 Blake J. Laham
Blake J. Laham Elizabeth Gould
Elizabeth Gould- Princeton Neuroscience Institute, Princeton University, Princeton, NJ, United States
Diffuse and structured extracellular matrix (ECM) comprise ∼20% of the brain’s volume and play important roles in development and adult plasticity. Perineuronal nets (PNNs), specialized ECM structures that surround certain types of neurons in the brain, emerge during the postnatal period, making their development and maintenance potentially sensitive to experience. Recent studies have shown that stress affects diffuse ECM as well as PNNs, and that such effects are dependent on life stage and brain region. Given that the ECM participates in synaptic plasticity, the generation of neuronal oscillations, and synchronous firing across brain regions, all of which have been linked to cognition and emotional regulation, ECM components may be candidate therapeutic targets for stress-induced neuropsychiatric disease. This review considers the influence of stress over diffuse and structured ECM during postnatal life with a focus on functional outcomes and the potential for translational relevance.
Introduction
In recent decades, it has been recognized that a comprehensive view of brain function requires considering the entirety of the brain’s microenvironment, including neurons, non-neuronal cells, the vasculature, and the extracellular matrix (ECM). Compelling evidence now suggests that each of these entities contributes not just to the brain’s structure, but to its function and its ability to respond to experience with adaptive changes (Song and Dityatev, 2018; Cope and Gould, 2019). Perhaps because of their malleability, each of these constituents has also been linked to neuropsychiatric disease. A growing literature suggests that the ECM is particularly important for a wide range of processes that are critical components of experience-dependent change in brain function, including synaptic plasticity (De Luca et al., 2020), neuronal oscillations (Carceller et al., 2020; Wingert and Sorg, 2021), and network connectivity (Bucher et al., 2021; Christensen et al., 2021). Since plasticity at the synapse, neuronal synchrony within a brain region, and coherence across brain regions are critical for healthy brain function, understanding ECM involvement in these processes may also elucidate its role in brain pathology.
The ECM includes two main categories: diffuse and structured (Nicholson and Syková, 1998; Krishnaswamy et al., 2019). Diffuse ECM fills the spaces among neurons, glia, and the brain’s microvasculature, and consists of polysaccharides, proteins, glycoproteins, and glycosaminoglycans. Although the ECM was initially thought to function primarily as a supportive substrate, essentially holding brain cells together (Celio, 1999; Cope and Gould, 2019), it is now known to play major roles in both the developing and adult brain, including the guidance of migrating neurons and growing axons, attracting and repelling astrocytes and microglia, and regulating neurotransmitter receptor availability at the synapse (Franco and Müller, 2011; Lubbers et al., 2014). Structured ECM has a similar general chemical composition to diffuse ECM, but due to a high concentration of chondroitin sulfate proteoglycans (CSPGs), it forms a lattice-like assembly around certain types of neurons, primarily inhibitory interneurons. These structures, known as perineuronal nets (PNNs), typically surround the cell bodies and proximal dendrites of neurons throughout the brain. Multiple functions have been ascribed to PNNs, including to protect neurons from free radical damage, to limit the formation of unnecessary synapses, and to functionally stabilize neuronal systems by increasing inhibitory tone after development has ended (Sorg et al., 2016; Bucher et al., 2021; Burket et al., 2021). Perhaps not surprisingly given the number of important neural processes linked to diffuse and structured ECM, studies suggest that both types are associated with key brain functions, including learning and memory, as well as emotional processing.
Stress is known to affect cognitive processes, as well as mood regulation (McEwen, 2010), raising questions about whether it does so, at least in part, by impacting the ECM. Indeed, a growing literature indicates that stress impacts the ECM both during development and in adulthood (Tables 1, 2). This mini-review first provides a general overview of studies linking the ECM to brain function during development and adulthood, and then focuses on how stress may influence both diffuse and structured ECM, including discussion about potential functional consequences.
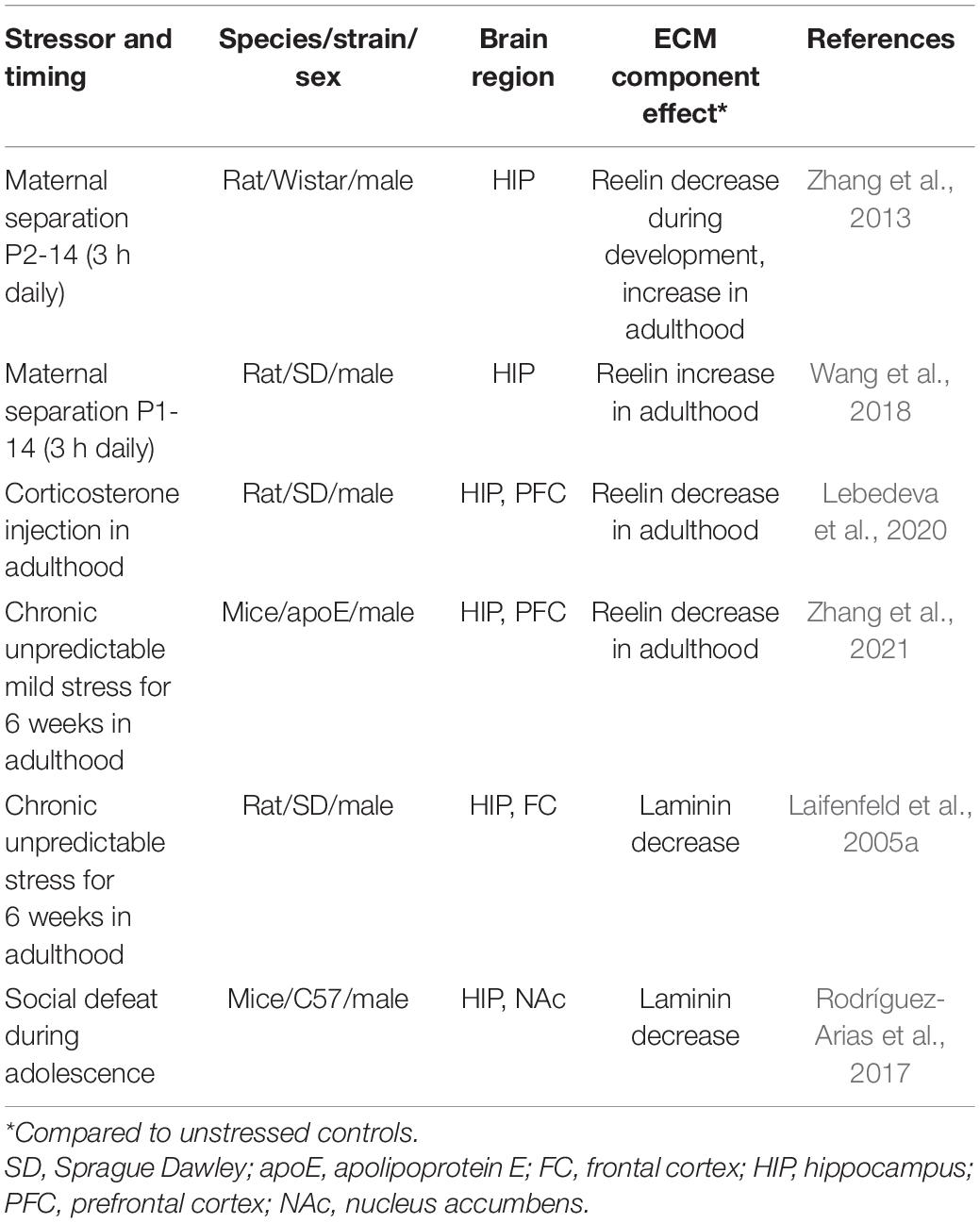
Table 1. Stress effects on diffuse ECM components.
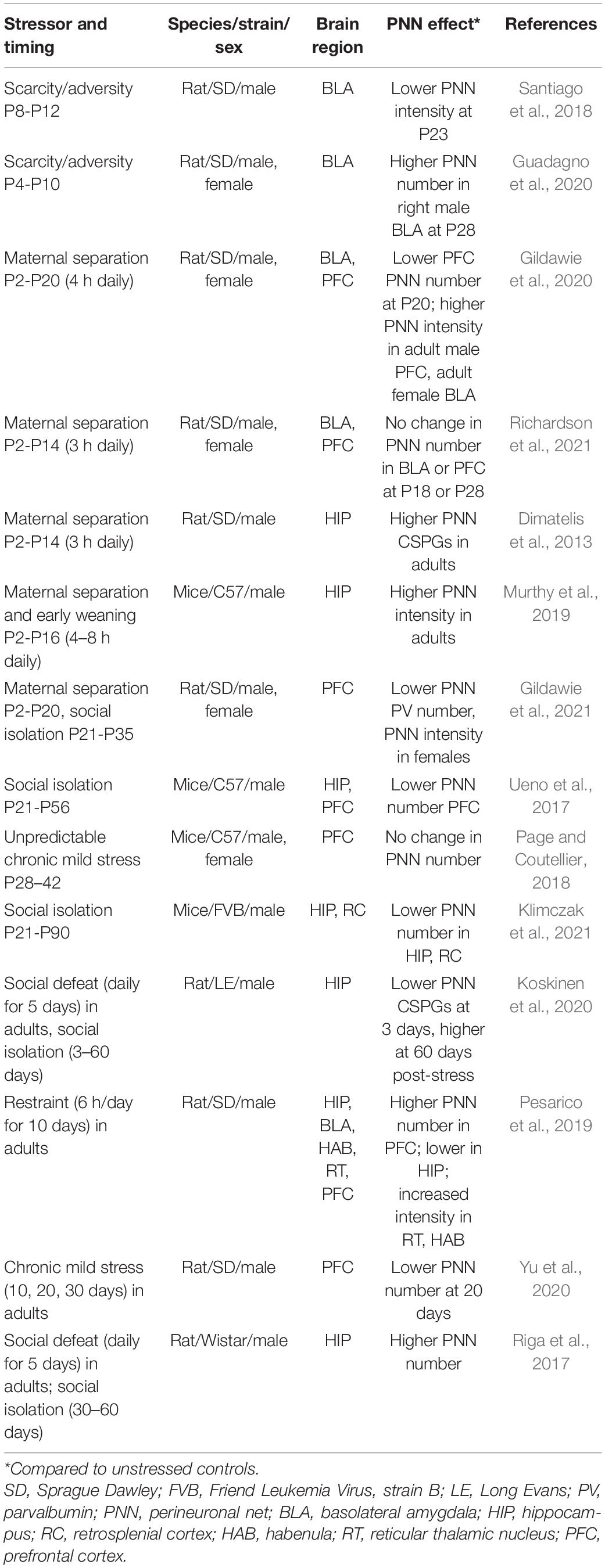
Table 2. Stress effects on PNNs.
Extracellular Matrix Function During Development and in Adulthood
Numerous studies have shown that diffuse ECM plays both subtle and vital roles in brain development. With regard to the latter, the glycoprotein laminin is critical for neural tube closure, making its knockout lethal (Miner et al., 1998). After the basic structure of the brain is formed, other ECM molecules, such as the glycoprotein reelin and the glycosaminoglycan hyaluronan, help to coordinate neuronal migration, axon guidance, and synaptogenesis (Borrell et al., 2007; Honda et al., 2011; Vaswani and Blaess, 2016). ECM molecules also sequester growth factors, chemokines, and additional molecules with attractant and repellent properties, all of which work to coordinate brain development. Diffuse CSPGs also play a role in multiple cellular events during development (Sirko et al., 2007; Zimmer et al., 2010), and their sulfation patterns are known to influence their involvement in events such as neuronal migration and maturation (Maeda et al., 2010, 2011).
As development proceeds, molecules of the diffuse ECM, such as reelin, undergo changes in abundance and function. In adulthood, reelin takes on a new function of enhancing synaptic plasticity at excitatory synapses. Reelin facilitates this function via its binding to the apolipoprotein E (apoE) receptor 2, which forms a complex with NMDA receptors (Beffert et al., 2005; Korwek et al., 2009). Reelin also functions in stimulating adult neurogenesis and dendritic spine formation in the hippocampus (Pujadas et al., 2010; Sibbe et al., 2015). Hyaluronan seems to have the opposite effect of reelin on adult neurogenesis in that its binding to the CD44 receptor reduces the production of new neurons in the hippocampus. Hyaluronan increases in the aging brain and may play a causal role in age-related reductions in adult neurogenesis (Su et al., 2017). Hyaluronan also participates in synaptic plasticity by regulating dendritic calcium channels (Kochlamazashvili et al., 2010).
During the postnatal period, the ECM surrounding a subset of neurons condense and forms PNNs, with CSPGs as major components. The lattice-like shape PNNs comes from the organization of CSPGs that bind the base of PNNs, which is hyaluronan, also a component of the diffuse ECM. The main CSPGs in the brain include aggrecan, neurocan, brevican, versican, and phosphocan, and their expression amounts are region-specific (Dauth et al., 2016; Pantazopoulos et al., 2021). Although speculative, the heterogeneous combination of PNN components may provide specialized function (Dauth et al., 2016). In addition, PNNs may have different functions depending on the sulfation patterns of their CSPGs, with some patterns conferring greater plasticity than others (Miyata et al., 2012; Yang et al., 2017).
The developmental appearance of PNNs in some brain regions coincides with the closure of critical periods, including the emergence of binocular vision in the visual cortex (Hensch and Quinlan, 2018) and leptin sensitivity in the hypothalamus (Mirzadeh et al., 2019). In adulthood, PNNs are thought to restrict plasticity, both by preventing the ingrowth of axons and the mobilization of neurotransmitter receptors at the synapse. In some cases, PNNs have been associated with increased firing of neurons they surround, which are primarily parvalbumin+ inhibitory interneurons in the neocortex and hippocampus (Sorg et al., 2016; Wingert and Sorg, 2021). The functional consequences of PNNs seem to differ depending on brain region, cell type, and behavioral task, with some studies showing that PNNs facilitate learning and memory, while others show they have an inhibitory effect on these processes (Paylor et al., 2018; Anderson et al., 2020; Carulli et al., 2020; Cope et al., 2021; Wingert and Sorg, 2021). The overall picture that is emerging is one that is common in biology—an inverted U-shaped curve exists where atypically low or high PNNs impair function. In addition to effects on behavior, PNNs have been shown to regulate neural correlates of cognitive function, including synaptic plasticity, neuronal oscillations, and neuronal synchrony across brain regions (Sorg et al., 2016; Bucher et al., 2021; Wingert and Sorg, 2021). These electrophysiological phenomena are also critical for behaviors associated with emotional regulation, and have been shown to be stress-sensitive (Murthy and Gould, 2020; Tomar et al., 2021), raising the possibility that stress-induced changes in brain function and behavior might occur through changes in the ECM. A growing body of evidence suggests that stress impacts both diffuse and structured ECM during development and in adulthood. The majority of studies investigating the effects of stress on the ECM have focused on stress-susceptible brain regions, including the hippocampus, the prefrontal cortex, and the amygdala, which play crucial roles in cognitive function and emotional processing (McEwen et al., 2015; Smith and Pollak, 2020).
Stress Effects on Diffuse Extracellular Matrix
Developmental stress exhibits different effects on reelin signaling depending on the age at which the brains are investigated (Table 1). Postnatal stress has been shown to first reduce reelin expression in the hippocampus, and then show a compensatory rebound and overshoot as animals reach adulthood (Zhang et al., 2013). The postnatal stress-induced increase in adult hippocampal reelin expression can be augmented by exposure to a stressful learning paradigm, such as contextual fear conditioning. This effect is accompanied by enhanced hippocampal LTP and dendritic spine density (Wang et al., 2018). These findings suggest that postnatal stress-induced latent increases in reelin expression may serve an adaptive function. In contrast, chronic stress or chronic glucocorticoid treatment in adulthood decrease reelin expression in the hippocampus and prefrontal cortex (Lebedeva et al., 2020; Zhang et al., 2021). Studies additionally suggest that reelin signaling through the apoE receptor is important for mitigating stress-induced behavioral dysfunction, especially in older mice (Zhang et al., 2021). These findings suggest that developmental and acute adult stress may produce adaptive stress responses through augmented reelin signaling, while chronic adult stress may lead to dysfunction via a reduction in reelin expression. Along these lines, it may be relevant that antidepressant drug treatment reverses stress-induced decreases in reelin expression (Fenton et al., 2015; Johnston et al., 2020), and that hippocampal reelin infusions can reverse stress-induced behavioral dysfunction (Brymer et al., 2020).
Laminin expression is also decreased after chronic stress in both the adult hippocampus and frontal cortex (Laifenfeld et al., 2005a; Rodríguez-Arias et al., 2017). Similar to what has been observed with reelin, the stress-induced decrease in laminin can be reversed with antidepressant treatment (Laifenfeld et al., 2005a). Hyaluronan signaling also seems to play a protective role in mediating stress effects, as mice lacking the hyaluronan receptor CD44 exhibit exacerbated stress-induced behavioral dysfunction, as well as reduced brain levels of the neuromodulators serotonin and dopamine (Barzilay et al., 2016). Collectively, the overall picture suggests that components of diffuse ECM are stress-sensitive and potentially involved in adaptive mechanisms enhancing the ability to appropriately respond to stress and buffer against stress-induced pathology.
Stress Effects on Perineuronal Nets
Several studies have investigated the effects of postnatal and adult stress on PNNs (Table 2). These studies have produced mixed results, likely due to differences in the developmental stage of stress exposure, the type of stressor, as well as the duration of time between stress and brain examination. An additional reason for potential discrepancies may be due to measures used to assess PNNs, which most commonly rely on the binding of an exogenous fluorophore-conjugated plant lectin Wisteria floribunda agglutinin (WFA). Using this approach, researchers most often quantify numbers of WFA+ cells or the intensity of WFA+ cells to assess whether PNNs have changed. WFA is not an endogenous component of PNNs, and although its binding site is known (Nadanaka et al., 2020), it does not label all PNNs (Yamada and Jinno, 2017; Ueno et al., 2018). Thus, changes in WFA labeling may be open to multiple interpretations. One study showed that postnatal stress using a scarcity/adversity model led to reduced PNN intensity in a subregion of the basolateral amygdala (Santiago et al., 2018), while another study using the same model reported increased PNN cell numbers but only on the right side of the amygdala in males (Guadagno et al., 2020). Two additional studies using maternal separation have reported either increased PNN intensity, but only in females (Gildawie et al., 2020), or no differences in PNN measures (Richardson et al., 2021). Although all of these studies used a similar method for identifying PNNs, they did not all use the same measure (WFA+ cell number vs. WFA intensity), and none used a label of a specific CSPG, such as aggrecan. It may be relevant that these studies also examined different time points after postnatal stress (Table 2), making it difficult to determine whether the effects would be more similar if the same time point had been examined.
Available evidence suggests a complex set of results from stress studies in the hippocampus and prefrontal cortex, with some studies showing decreased number or intensity of PNNs, others an increase, and others reporting no effects at all (Table 2). Examining the papers as a group, however, suggests that stress may have variable effects on PNNs depending on the duration of time after stress exposure. Several studies show decreased PNNs immediately after stress (Ueno et al., 2017; Pesarico et al., 2019; Gildawie et al., 2020, 2021; Koskinen et al., 2020; Yu et al., 2020; Klimczak et al., 2021), and a rebound increase in PNNs as time passes following stress cessation (Dimatelis et al., 2013; Riga et al., 2017; Murthy et al., 2019; Koskinen et al., 2020; Gildawie et al., 2021). Although some findings do not fit with this summary (Page and Coutellier, 2018; Richardson et al., 2021), the majority of studies suggest a stress-induced trajectory involving first suppression followed by an overshooting rebound, similar to what has been observed for reelin signaling (Zhang et al., 2013). This is in line with other theories of stress effects on the ECM, particularly the idea that depressive-like symptoms emerge after an “incubation period,” which involves latent increases in PNNs and accompanying plasticity reduction (Koskinen et al., 2020; Spijker et al., 2020). It should be recognized that outside of critical periods, PNNs are capable of rapid remodeling, as has been recently demonstrated in several studies (Marchand and Schwartz, 2020; Pantazopoulos et al., 2020; Uriarte et al., 2020). Evidence suggests that changes in PNNs can occur through alterations in neuronal activity both during development and in adulthood (Dityatev et al., 2007; Carstens et al., 2021; Devienne et al., 2021). Thus, stress-induced changes in neuronal activity (Della Valle et al., 2019; Murthy et al., 2019; Del Arco et al., 2020; Fee et al., 2020) could be a mechanism by which alterations in PNNs occur.
The extent to which stress-induced changes in PNNs represent adaptive or dysfunctional effects remains uncertain. Several studies show that reduced or increased PNN measures in the hippocampus, prefrontal cortex, and amygdala are associated with behavioral changes that are thought to reflect increased avoidance/threat and reduced stress coping behavior (Santiago et al., 2018; Murthy et al., 2019; Koskinen et al., 2020; Yu et al., 2020), as well as impaired cognitive function (Riga et al., 2017; Koskinen et al., 2020). Few studies have addressed causal relationships by including experimental manipulations of PNNs that restore healthy function after stress (Riga et al., 2017). Additional studies in non-stressed rodents have shown that reducing PNNs either by genetic manipulations or by enzymatic degradation can alter stress-susceptible behaviors in rodents, including avoidance, stress coping, cognitive function and substance use. For example, genetic deletion of neuronal membrane linking protein ankyrin-R or the transcription factor OTX2 reduces PNN expression and decreases avoidance of the open arms in an elevated plus maze task (Stevens et al., 2021; Vincent et al., 2021). Furthermore, degradation of PNNs using chondroitinase ABC facilitates extinction of drug-seeking behavior (Xue et al., 2014) and prevents both fear conditioning (Hylin et al., 2013) and cocaine-induced place preference (Slaker et al., 2015). However, studies have also shown that diminished PNNs can produce effects that mimic those of chronic stress, including increased threat responses (Santiago et al., 2018) and diminished cognitive function (Paylor et al., 2018). Since atypical behavioral states have been associated with both reduced or increased PNN measures in several brain regions, it seems likely that an optimal level of PNNs within a brain region may exist, which when disrupted produces behavioral dysfunction. Along these lines, it has been shown that antidepressant action on behavior and neuronal oscillations both require the presence of PNNs in the hippocampus (Donegan and Lodge, 2017), as well as the degradation of PNNs through the antidepressant-induced release of proteolytic enzymes by microglia (Alaiyed et al., 2019, 2020). Clearly, additional research is needed to better understand links among stress, PNNs, and behavior.
Bridging the Gap Between Stress-Induced Changes in Extracellular Matrix and Behavioral Outcomes
Despite the variability of outcomes regarding stress effects on diffuse and structured ECM, both forms are sensitive to stress and their changes have been linked to alterations in behaviors associated with cognition and emotional processing. These findings raise questions about the mechanisms by which stress-induced ECM remodeling give rise to behavioral change. It is likely that the larger literature on the role of the ECM in electrophysiological function of relevant brain regions may provide clues. As mentioned earlier, both diffuse ECM and PNNs regulate synaptic plasticity (Sorg et al., 2016; Jakob et al., 2017), which has clear links to cognitive function (Dringenberg, 2020), raising a scenario whereby stress produces cognitive dysfunction by disrupting ECM components associated with optimal synaptic plasticity. ECM has also been linked to neuronal oscillations in both the gamma and theta frequency ranges (Murthy and Gould, 2020), which are important for both cognitive function (Mably and Colgin, 2018; Zielinski et al., 2020) and behaviors associated with emotional processing, including avoidance behavior (Padilla-Coreano et al., 2019). These results suggest that stress may produce cognitive dysfunction and enhance avoidance behavior through ECM-induced changes in neuronal oscillations. The ECM has also been linked to synchrony in rhythmic signaling across brain regions (Bucher et al., 2021), which has been shown to play crucial roles in healthy brain function in terms of learning and memory, as well as stress responsivity (Adhikari et al., 2010; Del Arco et al., 2020). It is conceivable that stress-induced ECM changes in these electrophysiological properties are responsible for stress-induced behavioral outcomes. Since antidepressant treatment restores stress-induced ECM changes and behavioral dysfunction in rodents, it seems plausible that this occurs through reversal of atypical synaptic plasticity, neuronal oscillations and/or circuit-level coherence. Antidepressant treatment has been shown to influence all of these electrophysiological measures (Law et al., 2016; Alaiyed et al., 2019; Logue et al., 2021).
Future Studies
Future studies should directly investigate whether stress-induced changes in ECM lead to alterations in behavior through influences on electrophysiological properties at the synapse, among populations of neurons within a brain region, and across brain regions in the broader circuitry. Although these studies are likely to be informative, they will not provide a complete picture without considering other aspects of the brain’s microenvironment, such as glia, which are also known to be stress-responsive (Kaul et al., 2021) and participate in ECM remodeling (Strackeljan et al., 2021). Some studies have begun to test the involvement of microglia in regulating ECM in the context of both cognitive function (Alaiyed et al., 2019; Nguyen et al., 2020; Venturino et al., 2021) and emotional processing (Alaiyed et al., 2019; Venturino et al., 2021). Expanding these approaches to other brain regions, as well as other stress paradigms, should be illuminating. Lastly, finding ways to connect the experimental animal literature to humans will be important. Along these lines, it is relevant to note that the components of diffuse and structured ECM described in the rodent brain exist in the human brain (Fatemi et al., 2000; Laifenfeld et al., 2005b; Mauney et al., 2013). Furthermore, major depressive disorder and bipolar disorder, conditions linked to stress, have been associated with altered levels of diffuse ECM molecules, such as reelin, hyaluronan, and laminin (Fatemi et al., 2000; Guidotti et al., 2000; Laifenfeld et al., 2005b; Lubbers et al., 2014; Ventorp et al., 2016), and increased PNNs have been reported in the prefrontal cortex of suicide victims previously exposed to childhood maltreatment (Tanti et al., 2020). The similarities between rodent and human studies increase confidence that a better understanding of the connections between the ECM and stress-induced behavioral dysfunction in rodents may provide a window into stress-induced neuropsychiatric disease in humans.
Author Contributions
BL and EG wrote and edited the manuscript. Both authors contributed to the article and approved the submitted version.
Funding
This work was supported by the NIMH MH117459-01 (EG).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adhikari, A., Topiwala, M. A., and Gordon, J. A. (2010). Synchronized activity between the ventral hippocampus and the medial prefrontal cortex during anxiety. Neuron 65, 257–269. doi: 10.1016/j.neuron.2009.12.002
Alaiyed, S., Bozzelli, P. L., Caccavano, A., Wu, J. Y., and Conant, K. (2019). Venlafaxine stimulates PNN proteolysis and MMP-9-dependent enhancement of gamma power; relevance to antidepressant efficacy. J. Neurochem. 148, 810–821. doi: 10.1111/jnc.14671
Alaiyed, S., McCann, M., Mahajan, G., Rajkowska, G., Stockmeier, C. A., Kellar, K. J., et al. (2020). Venlafaxine stimulates an MMP-9-dependent increase in excitatory/inhibitory balance in a stress model of depression. J. Neurosci. 40, 4418–4431. doi: 10.1523/JNEUROSCI.2387-19.2020
Anderson, M. D., Paylor, J. W., Scott, G. A., Greba, Q., Winship, I. R., and Howland, J. G. (2020). ChABC infusions into medial prefrontal cortex, but not posterior parietal cortex, improve the performance of rats tested on a novel, challenging delay in the touchscreen TUNL task. Learn. Mem. 27, 222–235. doi: 10.1101/lm.050245.119
Barzilay, R., Ventorp, F., Segal-Gavish, H., Aharony, I., Bieber, A., Dar, S., et al. (2016). CD44 deficiency is associated with increased susceptibility to stress-induced anxiety-like behavior in mice. J. Mol. Neurosci. 60, 548–558. doi: 10.1007/s12031-016-0835-3
Beffert, U., Weeber, E. J., Durudas, A., Qiu, S., Masiulis, I., Sweatt, J. D., et al. (2005). Modulation of synaptic plasticity and memory by Reelin involves differential splicing of the lipoprotein receptor Apoer2. Neuron 47, 567–579. doi: 10.1016/j.neuron.2005.07.007
Borrell, V., Pujadas, L., Simó, S., Durà, D., Solé, M., Cooper, J. A., et al. (2007). Reelin and mDab1 regulate the development of hippocampal connections. Mol. Cell. Neurosci. 36, 158–173. doi: 10.1016/j.mcn.2007.06.006
Brymer, K. J., Johnston, J., Botterill, J. J., Romay-Tallon, R., Mitchell, M. A., Allen, J., et al. (2020). Fast-acting antidepressant-like effects of Reelin evaluated in the repeated-corticosterone chronic stress paradigm. Neuropsychopharm 45, 1707–1716. doi: 10.1038/s41386-020-0609-z
Bucher, E. A., Collins, J. M., King, A. E., Vickers, J. C., and Kirkcaldie, M. T. K. (2021). Coherence and cognition in the cortex: the fundamental role of parvalbumin, myelin, and the perineuronal net. Brain Struct. Funct. 226, 2041–2055. doi: 10.1007/s00429-021-02327-3
Burket, J. A., Webb, J. D., and Deutsch, S. I. (2021). Perineuronal nets and metal cation concentrations in the microenvironments of fast-spiking, parvalbumin-expressing GABAergic interneurons: relevance to neurodevelopment and neurodevelopmental disorders. Biomolecules 11:1235. doi: 10.3390/biom11081235
Carceller, H., Guirado, R., Ripolles-Campos, E., Teruel-Marti, V., and Nacher, J. (2020). Perineuronal nets regulate the inhibitory perisomatic input onto parvalbumin interneurons and γ activity in the prefrontal cortex. J. Neurosci. 40, 5008–5018. doi: 10.1523/JNEUROSCI.0291-20.2020
Carstens, K. E., Lustberg, D. J., Shaughnessy, E. K., McCann, K. E., Alexander, G. M., and Dudek, S. M. (2021). Perineuronal net degradation rescues CA2 plasticity in a mouse model of Rett syndrome. J. Clin. Invest. 131:e137221. doi: 10.1172/JCI137221
Carulli, D., Broersen, R., de Winter, F., Muir, E. M., Meşković, M., de Waal, M., et al. (2020). Cerebellar plasticity and associative memories are controlled by perineuronal nets. Proc. Natl. Acad. Sci. 117, 6855–6865. doi: 10.1073/pnas.1916163117
Celio, M. R. (1999). Evolution of the concept of “extracellular matrix’ in the brain. J. Hist. Neurosci. 8, 186–190. doi: 10.1076/jhin.8.2.186.1832
Christensen, A. C., Lensjø, K. K., Lepperød, M. E., Dragly, S. A., Sutterud, H., Blackstad, J. S., et al. (2021). Perineuronal nets stabilize the grid cell network. Nat. Commun. 12:253. doi: 10.1038/s41467-020-20241-w
Cope, E. C., and Gould, E. (2019). Adult neurogenesis, glia, and the extracellular matrix. Cell Stem Cell 24, 690–705. doi: 10.1016/j.stem.2019.03.023
Cope, E. C., Zych, A. D., Katchur, N. J., Waters, R. C., Laham, B. J., Diethorn, E. J., et al. (2021). Atypical perineuronal nets in the CA2 region interfere with social memory in a mouse model of social dysfunction. Mol. Psychiatry. doi: 10.1038/s41380-021-01174-2
Dauth, S., Grevesse, T., Pantazopoulos, H., Campbell, P. H., Maoz, B. M., Berretta, S., et al. (2016). Extracellular matrix protein expression is brain region dependent. J. Comp. Neurol. 524, 1309–1336. doi: 10.1002/cne.23965
De Luca, C., Colangelo, A. M., Virtuoso, A., Alberghina, L., and Papa, M. (2020). Neurons, glia, extracellular matrix and neurovascular unit: a systems biology approach to the complexity of synaptic plasticity in health and disease. Int. J. Mol. Sci. 21:1539. doi: 10.3390/ijms21041539
Del Arco, A., Park, J., and Moghaddam, B. (2020). Unanticipated stressful and rewarding experiences engage the same prefrontal cortex and ventral tegmental area neuronal populations. eNeuro 7, ENEURO.29-ENEURO.20. doi: 10.1523/ENEURO.0029-20.2020
Della Valle, R., Mohammadmirzaei, N., and Knox, D. (2019). Single prolonged stress alters neural activation in the periacqueductal gray and midline thalamic nuclei during emotional learning and memory. Learn. Mem. 26, 1–9. doi: 10.1101/lm.050310.119
Devienne, G., Picaud, S., Cohen, I., Piquet, J., Tricoire, L., Testa, D., et al. (2021). Regulation of perineuronal nets in the adult cortex by the activity of the cortical network. J. Neurosci. 41, 5779–5790. doi: 10.1523/jneurosci.0434-21.2021
Dimatelis, J. J., Hendricks, S., Hsieh, J., Vlok, N. M., Bugarith, K., Daniels, W. M., et al. (2013). Exercise partly reverses the effect of maternal separation on hippocampal proteins in 6-hydroxydopamine-lesioned rat brain. Exp. Physiol. 98, 233–244. doi: 10.1113/expphysiol.2012.066720
Dityatev, A., Brückner, G., Dityateva, G., Grosche, J., Kleene, R., and Schachner, M. (2007). Activity-dependent formation and functions of chondroitin sulfate-rich extracellular matrix of perineuronal nets. Dev. Neurobiol. 67, 570–588. doi: 10.1002/dneu.20361
Donegan, J. J., and Lodge, D. J. (2017). Hippocampal perineuronal nets are required for the sustained antidepressant effect of ketamine. Int. J. Neuropsychopharmacol. 20, 354–358. doi: 10.1093/ijnp/pyw095
Dringenberg, H. C. (2020). The history of long-term potentiation as a memory mechanism: controversies, confirmation, and some lessons to remember. Hippocampus 30, 987–1012. doi: 10.1002/hipo.23213
Fatemi, S. H., Earle, J. A., and McMenomy, T. (2000). Reduction in Reelin immunoreactivity in hippocampus of subjects with schizophrenia, bipolar disorder and major depression. Mol. Psychiatry 5, 654–663. doi: 10.1038/sj.mp.4000783
Fee, C., Prevot, T., Misquitta, K., Banasr, M., and Sibille, E. (2020). Chronic stress-induced behaviors correlate with exacerbated acute stress-induced cingulate cortex and ventral hippocampus activation. Neuroscience 440, 113–129. doi: 10.1016/j.neuroscience.2020.05.034
Fenton, E. Y., Fournier, N. M., Lussier, A. L., Romay-Tallon, R., Caruncho, H. J., and Kalynchuk, L. E. (2015). Imipramine protects against the deleterious effects of chronic corticosterone on depression-like behavior, hippocampal reelin expression, and neuronal maturation. Prog. Neuropsychopharm. Biol. Psych. 60, 52–59. doi: 10.1016/j.pnpbp.2015.02.001
Franco, S. J., and Müller, U. (2011). Extracellular matrix functions during neuronal migration and lamination in the mammalian central nervous system. Dev. Neurobiol. 71, 889–900. doi: 10.1002/dneu.20946
Gildawie, K. R., Honeycutt, J. A., and Brenhouse, H. C. (2020). Region-specific effects of maternal separation on perineuronal net and parvalbumin-expressing interneuron formation in male and female rats. Neuroscience 428, 23–37. doi: 10.1016/j.neuroscience.2019.12.010
Gildawie, K. R., Ryll, L. M., Hexter, J. C., Peterzell, S., Valentine, A. A., and Brenhouse, H. C. (2021). A two-hit adversity model in developing rats reveals sex-specific impacts on prefrontal cortex structure and behavior. Dev. Cog. Neurosci. 48:100924. doi: 10.1016/j.dcn.2021.100924
Guadagno, A., Verlezza, S., Long, H., Wong, T. P., and Walker, C. D. (2020). It is all in the right amygdala: increased synaptic plasticity and perineuronal nets in male, but not female, juvenile rat pups after exposure to early-life stress. J Neurosci. 40, 8276–8291. doi: 10.1523/JNEUROSCI.1029-20.2020
Guidotti, A., Auta, J., Davis, J. M., Di-Giorgi-Gerevini, V., Dwivedi, Y., Grayson, D. R., et al. (2000). Decrease in reelin and glutamic acid decarboxylase 67 (GAD67) expression in schizophrenia and bipolar disorder: a postmortem brain study. Arch. Gen. Psych. 57, 1061–1069. doi: 10.1001/archpsyc.57.11.1061
Hensch, T. K., and Quinlan, E. M. (2018). Critical periods in amblyopia. Vis. Neurosci. 35:E014. doi: 10.1017/S0952523817000219
Honda, T., Kobayashi, K., Mikoshiba, K., and Nakajima, K. (2011). Regulation of cortical neuron migration by the Reelin signaling pathway. Neurochem. Res. 36, 1270–1279. doi: 10.1007/s11064-011-0407-4
Hylin, M. J., Orsi, S. A., Moore, A. N., and Dash, P. K. (2013). Disruption of the perineuronal net in the hippocampus or medial prefrontal cortex impairs fear conditioning. Learn. Mem. 20, 267–273. doi: 10.1101/lm.030197.112
Jakob, B., Kochlamazashvili, G., Jäpel, M., Gauhar, A., Bock, H. H., Maritzen, T., et al. (2017). Intersectin 1 is a component of the Reelin pathway to regulate neuronal migration and synaptic plasticity in the hippocampus. Proc. Natl. Acad. Sci. U.S.A. 114, 5533–5538. doi: 10.1073/pnas.1704447114
Johnston, J. N., Thacker, J. S., Desjardins, C., Kulyk, B. D., Romay-Tallon, R., Kalynchuk, L. E., et al. (2020). Ketamine rescues hippocampal reelin expression and synaptic markers in the repeated-corticosterone chronic stress paradigm. Front. Pharmacol. 11:559627. doi: 10.3389/fphar.2020.559627
Kaul, D., Schwab, S. G., Mechawar, N., and Matosin, N. (2021). How stress physically re-shapes the brain: impact on brain cell shapes, numbers and connections in psychiatric disorders. Neurosci. Biobehav. Rev. 124, 193–215. doi: 10.1016/j.neubiorev.2021.01.025
Klimczak, P., Rizzo, A., Castillo-Gómez, E., Perez-Rando, M., Gramuntell, Y., Beltran, M., et al. (2021). Parvalbumin interneurons and perineuronal nets in the hippocampus and retrosplenial cortex of adult male mice after early social isolation stress and perinatal NMDA receptor antagonist treatment. Front. Synaptic Neurosci. 13:733989. doi: 10.3389/fnsyn.2021.733989
Kochlamazashvili, G., Henneberger, C., Bukalo, O., Dvoretskova, E., Senkov, O., Lievens, P. M., et al. (2010). The extracellular matrix molecule hyaluronic acid regulates hippocampal synaptic plasticity by modulating postsynaptic L-type Ca(2+) channels. Neuron 67, 116–128. doi: 10.1016/j.neuron.2010.05.030
Korwek, K. M., Trotter, J. H., Ladu, M. J., Sullivan, P. M., and Weeber, E. J. (2009). ApoE isoform-dependent changes in hippocampal synaptic function. Mol. Neurodegener. 4:21. doi: 10.1186/1750-1326-4-21
Koskinen, M.-K., Mourik, Y., van Smit, A. B., Riga, D., and Spijker, S. (2020). From stress to depression: development of extracellular matrix-dependent cognitive impairment following social stress. Sci. Rep. 10:17308. doi: 10.1038/s41598-020-73173-2
Krishnaswamy, V. R., Benbenishty, A., Blinder, P., and Sagi, I. (2019). Demystifying the extracellular matrix and its proteolytic remodeling in the brain: structural and functional insights. Cell. Mol. Life Sci. 76, 3229–3248. doi: 10.1007/s00018-019-03182-6
Laifenfeld, D., Karry, R., Grauer, E., Klein, E., and Ben-Shachar, D. (2005a). Antidepressants and prolonged stress in rats modulate CAM-L1, laminin, and pCREB, implicated in neuronal plasticity. Neurobiol. Dis. 20, 432–441. doi: 10.1016/j.nbd.2005.03.023
Laifenfeld, D., Karry, R., Klein, E., and Ben-Shachar, D. (2005b). Alterations in cell adhesion molecule L1 and functionally related genes in major depression: a postmortem study. Biol. Psych. 57, 716–725. doi: 10.1016/j.biopsych.2004.12.016
Law, J., Ibarguen-Vargas, Y., Belzung, C., and Surget, A. (2016). Decline of hippocampal stress reactivity and neuronal ensemble coherence in a mouse model of depression. Psychoneuroendocrinology 67, 113–123. doi: 10.1016/j.psyneuen.2016.01.028
Lebedeva, K. A., Allen, J., Kulhawy, E. Y., Caruncho, H. J., and Kalynchuk, L. E. (2020). Cyclical administration of corticosterone results in aggravation of depression-like behaviors and accompanying downregulations in reelin in an animal model of chronic stress relevant to human recurrent depression. Physiol. Behav. 224:113070. doi: 10.1016/j.physbeh.2020.113070
Logue, J., Schoepfer, K., Guerrero, A. B., Zhou, Y., and Kabbaj, M. (2021). Sex-specific effects of social isolation stress and ketamine on hippocampal plasticity. Neurosci. Lett. 766:136301. doi: 10.1016/j.neulet.2021.136301
Lubbers, B. R., Smit, A. B., Spijker, S., and van den Oever, M. C. (2014). Neural ECM in addiction, schizophrenia, and mood disorder. Prog. Brain Res. 214, 263–284. doi: 10.1016/B978-0-444-63486-3.00012-8
Mably, A. J., and Colgin, L. L. (2018). Gamma oscillations in cognitive disorders. Curr. Opin. Neurobiol. 52, 182–187. doi: 10.1016/j.conb.2018.07.009
Maeda, N., Fukazawa, N., and Ishii, M. (2010). Chondroitin sulfate proteoglycans in neural development and plasticity. Front. Biosci. 15, 626–644. doi: 10.2741/3637
Maeda, N., Ishii, M., Nishimura, K., and Kamimura, K. (2011). Functions of chondroitin sulfate and heparan sulfate in the developing brain. Neurochem. Res. 36, 1228–1240. doi: 10.1007/s11064-010-0324-y
Marchand, A., and Schwartz, C. (2020). Perineuronal net expression in the brain of a hibernating mammal. Brain Struct. Funct. 225, 45–56. doi: 10.1007/s00429-019-01983-w
Mauney, S. A., Athanas, K. M., Pantazopoulos, H., Shaskan, N., Passeri, E., Berretta, S., et al. (2013). Developmental pattern of perineuronal nets in the human prefrontal cortex and their deficit in schizophrenia. Biol. Psychiatry 74, 427–435. doi: 10.1016/j.biopsych.2013.05.007
McEwen, B. S. (2010). Stress, sex, and neural adaptation to a changing environment: mechanisms of neuronal remodeling. Ann. N. Y. Acad .Sci. 1204, E38–E59. doi: 10.1111/j.1749-6632.2010.05568.x
McEwen, B. S., Bowles, N. P., Gray, J. D., Hill, M. N., Hunter, R. G., Karatsoreos, I. N., et al. (2015). Mechanisms of stress in the brain. Nat. Neurosci. 18, 1353–1363. doi: 10.1038/nn.4086
Miner, J. H., Cunningham, J., and Sanes, J. R. (1998). Roles for laminin in embryogenesis: exencephaly, syndactyly, and placentopathy in mice lacking the laminin alpha5 chain. J. Cell Biol. 143, 1713–1723. doi: 10.1083/jcb.143.6.1713
Mirzadeh, Z., Alonge, K. M., Cabrales, E., Herranz-Pérez, V., Scarlett, J. M., Brown, J. M., et al. (2019). Perineuronal net formation during the critical period for neuronal maturation in the hypothalamic arcuate nucleus. Nat. Metab. 1, 212–221. doi: 10.1038/s42255-018-0029-0
Miyata, S., Komatsu, Y., Yoshimura, Y., Taya, C., and Kitagawa, H. (2012). Persistent cortical plasticity by upregulation of chondroitin 6-sulfation. Nat. Neurosci. 15, 414–122, S1–S2. doi: 10.1038/nn.3023
Murthy, S., and Gould, E. (2020). How early life adversity influences defensive circuitry. Trends Neurosci. 43, 200–212. doi: 10.1016/j.tins.2020.02.001
Murthy, S., Kane, G. A., Katchur, N. J., Mejia, P. S. L., Obiofuma, G., Buschman, T. J., et al. (2019). Perineuronal nets, inhibitory interneurons and anxiety-related ventral hippocampal neuronal oscillations are altered by early life adversity. Biol. Psych. 85, 1011–1020. doi: 10.1016/j.biopsych.2019.02.021
Nadanaka, S., Miyata, S., Yaqiang, B., Tamura, J. I., Habuchi, O., and Kitagawa, H. (2020). Reconsideration of the semaphorin-3A binding motif found in chondroitin sulfate using Galnac4s-6st-knockout mice. Biomolecules 10:1499. doi: 10.3390/biom10111499
Nguyen, P. T., Dorman, L. C., Pan, S., Vainchtein, I. D., Han, R. T., Nakao-Inoue, H., et al. (2020). Microglial remodeling of the extracellular matrix promotes synapse plasticity. Cell 182, 388–403.e15. doi: 10.1016/j.cell.2020.05.050
Nicholson, C., and Syková, E. (1998). Extracellular space structure revealed by diffusion analysis. Trends Neurosci. 21, 207–215. doi: 10.1016/s0166-2236(98)01261-2
Padilla-Coreano, N., Canetta, S., Mikofsky, R. M., Alway, E., Passecker, J., Myroshnychenko, M. V., et al. (2019). Hippocampal-prefrontal theta transmission regulates avoidance behavior. Neuron 104, 601–610.e4. doi: 10.1016/j.neuron.2019.08.006
Page, C. E., and Coutellier, L. (2018). Adolescent stress disrupts the maturation of anxiety-related behaviors and alters the developmental trajectory of the prefrontal cortex in a sex- and age-specific manner. Neuroscience. 390, 265–277. doi: 10.1016/j.neuroscience.2018.08.030
Pantazopoulos, H., Gisabella, B., Rexrode, L., Benefield, D., Yildiz, E., Seltzer, P., et al. (2020). Circadian rhythms of perineuronal net composition. eNeuro 7, ENEURO.34-ENEURO.19. doi: 10.1523/eneuro.0034-19.2020
Pantazopoulos, H., Katsel, P., Haroutunian, V., Chelini, G., Klengel, T., and Berretta, S. (2021). Molecular signature of extracellular matrix pathology in schizophrenia. Eur. J. Neurosci. 53, 3960–3987. doi: 10.1111/ejn.15009
Paylor, J. W., Wendlandt, E., Freeman, T. S., Greba, Q., Marks, W. N., Howland, J. G., et al. (2018). Impaired cognitive function after perineuronal net degradation in the medial prefrontal cortex. eNeuro 5, ENEURO.253-ENEURO.218. doi: 10.1523/ENEURO.0253-18.2018
Pesarico, A. P., Bueno-Fernandez, C., Guirado, R., Gómez-Climent, M. Á, Curto, Y., Carceller, H., et al. (2019). Chronic stress modulates interneuronal plasticity: effects on PSA-NCAM and perineuronal nets in cortical and extracortical regions. Front. Cell. Neurosci. 13:197. doi: 10.3389/fncel.2019.00197
Pujadas, L., Gruart, A., Bosch, C., Delgado, L., Teixeira, C. M., Rossi, D., et al. (2010). Reelin regulates postnatal neurogenesis and enhances spine hypertrophy and long-term potentiation. J. Neurosci. 30, 4636–4649. doi: 10.1523/JNEUROSCI.5284-09.2010
Richardson, R., Bowers, J., Callaghan, B. L., and Baker, K. D. (2021). Does maternal separation accelerate maturation of perineuronal nets and parvalbumin-containing inhibitory interneurons in male and female rats? Dev. Cogn. Neurosci. 47:100905. doi: 10.1016/j.dcn.2020.100905
Riga, D., Kramvis, I., Koskinen, M. K., van Bokhoven, P., van der Harst, J. E., Heistek, T. S., et al. (2017). Hippocampal extracellular matrix alterations contribute to cognitive impairment associated with a chronic depressive-like state in rats. Sci. Transl. Med. 9:eaai8753. doi: 10.1126/scitranslmed.aai8753
Rodríguez-Arias, M., Montagud-Romero, S., Rubio-Araiz, A., Aguilar, M. A., Martín-García, E., Cabrera, R., et al. (2017). Effects of repeated social defeat on adolescent mice on cocaine-induced CPP and self-administration in adulthood: integrity of the blood-brain barrier. Addict. Biol. 22, 129–141. doi: 10.1111/adb.12301
Santiago, A. N., Lim, K. Y., Opendak, M., Sullivan, R. M., and Aoki, C. (2018). Early life trauma increases threat response of peri-weaning rats, reduction of axo-somatic synapses formed by parvalbumin cells and perineuronal net in the basolateral nucleus of amygdala. J. Comp. Neurol. 16, 2647–2664. doi: 10.1002/cne.24522
Sibbe, M., Kuner, E., Althof, D., and Frotscher, M. (2015). Stem- and progenitor cell proliferation in the dentate gyrus of the reeler mouse. PLoS One 10:e0119643. doi: 10.1371/journal.pone.0119643
Sirko, S., von Holst, A., Wizenmann, A., Götz, M., and Faissner, A. (2007). Chondroitin sulfate glycosaminoglycans control proliferation, radial glia cell differentiation and neurogenesis in neural stem/progenitor cells. Development 134, 2727–2738. doi: 10.1242/dev.02871
Slaker, M., Churchill, L., Todd, R. P., Blacktop, J. M., Zuloaga, D. G., Raber, J., et al. (2015). Removal of perineuronal nets in the medial prefrontal cortex impairs the acquisition and reconsolidation of a cocaine-induced conditioned place preference memory. J. Neurosci. 35, 4190–4202. doi: 10.1523/JNEUROSCI.3592-14.2015
Smith, K. E., and Pollak, S. D. (2020). Early life stress and development: potential mechanisms for adverse outcomes. J. Neurodevelop. Dis. 12:34. doi: 10.1186/s11689-020-09337-y
Song, I., and Dityatev, A. (2018). Crosstalk between glia, extracellular matrix and neurons. Brain Res. Bull. 136, 101–108. doi: 10.1016/j.brainresbull.2017.03.003
Sorg, B. A., Berretta, S., Blacktop, J. M., Fawcett, J. W., Kitagawa, H., Kwok, J. C., et al. (2016). Casting a wide net: role of perineuronal nets in neural plasticity. J. Neurosci. 36, 11459–11468. doi: 10.1523/JNEUROSCI.2351-16.2016
Spijker, S., Koskinen, M. K., and Riga, D. (2020). Incubation of depression: ECM assembly and parvalbumin interneurons after stress. Neurosci. Biobehav. Rev. 118, 65–79. doi: 10.1016/j.neubiorev.2020.07.015
Stevens, S. R., Longley, C. M., Ogawa, Y., Teliska, L. H., Arumanayagam, A. S., Nair, S., et al. (2021). Ankyrin-R regulates fast-spiking interneuron excitability through perineuronal nets and Kv3.1b K+ channels. Elife 10:e66491. doi: 10.7554/eLife.66491
Strackeljan, L., Baczynska, E., Cangalaya, C., Baidoe-Ansah, D., Wlodarczyk, J., Kaushik, R., et al. (2021). Microglia depletion-induced remodeling of extracellular matrix and excitatory synapses in the hippocampus of adult mice. Cells 10:1862. doi: 10.3390/cells10081862
Su, W., Foster, S. C., Xing, R., Feistel, K., Olsen, R. H., Acevedo, S. F., et al. (2017). CD44 transmembrane receptor and hyaluronan regulate adult hippocampal neural stem cell quiescence and differentiation. J. Biol. Chem. 292, 4434–4445. doi: 10.1074/jbc.M116.774109
Tanti, A., Belliveau, C., Nagy, C., Maitra, M., Denux, F., Perlman, L., et al. (2020). Child abuse associates with increased recruitment of perineuronal nets in the ventromedial prefrontal cortex: a possible implication of oligodendrocyte progenitor cells. bioRxiv [Preprint]. doi: 10.1101/2020.10.19.345355
Tomar, A., Polygalov, D., Chattarji, S., and McHugh, T. J. (2021). Stress enhances hippocampal neuronal synchrony and alters ripple-spike interaction. Neurobiol. Stress 14:100327. doi: 10.1016/j.ynstr.2021.100327
Ueno, H., Suemitsu, S., Murakami, S., Kitamura, N., Wani, K., Matsumoto, Y., et al. (2018). Juvenile stress induces behavioral change and affects perineuronal net formation in juvenile mice. BMC Neurosci. 1:41. doi: 10.1186/s12868-018-0442-z
Ueno, H., Suemitsu, S., Murakami, S., Kitamura, N., Wani, K., Okamoto, M., et al. (2017). Region-specific impairments in parvalbumin interneurons in social isolation-reared mice. Neuroscience 359, 196–208. doi: 10.1016/j.neuroscience.2017.07.016
Uriarte, N., Ferreño, M., Méndez, D., and Nogueira, J. (2020). Reorganization of perineuronal nets in the medial preoptic area during the reproductive cycle in female rats. Sci. Rep. 10:5479. doi: 10.1038/s41598-020-62163-z
Vaswani, A. R., and Blaess, S. (2016). Reelin signaling in the migration of ventral brain stem and spinal cord neurons. Front. Cell. Neurosci. 10:62. doi: 10.3389/fncel.2016.00062
Ventorp, F., Barzilay, R., Erhardt, S., Samuelsson, M., Träskman-Bendz, L., Janelidze, S., et al. (2016). The CD44 ligand hyaluronic acid is elevated in the cerebrospinal fluid of suicide attempters and is associated with increased blood-brain barrier permeability. J. Affect. Disord. 193, 349–354. doi: 10.1016/j.jad.2015.12.069
Venturino, A., Schulz, R., Jesús-Cortés, H. D., Maes, M. E., Nagy, B., Reilly-Andújar, F., et al. (2021). Microglia enable mature perineuronal nets disassembly upon anesthetic ketamine exposure or 60-Hz light entrainment in the healthy brain. Cell Rep. 36:109313. doi: 10.1016/j.celrep.2021.109313
Vincent, C., Gilabert-Juan, J., Gibel-Russo, R., Alvarez-Fischer, D., Krebs, M. O., Le Pen, G., et al. (2021). Non-cell-autonomous OTX2 transcription factor regulates anxiety-related behavior in the mouse. Mol. Psychiatry doi: 10.1038/s41380-021-01132-y
Wang, R. H., Chen, Y. F., Chen, S., Hao, B., Xue, L., Wang, X. G., et al. (2018). Maternal deprivation enhances contextual fear memory via epigenetically programming second-hit stress-induced reelin expression in adult rats. Int. J. Neuropsychopharmacol. 21, 1037–1048. doi: 10.1093/ijnp/pyy078
Wingert, J. C., and Sorg, B. A. (2021). Impact of perineuronal nets on electrophysiology of parvalbumin interneurons, principal neurons, and brain oscillations: a review. Front. Synaptic Neurosci. 13:673210. doi: 10.3389/fnsyn.2021.673210
Xue, Y. X., Xue, L. F., Liu, J. F., He, J., Deng, J. H., Sun, S. C., et al. (2014). Depletion of perineuronal nets in the amygdala to enhance the erasure of drug memories. J. Neurosci. 34, 6647–6658. doi: 10.1523/JNEUROSCI.5390-13.2014
Yamada, J., and Jinno, S. (2017). Molecular heterogeneity of aggrecan-based perineuronal nets around five subclasses of parvalbumin-expressing neurons in the mouse hippocampus. J. Comp. Neurol. 525, 1234–1249. doi: 10.1002/cne.24132
Yang, S., Hilton, S., Alves, J. N., Saksida, L. M., Bussey, T., Matthews, R. T., et al. (2017). Antibody recognizing 4-sulfated chondroitin sulfate proteoglycans restores memory in tauopathy-induced neurodegeneration. Neurobiol. Aging 59, 197–209. doi: 10.1016/j.neurobiolaging.2017.08.002
Yu, Z., Chen, N., Hu, D., Chen, W., Yuan, Y., Meng, S., et al. (2020). Decreased density of perineuronal net in prelimbic cortex is linked to depressive-like behavior in young-aged rats. Front. Mol. Neurosci. 13:4. doi: 10.3389/fnmol.2020.00004
Zhang, J., Lin, L., Dai, X., Xiao, N., Ye, Q., and Chen, X. (2021). ApoE4 increases susceptibility to stress-induced age-dependent depression-like behavior and cognitive impairment. J. Psychiatr. Res. 143, 292–301. doi: 10.1016/j.jpsychires.2021.09.029
Zhang, J., Qin, L., and Zhao, H. (2013). Early repeated maternal separation induces alterations of hippocampus reelin expression in rats. J. Biosci. 38, 27–33. doi: 10.1007/s12038-012-9286-3
Zielinski, M. C., Tang, W., and Jadhav, S. P. (2020). The role of replay and theta sequences in mediating hippocampal-prefrontal interactions for memory and cognition. Hippocampus 30, 60–72. doi: 10.1002/hipo.22821
Keywords: perineuronal net, extracellular matrix, stress, hippocampus, prefrontal cortex, amygdala
Citation: Laham BJ and Gould E (2022) How Stress Influences the Dynamic Plasticity of the Brain’s Extracellular Matrix. Front. Cell. Neurosci. 15:814287. doi: 10.3389/fncel.2021.814287
Received: 13 November 2021; Accepted: 27 December 2021;
Published: 25 January 2022.
Edited by:
Chandrakanth Reddy Edamakanti, Northwestern Medicine, United StatesReviewed by:
Hiroshi Kitagawa, Kobe Pharmaceutical University, JapanBarbara Gisabella, University of Mississippi Medical Center, United States
Marta Miquel, University of Jaume I, Spain
Copyright © 2022 Laham and Gould. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elizabeth Gould, Z291bGRlQHByaW5jZXRvbi5lZHU=