-synaptic adrenoceptors. Under stress, when LC neurons fire in bursts, also galanin will be released, together with NA, from nerve endings and soma/dendrites of the NA LC neurons, galanin acting on somatic/dendritic GalR3 autoreceptors.
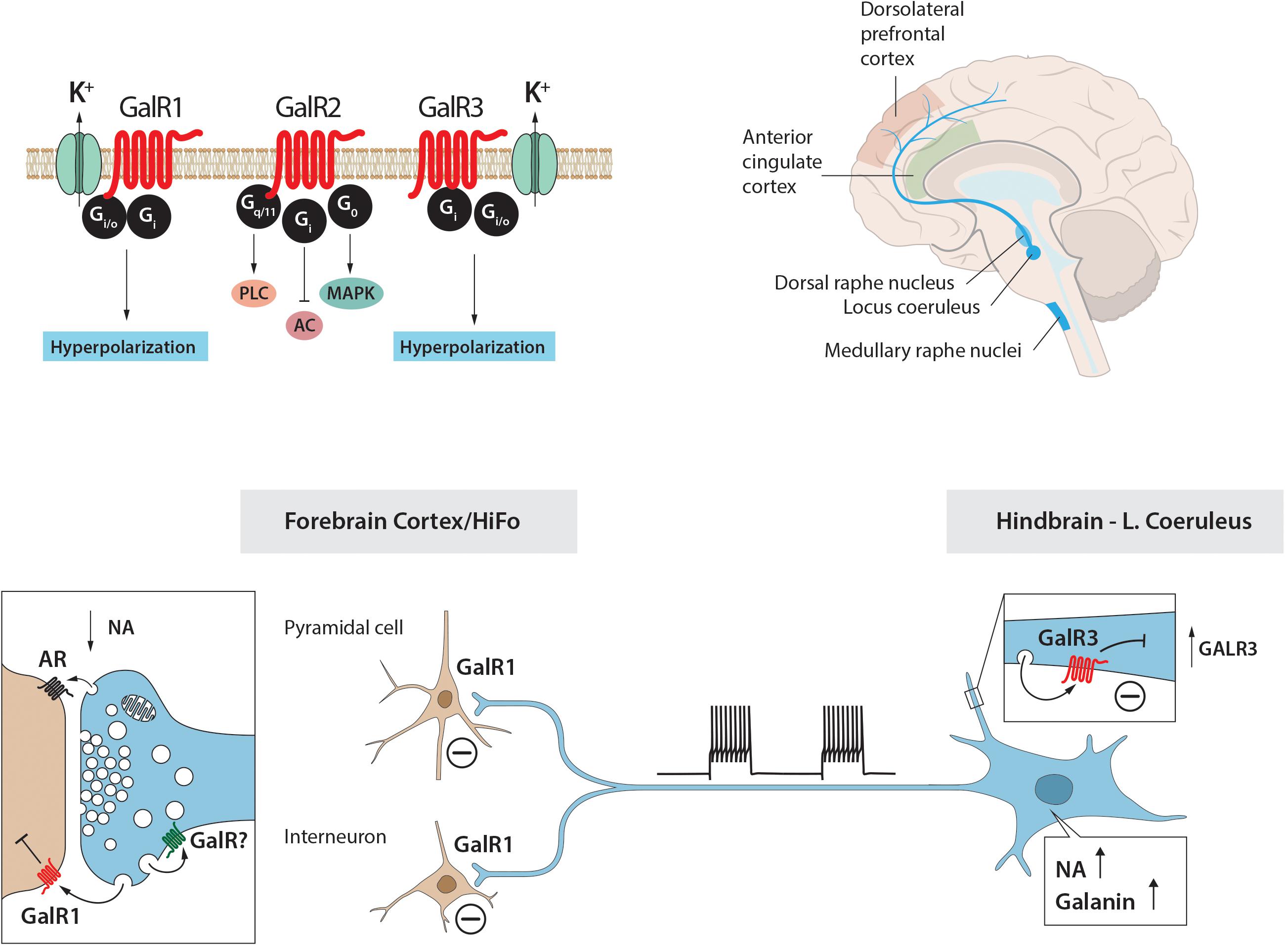
Figure 11. The galanin–locus coeruleus (LC) system in stress and depression: A hypothesis. The hypothesis is built on animal (rat) experiments showing that (i) galanin and GalR1 (top left) are present in LC NA neurons; (ii) galanin mRNA levels are increased during stress; (iii) galanin via GalR1 autoreceptors inhibits firing of LC neurons; and (iv) indirect evidence that galanin can be released from soma-dendrites of LC neurons. The second corner stone is results from two studies on human postmortem brain with ISH and qPCR. Five regions from postmortem brains from depressed subjects who committed suicide and controls were studied and are shown, including LC that projects to anterior cingulate and dorsolateral prefrontal cortices (top right). The results show that also in humans (i) the NA LC neurons express in any case galanin and GalR3 (top left). GalR1 and GalR3 probably have similar transduction mechanisms (top left). Under ‘normal’ firing only noradrenaline is released in forebrain. A situation after severe stress is depicted in the lower panel: LC neurons burst fire (lower panel, middle), NA and galanin are released from nerve endings in cortex (lower panel, left) and dendrites in the LC (lower panel, right), the latter in an attempt to prevent overexcitation (a resilience mechanism). To replace released peptide, galanin transcript levels and synthesis increase, and also GalR3 is upregulated (lower panel, right). The increased release, together with elevated galanin and GalR3 levels, result in a too strong inhibition and decreased NA levels in the forebrain (maladaptation) (lower panel, left), possibly contributing to depressive symptoms. HiFo, hippocampal formation. Drawing by Mattias Karlén.
GalR3 is, like GalR1, inhibitory (Smith et al., 1998) and causes hyperpolarization of the LC neurons, the purpose being to act as a ‘brake’ to prevent overexcitation, to keep the system in balance. This is similar to the proposed function of the 5-HT1A receptor as a “safety valve” of 5-HT neurons (Celada et al., 2013). As a consequence of increased firing and increased galanin release, synthesis of new peptide is initiated, reflected in increased mRNA levels. The fact that in dendrites sites of synthesis and release are close allows for rapid replacement. Thus, if mRNA is translated, increased galanin levels will be available for release from soma and dendrites, a feed-forward process. The additional increase in GalR3 transcript, presumably resulting in increased levels of receptor protein, could represent a robust increase in local galanin signaling. This seems unexpected, because intuitively one would expect downregulation of the receptor, following elevated levels of ligand. However, the inhibition may be strong and long-lasting leading to depletion of NA in the forebrain. The results suggest that mood disorders may be a consequence of a maladaption, an allostatic load (McEwen, 2003).
It may be emphasized that the prefrontal cortex has not been included in the discussion, even if significant changes in levels of galanin transcripts and methylation were recorded in this region of depressed subjects (Barde et al., 2016). This is because lack of knowledge about the cellular localization of the galanin system in this brain area. Finally, a similar scenario for an anti-depressive role of a GalR3 antagonist could be sketched for the 5-HT neurons in the DRN, since the galanin and GalR3 transcripts are upregulated both in the male and female DRN region from depressed patients who committed suicide, paralleled by decreased DNA methylation of the GalR3 gene in the female depressed subjects (Barde et al., 2016).
Of note, in the Barde et al. (2016) study the end stage of a mostly long development of the disorder is recorded, where all ‘resources’ have been mobilized to prevent overexcitation: increased ligand release plus increased receptor availability. Alternatively, the situation may reflect changes beyond patho-physiological regulatory mechanisms, especially when considering the considerable time it takes for depression to arise7.
Even if many humans are exposed to stress of various types and intensity, only comparatively few develop depression, thus displaying resilience to stress (Nestler et al., 2002; Southwick et al., 2005; Han and Nestler, 2017). Resilience appears to represent an active process involving several systems, including not only the mesolimbic dopamine neurons (Han and Nestler, 2017) but also other systems, such as the noradrenergic LC neurons (Charney, 2004; Feder et al., 2009; Krystal and Neumeister, 2009; Sciolino et al., 2015; Valentino and Van Bockstaele, 2015; Isingrini et al., 2016). Specific molecules, e.g., BDNF and neuropeptides like opioids and CRF, have also been implicated (Russo et al., 2012). Of particular interest in the present context is NPY (Kask et al., 2002; Morgan et al., 2002; Heilig, 2004; Krishnan et al., 2007; Zhou et al., 2008; Domschke et al., 2010; Cohen et al., 2012; Sabban et al., 2016; Kautz et al., 2017), a neuropeptide discovered in the Mutt laboratory (Tatemoto et al., 1982). NPY may be involved in the control of LC signaling in a similar way as galanin, but this will not be discussed further here, because limited information is available on the expression of NPY and NPY receptors in the brain of normal subjects and subjects who committed suicide.
The present data suggest that the GalR3-mediated ‘brake’ on the LC neurons is part of the resilience ‘machinery’ in humans. This is in agreement with animal experiments on rats by Sciolino et al. (2015) who show that exercise increases galanin levels in LC, and that exposure to stress reduces open arm exploration in sedentary rats. But this effect is not seen in exercise rats – and not in rats treated chronically with galanin given intraventricularly (i.c.v.); and it could be blocked by chronic administration of the galanin antagonist M40. Thus, increased galanin levels, presumably in LC, promotes resilience. This is also suggested by earlier animal experiments, as summarized by Sciolino et al. (2015): i.c.v. galanin protects against anxiety under stressful conditions (Bing et al., 1993), but not in the absence of stress (e.g. Holmes et al., 2005); i.c.v. M40 blocks fluoxetine-induced activity in the FST (Lu et al., 2005a); and transgenic mice overexpressing galanin under the dopamine B-hydroxylase promoter (the GalOE/D mouse), i.a. in LC, are protected against stress (Holmes et al., 2002).
The resilience hypothesis is further supported by comparison of the GalOE/D mouse (Steiner et al., 2001) with a mouse overexpressing galanin under the platelet growth factor B (PDGFB) promoter (the GalOE/P mouse) (Holmberg et al., 2005). Analyzing the effect of swim stress with microdialysis, it was shown that NA release in the forebrain is much lower in the GalOE/D than in the GalOE/P mouse (Yoshitake et al., 2004). The histochemical/qPCR analyses revealed that the galanin mRNA levels in the LC are 5 times higher in the GalOE/D than in wild type mice (Steiner et al., 2001). On the other hand, the GalOE/P mouse has a lower galanin expression in the LC and in (noradrenergic) nerve terminals in the forebrain (Kuteeva et al., 2004; Yoshitake et al., 2004). One possible explanation is that the higher galanin levels in the GalOE/D mouse suppress NA release in the forebrain by autoinhibition of the LC neurons, in agreement with the hypothesis.
The behavioral analyses of these two mouse lines reveal that the GalOE/P mouse displays an increased time of immobility in the FST, that is a depression-like behavior (Kuteeva et al., 2005). In contrast, there are no differences in immobility time on tail suspension between GalOE/D and wild type mice (Holmes et al., 2005). One interpretation is that the GalOE/D mouse is resilient to stress thanks to increased inhibitory galanin signaling in the LC, again, in agreement with the hypothesis.
The fact that neuropeptides routinely signal via GPCRs is promising from the perspective of drug development, since >30% of all prescription drugs act via such receptors (Luttrell et al., 2015; Hauser et al., 2017; Santos et al., 2017). Thus, >200 neuropeptide receptors are potential drug targets.
Peptide signaling in the rodent, and possibly primate, brain likely always means co-transmission with one or more small molecule transmitters – and other peptides. How could this affect treatment of patients? Are there any problems, or even advantages? Here some thoughts.
To fully block signaling at least two antagonists may be required. For example, a substance P antagonist is potentially a pain killer, since this excitatory peptide is released from nociceptors. However, the clinical tests failed for the reasons discussed (Hill, 2000; Herbert and Holzer, 2002). An obvious explanation would be that at least two additional excitatory transmitters are co-released with substance P from the same nerve endings in the dorsal horn: glutamate and CGRP, which are co-stored together with substance P in the same LDCVs (Merighi, 2002) (Figure 1E). So, blockade of one (the NK1) receptor may not be sufficient to achieve analgesia.
There could be advantages with peptide transmitters: First, neuropeptides are ’weak’ messengers. Thus, an antagonist will not have the potentially detrimental effects of blocking ‘strong’ and functionally essential fast transmitters, like those for GABA and glutamate. Although glutamate antagonists have many obvious indications for treating disease, it has been difficult to develop clinically efficacious and safe medicines. For example, glutamate is the major transmitter in nociceptors, and glutamate ligands have been strong candidates for pain treatment (Neugebauer, 2007), but glutamate antagonists like ketamine have yet to emerge as a clinically safe and widely used treatment alternative for pain (Bell et al., 2017). This said, and important in the context of the present review (MDD), the introduction of ketamine and ketamine analogs causing rapid antidepressant effects in subjects with treatment resistant depression has been, to say the least, an exciting advance (Zarate et al., 2006; Abdallah et al., 2015; Lener et al., 2017).
Furthermore, if neuropeptides are only released when neurons are firing at high frequency or firing in bursts, then antagonists would only have an effect under these circumstances, that is only affecting an activated system. For example, galanin is present in >20 nuclei in the rat brain. However, stress may primarily activate LC and 5-HT neurons, which in rats may be the only neurons releasing galanin. And an antagonist will consequently antagonize only the effects of galanin released from these two systems. The remaining galanin systems are silent and will therefore not be affected by the antagonist – so likely only few side effects. In contrast, a NA reuptake inhibitor will affect all NA neurons, resulting in increased extracellular amine levels in virtually all brain regions, probably leading to side effects. The same is of course true for SSRIs and serotonin, as well as for SNRIs and serotonin plus NA.
The discovery by the pharmaceutical company Merck of small non-peptide molecules passing the blood-brain-barrier (Uslaner et al., 2013) and acting as antagonists at orexin/hypocretin receptors (de Lecea et al., 1998; Sakurai et al., 1998) has resulted in a new medicine: Suvorex/Belsomra, approved by the federal drug administration (US FDA) for treatment of insomnia (Coleman et al., 2012; Yang, 2014): in less than 20 years from bench to patient! The small molecule co-transmitter in these orexin/hypocretin neurons is glutamate (Rosin et al., 2003), and these neurons also express the opioid peptide dynorphin (Chou et al., 2001). Furthermore, monoclonal antibodies8 to calcitonin gene-related peptide (CGRP) (Aimovig, erenumab), a peptide also present in nociceptors (Rosenfeld et al., 1983), are now approved by FDA and EMA for treatment of migraine (Silberstein et al., 2017; Edvinsson et al., 2018). In fact, antibodies to the CGRP receptor and CGRP antagonists are also efficacious in treatment of migraine (Silberstein et al., 2017; Edvinsson et al., 2018). Here some 35 years passed from the discovery to the clinic. Moreover, the NK1 antagonist (Aprepitant) mentioned in relation to depression is now used for treatment of chemotherapy-induced emesis (Pendergrass et al., 2004), a serendipitous finding
It has been difficult to generate small molecules that pass the blood-brain-barrier and act on central galanin receptors. Bartfai and associates made significant contributions, starting with chimeric peptide ligands (Bartfai et al., 1992). For several years, these were important tools in the galanin field, although they did not penetrate into the brain/spinal cord from the periphery. GalR3 antagonists (SNAP 39899 and related compounds) were then the first molecule acting on the brain after peripheral administration (Swanson et al., 2005; Barr et al., 2006; Konkel et al., 2006a,b). An allosteric modulator, a GalR2 agonist passing the blood-brain-barrier, was also reported (Lu et al., 2010), followed by further GalR2 ligands (Saar et al., 2013a,b). Several overviews of the field have been published (Mitsukawa et al., 2008; Hoyer and Bartfai, 2012; Webling et al., 2012; Freimann et al., 2015).
Based on the discussion above it appears that a GalR3 antagonist is a promising candidate for treatment of depression. Experiments in rats, suggest that the GalR1 receptor in LC also is a target for treatment of addiction (Picciotto, 2008; Genders et al., 2018a). However, in humans the correct receptor may be GalR3. In fact, in two genetic studies on alcoholism, both the galanin gene (Belfer et al., 2006) and, interestingly, the GalR3 gene, but not the other two receptor genes, have been implicated (Belfer et al., 2007). Of note, a GalR3 knockout mouse exhibits an alcohol-preferring phenotype (Genders et al., 2018b).
Why would a GalR3 antagonist be an advantageous choice over reuptake blockers? Analysis of regions of the postmortem MDD brains and controls (Table 1) reveals upregulation of galanin and GalR3 not only in the LC but also in the DRN (Barde et al., 2016). These changes are likely associated with higher levels of released galanin and of available receptors. Thus, a GalR3 antagonist could disinhibit blockade of two monoamine systems critical in mood disorders and restore both NA and 5-HT forebrain levels, relieving depressed mood. Since no changes are seen in the ACC, and since galanin and GalR3 are downregulated in the male DLPFC, these systems are likely ‘silent’. Thus, treatment with a GalR3 antagonist may overall have a high degree of selectivity with less side effects.
The key message of the present review is that the neuropeptide galanin and its subtype 3 (GalR3), both coexisting in noradrenergic LC neurons, are involved in MDD as part of the resilience machinery and GalR3 as a target for treatment. The hypothesis is based on solid and reproduced animal experiments from several laboratories. However, the translation to humans represents a major ‘jump,’ only involving one single (large) experiment on postmortem brains and a supporting candidate gene study. Thus, the hypothesis needs confirmation, preferably by other laboratories and methods. GWAS reports are negative, and no support based on imaging is published. A key experiment would be to label a GalR3 antagonist and carry out both in vitro autoradiography and positron emission tomography to analyze GalR3 binding sites/receptors. Moreover, the results are mainly based on transcript analysis, and it will be necessary to show translation into receptor protein, both in the rodent and human brain. The final proof would be to test a GalR3 antagonist in the clinical setting, but that would require generation of new, non-toxic molecules. Therefore, the retraction of major pharmaceutical companies from the neuroscience field represents a major disappointment. Further aspects on ‘limitations’ can be found in Barde et al. (2016).
The discovery of new drugs for treatment of mental illness has often been the result of serendipity (Celada et al., 2013; Millan et al., 2015a). The present review suggests that results from experimental animal studies can generate hypotheses that can be further validated by examining postmortem brains from relevant patient groups, perhaps leading to new pharmacological treatment strategies.
In animal studies the neuropeptide galanin has shown consistent changes in expression in response to a variety of stimuli, including stress. In agreement, results on the four galanin system genes (Juhasz et al., 2014), together with a recent study conducted with postmortem brains from depressed suicides (Barde et al., 2016), suggest involvement of galaninergic mechanisms in depression. On the basis of these studies, it is hypothesized that galanin may, via inhibitory GalR3 autoreceptors, act as a ‘brake’ to prevent overexcitation of LC neurons, representing a resilience mechanism to protect against depression. Galanin is, however, only one factor in a comprehensive network of built-in safeguards against overexcitation of LC neurons, reflecting the functional importance of a strict control of noradrenergic LC neurons which project to virtually all parts of the central nervous system.
It is now some 45 years since fluoxetine was generated, and 35 years since the first monoamine (serotonin)-reuptake inhibitor (Zimelidine) was launched (Spector et al., 2018). Since then SSRIs, NRIs and SNRIs have been the most widely used drugs for treatment of depression. Here we speculate that an antagonist at GalR3 receptors in noradrenergic LC neurons could lead to enhanced NA release in the forebrain – and recovery from disease. Since GalR3 is also likely present and upregulated in 5-HT neurons, it is possible that the same GalR3 antagonist also could normalize 5-HT release in the forebrain as well. Such a GalR3 antagonist has been developed (Swanson et al., 2005). However, the GalR3 antagonist SNAP 37889 has shown in vitro toxicity (Koller et al., 2016), and clinical trials have been terminated due to safety concerns.
If a GalR3 antagonist without toxic side effects will be developed and if ever tested in the clinic, the question might arise: Why would this drug be an advantage over a combined reuptake inhibitor, like Venlafaxine? Since the GalR3 antagonist works by a different mechanism it may, hypothetically, avoid some of the well-known side effects of reuptake inhibitors by a restricted site of action, versus the reuptake inhibitors increasing monoamine levels at all sites in the brain. It may be further speculated that the well-known delay of onset may be avoided, since the postulated disinhibition of the NA and 5-HT neurons via GalR3 antagonism should be a fast effect, perhaps without the complex ‘compensatory’ changes occurring after treatment with SSRIs and related to the 5-HT1A receptors (Celada et al., 2013). A third consideration is the consistently higher relevance of GalR3 gene variants compared to those of the serotonin transporter in stress-related depression, which might serve as a basis of personalized treatment. To what extent treatment resistant subjects will be helped is another issue that needs to be addressed. Finally, the use of agents acting at multiple sites, e.g., blocking monoamine re-uptake plus the NK1 receptor, may represent a way forward (Millan, 2009). In this respect, perhaps a drug combining blockade of a galanin receptor with another receptor/mechanism could represent an interesting alternative?
TH and SB wrote and revised the manuscript. GB, GJ, and NM wrote key sections of the manuscript. All authors contributed to manuscript revision, and read and approved the submitted versions.
The primary research from the host laboratory reviewed in this article was supported by the Swedish Research Council (04X-2887), the Marianne and Marcus Wallenberg Foundation, the European Community (NewMood, LSHM-CT-2004-503474; 2004-2008), Karolinska Institutet, AFA, the Swedish Brain Foundation, an Unrestricted Bristol-Myers-Squibb Neuroscience grant, NARSAD and the Torsten Söderberg Foundation.
TH has shares in Bioarctic, Stockholm, Sweden and Novo Nordisk, Copenhagen, Denmark.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Dr. Eric Kandel, Columbia University, New York, for critically reading and correcting the original draft of the manuscript, and Dr. Gilad Silberberg for valuable comments. We are particularly grateful to Reviewer AG for valuable suggestions and corrections of the final version. We express thanks to Drs. Marian DiFiglia (Figure 1D), Adalberto Merighi (Figure 1E), Dr. Mingdong Zhang and Ms. Joanne Bakker for allowing reproduction of photos (Figure 5).
Abbott, S. B., Kanbar, R., Bochorishvili, G., Coates, M. B., Stornetta, R. L., and Guyenet, P. G. (2012). C1 neurons excite locus coeruleus and A5 noradrenergic neurons along with sympathetic outflow in rats. J. Physiol. 590, 2897–2915. doi: 10.1113/jphysiol.2012.232157
Abdallah, C. G., Sanacora, G., Duman, R. S., and Krystal, J. H. (2015). Ketamine and rapid-acting antidepressants: a window into a new neurobiology for mood disorder therapeutics. Annu. Rev. Med. 66, 509–523. doi: 10.1146/annurev-med-053013-062946
Abercrombie, E. D., and Jacobs, B. L. (1987). Single-unit response of noradrenergic neurons in the locus coeruleus of freely moving cats. II. Adaptation to chronically presented stressful stimuli. J. Neurosci. 7, 2844–2848. doi: 10.1523/JNEUROSCI.07-09-02844.1987
Abercrombie, E. D., Keller, R. W. Jr., and Zigmond, M. J. (1988). Characterization of hippocampal norepinephrine release as measured by microdialysis perfusion: pharmacological and behavioral studies. Neuroscience 27, 897–904. doi: 10.1016/0306-4522(88)90192-3
Acher, R., and Chauvet, J. (1954). The structure of bovine vasopressin. Biochim. Biophys. Acta 14, 421–429. doi: 10.1016/0006-3002(54)90202-4
Aghajanian, G. K., Cedarbaum, J. M., and Wang, R. Y. (1977). Evidence for norepinephrine-mediated collateral inhibition of locus coeruleus neurons. Brain Res. 136, 570–577. doi: 10.1016/0006-8993(77)90083-X
Agnati, L. F., Ferre, S., Lluis, C., Franco, R., and Fuxe, K. (2003). Molecular mechanisms and therapeutical implications of intramembrane receptor/receptor interactions among heptahelical receptors with examples from the striatopallidal GABA neurons. Pharmacol. Rev. 55, 509–550. doi: 10.1124/pr.55.3.2
Agoston, D. V., Komoly, S., and Palkovits, M. (1994). Selective up-regulation of neuropeptide synthesis by blocking the neuronal activity: galanin expression in septohippocampal neurons. Exp. Neurol. 126, 247–255. doi: 10.1006/exnr.1994.1062
Ahmad, S., O’Donnell, D., Payza, K., Ducharme, J., Ménard, D., Brown, W., et al. (1998). Cloning and evaluation of the role of rat GALR-2, a novel subtype of galanin receptor, in the control of pain perception. Ann. N. Y. Acad. Sci. 21, 108–119. doi: 10.1111/j.1749-6632.1998.tb10688.x
Akil, H., Brenner, S., Kandel, E., Kendler, K. S., King, M. C., Scolnick, E., et al. (2010). Medicine. The future of psychiatric research: genomes and neural circuits. Science 327, 1580–1581. doi: 10.1126/science.1188654
Akil, H., Gordon, J., Hen, R., Javitch, J., Mayberg, H., McEwen, B., et al. (2018). Treatment resistant depression: a multi-scale, systems biology approach. Neurosci. Biobehav. Rev. 84, 272–288. doi: 10.1016/j.neubiorev.2017.08.019
Alhadeff, A. L., Su, Z., Hernandez, E., Klima, M. L., Phillips, S. Z., Holland, R. A., et al. (2018). A neural circuit for the suppression of pain by a competing need state. Cell 173, 140.e15–152.e15. doi: 10.1016/j.cell.2018.02.057
Allen, J. M., Adrian, T. E., Tatemoto, K., Polak, J. M., Hughes, J., and Bloom, S. R. (1982). Two novel related peptides, neuropeptide Y (NPY) and peptide YY (PYY) inhibit the contraction of the electrically stimulated mouse vas deferens. Neuropeptides 3, 71–77. doi: 10.1016/0143-4179(82)90001-4
Andersson, P. O., Bloom, S. R., Edwards, A. V., and Jarhult, J. (1982). Effects of stimulation of the chorda tympani in bursts on submaxillary responses in the cat. J. Physiol. 322, 469–483. doi: 10.1113/jphysiol.1982.sp014050
Andressen, C., Blumcke, I., and Celio, M. R. (1993). Calcium-binding proteins: selective markers of nerve cells. Cell Tissue Res. 271, 181–208. doi: 10.1007/BF00318606
Anisman, H., Du, L., Palkovits, M., Faludi, G., Kovacs, G. G., Szontagh-Kishazi, P., et al. (2008). Serotonin receptor subtype and p11 mRNA expression in stress-relevant brain regions of suicide and control subjects. J. Psychiatry Neurosci. 33, 131–141.
Armstrong, D. M., Ross, C. A., Pickel, V. M., Joh, T. H., and Reis, D. J. (1982). Distribution of dopamine-, noradrenaline-, and adrenaline-containing cell bodies in the rat medulla oblongata: demonstrated by the immunocytochemical localization of catecholamine biosynthetic enzymes. J. Comp. Neurol. 212, 173–187. doi: 10.1002/cne.902120207
Artigas, F. (2015). Developments in the field of antidepressants, where do we go now? Eur. Neuropsychopharmacol. 25, 657–670. doi: 10.1016/j.euroneuro.2013.04.013
Artigas, F., Bel, N., Casanovas, J. M., and Romero, L. (1996). Adaptative changes of the serotonergic system after antidepressant treatments. Adv. Exp. Med. Biol. 398, 51–59. doi: 10.1007/978-1-4613-0381-7_6
Asahina, K., Watanabe, K., Duistermars, B. J., Hoopfer, E., Gonzalez, C. R., Eyjólfsdóttir, E. A., et al. (2014). Tachykinin-expressing neurons control male-specific aggressive arousal in Drosophila. Cell 156, 221–235. doi: 10.1016/j.cell.2013.11.045
Aston, C., Jiang, L., and Sokolov, B. P. (2005). Transcriptional profiling reveals evidence for signaling and oligodendroglial abnormalities in the temporal cortex from patients with major depressive disorder. Mol. Psychiatry 10, 309–322. doi: 10.1038/sj.mp.4001565
Aston-Jones, G., Rajkowski, J., Kubiak, P., Valentino, R. J., and Shipley, M. T. (1996). Role of the locus coeruleus in emotional activation. Prog. Brain Res. 107, 379–402. doi: 10.1016/S0079-6123(08)61877-4
Aston-Jones, G., Shipley, M. T., Chouvet, G., Ennis, M., van Bockstaele, E., Pieribone, V., et al. (1991). Afferent regulation of locus coeruleus neurons: anatomy, physiology and pharmacology. Prog. Brain Res. 88, 47–75. doi: 10.1016/S0079-6123(08)63799-1
Aston-Jones, G., Shipley, M. T., and Grzanna, R. (1995). “Locus coeruleus, a5 and a7 noradrenergic cell groups,” in The Rat Nervous System, ed. G. Paxinos (San Diego, CA: Academic Press), 173–213.
Austin, M. C., Cottingham, S. L., Paul, S. M., and Crawley, J. N. (1990). Tyrosine hydroxylase and galanin mRNA levels in locus coeruleus neurons are increased following reserpine administration. Synapse 6, 351–357. doi: 10.1002/syn.890060407
Baimbridge, K. G., Celio, M. R., and Rogers, J. H. (1992). Calcium-binding proteins in the nervous system. Trends Neurosci. 15, 303–308. doi: 10.1016/0166-2236(92)90081-I
Baker, K. G., Tork, I., Hornung, J. P., and Halasz, P. (1989). The human locus coeruleus complex: an immunohistochemical and three dimensional reconstruction study. Exp. Brain Res. 77, 257–270. doi: 10.1007/BF00274983
Barde, S., Rüegg, J., Prud’homme, J., Ekstrom, T. J., Palkovits, M., Turecki, G., et al. (2016). Alterations in the neuropeptide galanin system in major depressive disorder involve levels of transcripts, methylation, and peptide. Proc. Natl. Acad. Sci. U.S.A. 113, E8472–E8481. doi: 10.1073/pnas.1617824113
Barde, Y. A. (1994). Neurotrophins: a family of proteins supporting the survival of neurons. Prog. Clin. Biol. Res. 390, 45–56.
Bargmann, C. I. (1993). Genetic and cellular analysis of behavior in C. elegans. Annu. Rev. Neurosci. 16, 47–71. doi: 10.1146/annurev.ne.16.030193.000403
Bargmann, W. (1949). [The neurosecretory connection between the hypothalamus and the neurohypophysis]. Z. Zellforsch. Mikrosk. Anat. 34, 610–634.
Bargmann, W., and Scharrer, E. (1951). The site of origin of the hormones of the posterior pituitary. Am. Sci. 39, 255–259.
Barr, A. M., Kinney, J. W., Hill, M. N., Lu, X., Biros, S. Jr., Rebek, J., et al. (2006). A novel, systemically active, selective galanin receptor type-3 ligand exhibits antidepressant-like activity in preclinical tests. Neurosci. Lett. 405, 111–115. doi: 10.1016/j.neulet.2006.06.033
Barrera, G., Echevarria, D. J., Poulin, J. F., Laforest, S., and Drolet, G. (2005). One for all or one for one: does co-transmission unify the concept of a brain galanin ”system” or clarify any consistent role in anxiety? Neuropeptides 39, 289–292. doi: 10.1016/j.npep.2004.12.008
Barry, J., Dubois, M. P., and Poulain, P. (1973). LRF producing cells of the mammalian hypothalamus. A fluorescent antibody study. Z. Zellforsch. Mikrosk. Anat. 146, 351–366. doi: 10.1007/BF02346227
Bartfai, T., Fisone, G., and Langel, U. (1992). Galanin and galanin antagonists: molecular and biochemical perspectives. Trends Pharmacol. Sci. 13, 312–317. doi: 10.1016/0165-6147(92)90098-Q
Bartfai, T., Iverfeldt, K., Fisone, G., and Serfozo, P. (1988). Regulation of the release of coexisting neurotransmitters. Annu. Rev. Pharmacol. Toxicol. 28, 285–310. doi: 10.1146/annurev.pa.28.040188.001441
Beal, M. F., MacGarvey, U., and Swartz, K. J. (1990). Galanin immunoreactivity is increased in the nucleus basalis of Meynert in Alzheimer’s disease. Ann. Neurol. 28, 157–161. doi: 10.1002/ana.410280207
Beas, B. S., Wright, B. J., Skirzewski, M., Leng, Y., Hyun, J. H., Koita, O., et al. (2018). The locus coeruleus drives disinhibition in the midline thalamus via a dopaminergic mechanism. Nat. Neurosci. 21, 963–973. doi: 10.1038/s41593-018-0167-4
Belfer, I., Hipp, H., Bollettino, A., McKnight, C., Evans, C., Virkkunen, M., et al. (2007). Alcoholism is associated with GALR3 but not two other galanin receptor genes. Genes Brain Behav. 6, 473–481. doi: 10.1111/j.1601-183X.2006.00275.x
Belfer, I., Hipp, H., McKnight, C., Evans, C., Buzas, B., Bollettino, A., et al. (2006). Association of galanin haplotypes with alcoholism and anxiety in two ethnically distinct populations. Mol. Psychiatry 11, 301–311. doi: 10.1038/sj.mp.4001768
Bell, R. F., Eccleston, C., and Kalso, E. A. (2017). Ketamine as an adjuvant to opioids for cancer pain. Cochrane Database Syst. Rev. 6:CD003351. doi: 10.1002/14651858.CD003351.pub3
Bellido, I., Diaz-Cabiale, Z., Jimenez-Vasquez, P. A., Andbjer, B., and Mathe, A. A. (2002). Increased density of galanin binding sites in the dorsal raphe in a genetic rat model of depression. Neurosci. Lett. 317, 101–105. doi: 10.1016/S0304-3940(01)02446-6
Bennett, M. R., and Lagopoulos, J. (2014). Stress and trauma: BDNF control of dendritic-spine formation and regression. Prog. Neurobiol. 112, 80–99. doi: 10.1016/j.pneurobio.2013.10.005
Bennet, W. M., Hill, S. F., Ghatei, M. A., and Bloom, S. R. (1991). Galanin in the normal human pituitary and brain and in pituitary adenomas. J. Endocrinol. 130, 463–467. doi: 10.1677/joe.0.1300463
Bernard, R., Kerman, I. A., Thompson, R. C., Jones, E. G., Bunney, W. E., Barchas, J. D., et al. (2011). Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol. Psychiatry 16, 634–646. doi: 10.1038/mp.2010.44
Berridge, C. W., and Waterhouse, B. D. (2003). The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res. Rev. 42, 33–84. doi: 10.1016/S0165-0173(03)00143-7
Berton, O., and Nestler, E. J. (2006). New approaches to antidepressant drug discovery: beyond monoamines. Nat. Rev. Neurosci. 7, 137–151. doi: 10.1038/nrn1846
Bing, O., Moller, C., Engel, J. A., Soderpalm, B., and Heilig, M. (1993). Anxiolytic-like action of centrally administered galanin. Neurosci. Lett. 164, 17–20. doi: 10.1016/0304-3940(93)90846-D
Bloem, B., Xu, L., Morava, E., Faludi, G., Palkovits, M., Roubos, E. W., et al. (2012). Sex-specific differences in the dynamics of cocaine- and amphetamine-regulated transcript and nesfatin-1 expressions in the midbrain of depressed suicide victims vs. controls. Neuropharmacology 62, 297–303. doi: 10.1016/j.neuropharm.2011.07.023
Boler, J., Enzmann, F., Folkers, K., Bowers, C. Y., and Schally, A. V. (1969). The identity of chemical and hormonal properties of the thyrotropin releasing hormone and pyroglutamyl-histidyl-proline amide. Biochem. Biophys. Res. Commun. 37, 705–710. doi: 10.1016/0006-291X(69)90868-7
Bondy, C. A., Gainer, H., and Russell, J. T. (1987). Effects of stimulus frequency and potassium channel blockade on the secretion of vasopressin and oxytocin from the neurohypophysis. Neuroendocrinology 46, 258–267. doi: 10.1159/000124829
Borroto-Escuela, D. O., Narvaez, M., Marcellino, D., Parrado, C., Narvaez, J. A., Tarakanov, A. O., et al. (2010). Galanin receptor-1 modulates 5-hydroxtryptamine-1A signaling via heterodimerization. Biochem. Biophys. Res. Commun. 393, 767–772. doi: 10.1016/j.bbrc.2010.02.078
Bost, A., Shaib, A. H., Schwarz, Y., Niemeyer, B. A., and Becherer, U. (2017). Large dense-core vesicle exocytosis from mouse dorsal root ganglion neurons is regulated by neuropeptide Y. Neuroscience 346, 1–13. doi: 10.1016/j.neuroscience.2017.01.006
Bouvier, M. (2001). Oligomerization of G-protein-coupled transmitter receptors. Nat. Rev. Neurosci. 2, 274–286. doi: 10.1038/35067575
Brahic, M., and Haase, A. T. (1978). Detection of viral sequences of low reiteration frequency by in situ hybridization. Proc. Natl. Acad. Sci. U.S.A. 75, 6125–6129. doi: 10.1073/pnas.75.12.6125
Branchek, T. A., Smith, K. E., Gerald, C., and Walker, M. W. (2000). Galanin receptor subtypes. Trends Pharmacol. Sci. 21, 109–117. doi: 10.1016/S0165-6147(00)01446-2
Brazeau, P., Ling, N., Bohlen, P., Esch, F., Ying, S. Y., and Guillemin, R. (1982). Growth hormone releasing factor, somatocrinin, releases pituitary growth hormone in vitro. Proc. Natl. Acad. Sci. U.S.A. 79, 7909–7913. doi: 10.1073/pnas.79.24.7909
Brazeau, P., Vale, W., Burgus, R., Ling, N., Butcher, M., Rivier, J., et al. (1973). Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179, 77–79. doi: 10.1126/science.179.4068.77
Bremner, J. D., Krystal, J. H., Southwick, S. M., and Charney, D. S. (1996a). Noradrenergic mechanisms in stress and anxiety: I. Preclinical studies. Synapse 23, 28–38. doi: 10.1002/(SICI)1098-2396(199605)23:1<28::AID-SYN4>3.0.CO;2-J
Bremner, J. D., Krystal, J. H., Southwick, S. M., and Charney, D. S. (1996b). Noradrenergic mechanisms in stress and anxiety: II. Clinical studies. Synapse 23, 39–51.
Broberger, C., Johansen, J., Johansson, C., Schalling, M., and Hokfelt, T. (1998). The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice. Proc. Natl. Acad. Sci. U.S.A. 95, 15043–15048. doi: 10.1073/pnas.95.25.15043
Brownstein, M., Arimura, A., Sato, H., Schally, A. V., and Kizer, J. S. (1975). The regional distribution of somatostatin in the rat brain. Endocrinology 96, 1456–1461. doi: 10.1210/endo-96-6-1456
Brownstein, M. J., and Mezey, E. (1986). Multiple chemical messengers in hypothalamic magnocellular neurons. Prog. Brain Res. 68, 161–168. doi: 10.1016/S0079-6123(08)60237-X
Brownstein, M. J., Saavedra, J. M., Axelrod, J., Zeman, G. H., and Carpenter, D. O. (1974). Coexistence of several putative neurotransmitters in single identified neurons of Aplysia. Proc. Natl. Acad. Sci. U.S.A. 71, 4662–4665. doi: 10.1073/pnas.71.12.4662
Brunner, S. M., Farzi, A., Locker, F., Holub, B. S., Drexel, M., Reichmann, F., et al. (2014). GAL3 receptor KO mice exhibit an anxiety-like phenotype. Proc. Natl. Acad. Sci. U.S.A. 111, 7138–7143. doi: 10.1073/pnas.1318066111
Brunner, S. M., Koller, A., Stockinger, J., Locker, F., Leis, S., Ernst, F., et al. (2018). Validation of antibody-based tools for galanin research. Peptides doi: 10.1016/j.peptides.2018.08.010 [Epub ahead of print].
Bunney, W. E. Jr., and Davis, J. M. (1965). Norepinephrine in depressive reactions. A review. Arch. Gen. Psychiatry 13, 483–494. doi: 10.1001/archpsyc.1965.01730060001001
Burazin, T. C., Larm, J. A., Ryan, M. C., and Gundlach, A. L. (2000). Galanin-R1 and -R2 receptor mRNA expression during the development of rat brain suggests differential subtype involvement in synaptic transmission and plasticity. Eur. J. Neurosci. 12, 2901–2917. doi: 10.1046/j.1460-9568.2000.00184.x
Burbach, J. P. (2010). Neuropeptides from concept to online database http://www.neuropeptides.nl/. Eur. J. Pharmacol. 626, 27–48. doi: 10.1016/j.ejphar.2009.10.015
Burgus, R., Dunn, T. F., Desiderio, D., Ward, D. N., Vale, W., and Guillemin, R. (1970). Characterization of ovine hypothalamic hypophysiotropic TSH-releasing factor. Nature 226, 321–325. doi: 10.1038/226321a0
Burnstock, G. (1976). Do some nerve cells release more than one transmitter? Neuroscience 1, 239–248.
Burnstock, G. (1978). Do some sympathetic neurones synthesize and release both noradrenaline and acetylcholine? Prog. Neurobiol. 11, 205–222. doi: 10.1016/0301-0082(78)90013-8
Burnstock, G. (2012). Purinergic signalling: its unpopular beginning, its acceptance and its exciting future. Bioessays 34, 218–225. doi: 10.1002/bies.201100130
Butzkueven, H., and Gundlach, A. L. (2010). Galanin in glia: expression and potential roles in the CNS. EXS 102, 61–69. doi: 10.1007/978-3-0346-0228-0_6
Calza, L., Giardino, L., and Hokfelt, T. (1998). Thyroid hormone-dependent regulation of galanin synthesis in neurons and glial cells after colchicine administration. Neuroendocrinology 68, 428–436. doi: 10.1159/000054393
Carlsson, A. (1975). Monoamine-depleting drugs. Pharmacol. Ther. B 1, 393–400. doi: 10.1016/0306-039X(75)90045-8
Carlsson, A., Falck, B., Hillarp, N. A., and Torp, A. (1962). Histochemical localization at the cellular level of hypothalamic noradrenaline. Acta Physiol. Scand. 54, 385–386. doi: 10.1111/j.1748-1716.1962.tb02364.x
Caspi, A., Sugden, K., Moffitt, T. E., Taylor, A., Craig, I. W., Harrington, H., et al. (2003). Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301, 386–389. doi: 10.1126/science.1083968
Castel, M., and Hochman, J. (1976). Ultrastructural immunohistochemical localization of vasopressin in the hypothalamic-neurohypophysial system of three murids. Cell Tissue Res. 174, 69–81. doi: 10.1007/BF00222151
Ceccatelli, S., Eriksson, M., and Hokfelt, T. (1989). Distribution and coexistence of corticotropin-releasing factor-, neurotensin-, enkephalin-, cholecystokinin-, galanin- and vasoactive intestinal polypeptide/peptide histidine isoleucine-like peptides in the parvocellular part of the paraventricular nucleus. Neuroendocrinology 49, 309–323. doi: 10.1159/000125133
Cedarbaum, J. M., and Aghajanian, G. K. (1976). Noradrenergic neurons of the locus coeruleus: inhibition by epinephrine and activation by the alpha-antagonist piperoxane. Brain Res. 112, 413–419. doi: 10.1016/0006-8993(76)90297-3
Cedarbaum, J. M., and Aghajanian, G. K. (1978). Afferent projections to the rat locus coeruleus as determined by a retrograde tracing technique. J. Comp. Neurol. 178, 1–16. doi: 10.1002/cne.901780102
Celada, P., Bortolozzi, A., and Artigas, F. (2013). Serotonin 5-HT1A receptors as targets for agents to treat psychiatric disorders: rationale and current status of research. CNS Drugs 27, 703–716. doi: 10.1007/s40263-013-0071-0
Chang, M. M., and Leeman, S. E. (1970). Isolation of a sialogogic peptide from bovine hypothalamic tissue and its characterization as substance P. J. Biol. Chem. 245, 4784–4790.
Chang, M. M., Leeman, S. E., and Niall, H. D. (1971). Amino-acid sequence of substance P. Nat. New Biol. 232, 86–87. doi: 10.1038/newbio232086a0
Chang, M. S., Sved, A. F., Zigmond, M. J., and Austin, M. C. (2000). Increased transcription of the tyrosine hydroxylase gene in individual locus coeruleus neurons following footshock stress. Neuroscience 101, 131–139. doi: 10.1016/S0306-4522(00)00352-3
Chan-Palay, V. (1988a). Galanin hyperinnervates surviving neurons of the human basal nucleus of Meynert in dementias of Alzheimer’s and Parkinson’s disease: a hypothesis for the role of galanin in accentuating cholinergic dysfunction in dementia. J. Comp. Neurol. 273, 543–557.
Chan-Palay, V. (1988b). Neurons with galanin innervate cholinergic cells in the human basal forebrain and galanin and acetylcholine coexist. Brain Res. Bull. 21, 465–472.
Chan-Palay, V., and Asan, E. (1989). Quantitation of catecholamine neurons in the locus coeruleus in human brains of normal young and older adults and in depression. J. Comp. Neurol. 287, 357–372. doi: 10.1002/cne.902870307
Chan-Palay, V., Jentsch, B., Lang, W., Höchli, M., and Asan, E. (1990). Distribution of neuropeptide y, c-terminal flanking peptide of npy and galanin and coexistence with catecholamine in the locus coeruleus of normal human, alzheimer’s dementia and parkinson’s disease brains. Dementia 1, 18–31.
Chan-Palay, V., Jonsson, G., and Palay, S. L. (1978). Serotonin and substance P coexist in neurons of the rat’s central nervous system. Proc. Natl. Acad. Sci. U.S.A. 75, 1582–1586. doi: 10.1073/pnas.75.3.1582
Chan-Palay, V., and Palay, S. (eds). (1984). Coexistence of Neuroactive Substances in Neurons. New York, NY: John Wiley and Sons.
Charney, D. S. (2004). Psychobiological mechanisms of resilience and vulnerability: implications for successful adaptation to extreme stress. Am. J. Psychiatry 161, 195–216. doi: 10.1176/appi.ajp.161.2.195
Chen, N. H., Reith, M. E., and Quick, M. W. (2004). Synaptic uptake and beyond: the sodium- and chloride-dependent neurotransmitter transporter family SLC6. Pflugers. Arch. 447, 519–531. doi: 10.1007/s00424-003-1064-5
Cheung, C. C., Hohmann, J. G., Clifton, D. K., and Steiner, R. A. (2001). Distribution of galanin messenger RNA-expressing cells in murine brain and their regulation by leptin in regions of the hypothalamus. Neuroscience 103, 423–432. doi: 10.1016/S0306-4522(01)00012-4
Chou, T. C., Lee, C. E., Lu, J., Elmquist, J. K., Hara, J., Willie, J. T., et al. (2001). Orexin (hypocretin) neurons contain dynorphin. J. Neurosci. 21:RC168. doi: 10.1523/JNEUROSCI.21-19-j0003.2001
Choudary, P. V., Molnar, M., Evans, S. J., Tomita, H., Li, J. Z., Vawter, M. P., et al. (2005). Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc. Natl. Acad. Sci. U.S.A. 102, 15653–15658. doi: 10.1073/pnas.0507901102
Christiansen, S. H., Olesen, M. V., Wortwein, G., and Woldbye, D. P. (2011). Fluoxetine reverts chronic restraint stress-induced depression-like behaviour and increases neuropeptide Y and galanin expression in mice. Behav. Brain Res. 216, 585–591. doi: 10.1016/j.bbr.2010.08.044
Chronwall, B. M., DiMaggio, D. A., Massari, V. J., Pickel, V. M., Ruggiero, D. A., O’Donohue, T. L., et al. (1985). The anatomy of neuropeptide-Y-containing neurons in rat brain. Neuroscience 15, 1159–1181. doi: 10.1016/0306-4522(85)90260-X
Cichon, J., Blanck, T. J. J., Gan, W. B., and Yang, G. (2017). Activation of cortical somatostatin interneurons prevents the development of neuropathic pain. Nat. Neurosci. 20, 1122–1132. doi: 10.1038/nn.4595
Cohen, H., Liu, T., Kozlovsky, N., Kaplan, Z., Zohar, J., and Mathé, A. A. (2012). The neuropeptide Y (NPY)-ergic system is associated with behavioral resilience to stress exposure in an animal model of post-traumatic stress disorder. Neuropsychopharmacology 37, 350–363. doi: 10.1038/npp.2011.230
Coleman, P. J., Schreier, J. D., Cox, C. D., Breslin, M. J., Whitman, D. B., Bogusky, M. J., et al. (2012). Discovery of [(2R,5R)-5-{[(5-Fluoropyridin-2-yl)oxy]methyl}-2-methylpiperidin-1-yl][5-methyl-2-(pyrimidin-2-yl)phenyl]methanone (MK-6096): A Dual Orexin Receptor Antagonist with Potent Sleep-Promoting Properties. ChemMedChem 7, 415–424. doi: 10.1002/cmdc.201200025
Consolo, S., Baldi, G., Russi, G., Civenni, G., Bartfai, T., and Vezzani, A. (1994). Impulse flow dependency of galanin release in vivo in the rat ventral hippocampus. Proc. Natl. Acad. Sci. U.S.A. 91, 8047–8051. doi: 10.1073/pnas.91.17.8047
Coons, A. H., Creech, H. J., Jones, R. N., and Berliner, E. (1942). The demonstration of pneumococcal antigen in tissues by the use of fluorescent antibody. J. Immunol. 45, 159–170.
Coppen, A. J. (1968). Depressed states and indolealkylamines. Adv. Pharmacol. 6, 283–291. doi: 10.1016/S1054-3589(08)60328-2
Cortes, R., Ceccatelli, S., Schalling, M., and Hokfelt, T. (1990a). Differential effects of intracerebroventricular colchicine administration on the expression of mRNAs for neuropeptides and neurotransmitter enzymes, with special emphasis on galanin: an in situ hybridization study. Synapse 6, 369–391.
Cortes, R., Villar, M. J., Verhofstad, A., and Hokfelt, T. (1990b). Effects of central nervous system lesions on the expression of galanin: a comparative in situ hybridization and immunohistochemical study. Proc. Natl. Acad. Sci. U.S.A. 87, 7742–7746.
Cottrell, G. A. (1976). Proceedings: does the giant cerebral neurone of Helix release two transmitters: ACh and serotonin? J. Physiol. 259, 44–45.
Crawley, J. N., Hommer, D. W., and Skirboll, L. R. (1984). Behavioral and neurophysiological evidence for a facilatory interaction between co-existing transmitters: cholecystokinin and dopamine. Neurochem. Int. 6, 755–760. doi: 10.1016/0197-0186(84)90007-X
Cuello, A. C. (ed.) (1982). Co-Transmission: Proceedings of a Symposium Held at Oxford During the 50th Anniversary Meeting of the British Pharmacological Society. Basingstoke: Palgrave Macmillan. doi: 10.1007/978-1-349-06239-3
Culverhouse, R. C., Saccone, N. L., Horton, A. C., Ma, Y., Anstey, K. J., Banaschewski, T., et al. (2018). Collaborative meta-analysis finds no evidence of a strong interaction between stress and 5-HTTLPR genotype contributing to the development of depression. Mol. Psychiatry 23, 133–142. doi: 10.1038/mp.2017.44
Dahlstrom, A., and Fuxe, K. (1964). Evidence for the existence of monoamine-containing neurons in the central nervous system. I. Demonstration of monoamines in the cell bodies of brain stem neurons. Acta Physiol. Scand. 62(Suppl. 232), 1–55.
De Camilli, P., and Jahn, R. (1990). Pathways to regulated exocytosis in neurons. Annu. Rev. Physiol. 52, 625–645. doi: 10.1146/annurev.ph.52.030190.003205
de Kloet, E. R., Joels, M., and Holsboer, F. (2005). Stress and the brain: from adaptation to disease. Nat. Rev. Neurosci. 6, 463–475. doi: 10.1038/nrn1683
de Kock, C. P., Wierda, K. D., Bosman, L. W., Min, R., Koksma, J. J., Mansvelder, H. D., et al. (2003). Somatodendritic secretion in oxytocin neurons is upregulated during the female reproductive cycle. J. Neurosci. 23, 2726–2734. doi: 10.1523/JNEUROSCI.23-07-02726.2003
de Lecea, L., Kilduff, T. S., Peyron, C., Gao, X., Foye, P. E., Danielson, P. E., et al. (1998). The hypocretins: hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. U.S.A. 95, 322–327. doi: 10.1073/pnas.95.1.322
de Souza, M. M., Silote, G. P., Herbst, L. S., Funck, V. R., Joca, S. R. L., Beijamini, V., et al. (2018). The antidepressant-like effect of galanin in the dorsal raphe nucleus of rats involves GAL2 receptors. Neurosci. Lett. 681, 26–30. doi: 10.1016/j.neulet.2018.05.029
de Wied, D., and Bohus, B. (1966). Long term and short term effects on retention of a conditioned avoidance response in rats by treatment with long acting pitressin and alpha-MSH. Nature 212, 1484–1486. doi: 10.1038/2121484a0
De Wied, D., and De Kloet, E. R. (1987). Pro-opiomelanocortin (Pomc) as homeostatic control-system. Ann. N. Y. Acad. Sci. 512, 328–337. doi: 10.1111/j.1749-6632.1987.tb24971.x
Descarries, L., and Saucier, G. (1972). Disappearance of the locus coeruleus in the rat after intraventricular 6-hydroxdopamine. Brain Res. 37, 310–316. doi: 10.1016/0006-8993(72)90676-2
Devi, L. A. (2001). Heterodimerization of G-protein-coupled receptors: pharmacology, signaling and trafficking. Trends Pharmacol. Sci. 22, 532–537. doi: 10.1016/S0165-6147(00)01799-5
Diaz-Cabiale, Z., Flores-Burgess, A., Parrado, C., Narvaez, M., Millon, C., Puigcerver, A., et al. (2014). Galanin receptor/neuropeptide y receptor interactions in the central nervous system. Curr. Protein Pept. Sci. 15, 666–672. doi: 10.2174/1389203715666140901111709
DiFiglia, M., Aronin, N., and Leeman, S. E. (1982). Light microscopic and ultrastructural localization of immunoreactive substance P in the dorsal horn of monkey spinal cord. Neuroscience 7, 1127–1139. doi: 10.1016/0306-4522(82)91120-4
Domschke, K., Dannlowski, U., Hohoff, C., Ohrmann, P., Bauer, J., Kugel, H., et al. (2010). Neuropeptide Y (NPY) gene: Impact on emotional processing and treatment response in anxious depression. Eur. Neuropsychopharmacol. 20, 301–309. doi: 10.1016/j.euroneuro.2009.09.006
Du, L., Merali, Z., Poulter, M. O., Palkovits, M., Faludi, G., Anisman, H., et al. (2014). Cortex of suicide brains. Prog. Neuropsychopharmacol. Biol. Psychiatry 50, 178–183. doi: 10.1016/j.pnpbp.2013.12.016
Du Vigneaud, V. (1954). Hormones of the posterior pituitary gland: oxytocin and vasopressin. Harvey Lect. 50, 1–26.
Dube, D., Leclerc, R., and Pelletier, G. (1976). Electron microscopic immunohistochemical localization of vasopressin and neurophysin in the median eminence of normal and adrenalectomized rats. Am. J. Anat. 147, 103–108. doi: 10.1002/aja.1001470109
Dubé, D., Leclerc, R., Pelletier, G., Arimura, A., and Schally, A. V. (1975). Immunohistochemical detection of growth hormone-release inhibiting hormone (somatostatin) in the guinea-pig brain. Cell Tissue Res. 161, 385–392. doi: 10.1007/BF00220006
Dubois, M. P., Barry, J., and Leonardelli, J. (1974). [Immunofluorescence demonstration and distribution of somatostatin (SRIF) in the median eminence of vertebrates (mammals, birds, amphibia, fishes)]. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 279, 1899–1902.
Eberwine, J., and Bartfai, T. (2011). Single cell transcriptomics of hypothalamic warm sensitive neurons that control core body temperature and fever response Signaling asymmetry and an extension of chemical neuroanatomy. Pharmacology & therapeutics 129, 241–259. doi: 10.1016/j.pharmthera.2010.09.010
Eccles, J. C. (1986). Chemical transmission and Dale’s principle. Prog. Brain Res. 68, 3–13. doi: 10.1016/S0079-6123(08)60227-7
Eccles, J. C., Fatt, P., and Koketsu, K. (1954). Cholinergic and inhibitory synapses in a pathway from motor-axon collaterals to motoneurones. J. Physiol. 126, 524–562. doi: 10.1113/jphysiol.1954.sp005226
Edvinsson, L., Haanes, K. A., Warfvinge, K., and Krause, D. N. (2018). CGRP as the target of new migraine therapies - successful translation from bench to clinic. Nat. Rev. Neurol. 14, 338–350. doi: 10.1038/s41582-018-0003-1
Eiden, L. E., Schafer, M. K., Weihe, E., and Schutz, B. (2004). The vesicular amine transporter family (SLC18): amine/proton antiporters required for vesicular accumulation and regulated exocytotic secretion of monoamines and acetylcholine. Pflugers. Arch. 447, 636–640. doi: 10.1007/s00424-003-1100-5
Ekblad, E., Edvinsson, L., Wahlestedt, C., Uddman, R., Hakanson, R., and Sundler, F. (1984). Neuropeptide Y co-exists and co-operates with noradrenaline in perivascular nerve fibers. Regul. Pept. 8, 225–235. doi: 10.1016/0167-0115(84)90064-8
Elde, R. P., and Parsons, J. A. (1975). Immunocytochemical localization of somatostatin in cell bodies of the rat hypothalamus. Am. J. Anat. 144, 541–548. doi: 10.1002/aja.1001440416
Erspamer, V., Melchiorri, P., Erspamer, C. F., and Negri, L. (1978). Polypeptides of the amphibian skin active on the gut and their mammalian counterparts. Adv. Exp. Med. Biol. 106, 51–64. doi: 10.1007/978-1-4684-7248-6_6
Evans, H. F., and Shine, J. (1991). Human galanin: molecular cloning reveals a unique structure. Endocrinology 129, 1682–1684. doi: 10.1210/endo-129-3-1682
Evans, S. J., Choudary, P. V., Neal, C. R., Li, J. Z., Vawter, M. P., Tomita, H., et al. (2004). Dysregulation of the fibroblast growth factor system in major depression. Proc. Natl. Acad. Sci. U.S.A. 101, 15506–15511. doi: 10.1073/pnas.0406788101
Everitt, B. J., Hokfelt, T., Terenius, L., Tatemoto, K., Mutt, V., and Goldstein, M. (1984). Differential co-existence of neuropeptide Y (NPY)-like immunoreactivity with catecholamines in the central nervous system of the rat. Neuroscience 11, 443–462. doi: 10.1016/0306-4522(84)90036-8
Everitt, B. J., and Robbins, T. W. (2005). Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat. Neurosci. 8, 1481–1489. doi: 10.1038/nn1579
Falck, B. (1962). Observations on the possibilities of the cellular localization of monoamines by a fluorescence method. Acta Physiol. Scand. Suppl. 56, 1–25.
Falck, B., Hillarp, N. A., Thieme, G., and Torp, A. (1962). Fluorescence of catecholamines and related compounds condensed with formaldehyde. J. Histochem. Cytochem. 10, 348–354. doi: 10.1177/10.3.348
Fathi, Z., Battaglino, P. M., Iben, L. G., Li, H., Baker, E., Zhang, D., et al. (1998). Molecular characterization, pharmacological properties and chromosomal localization of the human GALR2 galanin receptor. Brain Res. Mol. Brain Res. 58, 156–169. doi: 10.1016/S0169-328X(98)00116-8
Fathi, Z., Cunningham, A. M., Iben, L. G., Battaglino, P. B., Ward, S. A., Nichol, K. A., et al. (1997). Cloning, pharmacological characterization and distribution of a novel galanin receptor. Brain Res. Mol. Brain Res. 51, 49–59. doi: 10.1016/S0169-328X(97)00210-6
Feder, A., Nestler, E. J., and Charney, D. S. (2009). Psychobiology and molecular genetics of resilience. Nat. Rev. Neurosci. 10, 446–457. doi: 10.1038/nrn2649
Ferrari, A. J., Charlson, F. J., Norman, R. E., Patten, S. B., Freedman, G., Murray, C. J., et al. (2013). Burden of depressive disorders by country, sex, age, and year: findings from the global burden of disease study 2010. PLoS Med. 10:e1001547. doi: 10.1371/journal.pmed.1001547
Fisone, G., Berthold, M., Bedecs, K., Unden, A., Bartfai, T., Bertorelli, R., et al. (1989). N-terminal galanin-(1-16) fragment is an agonist at the hippocampal galanin receptor. Proc. Natl. Acad. Sci. U.S.A. 86, 9588–9591. doi: 10.1073/pnas.86.23.9588
Foote, S. L., Bloom, F. E., and Aston-Jones, G. (1983). Nucleus locus ceruleus: new evidence of anatomical and physiological specificity. Physiol. Rev. 63, 844–914. doi: 10.1152/physrev.1983.63.3.844
Freimann, K., Kurrikoff, K., and Langel, U. (2015). Galanin receptors as a potential target for neurological disease. Expert Opin. Ther. Targets 19, 1665–1676. doi: 10.1517/14728222.2015.1072513
Frick, A., Ahs, F., Palmquist, A. M., Pissiota, A., Wallenquist, U., Fernandez, M., et al. (2016). Alterations in the serotonergic and substance P systems in posttraumatic stress disorder. Mol. Psychiatry 21:1323. doi: 10.1038/mp.2016.159
Fried, G., Terenius, L., Hokfelt, T., and Goldstein, M. (1985). Evidence for differential localization of noradrenaline and neuropeptide Y in neuronal storage vesicles isolated from rat vas deferens. J. Neurosci. 5, 450–458. doi: 10.1523/JNEUROSCI.05-02-00450.1985
Fu, W., Le Maitre, E., Fabre, V., Bernard, J. F., David, XuZ. Q, Hökfelt, T., et al. (2010). Chemical neuroanatomy of the dorsal raphe nucleus and adjacent structures of the mouse brain. J. Comp. Neurol. 518, 3464–3494. doi: 10.1002/cne.22407
Furshpan, E. J., Macleish, P. R., Olague, P. H., and Potter, D. D. (1976). Chemical transmission between rat sympathetic neurons and cardiac myocytes developing in microcultures - evidence for cholinergic, adrenergic, and dual-function neurons. Proc. Natl. Acad. Sci. U.S.A. 73, 4225–4229. doi: 10.1073/pnas.73.11.4225
Fuxe, K., and Agnati, L. F. (1985). Receptor-receptor interactions in the central nervous system. A new integrative mechanism in synapses. Med. Res. Rev. 5, 441–482. doi: 10.1002/med.2610050404
Fuxe, K., Agnati, L. F., Kohler, C., Kuonen, D., Ogren, S. O., Andersson, K., et al. (1981). Characterization of normal and supersensitive dopamine receptors: effects of ergot drugs and neuropeptides. J. Neural. Transm. 51, 3–37. doi: 10.1007/BF01664003
Fuxe, K., Agnati, L., Von Euler, G., Lundgren, K., Zoli, M., Bjelke, B., et al. (1990). “Galanin/5-HT receptor interactions. a new integrative mechanism in the control of 5-ht neurotransmission in the central nervous system,” in Serotonin, eds R. V. H. Paoletti, P. Brunello, and N. Maggi (Netherlands: Springer), 169–185. doi: 10.1007/978-94-009-1912-9_23
Fuxe, K., Borroto-Escuela, D. O., Romero-Fernandez, W., Tarakanov, A. O., Calvo, F., Garriga, P., et al. (2012). On the existence and function of galanin receptor heteromers in the central nervous system. Front. Endocrinol. 3:127. doi: 10.3389/fendo.2012.00127
Fuxe, K., Dahlstrom, A. B., Jonsson, G., Marcellino, D., Guescini, M., Dam, M., et al. (2010). The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 90, 82–100. doi: 10.1016/j.pneurobio.2009.10.012
Fuxe, K., Hedlund, P., von Euler, G., Lundgren, K., Martire, M. S., Ögren, O., et al. (1991). “Galanin/5-HT interactions in the rat central nervous system. relevance for depression,” in Galanin: A New Multifunctional Peptide in the Neuro-endocrine System, eds T. Hökfelt, T. Bartfai, D. Jacobowitz, and D. Ottoson (London: Macmillan Education), 221–235. doi: 10.1007/978-1-349-12664-4_16
Fuxe, K., Jansson, A., Diaz-Cabiale, Z., Andersson, A., Tinner, B., Finnman, U. B., et al. (1998). Galanin modulates 5-Hydroxytryptamine functions. Focus on Galanin and Galanin Fragment/5-hydroxytryptamine1A Receptor Interactions in the Brain. New York, NY: Academy of Sciences.
Gabriel, S. M., Bierer, L. M., Davidson, M., Purohit, D. P., Perl, D. P., Harotunian, V., et al. (1994). Galanin-like immunoreactivity is increased in the postmortem cerebral cortex from patients with Alzheimer’s disease. J. Neurochem. 62, 1516–1523. doi: 10.1046/j.1471-4159.1994.62041516.x
Gainer, H., Yamashita, M., Fields, R. L., House, S. B., and Rusnak, M. (2002). The magnocellular neuronal phenotype: cell-specific gene expression in the hypothalamo-neurohypophysial system. Prog. Brain Res. 139, 1–14. doi: 10.1016/S0079-6123(02)39003-4
Gardier, A. M., Malagie, I., Trillat, A. C., Jacquot, C., and Artigas, F. (1996). Role of 5-HT1A autoreceptors in the mechanism of action of serotoninergic antidepressant drugs: recent findings from in vivo microdialysis studies. Fundam. Clin. Pharmacol. 10, 16–27. doi: 10.1111/j.1472-8206.1996.tb00145.x
Gaus, S. E., Strecker, R. E., Tate, B. A., Parker, R. A., and Saper, C. B. (2002). Ventrolateral preoptic nucleus contains sleep-active, galaninergic neurons in multiple mammalian species. Neuroscience 115, 285–294. doi: 10.1016/S0306-4522(02)00308-1
Gee, C. E., Chen, C. L., Roberts, J. L., Thompson, R., and Watson, S. J. (1983). Identification of proopiomelanocortin neurones in rat hypothalamus by in situ cDNA-mRNA hybridization. Nature 306, 374–376. doi: 10.1038/306374a0
Geffen, L. B., Livett, B. G., and Rush, R. A. (1969). Immunohistochemical localization of protein components of catecholamine storage vesicles. J. Physiol. 204, 593–605. doi: 10.1113/jphysiol.1969.sp008934
Genders, S. G., Scheller, K. J., and Djouma, E. (2018a). Neuropeptide modulation of addiction: Focus on galanin. Neurosci. Biobehav. Rev. doi: 10.1016/j.neubiorev.2018.06.021 [Epub ahead of print].
Genders, S. G., Scheller, K. J., Jaehne, E. J., Turner, B. J., Lawrence, A. J., Brunner, S. M., et al. (2018b). GAL3 receptor knockout mice exhibit an alcohol-preferring phenotype. Addict. Biol. doi: 10.1111/adb.12641 [Epub ahead of print].
Gentleman, S. M., Falkai, P., Bogerts, B., Herrero, M. T., Polak, J. M., Roberts, G. W., et al. (1989). Distribution of galanin-like immunoreactivity in the human brain. Brain Res. 505, 311–315. doi: 10.1016/0006-8993(89)91458-3
German, D. C., Walker, B. S., Manaye, K., Smith, W. K., Woodward, D. J., and North, A. J. (1988). The human locus coeruleus: computer reconstruction of cellular distribution. J. Neurosci. 8, 1776–1788. doi: 10.1523/JNEUROSCI.08-05-01776.1988
Goldman, G., and Coleman, P. D. (1981). Neuron numbers in locus coeruleus do not change with age in Fisher 344 rat. Neurobiol. Aging 2, 33–36. doi: 10.1016/0197-4580(81)90056-7
Goldsmith, P. C., and Ganong, W. F. (1975). Ultrastructural localization of luteinizing hormone-releasing hormone in the median eminence of the rat. Brain Res. 97, 181–193. doi: 10.1016/0006-8993(75)90444-8
Gonda, X., Hullam, G., Antal, P., Eszlari, N., Petschner, P., Riva, M. A., et al. (2018). Significance of risk polymorphisms for depression depends on stress exposure. Sci. Rep. 8:3946. doi: 10.1038/s41598-018-22221-z
Gottsch, M. L., Zeng, H., Hohmann, J. G., Weinshenker, D., Clifton, D. K., and Steiner, R. A. (2005). Phenotypic analysis of mice deficient in the type 2 galanin receptor (GALR2). Mol. Cell. Biol. 25, 4804–4811. doi: 10.1128/MCB.25.11.4804-4811.2005
Grabinski, T. M., Kneynsberg, A., Manfredsson, F. P., and Kanaan, N. M. (2015). A method for combining Inscape in situ hybridization with immunohistochemistry in thick free-floating brain sections and primary neuronal cultures. PLoS One 10:e0120120. doi: 10.1371/journal.pone.0120120
Grenhoff, J., Nisell, M., Ferre, S., Aston-Jones, G., and Svensson, T. H. (1993). Noradrenergic modulation of midbrain dopamine cell firing elicited by stimulation of the locus coeruleus in the rat. J. Neural Transm. Gen. Sect. 93, 11–25. doi: 10.1007/BF01244934
Griebel, G., and Holmes, A. (2013). 50 years of hurdles and hope in anxiolytic drug discovery. Nat. Rev. Drug Discov. 12, 667–687. doi: 10.1038/nrd4075
Griebel, G., and Holsboer, F. (2012). Neuropeptide receptor ligands as drugs for psychiatric diseases: the end of the beginning? Nat. Rev. Drug Discov. 11, 462–478. doi: 10.1038/nrd3702
Guillemin, R. (1978). Control of adenohypophysial functions by peptides of the central nervous system. Harvey Lect. 71, 71–131.
Guillemin, R. (2008). Somatostatin: the beginnings, 1972. Mol. Cell. Endocrinol. 286, 3–4. doi: 10.1016/j.mce.2008.02.025
Guiterrez, R. (ed.) (2009). Co-existence and Co-release of Classic Neurotransmitters. New York, NY: Springer. doi: 10.1007/978-0-387-09622-3
Gundlach, A. L., Rutherfurd, S. D., and Louis, W. J. (1990a). Increase in galanin and neuropeptide Y mRNA in locus coeruleus following acute reserpine treatment. Eur. J. Pharmacol. 184, 163–167.
Gundlach, A. L., Wisden, W., Morris, B. J., and Hunt, S. P. (1990b). Localization of preprogalanin mRNA in rat brain: in situ hybridization study with a synthetic oligonucleotide probe. Neurosci. Lett. 114, 241–247.
Guyenet, P. G., Stornetta, R. L., Bochorishvili, G., Depuy, S. D., Burke, P. G., and Abbott, S. B. (2013). C1 neurons: the body’s EMTs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 305, R187–R204. doi: 10.1152/ajpregu.00054.2013
Habert-Ortoli, E., Amiranoff, B., Loquet, I., Laburthe, M., and Mayaux, J. F. (1994). Molecular cloning of a functional human galanin receptor. Proc. Natl. Acad. Sci. U.S.A. 91, 9780–9783. doi: 10.1073/pnas.91.21.9780
Hahn, M. K., and Blakely, R. D. (2007). The functional impact of SLC6 transporter genetic variation. Annu. Rev. Pharmacol. Toxicol. 47, 401–441. doi: 10.1146/annurev.pharmtox.47.120505.105242
Hamberger, B., and Norberg, K. A. (1963). Monoamines in sympathetic ganglia studied with fluorescence microscopy. Experientia 19, 580–581. doi: 10.1007/BF02150999
Hamm, J. P., and Yuste, R. (2016). Somatostatin interneurons control a key component of mismatch negativity in mouse visual cortex. Cell Rep. 16, 597–604. doi: 10.1016/j.celrep.2016.06.037
Han, M. H., and Nestler, E. J. (2017). Neural substrates of depression and resilience. Neurotherapeutics 14, 677–686. doi: 10.1007/s13311-017-0527-x
Hanley, M. R., Cottrell, G. A., Emson, P. C., and Fonnum, F. (1974). Enzymatic synthesis of acetylcholine by a serotonin-containing neurone from Helix. Nature 251, 631–633. doi: 10.1038/251631a0
Harro, J., and Oreland, L. (2001). Depression as a spreading adjustment disorder of monoaminergic neurons: a case for primary implication of the locus coeruleus. Brain Res. Brain Res. Rev. 38, 79–128. doi: 10.1016/S0165-0173(01)00082-0
Haselton, J. R., and Guyenet, P. G. (1990). Ascending collaterals of medullary barosensitive neurons and C1 cells in rats. Am. J. Physiol. 258, R1051–R1063. doi: 10.1152/ajpregu.1990.258.4.R1051
Hauser, A. S., Attwood, M. M., Rask-Andersen, M., Schioth, H. B., and Gloriam, D. E. (2017). Trends in GPCR drug discovery: new agents, targets and indications. Nat. Rev. Drug Discov. 16, 829–842. doi: 10.1038/nrd.2017.178
Hawes, J. J., Brunzell, D. H., Wynick, D., Zachariou, V., and Picciotto, M. R. (2005). GalR1, but not GalR2 or GalR3, levels are regulated by galanin signaling in the locus coeruleus through a cyclic AMP-dependent mechanism. J. Neurochem. 93, 1168–1176. doi: 10.1111/j.1471-4159.2005.03105.x
Hayley, S., Du, L., Litteljohn, D., Palkovits, M., Faludi, G., Merali, Z., et al. (2015). Gender and brain regions specific differences in brain derived neurotrophic factor protein levels of depressed individuals who died through suicide. Neurosci. Lett. 600, 12–16. doi: 10.1016/j.neulet.2015.05.052
Hedlund, P. B., Yanaihara, N., and Fuxe, K. (1992). Evidence for specific N-terminal galanin fragment binding sites in the rat brain. Eur. J. Pharmacol. 224, 203–205. doi: 10.1016/0014-2999(92)90806-F
Heilig, M. (2004). The NPY system in stress, anxiety and depression. Neuropeptides 38, 213–224. doi: 10.1016/j.npep.2004.05.002
Henry, J. L. (1976). Effects of substance P on functionally identified units in cat spinal cord. Brain Res. 114, 439–451. doi: 10.1016/0006-8993(76)90965-3
Herbert, J. (1993). Peptides in the limbic system: neurochemical codes for co-ordinated adaptive responses to behavioural and physiological demand. Prog. Neurobiol. 41, 723–791. doi: 10.1016/0301-0082(93)90033-O
Herbert, M. K., and Holzer, P. (2002). [Why are substance P(NK1)-receptor antagonists ineffective in pain treatment?]. Anaesthesist 51, 308–319. doi: 10.1007/s00101-002-0296-7
Hill, R. (2000). NK1 (substance P) receptor antagonists–why are they not analgesic in humans? Trends Pharmacol. Sci. 21, 244–246.
Hillarp, N. A. (1949). The functional organization of the peripheral autonomic innervation. Acta Physiol. Scand. 17, 120–129. doi: 10.1111/j.1748-1716.1949.tb00558.x
Hnasko, T. S., Chuhma, N., Zhang, H., Goh, G. Y., Sulzer, D., Palmiter, R. D., et al. (2010). Vesicular glutamate transport promotes dopamine storage and glutamate corelease in vivo. Neuron 65, 643–656. doi: 10.1016/j.neuron.2010.02.012
Hnasko, T. S., and Edwards, R. H. (2012). Neurotransmitter corelease: mechanism and physiological role. Annu. Rev. Physiol. 74, 225–243. doi: 10.1146/annurev-physiol-020911-153315
Hobson, S. A., Bacon, A., Elliot-Hunt, C. R., Holmes, F. E., Kerr, N. C., Pope, R., et al. (2010). Galanin acts as a trophic factor to the central and peripheral nervous systems. EXS 102, 25–38. doi: 10.1007/978-3-0346-0228-0_3
Hoffmann, A., Sportelli, V., Ziller, M., and Spengler, D. (2017). Epigenomics of major depressive disorders and schizophrenia: early life decides. Int. J. Mol. Sci. 18:E1711. doi: 10.3390/ijms18081711
Hohmann, J. G., Jureus, A., Teklemichael, D. N., Matsumoto, A. M., Clifton, D. K., Steiner, R. A., et al. (2003). Distribution and regulation of galanin receptor 1 messenger RNA in the forebrain of wild type and galanin-transgenic mice. Neuroscience 117, 105–117. doi: 10.1016/S0306-4522(02)00798-4
Hokfelt, T. (1991). Neuropeptides in perspective: the last ten years. Neuron 7, 867–879. doi: 10.1016/0896-6273(91)90333-U
Hokfelt, T. (ed.) (2010). Galanin, galanin receptor subtypes and depression-like behaviour. EXS 102, 163–181. doi: 10.1007/978-3-0346-0228-0
Hokfelt, T., Bartfai, T., and Bloom, F. (2003). Neuropeptides: opportunities for drug discovery. Lancet Neurol. 2, 463–472. doi: 10.1016/S1474-4422(03)00482-4
Hökfelt, T., Bartfai, T., and Crawley, J. (eds). (1998). Galanin: Basic Research Discoveries and Therapeutic Implications. Ann. N. Y. Acad. Sci. 863, 1–469.
Hökfelt, T., Bartfai, T., Jacobowitz, D., and Ottoson, D. (eds). (1991). Galanin. A New Multifunctional Peptide in The Neuro-Endocrine System. London: Macmillan. doi: 10.1007/978-1-349-12664-4
Hökfelt, T., and Crawley, J. (eds). (2005). Special issue on galanin. Neuropeptides 39, 125–362. doi: 10.1016/j.npep.2005.03.002
Hokfelt, T., Efendic, S., Hellerstrom, C., Johansson, O., Luft, R., and Arimura, A. (1975a). Cellular localization of somatostatin in endocrine-like cells and neurons of the rat with special references to the A1-cells of the pancreatic islets and to the hypothalamus. Acta Endocrinol. Suppl. 200, 5–41. doi: 10.1530/acta.0.080S005
Hokfelt, T., Efendic, S., Johansson, O., Luft, R., and Arimura, A. (1974a). Immunohistochemical localization of somatostatin (growth hormone release-inhibiting factor) in the guinea pig brain. Brain Res. 80, 165–169. doi: 10.1016/0006-8993(74)90737-9
Hokfelt, T., Elfvin, L. G., Elde, R., Schultzberg, M., Goldstein, M., and Luft, R. (1977a). Occurrence of somatostatin-like immunoreactivity in some peripheral sympathetic noradrenergic neurons. Proc. Natl. Acad. Sci. U.S.A. 74, 3587–3591. doi: 10.1073/pnas.74.8.3587
Hokfelt, T., Fuxe, K., Goldstein, M., and Johansson, O. (1974b). Immunohistochemical evidence for the existence of adrenaline neurons in the rat brain. Brain Res. 66, 235–251. doi: 10.1016/0006-8993(74)90143-7
Hokfelt, T., Fuxe, K., and Pernow, P. (eds). (1986). Coexistence of neuronal messengers: a new principle in chemical transmission. Prog. Brain Res. 68, 1–411.
Hokfelt, T., Johansson, O., and Goldstein, M. (1984). “Central catcholamine neurons as revelaed by immunohistochemistry with special reference to adrenaline neurons,” in Handbook of Chemical Neuroanatomy. Vol. 2: Classical Transmitters in the CNS, eds A. Björklund and T. Hokfelt (Amsterdam: Elsevier), 157–276.
Hokfelt, T., Johansson, O., Ljungdahl, A., Lundberg, J. M., and Schultzberg, M. (1980a). Peptidergic neurones. Nature 284, 515–521. doi: 10.1038/284515a0
Hokfelt, T., and Jonsson, G. (1968). Studies on reaction and binding of monoamines after fixation and processing for electron microscopy with special reference to fixation with potassium permanganate. Histochemie 16, 45–67. doi: 10.1007/BF00306211
Hokfelt, T., Kellerth, J. O., Nilsson, G., and Pernow, B. (1975b). Substance P: localization in the central nervous system and in some primary sensory neurons. Science 190, 889–890.
Hokfelt, T., Ljungdahl, A., Steinbusch, H., Verhofstad, A., Nilsson, G., Brodin, E., et al. (1978). Immunohistochemical evidence of substance P-like immunoreactivity in some 5-hydroxytryptamine-containing neurons in the rat central nervous system. Neuroscience 3, 517–538. doi: 10.1016/0306-4522(78)90017-9
Hokfelt, T., Ljungdahl, A., Terenius, L., Elde, R., and Nilsson, G. (1977b). Immunohistochemical analysis of peptide pathways possibly related to pain and analgesia: enkephalin and substance P. Proc. Natl. Acad. Sci. U.S.A. 74, 3081–3085.
Hokfelt, T., Millhorn, D., Seroogy, K., Tsuruo, Y., Ceccatelli, S., Lindh, B., et al. (1987a). Coexistence of peptides with classical neurotransmitters. Experientia 43, 768–780. doi: 10.1007/BF01945354
Hokfelt, T., Skirboll, L., Rehfeld, J. F., Goldstein, M., Markey, K., and Dann, O. (1980b). A subpopulation of mesencephalic dopamine neurons projecting to limbic areas contains a cholecystokinin-like peptide: evidence from immunohistochemistry combined with retrograde tracing. Neuroscience 5, 2093–2124. doi: 10.1016/0306-4522(80)90127-X
Hokfelt, T., and Tatemoto, K. (2010). Galanin: a multitalented neuropeptide. EXS 102, 1–5. doi: 10.1007/978-3-0346-0228-0_1
Hokfelt, T., Wiesenfeld-Hallin, Z., Villar, M., and Melander, T. (1987b). Increase of galanin-like immunoreactivity in rat dorsal root ganglion cells after peripheral axotomy. Neurosci. Lett. 83, 217–220.
Holets, V. R., Hokfelt, T., Rokaeus, A., Terenius, L., and Goldstein, M. (1988). Locus coeruleus neurons in the rat containing neuropeptide Y, tyrosine hydroxylase or galanin and their efferent projections to the spinal cord, cerebral cortex and hypothalamus. Neuroscience 24, 893–906. doi: 10.1016/0306-4522(88)90076-0
Holmberg, K., Kuteeva, E., Brumovsky, P., Kahl, U., Karlstrom, H., Lucas, G. A., et al. (2005). Generation and phenotypic characterization of a galanin overexpressing mouse. Neuroscience 133, 59–77. doi: 10.1016/j.neuroscience.2005.01.062
Holmes, A., Heilig, M., Rupniak, N. M., Steckler, T., and Griebel, G. (2003). Neuropeptide systems as novel therapeutic targets for depression and anxiety disorders. Trends Pharmacol. Sci. 24, 580–588. doi: 10.1016/j.tips.2003.09.011
Holmes, A., Li, Q., Koenig, E. A., Gold, E., Stephenson, D., Yang, R. J., et al. (2005). Phenotypic assessment of galanin overexpressing and galanin receptor R1 knockout mice in the tail suspension test for depression-related behavior. Psychopharmacology 178, 276–285. doi: 10.1007/s00213-004-1997-1
Holmes, A., and Picciotto, M. R. (2006). Galanin: a novel therapeutic target for depression, anxiety disorders and drug addiction? CNS Neurol. Disord. Drug Targets 5, 225–232. doi: 10.2174/187152706776359600
Holmes, A., Yang, R. J., and Crawley, J. N. (2002). Evaluation of an anxiety-related phenotype in galanin overexpressing transgenic mice. J. Mol. Neurosci. 18, 151–165. doi: 10.1385/JMN:18:1-2:151
Holmes, P. V., Blanchard, D. C., Blanchard, R. J., Brady, L. S., and Crawley, J. N. (1995). Chronic social stress increases levels of preprogalanin mRNA in the rat locus coeruleus. Pharmacol. Biochem. Behav. 50, 655–660. doi: 10.1016/0091-3057(94)00334-3
Howard, A. D., Tan, C., Shiao, L. L., Palyha, O. C., McKee, K. K., Weinberg, D. H., et al. (1997). Molecular cloning and characterization of a new receptor for galanin. FEBS Lett. 405, 285–290. doi: 10.1016/S0014-5793(97)00196-8
Howe, P. R., Costa, M., Furness, J. B., and Chalmers, J. P. (1980). Simultaneous demonstration of phenylethanolamine N-methyltransferase immunofluorescent and catecholamine fluorescent nerve cell bodies in the rat medulla oblongata. Neuroscience 5, 2229–2238. doi: 10.1016/0306-4522(80)90139-6
Hoyer, D., and Bartfai, T. (2012). Neuropeptides and neuropeptide receptors: drug targets, and peptide and non-peptide ligands: a tribute to Prof. Dieter Seebach. Chem. Biodivers. 9, 2367–2387. doi: 10.1002/cbdv.201200288
Huang, H. P., Wang, S. R., Yao, W., Zhang, C., Zhou, Y., Chen, X. W., et al. (2007). Long latency of evoked quantal transmitter release from somata of locus coeruleus neurons in rat pontine slices. Proc. Natl. Acad. Sci. U.S.A. 104, 1401–1406. doi: 10.1073/pnas.0608897104
Hughes, J., Smith, T. W., Kosterlitz, H. W., Fothergill, L. A., Morgan, B. A., Morris, H. R., et al. (1975). Identification of two related pentapeptides from the brain with potent opiate agonist activity. Nature 258, 577–580. doi: 10.1038/258577a0
Hyde, C. L., Nagle, M. W., Tian, C., Chen, X., Paciga, S. A., Wendland, J. R., et al. (2016). Identification of 15 genetic loci associated with risk of major depression in individuals of European descent. Nat. Genet 48, 1031–1036. doi: 10.1038/ng.3623
Ihalainen, J. A., Riekkinen, P. Jr., and Feenstra, M. G. (1999). Comparison of dopamine and noradrenaline release in mouse prefrontal cortex, striatum and hippocampus using microdialysis. Neurosci. Lett. 277, 71–74. doi: 10.1016/S0304-3940(99)00840-X
Ihnatko, R., and Theodorsson, E. (2017). Short N-terminal galanin fragments are occurring naturally in vivo. Neuropeptides 63, 1–13. doi: 10.1016/j.npep.2017.03.005
Iismaa, T. P., and Shine, J. (1999). Galanin and galanin receptors. Results Probl. Cell Differ. 26, 257–291. doi: 10.1007/978-3-540-49421-8_12
Isingrini, E., Perret, L., Rainer, Q., Amilhon, B., Guma, E., Tanti, A., et al. (2016). Resilience to chronic stress is mediated by noradrenergic regulation of dopamine neurons. Nat. Neurosci. 19, 560–563. doi: 10.1038/nn.4245
Iwamoto, K., Kakiuchi, C., Bundo, M., Ikeda, K., and Kato, T. (2004). Molecular characterization of bipolar disorder by comparing gene expression profiles of postmortem brains of major mental disorders. Mol. Psychiatry 9, 406–416. doi: 10.1038/sj.mp.4001437
Jacobowitz, D. M., Kresse, A., and Skofitsch, G. (2004). Galanin in the brain: chemoarchitectonics and brain cartography–a historical review. Peptides 25, 433–464. doi: 10.1016/j.peptides.2004.02.015
Jacobowitz, D. M., and Skofitsch, G. (1991). “Localization of galanin cell bodies in the brain by immunocytochemistry and in situ hybridization histochemistry,” in Galanin: A New Multifunctional Peptide in the Neuro-endocrine System, eds T. Hökfelt, T. Bartfai, D. Jacobowitz, and D. Ottoson (Macmillan Education: London), 69–92. doi: 10.1007/978-1-349-12664-4_6
Jacoby, A. S., Webb, G. C., Liu, M. L., Kofler, B., Hort, Y. J., Fathi, Z., et al. (1997). Structural organization of the mouse and human GALR1 galanin receptor genes (Galnr and GALNR) and chromosomal localization of the mouse gene. Genomics 45, 496–508. doi: 10.1006/geno.1997.4960
Jaim-Etcheverry, G. (1994). Co-existence of chemical messengers in neurons. Prog. Brain Res. 100, 73–80. doi: 10.1016/S0079-6123(08)60771-2
Jaim-Etcheverry, G., and Zieher, L. M. (1973). Proceedings: coexistence of monoamines in adrenergic synaptic vesicles. Acta Physiol. Lat. Am. 23, 616–618.
James, D. J., and Martin, T. F. (2013). CAPS and munc13: CATCHRs that SNARE vesicles. Front. Endocrinol. 4:187. doi: 10.3389/fendo.2013.00187
Jan, L. Y., and Jan, Y. N. (1982). Peptidergic transmission in sympathetic ganglia of the frog. J. Physiol. 327, 219–246. doi: 10.1113/jphysiol.1982.sp014228
Jansson, A., Tinner, B., Andbjer, B., Razani, H., Wang, F., Schött, P. A., et al. (2000). Internalization of intracerebrally administered porcine galanin (1-29) by a discrete nerve cell population in the hippocampus of the rat. Exp. Neurol. 161, 153–166. doi: 10.1006/exnr.1999.7266
Jonas, P., Bischofberger, J., and Sandkuhler, J. (1998). Corelease of two fast neurotransmitters at a central synapse. Science 281, 419–424. doi: 10.1126/science.281.5375.419
Jordan, S., Kramer, G. L., Zukas, P. K., and Petty, F. (1994). Previous stress increases in vivo biogenic amine response to swim stress. Neurochem. Res. 19, 1521–1525. doi: 10.1007/BF00969000
Juhasz, G., Gonda, X., Hullam, G., Eszlari, N., Kovacs, D., Lazary, J., et al. (2015). Variability in the effect of 5-HTTLPR on depression in a large European population: the role of age, symptom profile, type and intensity of life stressors. PLoS One 10:e0116316. doi: 10.1371/journal.pone.0116316
Juhasz, G., Hullam, G., Eszlari, N., Gonda, X., Antal, P., Anderson, I. M., et al. (2014). Brain galanin system genes interact with life stresses in depression-related phenotypes. Proc. Natl. Acad. Sci. U.S.A. 111, E1666–E1673. doi: 10.1073/pnas.1403649111
Jungnickel, S. R., and Gundlach, A. L. (2005). [125I]-Galanin binding in brain of wildtype, and galanin- and GalR1-knockout mice: strain and species differences in GalR1 density and distribution. Neuroscience 131, 407–421. doi: 10.1016/j.neuroscience.2004.11.023
Jurek, B., and Neumann, I. D. (2018). The oxytocin receptor: from intracellular signaling to behavior. Physiol. Rev. 98, 1805–1908. doi: 10.1152/physrev.00031.2017
Kang, H. J., Adams, D. H., Simen, A., Simen, B. B., Rajkowska, G., Stockmeier, C. A., et al. (2007). Gene expression profiling in postmortem prefrontal cortex of major depressive disorder. J. Neurosci. 27, 13329–13340. doi: 10.1523/JNEUROSCI.4083-07.2007
Kanner, B. I. (1994). Sodium-coupled neurotransmitter transport: structure, function and regulation. J. Exp. Biol. 196, 237–249.
Karlsson, R. M., and Holmes, A. (2006). Galanin as a modulator of anxiety and depression and a therapeutic target for affective disease. Amino Acids 31, 231–239. doi: 10.1007/s00726-006-0336-8
Kask, A., Harro, J., von Horsten, S., Redrobe, J. P., Dumont, Y., and Quirion, R. (2002). The neurocircuitry and receptor subtypes mediating anxiolytic-like effects of neuropeptide Y. Neurosci. Biobehav. Rev. 26, 259–283. doi: 10.1016/S0149-7634(01)00066-5
Kautz, M., Charney, D. S., and Murrough, J. W. (2017). Neuropeptide Y, resilience, and PTSD therapeutics. Neurosci. Lett. 649, 164–169. doi: 10.1016/j.neulet.2016.11.061
Kawa, L., Arborelius, U. P., Yoshitake, T., Kehr, J., Hokfelt, T., Risling, M., et al. (2015). Neurotransmitter systems in a mild blast traumatic brain injury model: catecholamines and serotonin. J. Neurotrauma 32, 1190–1199. doi: 10.1089/neu.2014.3669
Kawa, L., Barde, S., Arborelius, U. P., Theodorsson, E., Agoston, D., Risling, M., et al. (2016). Expression of galanin and its receptors are perturbed in a rodent model of mild, blast-induced traumatic brain injury. Exp. Neurol. 279, 159–167. doi: 10.1016/j.expneurol.2016.02.019
Keller, M., Montgomery, S., Ball, W., Morrison, M., Snavely, D., Liu, G., et al. (2006). Lack of efficacy of the substance p (neurokinin1 receptor) antagonist aprepitant in the treatment of major depressive disorder. Biol. Psychiatry 59, 216–223. doi: 10.1016/j.biopsych.2005.07.013
Kempadoo, K. A., Mosharov, E. V., Choi, S. J., Sulzer, D., and Kandel, E. R. (2016). Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. U.S.A. 113, 14835–14840. doi: 10.1073/pnas.1616515114
Kendler, K. S. (2012). The dappled nature of causes of psychiatric illness: replacing the organic-functional/hardware-software dichotomy with empirically based pluralism. Mol. Psychiatry 17, 377–388. doi: 10.1038/mp.2011.182
Kendler, K. S. (2013). What psychiatric genetics has taught us about the nature of psychiatric illness and what is left to learn. Mol. Psychiatry 18, 1058–1066. doi: 10.1038/mp.2013.50
Kennedy, M. J., and Ehlers, M. D. (2011). Mechanisms and function of dendritic exocytosis. Neuron 69, 856–875. doi: 10.1016/j.neuron.2011.02.032
Kerkut, G. A., Sedden, C. B., and Walker, R. J. (1967). Uptake of DOPA and 5-hydroxytryptophan by monoamine-forming neurones in the brain of Helix aspersa. Comp. Biochem. Physiol. 23, 159–162. doi: 10.1016/0010-406X(67)90483-5
Kerman, I. A., Bernard, R., Bunney, W. E., Jones, E. G., Schatzberg, A. F., Myers, R. M., et al. (2012). Evidence for transcriptional factor dysregulation in the dorsal raphe nucleus of patients with major depressive disorder. Front. Neurosci. 6:135. doi: 10.3389/fnins.2012.00135
Kessler, R. C., Berglund, P., Demler, O., Jin, R., Koretz, D., Merikangas, K. R., et al. (2003). The epidemiology of major depressive disorder: results from the National Comorbidity Survey Replication (NCS-R). JAMA 289, 3095–3105. doi: 10.1001/jama.289.23.3095
Khoshbouei, H., Cecchi, M., Dove, S., Javors, M., and Morilak, D. A. (2002). Behavioral reactivity to stress: amplification of stress-induced noradrenergic activation elicits a galanin-mediated anxiolytic effect in central amygdala. Pharmacol. Biochem. Behav. 71, 407–417. doi: 10.1016/S0091-3057(01)00683-9
Kim, D. K., Yun, S., Son, G. H., Hwang, J. I, Park, C. R., Kim, J. I., et al. (2014). Coevolution of the spexin/galanin/kisspeptin family: spexin activates galanin receptor type II and III. Endocrinology 155, 1864–1873. doi: 10.1210/en.2013-2106
Kim, J. I, Ganesan, S., Luo, S. X., Wu, Y. W., Park, E., Huang, E. J., et al. (2015). Aldehyde dehydrogenase 1a1 mediates a GABA synthesis pathway in midbrain dopaminergic neurons. Science 350, 102–106. doi: 10.1126/science.aac4690
Kinney, J. W., Sanchez-Alavez, M., Barr, A. M., Criado, J. R., Crawley, J. N., Behrens, M. M., et al. (2009). Impairment of memory consolidation by galanin correlates with in vivo inhibition of both LTP and CREB phosphorylation. Neurobiol. Learn. Mem. 92, 429–438. doi: 10.1016/j.nlm.2009.06.005
Klempan, T. A., Sequeira, A., Canetti, L., Lalovic, A., Ernst, C., ffrench-Mullen, J., et al. (2009). Altered expression of genes involved in ATP biosynthesis and GABAergic neurotransmission in the ventral prefrontal cortex of suicides with and without major depression. Mol. Psychiatry 14, 175–189. doi: 10.1038/sj.mp.4002110
Kohler, C., and Chan-Palay, V. (1990). Galanin receptors in the post-mortem human brain. Regional distribution of 125I-galanin binding sites using the method of in vitro receptor autoradiography. Neurosci. Lett. 120, 179–182. doi: 10.1016/0304-3940(90)90032-5
Kohler, C., Persson, A., Melander, T., Theodorsson, E., Sedvall, G., Hökfelt, T., et al. (1989). Distribution of galanin-binding sites in the monkey and human telencephalon: preliminary observations. Exp. Brain Res. 75, 375–380. doi: 10.1007/BF00247944
Kohler, G., and Milstein, C. (1975). Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 256, 495–497. doi: 10.1038/256495a0
Kolakowski, L. F. Jr., O’Neill, G. P., Howard, A. D., Broussard, S. R., Sullivan, K. A., Feighner, S. D., et al. (1998). Molecular characterization and expression of cloned human galanin receptors GALR2 and GALR3. J. Neurochem. 71, 2239–2251. doi: 10.1046/j.1471-4159.1998.71062239.x
Koller, A., Rid, R., Beyreis, M., Bianchini, R., Holub, B. S., Lang, A., et al. (2016). In vitro toxicity of the galanin receptor 3 antagonist SNAP 37889. Neuropeptides 56, 83–88. doi: 10.1016/j.npep.2015.12.003
Komori, T., Nomura, J., Inoue, K., and Kitayama, I. (1990). Tyrosine hydroxylase activity in discrete brain regions of depression model rats. Jpn. J. Psychiatry Neurol. 44, 747–754. doi: 10.1111/j.1440-1819.1990.tb01654.x
Konkel, M. J., Lagu, B., Boteju, L. W., Jimenez, H., Noble, S., Walker, M. W., et al. (2006a). 3-arylimino-2-indolones are potent and selective galanin GAL3 receptor antagonists. J. Med. Chem. 49, 3757–3758. doi: 10.1021/jm060001n
Konkel, M. J., Packiarajan, M., Chen, H., Topiwala, U. P., Jimenez, H., Talisman, I. J., et al. (2006b). Amino substituted analogs of 1-phenyl-3-phenylimino-2-indolones with potent galanin Gal3 receptor binding affinity and improved solubility. Bioorg. Med. Chem. Lett. 16, 3950–3954. doi: 10.1016/j.bmcl.2006.05.025
Kordower, J. H., Le, H. K., and Mufson, E. J. (1992). Galanin immunoreactivity in the primate central nervous system. J. Comp. Neurol. 319, 479–500. doi: 10.1002/cne.903190403
Kordower, J. H., and Mufson, E. J. (1990). Galanin-like immunoreactivity within the primate basal forebrain: differential staining patterns between humans and monkeys. J. Comp. Neurol. 294, 281–292. doi: 10.1002/cne.902940211
Korf, J., Aghajanian, G. K., and Roth, R. H. (1973). Increased turnover of norepinephrine in the rat cerebral cortex during stress: role of the locus coeruleus. Neuropharmacology 12, 933–938. doi: 10.1016/0028-3908(73)90024-5
Kovacs, G. L., Szabo, G., Penke, B., and Telegdy, G. (1981). Effects of cholecystokinin octapeptide on striatal dopamine metabolism and on apomorphine-induced stereotyped cage-climbing in mice. Eur. J. Pharmacol. 69, 313–319. doi: 10.1016/0014-2999(81)90477-5
Kowall, N. W., and Beal, M. F. (1989). Galanin-like immunoreactivity is present in human substantia innominata and in senile plaques in Alzheimer’s disease. Neurosci. Lett. 98, 118–123. doi: 10.1016/0304-3940(89)90384-4
Kozicz, T., Tilburg-Ouwens, D., Faludi, G., Palkovits, M., and Roubos, E. (2008). Gender-related urocortin 1 and brain-derived neurotrophic factor expression in the adult human midbrain of suicide victims with major depression. Neuroscience 152, 1015–1023. doi: 10.1016/j.neuroscience.2007.12.050
Kozlovsky, N., Matar, M. A., Kaplan, Z., Zohar, J., and Cohen, H. (2009). The role of the galaninergic system in modulating stress-related responses in an animal model of posttraumatic stress disorder. Biol. Psychiatry 65, 383–391. doi: 10.1016/j.biopsych.2008.10.034
Kramer, M. S., Cutler, N., Feighner, J., Shrivastava, R., Carman, J., Sramek, J. J., et al. (1998). Distinct mechanism for antidepressant activity by blockade of central substance P receptors. Science 281, 1640–1645. doi: 10.1126/science.281.5383.1640
Krisch, B. (1976). Immunohistochemical and electron microscopic study of the rat hypothalamic nuclei and cell clusters under various experimental conditions. Possible sites of hormone release. Cell Tissue Res. 174, 109–127. doi: 10.1007/BF00222154
Krishnan, V., Han, M. H., Graham, D. L., Berton, O., Renthal, W., Russo, S. J., et al. (2007). Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell 131, 391–404. doi: 10.1016/j.cell.2007.09.018
Krystal, J. H., and Neumeister, A. (2009). Noradrenergic and serotonergic mechanisms in the neurobiology of posttraumatic stress disorder and resilience. Brain Res. 1293, 13–23. doi: 10.1016/j.brainres.2009.03.044
Kupfermann, I. (1991). Functional studies of cotransmission. Physiol. Rev. 71, 683–732. doi: 10.1152/physrev.1991.71.3.683
Kuteeva, E., Calza, L., Holmberg, K., Theodorsson, E., Ogren, S. O., Hökfelt, T., et al. (2004). Distribution of galanin and galanin transcript in the brain of a galanin-overexpressing transgenic mouse. J. Chem. Neuroanat. 28, 185–216. doi: 10.1016/j.jchemneu.2004.06.004
Kuteeva, E., Hokfelt, T., and Ogren, S. O. (2005). Behavioural characterisation of young adult transgenic mice overexpressing galanin under the PDGF-B promoter. Regul. Pept. 125, 67–78. doi: 10.1016/j.regpep.2004.07.028
Kuteeva, E., Hokfelt, T., Wardi, T., and Ogren, S. O. (2010). Galanin, galanin receptor subtypes and depression-like behaviour. EXS 102, 163–181. doi: 10.1007/978-3-0346-0228-0_12
Kuteeva, E., Wardi, T., Lundstrom, L., Sollenberg, U., Langel, U., Hökfelt, T., et al. (2008). Differential role of galanin receptors in the regulation of depression-like behavior and monoamine/stress-related genes at the cell body level. Neuropsychopharmacology 33, 2573–2585. doi: 10.1038/sj.npp.1301660
Labonte, B., Engmann, O., Purushothaman, I., Menard, C., Wang, J., Tan, C., et al. (2017). Sex-specific transcriptional signatures in human depression. Nat. Med. 23, 1102–1111. doi: 10.1038/nm.4386
Labonte, B., Suderman, M., Maussion, G., Lopez, J. P., Navarro-Sanchez, L., Yerko, V., et al. (2013). Genome-wide methylation changes in the brains of suicide completers. Am. J. Psychiatry 170, 511–520. doi: 10.1176/appi.ajp.2012.12050627
Landgraf, R., and Neumann, I. D. (2004). Vasopressin and oxytocin release within the brain: a dynamic concept of multiple and variable modes of neuropeptide communication. Front. Neuroendocrinol. 25, 150–176. doi: 10.1016/j.yfrne.2004.05.001
Landis, S. C., and Keefe, D. (1983). Evidence for neurotransmitter plasticity in vivo: developmental changes in properties of cholinergic sympathetic neurons. Dev. Biol. 98, 349–372. doi: 10.1016/0012-1606(83)90365-2
Landry, M., Aman, K., and Hokfelt, T. (1998). Galanin-R1 receptor in anterior and mid-hypothalamus: distribution and regulation. J. Comp. Neurol. 399, 321–340. doi: 10.1002/(SICI)1096-9861(19980928)399:3<321::AID-CNE3>3.0.CO;2-2
Lang, R., Gundlach, A. L., Holmes, F. E., Hobson, S. A., Wynick, D., Hökfelt, T., et al. (2015). Physiology, signaling, and pharmacology of galanin peptides and receptors: three decades of emerging diversity. Pharmacol. Rev. 67, 118–175. doi: 10.1124/pr.112.006536
Lang, R., Gundlach, A. L., and Kofler, B. (2007). The galanin peptide family: receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol. Ther. 115, 177–207. doi: 10.1016/j.pharmthera.2007.05.009
Larm, J. A., Shen, P. J., and Gundlach, A. L. (2003). Differential galanin receptor-1 and galanin expression by 5-HT neurons in dorsal raphe nucleus of rat and mouse: evidence for species-dependent modulation of serotonin transmission. Eur. J. Neurosci. 17, 481–493. doi: 10.1046/j.1460-9568.2003.02471.x
Le Maitre, E., Barde, S. S., Palkovits, M., Diaz-Heijtz, R., and Hokfelt, T. G. (2013). Distinct features of neurotransmitter systems in the human brain with focus on the galanin system in locus coeruleus and dorsal raphe. Proc. Natl. Acad. Sci. U.S.A. 110, E536–E545. doi: 10.1073/pnas.1221378110
Le Maitre, W., Xia, S., Le Maitre, E., Dun, X. P., Lu, J., Theodorsson, E., et al. (2011). Galanin receptor 2 overexpressing mice display an antidepressive-like phenotype: possible involvement of the subiculum. Neuroscience 190, 270–288. doi: 10.1016/j.neuroscience.2011.05.015
Lein, E. S., Hawrylycz, M. J., Ao, N., Ayres, M., Bensinger, A., Bernard, A., et al. (2007). Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176. doi: 10.1038/nature05453
Lembeck, F. (1957). Untersuchungen über die Auslösung afferenter Impulse. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmak. 230, 1–9. doi: 10.1007/BF00246068
Lener, M. S., Kadriu, B., and Zarate, C. A. Jr. (2017). Ketamine and beyond: investigations into the potential of glutamatergic agents to treat depression. Drugs 77, 381–401. doi: 10.1007/s40265-017-0702-8
Lesch, K. P., Bengel, D., Heils, A., Sabol, S. Z., Greenberg, B. D., Petri, S., et al. (1996). Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274, 1527–1531. doi: 10.1126/science.274.5292.1527
Lewis, D. A., Campbell, M. J., Foote, S. L., and Morrison, J. H. (1986). The monoaminergic innervation of primate neocortex. Hum. Neurobiol. 5, 181–188.
Li, J. Z., Bunney, B. G., Meng, F., Hagenauer, M. H., Walsh, D. M., Vawter, M. P., et al. (2013). Circadian patterns of gene expression in the human brain and disruption in major depressive disorder. Proc. Natl. Acad. Sci. U.S.A. 110, 9950–9955. doi: 10.1073/pnas.1305814110
Li, P., Janczewski, W. A., Yackle, K., Kam, K., Pagliardini, S., Krasnow, M. A., et al. (2016). The peptidergic control circuit for sighing. Nature 530, 293–297. doi: 10.1038/nature16964
Li, Y. W., Bayliss, D. A., and Guyenet, P. G. (1995). C1 neurons of neonatal rats: intrinsic beating properties and alpha 2-adrenergic receptors. Am. J. Physiol. 269, R1356–R1369. doi: 10.1152/ajpregu.1995.269.6.R1356
Liu, H. X., Brumovsky, P., Schmidt, R., Brown, W., Payza, K., Hodzic, L., et al. (2001). Receptor subtype-specific pronociceptive and analgesic actions of galanin in the spinal cord: selective actions via GalR1 and GalR2 receptors. Proc. Natl. Acad. Sci. U.S.A. 98, 9960–9964. doi: 10.1073/pnas.161293598
Liu, Y., and Edwards, R. H. (1997). The role of vesicular transport proteins in synaptic transmission and neural degeneration. Annu. Rev. Neurosci. 20, 125–156. doi: 10.1146/annurev.neuro.20.1.125
Liu, Z., Xu, Y., Wu, L., and Zhang, S. (2010). Evolution of galanin receptor genes: insights from the deuterostome genomes. J. Biomol. Struct. Dyn. 28, 97–106. doi: 10.1080/07391102.2010.10507346
Ljungdahl, A., Hokfelt, T., and Nilsson, G. (1978). Distribution of substance P-like immunoreactivity in the central nervous system of the rat–I. Cell bodies and nerve terminals. Neuroscience 3, 861–943. doi: 10.1016/0306-4522(78)90116-1
Lolak, S., Suwannarat, P., and Lipsky, R. H. (2014). Epigenetics of depression. Prog. Mol. Biol. Transl. Sci. 128, 103–137. doi: 10.1016/B978-0-12-800977-2.00005-X
Lopez, J. P., Fiori, L. M., Gross, J. A., Labonte, B., Yerko, V., Mechawar, N., et al. (2014a). Regulatory role of miRNAs in polyamine gene expression in the prefrontal cortex of depressed suicide completers. Int. J. Neuropsychopharmacol. 17, 23–32. doi: 10.1017/S1461145713000941
Lopez, J. P., Lim, R., Cruceanu, C., Crapper, L., Fasano, C., Labonte, B., et al. (2014b). miR-1202 is a primate-specific and brain-enriched microRNA involved in major depression and antidepressant treatment. Nat. Med. 20, 764–768. doi: 10.1038/nm.3582
Lopizzo, N., Bocchio Chiavetto, L., Cattane, N., Plazzotta, G., Tarazi, I., Pariante, C. M., et al. (2015). Gene-environment interaction in major depression: focus on experience-dependent biological systems. Front. Psychiatry 6:68. doi: 10.3389/fpsyt.2015.00068
Lorimer, D. D., Matkowskj, K., and Benya, R. V. (1997). Cloning, chromosomal location, and transcriptional regulation of the human galanin-1 receptor gene (GALN1R). Biochem. Biophys. Res. Commun. 241, 558–564. doi: 10.1006/bbrc.1997.7838
Lu, X., Barr, A. M., Kinney, J. W., Sanna, P., Conti, B., Behrens, M. M., et al. (2005a). A role for galanin in antidepressant actions with a focus on the dorsal raphe nucleus. Proc. Natl. Acad. Sci. U.S.A. 102, 874–879. doi: 10.1073/pnas.0408891102
Lu, X., Lundstrom, L., and Bartfai, T. (2005b). Galanin (2-11) binds to GalR3 in transfected cell lines: limitations for pharmacological definition of receptor subtypes. Neuropeptides 39, 165–167.
Lu, X., Roberts, E., Xia, F., Sanchez-Alavez, M., Liu, T., Baldwin, R., et al. (2010). GalR2-positive allosteric modulator exhibits anticonvulsant effects in animal models. Proc. Natl. Acad. Sci. U.S.A. 107, 15229–15234. doi: 10.1073/pnas.1008986107
Lu, X., Ross, B., Sanchez-Alavez, M., Zorrilla, E. P., and Bartfai, T. (2008). Phenotypic analysis of GalR2 knockout mice in anxiety- and depression-related behavioral tests. Neuropeptides 42, 387–397. doi: 10.1016/j.npep.2008.04.009
Lu, X., Sharkey, L., and Bartfai, T. (2007). The brain galanin receptors: targets for novel antidepressant drugs. CNS Neurol. Disord. Drug Targets 6, 183–192. doi: 10.2174/187152707780619335
Lu, X. Y., and Bartfai, T. (2009). Analyzing the validity of GalR1 and GalR2 antibodies using knockout mice. N. S. Arch. Pharmacol. 379, 417–420. doi: 10.1007/s00210-009-0394-z
Ludwig, M., Apps, D., Menzies, J., Patel, J. C., and Rice, M. E. (2016). Dendritic release of neurotransmitters. Compr. Physiol. 7, 235–252. doi: 10.1002/cphy.c160007
Ludwig, M., and Leng, G. (2006). Dendritic peptide release and peptide-dependent behaviours. Nat. Rev. Neurosci. 7, 126–136. doi: 10.1038/nrn1845
Lundberg, J. M. (1996). Pharmacology of cotransmission in the autonomic nervous system: integrative aspects on amines, neuropeptides, adenosine triphosphate, amino acids and nitric oxide. Pharmacol. Rev. 48, 113–178.
Lundberg, J. M., Anggard, A., Fahrenkrug, J., Hokfelt, T., and Mutt, V. (1980). Vasoactive intestinal polypeptide in cholinergic neurons of exocrine glands: functional significance of coexisting transmitters for vasodilation and secretion. Proc. Natl. Acad. Sci. U.S.A. 77, 1651–1655. doi: 10.1073/pnas.77.3.1651
Lundberg, J. M., Fried, G., Fahrenkrug, J., Holmstedt, B., Hokfelt, T., Lagercrantz, H., et al. (1981). Subcellular fractionation of cat submandibular gland: comparative studies on the distribution of acetylcholine and vasoactive intestinal polypeptide (VIP). Neuroscience 6, 1001–1010. doi: 10.1016/0306-4522(81)90066-X
Lundberg, J. M., and Hokfelt, T. (1983). Coexistence of peptides and classical neurotransmitters. TINS 6, 325–333. doi: 10.1016/0166-2236(83)90149-2
Lundberg, J. M., Hokfelt, T., Schultzberg, M., Uvnas-Wallensten, K., Kohler, C., Said, S. I., et al. (1979). Occurrence of vasoactive intestinal polypeptide (VIP)-like immunoreactivity in certain cholinergic neurons of the cat: evidence from combined immunohistochemistry and acetylcholinesterase staining. Neuroscience 4, 1539–1559. doi: 10.1016/0306-4522(79)90018-6
Lundberg, J. M., Terenius, L., Hokfelt, T., Martling, C. R., Tatemoto, K., Mutt, V., et al. (1982). Neuropeptide Y (NPY)-like immunoreactivity in peripheral noradrenergic neurons and effects of NPY on sympathetic function. Acta Physiol. Scand. 116, 477–480. doi: 10.1111/j.1748-1716.1982.tb07171.x
Lundstrom, L., Elmquist, A., Bartfai, T., and Langel, U. (2005). Galanin and its receptors in neurological disorders. Neuromol. Med. 7, 157–180. doi: 10.1385/NMM:7:1-2:157
Luttrell, L. M., Maudsley, S., and Bohn, L. M. (2015). Fulfilling the promise of ”Biased” G protein-coupled receptor agonism. Mol. Pharmacol. 88, 579–588. doi: 10.1124/mol.115.099630
Lutz, P. E., Tanti, A., Gasecka, A., Barnett-Burns, S., Kim, J. J., Zhou, Y., et al. (2017). Association of a history of child abuse with impaired myelination in the anterior cingulate cortex: convergent epigenetic, transcriptional, and morphological evidence. Am. J. Psychiatry 174, 1185–1194. doi: 10.1176/appi.ajp.2017.16111286
Ma, X., Tong, Y. G., Schmidt, R., Brown, W., Payza, K., Hodzic, L., et al. (2001). Effects of galanin receptor agonists on locus coeruleus neurons. Brain Res. 919, 169–174. doi: 10.1016/S0006-8993(01)03033-5
Machado-Vieira, R., Ibrahim, L., and Zarate, C. A. Jr. (2011). Histone deacetylases and mood disorders: epigenetic programming in gene-environment interactions. CNS Neurosci. Ther. 17, 699–704. doi: 10.1111/j.1755-5949.2010.00203.x
Maeda, T. (2000). The locus coeruleus: history. J. Chem. Neuroanat. 18, 57–64. doi: 10.1016/S0891-0618(99)00051-4
Maes, M., and Meltzer, H. Y. (1995). “The serotonin hypothesis of major depression,” in Psychopharmacology: The Fourth Generation of Progress, eds F. Bloom and D. Kupfer (New York, NY: Raven Press Ltd), 933–944.
Maheu, M., Lopez, J. P., Crapper, L., Davoli, M. A., Turecki, G., Mechawar, N., et al. (2015). MicroRNA regulation of central glial cell line-derived neurotrophic factor (GDNF) signalling in depression. Transl. Psychiatry 5:e511. doi: 10.1038/tp.2015.11
Mains, R. E., Cullen, E. I., May, V., and Eipper, B. A. (1987). The role of secretory granules in peptide biosynthesis. Ann. N. Y. Acad. Sci. 493, 278–291. doi: 10.1111/j.1749-6632.1987.tb27213.x
Major Depressive Disorder Working Group of the Psychiatric Gwas Consortium, Ripke, S., Wray, N. R., Lewis, C. M., Hamilton, S. P., Weissman, M. M., et al. (2013). A mega-analysis of genome-wide association studies for major depressive disorder. Mol. Psychiatry 18, 497–511. doi: 10.1038/mp.2012.21
Maldonado, R., and Koob, G. F. (1993). Destruction of the locus coeruleus decreases physical signs of opiate withdrawal. Brain Res. 605, 128–138. doi: 10.1016/0006-8993(93)91364-X
Mantyh, P. W., DeMaster, E., Malhotra, A., Ghilardi, J. R., Rogers, S. D., Mantyh, C. R., et al. (1995). Receptor endocytosis and dendrite reshaping in spinal neurons after somatosensory stimulation. Science 268, 1629–1632. doi: 10.1126/science.7539937
Maubach, K. A., Rupniak, N. M., Kramer, M. S., and Hill, R. G. (1999). Novel strategies for pharmacotherapy of depression. Curr. Opin. Chem. Biol. 3, 481–488. doi: 10.1016/S1367-5931(99)80070-2
McDevitt, R. A., Szot, P., Baratta, M. V., Bland, S. T., White, S. S., Maier, S. F., et al. (2009). Stress-induced activity in the locus coeruleus is not sensitive to stressor controllability. Brain Res. 1285, 109–118. doi: 10.1016/j.brainres.2009.06.017
McEwen, B. S. (2003). Mood disorders and allostatic load. Biol. Psychiatry 54, 200–207. doi: 10.1016/S0006-3223(03)00177-X
McEwen, B. S. (2007). Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol. Rev. 87, 873–904. doi: 10.1152/physrev.00041.2006
McEwen, B. S., Gray, J. D., and Nasca, C. (2015). 60 years of neuroendocrinology: redefining neuroendocrinology: stress, sex and cognitive and emotional regulation. J. Endocrinol. 226, T67–T83. doi: 10.1530/JOE-15-0121
Meaney, M. J., and Ferguson-Smith, A. C. (2010). Epigenetic regulation of the neural transcriptome: the meaning of the marks. Nat. Neurosci. 13, 1313–1318. doi: 10.1038/nn1110-1313
Meister, B. (1993). Gene expression and chemical diversity in hypothalamic neurosecretory neurons. Mol. Neurobiol. 7, 87–110. doi: 10.1007/BF02935638
Melander, T., Hokfelt, T., Nilsson, S., and Brodin, E. (1986a). Visualization of galanin binding sites in the rat central nervous system. Eur. J. Pharmacol. 124, 381–382. doi: 10.1016/0014-2999(86)90247-5
Melander, T., Hokfelt, T., and Rokaeus, A. (1986b). Distribution of galaninlike immunoreactivity in the rat central nervous system. J. Comp. Neurol. 248, 475–517.
Melander, T., Hokfelt, T., Rokaeus, A., Cuello, A. C., Oertel, W. H., Verhofstad, A., et al. (1986c). Coexistence of galanin-like immunoreactivity with catecholamines, 5-hydroxytryptamine, GABA and neuropeptides in the rat CNS. J. Neurosci. 6, 3640–3654. doi: 10.1523/JNEUROSCI.06-12-03640.1986
Melander, T., Kohler, C., Nilsson, S., Hokfelt, T., Brodin, E., Theodorsson, E., et al. (1988). Autoradiographic quantitation and anatomical mapping of 125I-galanin binding sites in the rat central nervous system. J. Chem. Neuroanat. 1, 213–233.
Melander, T., and Staines, W. A. (1986). A galanin-like peptide coexists in putative cholinergic somata of the septum-basal forebrain complex and in acetylcholinesterase-containing fibers and varicosities within the hippocampus in the owl monkey (Aotus trivirgatus). Neurosci. Lett. 68, 17–22. doi: 10.1016/0304-3940(86)90222-3
Melander, T., Staines, W. A., Hokfelt, T., Rokaeus, A., Eckenstein, F., Salvaterra, P. M., et al. (1985). Galanin-like immunoreactivity in cholinergic neurons of the septum-basal forebrain complex projecting to the hippocampus of the rat. Brain Res. 360, 130–138. doi: 10.1016/0006-8993(85)91228-4
Melander, T., Staines, W. A., and Rokaeus, A. (1986d). Galanin-like immunoreactivity in hippocampal afferents in the rat, with special reference to cholinergic and noradrenergic inputs. Neuroscience 19, 223–240.
Melia, K. R., Rasmussen, K., Terwilliger, R. Z., Haycock, J. W., Nestler, E. J., Duman, R. S., et al. (1992). Coordinate regulation of the cyclic AMP system with firing rate and expression of tyrosine hydroxylase in the rat locus coeruleus: effects of chronic stress and drug treatments. J. Neurochem. 58, 494–502. doi: 10.1111/j.1471-4159.1992.tb09748.x
Mennicken, F., Hoffert, C., Pelletier, M., Ahmad, S., and O’Donnell, D. (2002). Restricted distribution of galanin receptor 3 (GalR3) mRNA in the adult rat central nervous system. J. Chem. Neuroanat. 24, 257–268. doi: 10.1016/S0891-0618(02)00068-6
Merchenthaler, I., Lopez, F. J., and Negro-Vilar, A. (1993). Anatomy and physiology of central galanin-containing pathways. Prog. Neurobiol. 40, 711–769. doi: 10.1016/0301-0082(93)90012-H
Merighi, A. (2002). Costorage and coexistence of neuropeptides in the mammalian CNS. Prog. Neurobiol. 66, 161–190. doi: 10.1016/S0301-0082(01)00031-4
Merrillees, N. C., Burnstock, G., and Holman, M. E. (1963). Correlation of fine structure and physiology of the innervation of smooth muscle in the guinea pig vas deferens. J. Cell Biol. 19, 529–550. doi: 10.1083/jcb.19.3.529
Michel, M. C., Wieland, T., and Tsujimoto, G. (2009). How reliable are G-protein-coupled receptor antibodies? Naunyn Schmiedebergs Arch. Pharmacol. 379, 385–388. doi: 10.1007/s00210-009-0395-y
Mill, J., and Petronis, A. (2007). Molecular studies of major depressive disorder: the epigenetic perspective. Mol. Psychiatry 12, 799–814. doi: 10.1038/sj.mp.4001992
Millan, M. J. (2006). Multi-target strategies for the improved treatment of depressive states: Conceptual foundations and neuronal substrates, drug discovery and therapeutic application. Pharmacology & therapeutics 110, 135–370. doi: 10.1016/j.pharmthera.2005.11.006
Millan, M. J. (2009). Dual- and triple-acting agents for treating core and co-morbid symptoms of major depression: novel concepts, new drugs. Neurotherapeutics 6, 53–77. doi: 10.1016/j.nurt.2008.10.039
Millan, M. J., Goodwin, G. M., Meyer-Lindenberg, A., and Ogren, S. O. (2015a). 60 years of advances in neuropsychopharmacology for improving brain health, renewed hope for progress. Eur. Neuropsychopharmacol. 25, 591–598. doi: 10.1016/j.euroneuro.2015.01.015
Millan, M. J., Goodwin, G. M., Meyer-Lindenberg, A., and Ogren, S. O. (2015b). Learning from the past and looking to the future: Emerging perspectives for improving the treatment of psychiatric disorders. Eur. Neuropsychopharmacol. 25, 599–656. doi: 10.1016/j.euroneuro.2015.01.016
Miller, M. A., Kolb, P. E., Leverenz, J. B., Peskind, E. R., and Raskind, M. A. (1999). Preservation of noradrenergic neurons in the locus ceruleus that coexpress galanin mRNA in Alzheimer’s disease. J. Neurochem. 73, 2028–2036.
Millon, C., Flores-Burgess, A., Castilla-Ortega, E., Gago, B., Garcia-Fernandez, M., Serrano, A., et al. (2017a). Central administration of galanin N-terminal fragment 1-15 decreases the voluntary alcohol intake in rats. Addict. Biol. doi: 10.1111/adb.12582 [Epub ahead of print].
Millon, C., Flores-Burgess, A., Narvaez, M., Borroto-Escuela, D. O., Gago, B., Santín, L., et al. (2017b). The neuropeptides galanin and galanin(1-15) in depression-like behaviours. Neuropeptides 64, 39–45. doi: 10.1016/j.npep.2017.01.004
Millon, C., Flores-Burgess, A., Narvaez, M., Borroto-Escuela, D. O., Santin, L., Parrado, C., et al. (2014). A role for galanin N-terminal fragment (1-15) in anxiety- and depression-related behaviors in rats. Int. J. Neuropsychopharmacol. 18:yu064.
Milner, T. A., Abate, C., Reis, D. J., and Pickel, V. M. (1989). Ultrastructural localization of phenylethanolamine N-methyltransferase-like immunoreactivity in the rat locus coeruleus. Brain Res. 478, 1–15. doi: 10.1016/0006-8993(89)91471-6
Milstein, C. (1990). The Croonian lecture, 1989. Antibodies: a paradigm for the biology of molecular recognition. Proc. R. Soc. Lond. B Biol. Sci. 239, 1–16. doi: 10.1098/rspb.1990.0006
Mitchell, V., Bouret, S., Howard, A. D., and Beauvillain, J. C. (1999). Expression of the galanin receptor subtype Gal-R2 mRNA in the rat hypothalamus. J. Chem. Neuroanat. 16, 265–277. doi: 10.1016/S0891-0618(99)00011-3
Mitsukawa, K., Lu, X., and Bartfai, T. (2008). Galanin, galanin receptors and drug targets. Cell Mol. Life Sci. 65, 1796–1805. doi: 10.1007/s00018-008-8153-8
Mitsukawa, K., Lu, X., and Bartfai, T. (2009). Bidirectional regulation of stress responses by galanin in mice: involvement of galanin receptor subtype 1. Neuroscience 160, 837–846. doi: 10.1016/j.neuroscience.2009.02.063
Moller, C., Sommer, W., Thorsell, A., and Heilig, M. (1999). Anxiogenic-like action of galanin after intra-amygdala administration in the rat. Neuropsychopharmacology 21, 507–512. doi: 10.1016/S0893-133X(98)00102-X
Mongeau, R., Blier, P., and de Montigny, C. (1997). The serotonergic and noradrenergic systems of the hippocampus: their interactions and the effects of antidepressant treatments. Brain Res. Brain Res. Rev. 23, 145–195. doi: 10.1016/S0165-0173(96)00017-3
Montgomery, S. A. (2006). Why do we need new and better antidepressants? Int. Clin. Psychopharm 21, S1–S10. doi: 10.1097/01.yic.0000199455.39552.1c
Moore, L. D., Le, T., and Fan, G. (2013). DNA methylation and its basic function. Neuropsychopharmacology 38, 23–38. doi: 10.1038/npp.2012.112
Moore, R. Y., and Bloom, F. E. (1979). Central catecholamine neuron systems: anatomy and physiology of the norepinephrine and epinephrine systems. Annu. Rev. Neurosci. 2, 113–168. doi: 10.1146/annurev.ne.02.030179.000553
Moore, R. Y., and Gustafson, E. L. (1989). The distribution of dopamine-beta-hydroxylase, neuropeptide Y and galanin in locus coeruleus neurons. J. Chem. Neuroanat. 2, 95–106.
Moreau, J. L., Jenck, F., Martin, J. R., Mortas, P., and Haefely, W. E. (1992). Antidepressant treatment prevents chronic unpredictable mild stress-induced anhedonia as assessed by ventral tegmentum self-stimulation behavior in rats. Eur. Neuropsychopharmacol. 2, 43–49. doi: 10.1016/0924-977X(92)90035-7
Morgan, C. A. III, Rasmusson, A. M., Wang, S., Hoyt, G., Hauger, R. L., and Hazlett, G. (2002). Neuropeptide-Y, cortisol, and subjective distress in humans exposed to acute stress: replication and extension of previous report. Biol. Psychiatry 52, 136–142. doi: 10.1016/S0006-3223(02)01319-7
Morris, J. F., Budd, T. C., Epton, M. J., Ma, D., Pow, D. V., and Wang, H. (1998). Functions of the perikaryon and dendrites in magnocellular vasopressin-secreting neurons: new insights from ultrastructural studies. Prog. Brain Res. 119, 21–30. doi: 10.1016/S0079-6123(08)61559-9
Morrison, J. H., Grzanna, R., Molliver, M. E., and Coyle, J. T. (1978). The distribution and orientation of noradrenergic fibers in neocortex of the rat: an immunofluorescence study. J. Comp. Neurol. 181, 17–39. doi: 10.1002/cne.901810103
Mullis, K. B., Erlich, H. A., Arnheim, N., Horn, G. T., Saiki, R. K., and Scharf, S. J. (1987). Process for amplifying nucleic acid sequences. U.S. Patent No 4,683,202.
Murphy, R. B., and Schuster, D. I. (1982). Modulation of [3H]-dopamine binding by cholecystokinin octapeptide (CCK-8). Peptides 3, 539–543. doi: 10.1016/0196-9781(82)90123-1
Murray, C. J., and Lopez, A. D. (1997). Global mortality, disability, and the contribution of risk factors: global burden of disease study. Lancet 349, 1436–1442. doi: 10.1016/S0140-6736(96)07495-8
Mutt, V. (1989). Hormonal Peptides, Fidia Research Foundation Neuroscience Award Lectures. New York, NY: Raven Press, 125–218.
Nagy, C., Vaillancourt, K., and Turecki, G. (2018). A role for activity-dependent epigenetics in the development and treatment of major depressive disorder. Genes Brain Behav. 17:e12446. doi: 10.1111/gbb.12446
Nassel, D. R. (2018). Substrates for Neuronal Cotransmission With Neuropeptides and Small Molecule Neurotransmitters in Drosophila. Front. Cell. Neurosci. 12:83. doi: 10.3389/fncel.2018.00083
Nemeroff, C. B., and Vale, W. W. (2005). The neurobiology of depression: inroads to treatment and new drug discovery. J. Clin. Psychiatry 66(Suppl. 7), 5–13.
Nestler, E. J. (2015). Role of the brain’s reward circuitry in depression: transcriptional mechanisms. Int. Rev. Neurobiol. 124, 151–170. doi: 10.1016/bs.irn.2015.07.003
Nestler, E. J., Barrot, M., DiLeone, R. J., Eisch, A. J., Gold, S. J., and Russell, J. (2002). Neurobiology of depression. Neuron 34, 13–25. doi: 10.1016/S0896-6273(02)00653-0
Nestler, E. J.,and Carlezon, W. A. Jr. (2006). The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry 59, 1151–1159. doi: 10.1016/j.biopsych.2005.09.018
Noda, M., Teranishi, Y., Takahashi, H., Toyosato, M., Notake, M., Nakanishi, S., et al. (1982). Isolation and structural organization of the human preproenkephalin gene. Nature 297, 431–434. doi: 10.1038/297431a0
Nusbaum, M. P., Blitz, D. M., and Marder, E. (2017). Functional consequences of neuropeptide and small-molecule co-transmission. Nat. Rev. Neurosci. 18, 389–403. doi: 10.1038/nrn.2017.56
O’Donnell, D., Ahmad, S., Wahlestedt, C., and Walker, P. (1999). Expression of the novel galanin receptor subtype GALR2 in the adult rat CNS: distinct distribution from GALR1. J. Comp. Neurol. 409, 469–481. doi: 10.1002/(SICI)1096-9861(19990705)409:3<469::AID-CNE10>3.0.CO;2-Q
O’Donnell, D. F. M., Hoffert, C., Hubatsch, D., Pelletier, M., Walker, P., and Ahmad, S. (2003). “Localization of galanin receptor subtypes in the rat CNS,” in Handbook of Chemical Neuroanatomy, Vol. 20, Peptide Receptors, eds R. Quirion, A. Bjorklnud, and T. Hokfelt (Amsterdam: Elsevier), 195–244.
Ogren, S. O., Kuteeva, E., Elvander-Tottie, E., and Hokfelt, T. (2010). Neuropeptides in learning and memory processes with focus on galanin. Eur. J. Pharmacol. 626, 9–17. doi: 10.1016/j.ejphar.2009.09.070
Ogren, S. O., Kuteeva, E., Hokfelt, T., and Kehr, J. (2006). Galanin receptor antagonists : a potential novel pharmacological treatment for mood disorders. CNS Drugs 20, 633–654. doi: 10.2165/00023210-200620080-00003
Ogren, S. O., Razani, H., Elvander-Tottie, E., and Kehr, J. (2007). The neuropeptide galanin as an in vivo modulator of brain 5-HT1A receptors: possible relevance for affective disorders. Physiol. Behav. 92, 172–179. doi: 10.1016/j.physbeh.2007.05.022
Ohtaki, T., Kumano, S., Ishibashi, Y., Ogi, K., Matsui, H., Harada, M., et al. (1999). Isolation and cDNA cloning of a novel galanin-like peptide (GALP) from porcine hypothalamus. J. Biol. Chem. 274, 37041–37045. doi: 10.1074/jbc.274.52.37041
Okbay, A., Baselmans, B. M. L., De Neve, J. E., Turley, P., Nivard, M. G., Fontana, M. A., et al. (2016). Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses. Nat. Genet. 48, 624–633. doi: 10.1038/ng.3552
O’Neal, H. A., Van Hoomissen, J. D., Holmes, P. V., and Dishman, R. K. (2001). Prepro-galanin messenger RNA levels are increased in rat locus coeruleus after treadmill exercise training. Neurosci. Lett. 299, 69–72. doi: 10.1016/S0304-3940(00)01780-8
Ong, L. K., Guan, L., Damanhuri, H., Goodchild, A. K., Bobrovskaya, L., Dickson, P. W., et al. (2014). Neurobiological consequences of acute footshock stress: effects on tyrosine hydroxylase phosphorylation and activation in the rat brain and adrenal medulla. J. Neurochem. 128, 547–560. doi: 10.1111/jnc.12482
Ornung, G., Shupliakov, O., Ottersen, O. P., Storm-Mathisen, J., and Cullheim, S. (1994). Immunohistochemical evidence for coexistence of glycine and GABA in nerve terminals on cat spinal motoneurones: an ultrastructural study. Neuroreport 5, 889–892. doi: 10.1097/00001756-199404000-00009
Otsuka, M., Konishi, S., and Takahashi, T. (1975). Hypothalamic substance P as a candidate for transmitter of primary afferent neurons. Fed. Proc. 34, 1922–1928.
Ottersen, O. P., Storm-Mathisen, J., and Somogyi, P. (1988). Colocalization of glycine-like and GABA-like immunoreactivities in Golgi cell terminals in the rat cerebellum: a postembedding light and electron microscopic study. Brain Res. 450, 342–353. doi: 10.1016/0006-8993(88)91573-9
Ovsepian, S. V., and Dolly, J. O. (2011). Dendritic SNAREs add a new twist to the old neuron theory. Proc. Natl. Acad. Sci. U.S.A. 108, 19113–19120. doi: 10.1073/pnas.1017235108
Page, M. E., Berridge, C. W., Foote, S. L., and Valentino, R. J. (1993). Corticotropin-releasing factor in the locus coeruleus mediates EEG activation associated with hypotensive stress. Neurosci. Lett. 164, 81–84. doi: 10.1016/0304-3940(93)90862-F
Page, M. E., and Valentino, R. J. (1994). Locus coeruleus activation by physiological challenges. Brain Res. Bull. 35, 557–560. doi: 10.1016/0361-9230(94)90169-4
Palkovits, M. (1995). Neuropeptide messenger plasticity in the CNS neurons following axotomy. Mol. Neurobiol. 10, 91–103. doi: 10.1007/BF02740669
Parker, D., and Grillner, S. (1999). Long-lasting substance-P-mediated modulation of NMDA-induced rhythmic activity in the lamprey locomotor network involves separate RNA- and protein-synthesis-dependent stages. Eur. J. Neurosci. 11, 1515–1522. doi: 10.1046/j.1460-9568.1999.00565.x
Paul, B. D., and Snyder, S. H. (2015). H2S: a novel gasotransmitter that signals by sulfhydration. Trends Biochem. Sci. 40, 687–700. doi: 10.1016/j.tibs.2015.08.007
Pelletier, G., Leclerc, R., Dube, D., Labrie, F., Puviani, R., Arimura, A., et al. (1975). Localization of growth hormone-release-inhibiting hormone (somatostatin) in the rat brain. Am. J. Anat. 142, 397–401. doi: 10.1002/aja.1001420309
Pelletier, G., Steinbusch, H. W., and Verhofstad, A. A. (1981). Immunoreactive substance P and serotonin present in the same dense-core vesicles. Nature 293, 71–72. doi: 10.1038/293071a0
Pena, C. J., Kronman, H. G., Walker, D. M., Cates, H. M., Bagot, R. C., Purushothaman, I., et al. (2017). Early life stress confers lifelong stress susceptibility in mice via ventral tegmental area OTX2. Science 356, 1185–1188. doi: 10.1126/science.aan4491
Pendergrass, K., Hargreaves, R., Petty, K. J., Carides, A. D., Evans, J. K., and Horgan, K. J. (2004). Aprepitant: an oral NK1 antagonist for the prevention of nausea and vomiting induced by highly emetogenic chemotherapy. Drugs Today 40, 853–863. doi: 10.1358/dot.2004.40.10.863745
Perez, S. E., Wynick, D., Steiner, R. A., and Mufson, E. J. (2001). Distribution of galaninergic immunoreactivity in the brain of the mouse. J. Comp. Neurol. 434, 158–185. doi: 10.1002/cne.1171
Petschner, P., Juhasz, G., Tamasi, V., Adori, C., Tothfalusi, L., Hökfelt, T., et al. (2016). Chronic venlafaxine treatment fails to alter the levels of galanin system transcripts in normal rats. Neuropeptides 57, 65–70. doi: 10.1016/j.npep.2016.01.010
Pfenninger, K., Sandri, C., Akert, K., and Eugster, C. H. (1969). Contribution to the problem of structural organization of the presynaptic area. Brain Res. 12, 10–18. doi: 10.1016/0006-8993(69)90051-1
Picciotto, M. R. (2008). Galanin and addiction. Cell Mol. Life Sci. 65, 1872–1879. doi: 10.1007/s00018-008-8151-x
Picciotto, M. R., Brabant, C., Einstein, E. B., Kamens, H. M., and Neugebauer, N. M. (2010). Effects of galanin on monoaminergic systems and HPA axis: potential mechanisms underlying the effects of galanin on addiction- and stress-related behaviors. Brain Res. 1314, 206–218. doi: 10.1016/j.brainres.2009.08.033
Pieribone, V. A., Xu, Z. Q., Zhang, X., Grillner, S., Bartfai, T., and Hökfelt, T. (1995). Galanin induces a hyperpolarization of norepinephrine-containing locus coeruleus neurons in the brainstem slice. Neuroscience 64, 861–874. doi: 10.1016/0306-4522(94)00450-J
Pletscher, A., Shore, P. A., and Brodie, B. B. (1955). Serotonin release as a possible mechanism of reserpine action. Science 122, 374–375. doi: 10.1126/science.122.3165.374
Poulter, M. O., Du, L., Zhurov, V., Palkovits, M., Faludi, G., Merali, Z., et al. (2010). Altered organization of GABA(A) receptor mRNA expression in the depressed suicide brain. Front. Mol. Neurosci. 3:3. doi: 10.3389/neuro.02.003.2010
Priestley, J. V., Wotherspoon, G., Savery, D., Averill, S., and Rattray, M. (1993). A combined in situ hybridization and immunofluorescence procedure allowing visualisation of peptide mRNA and serotonin in single sections. J. Neurosci. Methods 48, 99–110. doi: 10.1016/S0165-0270(05)80011-1
Pudovkina, O. L., Kawahara, Y., de Vries, J., and Westerink, B. H. (2001). The release of noradrenaline in the locus coeruleus and prefrontal cortex studied with dual-probe microdialysis. Brain Res. 906, 38–45. doi: 10.1016/S0006-8993(01)02553-7
Pudovkina, O. L., and Westerink, B. H. (2005). Functional role of alpha1-adrenoceptors in the locus coeruleus: a microdialysis study. Brain Res. 1061, 50–56. doi: 10.1016/j.brainres.2005.08.049
Renaud, L. P., Martin, J. B., and Brazeau, P. (1975). Depressant action of TRH. LH-RH and somatostatin on activity of central neurones. Nature 255, 233–235. doi: 10.1038/255233a0
Richardson, K. C. (1966). Electron microscopic identification of autonomic nerve endings. Nature 210:756. doi: 10.1038/210756a0
Rivier, J., Spiess, J., Thorner, M., and Vale, W. (1982). Characterization of a growth hormone-releasing factor from a human pancreatic islet tumour. Nature 300, 276–278. doi: 10.1038/300276a0
Robinson, J., Smith, A., Sturchler, E., Tabrizifard, S., Kamenecka, T., and McDonald, P. (2013). Development of a high-throughput screening-compatible cell-based functional assay to identify small molecule probes of the galanin 3 receptor (GalR3). Assay Drug Dev. Technol. 11, 468–477. doi: 10.1089/adt.2013.526
Robinson, J. K., Bartfai, T., and Langel, U. (2006). Galanin/GALP receptors and CNS homeostatic processes. CNS Neurol. Disord. Drug Targets 5, 327–334. doi: 10.2174/187152706777452281
Robinson, J. K., and Brewer, A. (2008). Galanin: a potential role in mesolimbic dopamine-mediated instrumental behavior. Neurosci. Biobehav. Rev. 32, 1485–1493. doi: 10.1016/j.neubiorev.2008.05.025
Rokaeus, A., and Brownstein, M. J. (1986). Construction of a porcine adrenal medullary cDNA library and nucleotide sequence analysis of two clones encoding a galanin precursor. Proc. Natl. Acad. Sci. U.S.A. 83, 6287–6291. doi: 10.1073/pnas.83.17.6287
Rokaeus, A., Melander, T., Hokfelt, T., Lundberg, J. M., Tatemoto, K., Carlquist, M., et al. (1984). A galanin-like peptide in the central nervous system and intestine of the rat. Neurosci. Lett. 47, 161–166. doi: 10.1016/0304-3940(84)90423-3
Rosenfeld, M. G., Mermod, J. J., Amara, S. G., Swanson, L. W., Sawchenko, P. E., Rivier, J., et al. (1983). Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature 304, 129–135. doi: 10.1038/304129a0
Rosin, D. L., Weston, M. C., Sevigny, C. P., Stornetta, R. L., and Guyenet, P. G. (2003). Hypothalamic orexin (hypocretin) neurons express vesicular glutamate transporters VGLUT1 or VGLUT2. J. Comp. Neurol. 465, 593–603. doi: 10.1002/cne.10860
Rovin, M. L., Boss-Williams, K. A., Alisch, R. S., Ritchie, J. C., Weinshenker, D., West, C. H., et al. (2012). Influence of chronic administration of antidepressant drugs on mRNA for galanin, galanin receptors, and tyrosine hydroxylase in catecholaminergic and serotonergic cell-body regions in rat brain. Neuropeptides 46, 81–91. doi: 10.1016/j.npep.2012.01.001
Roy, B., Wang, Q., Palkovits, M., Faludi, G., and Dwivedi, Y. (2017). Altered miRNA expression network in Locus coeruleus of depressed suicide subjects. Sci. Rep. 7:4387. doi: 10.1038/s41598-017-04300-9
Rupniak, N. M. J., and Kramer, M. S. (2017). NK1 receptor antagonists for depression: Why a validated concept was abandoned. J. Affect. Disord. 223, 121–125. doi: 10.1016/j.jad.2017.07.042
Rusnak, M., Zorad, S., Buckendahl, P., Sabban, E. L., and Kvetnansky, R. (1998). Tyrosine hydroxylase mRNA levels in locus ceruleus of rats during adaptation to long-term immobilization stress exposure. Mol. Chem. Neuropathol. 33, 249–258. doi: 10.1007/BF02815186
Russo, S. J., Murrough, J. W., Han, M. H., Charney, D. S., and Nestler, E. J. (2012). Neurobiology of resilience. Nat. Neurosci. 15, 1475–1484. doi: 10.1038/nn.3234
Saar, I., Lahe, J., Langel, K., Runesson, J., Webling, K., Järv, J., et al. (2013a). Novel systemically active galanin receptor 2 ligands in depression-like behavior. J. Neurochem. 127, 114–123. doi: 10.1111/jnc.12274
Saar, I., Runesson, J., Jarv, J., Kurrikoff, K., and Langel, U. (2013b). Novel galanin receptor subtype specific ligand in depression like behavior. Neurochem. Res. 38, 398–404. doi: 10.1007/s11064-012-0933-8
Saavedra, K., Molina-Marquez, A. M., Saavedra, N., Zambrano, T., and Salazar, L. A. (2016). Epigenetic modifications of major depressive disorder. Int. J. Mol. Sci. 17:E1279. doi: 10.3390/ijms17081279
Sabban, E. L., Alaluf, L. G., and Serova, L. I. (2016). Potential of neuropeptide Y for preventing or treating post-traumatic stress disorder. Neuropeptides 56, 19–24. doi: 10.1016/j.npep.2015.11.004
Sadakata, T., Mizoguchi, A., Sato, Y., Katoh-Semba, R., Fukuda, M., Mikoshiba, K., et al. (2004). The secretory granule-associated protein CAPS2 regulates neurotrophin release and cell survival. J. Neurosci. 24, 43–52. doi: 10.1523/JNEUROSCI.2528-03.2004
Sajdyk, T. J., Shekhar, A., and Gehlert, D. R. (2004). Interactions between NPY and CRF in the amygdala to regulate emotionality. Neuropeptides 38, 225–234. doi: 10.1016/j.npep.2004.05.006
Sakurai, T., Amemiya, A., Ishii, M., Matsuzaki, I., Chemelli, R. M., Tanaka, H., et al. (1998). Orexins and orexin receptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 92, 573–585. doi: 10.1016/S0092-8674(00)80949-6
Salio, C., Lossi, L., Ferrini, F., and Merighi, A. (2006). Neuropeptides as synaptic transmitters. Cell Tissue Res. 326, 583–598. doi: 10.1007/s00441-006-0268-3
Samuels, E. R., and Szabadi, E. (2008). Functional neuroanatomy of the noradrenergic locus coeruleus: its roles in the regulation of arousal and autonomic function part II: physiological and pharmacological manipulations and pathological alterations of locus coeruleus activity in humans. Curr. Neuropharmacol. 6, 254–285. doi: 10.2174/157015908785777193
Santic, R., Fenninger, K., Graf, K., Schneider, R., Hauser-Kronberger, C., Schilling, F. H., et al. (2006). Gangliocytes in neuroblastic tumors express alarin, a novel peptide derived by differential splicing of the galanin-like peptide gene. J. Mol. Neurosci. 29, 145–152. doi: 10.1385/JMN:29:2:145
Santos, R., Ursu, O., Gaulton, A., Bento, A. P., Donadi, R. S., Bologa, C. G., et al. (2017). A comprehensive map of molecular drug targets. Nat. Rev. Drug Discov. 16, 19–34. doi: 10.1038/nrd.2016.230
Schally, A. V., Coy, D. H., and Meyers, C. A. (1978). Hypothalamic regulatory hormones. Annu. Rev. Biochem. 47, 89–128. doi: 10.1146/annurev.bi.47.070178.000513
Schank, J. R., and Heilig, M. (2017). Substance P and the neurokinin-1 receptor: the new CRF. Int. Rev. Neurobiol. 136, 151–175. doi: 10.1016/bs.irn.2017.06.008
Scharrer, E., and Scharrer, B. (1937). Über Drüsen-Nervenzellen und Neurosekretorische Organe bei Wirbellosen und Wirbeltieren. Biol. Rev. 12, 185–216. doi: 10.1111/j.1469-185X.1937.tb01229.x
Schatzberg, A. S., and Schildkraut, J. J. (1995). “Recent studies on norepinephrine systems im mood disorders,” in Psychopharmacology: The Fourth Generation of Progress, eds F. Bloom and D. Kupfer (New York, NY: Raven Press), 911–920.
Schildkraut, J. J. (1965). The catecholamine hypothesis of affective disorders: a review of supporting evidence. Am. J. Psychiatry 122, 509–522. doi: 10.1176/ajp.122.5.509
Schmidt, W. E., Kratzin, H., Eckart, K., Drevs, D., Mundkowski, G., Clemens, A., et al. (1991). Isolation and primary structure of pituitary human galanin, a 30-residue nonamidated neuropeptide. Proc. Natl. Acad. Sci. U.S.A. 88, 11435–11439. doi: 10.1073/pnas.88.24.11435
Schwab, Y., Mouton, J., Chasserot-Golaz, S., Marty, I., Maulet, Y., Jover, E., et al. (2001). Calcium-dependent translocation of synaptotagmin to the plasma membrane in the dendrites of developing neurones. Brain Res. Mol. Brain Res. 96, 1–13. doi: 10.1016/S0169-328X(01)00244-3
Sciolino, N. R., Dishman, R. K., and Holmes, P. V. (2012). Voluntary exercise offers anxiolytic potential and amplifies galanin gene expression in the locus coeruleus of the rat. Behav. Brain Res. 233, 191–200. doi: 10.1016/j.bbr.2012.05.001
Sciolino, N. R., Smith, J. M., Stranahan, A. M., Freeman, K. G., Edwards, G. L., Weinshenker, D., et al. (2015). Galanin mediates features of neural and behavioral stress resilience afforded by exercise. Neuropharmacology 89, 255–264. doi: 10.1016/j.neuropharm.2014.09.029
Seidah, N. G., and Chretien, M. (1999). Proprotein and prohormone convertases: a family of subtilases generating diverse bioactive polypeptides. Brain Res. 848, 45–62. doi: 10.1016/S0006-8993(99)01909-5
Seki, K., Yoshida, S., and Jaiswal, M. K. (2018). Molecular mechanism of noradrenaline during the stress-induced major depressive disorder. Neural Regen. Res. 13, 1159–1169. doi: 10.4103/1673-5374.235019
Sequeira, A., Mamdani, F., Ernst, C., Vawter, M. P., Bunney, W. E., Lebel, V., et al. (2009). Global brain gene expression analysis links glutamatergic and GABAergic alterations to suicide and major depression. PLoS One 4:e6585. doi: 10.1371/journal.pone.0006585
Sequeira, A., Morgan, L., Walsh, D. M., Cartagena, P. M., Choudary, P., Li, J., et al. (2012). Gene expression changes in the prefrontal cortex, anterior cingulate cortex and nucleus accumbens of mood disorders subjects that committed suicide. PLoS One 7:e35367. doi: 10.1371/journal.pone.0035367
Sergeyev, V., Fetissov, S., Mathe, A. A., Jimenez, P. A., Bartfai, T., Mortas, P., et al. (2005). Neuropeptide expression in rats exposed to chronic mild stresses. Psychopharmacology 178, 115–124. doi: 10.1007/s00213-004-2015-3
Seutin, V., Verbanck, P., Massotte, L., and Dresse, A. (1989). Galanin decreases the activity of locus coeruleus neurons in vitro. Eur. J. Pharmacol. 164, 373–376. doi: 10.1016/0014-2999(89)90481-0
Sevcik, J., Finta, E. P., and Illes, P. (1993). Galanin receptors inhibit the spontaneous firing of locus coeruleus neurones and interact with mu-opioid receptors. Eur. J. Pharmacol. 230, 223–230. doi: 10.1016/0014-2999(93)90806-S
Shaib, A. H., Staudt, A., Harb, A., Klose, M., Shaaban, A., Schirra, C., et al. (2018). Paralogs of the calcium-dependent activator protein for secretion differentially regulate synaptic transmission and peptide secretion in sensory neurons. Front. Cell Neurosci. 12:304. doi: 10.3389/fncel.2018.00304
Shen, P. J., Larm, J. A., and Gundlach, A. L. (2003). Expression and plasticity of galanin systems in cortical neurons, oligodendrocyte progenitors and proliferative zones in normal brain and after spreading depression. Eur. J. Neurosci. 18, 1362–1376. doi: 10.1046/j.1460-9568.2003.02860.x
Shen, P. J., Yuan, C. G., Ma, J., Cheng, S., Yao, M., Turnley, A. M., et al. (2005). Galanin in neuro(glio)genesis: expression of galanin and receptors by progenitor cells in vivo and in vitro and effects of galanin on neurosphere proliferation. Neuropeptides 39, 201–205. doi: 10.1016/j.npep.2004.12.021
Shimizu, N., Katoh, Y., Hida, T., and Satoh, K. (1979). The fine structural organization of the locus coeruleus in the rat with reference to noradrenaline contents. Exp. Brain Res. 37, 139–148. doi: 10.1007/BF01474260
Shinoda, Y., Sadakata, T., Nakao, K., Katoh-Semba, R., Kinameri, E., Furuya, A., et al. (2011). Calcium-dependent activator protein for secretion 2 (CAPS2) promotes BDNF secretion and is critical for the development of GABAergic interneuron network. Proc. Natl. Acad. Sci. U.S.A. 108, 373–378. doi: 10.1073/pnas.1012220108
Shumyatsky, G. P., Tsvetkov, E., Malleret, G., Vronskaya, S., Hatton, M., Hampton, L., et al. (2002). Identification of a signaling network in lateral nucleus of amygdala important for inhibiting memory specifically related to learned fear. Cell 111, 905–918. doi: 10.1016/S0092-8674(02)01116-9
Sibille, E., Wang, Y., Joeyen-Waldorf, J., Gaiteri, C., Surget, A., Oh, S., et al. (2009). A molecular signature of depression in the amygdala. Am. J. Psychiatry 166, 1011–1024. doi: 10.1176/appi.ajp.2009.08121760
Silberstein, S. D., Dodick, D. W., Bigal, M. E., Yeung, P. P., Goadsby, P. J., Blankenbiller, T., et al. (2017). Fremanezumab for the preventive treatment of chronic migraine. N. Engl. J. Med. 377, 2113–2122. doi: 10.1056/NEJMoa1709038
Sillard, R., Rokaeus, A., Xu, Y., Carlquist, M., Bergman, T., Jörnvall, H., et al. (1992). Variant forms of galanin isolated from porcine brain. Peptides 13, 1055–1060. doi: 10.1016/0196-9781(92)90005-N
Silote, G. P., Rosal, A. B., de Souza, M. M., and Beijamini, V. (2013). Infusion of galanin into the mid-caudal portion of the dorsal raphe nucleus has an anxiolytic effect on rats in the elevated T-maze. Behav. Brain Res. 252, 312–317. doi: 10.1016/j.bbr.2013.06.023
Simson, P. E., and Weiss, J. M. (1988). Altered activity of the locus coeruleus in an animal model of depression. Neuropsychopharmacology 1, 287–295.
Singewald, N., and Philippu, A. (1998). Release of neurotransmitters in the locus coeruleus. Prog. Neurobiol. 56, 237–267. doi: 10.1016/S0301-0082(98)00039-2
Skofitsch, G., and Jacobowitz, D. M. (1985). Immunohistochemical mapping of galanin-like neurons in the rat central nervous system. Peptides 6, 509–546. doi: 10.1016/0196-9781(85)90118-4
Skofitsch, G., and Jacobowitz, D. M. (1986). Quantitative distribution of galanin-like immunoreactivity in the rat central nervous system. Peptides 7, 609–613. doi: 10.1016/0196-9781(86)90035-5
Skofitsch, G., Sills, M. A., and Jacobowitz, D. M. (1986). Autoradiographic distribution of 125I-galanin binding sites in the rat central nervous system. Peptides 7, 1029–1042. doi: 10.1016/0196-9781(86)90133-6
Smith, K. E., Walker, M. W., Artymyshyn, R., Bard, J., Borowsky, B., Tamm, J. A., et al. (1998). Cloned human and rat galanin GALR3 receptors. Pharmacology and activation of G-protein inwardly rectifying K+ channels. J. Biol. Chem. 273, 23321–23326. doi: 10.1074/jbc.273.36.23321
Smith, M. A., Brady, L. S., Glowa, J., Gold, P. W., and Herkenham, M. (1991). Effects of stress and adrenalectomy on tyrosine hydroxylase mRNA levels in the locus ceruleus by in situ hybridization. Brain Res. 544, 26–32. doi: 10.1016/0006-8993(91)90881-U
Snyder, S. H., and Ferris, C. D. (2000). Novel neurotransmitters and their neuropsychiatric relevance. Am. J. Psychiatry 157, 1738–1751. doi: 10.1176/appi.ajp.157.11.1738
Somogyi, P., and Klausberger, T. (2005). Defined types of cortical interneurone structure space and spike timing in the hippocampus. J. Physiol. 562, 9–26. doi: 10.1113/jphysiol.2004.078915
Southwick, S. M., Vythilingam, M., and Charney, D. S. (2005). The psychobiology of depression and resilience to stress: implications for prevention and treatment. Annu. Rev. Clin. Psychol. 1, 255–291. doi: 10.1146/annurev.clinpsy.1.102803.143948
Spector, J. M., Harrison, R. S., and Fishman, M. C. (2018). Fundamental science behind today’s important medicines. Sci. Transl. Med. 10:eaaq1787.
Spiess, J., Rivier, J., Rivier, C., and Vale, W. (1981). Primary structure of corticotropin-releasing factor from ovine hypothalamus. Proc. Natl. Acad. Sci. U.S.A. 78, 6517–6521. doi: 10.1073/pnas.78.10.6517
Spiess, J., Rivier, J., and Vale, W. (1983). Characterization of rat hypothalamic growth hormone-releasing factor. Nature 303, 532–535. doi: 10.1038/303532a0
Spitzer, N. C. (2017). Neurotransmitter switching in the developing and adult brain. Annu. Rev. Neurosci. 40, 1–19. doi: 10.1146/annurev-neuro-072116-031204
Starr, M. S. (1982). Influence of peptides on (3)H-dopamine release from superfused rat striatal slices. Neurochem. Int. 4, 233–240. doi: 10.1016/0197-0186(82)90059-6
Steckler, T. (2008). “Peptide receptor ligands to treat anxiety disorders,” in Handbook of Anxiety and Fear, eds D. C. Blanchard, D. C. Blanchard, G. Griebel, and D. Nutt (Amsterdam: Elsevier), 157–217.
Steiner, R. A., Hohmann, J. G., Holmes, A., Wrenn, C. C., Cadd, G., Juréus, A., et al. (2001). Galanin transgenic mice display cognitive and neurochemical deficits characteristic of Alzheimer’s disease. Proc. Natl. Acad. Sci. U.S.A. 98, 4184–4189. doi: 10.1073/pnas.061445598
Stevens, D. R., and Rettig, J. (2009). The Ca(2+)-dependent activator protein for secretion CAPS: do I dock or do I prime? Mol. Neurobiol. 39, 62–72. doi: 10.1007/s12035-009-8052-5
Strand, F. (1991). Neuropeptides. Regulators of Physiological Processes. Cambridge, MA: The MIT Press.
Sudhof, T. C. (2014). The molecular machinery of neurotransmitter release (Nobel lecture). Angew. Chem. Int. Ed. Engl. 53, 12696–12717. doi: 10.1002/anie.201406359
Sudhof, T. C. (2017). Molecular neuroscience in the 21(st) century: a personal perspective. Neuron 96, 536–541. doi: 10.1016/j.neuron.2017.10.005
Sullivan, P. F., Neale, M. C., and Kendler, K. S. (2000). Genetic epidemiology of major depression: review and meta-analysis. Am. J. Psychiatry 157, 1552–1562. doi: 10.1176/appi.ajp.157.10.1552
Svensson, T. H. (1987). Stress, central neurotransmitters, and the mechanism of action of alpha 2-adrenoceptor agonists. J. Cardiovasc. Pharmacol. 10(Suppl. 12), S88–S92.
Svensson, T. H., Bunney, B. S., and Aghajanian, G. K. (1975). Inhibition of both noradrenergic and serotonergic neurons in brain by the alpha-adrenergic agonist clonidine. Brain Res. 92, 291–306. doi: 10.1016/0006-8993(75)90276-0
Swaab, D. F., Pool, C. W., and Nijveldt, F. (1975). Immunofluorescence of vasopressin and oxytocin in the rat hypothalamo-neurohypophypopseal system. J. Neural. Transm. 36, 195–215. doi: 10.1007/BF01253126
Swanson, C. J., Blackburn, T. P., Zhang, X., Zheng, K., Xu, Z. Q., Hökfelt, T., et al. (2005). Anxiolytic- and antidepressant-like profiles of the galanin-3 receptor (Gal3) antagonists SNAP 37889 and SNAP 398299. Proc. Natl. Acad. Sci. U.S.A. 102, 17489–17494. doi: 10.1073/pnas.0508970102
Swanson, L. W. (1976). The locus coeruleus: a cytoarchitectonic, Golgi and immunohistochemical study in the albino rat. Brain Res. 110, 39–56. doi: 10.1016/0006-8993(76)90207-9
Swanson, L. W., and Hartman, B. K. (1975). The central adrenergic system. An immunofluorescence study of the location of cell bodies and their efferent connections in the rat utilizing dopamine-beta-hydroxylase as a marker. J. Comp. Neurol. 163, 467–505. doi: 10.1002/cne.901630406
Swanson, L. W., Sawchenko, P. E., and Lind, R. W. (1986). Regulation of multiple peptides in CRF parvocellular neurosecretory neurons: implications for the stress response. Prog. Brain Res. 68, 169–190. doi: 10.1016/S0079-6123(08)60238-1
Sweerts, B. W., Jarrott, B., and Lawrence, A. J. (1999). Expression of preprogalanin mRNA following acute and chronic restraint stress in brains of normotensive and hypertensive rats. Brain Res. Mol. Brain Res. 69, 113–123. doi: 10.1016/S0169-328X(99)00095-9
Szot, P., Leverenz, J. B., Peskind, E. R., Kiyasu, E., Rohde, K., Miller, M. A., et al. (2000). Tyrosine hydroxylase and norepinephrine transporter mRNA expression in the locus coeruleus in Alzheimer’s disease. Brain Res. Mol. Brain Res. 84, 135–140. doi: 10.1016/S0169-328X(00)00168-6
Takahashi, T., and Otsuka, M. (1975). Regional distribution of substance P in the spinal cord and nerve roots of the cat and the effect of dorsal root section. Brain Res. 87, 1–11. doi: 10.1016/0006-8993(75)90774-X
Takeuchi, T., Duszkiewicz, A. J., Sonneborn, A., Spooner, P. A., Yamasaki, M., Watanabe, M., et al. (2016). Locus coeruleus and dopaminergic consolidation of everyday memory. Nature 537, 357–362. doi: 10.1038/nature19325
Tanti, A., Kim, J. J., Wakid, M., Davoli, M. A., Turecki, G., and Mechawar, N. (2017). Child abuse associates with an imbalance of oligodendrocyte-lineage cells in ventromedial prefrontal white matter. Mol. Psychiatry doi: 10.1038/mp.2017.231 [Epub ahead of print].
Tatemoto, K., Carlquist, M., and Mutt, V. (1982). Neuropeptide Y–a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 296, 659–660. doi: 10.1038/296659a0
Tatemoto, K., and Mutt, V. (1980). Isolation of two novel candidate hormones using a chemical method for finding naturally occurring polypeptides. Nature 285, 417–418. doi: 10.1038/285417a0
Tatemoto, K., Rokaeus, A., Jornvall, H., McDonald, T. J., and Mutt, V. (1983). Galanin - a novel biologically active peptide from porcine intestine. FEBS Lett. 164, 124–128. doi: 10.1016/0014-5793(83)80033-7
Thomas, M. J., Kalivas, P. W., and Shaham, Y. (2008). Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br. J. Pharmacol. 154, 327–342. doi: 10.1038/bjp.2008.77
Tochigi, M., Iwamoto, K., Bundo, M., Sasaki, T., Kato, N., and Kato, T. (2008). Gene expression profiling of major depression and suicide in the prefrontal cortex of postmortem brains. Neurosci. Res. 60, 184–191. doi: 10.1016/j.neures.2007.10.010
Todd, A. J., and Sullivan, A. C. (1990). Light microscope study of the coexistence of GABA-like and glycine-like immunoreactivities in the spinal cord of the rat. J. Comp. Neurol. 296, 496–505. doi: 10.1002/cne.902960312
Torres, G. E., and Amara, S. G. (2007). Glutamate and monoamine transporters: new visions of form and function. Curr. Opin. Neurobiol. 17, 304–312. doi: 10.1016/j.conb.2007.05.002
Torres-Platas, S. G., Nagy, C., Wakid, M., Turecki, G., and Mechawar, N. (2016). Glial fibrillary acidic protein is differentially expressed across cortical and subcortical regions in healthy brains and downregulated in the thalamus and caudate nucleus of depressed suicides. Mol. Psychiatry 21, 509–515. doi: 10.1038/mp.2015.65
Tortorella, C., Neri, G., and Nussdorfer, G. G. (2007). Galanin in the regulation of the hypothalamic-pituitary-adrenal axis (Review). Int. J. Mol. Med. 19, 639–647. doi: 10.3892/ijmm.19.4.639
Tritsch, N. X., Ding, J. B., and Sabatini, B. L. (2012). Dopaminergic neurons inhibit striatal output through non-canonical release of GABA. Nature 490, 262–266. doi: 10.1038/nature11466
Trivedi, M. H., Rush, A. J., Wisniewski, S. R., Nierenberg, A. A., Warden, D., Ritz, L., et al. (2006). Evaluation of outcomes with citalopram for depression using measurement-based care in STAR∗D: implications for clinical practice. Am. J. Psychiatry 163, 28–40. doi: 10.1176/appi.ajp.163.1.28
Trudeau, L. E., Hnasko, T. S., Wallen-Mackenzie, A., Morales, M., Rayport, S., Sulzer, D., et al. (2014). The multilingual nature of dopamine neurons. Prog. Brain Res. 211, 141–164. doi: 10.1016/B978-0-444-63425-2.00006-4
Ungerstedt, U. (1971). Stereotaxic mapping of the monoamine pathways in the rat brain. Acta Physiol. Scand. Suppl. 367, 1–48. doi: 10.1111/j.1365-201X.1971.tb10998.x
Ungerstedt, U. (1984). Measurement of neurotransmitter release by intracranial dialysis. Methods Neurosci. 6, 81–105.
Unschuld, P. G., Ising, M., Roeske, D., Erhardt, A., Specht, M., Kloiber, S., et al. (2010). Gender-specific association of galanin polymorphisms with HPA-axis dysregulation, symptom severity, and antidepressant treatment response. Neuropsychopharmacology 35, 1583–1592. doi: 10.1038/npp.2010.30
Uslaner, J. M., Tye, S. J., Eddins, D. M., Wang, X., Fox, S. V., Savitz, A. T., et al. (2013). Orexin receptor antagonists differ from standard sleep drugs by promoting sleep at doses that do not disrupt cognition. Sci. Transl. Med. 5:179ra144. doi: 10.1126/scitranslmed.3005213
Vahabzadeh, A., and Fillenz, M. (1994). Comparison of stress-induced changes in noradrenergic and serotonergic neurons in the rat hippocampus using microdialysis. Eur. J. Neurosci. 6, 1205–1212. doi: 10.1111/j.1460-9568.1994.tb00619.x
Vale, W., Brazeau, P., Rivier, C., Brown, M., Boss, B., Rivier, J., et al. (1975). Somatostatin. Recent Prog. Horm. Res. 31, 365–397. doi: 10.1016/B978-0-12-571131-9.50014-1
Vale, W., Spiess, J., Rivier, C., and Rivier, J. (1981). Characterization of a 41-residue ovine hypothalamic peptide that stimulates secretion of corticotropin and beta-endorphin. Science 213, 1394–1397. doi: 10.1126/science.6267699
Valentino, R. J., and Van Bockstaele, E. (2015). Endogenous opioids: the downside of opposing stress. Neurobiol. Stress 1, 23–32. doi: 10.1016/j.ynstr.2014.09.006
Van Bockstaele, E. J. (1998). Morphological substrates underlying opioid, epinephrine and gamma-aminobutyric acid inhibitory actions in the rat locus coeruleus. Brain Res. Bull. 47, 1–15. doi: 10.1016/S0361-9230(98)00062-8
Van Bockstaele, E. J., and Valentino, R. J. (2013). Neuropeptide regulation of the locus coeruleus and opiate-induced plasticity of stress responses. Adv. Pharmacol. 68, 405–420. doi: 10.1016/B978-0-12-411512-5.00019-1
van den Pol, A. N. (2012). Neuropeptide transmission in brain circuits. Neuron 76, 98–115. doi: 10.1016/j.neuron.2012.09.014
Vandesande, F., and Dierickx, K. (1975). Identification of the vasopressin producing and of the oxytocin producing neurons in the hypothalamic magnocellular neurosecretroy system of the rat. Cell Tissue Res. 164, 153–162. doi: 10.1007/BF00218970
Veit, J., Hakim, R., Jadi, M. P., Sejnowski, T. J., and Adesnik, H. (2017). Cortical gamma band synchronization through somatostatin interneurons. Nat. Neurosci. 20, 951–959. doi: 10.1038/nn.4562
Verhage, M., McMahon, H. T., Ghijsen, W. E., Boomsma, F., Scholten, G., Wiegant, V. M., et al. (1991). Differential release of amino acids, neuropeptides, and catecholamines from isolated nerve terminals. Neuron 6, 517–524. doi: 10.1016/0896-6273(91)90054-4
Vialou, V., Feng, J., Robison, A. J., and Nestler, E. J. (2013). Epigenetic mechanisms of depression and antidepressant action. Annu. Rev. Pharmacol. Toxicol. 53, 59–87. doi: 10.1146/annurev-pharmtox-010611-134540
Vila-Porcile, E., Xu, Z. Q., Mailly, P., Nagy, F., Calas, A., Hökfelt, T., et al. (2009). Dendritic synthesis and release of the neuropeptide galanin: morphological evidence from studies on rat locus coeruleus neurons. J. Comp. Neurol. 516, 199–212. doi: 10.1002/cne.22105
Villar, M. J., Meister, B., Cortes, R., Schalling, M., Morris, M., Hökfelt, T., et al. (1990). Neuropeptide gene expression in hypothalamic magnocellular neurons of normal and hypophysectomized rats: a combined immunohistochemical and in situ hybridization study. Neuroscience 36, 181–199. doi: 10.1016/0306-4522(90)90361-7
von Euler, U. S. (1948). Identification of the Sympathomimetic ergone in adrenergic nerves of cattle (Sympathin N) with laevo-noradrenaline. Acta Physiol. Scand. 16:63. doi: 10.1111/j.1748-1716.1948.tb00526.x
von Euler, U. S., and Gaddum, J. H. (1931). An unidentified depressor substance in certain tissue extracts. J. Physiol. 72, 74–87. doi: 10.1113/jphysiol.1931.sp002763
Walent, J. H., Porter, B. W., and Martin, T. F. (1992). A novel 145 kd brain cytosolic protein reconstitutes Ca(2+)-regulated secretion in permeable neuroendocrine cells. Cell 70, 765–775. doi: 10.1016/0092-8674(92)90310-9
Walker, L. C., Rance, N. E., Price, D. L., and Young, 3rd (1991). Galanin mRNA in the nucleus basalis of Meynert complex of baboons and humans. J. Comp. Neurol. 303, 113–120. doi: 10.1002/cne.903030110
Walton, K. M., Chin, J. E., Duplantier, A. J., and Mather, R. J. (2006). Galanin function in the central nervous system. Curr. Opin. Drug Discov. Dev. 9, 560–570.
Wang, P., Li, H., Barde, S., Zhang, M. D., Sun, J., Wang, T., et al. (2016). Depression-like behavior in rat: Involvement of galanin receptor subtype 1 in the ventral periaqueductal gray. Proc. Natl. Acad. Sci. U.S.A. 113, E4726–E4735. doi: 10.1073/pnas.1609198113
Wang, S., He, C., Hashemi, T., and Bayne, M. (1997). Cloning and expressional characterization of a novel galanin receptor. Identification of different pharmacophores within galanin for the three galanin receptor subtypes.J. Biol. Chem. 272, 31949–31952. doi: 10.1074/jbc.272.51.31949
Wang, Y. J., Li, H., Yang, Y. T., Tie, C. L., Li, F., David, et al. (2013). Association of galanin and major depressive disorder in the Chinese Han population. PLoS One 8:e64617. doi: 10.1371/journal.pone.0064617
Waters, S. M., and Krause, J. E. (2000). Distribution of galanin-1, -2 and -3 receptor messenger RNAs in central and peripheral rat tissues. Neuroscience 95, 265–271. doi: 10.1016/S0306-4522(99)00407-8
Webling, K. E., Runesson, J., Bartfai, T., and Langel, U. (2012). Galanin receptors and ligands. Front. Endocrinol. 3:146. doi: 10.3389/fendo.2012.00146
Weinshenker, D., and Holmes, P. V. (2016). Regulation of neurological and neuropsychiatric phenotypes by locus coeruleus-derived galanin. Brain Res. 1641, 320–337. doi: 10.1016/j.brainres.2015.11.025
Weiss, J. M., Bonsall, R. W., Demetrikopoulos, M. K., Emery, M. S., and West, C. H. (1998). Galanin: a significant role in depression? Ann. N. Y. Acad. Sci. 863, 364–382. doi: 10.1111/j.1749-6632.1998.tb10707.x
Weiss, J. M., Boss-Williams, K. A., Moore, J. P., Demetrikopoulos, M. K., Ritchie, J. C., West, C. H., et al. (2005). Testing the hypothesis that locus coeruleus hyperactivity produces depression-related changes via galanin. Neuropeptides 39, 281–287. doi: 10.1016/j.npep.2004.12.028
Weiss, J. M., Demetrikopoulos, M. K., West, K. A., and Bonsall, R. W. (1996). Hypothesis linking the noradrenergic and dopaminergic systems in depression. Depression 3, 225–245. doi: 10.1002/depr.3050030503
Weiss, J. M., Goodman, P. A., Losito, B. G., Corrigan, S., Charry, J. M., Bailey, W. H., et al. (1981). Behavioral depression produced by an uncontrollable stressor: relationship to norepinephrine, dopamine, and serotonin levels in various regions of rat brain. Brain Res. Rev. 3, 167–205. doi: 10.1016/0165-0173(81)90005-9
Weiss, J. M., Stout, J. C., Aaron, M. F., Quan, N., Owens, M. J., Butler, P. D., et al. (1994). Depression and anxiety: role of the locus coeruleus and corticotropin-releasing factor. Brain Res. Bull. 35, 561–572. doi: 10.1016/0361-9230(94)90170-8
Wilkinson, L. O., and Jacobs, B. L. (1988). Lack of response of serotonergic neurons in the dorsal raphe nucleus of freely moving cats to stressful stimuli. Exp. Neurol. 101, 445–457. doi: 10.1016/0014-4886(88)90055-6
Willner, P., Towell, A., Sampson, D., Sophokleous, S., and Muscat, R. (1987). Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 93, 358–364. doi: 10.1007/BF00187257
Wittchen, H. U., Jacobi, F., Rehm, J., Gustavsson, A., Svensson, M., Jönsson, B., et al. (2011). The size and burden of mental disorders and other disorders of the brain in Europe 2010. Eur. Neuropsychopharmacol. 21, 655–679. doi: 10.1016/j.euroneuro.2011.07.018
Wray, N. R., Pergadia, M. L., Blackwood, D. H., Penninx, B. W., Gordon, S. D., Nyholt, D. R., et al. (2012). Genome-wide association study of major depressive disorder: new results, meta-analysis, and lessons learned. Mol. Psychiatry 17, 36–48. doi: 10.1038/mp.2010.109
Wray, N. R., Ripke, S., Mattheisen, M., Trzaskowski, M., Byrne, E. M., Abdellaoui, A., et al. (2018). Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 50, 668–681. doi: 10.1038/s41588-018-0090-3
Wrenn, C. C., and Holmes, A. (2006). The role of galanin in modulating stress-related neural pathways. Drug News Perspect. 19, 461–467. doi: 10.1358/dnp.2006.19.8.1043963
Wu, G., Feder, A., Wegener, G., Bailey, C., Saxena, S., Charney, D., et al. (2011). Central functions of neuropeptide Y in mood and anxiety disorders. Expert Opin. Ther. Targets 15, 1317–1331. doi: 10.1517/14728222.2011.628314
Wu, Z., Autry, A. E., Bergan, J. F., Watabe-Uchida, M., and Dulac, C. G. (2014). Galanin neurons in the medial preoptic area govern parental behaviour. Nature 509, 325–330. doi: 10.1038/nature13307
Xia, X., Lessmann, V., and Martin, T. F. (2009). Imaging of evoked dense-core-vesicle exocytosis in hippocampal neurons reveals long latencies and kiss-and-run fusion events. J. Cell Sci. 122, 75–82. doi: 10.1242/jcs.034603
Xiao, X., Wang, L., Wang, C., Yuan, T. F., Zhou, D., Zheng, F., et al. (2017). Common variants at 2q11.2, 8q21.3, and 11q13.2 are associated with major mood disorders. Transl. Psychiatry 7:1273. doi: 10.1038/s41398-017-0019-0
Xu, X. J., Hokfelt, T., and Wiesenfeld-Hallin, Z. (2008). Galanin and spinal pain mechanisms: where do we stand in 2008? Cell Mol. Life Sci. 65, 1813–1819. doi: 10.1007/s00018-008-8155-6
Xu, Z., Cortes, R., Villar, M., Morino, P., Castel, M. N., and Hökfelt, T. (1992). Evidence for upregulation of galanin synthesis in rat glial cells in vivo after colchicine treatment. Neurosci. Lett. 145, 185–188. doi: 10.1016/0304-3940(92)90018-3
Xu, Z. Q., and Hokfelt, T. (1997). Expression of galanin and nitric oxide synthase in subpopulations of serotonin neurons of the rat dorsal raphe nucleus. J. Chem. Neuroanat. 13, 169–187. doi: 10.1016/S0891-0618(97)00043-4
Xu, Z. Q., Ma, X., Soomets, U., Langel, U., and Hokfelt, T. (1999). Electrophysiological evidence for a hyperpolarizing, galanin (1-15)-selective receptor on hippocampal CA3 pyramidal neurons. Proc. Natl. Acad. Sci. U.S.A. 96, 14583–14587. doi: 10.1073/pnas.96.25.14583
Xu, Z. Q., Shi, T. J., and Hokfelt, T. (1998). Galanin/GMAP- and NPY-like immunoreactivities in locus coeruleus and noradrenergic nerve terminals in the hippocampal formation and cortex with notes on the galanin-R1 and -R2 receptors. J. Comp. Neurol. 392, 227–251. doi: 10.1002/(SICI)1096-9861(19980309)392:2<227::AID-CNE6>3.0.CO;2-4
Xu, Z. Q., Tong, Y. G., and Hokfelt, T. (2001). Galanin enhances noradrenaline-induced outward current on locus coeruleus noradrenergic neurons. Neuroreport 12, 1779–1782. doi: 10.1097/00001756-200106130-00052
Xu, Z. Q., Zheng, K., and Hokfelt, T. (2005). Electrophysiological studies on galanin effects in brain–progress during the last six years. Neuropeptides 39, 269–275. doi: 10.1016/j.npep.2005.02.003
Yalow, R. S., and Berson, S. A. (1959). Assay of plasma insulin in human subjects by immunological methods. Nature 184(Suppl. 21), 1648–1649. doi: 10.1038/1841648b0
Yamada, M., Makino, Y., Hashimoto, T., Sugiyama, A., Oka, J., Inagaki, M., et al. (2013). Induction of galanin after chronic sertraline treatment in mouse ventral dentate gyrus. Brain Res. 1516, 76–82. doi: 10.1016/j.brainres.2013.04.002
Yamazoe, M., Shiosaka, S., Emson, P. C., and Tohyama, M. (1985). Distribution of neuropeptide Y in the lower brainstem: an immunohistochemical analysis. Brain Res. 335, 109–120. doi: 10.1016/0006-8993(85)90281-1
Yang, L. P. (2014). Suvorexant: first global approval. Drugs 74, 1817–1822. doi: 10.1007/s40265-014-0294-5
Yoshitake, T., Reenila, I., Ogren, S. O., Hokfelt, T., and Kehr, J. (2003). Galanin attenuates basal and antidepressant drug-induced increase of extracellular serotonin and noradrenaline levels in the rat hippocampus. Neurosci. Lett. 339, 239–242.
Yoshitake, T., Wang, F. H., Kuteeva, E., Holmberg, K., Yamaguchi, M., Crawley, J. N., et al. (2004). Enhanced hippocampal noradrenaline and serotonin release in galanin-overexpressing mice after repeated forced swimming test. Proc. Natl. Acad. Sci. U.S.A. 101, 354–359. doi: 10.1073/pnas.0307042101
Zachariou, V., Brunzell, D. H., Hawes, J., Stedman, D. R., Bartfai, T., Steiner, R. A., et al. (2003). The neuropeptide galanin modulates behavioral and neurochemical signs of opiate withdrawal. Proc. Natl. Acad. Sci. U.S.A. 100, 9028–9033. doi: 10.1073/pnas.1533224100
Zarate, C. A. Jr., Singh, J. B., Carlson, P. J., Brutsche, N. E., Ameli, R., Luckenbaugh, D. A., et al. (2006). A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 63, 856–864. doi: 10.1001/archpsyc.63.8.856
Zelikowsky, M., Hui, M., Karigo, T., Choe, A., Yang, B., Blanco, M. R., et al. (2018). The neuropeptide Tac2 controls a distributed brain state induced by chronic social isolation stress. Cell 173, 1265.e19–1279.e19. doi: 10.1016/j.cell.2018.03.037
Zhang, T. Y., and Meaney, M. J. (2010). Epigenetics and the environmental regulation of the genome and its function. Annu. Rev. Psychol. 61, C431–C433. doi: 10.1146/annurev.psych.60.110707.163625
Zhou, Z., Zhu, G., Hariri, A. R., Enoch, M. A., Scott, D., Sinha, R., et al. (2008). Genetic variation in human NPY expression affects stress response and emotion. Nature 452, 997–1001. doi: 10.1038/nature06858
Zhu, P. C., Thureson-Klein, A., and Klein, R. L. (1986). Exocytosis from large dense cored vesicles outside the active synaptic zones of terminals within the trigeminal subnucleus caudalis: a possible mechanism for neuropeptide release. Neuroscience 19, 43–54. doi: 10.1016/0306-4522(86)90004-7
Zhurov, V., Stead, J. D., Merali, Z., Palkovits, M., Faludi, G., Schild-Poulter, C., et al. (2012). Molecular pathway reconstruction and analysis of disturbed gene expression in depressed individuals who died by suicide. PLoS One 7:e47581. doi: 10.1371/journal.pone.0047581
Keywords: allostatic load, epigenetics, galanin, locus coeruleus, major depression disorder, neuropeptides, resilience
Citation: Hökfelt T, Barde S, Xu Z-QD, Kuteeva E, Rüegg J, Le Maitre E, Risling M, Kehr J, Ihnatko R, Theodorsson E, Palkovits M, Deakin W, Bagdy G, Juhasz G, Prud’homme HJ, Mechawar N, Diaz-Heijtz R and Ögren SO (2018) Neuropeptide and Small Transmitter Coexistence: Fundamental Studies and Relevance to Mental Illness. Front. Neural Circuits 12:106. doi: 10.3389/fncir.2018.00106
Received: 28 June 2018; Accepted: 05 November 2018;
Published: 21 December 2018.
Edited by:
Miguel Angel Morales, Universidad Nacional Autónoma de México, MexicoReviewed by:
Istvan Jozsef Merchenthaler, University of Maryland, Baltimore, United StatesCopyright © 2018 Hökfelt, Barde, Xu, Kuteeva, Rüegg, Le Maitre, Risling, Kehr, Ihnatko, Theodorsson, Palkovits, Deakin, Bagdy, Juhasz, Prud’homme, Mechawar, Diaz-Heijtz and Ögren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tomas Hökfelt, VG9tYXMuSG9rZmVsdEBraS5zZQ==
†Present address: Eugenia Kuteeva, Atlas Antibodies AB, Bromma, Sweden
Erwan Le Maitre, Unit of Rheumatology, Center for Molecular Medicine, Department of Medicine, Karolinska Institutet, Karolinska University Hospital, Stockholm, Sweden
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.