 Anwesha Bhattacharyya
Anwesha Bhattacharyya- Amity Institute of Neuropsychology and Neurosciences, Amity University, Noida, UP, India
The leading cause of blindness in inherited and age-related retinal degeneration (RD) is the death of retinal photoreceptors such as rods and cones. The most prevalent form of RD is age-related macular degeneration (AMD) which affects the macula resulting in an irreversible loss of vision. The other is a heterogenous group of inherited disorders known as Retinitis Pigmentosa (RP) caused by the progressive loss of photoreceptors. Several approaches have been developed in recent years to artificially stimulate the remaining retinal neurons using optogenetics, retinal prostheses, and chemical photoswitches. However, the outcome of these strategies has been limited. The success of these treatments relies on the morphology, physiology, and proper functioning of the remaining intact structures in the downstream visual pathway. It is not completely understood what all alterations occur in the visual cortex during RD. In this review, I will discuss the known information in the literature about morphological and functional changes that occur in the visual cortex in rodents and humans during RD. The aim is to highlight the changes in the visual cortex that will be helpful for developing tools and strategies directed toward the restoration of high-resolution vision in patients with visual impairment.
Significance statement
To achieve successful therapeutic intervention in RD, the structures downstream of the retina (lateral geniculate nucleus, LGN; and primary visual cortex, V1) should remain intact. Long-term sensory deprivation resulting from degenerating retina is associated with potential changes in the cortical circuit posing significant constraints on the success of RD treatment. Therefore, it is important to understand the consequences of RD on the state of the visual cortex for the successful implementation of restorative techniques. This review provides an up-to-date overview of the morphological and physiological changes in the visual cortex from various rodent and human studies.
Introduction
The retina is a light-sensitive tissue layer at the back of the eye equipped with the necessary machinery to process light information to create an informative image of the external world. During retinal degeneration (RD) the retina undergoes deterioration with the consequent death of photoreceptors such as rods and cones. The progressive loss of receptors and loss of vision has a greater impact on daily life such as not being able to recognize faces, read and find objects. The most common form of RD is age-related macular degeneration (AMD) with vision loss in the elderly population. A recent report on numerous population-based studies indicates the emerging global burden arising with the increase in the number of people suffering from AMD from 196 million in 2020 to 288 million in 2040 (Wong et al., 2014). Another cause of RD is Retinitis Pigmentosa resulting from progressive loss of photoreceptors and the reginal pigment epithelium. An estimated 1.5 million individuals are affected with RP in the age group of 20–64 years (Bovolenta and Cisneros, 2009; Moore et al., 2020). Visual impairment not only has a personal impact but also represents a significant societal and economic burden. It thus poses a major health problem with an urgent medical need.
Several therapeutic strategies have been developed to combat RD, arising hope for patients. Such strategies include stem cell therapy (Ramsden et al., 2013), retinal prostheses (Shim et al., 2020), light-activated ion pump (Busskamp et al., 2010), photoswitch approaches modulating intrinsic channel (Polosukhina et al., 2012), and optogenetic gene therapy (van Wyk et al., 2015; Sahel et al., 2021). However, until now, all of them have had limitations in achieving the high-resolution vision in pre-clinical models as well as in patients. The success of these therapies relies heavily on the integrity of the visual system along the downstream region of the visual pathway (Kien et al., 2012).
There is abundant information about the progressive remodeling happening in the inner retinal neurons and glial cells following the death of photoreceptors (Jones et al., 1995; Marc et al., 2003; Anderson et al., 2016). Severe remodeling gradually affects the retinal ganglion cells (RGCs) at later stages (Garcia-Ayuso et al., 2019). On contrary, one finding shows that the dendritic structures of the RGCs remain intact and maintain stability even into the late stages of RD (Damiani et al., 2012; Lin and Peng, 2013; Anderson et al., 2016). For successful visual restoration, it is important to understand how the structures downstream in the visual pathway change over time. This review highlights the work that has examined the morphological and physiological changes in the visual pathway during RD.
Murine and rat models are widely used for RD research and have been advantageous in assisting the development of new therapies (Chung, 2017). There are a wide variety of animal models that have genetic mutations having resemblances with humans (Chang et al., 2002). These models have been very useful in understanding morphological and physiological changes. In contrast, rodent models of AMD are limited as they lack anatomical macula, have a lower percentage of cones, and the disease is not caused by a single genetic defect unlike RP but by several genetic polymorphisms. Further adding to the complexity are the anatomical differences between the human retina and AMD models (Pennesi et al., 2012; Soundara Pandi et al., 2021). Until now there are no animal models that can entirely represent the complete disease phenotype of AMD.
Morphological changes in the visual cortex during RD
During early postnatal development the visual cortex undergoes dynamic changes (Himmelhan et al., 2018). Mice with RD in postnatal ages, P3–P28 show the same expression of ontogenetic markers, such as Nestin, Doublecortin, MAP2, Parvalbumin, and NeuN similar to wild-type mice. These proteins are expressed at different stages of development and some of them continue their expression until adulthood. Results demonstrate that these markers do not show any obvious deviation in cortical development during RD (Himmelhan et al., 2018). Until now, it is not completely clear if the visual cortex remains intact or undergoes structural changes following defects at the level of the retina. Humans suffering from MD undergo loss of vision in the central visual field. People who have an early onset of MD known as Juvenile Macular Degeneration (JMD) have different morphological changes compared to AMD. In AMD, only the structures downstream to the lateral geniculate bodies and frontal lobe white matter are affected while in JMD all the structures in the entire visual pathway (optic nerve, chiasm, lateral geniculate bodies, visual cortex) undergo volumetric reduction (Nuzzi et al., 2020). Additionally, AMD patients also have a reduction in the gray matter density following prolonged visual deprivation (Boucard et al., 2009). During MD the corresponding regions of the primary visual cortex (V1) deprived of retinal input result in the formation of silent cortical zones termed lesion projection zone (LPZ) (Ferreira et al., 2017). There is a discrepancy in the existing literature on whether new anatomical connections are generated with changes in the strength of the existing connections (plastic reorganization) with time (Wandell and Smirnakis, 2009). The impact of MD within the LPZ was assessed in a primate study and humans to see if the visual cortex undergoes reorganization by remapping visual inputs. The effects were assessed in a macaque monkey suffering from chronic MD (Shao et al., 2013). There was no significant change in the extent of the LPZ border and the deafferented region of V1. Consistently, the secondary visual area, V2 also had a similar trend with a stable LPZ border. However, it was interesting to note that area V5/MT showed significant activation suggesting marked reorganization. This could arise from the population receptive fields of the deafferented region of V5 that attain the capacity to be modulated by the visual stimulus presented outside the area of the retinal lesion. However, with due course of disease progression, these ectopic population receptive fields do not return to their original size indicating pronounced reorganization. Similar results were also obtained from AMD and JMD individuals where remapping was largely absent in the early visual areas (Baseler et al., 2011). These results corroborate the fact that there is limited reorganization in the early visual areas due to the absence of significant visual modulation despite the loss of vision.
It has been shown that the visual white matter pathway is greatly affected in age-related diseases as well as in several eye diseases (Wang et al., 2012). The white matter tissue organization within the optic radiation in AMD subjects has profound damage correlating with the loss in visual acuity (Yoshimine et al., 2018). Another form of AMD such as the neovascular form of AMD has an acute onset with leakage of fluid and hemorrhage in the central macula and these patients have different short- and long-term changes in the visual cortex (Hanson et al., 2019). The patients affected show no changes in the cortical volume of the occipital lobe of the brain within 3 months of disease diagnosis. However, these patients showed a decrease in cortical volume after long-term assessment (~5 years) in the LPZ region of the visual cortex. The central cortical region of V1 deprived of visual input also undergoes significant thinning, unlike the peripherally responsive area that becomes thicker as a compensatory mechanism of improvement in peripheral vision (Burge et al., 2016).
In RP, the degeneration starts in the periphery and advances toward the central retina. These patients display no changes in the thickness of gray matter but undergo a shift of central retinal representation to further peripheral locations (Ferreira et al., 2017). One plausible reason for the remapping could be rapid adaptation following degeneration. The neurons from the central visual field exert influence on the deafferented peripheral visual field by strengthening the long-range horizontal connections within V1. The retinotopic changes thus help in the stabilization of central vision increasing the necessity to facilitate visual plasticity. In addition, there are changes in the anatomical connectivity across the visual areas V1–V3 (Fine and Park, 2018). In the absence of visual input, the occipital cortex is recruited for various auditory and tactile tasks resulting in cross-modal plasticity (Cunningham et al., 2015). The human medial temporal complex (hMT+) present in the dorsal visual stream gets tuned to perform comparable computations as in the visual cortex. The corpus callosum which is a channel of information processing between two hemispheres is induced by plastic rearrangements due to visual dysfunction.
The V1 and the association cortices undergo significant changes in the gray matter volume restricted to the occipital cortex of patients with RP. The extent of loss is associated with the extent of peripheral visual degeneration (Rita Machado et al., 2017). The differences observed in spatial reorganization in RP and AMD patients are most likely due to distinct processing of central and peripheral vision, and different connections of the central and peripheral visual neurons with subcortical and higher-order cortical areas. Another reason could be the death of the RGCs leading to volumetric reduction of the visual cortex weakening the chances of functional reorganization in AMD patients (Hernowo et al., 2014). The summary of the morphological changes in the downstream visual pathway during RD in humans and rodents is listed in Table 1. These studies indicate that the changes in the visual cortex following RD may not be apparent initially after disease onset but may emerge slowly over time resulting from prolonged loss of input.
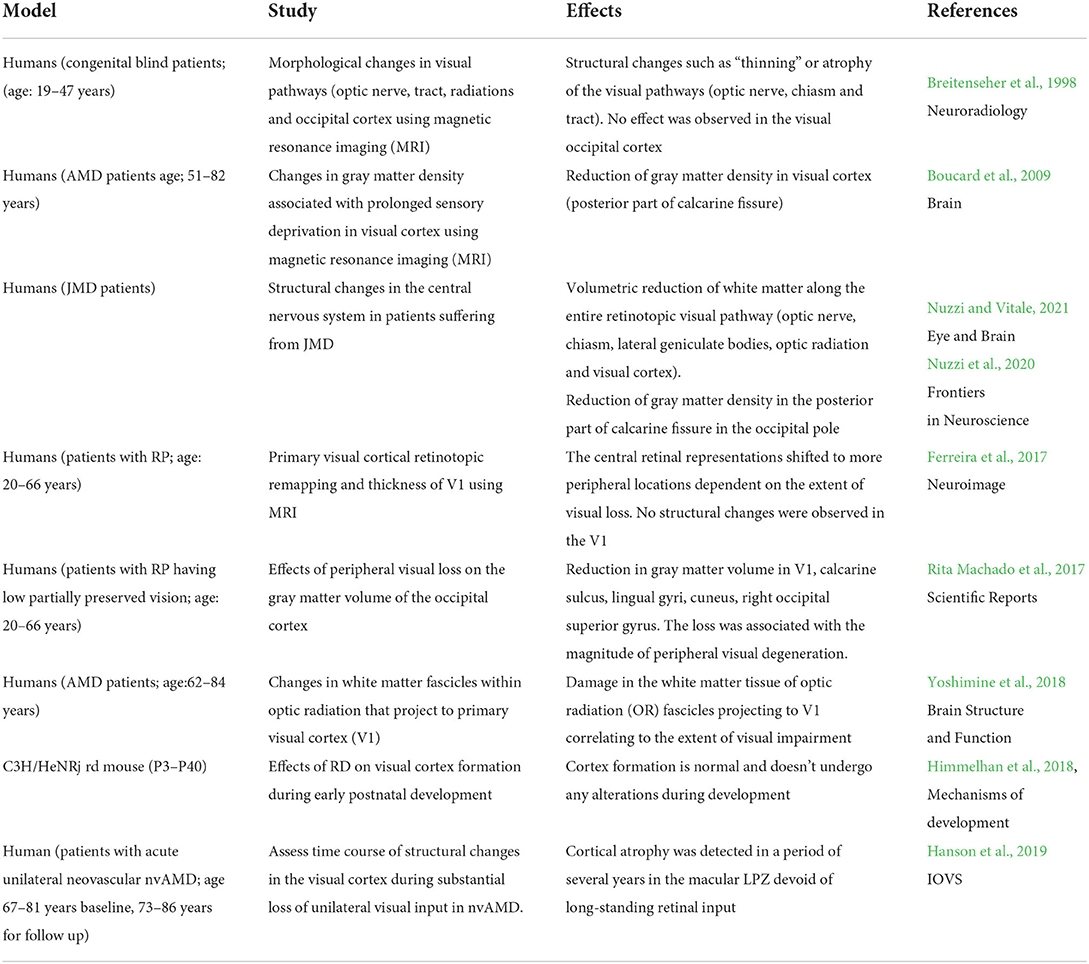
Table 1. Summary of morphological studies of retinal degeneration effects on the visual cortex.
Physiological changes in the visual cortex during RD
Animal studies
In a rat model of retinal dystrophy (RCS rat), the extent of pattern-vision degeneration was demonstrated using a short pulse of bright white light and gratings of different spatial frequencies (patterns) (Gias et al., 2011). Analysis of both multiunit activity (MUA) and local field potentials (LFPs) showed a marked decrease in responses and increased latency across different layers of the visual cortex. The degenerated group had reduced responses across different spatial frequencies and the observations were pronounced as early as 4 weeks of age. This demonstrates that conscious perception can get affected even at the very early stages of RD, i.e., within the first postnatal months.
Another study in a transgenic rat model of RD, S334ter-3 rats, aged between 2 and 3 months (has a rhodopsin mutation like human RP patients) evaluated the changes in the electrophysiological properties of the V1 such as orientation selectivity, spatial, and temporal frequency tuning and receptive field size. Degenerated rats had diminished orientation selectivity mostly in the lower layers of the cortex (layers V-VI) with better responses only at lower spatial and temporal frequencies. The size of the receptive field was smaller compared to normal seeing rats (Chen et al., 2016). Recently, Chen and colleagues have done an elaborate study to examine how the contrast response properties in V1 neurons are affected by degeneration. The contrast response function was analyzed for each cell where higher spontaneous activity was observed in absence of contrast and weak stimulus-evoked activity at both medium and higher contrasts (30 and 100%). The firing rate was low across three different contrast levels. It is surprising to note that the contrast sensitivity (CS) semi-saturation contrast (C50) did not show any significant alterations in response to spatial-temporal variations in luminance contrast change. Overall these results indicate the diminished capacity of discriminating stimuli under different contrast conditions and adaptation of the visual system to environmental contrast (Chen et al., 2020).
Spontaneous activity is essential in establishing appropriate connectivity during the early development of the visual circuit and maintenance of topographic maps (Tritsch et al., 2007; Wosniack et al., 2021). The V1 of S334ter rats demonstrates an increase in spontaneous activity and a reduced capacity to elicit responses generated by novel stimuli (Wang et al., 2016). The elevated activity could be due to morphological changes in the visual pathway and altered function of the V1 (Hubel and Wiesel, 1964). Despite this, the visual cortex has a remarkable capacity of undergoing plastic changes even at later stages of degeneration to improve the signal quality of degraded input (Begenisic et al., 2020). As a compensatory mechanism excitation/inhibition (E/I) balance in the V1 is altered following an increase in the inhibitory drive thus improving the signal-to-noise ratio (Pietra et al., 2021). This could be due to the activation of parvalbumin-containing Gamma-aminobutyric acid (GABA) neurons that inhibit pyramidal cell activity (Bhattacharyya et al., 2013).
To gain better insights into the cortical effects of RD, neural activity in response to light flashes of different intensity were measured from V1. For this mutual information (MI) was used as a measure to determine the efficacy of information transmission between visual stimulus and neuronal activity (Wang et al., 2018). The MI of spiking activity and LFPs were compared as a better predictor of how well the visual information can be decoded by the V1 neurons. This is also helpful to understand the ability of the cortex to respond under pathological conditions in the retina. There was a substantial decrease in firing rate compared to control, a variability of responses with different light intensities, and an overall decrease in MI of responsive activities. The LFP signals were able to encode the light intensity consistently and reliably across several trials, especially in the delta and beta bands. This emphasizes that the low-frequency activity is possibly a better predictor of information processing in the visual cortex during cortical reorganization. The work reviewed suggests that while vision is impaired there are remaining neurons in the visual pathway that retain the ability to capture visual information.
Human studies
The signals from the retina are relayed to the primary visual cortex (V1) for processing visual information and conscious visual perception. During RD the retina undergoes a profound increase in spontaneous activity as a result of increased glutamate concentration, and altered synaptic input due to the death of photoreceptors (Marc et al., 2007). It is important to understand how these changes in the retina affect the functioning and information processing in the downstream visual pathway (Table 2). The loss of retinal input causes the visual cortex to undergo functional adaptations in its circuitry, which can have significant repercussions on the current therapeutic approaches. A summary of the salient effects in the visual cortex during RP and AMD disease conditions in humans is briefly summarized in Table 2.
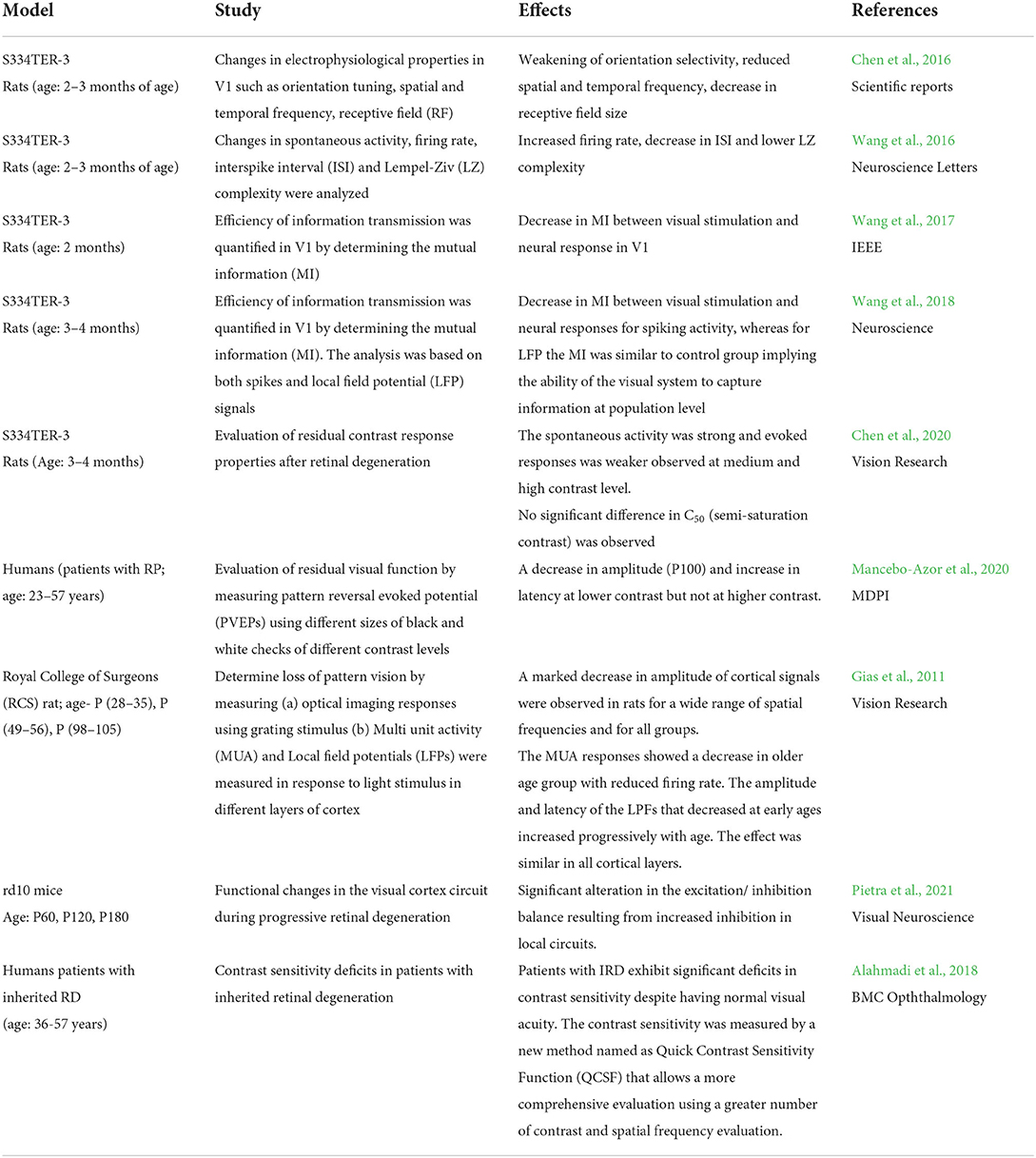
Table 2. Summary of physiological studies of retinal degeneration effects on the visual cortex.
A lack of stimulation in the visual cortex due to a retinal lesion in AMD is characterized by a central scotoma, loss of central vision, and impaired visual acuity. The loss of central vision can result in the formation of a new point of focus through eccentric fixation called preferred retinal locus (PRL). Visual stimulation in these affected individuals shows decreased activity of the visual cortex and elevated activity in a few associative areas outside the visual field responsible for eye coordination such as frontal and supplementary eye fields, prefrontal cortex, intraparietal sulci, and parietal lobule (Alahmadi et al., 2018). Another finding has also shown that AMD patients are unable to process fine details of the daily visual scenes with high spatial frequency (Ramanoel et al., 2018). Increasing the contrast level enhances the cortical responses and improves the processing of high spatial frequencies in scenes. CS is an important aspect of visual function useful for providing information about defects in perception. Deficits in CS contribute extensively to poor visual function even when visual acuity is unaffected. Implementing new methods such as Quick Contrast Sensitivity Function (QCSF) provides a comprehensive evaluation of deterioration in CS (Alahmadi et al., 2018). This test can use a wide range of contrast and spatial frequency combinations. Patients diagnosed with genetic inherited RD exhibit visual deficits despite no significant alterations in their visual acuity. This arises due to significant decreases in the CS at all spatial frequencies with pronounced effects at higher spatial frequencies.
The effect of degeneration on contrast processing was also determined in patients with RP by measuring pattern reversal visual evoked potentials (PVEPs) (Mancebo-Azor et al., 2020). The patients had a reduced amplitude and increased latency of P100 amplitude for different sizes of checks at low contrasts (16% and 6%) but not at higher contrasts. The authors hypothesized that the observed differences between low and high contrast could be due to different processing streams (parvocellular and magnocellular) with different functional properties contributing to this effect. The results are in line with previous studies in rodents and corroborate the fact that degeneration affects the functioning of the cortex in a wider manner. Recent studies in rodents and humans show that the adult brain has the ability to undergo short-term plasticity to adapt to visual changes (Lunghi et al., 2019; Begenisic et al., 2020). This is promising since it can be advantageous for possible therapies to restore vision (Castaldi et al., 2016). Taken together, the above findings suggest that although morphological changes happen in the early and late stages of the visual pathway the capability of stimulus-dependent remodeling until advanced stages of degeneration can be beneficial in the development of visual prosthesis.
Conclusion
Understanding the status of the visual cortex during the progressive death of the photoreceptors is essential for effective restorative techniques. The loss of photoreceptor input upon degeneration does not have any specific impact on the maturation and structure of the visual cortex but more generally affects the brain tissue properties. On the other hand, visual cortex neurons exhibit substantial changes in the electrophysiological properties during RD affecting their physiological activity. The retina undergoes profound morphological and functional changes resulting in reduced visual sensitivity that can potentially disrupt the efficiency of information processing in the visual cortex.
The preservation of the lower order neurons and the downstream retinal circuitry have encouraged scientists and clinicians to develop several therapeutic approaches such as retinal prosthetics and optogenetic gene therapy in restoring vision (McClements et al., 2020; Shim et al., 2020). Despite several advancements, the quality of the visual percept obtained after therapy is of relatively low quality and not enough to lead an independent life. One plausible reason for this could be the age at which therapeutic intervention is initiated. At advanced stages of degeneration when the rods are completely absent and with remnant cones, rescue strategies to obtain high acuity vision may be hindered due to disruption of excitation-inhibition balance. It is therefore necessary and significant to evaluate the changes in neural activities in the primary visual cortex during RD. Despite the fact that the rd1 mouse model of RD is widely used for optogenetic gene therapy and pre-clinical testing of inherited retinal disease, there is hardly any study evaluating changes in the visual cortex of these mice, most of the cortical work having been performed in rats. To make further advancements in pre-clinical testing it is essential to evaluate the cortical activity at different stages of RD facilitating the development of optogenetic gene therapy or visual prosthesis to achieve successful vision restoration.
Author contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Funding
AB was supported by the Ramanujan Fellowship of Science and Engineering Research Board (SERB), Department of Science and Technology (DST), Govt. of India (Grant: RJF/2019/000040), and the Core Research Grant (CRG) of SERB, DST (CRG/2021/003472).
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alahmadi, B. O., Omari, A. A., Abalem, M. F., Andrews, C., Schlegel, D., Branham, K. H., et al. (2018). Contrast sensitivity deficits in patients with mutation-proven inherited retinal degenerations. BMC Ophthalmol. 18, 313. doi: 10.1186/s12886-018-0982-0
Anderson, E. E., Greferath, U., and Fletcher, E. L. (2016). Changes in morphology of retinal ganglion cells with eccentricity in retinal degeneration. Cell Tissue Res. 364, 263–271. doi: 10.1007/s00441-015-2337-y
Baseler, H. A., Gouws, A., Haak, K. V., Racey, C., Crossland, M. D., Tufail, A., et al. (2011). Large-scale remapping of visual cortex is absent in adult humans with macular degeneration. Nat. Neurosci. 14, 649–655. doi: 10.1038/nn.2793
Begenisic, T., Mazziotti, R., Sagona, G., Lupori, L., Sale, A., Galli, L., et al. (2020). Preservation of visual cortex plasticity in retinitis pigmentosa. Neuroscience 424, 205–210. doi: 10.1016/j.neuroscience.2019.10.045
Bhattacharyya, A., Veit, J., Kretz, R., Bondar, I., and Rainer, G. (2013). Basal forebrain activation controls contrast sensitivity in primary visual cortex. BMC Neurosci. 14, 55. doi: 10.1186/1471-2202-14-55
Boucard, C. C., Hernowo, A. T., Maguire, R. P., Jansonius, N. M., Roerdink, J. B., Hooymans, J. M., et al. (2009). Changes in cortical grey matter density associated with long-standing retinal visual field defects. Brain 132, 1898–1906. doi: 10.1093/brain/awp119
Bovolenta, P., and Cisneros, E. (2009). Retinitis pigmentosa: cone photoreceptors starving to death. Nat. Neurosci. 12, 5–6. doi: 10.1038/nn0109-5
Breitenseher, M., Uhl, F., Wimberger, D. P., Deecke, L., Trattnig, S., and Kramer, J. (1998). Morphological dissociation between visual pathways and cortex: MRI of visually-deprived patients with congenital peripheral blindness. Neuroradiology. 40, 424–427. doi: 10.1007/s002340050616
Burge, W. K., Griffis, J. C., Nenert, R., Elkhetali, A., DeCarlo, D. K., ver Hoef, L. W., et al. (2016). Cortical thickness in human V1 associated with central vision loss. Sci. Rep. 6, 23268. doi: 10.1038/srep23268
Busskamp, V., Duebel, J., Balya, D., Fradot, M, Viney, T. J., Siegert, S., et al. (2010). Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa. Science 329, 413–417. doi: 10.1126/science.1190897
Castaldi, E., Cicchini, G. M., Cinelli, L., Biagi, L., Rizzo, S., and Morrone, M. C. (2016). Visual BOLD Response in Late Blind Subjects with Argus II Retinal Prosthesis. PLoS Biol. 14, e1002569. doi: 10.1371/journal.pbio.1002569
Chang, B., Hawes, N. L., Hurd, R. E., Davisson, M. T., Nusinowitz, S., and Heckenlively, J. R. (2002). Retinal degeneration mutants in the mouse. Vision Res. 42, 517–525. doi: 10.1016/S0042-6989(01)00146-8
Chen, K., Hou, B., Zhao, Y., Yuan, P., Yao, D., and Chan, L. L. H. (2020). Residual contrast response in primary visual cortex of rats with inherited retinal degeneration. Vision Res. 177, 6–11. doi: 10.1016/j.visres.2020.08.007
Chen, K., Wang, Y., Liang, X., Zhang, Y., Ng, T. K., and Chan, L. L. (2016). Electrophysiology alterations in primary visual cortex neurons of retinal degeneration (S334ter-line-3) rats. Sci. Rep. 6, 26793. doi: 10.1038/srep26793
Chung, C. Y. (2017). Animal models of retinal degeneration. J. Ophthalmic. Res. Ocular Care 1, 19–24. doi: 10.36959/936/562
Cunningham, S. I., Weiland, J. D., Bao, P., Lopez-Jaime, G. R., and Tjan, B. S. (2015). Correlation of vision loss with tactile-evoked V1 responses in retinitis pigmentosa. Vision Res. 111, 197–207. doi: 10.1016/j.visres.2014.10.015
Damiani, D., Novelli, E., Mazzoni, F., and Strettoi, E. (2012). Undersized dendritic arborizations in retinal ganglion cells of the rd1 mutant mouse: a paradigm of early onset photoreceptor degeneration. J. Comp. Neurol. 520, 1406–1423. doi: 10.1002/cne.22802
Ferreira, S., Pereira, A. C., Quendera, B., Reis, A., Silva, E. D., and Castelo-Branco, M. (2017). Primary visual cortical remapping in patients with inherited peripheral retinal degeneration. Neuroimage Clin. 13, 428–438. doi: 10.1016/j.nicl.2016.12.013
Fine, I., and Park, J. M. (2018). Blindness and human brain plasticity. Annu. Rev. Vis. Sci. 4, 337–356. doi: 10.1146/annurev-vision-102016-061241
Garcia-Ayuso, D., Di Pierdomenico, J., Vidal-Sanz, M., and Villegas-Perez, M. P. (2019). Retinal ganglion cell death as a late remodeling effect of photoreceptor degeneration. Int. J. Mol. Sci. 20, 4649. doi: 10.3390/ijms20184649
Gias, C., Vugler, A., Lawrence, J., Carr, A. J., Chen, L. L., Ahmado, A., et al. (2011). Degeneration of cortical function in the royal college of surgeons rat. Vision Res. 51, 2176–2185. doi: 10.1016/j.visres.2011.08.012
Hanson, R. L. W., Gale, R. P., Gouws, A. D., Airody, A., Scott, M. T. W., Akhtar, F., et al. (2019). Following the status of visual cortex over time in patients with macular degeneration reveals atrophy of visually deprived brain regions. Invest. Ophthalmol. Vis. Sci. 60, 5045–5051. doi: 10.1167/iovs.18-25823
Hernowo, A. T., Prins, D., Baseler, H. A., Plank, T., Gouws, A. D., Hooymans, J. M., et al. (2014). Morphometric analyses of the visual pathways in macular degeneration. Cortex 56, 99–110. doi: 10.1016/j.cortex.2013.01.003
Himmelhan, D. K., Rawashdeh, O., and Oelschlager, H. H. A. (2018). Early postnatal development of the visual cortex in mice with retinal degeneration. Mech. Dev. 151, 1–9. doi: 10.1016/j.mod.2018.03.002
Hubel, D. H., and Wiesel, T. N. (1964). Effects of monocular deprivation in kittens. Naunyn Schmiedebergs Arch. Exp. Pathol. Pharmakol. 248, 492–497. doi: 10.1007/BF00348878
Jones, B. W., Marc, R. E., and Pfeiffer, R. L. (1995). “Retinal degeneration, remodeling and plasticity,” in Webvision: The Organization of the Retina and Visual System, eds H. Kolb, E. Fernandez, and R. Nelson (Salt Lake City, UT: University of Utah Health Sciences Center).
Kien, T. T., Maul, T., and Bargiela, A. (2012). A review of retinal prosthesis approaches. Int. J. Modern Phys. 9, 209–231. doi: 10.1142/S2010194512005272
Lin, B., and Peng, E. B. (2013). Retinal ganglion cells are resistant to photoreceptor loss in retinal degeneration. PLoS ONE 8, e68084. doi: 10.1371/journal.pone.0068084
Lunghi, C., Galli-Resta, L., Binda, P., Cicchini, G. M., Placidi, G., Falsini, B., et al. (2019). Visual cortical plasticity in retinitis pigmentosa. Invest. Ophthalmol. Vis. Sci. 60, 2753–2763. doi: 10.1167/iovs.18-25750
Mancebo-Azor, R., Sáez-Moreno, J. A., and Rodríguez-Ferrer, J. M. (2020). Cortical contrast processing in retinitis pigmentosa: evidence of PVEPs spatial functions. Invest. Health, Psychol. Educ. 10, 1010–1019. doi: 10.3390/ejihpe10040071
Marc, R. E., Jones, B. W., Anderson, J. R., Kinard, K., Marshak, D. W., Wilson, J. H., et al. (2007). Neural reprogramming in retinal degeneration. Invest. Ophthalmol. Vis. Sci. 48, 3364–3371. doi: 10.1167/iovs.07-0032
Marc, R. E., Jones, B. W., Watt, C. B., and Strettoi, E. (2003). Neural remodeling in retinal degeneration. Prog. Retin. Eye Res. 22, 607–655. doi: 10.1016/S1350-9462(03)00039-9
McClements, M. E., Staurenghi, F., MacLaren, R. E., and Cehajic-Kapetanovic, J. (2020). Optogenetic gene therapy for the degenerate retina: recent advances. Front. Neurosci. 14, 570909. doi: 10.3389/fnins.2020.570909
Moore, S. M., Skowronska-Krawczyk, D., and Chao, D. L. (2020). Targeting of the NRL pathway as a therapeutic strategy to treat retinitis pigmentosa. J. Clin. Med. 9, 2224. doi: 10.3390/jcm9072224
Nuzzi, R., Dallorto, L., and Vitale, A. (2020). Cerebral modifications and visual pathway reorganization in maculopathy: a systematic review. Front. Neurosci. 14, 755. doi: 10.3389/fnins.2020.00755
Nuzzi, R., and Vitale, A. (2021). Cerebral modifications in glaucoma and macular degeneration: Analysis of current evidence in literature and their implications on therapeutic perspectives. Eye Brain. 13, 159–173. doi: 10.2147/EB.S307551
Pennesi, M. E., Neuringer, M., and Courtney, R. J. (2012). Animal models of age related macular degeneration. Mol. Aspects Med. 33, 487–509. doi: 10.1016/j.mam.2012.06.003
Pietra, G., Bonifacino, T., Talamonti, D., Bonanno, G., Sale, A., Galli, L., et al. (2021). Visual cortex engagement in retinitis pigmentosa. Int. J. Mol. Sci. 22, 9412. doi: 10.3390/ijms22179412
Polosukhina, A., Litt, J., Tochitsky, I., Nemargut, J., Sychev, Y., De Kouchkovsky, I., et al. (2012). Photochemical restoration of visual responses in blind mice. Neuron 75, 271–282. doi: 10.1016/j.neuron.2012.05.022
Ramanoel, S., Chokron, S., Hera, R., Kauffmann, L., Chiquet, C., Krainik, A., et al. (2018). Age-related macular degeneration changes the processing of visual scenes in the brain. Vis. Neurosci. 35, E006. doi: 10.1017/S0952523817000372
Ramsden, C. M., Powner, M. B., Carr, A. J., Smart, M. J., da Cruz, L., and Coffey, P. J. (2013). Stem cells in retinal regeneration: past, present and future. Development 140, 2576–2585. doi: 10.1242/dev.092270
Rita Machado, A., Carvalho Pereira, A., Ferreira, F., Ferreira, S., Quendera, B., Silva, E., et al. (2017). Structure-function correlations in retinitis pigmentosa patients with partially preserved vision: a voxel-based morphometry study. Sci. Rep. 7, 11411. doi: 10.1038/s41598-017-11317-7
Sahel, J. A., Boulanger-Scemama, E., Pagot, C., Arleo, A., Galluppi, F., Martel, J. N., et al. (2021). Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 27, 1223–1229. doi: 10.1038/s41591-021-01351-4
Shao, Y., Keliris, G. A., Papanikolaou, A., Fischer, M. D., Zobor, D., Jagle, H., et al. (2013). Visual cortex organisation in a macaque monkey with macular degeneration. Eur. J. Neurosci. 38, 3456–3464. doi: 10.1111/ejn.12349
Shim, S., Eom, K., Jeong, J., and Kim, S. J. (2020). Retinal prosthetic approaches to enhance visual perception for blind patients. Micromachines 11, 535. doi: 10.3390/mi11050535
Soundara Pandi, S. P., Ratnayaka, J. A., Lotery, A. J., and Teeling, J. L. (2021). Progress in developing rodent models of age-related macular degeneration (AMD). Exp. Eye Res. 203, 108404. doi: 10.1016/j.exer.2020.108404
Tritsch, N. X., Yi, E., Gale, J. E., Glowatzki, E., and Bergles, D. E. (2007). The origin of spontaneous activity in the developing auditory system. Nature 450, 50–55. doi: 10.1038/nature06233
van Wyk, M., Pielecka-Fortuna, J., Lowel, S., and Kleinlogel, S. (2015). Restoring the ON switch in blind retinas: opto-mGluR6, a next-generation, cell-tailored optogenetic tool. PLoS Biol. 13, e1002143. doi: 10.1371/journal.pbio.1002143
Wandell, B. A., and Smirnakis, S. M. (2009). Plasticity and stability of visual field maps in adult primary visual cortex. Nat. Rev. Neurosci. 10, 873–884. doi: 10.1038/nrn2741
Wang, Y., Chen, K., and Chan, L. L. H. (2017). Information transmission in the primary visual cortex of retinal degenerated rats. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2017, 3632–3635. doi: 10.1109/EMBC.2017.8037644
Wang, Y., Chen, K., and Chan, L. L. H. (2018). Responsive neural activities in the primary visual cortex of retina-degenerated rats. Neuroscience 383, 84–97. doi: 10.1016/j.neuroscience.2018.05.006
Wang, Y., Chen, K., Xu, P., Ng, T. K., and Chan, L. L. (2016). Spontaneous neural activity in the primary visual cortex of retinal degenerated rats. Neurosci. Lett. 623, 42–46. doi: 10.1016/j.neulet.2016.04.062
Wang, Y., West, J. D., Flashman, L. A., Wishart, H. A., Santulli, R. B., Rabin, L. A., et al. (2012). Selective changes in white matter integrity in MCI and older adults with cognitive complaints. Biochim. Biophys Acta 1822, 423–430. doi: 10.1016/j.bbadis.2011.08.002
Wong, W. L., Su, X., Li, X., Cheung, C. M., Klein, R., Cheng, C. Y., et al. (2014). Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: a systematic review and meta-analysis. Lancet Glob. Health 2, e106–116. doi: 10.1016/S2214-109X(13)70145-1
Wosniack, M. E., Kirchner, J. H., Chao, L. Y., Zabouri, N., Lohmann, C., and Gjorgjieva, J. (2021). Adaptation of spontaneous activity in the developing visual cortex. Elife. 10, e61619. doi: 10.7554/eLife.61619
Keywords: primary visual cortex, retina, retinal degeneration, electrophysiology, morphology, rodents, humans
Citation: Bhattacharyya A (2022) The detrimental effects of progression of retinal degeneration in the visual cortex. Front. Cell. Neurosci. 16:904175. doi: 10.3389/fncel.2022.904175
Received: 25 March 2022; Accepted: 04 July 2022;
Published: 29 July 2022.
Edited by:
Michel Joseph Roux, INSERM U964 Institut de Génétique et de Biologie Moléculaire et Cellulaire (IGBMC), FranceReviewed by:
Christianne E. Strang, University of Alabama at Birmingham, United StatesCopyright © 2022 Bhattacharyya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anwesha Bhattacharyya, YWJoYXR0YWNoYXJ5eWFAYW1pdHkuZWR1; YW53ZXNoYS5iaGF0dGFjaGFyeXlhMkBnbWFpbC5jb20=