 Wenxue Dong
Wenxue Dong Fanghe Gong1†
Fanghe Gong1† Hongmin Bai
Hongmin Bai Ruixin Yang
Ruixin Yang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Neurosci. , 09 August 2023
Sec. Cellular Neuropathology
Volume 17 - 2023 | https://doi.org/10.3389/fncel.2023.1228968
Acute central nervous system injuries (ACNSI), encompassing traumatic brain injury (TBI), non-traumatic brain injury like stroke and encephalomeningitis, as well as spinal cord injuries, are linked to significant rates of disability and mortality globally. Nevertheless, effective and feasible treatment plans are still to be formulated. There are primary and secondary injuries occurred after ACNSI. Most ACNSIs exhibit comparable secondary injuries, which offer numerous potential therapeutic targets for enhancing clinical outcomes. Ferroptosis, a newly discovered form of cell death, is characterized as a lipid peroxidation process that is dependent on iron and oxidative conditions, which is also indispensable to mitochondria. Ferroptosis play a vital role in many neuropathological pathways, and ACNSIs may induce mitochondrial dysfunction, thereby indicating the essentiality of the mitochondrial connection to ferroptosis in ACNSIs. Nevertheless, there remains a lack of clarity regarding the involvement of mitochondria in the occurrence of ferroptosis as a secondary injuries of ACNSIs. In recent studies, anti-ferroptosis agents such as the ferroptosis inhibitor Ferrostain-1 and iron chelation therapy have shown potential in ameliorating the deleterious effects of ferroptosis in cases of traumatic ACNSI. The importance of this evidence is extremely significant in relation to the research and control of ACNSIs. Therefore, our review aims to provide researchers focusing on enhancing the therapeutic outcomes of ACNSIs with valuable insights by summarizing the physiopathological mechanisms of ACNSIs and exploring the correlation between ferroptosis, mitochondrial dysfunction, and ACNSIs.
Acute central nervous system injury (ACNSI) is the result of sudden damage to the brain or spinal cord due to different factors, including stroke, traumatic brain injury (TBI), and spinal cord injury (SCI) (Shein and Shohami, 2011). ACNSI is a prevalent condition that poses significant risks to human health and life (Shein and Shohami, 2011). This condition is marked by elevated occurrence and death rates, unfavorable treatment results, and may lead to significant impairments and societal impact (Shein and Shohami, 2011). ACNSI involves different types of cell death, such as apoptosis, necrosis, autophagy, and ferroptosis, which contribute to its onset and progression (Walker et al., 2009; El Sayed and Ghoneum, 2020; Bai and Bian, 2022; Sun et al., 2022). Ferroptosis, a form of programmed cell death, differs from other apoptosis as it relies on iron and the buildup of lipid peroxides and their byproducts (Zhang C. et al., 2022). Ferroptosis is intricately linked to various biological processes, encompassing cellular iron levels, metabolism of amino acids and polyunsaturated fatty acids, glutathione (GSH), and biosynthesis of phospholipids (Balihodzic et al., 2022; Zhang C. et al., 2022). The role of this procedure is vital in various illnesses, such as neurologic conditions, malignancies, and ischemia-reperfusion damage (Wang Y. et al., 2020; Jiang et al., 2021). Recent evidence suggests that ferroptosis also plays a significant role in ACNSI (Lei et al., 2020). Following an ACNSI occurrence, there is a rise in intracellular iron levels, which results in heightened lipid peroxidation and subsequent initiation of ferroptosis (Xie et al., 2016). In addition to impacting the viability of neurons, ferroptosis also has an impact on the functionality of glial cells, vascular endothelial cells, and immune cells (Wang W. et al., 2019; Cui et al., 2021; Gao et al., 2022). Moreover, ferroptosis is closely associated with neurological functional deficits, neurodegenerative changes, neuroinflammatory responses, and blood-brain barrier (BBB) damage following ACNSI (Yu J. et al., 2020; Dong et al., 2022; Wang X. et al., 2022). Hence, it is crucial to investigate the mechanisms and functions of ferroptosis in ACNSI and create potent inhibitors or regulators of ferroptosis to prevent and treat ACNSI and its associated complications. The main objective of this article is to thoroughly examine the mechanisms and approaches for treating ferroptosis in ACNSI, specifically emphasizing the advancements in targeted mitochondrial therapy. The results of this investigation will offer valuable perspectives for subsequent studies and aid in the formulation of efficient treatment strategies for ACNSI.
ACNSI refers to sudden and unexpected damage to the brain or spinal cord, often caused by trauma, infection, disease, or the administration of certain drugs or chemicals (Slovinska et al., 2022). ACNSI is a frequent reason for mortality or enduring impairment (Shein and Shohami, 2011). ACNSI can be classified into primary and secondary injuries, depending on the mechanical stress situation at the site of injury (Jarrahi et al., 2020). The primary injuries occurs when tissues and blood vessels are directly injured, leading to the death of neurons and glial cells, damage to axons, and hemorrhagic or ischemic injuries due to problems with blood vessels (Ruhatiya et al., 2020). This includes brain contusion, intracranial hemorrhage, brain infarction, axon dysfunction, and other direct injuries caused by pathogens (Lutton et al., 2017; Li Z. et al., 2023). The secondary damage encompasses a range of intricate pathophysiological processes initiated by the initial injury, encompassing excitotoxicity in neurons, activation of neuroglial cells and neuroinflammation, disruption of the BBB, brain swelling, injury caused by ischemia-reperfusion, oxidative stress, and apoptosis (Wang D. et al., 2020; Zhang et al., 2023). Primary and secondary injury interact with each other and jointly influence the clinical outcome of ACNSI.
TBI is an intricate neurological condition caused by physical impact that results in harm to brain function or tissue (Lutton et al., 2017). In addition to 60 million people suffering TBI every year, about half of the world population has suffered one or more TBI over the course of their lifetime (Tanaka and Zhang, 2022). The intricacy of TBI becomes apparent through its various injury mechanisms, which include rapid acceleration/deceleration, direct impact, penetrating injury, and blast waves (Lutton et al., 2017). The outcome is the occurrence of physical harm to structures within the skull and direct harm to the brain tissue, which encompasses hematomas, contusions, diffused axon injury (DAI), and lacerations (Ellis et al., 2016). The primary cause of secondary brain injury is primarily due to heightened levels of excitatory neurotransmitters, generation of reactive oxygen species (ROS), malfunctioning mitochondria, and the release of proinflammatory cytokines (Glotfelty et al., 2019; Lorente et al., 2019; Stelmashook et al., 2019). These factors collectively contribute to the harm and potential demise of neuronal cells. After experiencing additional harm to the nerve tissue, there is a possibility of production of cerebral edema, increased intracranial pressure (ICP), impairment of BBB, and alterations in cerebrovascular reactivity (Lutton et al., 2017). Although there is a clear necessity for efficient and all-encompassing therapies to enhance results and minimize long-lasting impacts, there is currently no specialized and effective treatment for brains that have suffered traumatic injuries. Hence, it is imperative to prioritize the advancement of novel therapies for TBI.
Stroke is a cerebrovascular disease encompassing both hemorrhagic and ischemic strokes. This happens when the blood vessels in the brain are blocked or ruptured, resulting in a sudden decline in brain function (O’Reilly et al., 2014). Intracerebral hemorrhage (ICH) occurs in approximately 10–20% of cases, causing severe disability and, in some instances, death (Hanley et al., 2013). Brain injury following ICH involves primary injury, caused by direct compression and stimulation of the hematoma, and secondary injury, caused by ischemia and hypoxia in the penumbra surrounding the hematoma (Peng et al., 2017). On the contrary, cerebral infarction is a type of brain injury caused by the blockage of blood vessels in the brain. Hypoxia leads to irreversible necrotic death of neurons, glial cells, and vascular endothelial cells in the ischemic region, which is known as the primary injury (Zhou and Huang, 2019). The primary injury can also cause additional harm, such as excitatory neurotoxicity, oxidative stress, inflammation, and cell apoptosis, resulting in expansion of ischemic regions and neurological dysfunction (Dhungana et al., 2017). In the past, stroke treatment has typically included the utilization of oral interventions like tranexamic acid (TXA) and intravenous administrations of edaravone, which have produced disappointing outcomes (Sprigg et al., 2018; Uchida et al., 2022). To effectively treat stroke, it is necessary to protect the well-being of brain tissues and facilitate the healing process in the surrounding and infarcted areas.
In addition, the ACNSI encompasses infective meningitis where microbial pathogens directly invade the central nervous system (CNS) tissues, leading to inflammation and harm (Koelman et al., 2019). The CNS is at significant risk from bacterial meningitis, which could result in fatal outcomes. Annually, the three predominant types of bacteria, namely Meningococci, Haemophilus influenzae, and Streptococcus pneumoniae, contribute to an estimated 1,000,000 cases of meningitis (Chanteau et al., 2006). These bacteria are responsible for causing meningococcal meningitis. Furthermore, meningitis can also be caused by mold, yeast, and fungi that have the ability to change their shape (Davis et al., 2020). The opportunistic infection caused by fungal pathogens, particularly Cryptococcus neoformans, known as cryptococcal meningitis (CM), can result in fatality (Sridhar et al., 2021). Despite receiving appropriate antibiotic treatment, mortality remains a significant concern in cases of meningitis.
Different levels and degrees of neurological dysfunction that result from external or internal factors affecting the spinal cord are included in SCI. Direct force applied to the spinal cord causes the primary injury, resulting in the rupture of cell membranes, vascular harm, bleeding, localized lack of blood supply, and embolism (Zavodska et al., 2018). After the primary injury, a secondary injury takes place. The local area experiences the liberation and buildup of a substantial amount of catecholamine neurotransmitters, resulting in microvascular spasm, reduced blood flow, heightened permeability of blood vessels, rupture of small veins, and secondary hemorrhagic necrosis (Liu et al., 2021). SCI causes extensive neuronal demise and axonal disturbance, leading to a state where the functionality of the spinal cord is modified either temporarily or permanently (Chen et al., 2020). Due to the limited regenerative capabilities of the spinal cord, SCI frequently results in enduring impairment to the patient sensory and motor functions (Xi et al., 2020). As a result, SCI continues to be one of the foremost unresolved obstacles in the field of medicine. People who have SCI go through both psychological and physical distress, while medical professionals and patients face significant financial hardships.
Cell death via ferroptosis is distinct from other cell death types. The origin of its name is derived from the iron ions and the lipid peroxides that are linked to it (Ye et al., 2021). In ferroptosis, the cell lipid bilayers undergo oxidation and degradation, leading to the build-up of peroxides and oxidative stress within the cellular environment (Fujiki et al., 2019; Wang T.-X. et al., 2019; Wu Y. et al., 2020). This eventually leads to cell death. The presence of ferroptosis is closely connected to cellular elements like GSH metabolism, lipid metabolism, and iron metabolism. In particular, the build-up of lipid peroxides and the depletion of GPX4 play a crucial role in ferroptosis (Fujiki et al., 2019; Miao et al., 2022; Zhong et al., 2022). Research on ferroptosis offers fresh insights into the mechanisms of cellular demise and unveils innovative therapeutic pathways for delaying or thwarting the progression of diseases (Chen et al., 2022).
There are several mechanisms by which excess iron results to cellular damage. The Haber Weiss reaction can be catalyzed by iron, meaning it has the ability to produce hydroxyl radical (OH) by combining the superoxide anion and hydrogen peroxide, which can cause harm to lipids, nucleic acids, and proteins (Azad et al., 2008; Jabłońska-Trypuć et al., 2017; Lewandowska et al., 2019; Scott et al., 2021). Iron can also bind to transition metals, forming non-biological iron-sulfur clusters. These clusters can transfer electrons and increase ROS in the mitochondria (Chen et al., 2018; Nakamura et al., 2021). Iron can affect neurotransmitter metabolism and release, leading to excessive accumulation or depletion of neurotransmitters such as dopamine, norepinephrine, and serotonin that can interfere with neural signal transmission (Zhou and Tan, 2017; Kletetschka et al., 2021).
Ferroptosis and mitochondria are closely related. In the cell, mitochondria are the primary energy producers and are crucial for controlling cell death (Perez-Pinzon et al., 2012; Law et al., 2018). Mitochondrial dysfunction and damage resulting in ferroptosis, can be caused by the abnormal buildup of iron ions as observed in several studies (Dong et al., 2022). Ferroptosis displays unique morphological features, such as reduced mitochondria and increased density of mitochondrial membrane, leading to impaired mitochondrial function (Wang T.-X. et al., 2019). Iron ions can affect mitochondrial function in several ways, including inhibition of electron transport chain enzyme activity, increased ROS production, mitochondrial membrane peroxidation, and loss of inner membrane potential (Hu et al., 2016; Jiang et al., 2020; Jing et al., 2021). The inhibition of GPX4 can occur as a consequence of ferroptosis, either through direct or indirect means (Cao and Dixon, 2016). Consequently, this can result in harm to the intracellular antioxidant mechanism and lead to the buildup of ROS in mitochondria, ultimately causing cellular impairment (Dong et al., 2020). Due to malfunctioning mitochondria, the impairment of GSH, a GPX4 substrate is exacerbated, resulting in the accumulation of iron overload and ROS in membrane lipids (Wang et al., 2022b). Moreover, ferri-liposomes possess the capability to trigger apoptosis and ferroptosis through the liberation of citric acid iron oxide nanoparticles (IONPs-Ac) within the cellular cytosol. This action enhances mitochondrial dysfunction, ultimately resulting in the onset of apoptosis/ferroptosis (de Souza et al., 2021). The formation of mitochondrial dysfunction in CNS disease has been identified as triggered by ferroptosis, according to recent discoveries on cell death mechanisms (Krabbendam et al., 2020).
TBI has the ability to trigger a recently discovered type of cellular demise called ferroptosis, which is distinguished by significant buildup of iron and heightened oxidative stress in the impacted area (Cheng et al., 2023). Mitochondria, which are essential controllers of cellular destiny, have a vital function in the progression of both TBI and ferroptosis (Shen et al., 2019). Nonetheless, the exact function of mitochondria in ferroptosis is still a subject of debate. Certain academics have put forth proof indicating that malfunction in mitochondria may amplify the generation of ROS, consequently intensifying the incidence of ferroptosis (Shojaie et al., 2020). Furthermore, TBI is recognized to cause long-lasting cognitive and motor disabilities, and there is a lack of effective therapies for these individuals. Nevertheless, with the application of our existing knowledge about ferroptosis, we can develop innovative treatment approaches focused on addressing the crucial molecules and pathways involved in this mechanism. These strategies promise in reducing neuronal damage and enhancing functional recovery following TBI.
Neuronal membrane lipids are harmed by ferroptosis, which is a form of cellular demise caused by oxidative stress and lipid peroxidation. ROS initiates ferroptosis-mediated oxidative stress, which is the primary mechanism of ferroptosis and results in lipid peroxidation (Yang et al., 2022). Mitochondria are both major sources and targets of ROS, which play a vital part in starting and carrying out ferroptosis (Shojaie et al., 2020). Multiple pieces of evidence substantiate the role of mitochondria in ferroptosis. In mouse embryonic fibroblasts (MEF) and HT-22 hippocampal cells (HT-22) cells, the production of mitochondrial ROS is significantly enhanced by erastin and Ras selective lethal 3 (RSL3) (Xia et al., 2016; Fang et al., 2017; Rodríguez-Graciani et al., 2022). In addition, the reduction of ferroptosis can be achieved by using mitochondria-specific ROS scavengers like mitoquinone (MitoQ), XJB-5-131, and triphenylphosphonium chloride (Mito-TEMPO) (Park et al., 2017; Broome et al., 2018; Guigni et al., 2018). Following TBI, excessive glutamate release and impaired uptake result in excitotoxicity, leading to calcium overload and ROS generation in neurons (Shen et al., 2019). These factors trigger mitochondrial dysfunction, including the decline in membrane potential, the opening of the mitochondrial permeability transition pore, the liberation of cytochrome c (Cyt c), and the initiation of caspases (Wang P. et al., 2022). The after-effects of trauma lead to situations where the reduced usual number of mitochondria face higher metabolic requirements, leading to a significant increase in the production of ROS (Wang P. et al., 2022). Excitotoxicity, free iron, and interactions among ROS contribute to the excessive production of ROS (Lee et al., 2017; Liu et al., 2023). Mitochondrial damage exacerbates the ferroptosis process by inducing additional production of ROS and abnormal release of ROS (Taeubert et al., 2021). To elaborate, CNS injury causes mitochondrial damage, affecting the balance of mitochondrial dynamics (Chang et al., 2019). The disruption in mitochondrial dynamics results in decreased quantity, integrity, and performance of mitochondria, consequently hindering their capacity to produce energy and withstand oxidative stress (Di Pietro et al., 2017). Consequently, neurons become more susceptible to ferroptosis. Hence, focusing on the impairment of mitochondria presents a hopeful approach to hinder or counteract the consequences of TBI and ferroptosis.
The research carried out by Gao et al. (2019) demonstrated that the suppression of mitochondrial function effectively inhibited ferroptosis induced by cysteine deprivation. A substantial rise in mitochondrial metabolism occurred when cysteine levels were depleted, leading to an increased utilization of GSH. As a result, there was an ensuing increase in the generation of lipid ROS, ultimately resulting in ferroptosis (Gao et al., 2019). By increasing GSH levels, reducing oxidative stress, and regulating neurotransmitter systems, among other mechanisms, the inclusion of N-acetyl cysteine (NAC), a cysteine supplement, has the capacity to improve cognitive and behavioral functions after TBI (Huang et al., 2018). The decrease in cysteine concentrations after TBI may be due to its use in fighting against oxidative stress, inflammatory reaction, neuronal demise, and imbalances in neurotransmitters during pathological circumstances (Chen et al., 2018). Although active mitochondria may enhance ferroptosis, it has been observed that impaired mitochondrial function alone is not enough to effectively alleviate this process (Gao et al., 2019; Pandur et al., 2019). This may be caused by other processes that produce lipid ROS, which are quickly intensified by the Fenton reaction, ultimately leading to complete ferroptosis cell demise (Gao et al., 2019; Zhang et al., 2021). Nonetheless, the precise role of mitochondrial function in ACNSI necessitates further substantial evidence to substantiate its mechanisms.
The disparity between an overabundance of iron ions being consumed and released leads to an accumulation of iron within cells, ultimately leading to the production of extremely reactive free radicals OH via the Fenton reaction, thereby resulting in heightened oxidative harm. The activation of the Fenton reaction occurs when cells take in an excess of ferrous ions, resulting in lipid peroxidation (Jing et al., 2021). The presence of this mechanism plays a crucial role in triggering ferroptosis, thus emphasizing the important outcomes resulting from the excessive generation of lipid peroxides and the initiation of free radicals (Jing et al., 2021). Following TBI, axons in the brain white matter become more vulnerable to damage (Han et al., 2015). The main component of DAI is the physical breakage of the axonal cytoskeleton, which disrupts the transportation of axons and leads to swelling and breakdown of proteins (Lazarus et al., 2015). Secondary trauma pertains to subsequent molecular and chemical inflammatory responses, like the liberation of hemoglobin and iron, impaired brain metabolism, and cerebral blood flow (Wei et al., 2012). These reactions further trigger neuroinflammatory processes, oxidative stress, glutamatergic excitotoxicity, and mitochondrial malfunction, ultimately resulting in further brain damage (Lipponen et al., 2019; Liao et al., 2020). The BBB is compromised by primary and secondary injuries caused by TBI, enabling iron to enter brain tissue and create deposits of iron (Yauger et al., 2019). The presence of iron initiates the Fenton reaction, resulting in the production of extremely reactive OH free radicals, which in turn leads to the occurrence of oxidative stress and lipid peroxidation (Zhu et al., 2021). These processes represent key features of ferroptosis. Moreover, TBI also disrupts the balance of iron uptake, storage, and efflux, resulting in intracellular iron overload and dysfunction (Cheng et al., 2023). It is well-established that ROS, including superoxide anion free radicals and OH, can contribute to brain injury in TBI patients (Shi et al., 2018). The overabundance of iron and malfunction increase the activation of heme oxygenase-1 (HO-1), subsequently enhancing the onset of ferroptosis (Chen et al., 2023). Furthermore, potential elements that may contribute to brain iron metabolism disorder could include dysfunction of mitochondria and lysosomes. Under pathological conditions of TBI, the MCU protein situated on the inner membrane of mitochondria has the ability to convey significant quantities of calcium ions into the mitochondria, causing disruption to their regular operation (Zhang L. et al., 2019). The rapid transportation of calcium ions into mitochondria by MCU is accompanied by the simultaneous transportation of significant amounts of iron into the mitochondrial matrix, which could potentially result in a disorder of iron homeostasis (Zhang L. et al., 2019). The overabundance of iron triggers the generation of ROS through the Fenton reaction, worsening the impairment of mitochondria and creating an ongoing destructive cycle that intensifies iron-triggered cellular demise (Jing et al., 2021). Regulating iron metabolism, scavenging free radicals, and safeguarding the normal functioning of mitochondria are potential strategies to prevent iron-induced cell death following TBI.
By altering the ferroptosis pathway, TBI could be reduced in the following ways (Table 1).
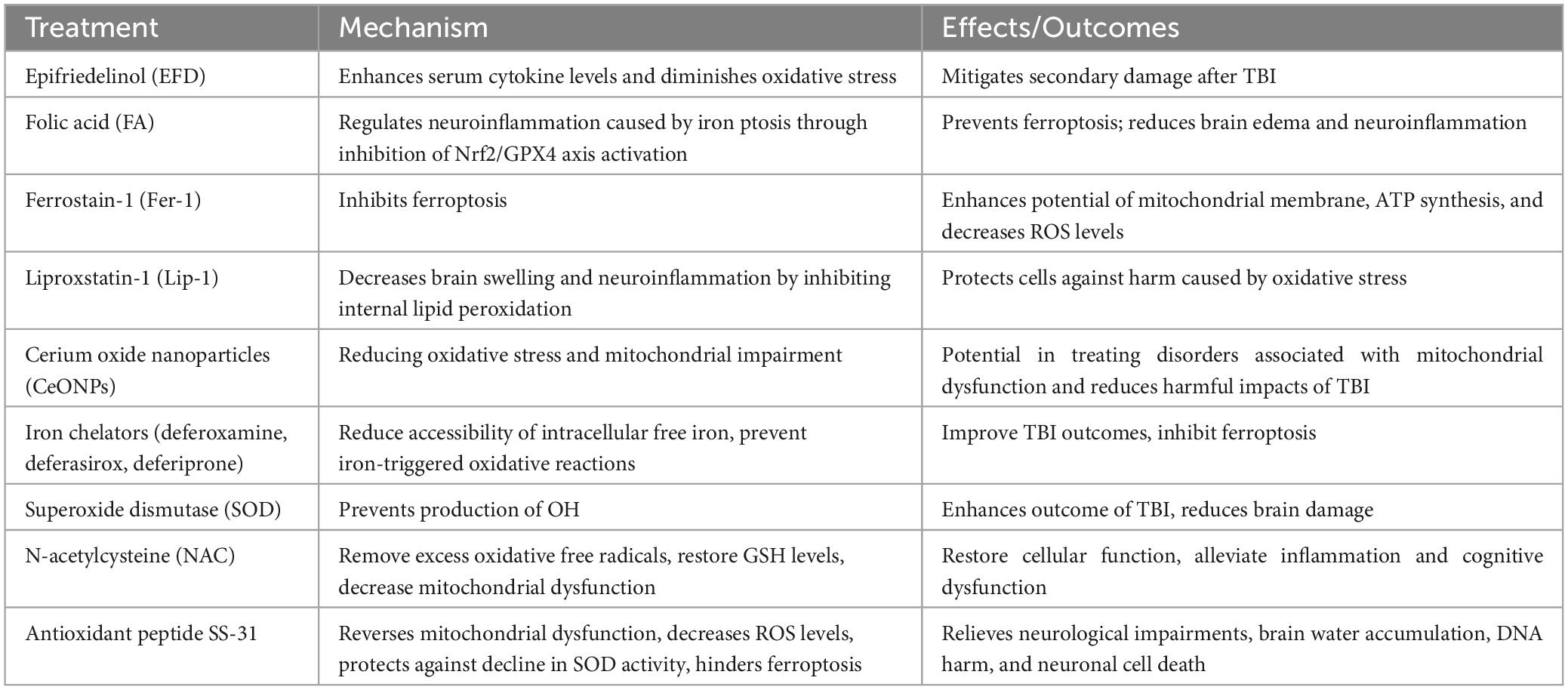
Table 1. Therapeutic strategies targeting ferroptosis in TBI.
Reducing oxidative stress is regarded as a crucial goal in the management of TBI. Epifriedelinol (EFD) has demonstrated its ability to mitigate secondary damage in rats with TBI by enhancing serum cytokine levels and diminishing oxidative stress (Li et al., 2018). The Nrf2 gene expression, accountable for the response to oxidative stress, has been discovered as a potential mechanism for averting ferroptosis. Research has demonstrated that the use of folic acid (FA) therapy can regulate neuroinflammation caused by iron ptosis through the inhibition of Nrf2/GPX4 axis activation (Wang C. et al., 2022). Noteworthy, Wu et al. (2018) Discovered that Ferrostain-1 (Fer-1), a substance that inhibits ferroptosis, has the ability to successfully restore mitochondrial malfunction. Evidence suggests that Fer-1 enhances the potential of the mitochondrial membrane and the synthesis of Adenosine Triphosphate (ATP), while also decreasing ROS levels (Wu et al., 2018). This has also been actively used in subarachnoid hemorrhage (SAH) to protect against brain edema and neuroinflammation (Gao et al., 2022). Liproxstatin-1 (lip-1), an additional iron inhibitor, is employed to decrease brain swelling and neuroinflammation. It hinders internal lipid peroxidation and safeguards cells against harm caused by oxidative stress (Gao et al., 2022). Furthermore, considering the link between increased oxidative stress and mitochondrial impairment as underlying causes for the neurological outcomes of TBI, the utilization of cerium oxide nanoparticles (CeONPs) could potentially be effective in reducing the harmful impacts of TBI (Bailey et al., 2020). These discoveries have important implications in the treatment of disorders associated with mitochondrial dysfunction.
Inhibiting the presence of unbound iron ions and free radicals is a feasible strategy to impede ferroptosis in TBI. The use of iron chelation treatment has shown promise in improving the negative consequences of TBI. Iron chelators, such as deferoxamine (DFO), deferasirox (DFX) and deferiprone (DFP), effectively limit the accessibility of intracellular free iron, thereby reducing the likelihood of iron-triggered oxidative reactions (Votavova et al., 2021; Zhou et al., 2022). Recent investigations indicate that DFO treatment also inhibits the emergence of free radicals and prevents ferroptosis (Zhou et al., 2022). Besides the use of iron chelation, superoxide dismutase (SOD) has been recognized as a hopeful strategy to enhance the outcome of TBI (Kobeissy et al., 2016). The function of SOD is to prevent the production of OH, thereby reducing brain damage (Lv et al., 2016). To decrease the occurrence of ferroptosis, cells make use of antioxidants like GSH and NAC to remove an excess of oxidative free radicals. Studies have shown that the sole administration of NAC systemically can restore GSH levels and decrease mitochondrial dysfunction in rats with TBI (Gomez et al., 2013; Du et al., 2016). On the other hand, when NAC is combined with minocycline, it can alleviate inflammation and cognitive dysfunction in TBI rats (Margulies et al., 2016). Several research studies have indicated a favorable pattern in the utilization of antioxidant treatment, and traditional antioxidants can function as supplementary therapy for TBI. Nevertheless, these traditional antioxidants lack specificity in their actions, frequently requiring high levels to attain desired therapeutic outcomes. Effective mitochondrial targeting requires the crucial delivery of antioxidants specifically to the mitochondria. Administration of the antioxidant peptide SS-31, 30 min after mild TBI, significantly reverses the mitochondrial dysfunction caused by TBI and improves secondary brain injury (Shen et al., 2019). SS-31 directly decreases the level of ROS in brain tissue, protects against the decline in SOD activity, and hinders ferroptosis (Righi et al., 2013). As a result, it relieves neurological impairments, brain water accumulation, DNA harm, and neuronal cell death (Shen et al., 2019). Furthermore, the peroxisome proliferator-activated receptor γ (PPARγ) signaling pathway is crucial for cellular function. Enhancing the cell antioxidant defense mechanism is one of the ways it helps to facilitate the expression and activity of GPX4 in cells (Wang M.-J. et al., 2023). Through ongoing research and innovative treatment techniques like these, healthcare professionals can aspire to mitigate the ferroptosis caused by TBI and improve patient outcomes.
Stroke is a neurological disorder that mainly happens when brain cells are harmed and die because of a lack of oxygen and inadequate blood flow in the affected region. This occurs due to the blockage of arteries in the brain. There are two types of stroke: ischemic and hemorrhagic. Globally, it ranks second in terms of mortality and is the main reason for fatalities in China, which carries the highest burden of stroke cases worldwide (Mei et al., 2020).
In instances of hemorrhagic stroke and advanced cerebral ischemia, Ferroptosis has been identified. The presence of iron-rich blood and ferritin-rich brain tissue is a result of the compromised BBB and the primary brain injury caused by direct compression and stimulation of the hematoma (Liu et al., 2019). Following ICH, it takes a few weeks for brain heme iron levels to significantly increase, while levels of transferrin (TF), transferrin receptor (TFR), and ferritin (FT) in the brain take a few days to rise (Wu et al., 2003). The disproportionate rise in proteins responsible for iron handling and excessive accumulation of iron can result in harmful reactions that cause damage to the brain (Park et al., 2015; Gill et al., 2018). The release of iron from the hematoma following ICH could potentially lead to oxidative stress, perihematomal edema, and an elevation in ROS levels, ultimately leading to ferroptosis (Zhang H.-Y. et al., 2022).
Following the onset of cerebral hemorrhage, the surrounding region of the hematoma experiences secondary brain damage due to ischemia and lack of oxygen. Likewise, cerebral infarction leads to primary damage and triggers additional mechanisms of injury, including excitotoxicity, oxidative stress, inflammation, and apoptosis (Dhungana et al., 2017). These mechanisms play a role in the enlargement of the ischemic region and subsequent impairment of neurological function. Children with hypoxic-ischemic encephalopathy often exhibit iron deposition in the basal ganglia, thalamus, and periventricular white matter (Dietrich and Bradley, 1988). The presence of iron in the brain tissue could be due to the breakdown of the BBB, which allows iron from the blood to enter the brain. The function of iron transport and storage proteins is impacted by hypoxia, which leads to an increased susceptibility of neurons to iron (Gill et al., 2018). The Fenton reaction is responsible for the aggravation of lipid peroxidation-induced ferroptosis due to an excess of iron, leading to increased infarct sizes (Castellanos et al., 2002; Jing et al., 2021). Furthermore, in a mouse model of ischemic stroke, it was observed that free radicals and an excess of iron resulted in the prolonged upregulation of transferrin receptor 1 (TFR1), which subsequently led to an elevation in peripheral iron absorption, oxidative stress, and the demise of neurons (Yu Y. et al., 2020). Moreover, diminished GSH levels, which are evident in both individuals with stroke and animal models of middle cerebral artery occlusion (MCAO), additionally amplify ferroptosis (Liu et al., 2020). Cerebral ischemia-reperfusion injury (CIRI) exhibits an imbalance in lipid and amino acid metabolism. For example, the levels of malondialdehyde (MDA) and nitric oxide (NO) are increased, whereas the levels of SOD and GPX4 are decreased in mouse models of CIRI and oxygen-glucose deprivation/reoxygenation (OGD/R) cell models (Zeng et al., 2019). Furthermore, an imbalance in the metabolism of amino acids contributes to the promotion of ferroptosis. In rats with middle MCAO, the levels of extracellular glutamate significantly increased. This led to the acceleration of neuronal iron absorption and subsequent cell death due to excitatory toxicity (Shu et al., 2021). This could be attributed to the regulation of glutaminase 2 by the p53 gene, which triggers ferroptosis when glutaminase increased (Jennis et al., 2016). Furthermore, mitochondria have a vital function in upholding cellular balance and operation, and their impairment is linked to the development of heart and brain disorders (Horvath et al., 2008). In individuals suffering from acute ischemic stroke, plasma analysis indicated an increase in the expression of lncRNA PVT1 and a decrease in the expression of miR-214 (Lu et al., 2020). In CIRI mice, the infarct area was reduced and ferroptosis was suppressed by either silencing PVT1 or increasing the expression of miR-214 (Lu et al., 2020).
The phenomenon of ferroptosis has been observed to intensify the severity of stroke by inducing oxidative stress and lipid peroxidation in the brain (Table 2), similar to the mechanisms underlying TBI. GSH levels and GPX4 levels have been strongly associated with stroke. Following a stroke, the levels of GSH and GPX4 expression decrease, impeding the prompt elimination of the oxidative stress reaction and worsening ferroptosis (Alim et al., 2019). By activating the transcription factors TFAP2c and Sp1, Selenium has the potential to enhance GPX4 expression, offering a possible therapeutic strategy (Alim et al., 2019). Studies have demonstrated that drugs like metformin can safeguard the brain by inhibiting ferroptosis, either by decreasing the production of lipid peroxidation or by elevating GPX4 levels, ultimately reducing hemorrhagic or ischemic brain damage in mice (Abd-Elsameea et al., 2014; Ismail Hassan et al., 2020). This process helps prevent ferroptosis and glutamic acid (Glu) excitotoxicity, which in turn reduces hemorrhagic or ischemic brain damage in mice (Alim et al., 2019). The quantity or activity of LOX-5 has been observed to rise in both ischemic and hemorrhagic strokes, potentially initiating lipid peroxidation and ultimately causing neuronal ferroptosis (Yigitkanli et al., 2017; Karuppagounder et al., 2018). A different research study indicated that the antioxidative characteristics of NAC have the ability to neutralize the harmful effects caused by nuclear LOX-5 and hinder neuronal ferroptosis (Karuppagounder et al., 2018). Extracted from safflower, carthamin yellow, a compound of flavonoid nature, has been found to hinder the buildup of iron and ROS in the rat brain suffering from CIRI. Additionally, it can restore the protein expression levels of ACSL4, TFR1, GPX4, and ferritin heavy chain 1 (Guo et al., 2021). Furthermore, inhibition of Tau was found to protect young animals from ferroptosis during ischemia-reperfusion following ischemic stroke, subsequently suppressing neural damage (Tuo et al., 2017). It is crucial to address oxidative stress and lipid peroxides in individuals who have experienced a stroke in order to avoid additional harm to cells.
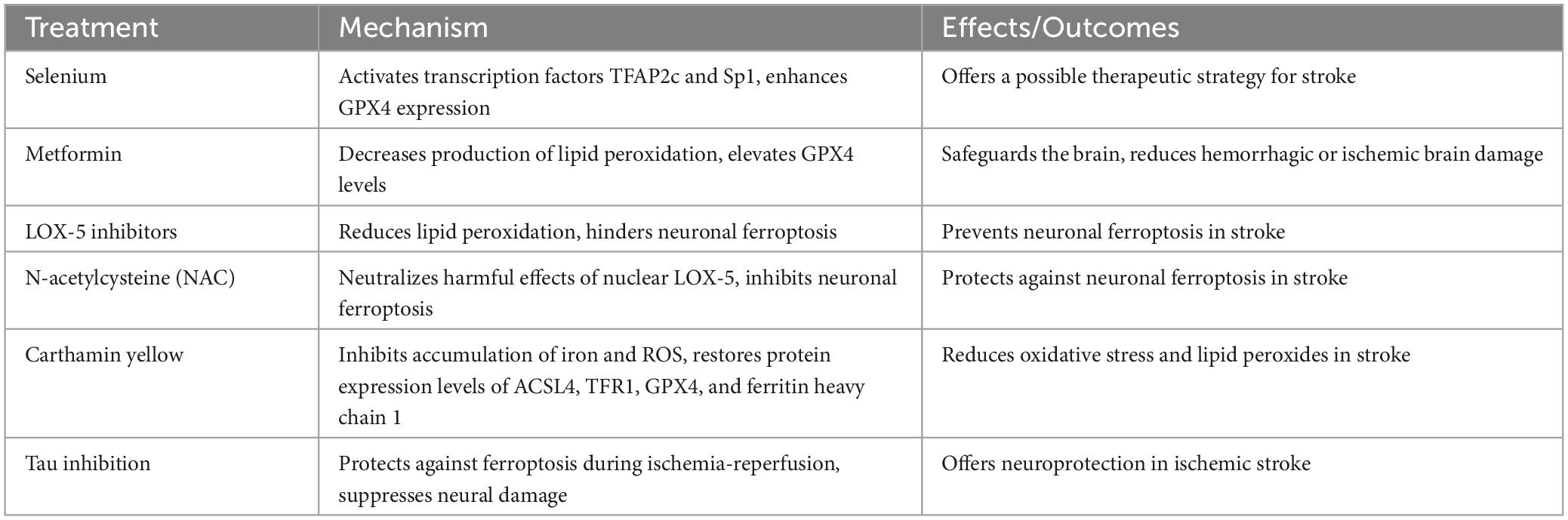
Table 2. Therapeutic strategies targeting ferroptosis in stroke.
Neisseria meningitidis is a prominent etiological agent of bacterial meningitis, as it colonizes the human pharynx and leads to severe diseases such as meningitis (Harrison, 2006). Moreover, in Africa, CM is the primary cause of adult meningitis (Kwizera et al., 2021). CM is a disease caused by the fungus Cryptococcus, which is usually more common in people with weakened immune systems (Kwizera et al., 2021). These pathogens can infect the patient body through inhalation, causing manifestations such as pneumonia. In some cases, Cryptococcus can circulate through the bloodstream and reach the brain, causing meningitis (Kwizera et al., 2021; Retchless et al., 2022). The disease presents with a range of symptoms, including headache, fever, nausea and vomiting, and stiff neck, among others (Haidrani, 2016; Bårnes et al., 2018; Lin and Su, 2020). If left untreated, the disease can pose a significant risk to the patient life.
Recent studies have increasingly demonstrated a correlation between meningitis and ferroptosis, yet the precise mechanisms that underlie this association remain unclear. It is hypothesized that ferroptosis may impact the progression of meningitis by inducing iron accumulation and alterations in lipid peroxidation. Individuals with CM have elevated ferritin levels in their cerebrospinal fluid (CSF) (Deisenhammer et al., 1997). Furthermore, the increase in ferritin levels can be considered a sign of meningitis (Campbell et al., 1986). Another study has indicated that iron overload exacerbates experimental CM (Barluzzi et al., 2002). The process of ferritinophagy, which involves the release of iron into the labile iron pool (LIP) through ferritin, has been found to increase ferroptosis sensitivity (Xu et al., 2021). As demonstrated in multiple studies, both the vasculature and brain are susceptible to C. neoformans-induced lipid peroxidation. Notably, Hall et al. observed a significant increase in cellular lipid peroxidation levels in rabbits inoculated with C. neoformans (Hall et al., 2010).
An extremely devastating traumatic injury is SCI, characterized by massive neuronal death and axonal disruption (Wang R. et al., 2023). After suffering from SCI, the individual might encounter lasting limitations in both their motor and sensory abilities due to the spinal cord restricted ability to regenerate (Xi et al., 2020). Treating SCI efficiently continues to be one of the most significant medical obstacles to this day (Simard et al., 2010).
Iron overload and ROS accumulation result in SCI-induced downward translocation of motor neuron iron and impaired recovery. By quantifying iron deposition, demyelination, and atrophy over 2 years after SCI, researchers found that cord atrophy and cerebellar loss decreased, and while brain white and gray matter atrophy was sustained, the myelin content in the spinal cord and cortex reduced progressively. As the sustained atrophy in the thalamus progressed, iron deposition was significant. Feng et al. showed that motor cortex microglia respond to iron overload by accumulating ROS, resulting in ferroptosis of motor neurons and thereby hindering recovery following SCI (Feng et al., 2021). The augmented accumulation of iron was confirmed in both rats and individuals with SCI. Furthermore, the use of iron chelators, inhibitors of ROS, and inhibitors of ferroptosis was found to decrease the death of motor neurons and promote the restoration of function. Furthermore, activated microglia were discovered to release NO, which regulates the balance of cellular iron and causes an excessive accumulation of iron in motor neurons (Feng et al., 2021). In a separate investigation, scientists employed a modified Allen method to create a rat model for DFO treatment at the thoracic T10 section, and compared it to a mouse model for SCI. Examination with electron microscopy revealed diminished mitochondria in the SCI cohort. Unlike the SCI group, the DFO group showed significantly better Basso, Beattie, and Bresnahan locomotor scores, lower iron levels, and higher levels of GPX4, xCT, and GSH expression. In addition, the DFO group exhibited increased expression of mRNA for genes associated with ferroptosis, specifically Acyl-CoA synthetase family member 2 (ACSF2) and iron-responsive element-binding protein 2 (IREB2). Additionally, DFO was observed to promote neuronal survival and suppress gliosis (Yao et al., 2019). A distinct investigation discovered that there was an increase in the manifestation of the ferroptosis indicator ACSL4 and the oxidative stress indicator MDA in the subset of individuals who had experienced SCI within 24 h of the incident. Moreover, there was an observed decline in GSH levels throughout this timeframe. The results, when combined with bioinformatics analysis, suggested that the initial day after SCI played a vital role in the advancement of ferroptosis. Through bioinformatics analysis, the study additionally discovered 10 crucial hub genes associated with ferroptosis, specifically STAT3, JUN, TLR4, ATF3, HMOX1, MAPK1, MAPK9, PTGS2, VEGFA, and RELA. Following injury, the SCI group displayed increased levels of STAT3, JUN, TLR4, ATF3, HMOX1, PTGS2, and RELA mRNA, while VEGFA, MAPK1, and MAPK9 mRNA levels were found to be decreased, as confirmed by real-time PCR analysis, in comparison to the sham group, 1 day post-injury. This pattern was not observed in the sham group (Li et al., 2023). DHODH expression and enzymatic activity have been implicated in cancer progression in previous studies. Currently known inhibitors such as leflunomide metabolites and teriflunomide are effective therapeutic agents for the treatment of autoimmune diseases such as rheumatoid arthritis (RA) (Li et al., 2023). DHODH is targeted by immunosuppressive agents and anticancer drugs, which have shown potential in protecting the CNS, promoting neuronal survival, and facilitating axon regeneration (Li et al., 2023). DHODH, located on the outer surface of the inner mitochondrial membrane, plays a crucial role in detoxifying lipid peroxyl radicals in mitochondria by catalyzing the oxidation of dihydroorotate and reducing coenzyme Q (CoQ) to CoQH2 (Lyamzaev et al., 2023). Recent studies have also identified a similar antiferroptotic mechanism involving CoQ reduction by mitochondrial glycerol-3-phosphate dehydrogenase 2 (GPD2) (Lyamzaev et al., 2023). In a study conducted by Li et al. (2023) molecular evidence was provided indicating that DHODH acts as an inhibitor, preventing the activation of molecules associated with ferroptosis. This inhibition leads to a decrease in the production of lipid peroxides and mitochondrial damage, which ultimately results in the reduction of neuronal ferroptosis (Li et al., 2023). While the exact role of DHODH in the other CNS remains unclear, exploring its association with neurodegenerative diseases and iron-dependent cell death in ACNSI provides new possibilities for therapeutic interventions. These opens up a new idea for the relationship between DHODH and iron death in ACNSI.
The therapeutic strategies aimed at reversing or delaying neurological damage following SCI by countering ferroptosis have been achieved through the targeting of the ferroptosis pathway (Table 3). One approach includes utilizing DFO, a compound that binds to iron, leading to an upregulation of GPX4, systemic Xc–, and GSH levels. Consequently, this leads to a decrease in excessive iron accumulation and neuronal ferroptosis within the motor cortex following SCI, thus aiding in the restoration of motor function (Zhang et al., 2020). Another method is to utilize the ferroptosis inhibitor Fer-1, which can improve SCI by reducing iron and ROS deposition, thereby inhibiting ferroptosis in oligodendrocytes (Wang F. et al., 2022). Furthermore, an innovative compound named SRS 16-86 has shown promising outcomes in SCI models by enhancing GPX4, GSH, and xCT levels, as well as diminishing lipid oxidation, gliosis, and neuronal harm (Zhang Y. et al., 2019). Proanthocyanidins, which are potent free radical scavengers, have also shown promise in treating nerve damage in SCI. They efficiently decrease the amounts of iron and substances that react with thiobarbituric acid, inhibit the expression of ALOX15 and ACSL4, and enhance the levels of HO-1, NRF2, GSH, and GPX4 (Zhou et al., 2020). Moreover, zinc gluconate has shown the capacity to reduce ferroptosis in SCI by enhancing NRF2/HO-1 signaling. As a consequence, there is a rise in GSH, SOD, and GPX4 levels, accompanied by a decrease in ROS and lipid peroxides. Zinc gluconate also promotes the healing of injured mitochondria and inflammation, making it a potential future option for promoting both behavioral and structural recovery after SCI (Li et al., 2019). Notably, a study investigating the DHODH inhibitor teriflunomide in a SCI model revealed significant neuroprotective effects, including improved motor function recovery, reduced inflammation, increased neuronal survival, and inhibition of apoptosis, autophagy, mitochondrial dysfunction, and oxidative stress (Li et al., 2023). These evidence that terfenamine may protect Nervous tissue after SCI through various ways.
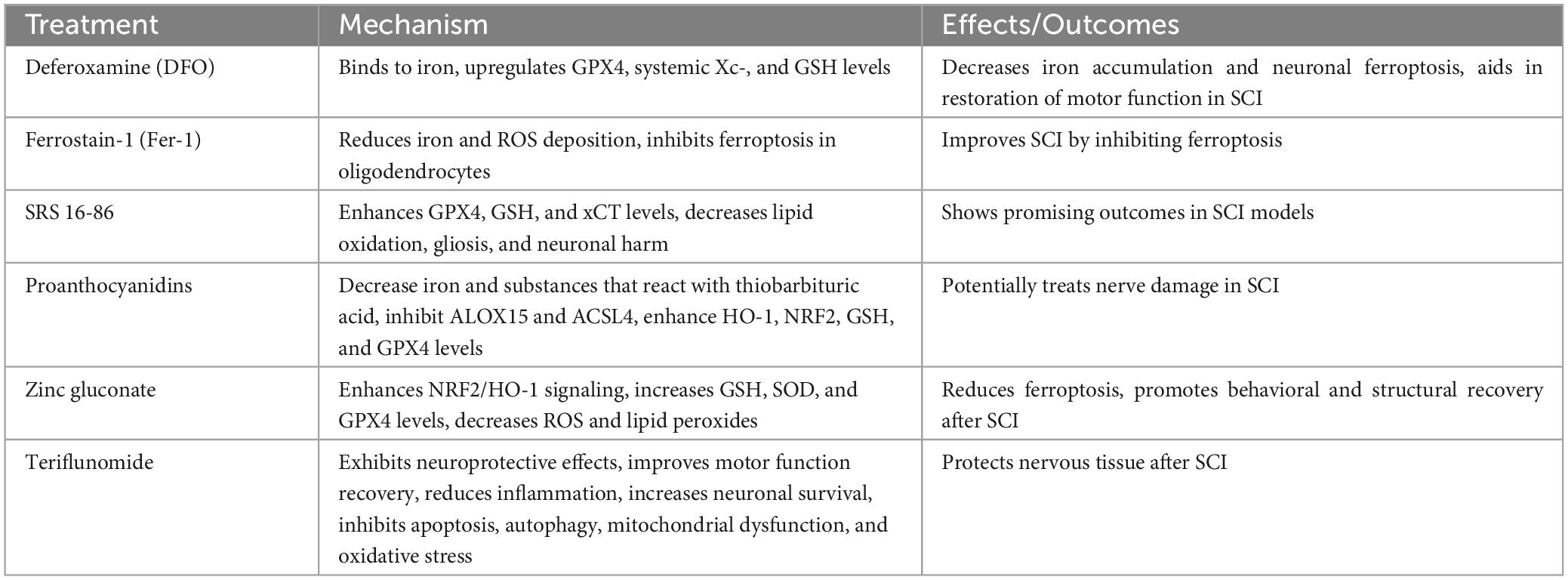
Table 3. Therapeutic strategies targeting ferroptosis in SCI.
Numerous studies have shown the significance of ferroptosis in ACNSI, specifically in relation to secondary injury following ACNSI. In this process, the significant role of mitochondria has been discovered and confirmed by researchers, despite the existence of controversial mechanisms (Wang et al., 2022a). Numerous researches have verified that malfunctioning mitochondria can trigger oxidative stress and additional elements that contribute to ferroptosis and neuronal cell death (Bécquer-Viart et al., 2021). Scientific evidence demonstrates that mitigating ferroptosis has the potential to postpone or even reverse neural damage. Different methods can be employed to accomplish this, including the use of iron chelators, lipid antioxidants, boosting GPX4 activity or stability, elevating GSH levels, or suppressing Ferroptosis Suppressor Protein 1 (FSP1) activity (Singh et al., 2014; Hayano et al., 2016; Serra et al., 2020; Wu L. et al., 2020; Wang F. et al., 2022). Conventional antioxidants typically lack selectivity and require high concentrations for therapeutic effects. Therefore, it is essential to direct antioxidants specifically toward mitochondria in order to achieve a targeted effect on mitochondria. Mitochondrial-targeted therapy involves protective strategies for CNS cells against ferroptosis by improving or replacing damaged mitochondria. One example is DHODH inhibitor, called teriflunomide targets mitochondrial ferroptosis (Malla et al., 2020). Research has also been conducted on nanomaterials for precise delivery of drugs to mitochondria, although there is still a need for accurate regulation of drug release amount and location. The regulation of the dynamics of mitochondria, which involves encouraging the splitting and merging of mitochondria, as well as the removal of damaged mitochondria through mitophagy, holds potential in counteracting ferroptosis. Nevertheless, the regulation of mitochondrial dynamics necessitates the consideration of various elements, including but not limited to timing, dosage, and duration. Moreover, there have been reports on the potential use of mitochondrial transplantation as a remedy for different ailments, providing a chance to combat ferroptosis and reverse neural damage. Nevertheless, additional investigation is necessary to explore the molecular signaling pathways, regulatory elements, and factors that exert influence. The potential for treating ACNSI by targeting mitochondria to counteract ferroptosis is significant. However, further fundamental investigations and clinical experiments are required to confirm effectiveness and security, as well as ascertain the most suitable treatment plan and indications.
Undoubtedly, the process of ferroptosis in instances of ACNSI is a highly debated and promisingly investigated subject. Suppressing ferroptosis offers a promising avenue for treating the prognosis of ACNSI in the future. However, the implementation of these ideas is currently limited to experimental settings, although it is expected that they will eventually be expanded to a broader range of individuals.
RY and HB designed and coordinated the study. WD and YZ collected the references in related fields and wrote the first draft of the manuscript. FG and RY revised the manuscript. All authors reviewed and approved the final manuscript.
This work was supported by the Science and Technology Projects of Guangzhou (202102021271).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abd-Elsameea, A. A., Moustaf, A. A., and Mohamed, A. M. (2014). Modulation of the oxidative stress by metformin in the cerebrum of rats exposed to global cerebral ischemia and ischemia/reperfusion. Eur. Rev. Med. Pharmacol. Sci. 18, 2387–2392.
Alim, I., Caulfield, J. T., Chen, Y., Swarup, V., Geschwind, D. H., Ivanova, E., et al. (2019). Selenium drives a transcriptional adaptive program to block ferroptosis and treat stroke. Cell 177, 1262–1279.e25. doi: 10.1016/j.cell.2019.03.032
Azad, N., Iyer, A. K. V., Manosroi, A., Wang, L., and Rojanasakul, Y. (2008). Superoxide-mediated proteasomal degradation of Bcl-2 determines cell susceptibility to Cr(VI)-induced apoptosis. Carcinogenesis 29, 1538–1545. doi: 10.1093/carcin/bgn137
Bai, X., and Bian, Z. (2022). MicroRNA-21 is a versatile regulator and potential treatment target in central nervous system disorders. Front. Mol. Neurosci. 15:842288. doi: 10.3389/fnmol.2022.842288
Bailey, Z. S., Nilson, E., Bates, J. A., Oyalowo, A., Hockey, K. S., Sajja, V. S. S. S., et al. (2020). Cerium oxide nanoparticles improve outcome after in vitro and in vivo mild traumatic brain injury. J. Neurotrauma 37, 1452–1462. doi: 10.1089/neu.2016.4644
Balihodzic, A., Prinz, F., Dengler, M. A., Calin, G. A., Jost, P. J., and Pichler, M. (2022). Non-coding RNAs and ferroptosis: Potential implications for cancer therapy. Cell Death Differ. 29, 1094–1106. doi: 10.1038/s41418-022-00998-x
Barluzzi, R., Saleppico, S., Nocentini, A., Boelaert, J. R., Neglia, R., Bistoni, F., et al. (2002). Iron overload exacerbates experimental meningoencephalitis by Cryptococcus neoformans. J. Neuroimmunol. 132, 140–146. doi: 10.1016/s0165-5728(02)00324-7
Bårnes, G. K., Gudina, E. K., Berhane, M., Abdissa, A., Tesfaw, G., Abebe, G., et al. (2018). New molecular tools for meningitis diagnostics in Ethiopia – A necessary step towards improving antimicrobial prescription. BMC Infect. Dis. 18:684. doi: 10.1186/s12879-018-3589-4
Bécquer-Viart, M. Á, Armentero-López, A., Alvarez-Almiñaque, D., Fernández-Acosta, R., Matos-Peralta, Y., D’Vries, R. F., et al. (2021). Gossypitrin, a naturally occurring flavonoid, attenuates iron-induced neuronal and mitochondrial damage. Molecules 26:3364. doi: 10.3390/molecules26113364
Broome, S. C., Woodhead, J. S. T., and Merry, T. L. (2018). Mitochondria-targeted antioxidants and skeletal muscle function. Antioxidants 7:107. doi: 10.3390/antiox7080107
Campbell, D. R., Skikne, B. S., and Cook, J. D. (1986). Cerebrospinal fluid ferritin levels in screening for meningism. Arch. Neurol. 43, 1257–1260. doi: 10.1001/archneur.1986.00520120041014
Cao, J. Y., and Dixon, S. J. (2016). Mechanisms of ferroptosis. Cell Mol. Life Sci. 73, 2195–2209. doi: 10.1007/s00018-016-2194-1
Castellanos, M., Puig, N., Carbonell, T., Castillo, J., Martinez, J., Rama, R., et al. (2002). Iron intake increases infarct volume after permanent middle cerebral artery occlusion in rats. Brain Res. 952, 1–6. doi: 10.1016/s0006-8993(02)03179-7
Chang, C.-Y., Liang, M.-Z., and Chen, L. (2019). Current progress of mitochondrial transplantation that promotes neuronal regeneration. Transl. Neurodegener. 8:17. doi: 10.1186/s40035-019-0158-8
Chanteau, S., Dartevelle, S., Mahamane, A. E., Djibo, S., Boisier, P., and Nato, F. (2006). New rapid diagnostic tests for Neisseria meningitidis serogroups A. W135, C, and Y. PLoS Med. 3, e337. doi: 10.1371/journal.pmed.0030337
Chen, H., Li, Z., Xu, J., Zhang, N., Chen, J., Wang, G., et al. (2023). Curcumin induces ferroptosis in follicular thyroid cancer by upregulating HO-1 expression. Oxid. Med. Cell Longev. 2023:6896790. doi: 10.1155/2023/6896790
Chen, H.-C., Tang, H.-H., Hsu, W.-H., Wu, S.-Y., Cheng, W.-H., Wang, B.-Y., et al. (2022). Vulnerability of triple-negative breast cancer to saponin formosanin C-Induced ferroptosis. Antioxidants 11:298. doi: 10.3390/antiox11020298
Chen, X., Pan, Z., Fang, Z., Lin, W., Wu, S., Yang, F., et al. (2018). Omega-3 polyunsaturated fatty acid attenuates traumatic brain injury-induced neuronal apoptosis by inducing autophagy through the upregulation of SIRT1-mediated deacetylation of Beclin-1. J. Neuroinflammation 15:310. doi: 10.1186/s12974-018-1345-8
Chen, Y., Wang, D., Cao, S., Hou, G., Ma, H., and Shi, B. (2020). Association between Serum IL-37 and spinal cord injury: A prospective observational study. Biomed. Res. Int. 2020:6664313. doi: 10.1155/2020/6664313
Cheng, H., Wang, P., Wang, N., Dong, W., Chen, Z., Wu, M., et al. (2023). Neuroprotection of NRF2 against ferroptosis after traumatic brain injury in mice. Antioxidants 12:731. doi: 10.3390/antiox12030731
Cui, Y., Zhang, Z., Zhou, X., Zhao, Z., Zhao, R., Xu, X., et al. (2021). Microglia and macrophage exhibit attenuated inflammatory response and ferroptosis resistance after RSL3 stimulation via increasing Nrf2 expression. J. Neuroinflammation 18:249. doi: 10.1186/s12974-021-02231-x
Davis, C., Wheat, L. J., Myint, T., Boulware, D. R., and Bahr, N. C. (2020). Efficacy of cerebrospinal fluid beta-d-glucan diagnostic testing for fungal meningitis: A systematic review. J. Clin. Microbiol. 58, e2094–e2019. doi: 10.1128/JCM.02094-19
de Souza, M. G. F., de Jesus Guedes, F. N., Tebaldi, M. L., do Nascimento Alencar, É, Amaral-Machado, L., do Egito, E. S. T., et al. (2021). Ferri-Liposomes: Preformulation and selective cytotoxicity against A549 lung cancer cells. Pharmaceutics 13:712. doi: 10.3390/pharmaceutics13050712
Deisenhammer, F., Miller, R. F., Brink, N. S., Harrison, M. J., and Thompson, E. J. (1997). Cerebrospinal fluid ferritin in HIV infected patients with acute neurological episodes. Genitourin. Med. 73, 181–183. doi: 10.1136/sti.73.3.181
Dhungana, H., Huuskonen, M. T., Jaronen, M., Lemarchant, S., Ali, H., Keksa-Goldsteine, V., et al. (2017). Sulfosuccinimidyl oleate sodium is neuroprotective and alleviates stroke-induced neuroinflammation. J. Neuroinflammation 14:237. doi: 10.1186/s12974-017-1010-7
Di Pietro, V., Lazzarino, G., Amorini, A. M., Signoretti, S., Hill, L. J., Porto, E., et al. (2017). Fusion or fission: The destiny of mitochondria in traumatic brain injury of different severities. Sci. Rep. 7:9189. doi: 10.1038/s41598-017-09587-2
Dietrich, R. B., and Bradley, W. G. (1988). Iron accumulation in the basal ganglia following severe ischemic-anoxic insults in children. Radiology 168, 203–206. doi: 10.1148/radiology.168.1.3380958
Dong, H., Qiang, Z., Chai, D., Peng, J., Xia, Y., Hu, R., et al. (2020). Nrf2 inhibits ferroptosis and protects against acute lung injury due to intestinal ischemia reperfusion via regulating SLC7A11 and HO-1. Aging 12, 12943–12959. doi: 10.18632/aging.103378
Dong, X., Tan, Y., Zhuang, D., Hu, T., and Zhao, M. (2022). Global characteristics and trends in research on ferroptosis: A data-driven bibliometric study. Oxid. Med. Cell Longev. 2022:8661864. doi: 10.1155/2022/8661864
Du, X., West, M. B., Cheng, W., Ewert, D. L., Li, W., Saunders, D., et al. (2016). Ameliorative effects of antioxidants on the hippocampal accumulation of pathologic tau in a rat model of blast-induced traumatic brain injury. Oxid. Med. Cell Longev. 2016:4159357. doi: 10.1155/2016/4159357
El Sayed, N. S., and Ghoneum, M. H. (2020). Antia, a natural antioxidant product, attenuates cognitive dysfunction in streptozotocin-induced mouse model of sporadic Alzheimer’s disease by targeting the amyloidogenic, inflammatory, autophagy, and oxidative stress pathways. Oxid. Med. Cell Longev. 2020:4386562. doi: 10.1155/2020/4386562
Ellis, M. U., Marion, S. D., McArthur, D. L., Babikian, T., Giza, C., Kernan, C. L., et al. (2016). The UCLA study of children with moderate-to-severe traumatic brain injury: Event-related potential measure of interhemispheric transfer time. J. Neurotrauma 33, 990–996. doi: 10.1089/neu.2015.4023
Fang, W., Zhao, D., Wang, F., Li, M., Fan, S., Liao, W., et al. (2017). Neurotropin® alleviates hippocampal neuron damage through a HIF-1α/MAPK pathway. CNS Neurosci. Ther. 23, 428–437. doi: 10.1111/cns.12689
Feng, Z., Min, L., Chen, H., Deng, W., Tan, M., Liu, H., et al. (2021). Iron overload in the motor cortex induces neuronal ferroptosis following spinal cord injury. Redox. Biol. 43:101984. doi: 10.1016/j.redox.2021.101984
Fujiki, K., Inamura, H., Sugaya, T., and Matsuoka, M. (2019). Blockade of ALK4/5 signaling suppresses cadmium and erastin-induced cell death in renal proximal tubular epithelial cells via distinct signaling mechanisms. Cell Death Differ. 26, 2371–2385. doi: 10.1038/s41418-019-0307-8
Gao, M., Yi, J., Zhu, J., Minikes, A. M., Monian, P., Thompson, C. B., et al. (2019). Role of mitochondria in ferroptosis. Mol. Cell 73, 354–363.e3. doi: 10.1016/j.molcel.2018.10.042
Gao, S., Zhou, L., Lu, J., Fang, Y., Wu, H., Xu, W., et al. (2022). Cepharanthine attenuates early brain injury after subarachnoid hemorrhage in mice via inhibiting 15-lipoxygenase-1-mediated microglia and endothelial cell ferroptosis. Oxid. Med. Cell Longev. 2022:4295208. doi: 10.1155/2022/4295208
Gill, D., Monori, G., Tzoulaki, I., and Dehghan, A. (2018). Iron status and risk of stroke. Stroke 49, 2815–2821. doi: 10.1161/STROKEAHA.118.022701
Glotfelty, E. J., Delgado, T. E., Tovar-y-Romo, L. B., Luo, Y., Hoffer, B. J., Olson, L., et al. (2019). Incretin mimetics as rational candidates for the treatment of traumatic brain injury. ACS Pharmacol. Transl. Sci. 2, 66–91. doi: 10.1021/acsptsci.9b00003
Gomez, S. A., Abrey-Recalde, M. J., Panek, C. A., Ferrarotti, N. F., Repetto, M. G., Mejías, M. P., et al. (2013). The oxidative stress induced in vivo by Shiga toxin-2 contributes to the pathogenicity of haemolytic uraemic syndrome. Clin. Exp. Immunol. 173, 463–472. doi: 10.1111/cei.12124
Guigni, B. A., Callahan, D. M., Tourville, T. W., Miller, M. S., Fiske, B., Voigt, T., et al. (2018). Skeletal muscle atrophy and dysfunction in breast cancer patients: Role for chemotherapy-derived oxidant stress. Am. J. Physiol. Cell Physiol. 315, C744–C756. doi: 10.1152/ajpcell.00002.2018
Guo, H., Zhu, L., Tang, P., Chen, D., Li, Y., Li, J., et al. (2021). Carthamin yellow improves cerebral ischemia-reperfusion injury by attenuating inflammation and ferroptosis in rats. Int. J. Mol. Med. 47:52. doi: 10.3892/ijmm.2021.4885
Haidrani, L. (2016). Meningitis signs and symptoms. Nurs. Stand. 30:31. doi: 10.7748/ns.30.32.31.s33
Hall, C. J., Bouhafs, L., Dizcfalusy, U., and Sandstedt, K. (2010). Cryptococcus neoformans causes lipid peroxidation; therefore it is a potential inducer of atherogenesis. Mycologia 102, 546–551. doi: 10.3852/08-110
Han, K., Chapman, S. B., and Krawczyk, D. C. (2015). Altered amygdala connectivity in individuals with chronic traumatic brain injury and comorbid depressive symptoms. Front. Neurol. 6:231. doi: 10.3389/fneur.2015.00231
Hanley, D. F., Awad, I. A., Vespa, P. M., Martin, N. A., and Zuccarello, M. (2013). Hemorrhagic stroke: Introduction. Stroke 44, S65–S66. doi: 10.1161/STROKEAHA.113.000856
Harrison, L. H. (2006). Prospects for vaccine prevention of meningococcal infection. Clin. Microbiol. Rev. 19, 142–164. doi: 10.1128/CMR.19.1.142-164.2006
Hayano, M., Yang, W. S., Corn, C. K., Pagano, N. C., and Stockwell, B. R. (2016). Loss of cysteinyl-tRNA synthetase (CARS) induces the transsulfuration pathway and inhibits ferroptosis induced by cystine deprivation. Cell Death Differ. 23, 270–278. doi: 10.1038/cdd.2015.93
Horvath, R., Gorman, G., and Chinnery, P. F. (2008). How can we treat mitochondrial encephalomyopathies? Approaches to therapy. Neurotherapeutics 5, 558–568. doi: 10.1016/j.nurt.2008.07.002
Hu, L., Wang, H., Huang, L., Zhao, Y., and Wang, J. (2016). The protective roles of ROS-Mediated mitophagy on 125I seeds radiation induced cell death in HCT116 cells. Oxid. Med. Cell Longev. 2016:9460462. doi: 10.1155/2016/9460462
Huang, Y.-N., Yang, L.-Y., Greig, N. H., Wang, Y.-C., Lai, C.-C., and Wang, J.-Y. (2018). Neuroprotective effects of pifithrin-α against traumatic brain injury in the striatum through suppression of neuroinflammation, oxidative stress, autophagy, and apoptosis. Sci. Rep. 8:2368. doi: 10.1038/s41598-018-19654-x
Ismail Hassan, F., Didari, T., Khan, F., Niaz, K., Mojtahedzadeh, M., and Abdollahi, M. (2020). A review on the protective effects of metformin in sepsis-induced organ failure. Cell J. 21, 363–370. doi: 10.22074/cellj.2020.6286
Jabłońska-Trypuć, A., Świderski, G., Krętowski, R., and Lewandowski, W. (2017). Newly synthesized doxorubicin complexes with selected metals-synthesis, structure and anti-breast cancer activity. Molecules 22:1106. doi: 10.3390/molecules22071106
Jarrahi, A., Braun, M., Ahluwalia, M., Gupta, R. V., Wilson, M., Munie, S., et al. (2020). Revisiting traumatic brain injury: From molecular mechanisms to therapeutic interventions. Biomedicines 8:389. doi: 10.3390/biomedicines8100389
Jennis, M., Kung, C.-P., Basu, S., Budina-Kolomets, A., Leu, J. I.-J., Khaku, S., et al. (2016). An African-specific polymorphism in the TP53 gene impairs p53 tumor suppressor function in a mouse model. Genes Dev. 30, 918–930. doi: 10.1101/gad.275891.115
Jiang, H., Qi, Y.-T., Wu, W.-T., Wen, M.-Y., Liu, Y.-L., and Huang, W.-H. (2020). Intracellular monitoring of NADH release from mitochondria using a single functionalized nanowire electrode. Chem. Sci. 11, 8771–8778. doi: 10.1039/d0sc02787a
Jiang, X., Stockwell, B. R., and Conrad, M. (2021). Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 22, 266–282. doi: 10.1038/s41580-020-00324-8
Jing, X., Du, T., Li, T., Yang, X., Wang, G., Liu, X., et al. (2021). The detrimental effect of iron on OA chondrocytes: Importance of pro-inflammatory cytokines induced iron influx and oxidative stress. J. Cell Mol. Med. 25, 5671–5680. doi: 10.1111/jcmm.16581
Karuppagounder, S. S., Alin, L., Chen, Y., Brand, D., Bourassa, M. W., Dietrich, K., et al. (2018). N-acetylcysteine targets 5 lipoxygenase-derived, toxic lipids and can synergize with prostaglandin E2 to inhibit ferroptosis and improve outcomes following hemorrhagic stroke in mice. Ann. Neurol. 84, 854–872. doi: 10.1002/ana.25356
Kletetschka, G., Bazala, R., Takáč, M., and Svecova, E. (2021). Magnetic domains oscillation in the brain with neurodegenerative disease. Sci. Rep. 11:714. doi: 10.1038/s41598-020-80212-5
Kobeissy, F. H., Guingab-Cagmat, J. D., Zhang, Z., Moghieb, A., Glushakova, O. Y., Mondello, S., et al. (2016). Neuroproteomics and systems biology approach to identify temporal biomarker changes post experimental traumatic brain injury in rats. Front. Neurol. 7:198. doi: 10.3389/fneur.2016.00198
Koelman, D. L. H., Brouwer, M. C., and van de Beek, D. (2019). Targeting the complement system in bacterial meningitis. Brain 142, 3325–3337. doi: 10.1093/brain/awz222
Krabbendam, I. E., Honrath, B., Dilberger, B., Iannetti, E. F., Branicky, R. S., Meyer, T., et al. (2020). SK channel-mediated metabolic escape to glycolysis inhibits ferroptosis and supports stress resistance in C. elegans. Cell Death Dis. 11:263. doi: 10.1038/s41419-020-2458-4
Kwizera, R., Omali, D., Tadeo, K., Kasibante, J., Rutakingirwa, M. K., Kagimu, E., et al. (2021). Evaluation of the dynamiker cryptococcal antigen lateral flow assay for the diagnosis of HIV-associated cryptococcosis. J. Clin. Microbiol. 59, e2421–e2420. doi: 10.1128/JCM.02421-20
Law, Y.-S., Ngan, L., Yan, J., Kwok, L. Y., Sun, Y., Cheng, S., et al. (2018). Multiple kinases can phosphorylate the N-Terminal sequences of mitochondrial proteins in Arabidopsis thaliana. Front. Plant Sci. 9:982. doi: 10.3389/fpls.2018.00982
Lazarus, C., Soheilypour, M., and Mofrad, M. R. K. (2015). Torsional behavior of axonal microtubule bundles. Biophys. J. 109, 231–239. doi: 10.1016/j.bpj.2015.06.029
Lee, S. H., Choi, B. Y., Lee, S. H., Kho, A. R., Jeong, J. H., Hong, D. K., et al. (2017). Administration of protocatechuic acid reduces traumatic brain injury-induced neuronal death. Int. J. Mol. Sci. 18:2510. doi: 10.3390/ijms18122510
Lei, J., Chen, Z., Song, S., Sheng, C., Song, S., and Zhu, J. (2020). Insight into the role of ferroptosis in non-neoplastic neurological diseases. Front. Cell Neurosci. 14:231. doi: 10.3389/fncel.2020.00231
Lewandowska, M., Sajdak, S., and Lubiński, J. (2019). Can serum iron concentrations in early healthy pregnancy be risk marker of pregnancy-induced hypertension? Nutrients 11:1086. doi: 10.3390/nu11051086
Li, D., Lu, X., Xu, G., Liu, S., Gong, Z., Lu, F., et al. (2023). Dihydroorotate dehydrogenase regulates ferroptosis in neurons after spinal cord injury via the P53-ALOX15 signaling pathway. CNS Neurosci. Ther. 29, 1923–1939. doi: 10.1111/cns.14150
Li, J.-Z., Fan, B.-Y., Sun, T., Wang, X.-X., Li, J.-J., Zhang, J.-P., et al. (2023). Bioinformatics analysis of ferroptosis in spinal cord injury. Neural Regen. Res. 18, 626–633. doi: 10.4103/1673-5374.350209
Li, Z., Lin, Y., Mao, L., and Zhang, L. (2023). Expression characteristics of circular RNA in human traumatic brain injury. Front. Neurol. 13:1086553. doi: 10.3389/fneur.2022.1086553
Li, S., Zhang, Q., and Li, P. (2018). Protective effects of epifriedelinol in a rat model of traumatic brain injury assessed with histological and hematological markers. Transl. Neurosci. 9, 38–42. doi: 10.1515/tnsci-2018-0008
Li, X., Chen, S., Mao, L., Li, D., Xu, C., Tian, H., et al. (2019). Zinc improves functional recovery by regulating the secretion of granulocyte colony stimulating factor from microglia/macrophages after spinal cord injury. Front. Mol. Neurosci. 12:18. doi: 10.3389/fnmol.2019.00018
Liao, R., Wood, T. R., and Nance, E. (2020). Superoxide dismutase reduces monosodium glutamate-induced injury in an organotypic whole hemisphere brain slice model of excitotoxicity. J. Biol. Eng. 14:3. doi: 10.1186/s13036-020-0226-8
Lin, N.-H., and Su, Y.-J. (2020). A febrile old man with xanthochromic hypoglycorrhachia. Germs 10, 55–57. doi: 10.18683/germs.2020.1186
Lipponen, A., Natunen, T., Hujo, M., Ciszek, R., Hämäläinen, E., Tohka, J., et al. (2019). In vitro and in vivo pipeline for validation of disease-modifying effects of systems biology-derived network treatments for traumatic brain injury—lessons learned. Int. J. Mol. Sci. 20:5395. doi: 10.3390/ijms20215395
Liu, H., Hua, Y., Keep, R. F., and Xi, G. (2019). Brain ceruloplasmin expression after experimental intracerebral hemorrhage and protection against iron-induced brain injury. Transl. Stroke Res. 10, 112–119. doi: 10.1007/s12975-018-0669-0
Liu, J.-H., Wang, T.-W., Lin, Y.-Y., Ho, W.-C., Tsai, H.-C., Chen, S.-P., et al. (2020). Acrolein is involved in ischemic stroke-induced neurotoxicity through spermidine/spermine-N1-acetyltransferase activation. Exp. Neurol. 323:113066. doi: 10.1016/j.expneurol.2019.113066
Liu, Y., Hammel, G., Shi, M., Cheng, Z., Zivkovic, S., Wang, X., et al. (2021). Myelin debris stimulates NG2/CSPG4 expression in bone marrow-derived macrophages in the injured spinal cord. Front. Cell Neurosci. 15:651827. doi: 10.3389/fncel.2021.651827
Liu, Y.-S., Zhang, G.-Y., and Hou, Y. (2023). Theoretical and experimental investigation of the antioxidation mechanism of Loureirin C by radical scavenging for treatment of stroke. Molecules 28:380. doi: 10.3390/molecules28010380
Lorente, L., Martín, M. M., Abreu-González, P., Ramos, L., Cáceres, J. J., Argueso, M., et al. (2019). Maintained high sustained serum malondialdehyde levels after severe brain trauma injury in non-survivor patients. BMC Res. Notes 12:789. doi: 10.1186/s13104-019-4828-5
Lu, J., Xu, F., and Lu, H. (2020). LncRNA PVT1 regulates ferroptosis through miR-214-mediated TFR1 and p53. Life Sci. 260:118305. doi: 10.1016/j.lfs.2020.118305
Lutton, E. M., Razmpour, R., Andrews, A. M., Cannella, L. A., Son, Y.-J., Shuvaev, V. V., et al. (2017). Acute administration of catalase targeted to ICAM-1 attenuates neuropathology in experimental traumatic brain injury. Sci. Rep. 7:3846. doi: 10.1038/s41598-017-03309-4
Lv, Q.-B., Gao, X., Lin, D.-S., Chen, Y., Cao, B., and Zhou, K.-L. (2016). Effects of diammonium glycyrrhizinate on random skin flap survival in rats: An experimental study. Biomed. Rep. 5, 383–389. doi: 10.3892/br.2016.733
Lyamzaev, K. G., Panteleeva, A. A., Simonyan, R. A., Avetisyan, A. V., and Chernyak, B. V. (2023). Mitochondrial lipid peroxidation is responsible for ferroptosis. Cells 12:611. doi: 10.3390/cells12040611
Malla, B., Cotten, S., Ulshoefer, R., Paul, F., Hauser, A. E., Niesner, R., et al. (2020). Teriflunomide preserves peripheral nerve mitochondria from oxidative stress-mediated alterations. Therap. Adv. Chron. Dis. 11:2040622320944773. doi: 10.1177/2040622320944773
Margulies, S., Anderson, G., Atif, F., Badaut, J., Clark, R., Empey, P., et al. (2016). Combination therapies for traumatic brain injury: Retrospective considerations. J. Neurotrauma 33, 101–112. doi: 10.1089/neu.2014.3855
Mei, Z.-G., Huang, Y.-G., Feng, Z.-T., Luo, Y.-N., Yang, S.-B., Du, L.-P., et al. (2020). Electroacupuncture ameliorates cerebral ischemia/reperfusion injury by suppressing autophagy via the SIRT1-FOXO1 signaling pathway. Aging 12, 13187–13205. doi: 10.18632/aging.103420
Miao, Y., Chen, Y., Xue, F., Liu, K., Zhu, B., Gao, J., et al. (2022). Contribution of ferroptosis and GPX4’s dual functions to osteoarthritis progression. EBioMed. 76:103847. doi: 10.1016/j.ebiom.2022.103847
Nakamura, T., Ogawa, M., Kojima, K., Takayanagi, S., Ishihara, S., Hattori, K., et al. (2021). The mitochondrial Ca2+ uptake regulator, MICU1, is involved in cold stress-induced ferroptosis. EMBO Rep. 22, e51532. doi: 10.15252/embr.202051532
O’Reilly, C., Plamondon, R., and Lebrun, L.-H. (2014). Linking brain stroke risk factors to human movement features for the development of preventive tools. Front. Aging Neurosci. 6:150. doi: 10.3389/fnagi.2014.00150
Pandur, E., Tamási, K., Pap, R., Varga, E., Miseta, A., and Sipos, K. (2019). Fractalkine induces hepcidin expression of BV-2 microglia and causes iron accumulation in SH-SY5Y cells. Cell Mol. Neurobiol. 39, 985–1001. doi: 10.1007/s10571-019-00694-4
Park, J., Choi, W., Lee, P., Chung, S., Kim, B., Chung, H., et al. (2017). The novel resveratrol derivative 3,5-diethoxy-3′,4′-dihydroxy-trans-stilbene induces mitochondrial ROS-mediated ER stress and cell death in human hepatoma cells in vitro. Acta Pharmacol. Sin. 38, 1486–1500. doi: 10.1038/aps.2017.106
Park, J., Lee, D. G., Kim, B., Park, S.-J., Kim, J.-H., Lee, S.-R., et al. (2015). Iron overload triggers mitochondrial fragmentation via calcineurin-sensitive signals in HT-22 hippocampal neuron cells. Toxicology 337, 39–46. doi: 10.1016/j.tox.2015.08.009
Peng, W.-J., Reis, C., Reis, H., Zhang, J., and Yang, J. (2017). Predictive value of CTA spot sign on hematoma expansion in intracerebral hemorrhage patients. Biomed. Res. Int. 2017:4137210. doi: 10.1155/2017/4137210
Perez-Pinzon, M. A., Stetler, R. A., and Fiskum, G. (2012). Novel mitochondrial targets for neuroprotection. J. Cereb. Blood Flow Metab. 32, 1362–1376. doi: 10.1038/jcbfm.2012.32
Retchless, A. C., Itsko, M., Bazan, J. A., Turner, A. N., Hu, F., Joseph, S. J., et al. (2022). Evaluation of urethrotropic-clade meningococcal infection by urine metagenomic shotgun sequencing. J. Clin. Microbiol. 60, e0173221. doi: 10.1128/JCM.01732-21
Righi, V., Constantinou, C., Mintzopoulos, D., Khan, N., Mupparaju, S. P., Rahme, L. G., et al. (2013). Mitochondria-targeted antioxidant promotes recovery of skeletal muscle mitochondrial function after burn trauma assessed by in vivo 31P nuclear magnetic resonance and electron paramagnetic resonance spectroscopy. FASEB J. 27, 2521–2530. doi: 10.1096/fj.12-220764
Rodríguez-Graciani, K. M., Chapa-Dubocq, X. R., Ayala-Arroyo, E. J., Chaves-Negrón, I., Jang, S., Chorna, N., et al. (2022). Effects of ferroptosis on the metabolome in cardiac cells: The role of glutaminolysis. Antioxidants 11:278. doi: 10.3390/antiox11020278
Ruhatiya, R. S., Adukia, S. A., Manjunath, R. B., and Maheshwarappa, H. M. (2020). Current status and recommendations in multimodal neuromonitoring. Indian J. Crit. Care Med. 24, 353–360. doi: 10.5005/jp-journals-10071-23431
Scott, C., Arora, G., Dickson, K., and Lehmann, C. (2021). Iron chelation in local infection. Molecules 26:189. doi: 10.3390/molecules26010189
Serra, D., Henriques, J. F., Serra, T., Silva, A. B., Bronze, M. R., Dinis, T. C. P., et al. (2020). An anthocyanin-rich extract obtained from Portuguese blueberries maintains its efficacy in reducing microglia-driven neuroinflammation after simulated digestion. Nutrients 12:3670. doi: 10.3390/nu12123670
Shein, N. A., and Shohami, E. (2011). Histone deacetylase inhibitors as therapeutic agents for acute central nervous system injuries. Mol. Med. 17, 448–456. doi: 10.2119/molmed.2011.00038
Shen, J., Xin, W., Li, Q., Gao, Y., Yuan, L., and Zhang, J. (2019). Methylene blue reduces neuronal apoptosis and improves blood-brain barrier integrity after traumatic brain injury. Front. Neurol. 10:1133. doi: 10.3389/fneur.2019.01133
Shi, Z., Qiu, W., Xiao, G., Cheng, J., and Zhang, N. (2018). Resveratrol attenuates cognitive deficits of traumatic brain injury by activating p38 signaling in the brain. Med. Sci. Monit. 24, 1097–1103. doi: 10.12659/MSM.909042
Shojaie, L., Iorga, A., and Dara, L. (2020). Cell death in liver diseases: A review. Int. J. Mol. Sci. 21:9682. doi: 10.3390/ijms21249682
Shu, R., Zhang, L., Zhang, H., Li, Y., Wang, C., Su, L., et al. (2021). NMDA receptor modulates spinal iron accumulation via activating DMT1(-)IRE in remifentanil-induced hyperalgesia. J. Pain 22, 32–47. doi: 10.1016/j.jpain.2020.03.007
Simard, J. M., Woo, S. K., Norenberg, M. D., Tosun, C., Chen, Z., Ivanova, S., et al. (2010). Brief suppression of Abcc8 prevents autodestruction of spinal cord after trauma. Sci. Transl. Med. 2:28ra29. doi: 10.1126/scitranslmed.3000522
Singh, N., Haldar, S., Tripathi, A. K., Horback, K., Wong, J., Sharma, D., et al. (2014). Brain iron homeostasis: From molecular mechanisms to clinical significance and therapeutic opportunities. Antioxid. Redox Signal 20, 1324–1363. doi: 10.1089/ars.2012.4931
Slovinska, L., Harvanova, D., Janockova, J., Matejova, J., Cibur, P., Moravek, M., et al. (2022). Mesenchymal stem cells in the treatment of human spinal cord injury: The effect on individual values of pNF-H, GFAP, S100 proteins and selected growth factors, cytokines and chemokines. Curr. Issues Mol. Biol. 44, 578–596. doi: 10.3390/cimb44020040
Sprigg, N., Flaherty, K., Appleton, J. P., Salman, R. A.-S., Bereczki, D., Beridze, M., et al. (2018). Tranexamic acid for hyperacute primary IntraCerebral Haemorrhage (TICH-2): An international randomised, placebo-controlled, phase 3 superiority trial. Lancet 391, 2107–2115. doi: 10.1016/S0140-6736(18)31033-X
Sridhar, S., Hori, T., Nakagawa, R., Fukagawa, T., and Sanyal, K. (2021). Bridgin connects the outer kinetochore to centromeric chromatin. Nat. Commun. 12:146. doi: 10.1038/s41467-020-20161-9
Stelmashook, E. V., Isaev, N. K., Genrikhs, E. E., and Novikova, S. V. (2019). Mitochondria-targeted antioxidants as potential therapy for the treatment of traumatic brain injury. Antioxidants 8:124. doi: 10.3390/antiox8050124
Sun, Y., Li, Q., Guo, H., and He, Q. (2022). Ferroptosis and iron metabolism after intracerebral hemorrhage. Cells 12:90. doi: 10.3390/cells12010090
Taeubert, M. J., Wiertsema, C. J., Vermeulen, M. J., Quezada-Pinedo, H. G., Reiss, I. K., Muckenthaler, M. U., et al. (2021). Maternal iron status in early pregnancy and blood pressure throughout pregnancy, placental hemodynamics, and the risk of gestational hypertensive disorders. J. Nutr. 152, 525–534. doi: 10.1093/jn/nxab368
Tanaka, M., and Zhang, Y. (2022). Preclinical studies of posttraumatic headache and the potential therapeutics. Cells 12:155. doi: 10.3390/cells12010155
Tuo, Q.-Z., Lei, P., Jackman, K. A., Li, X.-L., Xiong, H., Li, X.-L., et al. (2017). Tau-mediated iron export prevents ferroptotic damage after ischemic stroke. Mol. Psychiatry 22, 1520–1530. doi: 10.1038/mp.2017.171
Uchida, H. A., Takatsuka, T., Hada, Y., Umebayashi, R., Takeuchi, H., Shikata, K., et al. (2022). Edaravone attenuated angiotensin II-induced atherosclerosis and abdominal aortic aneurysms in apolipoprotein E-deficient mice. Biomolecules 12:1117. doi: 10.3390/biom12081117
Votavova, H., Urbanova, Z., Kundrat, D., Dostalova Merkerova, M., Vostry, M., Hruba, M., et al. (2021). Modulation of the immune response by deferasirox in myelodysplastic syndrome patients. Pharmaceuticals 14:41. doi: 10.3390/ph14010041
Walker, P. A., Aroom, K. R., Jimenez, F., Shah, S. K., Harting, M. T., Gill, B. S., et al. (2009). Advances in progenitor cell therapy using scaffolding constructs for central nervous system injury. Stem Cell Rev. 5, 283–300. doi: 10.1007/s12015-009-9081-1
Wang, C., Chen, S., Guo, H., Jiang, H., Liu, H., Fu, H., et al. (2022). Forsythoside a mitigates Alzheimer’s-like pathology by inhibiting ferroptosis-mediated neuroinflammation via Nrf2/GPX4 axis activation. Int. J. Biol. Sci. 18, 2075–2090. doi: 10.7150/ijbs.69714
Wang, F., Li, J., Zhao, Y., Guo, D., Liu, D., Chang, S., et al. (2022). miR-672-3p promotes functional recovery in rats with contusive spinal cord injury by inhibiting ferroptosis suppressor protein 1. Oxid. Med. Cell Longev. 2022:6041612. doi: 10.1155/2022/6041612
Wang, P., Zhang, S., Liu, W., Chen, S., Lv, X., Hu, B., et al. (2022). Selenium attenuates TBHP-induced apoptosis of nucleus pulposus cells by suppressing mitochondrial fission through activating nuclear factor erythroid 2-related factor 2. Oxid. Med. Cell Longev. 2022:7531788. doi: 10.1155/2022/7531788
Wang, X., Sun, H., Cui, L., Wang, X., Ren, C., Tong, Z., et al. (2022). Acute high-altitude hypoxia exposure causes neurological deficits via formaldehyde accumulation. CNS Neurosci. Ther. 28, 1183–1194. doi: 10.1111/cns.13849
Wang, Y., Dong, Z., Zhang, Z., Wang, Y., Yang, K., and Li, X. (2022a). Postconditioning with irisin attenuates lung ischemia/reperfusion injury by suppressing ferroptosis via induction of the Nrf2/HO-1 signal axis. Oxid. Med. Cell Longev. 2022:9911167. doi: 10.1155/2022/9911167
Wang, Y., Feng, F., He, W., Sun, L., He, Q., and Jin, J. (2022b). miR-188-3p abolishes germacrone-mediated podocyte protection in a mouse model of diabetic nephropathy in type I diabetes through triggering mitochondrial injury. Bioengineered 13, 774–788. doi: 10.1080/21655979.2021.2012919
Wang, D., Wang, K., Liu, Z., Wang, Z., and Wu, H. (2020). Valproic acid-labeled chitosan nanoparticles promote recovery of neuronal injury after spinal cord injury. Aging 12, 8953–8967. doi: 10.18632/aging.103125
Wang, Y., Quan, F., Cao, Q., Lin, Y., Yue, C., Bi, R., et al. (2020). Quercetin alleviates acute kidney injury by inhibiting ferroptosis. J. Adv. Res. 28, 231–243. doi: 10.1016/j.jare.2020.07.007
Wang, M.-J., Liu, X., Chi, S., Wang, F.-Y., Zhang, L., Yao, M.-Y., et al. (2023). Activation of PPARγ prevents ferroptosis-induced damage through Nrf2/Gpx4 signaling pathway in rat hippocampus following epilepsy. Neuroreport 34, 368–374. doi: 10.1097/WNR.0000000000001892
Wang, R., Wu, X., Tian, Z., Hu, T., Cai, C., Wu, G., et al. (2023). Sustained release of hydrogen sulfide from anisotropic ferrofluid hydrogel for the repair of spinal cord injury. Bioact. Mater. 23, 118–128. doi: 10.1016/j.bioactmat.2022.10.020
Wang, T.-X., Liang, J.-Y., Zhang, C., Xiong, Y., Guan, K.-L., and Yuan, H.-X. (2019). The oncometabolite 2-hydroxyglutarate produced by mutant IDH1 sensitizes cells to ferroptosis. Cell Death Dis. 10:755. doi: 10.1038/s41419-019-1984-4
Wang, W., Green, M., Choi, J. E., Gijón, M., Kennedy, P. D., Johnson, J. K., et al. (2019). CD8+ T cells regulate tumor ferroptosis during cancer immunotherapy. Nature 569, 270–274. doi: 10.1038/s41586-019-1170-y
Wei, X.-E., Zhang, Y.-Z., Li, Y.-H., Li, M.-H., and Li, W.-B. (2012). Dynamics of rabbit brain edema in focal lesion and perilesion area after traumatic brain injury: A MRI study. J. Neurotrauma 29, 2413–2420. doi: 10.1089/neu.2010.1510
Wu, C., Zhao, W., Yu, J., Li, S., Lin, L., and Chen, X. (2018). Induction of ferroptosis and mitochondrial dysfunction by oxidative stress in PC12 cells. Sci. Rep. 8:574. doi: 10.1038/s41598-017-18935-1
Wu, J., Hua, Y., Keep, R. F., Nakamura, T., Hoff, J. T., and Xi, G. (2003). Iron and iron-handling proteins in the brain after intracerebral hemorrhage. Stroke 34, 2964–2969. doi: 10.1161/01.STR.0000103140.52838.45
Wu, L., Xiong, X., Wu, X., Ye, Y., Jian, Z., Zhi, Z., et al. (2020). Targeting oxidative stress and inflammation to prevent ischemia-reperfusion injury. Front. Mol. Neurosci. 13:28. doi: 10.3389/fnmol.2020.00028
Wu, Y., Zhang, S., Gong, X., Tam, S., Xiao, D., Liu, S., et al. (2020). The epigenetic regulators and metabolic changes in ferroptosis-associated cancer progression. Mol. Cancer 19:39. doi: 10.1186/s12943-020-01157-x
Xi, K., Gu, Y., Tang, J., Chen, H., Xu, Y., Wu, L., et al. (2020). Microenvironment-responsive immunoregulatory electrospun fibers for promoting nerve function recovery. Nat. Commun. 11:4504. doi: 10.1038/s41467-020-18265-3
Xia, N., Cai, J., Wang, F., Dong, B., Liu, S., Chen, F., et al. (2016). SENP1 Is a Crucial Regulator for Cell Senescence through DeSUMOylation of Bmi1. Sci. Rep. 6:34099. doi: 10.1038/srep34099
Xie, Y., Song, X., Sun, X., Huang, J., Zhong, M., Lotze, M. T., et al. (2016). Identification of baicalein as a ferroptosis inhibitor by natural product library screening. Biochem. Biophys. Res. Commun. 473, 775–780. doi: 10.1016/j.bbrc.2016.03.052
Xu, X., Lin, D., Tu, S., Gao, S., Shao, A., and Sheng, J. (2021). Is ferroptosis a future direction in exploring cryptococcal meningitis? Front. Immunol. 12:598601. doi: 10.3389/fimmu.2021.598601
Yang, X., Yin, F., Liu, Q., Ma, Y., Zhang, H., Guo, P., et al. (2022). Ferroptosis-related genes identify tumor immune microenvironment characterization for the prediction of prognosis in cervical cancer. Ann. Transl. Med. 10:123. doi: 10.21037/atm-21-6265
Yao, X., Zhang, Y., Hao, J., Duan, H.-Q., Zhao, C.-X., Sun, C., et al. (2019). Deferoxamine promotes recovery of traumatic spinal cord injury by inhibiting ferroptosis. Neural Regen. Res. 14, 532–541. doi: 10.4103/1673-5374.245480
Yauger, Y. J., Bermudez, S., Moritz, K. E., Glaser, E., Stoica, B., and Byrnes, K. R. (2019). Iron accentuated reactive oxygen species release by NADPH oxidase in activated microglia contributes to oxidative stress in vitro. J Neuroinflammation 16:41. doi: 10.1186/s12974-019-1430-7
Ye, Z., Zhuo, Q., Hu, Q., Xu, X., Liu, M., Zhang, Z., et al. (2021). FBW7-NRA41-SCD1 axis synchronously regulates apoptosis and ferroptosis in pancreatic cancer cells. Redox Biol. 38:101807. doi: 10.1016/j.redox.2020.101807
Yigitkanli, K., Zheng, Y., Pekcec, A., Lo, E. H., and van Leyen, K. (2017). Increased 12/15-lipoxygenase leads to widespread brain injury following global cerebral ischemia. Transl. Stroke Res. 8, 194–202. doi: 10.1007/s12975-016-0509-z
Yu, J., Zhang, Y., Ma, H., Zeng, R., Liu, R., Wang, P., et al. (2020). Epitranscriptomic profiling of N6-methyladenosine-related RNA methylation in rat cerebral cortex following traumatic brain injury. Mol. Brain 13:11. doi: 10.1186/s13041-020-0554-0
Yu, Y., Jiang, L., Wang, H., Shen, Z., Cheng, Q., Zhang, P., et al. (2020). Hepatic transferrin plays a role in systemic iron homeostasis and liver ferroptosis. Blood 136, 726–739. doi: 10.1182/blood.2019002907
Zavodska, M., Galik, J., Marsala, M., Papcunova, S., Pavel, J., Racekova, E., et al. (2018). Hypothermic treatment after computer-controlled compression in minipig: A preliminary report on the effect of epidural vs. direct spinal cord cooling. Exp. Ther. Med. 16, 4927–4942. doi: 10.3892/etm.2018.6831
Zeng, J., Zhu, L., Liu, J., Zhu, T., Xie, Z., Sun, X., et al. (2019). Metformin protects against oxidative stress injury induced by ischemia/reperfusion via regulation of the lncRNA-H19/miR-148a-3p/Rock2 axis. Oxid. Med. Cell Longev. 2019:8768327. doi: 10.1155/2019/8768327
Zhang, C., Liu, X., Jin, S., Chen, Y., and Guo, R. (2022). Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 21:47. doi: 10.1186/s12943-022-01530-y
Zhang, H.-Y., Lu, X., Hao, Y.-H., Tang, L., and He, Z.-Y. (2022). Oxidized low-density lipoprotein receptor 1: A novel potential therapeutic target for intracerebral hemorrhage. Neural Regen. Res. 17, 1795–1801. doi: 10.4103/1673-5374.332157
Zhang, H., Chen, Y., Li, F., Wu, C., Cai, W., Ye, H., et al. (2023). Elamipretide alleviates pyroptosis in traumatically injured spinal cord by inhibiting cPLA2-induced lysosomal membrane permeabilization. J. Neuroinflammation 20:6. doi: 10.1186/s12974-023-02690-4
Zhang, L., Wang, H., Zhou, X., Mao, L., Ding, K., and Hu, Z. (2019). Role of mitochondrial calcium uniporter-mediated Ca2+ and iron accumulation in traumatic brain injury. J. Cell Mol. Med. 23, 2995–3009. doi: 10.1111/jcmm.14206
Zhang, Y., Sun, C., Zhao, C., Hao, J., Zhang, Y., Fan, B., et al. (2019). Ferroptosis inhibitor SRS 16-86 attenuates ferroptosis and promotes functional recovery in contusion spinal cord injury. Brain Res. 1706, 48–57. doi: 10.1016/j.brainres.2018.10.023
Zhang, Y., Fan, B.-Y., Pang, Y.-L., Shen, W.-Y., Wang, X., Zhao, C.-X., et al. (2020). Neuroprotective effect of deferoxamine on erastin-induced ferroptosis in primary cortical neurons. Neural Regen. Res. 15, 1539–1545. doi: 10.4103/1673-5374.274344
Zhang, Y., Xia, M., Zhou, Z., Hu, X., Wang, J., Zhang, M., et al. (2021). p53 Promoted ferroptosis in ovarian cancer cells treated with human serum incubated-superparamagnetic iron oxides. Int. J. Nanomed. 16, 283–296. doi: 10.2147/IJN.S282489
Zhong, F.-M., Yao, F.-Y., Liu, J., Zhang, H.-B., Zhang, J., Zhang, N., et al. (2022). Ferroptosis-related molecular patterns reveal immune escape, inflammatory development and lipid metabolism characteristics of the tumor microenvironment in acute myeloid leukemia. Front. Oncol. 12:888570. doi: 10.3389/fonc.2022.888570
Zhou, H., Yin, C., Zhang, Z., Tang, H., Shen, W., Zha, X., et al. (2020). Proanthocyanidin promotes functional recovery of spinal cord injury via inhibiting ferroptosis. J. Chem. Neuroanat. 107:101807. doi: 10.1016/j.jchemneu.2020.101807
Zhou, L., Yu, P., Wang, T.-T., Du, Y.-W., Chen, Y., Li, Z., et al. (2022). Polydatin attenuates cisplatin-induced acute kidney injury by inhibiting ferroptosis. Oxid. Med. Cell Longev. 2022:9947191. doi: 10.1155/2022/9947191
Zhou, M., and Huang, Z. (2019). Effect of comprehensive cerebral protection program on cerebral oxygen metabolism and vascular endothelial function in elderly patients with acute cerebral infarction. Iran J. Public Health 48, 299–304.
Zhou, Z. D., and Tan, E.-K. (2017). Iron regulatory protein (IRP)-iron responsive element (IRE) signaling pathway in human neurodegenerative diseases. Mol. Neurodegener. 12:75. doi: 10.1186/s13024-017-0218-4
Keywords: ferroptosis, acute central nervous system injury, traumatic brain injury, spinal cord injury, mitochondrial homeostasis
Citation: Dong W, Gong F, Zhao Y, Bai H and Yang R (2023) Ferroptosis and mitochondrial dysfunction in acute central nervous system injury. Front. Cell. Neurosci. 17:1228968. doi: 10.3389/fncel.2023.1228968
Received: 25 May 2023; Accepted: 20 July 2023;
Published: 09 August 2023.
Edited by:
Diogo Trigo, University of Aveiro, PortugalReviewed by:
Gilberto L. Pardo Andreu, University of Havana, CubaCopyright © 2023 Dong, Gong, Zhao, Bai and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ruixin Yang, eWFuZ3J1aXhpbmZtbXVAMTI2LmNvbQ==; Hongmin Bai, YmFpaG1mbW11QHZpcC4xNjMuY29t
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.