 Maya Junghans1†
Maya Junghans1† Felix John1†
Felix John1† Hilal Cihankaya1
Hilal Cihankaya1 Daniel Schliebs1
Daniel Schliebs1 Konstanze F. Winklhofer2,3
Konstanze F. Winklhofer2,3 Verian Bader2,4
Verian Bader2,4 Johann Matschke5
Johann Matschke5 Carsten Theiss1
Carsten Theiss1 Veronika Matschke1*
Veronika Matschke1*- 1Department of Cytology, Institute of Anatomy, Ruhr University Bochum, Bochum, Germany
- 2Department of Molecular Cell Biology, Institute of Biochemistry and Pathobiochemistry, Ruhr University Bochum, Bochum, Germany
- 3Cluster of Excellence RESOLV, Bochum, Germany
- 4Department of Biochemistry of Neurodegenerative Diseases, Institute of Biochemistry and Pathobiochemistry, Ruhr University Bochum, Bochum, Germany
- 5Institute of Cell Biology (Cancer Research), University Hospital Essen, University of Duisburg-Essen, Essen, Germany
Background: Amyotrophic lateral sclerosis (ALS) is an incurable neurodegenerative disease characterized by the loss of motor neurons in cerebral cortex, brainstem and spinal cord. Numerous studies have demonstrated signs of oxidative stress in postmortem neuronal tissue, cerebrospinal fluid, plasma and urine of ALS patients, without focusing on the specific processes within motor neurons. Thus, we aimed to investigate the relevance of reactive oxygen species (ROS) detoxification mechanisms and its consequences on the formation of toxic/lethal DNA double strand breaks (DSBs) in the ALS model of the Wobbler mouse.
Methods: Live cell imaging in dissociated motor neuronal cultures was used to investigate the production of ROS using Dihydroethidium (DHE). The expression levels of ROS detoxifying molecules were investigated by qPCR as well as Western blots. Furthermore, the expression levels of DNA damage response proteins p53bp1 and H2ax were investigated using qPCR and immunofluorescence staining. Proof-of-principle experiments using ROS scavengers were performed in vitro to decipher the influence of ROS on the formation of DNA double strand breaks quantifying the γH2ax spots formation.
Results: Here, we verified an elevated ROS-level in spinal motor neurons of symptomatic Wobbler mice in vitro. As a result, an increased number of DNA damage response proteins p53bp1 and γH2ax in dissociated motor neurons of the spinal cord of Wobbler mice was observed. Furthermore, we found a significantly altered expression of several antioxidant molecules in the spinal cord of Wobbler mice, suggesting a deficit in ROS detoxification mechanisms. This hypothesis could be verified by using ROS scavenger molecules in vitro to reduce the number of γH2ax foci in dissociated motor neurons and thus counteract the harmful effects of ROS.
Conclusion: Our data indicate that maintenance of redox homeostasis may play a key role in the therapy of the neurodegenerative disease ALS. Our results underline a necessity for multimodal treatment approaches to prolong the average lifespan of motor neurons and thus slow down the progression of the disease, since a focused intervention in one pathomechanism seems to be insufficient in ALS therapy.
Introduction
Amyotrophic lateral sclerosis (ALS) is the most common motor neuron disease in adults with a prevalence of 3–5/100,000 in Europe and the United States (Brown and Al-Chalabi, 2017). About 5%–10% of ALS patients are diagnosed with the hereditary, familial form (fALS) and 90%–95% with the sporadic form of ALS (sALS). In individual cases a clear classification to a specific subtype is often difficult (Al-Chalabi et al., 2016). This neurodegenerative disease is characterized by the loss of both, the upper motor neurons in the cerebral cortex and the lower motor neurons in the brainstem and spinal cord (Brown and Al-Chalabi, 2017; Petrov et al., 2017).
ALS is rapidly progressive and 50% of all ALS patients die in the first 30 months after symptoms appear, often due to respiratory failure (Kiernan et al., 2011). Although ALS was first described by Charcot and Joffroy in 1869, the causality of the disease is still poorly understood (Petrov et al., 2017). In recent decades, ALS research has identified a great heterogeneity in pathological abnormalities between different ALS subgroups like modified protein homeostasis, excitotoxicity, modified RNA homeostasis, neuroinflammation, dysfunctional axonal transport, mitochondrial dysfunction, and oxidative stress (Brown and Al-Chalabi, 2017; Tam et al., 2019). Currently there are only two drugs approved for the treatment of ALS: Riluzole, an anti-glutamatergic agent, extending the life of ALS patients for approximately 3 months (Jaiswal, 2019), and Edaravone, an antioxidant and free-radical scavenger (Brown and Al-Chalabi, 2017; Jaiswal, 2019), with its capability to inhibit ALS motor deterioration and thus slowing disease progression in a subpopulation of ALS patients, especially patients with shorter duration and milder symptoms (Abe et al., 2014; Yoshino, 2019).
Various mouse models with different ALS-associated mutations are currently available to study the pathomechanisms of ALS (Stephenson and Amor, 2017; Cihankaya et al., 2021), including the Wobbler mouse, which was first described by Falconer in 1956 (Andrews et al., 1974; Giorgio et al., 2019). The Wobbler mouse displays almost the same symptoms as human ALS patients at the clinical-symptomatic, cellular and metabolic levels (Boillée et al., 2003; Moser et al., 2013; Ott et al., 2015). A spontaneous autosomal recessive missense mutation L967Q located in the Vps54 gene, mapped to mouse chromosome 11, is responsible for the Wobbler phenotype (Kaupmann et al., 1992; Schmitt-John et al., 2005). The development of the clinical phenotype of Wobbler mice can be divided into three different phases (Ott et al., 2015). During the first pre-symptomatic phase, from p0 to p19, Wobbler mice cannot be distinguished from their wild-type and heterozygous siblings (Boillée et al., 2003). This is followed by the evolutionary phase from p20 to p39 which is characterized by a development of symptoms. After forty days the disease development proceeds with the stabilized clinical phase which is characterized by a stagnation of symptoms (Duchen and Strich, 1968; Ott et al., 2015; Saberi et al., 2016). The fully developed phenotype of the homozygous Wobbler mouse shows a head tremor, a wobbly gait pattern, an inability to climb and an overall muscle atrophy as well as a reduced body size and weight compared to wild-type mice (Ott et al., 2015).
Recent studies described the importance of oxidative stress for the pathogenesis, both in Wobbler mice and in ALS patients (Petrov et al., 2017; Röderer et al., 2018; Matschke et al., 2019; Tam et al., 2019; Wang et al., 2019; Zwilling et al., 2020; Stein et al., 2021). Our previous studies identified significantly increased levels of reactive oxygen species (ROS) as a marker for oxidative stress in the stable clinical phase (p40) of the Wobbler spinal cord (Röderer et al., 2018). In many cases, ROS arise as physiological molecules (Gorrini et al., 2013; Pourahmad et al., 2016; Matschke et al., 2019). Depending on their concentration, ROS influence various biological processes, and a balanced intracellular ROS concentration is essential for a continuously functioning cell metabolism (Rhee, 2006; Janssen-Heininger et al., 2008; Brieger et al., 2012; Matschke et al., 2019). However, excessive ROS production or a failure in defense mechanisms is extremely harmful to the cell as components such as DNA, RNA, proteins and lipids are oxidized. ROS can lead to single-strand (SSB) or double strand breaks (DSB) of the DNA via various factors. Among others, ROS can directly lead to DNA SSB through the oxidation of bases. In case of strongly increased ROS concentrations, these types of damage can occur in close proximity to each other on both DNA strands, resulting in DNA DSB (Cannan et al., 2014; Henrikus et al., 2019; Srinivas et al., 2019). Mitotic cells can counter such damage effectively and accurately by homologous recombination. However, post-mitotic cells, such as neurons, may respond to such damage only through rather error-prone but essential mechanisms such as non-homologous end joining (NHEJ; Lieber, 2010; Iyama and Wilson, 2013; Shanbhag et al., 2019). Oxidative DNA damage in post-mitotic cells might therefore be particularly dangerous for the survival of neuronal cells, as they can accumulate with age and lead to cell death (Islam, 2017). To counteract oxidative stress and thus oxidative DNA damage under physiological conditions the cell is equipped with various antioxidant mechanisms to scavenge ROS and maintain redox homeostasis, which is essential for normal cell function (Lagouge and Larsson, 2013; Pourahmad et al., 2016; Matschke et al., 2019).
Recent studies have already tried to overcome oxidative stress in Wobbler mice. Edaravone, as a free radical scavenger delays the disease progression in Wobbler mice and is already an approved drug for ALS patients in Japan, South Korea, Canada and the US (Ikeda and Iwasaki, 2015; Yoshino, 2019). N-Acetyl-L-Cysteine (NAC) can also act as an antioxidant—either directly or as a precursor for the antioxidant glutathione (Henderson et al., 1996). The degeneration of Wobbler mice motor neurons, shown by axon caliber, muscle size, muscle area and forelimb functionality, could significantly be decreased by giving the mice NAC-supplemented water (Henderson et al., 1996). In addition, Wobbler mice exposed to intraperitoneally high doses of the antioxidant Methylcarbylamine experienced a reduced rate of symptom progression compared to non-treated mice (Ikeda et al., 2015).
Since several studies observed a delay in symptomatic and pathological progression of motor neuron disease in Wobbler mice by the use of antioxidant molecules, we hypothesize that a dysfunction of antioxidant mechanisms may be the reason for increased ROS levels in spinal cord of this mouse model. Therefore, we aimed to investigate the underlying processes and mechanisms for the increased oxidative stress in motor neurons of Wobbler mice. Here, we deciphered the role of the redox homeostasis in Wobbler spinal cord and dissociated motor neuronal enriched cell culture on the formation of toxic DNA double strand breaks (DNBs) as well as the impact of ROS scavengers on the prevention of integrity of Wobbler motor neuron DNA.
Materials and Methods
Animals
All procedures were conducted under established standards of the German federal state of North Rhine Westphalia, in accordance with the European Communities Council Directive 2010/63/EU on the protection of animals used for scientific purposes. Animal experiments were carried out according to the German animal welfare regulations and approved by the local authorities (registration number Az. 84-02.04.2017.A085). As described earlier, for all experiments the mouse strain C57BL/Fa carrying the Wobbler mutation was used (Ott et al., 2015). Breeding, handling and genotyping of mice was performed as previously described (Ott et al., 2015). For each experiment, the spinal cord of both homozygous genotypes (wild-type and Wobbler) and gender was collected. Since our focus was to investigate oxidative DNA damage, only tissue from the stable clinical stage (p40) was used, as we previously showed no oxidative stress in the Wobbler spinal cord at earlier time points (Röderer et al., 2018). Heterozygous animals were only used for breeding.
Immunofluorescence staining of DNA damage response proteins
To investigate whether the cervical spinal cord of Wobbler mice exhibits increased expression of DNA damage response proteins p53bp1 and phosphorylated H2ax (γH2ax), immunofluorescence staining on sections of five animals per genotype was performed. For this purpose animals (p40) were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg) and perfused with 4% PFA in PBS, as described before (Klatt et al., 2019). After decapitation and dissection of the cervical spinal cord, the tissue was post fixated with 4% PFA in PBS at 4°C for 24 h, transferred to 30% sucrose for 48 h and frozen in isopentane at −45°C (Klatt et al., 2019). Afterwards the tissue was embedded in tissue freezing medium (#14020108926, Leica Biosystems Inc., Buffalo Grove, Illinois, USA). Ten μm thick slices of cervical spinal cord were cut on a cryostat (Thermo Fisher Scientific, CryoStar NX50 Cryostat, chamber temperature −18°C; stage temperature −20°C). Cryosections were applied on SuperFrost® Plus objective slides (Thermo Fisher Scientific, Waltham, Massachusetts, USA) and dried at 37°C for 30 min.
For immunofluorescence staining, cryosections were incubated for 1 min with PBS followed by 60 min incubation with 0.3% Triton-X100 and 5% goat serum in PBS. Further, the sections were incubated with the primary antibody p53bp1 (1:100 in 5% goat serum in PBS, #sc-22760, Santa-Cruz Biotechnology, Dallas, Texas, USA) or γH2ax (1:200 in 5% goat serum in PBS, #2577, Cell Signaling Technology, Danvers, Massachusetts, USA) at 4°C over night. After washing with PBS, the sections were reacted with the secondary antibody anti-rabbit-AlexaFluor488 (1:200 in PBS, #A11008, Thermo Fisher Scientific, Waltham, Massachusetts, USA) for 2 h at room temperature. Nuclei were stained with DAPI (1μg/ml; #D9542, Sigma-Aldrich, Danvers, Massachusetts, USA) for 30 min. Finally, the slices were covered with fluoroshield (#F6937, Sigma-Aldrich, St. Louis, Missouri, USA) and stored at 4°C.
Samples were imaged using an inverted confocal Laser Scanning Microscope (LSM 800, Carl Zeiss Microscopy GmbH, Jena, Germany) equipped with the respective filter sets in combination with a 10× (Plan-Apochromat 10×/0.45, Carl Zeiss Microscopy GmbH, Jena, Germany) and 40× objective (Plan-Apochromat 40×/1.4 Oil, Carl Zeiss Microscopy GmbH, Jena, Germany). Fluorescence image acquisition settings were kept the same for all conditions to achieve quantitative comparability of p53bp1 or γH2ax signals. Secondary antibodies were tested for specificity and showed no unspecific binding. ImageJ 1.53f51 (National Institute of Health, USA) was used to analyze the fluorescence intensity of p53bp1 and γH2ax in cervical spinal cord sections. Fluorescence signals of p53bp1 or γH2ax were measured in the nuclear area of individual cells, normalized to the respective DNA content (integrated DAPI signals) and expressed as relative fluorescence units (RFU) representing the relative levels of p53bp1 or γH2ax.
RNA isolation, reverse transcription, and quantitative real-time PCR
RNA isolation, Reverse Transcription (RT) and qPCR were performed as described earlier (Rohm et al., 2019). Total RNA was extracted from the cervical spinal cord tissue of five Wobbler and five wild-type mice at stable clinical phase (p40) using the NucleoSpin® miRNA isolation kit (# 740304, Macherey-Nagel, Düren, Germany) according to the manufacturer’s protocol for animal tissue and cultured cells. cDNA was synthesized using qScript cDNA SuperMix (#95048, QuantaBio, Beverly, Massachusetts, USA) according to the manufacturer’s protocol using 500 ng total RNA and oligo(dT) primer. Quantitative real-time PCR was performed on a CFX96 real-time PCR Detection System (Bio-Rad, Hercules, California, USA) using GoTaq qPCR Master Mix (#A6001, Promega, Madison, Wisconsin, USA). Specific primer used are shown in Table 1. Expression levels were analyzed in three independent qPCR runs in triplicates and normalized to GAPDH. The obtained Ct-values were analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).

Table 1. Primer sequences used for quantitative real-time PCR.
SDS-gel electrophoresis and Western blotting
SDS-gel electrophoresis and Western blotting were performed as previously described (Stein et al., 2021). For SDS-gel electrophoresis proteins were isolated from four to eight cervical spinal cord samples (p40) per genotype using Cell lysis buffer (#9803S, Cell Signaling Technology, Danvers, Massachusetts, USA) supplemented with protease inhibitor (#11697498001, Merck, Darmstadt, Germany). Protein concentrations were determined by PierceTM BCA Protein Assay Kit (#23225, Thermo Fisher Scientific, Waltham, Massachusetts, USA). Fifty microgram of total protein was separated by SDS-gel electrophoresis and transferred to a nitrocellulose membrane. Blots were blocked with 1% RotiBlock (#A151, Roth, Karlsruhe, Germany) in 1× TBS for 1 h at room temperature. Primary antibodies (Table 2) were incubated over night at 4°C. HRP-coupled secondary antibodies (Table 2) were incubated for 1 h at room temperature. Finally, Immuno Cruz Luminol Agent (#sc-2048, Santa Cruz Biotechnology, Dallas, Texas, USA) was used for signal detection with an imaging system (ChemiDoc XRS+, Bio-Rad, Hercules, California, USA). For arithmetic analysis of the band intensity ImageJ 1.53f51 (National Institute of Health, USA) software was used. Band intensities of proteins of interest were normalized to the housekeeper actin. Normalized protein levels were compared between different genotypes.

Table 2. Primary and secondary antibodies used for Western blotting.
Motor neuron enriched dissociated cell cultures of the ventral horn
To study the ROS amount and the expression of the DNA damage response proteins in motor neurons ex vivo, the ventral horn of the cervical spinal cord of Wobbler and wild-type mice from stable clinical phase (p40) was isolated and a dissociated motor neuronal cell culture was prepared according to Zwilling et al. (2020). Cells were seeded on glass slides and kept in culture for 10 days in vitro (div) with periodic medium changes every 48 h. For the entire time, the temperature was kept at 37°C and the CO2 level at 5%. The motor neurons to be studied were selected on the basis of their morphology (multipolar neurons) with a soma size of 60–90 μm and a nucleus size of about 20 μm as described earlier (Weber et al., 1997; Thomson et al., 2012). To investigate whether the application of ROS scavengers affects the amount of DNA damage response protein γH2ax in motor neurons, the following supplements were added to the cell culture medium:
(i) N-Acetyl-L-Cysteine (NAC; #sc202232, Santa Cruz, Dallas, Texas, USA) was dissolved in PBS at a concentration of 10 M and diluted to the final concentration of 20 μM directly in culture medium. Dissociated cells of the ventral horn were cultured for 8 days in normal culturing medium and for 48 h with supplemented medium.
(ii) Glutathione ethyl ester (#14953, Cayman Chemical, Ann Arbor, Michigan, USA) was dissolved in water at a concentration of 74.5 mM and diluted to the final concentration of 500 μM directly in culture medium. The dissociated cells of the ventral horn were cultured for 9 days in normal culturing medium and for 24 h with supplemented medium.
(iii) Mito-TEMPO (#sc-221945, Santa Cruz, Dallas, Texas, USA) was dissolved in DMSO at a concentration of 10 mM and diluted to the final concentration of 1 μM directly in culture medium. The dissociated cells of the ventral horn were cultured for 9 days in normal culturing medium and for 24 h with supplemented medium.
At least four animals per genotype were used for one procedure of motor neuron isolation. The entire experimental set-up was repeated at least five times per supplement.
ROS measurement in dissociated motor neuronal cultures
For measurement of ROS amount in motor neurons in vitro, live-cell imaging was performed 10 days after cell isolation. The motor neurons to be studied were identified as described above (see Section “Motor neuron enriched dissociated cell cultures of the ventral horn”). After 15 min of live-cell imaging under normal conditions, medium was replaced by culturing medium including Dihydroethidium (DHE; final concentration: 10 μM) which reacts with superoxide anions resulting in a red immunofluorescence (Owusu-Ansah et al., 2008). The fluorescence signals of the cells were documented every 5 min for 1 h in DHE-including medium, 37°C and 5% CO2. Live-cell imaging was performed using a spinning disc confocal microscope (VisiScope Confocal-Cell Explorer, Visitron Systems GmbH, Puchheim, Germany) with a 20× objective (Plan-Apo 20×/0.75, Nikon Europe BV, Amsterdam, Netherlands). ImageJ 1.53f51 (National Institute of Health, USA) was used to analyze the average red fluorescence intensity within the soma of a motor neuron, while motor neurons themselves were detected in transmitted-light channel. A mean value of background fluorescence intensity without DHE addition was used for normalization. We prepared five different cell cultures per genotype for live cell imaging. Four different motor neurons of each motor neuronal preparation were subsequently documented over a period of 80 min.
Immunofluorescence staining of DNA damage response proteins in vitro
To investigate the number of DNA damage response proteins in the nucleus of motor neurons, after 10 days in culture, the cells were fixed using 4% PFA for 15 min and washed with PBS afterwards. For permeabilization, glass slices with dissociated cells of the ventral horn were incubated with 0.3% Triton in PBS for 15 min at room temperature. After washing with PBS, unspecific binding sites were blocked with 2% goat serum in PBS for 30 min. The slices were then incubated with primary antibodies (rabbit-α-phospho-histone H2A.X (Ser139) antibody, #2577, Cell Signaling Technology, Danvers, Massachusetts, USA, 1:500 in 2% goat serum in PBS; chicken-α-NEFM Antibody, #PA1-16758, Thermo Fisher Scientific, Waltham, Massachusetts, USA, 1:1,000 in 10% goat serum in PBS) at 4°C over night. Thereafter, samples were reacted with secondary antibodies (α-rabbit-Alexa488, #11008, Thermo Fisher Scientific, Waltham, Massachusetts, USA, 1:300 in 2% goat serum in PBS; α-chicken-Alexa568, #A11041, Thermo Fisher Scientific, Waltham, Massachusetts, USA, 1:1,000 in 10% goat serum in PBS) for 2 h at room temperature. Nuclei were stained with DAPI (1 μg/ml; #D9542, Sigma-Aldrich, St. Louis, Missouri, USA) for 30 min at room temperature. Finally, the slices were coverslipped in fluoroshield (#F6937, Sigma-Aldrich, St. Louis, Missouri, USA) and stored at 4°C. Secondary antibodies were tested for specificity and showed no unspecific binding.
Samples were imaged using an inverted confocal Laser Scanning Microscope (LSM 800, Carl Zeiss Microscopy GmbH, Jena, Germany) equipped with the respective filter sets in combination with a 40× objective (Plan-Apochromat 40× /1.4 Oil, Carl Zeiss Microscopy GmbH, Jena, Germany). The motor neurons to be studied were selected as described above (see Section “Motor neuron enriched dissociated cell cultures of the ventral horn”). γH2ax in nuclei of the motor neurons was analyzed with aid of the microscopy image analysis software Imaris x64 (Version 9.3.1, Oxford Instruments, UK). The nuclear surface was segmented using the surface function. By means of the spots function (diameter over 0.2 μm and intensity quality above 19.2) immunosignals of the phospho-histone H2A.X signal were measured to analyze the amount of DNA damage response and thus double strand breaks in the nucleus of motor neurons (Dumur et al., 2019). At least 35 cell nuclei from at least five independent experiments were analyzed per genotype.
GSH/GSSG assay
For measurement of the amount of GSH and GSSG in wild-type and Wobbler spinal cord (p40) the GSH/GSSG detection assay kit (#ab138881, Abcam, Cambridge, UK) was used according to manufacturer’s protocol. In brief, spinal cord tissue was lysed and homogenized in extraction buffer (#ab65349, Abcam, Cambridge, UK), centrifuged for 15 min at 4°C at 12,700 rpm and the supernatant was collected. For measurement of reduced glutathione (GSH) and total glutathione (GSH+GSSG), 50 μl of the samples were plated in duplicates in a 96-well plate suitable for fluorescence measurements. For GSH detection, 50 μl of GSH assay mixture were added to samples, for total glutathione (GSH+GSSG), 50 μl of total glutathione assay mixture were added to samples. After incubation at room temperature for 15 min protected from light, the fluorescence was measured at 490 nm excitation and 520 nm emission with a TriStar2 Multimode Reader LB 942 (Berthold Technologies GmbH and Co. KG, Bad Wildbad, Germany). Twelve animals per genotype were used.
Statistical analysis
Statistical data analysis was performed with Prism 7.0 (GraphPad Inc., La Jolla, California, USA). Data represent mean values of at least three to seven independent experiments ± standard deviation (SD) or standard error of the mean (SEM). Kolmogorov-Smirnov normality test was used to confirm normal distribution. Data were tested for significance using Student’s t test or two-way ANOVA with a Sidak post-hoc test. Results with p < 0.05 were considered statistically significant.
Results
Increased expression of DNA damage response proteins—p53bp1 and γH2ax in spinal cord of symptomatic Wobbler mice
The initial question was whether an increased amount of ROS in the cervical part of the spinal cord of Wobbler mice at stable clinical stage observed in previous studies (Röderer et al., 2018) has a harmful effect on DNA by inducing the formation of toxic DNA DSBs. To get further insights into the cellular response to DNA DSB formation in cervical spinal cord of wild-type and Wobbler mice, we subsequently investigated the expression on protein- and mRNA-levels of two described participants in the response to DNA DSB formation—p53bp1 and γH2ax (Schultz et al., 2000).
For investigation of the amount of p53bp1 and γH2ax protein, frozen sections were prepared, followed by immunofluorescence staining for the appropriate marker (Figures 1A,B). Thereby, a significantly higher relative signal intensity of both investigated DNA damage response proteins was observed in cervical spinal cord sections of Wobbler animals (Figure 1C). Noticeably, very large nuclei with a diameter of about 20 μm predominantly showed a fluorescent signal of p53bp1 or γH2ax staining (Figure 1A, enlarged images on the right). In addition, the mRNA expression of Tp53bp1 and H2ax was analyzed by qPCR in cervical spinal cord samples. Concordantly, a significant upregulation of both mRNA level could be detected in Wobbler compared to wild-type spinal cord samples (Figure 1D).
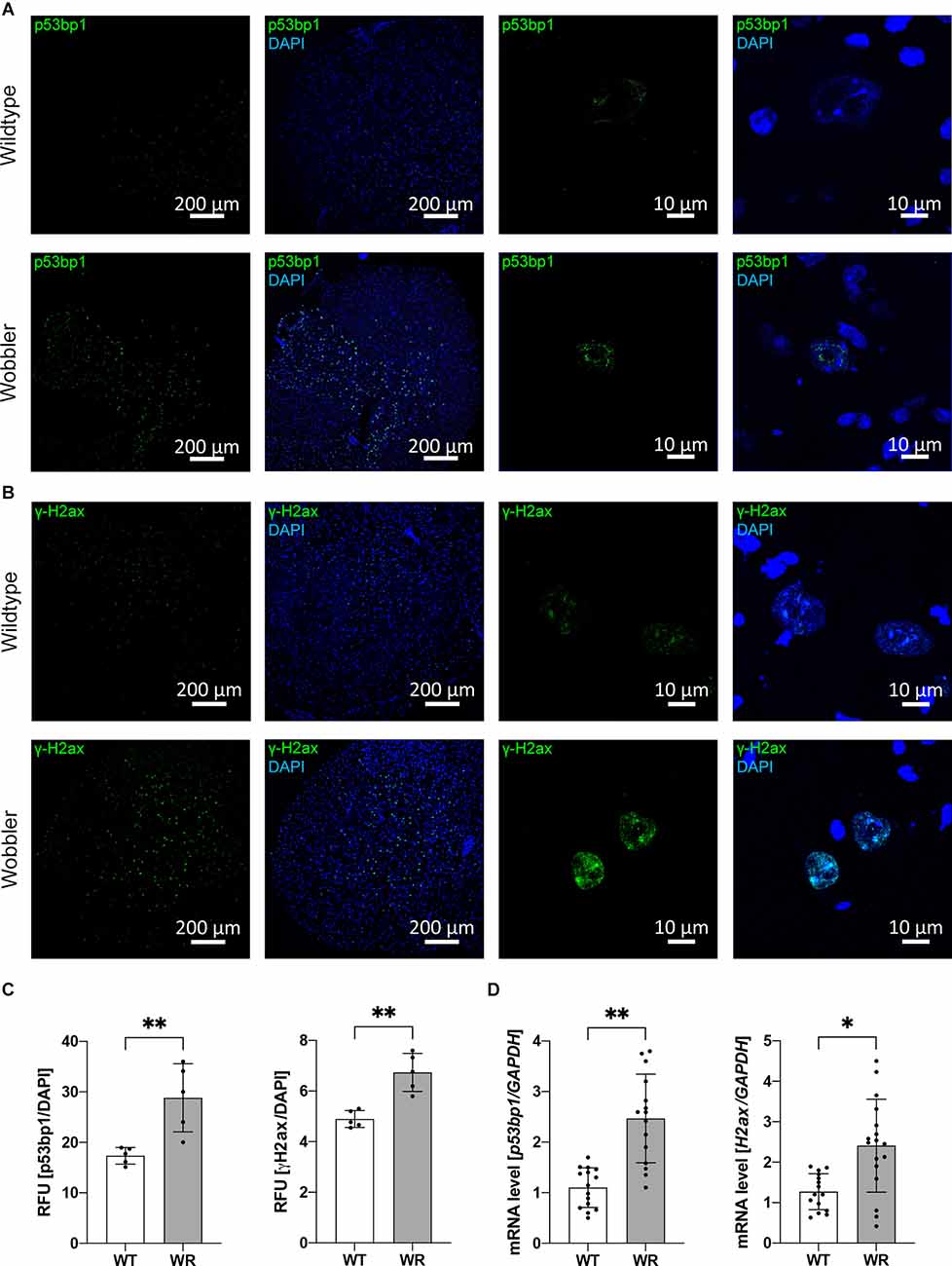
Figure 1. Increased expression of DNA damage response proteins, p53bp1 and γH2ax, in cervical Wobbler spinal cord. (A,B) Exemplary immunofluorescence staining of p53bp1 (green, A) and γH2ax (green, B) and DAPI (blue) in cryo-sections of cervical wild-type and Wobbler spinal cord (p40). (C) Relative fluorescence units of p53bp1 and γH2ax in cryo-sections of wild-type and Wobbler spinal cords (p40). ImageJ 1.53f51 (National Institute of Health, USA) was used to analyze the fluorescence intensity. Intensity of DAPI channel was used for normalization. Data are provided as means ± SD including individual data points. Data were tested for significance using Student’s t-test. Significant differences are indicated by **p < 0.01, n = 5. (D) Tp53bp1- and H2ax-mRNA expression level in wild-type and Wobbler mice at p40. The qPCR was performed using five samples for each genotype. For relative quantification of p53bp1-, and H2ax-mRNA expression, the 2−ΔΔCt method was conducted using the housekeeping gene GAPDH for normalization; data are provided as means ± SD including individual data points. Data were tested for significance using Student’s t-test. Significant differences are indicated by *p < 0.05, **p < 0.01, n = 5.
Elevated ROS levels and γH2ax spots in dissociated motor neurons of symptomatic Wobbler mice
In order to investigate the ROS levels in motor neurons in vitro without exogenous influences of the naturally occurring micromilieu, the spinal cord of 40-days old Wobbler and wild-type mice was isolated, and the cells were cultivated in dissociated cell culture for 10 days in vitro (div).
To investigate intracellular ROS levels in isolated motor neurons, fluorescence intensity of DHE was measured in 20 motor neurons from four different cultures (p40+10 div) per genotype (Figure 2A). On average there was no autofluorescence in cell soma of wild-type and Wobbler motor neurons without DHE (Figure 2B, time period: −10 min to 0 min). After exchange to DHE-containing medium (Figure 2B, timepoint 0 min), the average intensity of red fluorescence in the cell soma of Wobbler motor neurons showed a significant progressive increase compared to the cell soma of wild-type motor neurons. After an hour of observation, the average fluorescence intensity in soma of wild-type motor neurons indicated an increase to 1.87 in relative comparison to the baseline, while Wobbler motor neurons showed a relative intensity of 6.3 (Figure 2B). Thus, Wobbler motor neurons were characterized by a strong and significant increase in DHE associated fluorescence 1 h after DHE addition in vitro compared to wild-type motor neurons. Thus, it can be concluded that motor neurons from Wobbler mice experience a significantly increased ROS level in vitro compared to wild-type.
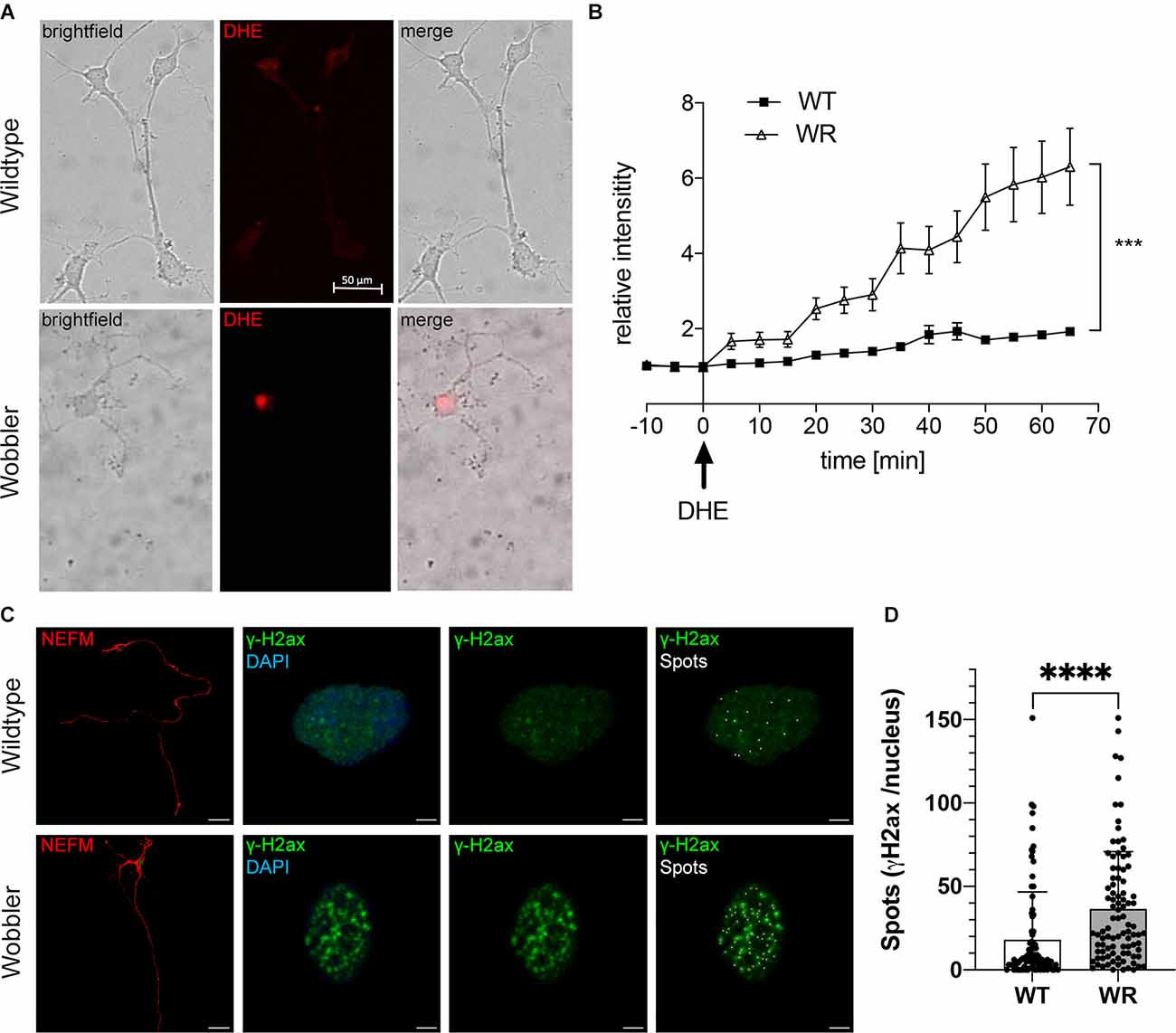
Figure 2. Elevated ROS levels in motor neurons of Wobbler mice correlate with increased abundance of the DNA damage response protein γH2ax. (A) Exemplary images of dissociated motor neurons from wild-type and Wobbler animals (p40+10div) 65 min after DHE (red fluorescence) addition and live cell imaging. Live-cell imaging was performed using a spinning disc confocal microscope with a 20× objective. The fluorescence signals of the cells were documented every 5 min for 1 h in DHE-including medium, 37°C and 5% CO2. Scale bar = 50 μm. (B) Relative red fluorescence intensity in five dissociated motor neuronal cell cultures from p40 wild-type and Wobbler mice documented in 5 min intervals of 20 cells per genotype. At timepoint 0 the normal medium was exchanged for DHE including medium. ImageJ 1.53f51 (National Institute of Health, USA) was used to analyze the average red fluorescence intensity within the soma of a motor neuron, while motor neurons themselves were detected in transmitted-light channel. A mean value of background fluorescence intensity without DHE addition was used for normalization. Data are provided as means ± SEM. Data were tested for significance using two-way ANOVA with a Sidak post-hoc test. Significant differences are indicated by ***p < 0.001, n = 20 cells from five dissociated cultures per genotype. (C) Exemplary images of wild-type and Wobbler motor neurons (p40+10div) stained with anti-NEFM antibody (red), anti-phospho-Histone H2A.X (green) and DAPI (blue). The overview images (1st row) show an exemplary motor neuron, with rows 2–4 focusing on the nucleus (blue) of the exemplary cell to see the H2ax signals (green) and spots (white) created using Imaris 9.3.1. Pictures were taken with a confocal fluorescent microscope equipped with a 40× oil objective. Scale bar (1st row) = 30 μm. Scale bar (2nd–4th raw) = 2 μm. (D) Detected spots of γH2ax staining in wild-type and Wobbler motor neuronal cell nuclei. Wobbler mice show a significantly higher number of spots with an average of 36.56 while wild-type motor neurons have an average number of spots of 18.09. Data are provided as means ± SD including individual data points. Data were tested for significance using Student’s t-test. Significant differences are indicated by ****p < 0.0001, n = 90–94 nuclei per genotype of a least five experiments.
To test whether elevated ROS levels in Wobbler motor neurons cause increased DNA damage, the protein γH2ax was investigated by immunofluorescent staining in dissociated motor neuronal cultures as a marker for the formation of DNA DSBs. With the aid of Imaris spots function the punctate signals of γH2ax were converted into spots (Figure 2C). Comparing γH2ax signals in wild-type and Wobbler motor neurons from 40-days old mice (p40+10 div), striking differences emerged. Nuclei of Wobbler motor neurons displayed a significantly higher number of spots than motor neurons of wild-type mice and thus more DSBs (Figures 2C,D). Based on these results, it can be hypothesized that the increased ROS levels observed in Wobbler animals give rise to the increased DNA damage and further to accumulation of γH2ax signal as a marker of DNA damage and repair.
Dysregulated mediators of ROS detoxification mechanisms in Wobbler motor neurons
As we detected increased DHE intensity in Wobbler motor neurons in vitro, indicating increased ROS levels, two hypotheses can be raised: (i) do motor neurons of Wobbler animals exhibit increased ROS production; or (ii) are there deficits in ROS detoxification? Therefore, further focus was placed on the analysis of important intracellular mediators of ROS detoxification mechanisms. For this purpose, mRNA and protein expression of several molecules for ROS detoxification were analyzed in cervical spinal cord of 40-days old wild-type and Wobbler mice (Figure 3). Evaluation of the expression of superoxide anion detoxifying proteins SOD1 and SOD2 showed no alteration in protein expression, although a significantly decreased mRNA level of SOD2 was detected in the cervical spinal cord of Wobbler mice (Figure 3A). Detoxification of hydrogen peroxide, the product of SOD, is carried out by catalase or the glutathione system. Here, the analysis of the mRNA expression of catalase revealed a strongly significant reduction of catalase, which, however, was not reflected at the protein level (Figure 3B). In contrast, the expression of GPX4, an important member of the glutathione system especially for motor neurons, was significantly increased at both the mRNA and protein level in cervical spinal cord of Wobbler mice (Figure 3C).
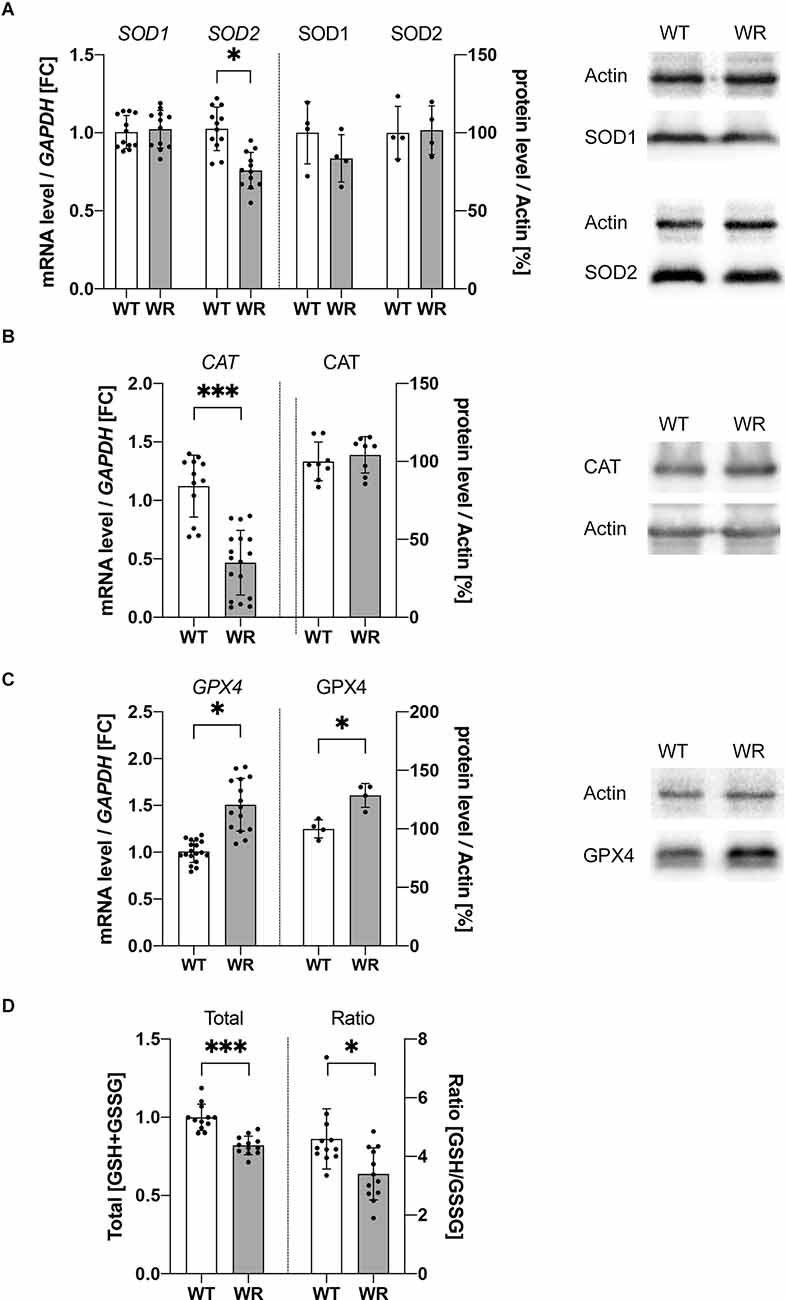
Figure 3. Dysregulated expression of ROS detoxifying molecules in cervical Wobbler spinal cord. (A–C) mRNA and protein expression levels of SOD1 and 2 (A), CAT (B), and GPX4 (C) at stable clinical stage (p40) in cervical spinal cord of wild-type (WT) and Wobbler (WR) mouse. For relative quantification of mRNA level, the 2−ΔΔCt method was conducted using GAPDH for normalization. For semiquantitative analysis of protein expression levels, arithmetic analysis of the band intensity with ImageJ 1.53f51 (National Institute of Health, USA) software was used. Band intensities of proteins of interest were normalized to the housekeeper actin. Normalized protein levels were compared between different genotypes. All data are presented as the mean values ± SD including individual data points, and Student’s t-test was performed for significance testing between WT and WR. Significant differences are indicated by *p < 0.05, ***p < 0.001, n(qPCR) = 5; n(WB) = 4–8. (D) The total amount of reduced and oxidized glutathione was measured in the spinal cord of 40-days old wild-type and Wobbler mice using a GSH/GSSG detection assay kit. Total [GSH+GSSG] was normalized to WT. Using the same GSH/GSSG detection assay kit a significantly decreased ratio of [GSH/GSSG] could be detected in Wobbler spinal cord compared to wild-type spinal cord. All data are presented as the mean values ± SD including individual data points, and Student’s t-test was performed for significance testing between different genotypes. Significant differences are indicated by *p < 0.05, ***p < 0.001, n = 12.
After becoming apparent that an increased expression of glutathione-consuming peroxidase was present in the spinal cord of Wobbler mice, investigations concerning the cofactor glutathione were performed. Therefore, the total amount and the ratio of oxidized (GSSG) and reduced glutathione (GSH) in Wobbler and wild-type cervical spinal cord was quantified. Wobbler spinal cord showed a significantly decreased amount of total glutathione (GSH+GSSG), in combination with a significantly decreased ratio of GSH/GSSG (Figure 3D) indicating deficits in ROS detoxification and oxidative stress in this tissue.
Effects of ROS scavengers N-Acetyl-L-Cysteine (NAC), glutathione ethyl ester (GSHe), and Mito-TEMPO on Wobbler motor neurons in vitro
After observing a significant difference in the expression of several key ROS detoxification representatives including an altered ratio of the redox partners GSH/GSSG in the Wobbler spinal cord, dissociated motor neurons of 40-days old wild-type and Wobbler mice were cultured and treated with the ROS scavengers N-acetyl-L-cysteine (NAC), glutathione ethyl ester (GSHe), and Mito-TEMPO after a defined time period (see above). Here, immunofluorescence staining for γH2ax as a DNA double-strand-break-repair-related protein was analyzed and the intensity of the signals was evaluated using the spot function of Imaris 9.3.1 (Figures 4A,B).
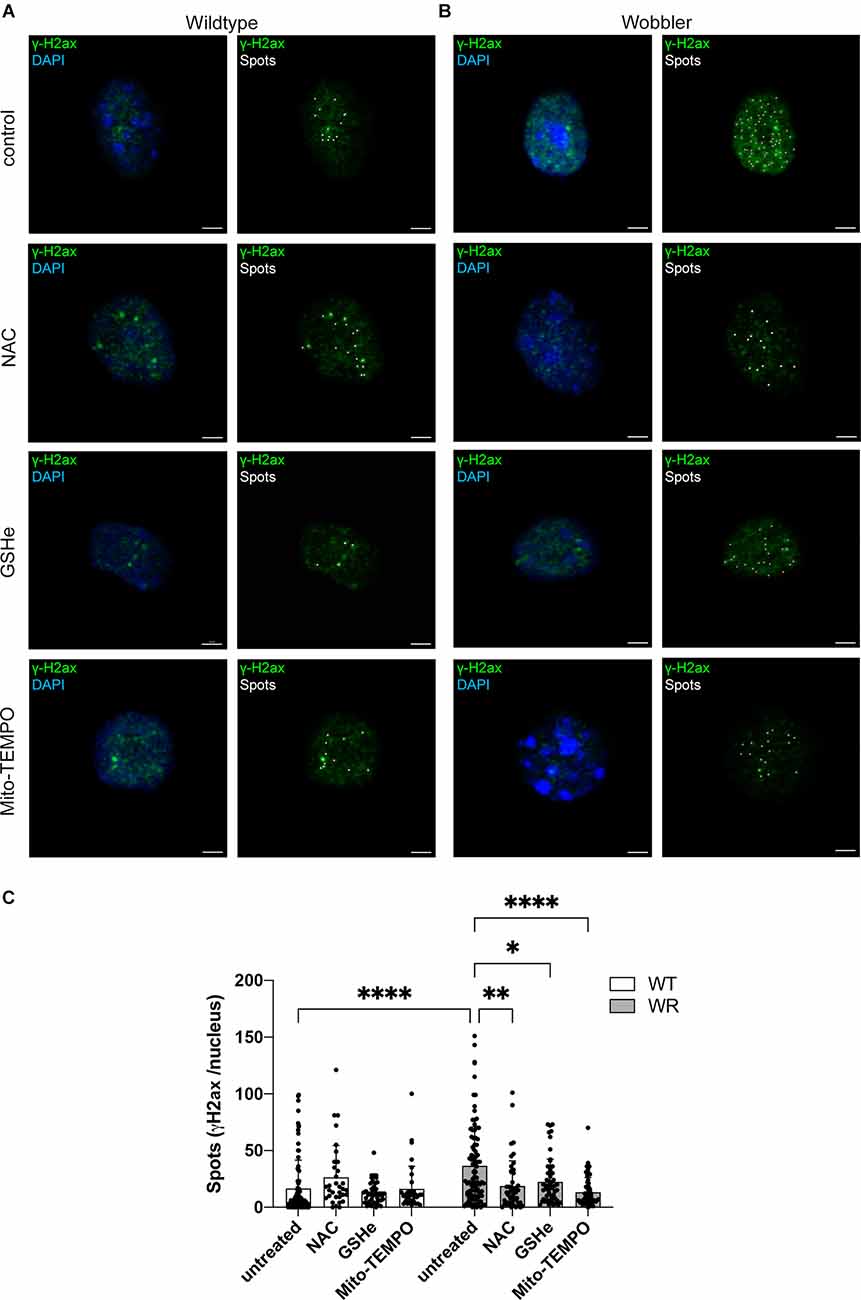
Figure 4. Altered γH2ax Spots in Wobbler motor neuronal nuclei after treatment with ROS scavengers NAC, GSHe, and Mito-TEMPO. (A,B) Exemplary images of untreated control and NAC-, GSHe-, and Mito-TEMPO-treated wild-type (A) and Wobbler (B) motor neurons. Nuclei were stained with DAPI (blue) and DNA damage response protein γH2ax (green). Pictures were taken with a confocal fluorescent microscope equipped with a 40× oil objective. Amount of γH2ax and thus double strand breaks was analyzed with the spots function (white) of Imaris 9.3.1. Scale bar = 2 μm. (C) Evaluation of the spots number of untreated control and NAC-, GSHe- and Mito-TEMPO-treated wild-type (WT) and Wobbler (WR) motor neurons at stable clinical phase (p40+10div). Data are provided as means ± SD including individual data points. Data were tested for significance using two-way ANOVA with a Sidak post-hoc test. Significant differences are indicated by *p < 0.05, **p < 0.01, ****p < 0.0001, n = 35–55 nuclei per genotype of a least five experiments.
Considering the evaluated spots in motor neurons from wild-type animals, no significant difference in the number of spots after treatment with any of applied ROS scavenger was observed within this genotype (Figure 4C). In contrast, looking at the influence of ROS scavenger treatment within Wobbler motor neurons, it was obvious that all tested ROS scavengers displayed a significant decrease in the number of detected γH2ax spots (Figure 4C).
Discussion
ALS is a highly heterogenous disease considering the underlying pathology. To date, no significant therapeutic option exists that could stop the progressive loss of motor neurons over a prolonged period of time (Brown and Al-Chalabi, 2017). However, it is known that the majority of all ALS patients exhibit uniform phenotypes reflected in signs of oxidative stress (Tam et al., 2019), making it an important factor in the pathogenesis of ALS (Matschke et al., 2019; Singh et al., 2019). Protein and lipid peroxidation as well as oxidative DNA damage could be observed in the spinal cord of ALS patients (Ferrante et al., 1997; Shibata et al., 2001). Previous studies also indicate changes in the ROS homeostasis in an animal model for ALS, the Wobbler mouse (Moser et al., 2013; Röderer et al., 2018). Thus, own previous work identified increased ROS level in the cervical spinal cord of the Wobbler mouse (Röderer et al., 2018). A neuroprotective and symptom-improving effect of several ROS scavengers on the Wobbler mouse has already been described (Henderson et al., 1996; Abe et al., 1997; Ikeda et al., 2015; Zwilling et al., 2020). However, an accurate analysis of the ROS homeostasis, especially in Wobbler mouse motor neurons, was still lacking. The present study was conceptualized to further substantiate and concretize existing findings and to be able to assess the consequences of an altered ROS homeostasis for single motor neurons.
The DNA of the cell is highly susceptible to changes in the intracellular ROS level, and it is essential for cell survival to detect and effectively repair oxidative damage (Ba and Boldogh, 2018). Base oxidation, DNA single-strand-breaks (SSBs) and consecutive DNA DSBs are a result of increased exposure of DNA to elevated ROS concentrations with a consequence of increased overall DNA instability (Kobayashi et al., 2017; Ba and Boldogh, 2018; Qi et al., 2019). Two central biomarkers for detecting DNA DSBs have been described in the past: γH2ax and p53bp1 (Schultz et al., 2000; Mah et al., 2010; Kannan et al., 2018; Williamson et al., 2020). In the presence of DNA damage one of the earliest key factors of the DNA repair Histone H2ax is phosphorylated to γH2ax thereby allowing the assembly of DNA repair-related proteins to DNA (Kobayashi, 2004; Kuo and Yang, 2008; Podhorecka et al., 2010). Moreover, hyperphosphorylated p53bp1 attaches to the newly formed DNA DSB and initiates non-homologous end joining (NHEJ) via the recruitment of repair factors, which play a major role especially in non-dividing cells such as neurons (Iyama and Wilson, 2013; Panier and Boulton, 2014).
On mRNA level, we could observe a significantly increased expression of both p53bp1 and unphosphorylated H2ax in the cervical spinal cord of the Wobbler mouse from stabilized clinical phase (p40). This observation is verified by evaluation of protein expression of p53bp1 and γH2ax in the cervical spinal cord, where an increased signal of p53bp1 or γH2ax was detected mainly in very large nuclei with a diameter of about 20 μm of the spinal cord. Although a statement on the colocalization of p53bp1 and γH2ax is not provided by our experimental setup, signals of both investigated proteins are detectable in cells with the same morphology, which can be identified as motor neurons due to their large nucleus. Although elevated phosphorylation of H2ax does not necessarily correlate to increased mRNA expression of H2ax, this result may indicate a functional work flow of DNA DSB detection and accumulation (Zhang et al., 2016). All in all, the increased mRNA expression of both p53bp1 and unphosphorylated H2ax in the spinal cord as well as the increased protein level in cervical spinal cord of Wobbler mice strongly suggest that there is an increased accumulation of DNA DSBs in Wobbler tissue presumably due to oxidation of DNA.
To investigate these results more extensively in motor neuronal cells, dissociated motor neuronal cell cultures were performed. Due to the isolated cultivation of the cells in vitro, the motor neurons do not experience any influence from neighboring cells, like microglia, and thus the endogenous factors of cell homeostasis can be observed (Zwilling et al., 2020). Wobbler motor neurons from stabilized clinical phase (p40+10div) were incubated with dihydroethidium (DHE). DHE diffuses well through cell membranes and reacts most strongly with superoxide anions but weaker also with other ROS species and NO resulting in a red immunofluorescence and thus is a marker for oxidative stress (Owusu-Ansah et al., 2008; Gardiner et al., 2020). Wobbler motor neurons show an almost three times higher increase in fluorescence intensity after 65 min in vitro compared to wild-type motor neurons. This suggests that the increased ROS levels measured in the Wobbler spinal cord by Röderer et al. (2018) is also found in isolated motor neurons.
As a consequence of increased ROS levels, we next examined the occurrence of DNA damage in isolated motor neuronal cultures. Therefore, we measured the immunofluorescence signals of γH2ax in the nucleus of dissociated motor neurons of Wobbler and wild-type mice. Wobbler motor neurons in dissociated cell culture displayed a significantly higher number of γH2ax foci compared to wild-type. This result is interesting in two aspects: first, it provides further evidence of increased DNA DSB abundance in motor neurons of Wobbler mice and confirms the overexpression of H2ax at the mRNA level and increased γH2ax fluorescence signal intensity in spinal cord sections of Wobbler mice. Second, our observation provides evidence for the influence of endogenous factors, like ROS formation, in the emergence of toxic DNA DSBs in motor neurons. Here, a link can be drawn to the increased ROS levels in dissociated motor neuronal cells of Wobbler mice. These results may be indicative of increased oxidative stress in Wobbler spinal cord, particularly in motor neurons, and its dependence on endogenous factors.
The combination of increased expression of both p53bp1 and H2ax/γH2ax generally indicates an increased number of DNA DSBs (Kannan et al., 2018). In this study, we investigated p40 mice, as previous studies from our group have shown increased ROS levels in the spinal cord only at this stage. Based on the unchanged ROS level of p0 and p20 Wobbler animals (Röderer et al., 2018), we assume that ROS-related DNA damage is not yet detectible at the time points of the development earlier than p40. However, further investigations on the time-resolved development of the Wobbler phenotype are necessary to clarify the relation between ROS induction and DNA damage in ALS.
Various studies have already shown an increased activation of DNA damage response in human ALS and suggest that the accumulation of DNA damage response markers may play a role in premature cell death (Lopez-Gonzalez et al., 2016; Farg et al., 2017; Kim et al., 2020). Among others, the analysis of Kim et al. (2020) in postmortem tissue samples of ALS patients and induced pluripotent stem cells interestingly showed an accumulation of DNA damage like single-stranded DNA and 8-oxoguanin as well as an increased recruitment of the DNA damage and repair components like ATM Serine/Threonine Kinase (ATM) and Breast Cancer Anti-Estrogen Resistance 1 Cas Family Scaffold Protein (BCRC1). Similar observations of increased numbers of DNA damage, like DNA double strand breaks, were shown in different ALS subgroups. Thus, an increased number of p53bp1 and γH2ax expression was shown to be associated with specific C9Orf72 variants (Farg et al., 2017). This increased number of DNA damage response proteins was also associated with increased oxidative stress in an iPSC model with C9Orf72 variants (Lopez-Gonzalez et al., 2016). Konopka et al. (2020) demonstrated that TDP-43 ALS-associated mutations lead to a reduced activity of NHEJ, which in consequence leads to increased DNA damage. Therefore, it is important to emphasize that despite different genetic backgrounds, similar effects on genomic DNA have been described in the different ALS models. Histone 2ax has also been shown to play a key role, in both, the registration of increased ROS levels and in the initiation of various DNA damage response components (Kuo and Yang, 2008; Podhorecka et al., 2010; Barral et al., 2014; Merighi et al., 2021). Interestingly, H2ax-knock out mice exhibit dysfunctional motor characteristics, such as an impaired overall motor activity as well as impaired motor balance, among others (Weyemi et al., 2018). Thus, the increased expression and phosphorylation of H2AX/γH2ax in Wobbler mice observed in this study indicates a response to existing oxidative stress to trigger a mechanism that, as a consequence, should counteract the loss of function of motor neurons.
Maintaining ROS-homeostasis is essential to maintain normal cell function. Therefore, the cell is equipped with various antioxidant mechanisms to scavenge ROS (Lagouge and Larsson, 2013; Pourahmad et al., 2016; Matschke et al., 2019). Physiologically occurring superoxide radicals are converted to hydrogen peroxide by two intracellular superoxide dismutases (SODs), cytosolic Cu-Zn SOD (SOD1) and mitochondrial MnSOD (SOD2; van Remmen et al., 2003; Lagouge and Larsson, 2013). Another important factor for neuroprotection is the detoxification of hydrogen peroxide produced by SODs (Ren et al., 2017; Matschke et al., 2019). The highly reactive hydrogen peroxide is detoxified by catalases and the glutathione system (Ren et al., 2017; Matschke et al., 2019).
The expression and activity level of SODs plays an important role in balancing the concentration of ROS (Miao and St Clair, 2009). With regard to human ALS, a primary mutation in the SOD1 gene can be found in nearly 15% of all European fALS and about 1% of European sALS cases (Zou et al., 2017; Mejzini et al., 2019). Interestingly, our results show no difference in the SOD1 expression between Wobbler and wild-type spinal cord. Although SOD2 mRNA is significantly downregulated in Wobbler animals, there is no effect on protein expression in symptomatic stage Wobbler tissue. SOD2 plays a role in the mitochondrial antioxidant system and a reduction in SOD2 activity has already been associated with increased oxidative levels and cancer potential (van Remmen et al., 2003; Celotto et al., 2012). The question arises whether protein expression of SOD2 could also be found decreased at a later stage of age leading to mitochondrial oxidative stress and consequently to motor neuronal death. However, as we detect decreased SOD2 mRNA and no deregulation of the protein level at a stage when all symptoms of ALS are already developed in the Wobbler animals and motoneuronal dysfunctionalities are already apparent, an underlying influence of a possible under-expression of the SOD2 protein at later age stages can be excluded.
Our results demonstrated a strong downregulation of CAT mRNA expression in cervical Wobbler spinal cord again without affecting protein expression. It should be emphasized that catalase activity is enhanced by ATM and p53, among others (Glorieux et al., 2015). Based on our results regarding the increased amount of γH2ax and p53bp1 in Wobbler, it is highly likely that catalase could receive a strong induction of its activity by activated ATM and p53.
Another important component in the detoxification of hydrogen peroxide are glutathione peroxidases, which consist of a group of seven isoforms (GPX 1–7). Of particular interest for our study was the increased expression of the ubiquitous GPX4 isoform, as it has been described to be strongly neuroprotective in vitro and in vivo (Seiler et al., 2008). Moreover, GPX4 is the only glutathione peroxidase that accepts protein thiol groups as reducing substrates under GSH deficiency (Seiler et al., 2008). Here, we show a significantly increased GPX4 expression at both, the mRNA and protein level in the spinal cord of p40 Wobbler mice. This may be an attempt of the cell to compensate for the reduced amount of GSH and reduced GSH/GSSG ratio that we have demonstrated, since GPX4 is the only isoform that does not rely on GSH as a reactant (Seiler et al., 2008).
Considering the general increase in ROS levels throughout the spinal cord and the increased ROS levels in isolated motor neurons, it is possible to hypothesize that the mechanisms of ROS-detoxification are not effective enough to detoxify ROS and thus ensure neuroprotection. It is striking here that catalase, in contrast to glutathione peroxidases, is not increased in expression at the protein level, although it receives an expression stimulus via increased ROS concentration. The non-increased expression of catalase is even more remarkable because it has been shown in the past that 50% of hydrogen peroxides are detoxified by catalase, whereas the other half are detoxified by glutathione peroxidases (Sepasi Tehrani and Moosavi-Movahedi, 2018). Catalases in Wobbler mice should receive an additional expression stimulus from p53. Thus, increased expression of p53 has been described in previous publications in the Wobbler mouse and is considered an important inducer of catalase expression (Eve et al., 2007; Kang et al., 2013).
Furthermore, we also analyzed the relative and absolute concentrations of oxidized and reduced glutathione, which is an important redox partner for glutathione peroxidase in particular. Glutathione (GSH), a necessary cofactor for GPX function, is synthetized by the cytosolic y-glutamylcysteine ligase and GSH synthetase in virtually all cell types with the liver as the major producer and exporter (Wu et al., 2004; Chi et al., 2007; Aoyama and Nakaki, 2013). Neurons do not have an uptake mechanism for GSH and rely on the uptake of cysteine and thus their own synthesis of GSH to maintain adequate concentrations (Johnson et al., 2012; Aoyama and Nakaki, 2013; Rae and Williams, 2017). GSH is involved in maintaining reducing conditions in all cells as an important antioxidant (Townsend et al., 2003). In case of reduction, the thiol-group of the cysteine is oxidized, and two glutathione molecules react to glutathione disulfide (GSSG). GSH/GSSG is the major redox couple in neurons (Wu et al., 2004; Ghosh et al., 2014). In p40 Wobbler spinal cord we measured a significantly decreased level of total glutathione [GSH+GSSG] as well as a significantly decreased ratio [GSH/GSSG]. This reconfirms the underlying oxidative stress in cervical Wobbler spinal cord, with a reduced amount of total glutathione and a shifted ratio to the oxidized form. This reduced glutathione level also suggests that there is not enough GSH available as a coenzyme for proper glutathione peroxidase function. On the one hand, GPX4 could be reflectively upregulated to compensate the lacking glutathione, and on the other hand the assumption arises that lacking reduced glutathione nevertheless leads to a reduced efficiency of glutathione peroxidase and thus to an increased ROS concentration.
Based on the abnormalities we observed in the GSH-dependent ROS detoxification system, we further specified these results using glutathione-based ROS scavengers. Therefore, we treated dissociated motor neurons of wild-type and Wobbler spinal cord (p40+10div) with N-acetyl-L-cysteine and glutathione ethyl ester (GSHe) for a defined period of time and measured the number of γH2ax spots and thus the DNA damage response afterwards.
N-acetyl-L-cysteine (NAC) is a well-described antioxidant. It has direct antioxidant effects via the modulation of the transcription factor NF-KB (Staal et al., 1995; Oka et al., 2000) and indirect antioxidant effects as a precursor of cysteine and thus GSH (Henderson et al., 1996; Aldini et al., 2018). After NAC is transported into the cells, it is deacetylated to cysteine (Aldini et al., 2018). As neurons rely on the uptake of cysteine for GSH production and thus, cysteine is the rate-limiting factor in the glutathione synthesis, NAC can replenish the GSH content (Rushworth and Megson, 2014). In a prior study the degeneration of Wobbler mice motor neurons could significantly be decreased by treating the mice directly with NAC (Henderson et al., 1996). Furthermore, a ROS-reducing effect of NAC in primary neuronal cells has already been shown (Wu et al., 2018). In this study, we observed a significantly decreased number of γH2ax spots and thus DNA damage in Wobbler motor neurons after treatment with NAC. Assuming an increased ROS-level and a decreased level of glutathione, especially the reduced form, as an important antioxidant in Wobbler motor neurons, it can be concluded that NAC, as a precursor of glutathione, can replenish the GSH content and thus reduce oxidative stress in Wobbler motor neurons resulting in less oxidative DNA damage. Furthermore, direct effects of NAC, through its influence on gene modulation, could also lead to a decrease in γH2ax spots.
Treating motor neurons with GSHe also lead to a reduced number of γH2ax spots and thus, DNA damage in Wobbler motor neurons (p40+10div). GSHe is a ROS scavenger which increases intracellular GSH and bypasses the need of GSH synthesis. GSHe is cell-permeable and readily de-esterified by intracellular esterases (Anderson et al., 1985; Dringen et al., 1999; Chen et al., 2018). In prior studies, a neuroprotective effect of GSHe-treatment on ischemic damaged brain and spinal cord injuries in rats has already been shown (Anderson et al., 2004; Guízar-Sahagún et al., 2005). This neuroprotective effect is associated with a reduction in ROS levels (Ross et al., 2012; Rush et al., 2012). GSHe treatment of ALS mouse models expressing the ALS-associated SOD1 mutation delayed early degenerative changes in motor neurons and reduced oxidative stress (Winkler et al., 2014).
Thus, our results confirm the hypothesis that increased ROS levels in motor neurons of Wobbler animals are associated with impaired ROS detoxification by the glutathione system.
Finally, it remained to consider whether motor neurons from Wobbler animals also show increased ROS production. As mitochondria are physiologically the major producers of intracellular ROS, mitochondrial dysfunction can lead to many cell damaging processes including an excessive production of this harmful species (Smith et al., 2019). Since we demonstrated in a previous study that mitochondria in Wobbler motor neurons have abnormal morphology (Stein et al., 2021) and mitochondrial malfunction is known to be present in Wobbler animals (Xu et al., 2001; Dave et al., 2003; Santoro et al., 2004; Gonzalez Deniselle et al., 2012), it is tempting to assume an increased mitochondrial ROS production in a Wobbler phenotype. Therefore, an additional ROS scavenger, Mito-TEMPO, was used to specifically scavenge mitochondrially produced ROS. Mito-TEMPO consists of the antioxidant piperidine nitroxide (TEMPO) and the lipophilic cation triphenylphosphonium (TPP+). Due to the combination with TPP as a membrane-permeant cation that is accumulated several hundred-fold within mitochondria driven by the membrane potential the antioxidant TEMPO as a SOD mimetic can accumulate within mitochondria (Trnka et al., 2008). Therefore, it helps protect against oxidative damage to the mitochondria and has superoxide and alkyl radical scavenging properties (Trnka et al., 2008; Du et al., 2017). Mito-TEMPO has already been shown to have protective effects on neuronal cells in the neurodegenerative disease Alzheimer’s disease, in which oxidative stress also plays an important role (Lu et al., 2015; Hu and Li, 2016). When primary cortical neurons were treated in cell culture with Mito-TEMPO, a reduction in ROS concentration could also be observed (Zhou et al., 2015). More specifically, in primary cultured neurons, Aß-promoted mitochondrial superoxide production and neuronal lipid peroxidation were significantly suppressed by Mito-TEMPO (Hu and Li, 2016). In this study, we demonstrated that treating Wobbler motor neurons with Mito-TEMPO also leads to a significantly decreased number of γH2ax foci and thus DNA damage in motor neurons. Hence, counteracting mitochondrial ROS production seems to be an important part in reducing DNA damage in Wobbler spinal cord. In conclusion, not only glutathione-based but also mitochondria-targeted ROS scavengers might be involved in reducing oxidative stress and thus the extent of DNA damage in the Wobbler spinal cord.
Using ROS scavengers, we were able to detect a reduced number of DNA damage response proteins in the nuclei of dissociated motor neurons from Wobbler mice. Thus, we hypothesize that scavenging mitochondria-produced ROS or enhancing the GSH system with GSH precursors or derivatives may reduce oxidative stress in motor neuron cells. As a result, the cellular ROS level and thus oxidation of DNA bases are reduced. This would lead to decreased number of SSBs and reduce the probability for an occurrence of DSBs resulting from the accumulation of SSBs. Furthermore, DSBs were described in the cell to trigger apoptosis (Ghardi et al., 2012). Since several studies revealed a positive effect of ROS scavengers against apoptotic cell death, it is probable to hypothesize that ROS scavengers not only reduce the number of toxic DSBs in the post-mitotic cell, but also reduce apoptotic cell death. However, this correlation still needs to be investigated in more detail in the ALS model of Wobbler mouse.
Conclusion
ALS is the most common motor neuronal disease, which has been challenging both basic and clinical research and clinical therapy for decades. In this study, we present for the first-time evidence for increased DNA damage of symptomatic Wobbler mice, which might be a consequence of exposure to oxidative stress. In dissociated motor neuronal cultures, Wobbler motor neurons exhibit not only increased ROS levels, but also increased numbers of γH2ax foci and thus DNA DSBs, which could be counteracted by using glutathione precursors, derivatives, and a mitochondria-specific ROS scavenger Mito-TEMPO. The beneficial effect of Mito-TEMPO observed in our study also emphasizes the role of mitochondria in the pathology of increased ROS levels. In addition, a decreased amount of total glutathione and a decreased ratio of GSH/GSSG were detected in Wobbler spinal cord tissue, verifying the presence of oxidative stress presumably due to a fundamental glutathione deficiency.
Overall, this work reveals several pathologies in the Wobbler spinal cord and especially motor neurons, all of which are closely related to and might cause progressive, premature cell death. This substantiates previous work on the Wobbler mouse, which only described the benefit of antioxidant molecules in vivo without showing a possible link to oxidative damage to cellular components. Our work here focuses on the damage of DNA by oxidative stress in motoneurons and at the same time presents that it is a reversible process.
Our results indicate that a focused intervention in one pathomechanism might be insufficient in ALS therapy, as multiple systems are affected, so that multimodal therapy will be necessary to prolong the average lifespan of motor neurons and thus slow down the progression of the disease.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (LANUV).
Author Contributions
VM designed, conceptualized, and supervised the research. MJ, FJ, HC, DS, and VM performed all experiments with support of VB and KW (IF visualization and analysis). MJ, FJ, HC, and VM analyzed, validated, and visualized the results. CT and JM contributed expertise (IF) and intellectual feedback. MJ, FJ, and VM wrote the original manuscript draft. CT and JM critically revised and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
KW is supported by the German Research Foundation (FOR 2848 and Germany’s Excellence Strategy—EXC 2033—390677874—RESOLV). This study was supported by German Academic Exchange Service and International Graduate School of Neuroscience (IGSN), Ruhr University Bochum to HC. JM is supported by the Federal Ministry of Education and Research (BMBF, 02NUK061B).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We gratefully acknowledge A. Lodwig, C. Grzelak, and A. Harbecke for technical assistance, as well as A. Lenz and T. Kolek-Vahstall for secretarial work. We acknowledge support by the Open Access Publication Funds of the Ruhr-Universität Bochum.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fncel.2022.963169/full#supplementary-material.
References
Abe, K., Itoyama, Y., Sobue, G., Tsuji, S., Aoki, M., Doyu, M., et al. (2014). Confirmatory double-blind, parallel-group, placebo-controlled study of efficacy and safety of edaravone (MCI-186) in amyotrophic lateral sclerosis patients. Amyotroph. Lateral Scler. Frontotemporal Degener. 15, 610–617. doi: 10.3109/21678421.2014.959024
Abe, K., Morita, S., Kikuchi, T., and Itoyama, Y. (1997). Protective effect of a novel free radical scavenger, OPC-14117, on wobbler mouse motor neuron disease. J. Neurosci. Res. 48, 63–70. doi: 10.1002/(SICI)1097-4547(19970401)48:1%3C63::AID-JNR6%3E3.0.CO;2-C
Al-Chalabi, A., Hardiman, O., Kiernan, M. C., Chiò, A., Rix-Brooks, B., and van den Berg, L. H. (2016). Amyotrophic lateral sclerosis: moving towards a new classification system. Lancet Neurol. 15, 1182–1194. doi: 10.1016/S1474-4422(16)30199-5
Aldini, G., Altomare, A., Baron, G., Vistoli, G., Carini, M., Borsani, L., et al. (2018). N-Acetylcysteine as an antioxidant and disulphide breaking agent: the reasons why. Free Radic. Res. 52, 751–762. doi: 10.1080/10715762.2018.1468564
Anderson, M. F., Nilsson, M., Eriksson, P. S., and Sims, N. R. (2004). Glutathione monoethyl ester provides neuroprotection in a rat model of stroke. Neurosci. Lett. 354, 163–165. doi: 10.1016/j.neulet.2003.09.067
Anderson, M. E., Powrie, F., Puri, R. N., and Meister, A. (1985). Glutathione monoethyl ester: preparation, uptake by tissues and conversion to glutathione. Arch. Biochem. Biophys. 239, 538–548. doi: 10.1016/0003-9861(85)90723-4
Andrews, J. M., Gardner, M. B., Wolfgram, F. J., Ellison, G. W., Porter, D. D., and Brandkamp, W. W. (1974). Studies on a murine form of spontaneous lower motor neuron degeneration - the wobbler (wr) mouse. Am. J. Pathol. 76, 63–78.
Aoyama, K., and Nakaki, T. (2013). Impaired glutathione synthesis in neurodegeneration. Int. J. Mol. Sci. 14, 21021–21044. doi: 10.3390/ijms141021021
Ba, X., and Boldogh, I. (2018). 8-Oxoguanine DNA glycosylase 1: beyond repair of the oxidatively modified base lesions. Redox Biol. 14, 669–678. doi: 10.1016/j.redox.2017.11.008
Barral, S., Beltramo, R., Salio, C., Aimar, P., Lossi, L., and Merighi, A. (2014). Phosphorylation of histone H2AX in the mouse brain from development to senescence. Int. J. Mol. Sci. 15, 1554–1573. doi: 10.3390/ijms15011554
Boillée, S., Peschanski, M., and Junier, M.-P. (2003). The wobbler mouse: a neurodegeneration jigsaw puzzle. Mol. Neurobiol. 28, 65–106. doi: 10.1385/MN:28:1:65
Brieger, K., Schiavone, S., Miller, F. J., and Krause, K.-H. (2012). Reactive oxygen species: from health to disease. Swiss Med. Wkly. 142:w13659. doi: 10.4414/smw.2012.13659
Brown, R. H., and Al-Chalabi, A. (2017). Amyotrophic lateral sclerosis. N. Engl. J. Med. 377, 162–172. doi: 10.1056/NEJMra1603471
Cannan, W. J., Tsang, B. P., Wallace, S. S., and Pederson, D. S. (2014). Nucleosomes suppress the formation of double-strand DNA breaks during attempted base excision repair of clustered oxidative damages. J. Biol. Chem. 289, 19881–19893. doi: 10.1074/jbc.M114.571588
Celotto, A. M., Liu, Z., Vandemark, A. P., and Palladino, M. J. (2012). A novel Drosophila SOD2 mutant demonstrates a role for mitochondrial ROS in neurodevelopment and disease. Brain Behav. 2, 424–434. doi: 10.1002/brb3.73
Chen, T., Turner, B. J., Beart, P. M., Sheehan-Hennessy, L., Elekwachi, C., and Muyderman, H. (2018). Glutathione monoethyl ester prevents TDP-43 pathology in motor neuronal NSC-34 cells. Neurochem. Int. 112, 278–287. doi: 10.1016/j.neuint.2017.08.009
Chi, L., Ke, Y., Luo, C., Gozal, D., and Liu, R. (2007). Depletion of reduced glutathione enhances motor neuron degeneration in vitro and in vivo. Neuroscience 144, 991–1003. doi: 10.1016/j.neuroscience.2006.09.064
Cihankaya, H., Theiss, C., and Matschke, V. (2021). Little helpers or mean rogue-role of microglia in animal models of amyotrophic lateral sclerosis. Int. J. Mol. Sci. 22:993. doi: 10.3390/ijms22030993
Dave, K. R., Bradley, W. G., and Pérez-Pinzón, M. A. (2003). Early mitochondrial dysfunction occurs in motor cortex and spinal cord at the onset of disease in the Wobbler mouse. Exp. Neurol. 182, 412–420. doi: 10.1016/s0014-4886(03)00091-8
Dringen, R., Kussmaul, L., Gutterer, J. M., Hirrlinger, J., and Hamprecht, B. (1999). The glutathione system of peroxide detoxification is less efficient in neurons than in astroglial cells. J. Neurochem. 72, 2523–2530. doi: 10.1046/j.1471-4159.1999.0722523.x
Du, K., Farhood, A., and Jaeschke, H. (2017). Mitochondria-targeted antioxidant Mito-Tempo protects against acetaminophen hepatotoxicity. Arch. Toxicol. 91, 761–773. doi: 10.1007/s00204-016-1692-0
Duchen, L. W., and Strich, S. J. (1968). An hereditary motor neurone disease with progressive denervation of muscle in the mouse: the mutant “wobbler”. J. Neurol. Neurosurg. Psychiatry 31, 535–542. doi: 10.1136/jnnp.31.6.535
Dumur, T., Duncan, S., Graumann, K., Desset, S., Randall, R. S., Scheid, O. M., et al. (2019). Probing the 3D architecture of the plant nucleus with microscopy approaches: challenges and solutions. Nucleus 10, 181–212. doi: 10.1080/19491034.2019.1644592
Eve, D. J., Dennis, J. S., and Citron, B. A. (2007). Transcription factor p53 in degenerating spinal cords. Brain Res. 1150, 174–181. doi: 10.1016/j.brainres.2007.02.088
Farg, M. A., Konopka, A., Soo, K. Y., Ito, D., and Atkin, J. D. (2017). The DNA damage response (DDR) is induced by the C9orf72 repeat expansion in amyotrophic lateral sclerosis. Hum. Mol. Genet. 26, 2882–2896. doi: 10.1093/hmg/ddx170
Ferrante, R. J., Browne, S. E., Shinobu, L. A., Bowling, A. C., Baik, M. J., MacGarvey, U., et al. (1997). Evidence of increased oxidative damage in both sporadic and familial amyotrophic lateral sclerosis. J. Neurochem. 69, 2064–2074. doi: 10.1046/j.1471-4159.1997.69052064.x
Gardiner, B., Dougherty, J. A., Ponnalagu, D., Singh, H., Angelos, M., Chen, C.-A., et al. (2020). “Measurement of oxidative stress markers in vitro using commercially available kits,” in Measuring Oxidants and Oxidative Stress in Biological Systems, eds L. J. Berliner and N. L. Parinandi (Cham: Springer), 39–60.
Ghardi, M., Moreels, M., Chatelain, B., Chatelain, C., and Baatout, S. (2012). Radiation-induced double strand breaks and subsequent apoptotic DNA fragmentation in human peripheral blood mononuclear cells. Int. J. Mol. Med. 29, 769–780. doi: 10.3892/ijmm.2012.907
Ghosh, D., Levault, K. R., and Brewer, G. J. (2014). Relative importance of redox buffers GSH and NAD(P)H in age-related neurodegeneration and Alzheimer disease-like mouse neurons. Aging Cell 13, 631–640. doi: 10.1111/acel.12216
Giorgio, F. de., Maduro, C., Fisher, E. M. C., and Acevedo-Arozena, A. (2019). Transgenic and physiological mouse models give insights into different aspects of amyotrophic lateral sclerosis. Dis. Models Mech. 12:dmm037424. doi: 10.1242/dmm.037424
Glorieux, C., Zamocky, M., Sandoval, J. M., Verrax, J., and Calderon, P. B. (2015). Regulation of catalase expression in healthy and cancerous cells. Free Radic. Biol. Med. 87, 84–97. doi: 10.1016/j.freeradbiomed.2015.06.017
Gonzalez Deniselle, M. C., Carreras, M. C., Garay, L., Gargiulo-Monachelli, G., Meyer, M., Poderoso, J. J., et al. (2012). Progesterone prevents mitochondrial dysfunction in the spinal cord of wobbler mice. J. Neurochem. 122, 185–195. doi: 10.1111/j.1471-4159.2012.07753.x
Gorrini, C., Harris, I. S., and Mak, T. W. (2013). Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 12, 931–947. doi: 10.1038/nrd4002
Guízar-Sahagún, G., Ibarra, A., Espitia, A., Martínez, A., Madrazo, I., and Franco-Bourland, R. E. (2005). Glutathione monoethyl ester improves functional recovery, enhances neuron survival and stabilizes spinal cord blood flow after spinal cord injury in rats. Neuroscience 130, 639–649. doi: 10.1016/j.neuroscience.2004.09.056
Henderson, J. T., Javaheri, M., Kopko, S., and Roder, J. C. (1996). Reduction of lower motor neuron degeneration in wobbler mice by N-acetyl-L-cysteine. J. Neurosci. 16, 7574–7582. doi: 10.1523/JNEUROSCI.16-23-07574.1996
Henrikus, S. S., Henry, C., McDonald, J. P., Hellmich, Y., Bruckbauer, S. T., Cherry, M. E., et al. (2019). DNA double-strand breaks induced by reactive oxygen species promote DNA polymerase IV activity in Escherichia Coli. bioRXiv.[Preprint]. doi: 10.1101/533422
Hu, H., and Li, M. (2016). Mitochondria-targeted antioxidant mitotempo protects mitochondrial function against amyloid beta toxicity in primary cultured mouse neurons. Biochem. Biophys. Res. Commun. 478, 174–180. doi: 10.1016/j.bbrc.2016.07.071
Ikeda, K., and Iwasaki, Y. (2015). Edaravone, a free radical scavenger, delayed symptomatic and pathological progression of motor neuron disease in the wobbler mouse. PLoS One 10:e0140316. doi: 10.1371/journal.pone.0140316
Ikeda, K., Iwasaki, Y., and Kaji, R. (2015). Neuroprotective effect of ultra-high dose methylcobalamin in wobbler mouse model of amyotrophic lateral sclerosis. J. Neurol. Sci. 354, 70–74. doi: 10.1016/j.jns.2015.04.052
Islam, M. T. (2017). Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 39, 73–82. doi: 10.1080/01616412.2016.1251711
Iyama, T., and Wilson, D. M., III (2013). DNA repair mechanisms in dividing and non-dividing cells. DNA Repair (Amst) 12, 620–636. doi: 10.1016/j.dnarep.2013.04.015
Jaiswal, M. K. (2019). Riluzole and edaravone: a tale of two amyotrophic lateral sclerosis drugs. Med. Res. Rev. 39, 733–748. doi: 10.1002/med.21528
Janssen-Heininger, Y. M. W., Mossman, B. T., Heintz, N. H., Forman, H. J., Kalyanaraman, B., Finkel, T., et al. (2008). Redox-based regulation of signal transduction: principles, pitfalls and promises. Free Radic. Biol. Med. 45, 1–17. doi: 10.1016/j.freeradbiomed.2008.03.011
Johnson, W. M., Wilson-Delfosse, A. L., and Mieyal, J. J. (2012). Dysregulation of glutathione homeostasis in neurodegenerative diseases. Nutrients 4, 1399–1440. doi: 10.3390/nu4101399
Kang, M. Y., Kim, H.-B., Piao, C., Lee, K. H., Hyun, J. W., Chang, I.-Y., et al. (2013). The critical role of catalase in prooxidant and antioxidant function of p53. Cell Death Differ. 20, 117–129. doi: 10.1038/cdd.2012.102
Kannan, A., Bhatia, K., Branzei, D., and Gangwani, L. (2018). Combined deficiency of Senataxin and DNA-PKcs causes DNA damage accumulation and neurodegeneration in spinal muscular atrophy. Nucleic Acids Res. 46, 8326–8346. doi: 10.1093/nar/gky641
Kaupmann, K., Simon-Chazottes, D., Guénet, J.-L., and Jokusch, H. (1992). Wobbler, a mutation affecting motoneuron survival and gonadal functions in the mouse, maps to proximal chromosome 11. Genomics 13, 39–43. doi: 10.1016/0888-7543(92)90199-3
Kiernan, M. C., Vucic, S., Cheah, B. C., Turner, M. R., Eisen, A., Hardiman, O., et al. (2011). Amyotrophic lateral sclerosis. Lancet 377, 942–955. doi: 10.1016/S0140-6736(10)61156-7
Kim, B. W., Jeong, Y. E., Wong, M., and Martin, L. J. (2020). DNA damage accumulates and responses are engaged in human ALS brain and spinal motor neurons and DNA repair is activatable in iPSC-derived motor neurons with SOD1 mutations. Acta Neuropathol. Commun. 8:7. doi: 10.1186/s40478-019-0874-4
Klatt, C. L., Theis, V., Hahn, S., Theiss, C., and Matschke, V. (2019). Deregulated miR-29b-3p correlates with tissue-specific activation of intrinsic apoptosis in an animal model of amyotrophic lateral sclerosis. Cells 8:1077. doi: 10.3390/cells8091077
Kobayashi, J. (2004). Molecular mechanism of the recruitment of NBS1/hMRE11/hRAD50 complex to DNA double-strand breaks: NBS1 binds to γ-H2AX through FHA/BRCT domain. J. Radiat. Res. 45, 473–478. doi: 10.1269/jrr.45.473
Kobayashi, H., Ogawa, K., Kawahara, N., Iwai, K., Niiro, E., Morioka, S., et al. (2017). Sequential molecular changes and dynamic oxidative stress in high-grade serous ovarian carcinogenesis. Free Radic. Res. 51, 755–764. doi: 10.1080/10715762.2017.1383605
Konopka, A., Whelan, D. R., Jamali, M. S., Perri, E., Shahheydari, H., Toth, R. P., et al. (2020). Impaired NHEJ repair in amyotrophic lateral sclerosis is associated with TDP-43 mutations. Mol. Neurodegener. 15:51. doi: 10.1186/s13024-020-00386-4
Kuo, L. J., and Yang, L.-X. (2008). Gamma-H2AX - A Novel Biomarker for DNA double-strand Breaks. In vivo 22, 305–309.
Lagouge, M., and Larsson, N.-G. (2013). The role of mitochondrial DNA mutations and free radicals in disease and ageing. J. Intern. Med. 273, 529–543. doi: 10.1111/joim.12055
Lieber, M. R. (2010). The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 79, 181–211. doi: 10.1146/annurev.biochem.052308.093131
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lopez-Gonzalez, R., Lu, Y., Gendron, T. F., Karydas, A., Tran, H., Yang, D., et al. (2016). Poly(GR) in C9ORF72-related ALS/FTD compromises mitochondrial function and increases oxidative stress and DNA damage in iPSC-derived motor neurons. Neuron 92, 383–391. doi: 10.1016/j.neuron.2016.09.015
Lu, L., Guo, L., Gauba, E., Tian, J., Wang, L., Tandon, N., et al. (2015). Transient cerebral ischemia promotes brain mitochondrial dysfunction and exacerbates cognitive impairments in young 5xFAD mice. PLoS One 10:e0144068. doi: 10.1371/journal.pone.0144068
Mah, L.-J., El-Osta, A., and Karagiannis, T. C. (2010). γH2AX: a sensitive molecular marker of DNA damage and repair. Leukemia 24, 679–686. doi: 10.1038/leu.2010.6
Matschke, V., Theiss, C., and Matschke, J. (2019). Oxidative stress: the lowest common denominator of multiple diseases. Neural Regen. Res. 14, 238–241. doi: 10.4103/1673-5374.244780
Mejzini, R., Flynn, L. L., Pitout, I. L., Fletcher, S., Wilton, S. D., and Akkari, P. A. (2019). ALS genetics, mechanisms and therapeutics: where are we now? Front. Neurosci. 13:1310. doi: 10.3389/fnins.2019.01310
Merighi, A., Gionchiglia, N., Granato, A., and Lossi, L. (2021). The phosphorylated form of the histone H2AX (γH2AX) in the brain from embryonic life to old age. Molecules 26:7198. doi: 10.3390/molecules26237198
Miao, L., and St Clair, D. K. (2009). Regulation of superoxide dismutase genes: implications in disease. Free Radic. Biol. Med. 47, 344–356. doi: 10.1016/j.freeradbiomed.2009.05.018
Moser, J. M., Bigini, P., and Schmitt-John, T. (2013). The wobbler mouse, an ALS animal model. Mol. Genet. Genomics 288, 207–229. doi: 10.1007/s00438-013-0741-0
Oka, S., Kamata, H., Kamata, K., Yagisawa, H., and Hirata, H. (2000). N-Acetylcysteine suppresses TNF-induced NF-κB activation through inhibition of IκB kinases. FEBS Lett. 472, 196–202. doi: 10.1016/S0014-5793(00)01464-2
Ott, B., Dahlke, C., Meller, K., Napirei, M., Schmitt-John, T., Brand-Saberi, B., et al. (2015). Implementation of a manual for working with wobbler mice and criteria for discontinuation of the experiment. Ann. Anat. 200, 118–124. doi: 10.1016/j.aanat.2015.03.007
Owusu-Ansah, E., Yavari, A., and Banerjee, U. (2008). A protocol for in vivo detection of reactive oxygen species. Nat. Protocols doi: 10.1038/NPROT.2008.23
Panier, S., and Boulton, S. J. (2014). Double-strand break repair: 53BP1 comes into focus. Nat. Rev. Mol. Cell Biol. 15, 7–18. doi: 10.1038/nrm3719
Petrov, D., Mansfield, C., Moussy, A., and Hermine, O. (2017). ALS clinical trials review: 20 years of failure. Are we any closer to registering a new treatment? Front. Aging Neurosci. 9:68. doi: 10.3389/fnagi.2017.00068
Podhorecka, M., Skladanowski, A., and Bozko, P. (2010). H2AX phosphorylation: its role in DNA damage response and cancer therapy. J. Nucleic Acids 2010:920161. doi: 10.4061/2010/920161
Pourahmad, J., Salimi, A., and Seydi, E. (2016). “Role of oxygen free radicals in cancer development and treatment,” in Free Radicals and Diseases, ed R. Ahmad (UK: IntechOpen), 347–361.
Qi, L., Wu, X.-C., and Zheng, D.-Q. (2019). Hydrogen peroxide, a potent inducer of global genomic instability. Curr. Genet. 65, 913–917. doi: 10.1007/s00294-019-00969-9
Rae, C. D., and Williams, S. R. (2017). Glutathione in the human brain: review of its roles and measurement by magnetic resonance spectroscopy. Anal. Biochem. 529, 127–143. doi: 10.1016/j.ab.2016.12.022
Ren, X., Zou, L., Zhang, X., Branco, V., Wang, J., Carvalho, C., et al. (2017). Redox signaling mediated by thioredoxin and glutathione systems in the central nervous system. Antioxid. Redox Signal. 27, 989–1010. doi: 10.1089/ars.2016.6925
Rhee, S. G. (2006). Cell signaling. H2O2, a necessary evil for cell signaling. Science 312, 1882–1883. doi: 10.1126/science.1130481
Röderer, P., Klatt, L., John, F., Theis, V., Winklhofer, K. F., Theiss, C., et al. (2018). Increased ROS level in spinal cord of wobbler mice due to Nmnat2 downregulation. Mol. Neurobiol. 55, 8414–8424. doi: 10.1007/s12035-018-0999-7
Rohm, M., May, C., Marcus, K., Steinbach, S., Theis, V., Theiss, C., et al. (2019). The microRNA miR-375-3p and the tumor suppressor NDRG2 are involved in sporadic amyotrophic lateral sclerosis. Cell Physiol. Biochem. 52, 1412–1426. doi: 10.3892/or.2020.7738
Ross, E. K., Gray, J. J., Winter, A. N., and Linseman, D. A. (2012). Immunocal® and preservation of glutathione as a novel neuroprotective strategy for degenerative disorders of the nervous system. Recent Patents CNS Drug Discov. 7, 230–235. doi: 10.2174/157488912803252014
Rush, T., Liu, X., Nowakowski, A. B., Petering, D. H., and Lobner, D. (2012). Glutathione-mediated neuroprotection against methylmercury neurotoxicity in cortical culture is dependent on MRP1. Neurotoxicology 33, 476–481. doi: 10.1016/j.neuro.2012.03.004
Rushworth, G. F., and Megson, I. L. (2014). Existing and potential therapeutic uses for N-acetylcysteine: the need for conversion to intracellular glutathione for antioxidant benefits. Pharmacol. Ther. 141, 150–159. doi: 10.1016/j.pharmthera.2013.09.006
Saberi, D., Ott, B., Dahlke, C., Matschke, V., Schmitt-John, T., and Theiss, C. (2016). The spatiotemporal pattern of degeneration in the cerebellum of the wobbler mouse. J. Neuropathol. Exp. Neurol. 75, 347–357. doi: 10.1093/jnen/nlw005
Santoro, B., Bigini, P., Levandis, G., Nobile, V., Biggiogera, M., Botti, F., et al. (2004). Evidence for chronic mitochondrial impairment in the cervical spinal cord of a murine model of motor neuron disease. Neurobiol. Dis. 17, 349–357. doi: 10.1016/j.nbd.2004.07.003
Schmitt-John, T., Drepper, C., Mussmann, A., Hahn, P., Kuhlmann, M., Thiel, C., et al. (2005). Mutation of Vps54 causes motor neuron disease and defective spermiogenesis in the wobbler mouse. Nat. Genet. 37, 1213–1215. doi: 10.1038/ng1661
Schultz, L. B., Chehab, N. H., Malikzay, A., and Halazonetis, T. D. (2000). p53 binding protein 1 (53BP1) is an early participant in the cellular response to DNA double-strand breaks. J. Cell Biol. 151, 1381–1390. doi: 10.1083/jcb.151.7.1381
Seiler, A., Schneider, M., Förster, H., Roth, S., Wirth, E. K., Culmsee, C., et al. (2008). Glutathione peroxidase 4 senses and translates oxidative stress into 12/15-lipoxygenase dependent- and AIF-mediated cell death. Cell Metab. 8, 237–248. doi: 10.1016/j.cmet.2008.07.005
Sepasi Tehrani, H., and Moosavi-Movahedi, A. A. (2018). Catalase and its mysteries. Prog. Biophys. Mol. Biol. 140, 5–12. doi: 10.1016/j.pbiomolbio.2018.03.001
Shanbhag, N. M., Evans, M. D., Mao, W., Nana, A. L., Seeley, W. W., Adame, A., et al. (2019). Early neuronal accumulation of DNA double strand breaks in Alzheimer’s disease. Acta Neuropathol. Commun. 7:77. doi: 10.1186/s40478-019-0723-5
Shibata, N., Nagai, R., Uchida, K., Horiuchi, S., Yamada, S., Hirano, A., et al. (2001). Morphological evidence for lipid peroxidation and protein glycoxidation in spinal cords from sporadic amyotrophic lateral sclerosis patients. Brain Res. 917, 97–104. doi: 10.1016/s0006-8993(01)02926-2
Singh, A., Kukreti, R., Saso, L., and Kukreti, S. (2019). Oxidative stress: a key modulator in neurodegenerative diseases. Molecules 24:1583. doi: 10.3390/molecules24081583
Smith, E. F., Shaw, P. J., and de Vos, K. J. (2019). The role of mitochondria in amyotrophic lateral sclerosis. Neurosci. Lett. 710:132933. doi: 10.1016/j.neulet.2017.06.052
Srinivas, U. S., Tan, B. W. Q., Vellayappan, B. A., and Jeyasekharan, A. D. (2019). ROS and the DNA damage response in cancer. Redox Biol. 25:101084. doi: 10.1016/j.redox.2018.101084
Staal, F. J., Anderson, M. T., and Herzenberg, L. A. (1995). Redox regulation of activation of NF-KB transcription factor complex: effects of N-acetylcysteine. Methods Enzymol. 252, 168–174. doi: 10.1016/0076-6879(95)52019-8
Stein, J., Walkenfort, B., Cihankaya, H., Hasenberg, M., Bader, V., Winklhofer, K. F., et al. (2021). Increased ROS-dependent fission of mitochondria causes abnormal morphology of the cell powerhouses in a murine model of amyotrophic lateral sclerosis. Oxid. Med. Cell. Longev. 2021:6924251. doi: 10.1155/2021/6924251
Stephenson, J., and Amor, S. (2017). Modelling amyotrophic lateral sclerosis in mice. Drug Discov. Dis. Models 25–26, 35–44. doi: 10.1016/j.ddmod.2018.10.001
Tam, O. H., Rozhkov, N. V., Shaw, R., Kim, D., Hubbard, I., Fennessey, S., et al. (2019). Postmortem cortex samples identify distinct molecular subtypes of ALS: retrotransposon activation, oxidative stress and activated glia. Cell Rep. 29, 1164–1177.e5. doi: 10.1016/j.celrep.2019.09.066
Thomson, S. R., Nahon, J. E., Mutsaers, C. A., Thomson, D., Hamilton, G., Parson, S. H., et al. (2012). Morphological characteristics of motor neurons do not determine their relative susceptibility to degeneration in a mouse model of severe spinal muscular atrophy. PLoS One 7:e52605. doi: 10.1371/journal.pone.0052605
Townsend, D. M., Tew, K. D., and Tapiero, H. (2003). The importance of glutathione in human disease. Biomed. Pharmacother. 57, 145–155. doi: 10.1016/s0753-3322(03)00043-x
Trnka, J., Blaikie, F. H., Smith, R. A. J., and Murphy, M. P. (2008). A mitochondria-targeted nitroxide is reduced to its hydroxylamine by ubiquinol in mitochondria. Free Radic. Biol. Med. 44, 1406–1419. doi: 10.1016/j.freeradbiomed.2007.12.036
van Remmen, H., Ikeno, Y., Hamilton, M., Pahlavani, M., Wolf, N., Thorpe, S. R., et al. (2003). Life-long reduction in MnSOD activity results in increased DNA damage and higher incidence of cancer but does not accelerate aging. Physiol. Genomics 16, 29–37. doi: 10.1152/physiolgenomics.00122.2003
Wang, Z., Bai, Z., Qin, X., and Cheng, Y. (2019). Aberrations in oxidative stress markers in amyotrophic lateral sclerosis: a systematic review and meta-analysis. Oxid. Med. Cell. Longev. 2019:1712323. doi: 10.1155/2019/1712323
Weber, U. J., Bock, T., Buschard, K., and Pakkenberg, B. (1997). Total number and size distribution of motor neurons in the spinal cord of normal and EMC-virus infected mice–a stereological study. J. Anat. 191, 347–353. doi: 10.1046/j.1469-7580.1997.19130347.x
Weyemi, U., Paul, B. D., Snowman, A. M., Jailwala, P., Nussenzweig, A., Bonner, W. M., et al. (2018). Histone H2AX deficiency causes neurobehavioral deficits and impaired redox homeostasis. Nat. Commun. 9:1526. doi: 10.1038/s41467-018-03948-9
Williamson, J., Hughes, C. M., Burke, G., and Davison, G. W. (2020). A combined γ-H2AX and 53BP1 approach to determine the DNA damage-repair response to exercise in hypoxia. Free Radic. Biol. Med. 154, 9–17. doi: 10.1016/j.freeradbiomed.2020.04.026
Winkler, E. A., Sengillo, J. D., Sagare, A. P., Zhao, Z., Ma, Q., Zuniga, E., et al. (2014). Blood-spinal cord barrier disruption contributes to early motor-neuron degeneration in ALS-model mice. Proc. Natl. Acad. Sci. U S A 111, E1035–E1042. doi: 10.1073/pnas.1401595111
Wu, G., Fang, Y.-Z., Yang, S., Lupton, J. R., and Turner, N. D. (2004). Glutathione metabolism and its implications for health. J. Nutr. 134, 489–492. doi: 10.1093/jn/134.3.489
Wu, W., Liu, B.-H., Xie, C.-L., Xia, X.-D., and Zhang, Y.-M. (2018). Neuroprotective effects of N-acetyl cysteine on primary hippocampus neurons against hydrogen peroxide-induced injury are mediated via inhibition of mitogen-activated protein kinases signal transduction and antioxidative action. Mol. Med. Rep. 17, 6647–6654. doi: 10.3892/mmr.2018.8699
Xu, G.-P., Dave, K. R., Moraes, C. T., Busto, R., Sick, T. J., Bradley, W. G., et al. (2001). Dysfunctional mitochondrial respiration in the wobbler mouse brain. Neurosci. Lett. 300, 141–144. doi: 10.1016/s0304-3940(01)01575-0
Yoshino, H. (2019). Edaravone for the treatment of amyotrophic lateral sclerosis. Expert Rev. Neurother. 19, 185–193. doi: 10.1080/14737175.2019.1581610
Zhang, J., He, Y., Shen, X., Jiang, D., Wang, Q., Liu, Q., et al. (2016). γ-H2AX responds to DNA damage induced by long-term exposure to combined low-dose-rate neutron and γ-ray radiation. Mutat Res. Genet. Toxicol. Environ. Mutagen. 795, 36–40. doi: 10.1016/j.mrgentox.2015.11.004
Zhou, Q., Liu, C., Liu, W., Zhang, H., Zhang, R., Liu, J., et al. (2015). Rotenone induction of hydrogen peroxide inhibits mTOR-mediated S6K1 and 4E-BP1/eIF4E pathways, leading to neuronal apoptosis. Toxicol. Sci. 143, 81–96. doi: 10.1093/toxsci/kfu211
Zou, Z.-Y., Zhou, Z.-R., Che, C.-H., Liu, C.-Y., He, R.-L., and Huang, H.-P. (2017). Genetic epidemiology of amyotrophic lateral sclerosis: a systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 88, 540–549. doi: 10.1136/jnnp-2016-315018
Keywords: Wobbler, double strand breaks, glutathione ethyl ester, N-Acetyl-L-Cysteine, Mito-TEMPO, p53bp1, neuroprotection
Citation: Junghans M, John F, Cihankaya H, Schliebs D, Winklhofer KF, Bader V, Matschke J, Theiss C and Matschke V (2022) ROS scavengers decrease γH2ax spots in motor neuronal nuclei of ALS model mice in vitro. Front. Cell. Neurosci. 16:963169. doi: 10.3389/fncel.2022.963169
Received: 07 June 2022; Accepted: 08 August 2022;
Published: 31 August 2022.
Edited by:
Anwen Shao, Zhejiang University, ChinaReviewed by:
Anna Konopka, Macquarie University, AustraliaJoy Mitra, Houston Methodist Research Institute, United States
Copyright © 2022 Junghans, John, Cihankaya, Schliebs, Winklhofer, Bader, Matschke, Theiss and Matschke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Veronika Matschke, VmVyb25pa2EuTWF0c2Noa2VAcnViLmRl
† These authors have contributed equally to this work and share first authorship