 Yu-hua Gong
Yu-hua Gong Shi-lei Hao
Shi-lei Hao Bo-chu Wang
Bo-chu Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Neurosci. , 24 March 2021
Sec. Cellular Neurophysiology
Volume 15 - 2021 | https://doi.org/10.3389/fncel.2021.653367
This article is part of the Research Topic Pluripotent Cells for Stroke: from Mechanism to Therapeutic Strategies View all 13 articles
Intracerebral hemorrhage (ICH) is one of the leading causes of death and long-term disability worldwide. Mesenchymal stem cell (MSC) therapies have demonstrated improved outcomes for treating ICH-induced neuronal defects, and the neural network reconstruction and neurological function recovery were enhanced in rodent ICH models through the mechanisms of neurogenesis, angiogenesis, anti-inflammation, and anti-apoptosis. However, many key issues associated with the survival, differentiation, and safety of grafted MSCs after ICH remain to be resolved, which hinder the clinical translation of MSC therapy. Herein, we reviewed an overview of the research status of MSC transplantation after ICH in different species including rodents, swine, monkey, and human, and the challenges for MSC-mediated ICH recovery from pathological microenvironment have been summarized. Furthermore, some efficient strategies for the outcome improvement of MSC transplantation were proposed.
Intracerebral hemorrhage (ICH) is one of the most devastating and life-threatening neurological diseases, which has led to high disability and mortality worldwide. Although ICH only accounts for ∼15% of all strokes, its mortality rate within 28 days is as high as ∼47%, and the surviving patients up to 25% are at risk of relapse ICH in the next 5 years (Chen Y. et al., 2020). Worse still, less than 12% of patients would have independent living ability at 1 year after ICH (van Asch et al., 2010). The incidence rate of ICH has not decreased with the development of medical and healthcare level, but shows an increasing trend in low- and middle-income countries (Wu and Anderson, 2020). This is probably due to the increased number of elderly people and the use of antiplatelet, anticoagulants, and thrombolytics (Carpenter et al., 2016; Wu et al., 2019). As of yet, considerable progress for ICH treatment has been made in preclinical research, however, there is still a lack of effective therapeutic strategies for the acute and long-term treatment of ICH in clinic, except for active rehabilitation.
Mesenchymal stem cell (MSC) therapy holds significant promise in regenerative medicine research and tissue engineering due to its unique properties, including extensive proliferation capacity, multiple differentiation potential, ease of isolation from various tissues, low immunogenicity, paracrine activity, immunomodulatory function, and fewer ethical disputes (Zheng et al., 2018). The application of MSC therapies has represented an exciting treatment for stroke, especially for ICH patients suffering from neurological deficits and motor dysfunction. To date, ICH researchers have conducted much pre-clinical and clinical research based on MSC treatment with promising results (Bedini et al., 2018; Gao et al., 2018).
However, the prognostic differences between individuals, the influence of pathological environment on the characteristics of implanted MSCs, and the long-term safety based on MSC therapies are undefined, which is still a non-negligible obstacle to clinical translation. When emphasizing that MSC replacement therapy is a very promising treatment option for ICH, the interaction between the pathological microenvironment of the individual and the implanted MSCs should also be strictly considered. Therefore, this review summarized the application of MSCs in the treatment of ICH in different species (e.g., rodent, swine, monkey, and human) and their mechanism of promoting neurological recovery. Meanwhile, we emphasized the existing challenges faced by MSCs planted into the microenvironment of ICH injured brain tissues, including the impacts of mass effect, iron overload, and oxidative stress. Furthermore, we further discussed the MSC-based potential therapeutic methods that might have an optimized clinical transformation effect in the treatment of ICH.
The cascade brain injury initiated by ICH is conventionally described as two consecutive pathological processes. In the first stage of primary brain injury, the resulting hematoma from the ruptured vessel could form a persistent mass effect, and mechanically stretch and compress the brain tissue to cause brain tissue damage. Subsequently, the secondary brain injury was mainly caused by the hematoma and its degradation products (i.e., hemoglobin, heme, and iron), which could induce microglia activation, inflammatory response, oxidative stress, brain edema, and blood-brain barrier (BBB) leakage. The secondary brain injury will further lead to permanent death of brain tissue and severe neurological deficits (Qureshi et al., 2009; Wilkinson et al., 2018).
Mesenchymal stem cells represent a versatile class of multipotent stem cells that hold promise to directly regenerate damaged tissues due to their potential of differentiation into almost any end-stage lineage cells (Han et al., 2019). Bone marrow-derived MSCs were the firstly-discovered and extensively studied MSCs, although they also had been isolated and characterized from tissues including adipose, dermis, cord blood, peripheral blood, synovial fluid, umbilical cord, placenta, amniotic fluid, fetal tissues, dental pulp, periosteum, and skeletal muscle (Murphy et al., 2013; Bedini et al., 2018). Various minimally invasive procedures had been used to isolate and expand MSCs in vitro, including abdominoplasty, bone marrow aspiration, and placental collection (Sherman et al., 2019). Therefore, MSCs could be obtained in quantity from the patients or third-party donors. These characteristics of easy availability and no ethical concerns make them widely used in regenerative medicine research (Wang et al., 2018). It should be noted that although these cells have many similarities, MSCs still exist with different gene expression patterns, differentiation profile, and clinical application potential according to their origin (Ghanta et al., 2018).
Although the exact mechanism behind MSC-based therapies was still largely ambiguous, numerous studies had been conducted in animals and humans, and some therapeutic effects had been achieved with various therapy mechanisms. However, differences about the focus point and outcomes of MSC treatment in ICH still exist among species. Detailed research findings of MSC-based ICH intervention in different species will be introduced in subsequent sections, and a summary of the relevant studies is reported in Table 1.
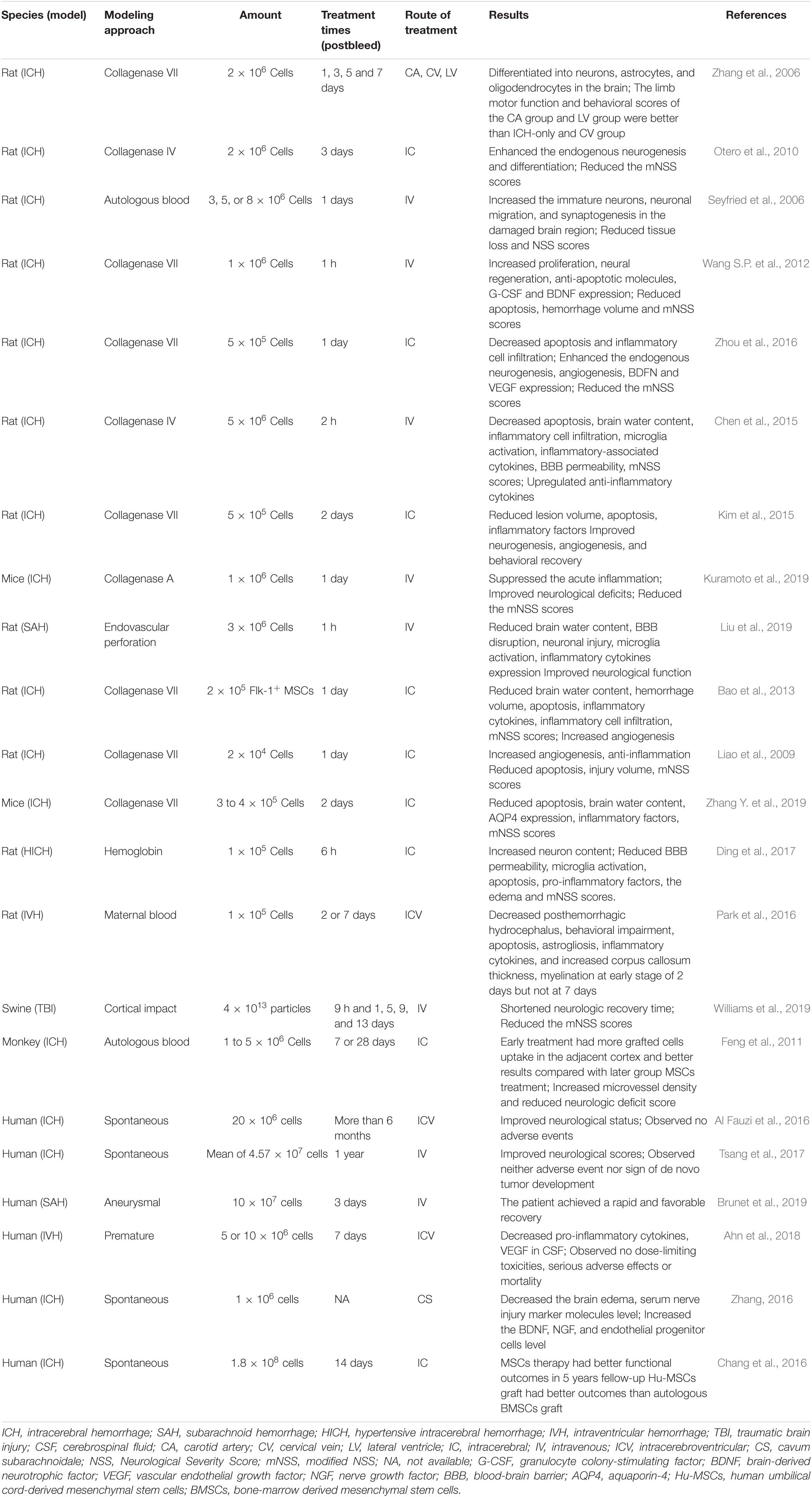
Table 1. The application of MSC-based therapy in different species of hemorrhage stroke.
At present, plenty of experimental research has been conducted on the treatment of rodent ICH with MSC transplantation, probably due to the versatility of rodent models. Large amounts of evidence confirmed that MSC transplantation is a promising treatment for neuronal recovery after ICH (Ma et al., 2015; Hu et al., 2016; Zheng et al., 2018). In view of the neurological function improvement effect of MSCs in ischemic rat brain, it was no doubt that the first MSC application in the rat ICH model also showed significant improvement in motor function (Zhang et al., 2006). To investigate the role of MSCs in ICH therapy, BrdU-labeled MSCs were delivered into the brain through carotid artery (CA), cervical vein (CV), or lateral ventricle (LV), respectively, at days 1, 3, 5, and 7 following ICH. Thereafter, BrdU-labeled MSCs were found in the brain of CA and LV delivery groups except CV injection. The majority of labeled MSCs could migrate into the ipsilateral cortex, bleeding area, and hippocampus. Additionally, double staining revealed that MSCs mainly differentiated into neurons in the hippocampus, while MSCs differentiated into neurons and astrocytes around the bleeding area. In the ipsilateral cortex, MSCs could differentiate into neurons, astrocytes, and oligodendrocytes. Taken together, these results demonstrated that the functional improvement of MSCs in ICH was probably related to their ability to differentiate into appropriate cell types. Furthermore, the nestin and doublecortin positive cells had been observed in the subventricular zone (SVZ) and nearby the lesion zone in the ICH animals received MSCs, indicating that the MSC transplantation could also enhance endogenous neurogenesis and differentiation (Otero et al., 2010). Therefore, MSC therapy could mediate the transplanted and endogenous neurogenesis to reorganize the lost neuronal circuity after ICH. The observation of immature neurons and decreased tissue loss of striatum further confirmed the organizational restructuring role of MSCs in ICH (Seyfried et al., 2006, 2008).
The molecular mechanisms underlying MSC-mediated neurogenesis behavior was partly attributed to the trophic properties of MSCs, which could secrete neurotrophic and other chemokines to promote transplanted or endogenous stem cells proliferation and differentiation (Murphy et al., 2013; Baker et al., 2019). Previous studies demonstrated that tail vein delivered bone marrow-derived MSCs had increased the expression of brain-derived neurotrophic factor (BDNF), which was a crucial growth factor for the growth and differentiation of central nervous systems (Wang S.P. et al., 2012). Knockout of BDNF in MSCs had significant suppression in the effective treatment results of transplanted cells in intraventricular hemorrhage (IVH) (Ahn et al., 2017). Vascular endothelial growth factor (VEGF), which was closely related to angiogenesis, was also significantly upregulated in ICH brain tissues with transplanted MSCs (Zhou et al., 2016). In addition, glial cell-derived neurotrophic factor (GDNF) was also recognized as an important neurotrophic factor in the growth, differentiation, development, maintenance, and injury repair for several types of neurons. Not only transfection but also drug-induced increase of GDNF could significantly enhance the functional improvement of grafted MSCs on ICH (Yang et al., 2011; Lee et al., 2015; Deng et al., 2019). Furthermore, delivered hypoxia-preconditioned MSCs could reduce the tissue loss of the ipsilateral striatum, and enhance neuroregeneration and neurological functional recovery after ICH, which was attributed to the neuronal nourish and protect effect of MSC-mediated upregulation of GDNF, VEGF, and BDNF (Sun et al., 2015).
Neuronal cells insult caused by the primary brain injury and secondary brain injury after ICH was mainly located around the hematoma. Therefore, whether endogenous stem cells or exogenous stem cells could migrate to the perihematoma was a key factor for neuronal remodeling. To promote cells viability and migration, platelet-rich plasma (PRP)-derived scaffold was employed to deliver MSCs, and the contained fibronectin, fibrin, and adhesive proteins in the scaffold were expected to augment cell interaction and promote cell migration. Apparently, (PRP)-derived scaffold increased the biologic activity of donor cells and its integration in the injured tissue (Vaquero et al., 2013).
Although the potential neural recovery mechanism of MSCs in ICH is still largely undefined, apparently, it is not limited to the differentiation and remodeling of brain tissue. In fact, mechanisms involved in tissue repair, such as neurogenesis, angiogenesis, anti-inflammation, and anti-apoptosis, were all suggested to be associated with the functional recovery effect of transplanted MSCs in ICH (Chen et al., 2015; Kim et al., 2015; Xie et al., 2016; Kuramoto et al., 2019; Liu et al., 2019). It was well characterized that ICH-induced inflammation response was a key factor in secondary brain injury, which involved microglia activation, and infiltrating neutrophils release could induce BBB breakdown, vasogenic edema, and apoptosis of glia and neurons (Keep et al., 2012). Thus, a previous study had proposed a hypothesis that the behavioral recovery improved by MSC therapy might derive from the inhibition of inflammation cascade in ICH. In order to verify this hypothesis, Flk-1+ MSCs was introduced for ICH (Bao et al., 2013). Results showed that Flk-1+ MSCs-treatment dramatically decreased the neurons apoptosis, brain water content, inflammatory cells expression (e.g., microglia and neutrophils), and the mRNA levels of inflammatory mediators (e.g., IL-1β, IL-2, IL-4, IL-6, and TNF-α). Moreover, umbilical cord-derived MSCs administration had statistically increased vessel density, and reduced microglial activation and leukocytes infiltration at 3 days after ICH compared to the control group. These suggested that, except for neuronal regeneration, the underlying mechanisms of MSCs to accelerate neurological function recovery after ICH were also attributed to its effects of promoting angiogenesis and inhibiting inflammation (Liao et al., 2009). Other therapeutic mechanisms revealed that MSC transplantation-mediated inflammation suppression and down-regulation of aquaporin-4 (AQP4) protein expression could alleviate brain edema of ICH (Zhang Y. et al., 2019). The mechanism of transplanted MSCs inhibiting inflammation might be partly involved in the inhibition of iNOS expression (Ding et al., 2017). In addition, the microglial M2 polarization mediated by the secretory factors, insulin-like growth factor-1 (IGF-1) of stem cells, was considered to be a possible mechanism for stem cells improved hemorrhage stroke prognosis (Chen et al., 2019; Sun et al., 2020). Taken together, these studies demonstrated that anti-inflammation is a crucial mechanism involved in MSCs-mediated functional and tissue recovery after ICH.
Moreover, Adipose-derived stem cells administration rats of ICH found significantly decreased brain water content, brain atrophy, and apoptosis, indicating the multiple mechanisms of MSCs therapy in ICH recovery (Kim et al., 2007). Furthermore, other studies had demonstrated that MSCs transplantation improved BBB integrity after ICH, which was associated with the increased expression of TNF-α stimulated gene/protein 6 (TSG-6) and tight junction proteins (claudin-5 and zonula occludens-1) (Chen et al., 2015; Choi et al., 2018). As hypertension is the most common cause of cerebral hemorrhage clinically, a spontaneously hypertensive model was created to evaluate the long-term neuroprotective effects of MSCs in ICH, which would have great value for pre-clinical study. This study demonstrated that MSC treatment could reverse the BBB permeability and improve the neurological function after hypertensive ICH (Wang et al., 2015). For stem cell therapy, the conventional mode of administration is injection of cells into the brain directly or through blood vessels. However, the method of cells being injected into the brain is invasive, and regarding the method of vascular administration, it has been demonstrated that there were only a few cells homing to the brain or had embolization and possible tumorigenesis risk (Li P. et al., 2019; Mello et al., 2020). Thus, a novel way of intranasal route was applied for MSC transplantation therapy in collagenase IV injected mice (Sun et al., 2015). In addition, MSC-derived exosomes were also considered as a promising alternative strategy to solve the obstacle in MSC application (Bedini et al., 2018).
Although different from the etiology of spontaneous ICH, the rehabilitation research of traumatic cerebral hemorrhage also had an important reference value for the clinical transformation of ICH intervention, especially in large animals. Exosomes were membrane-enclosed nanovesicles that contained numerous molecular constituents including cellular proteins, lipids, microRNAs and mRNAs, and had the properties of low immunogenicity, BBB penetrability, intercellular communication and neural regeneration mediation (Yang et al., 2017). Meanwhile, due to the neuroprotective effect of MSCs-generated exosomes in small animal traumatic brain injury (TBI) model, they were also highly expected in the treatment of traumatic cerebral hemorrhage in large animals (Zhang Y. et al., 2015). When Yorkshire swine were treated with MSCs-derived exosomes, they appeared to have faster neurologic recovery and obvious reduction of neurologic injury (Williams et al., 2019). The benefit outcomes in large animal models would provide a reliable evidence for the clinical application of preclinical research.
The first study of MSC treatment for primates was conducted in an ICH model of intracranial autologous injected macaca fascicularis monkey (Feng et al., 2011). The MSC-treated group showed significantly improved neurologic deficit and microvessel density compared with the control group. Additionally, the serial 18F-FDG PET scans found that the 18F-FDG uptake in the adjacent cortex of the early treatment was increased significantly when compared with control group during recovery phase, and better treatment results were obtained in the MSC early treatment group (7 days) compared with late treatment group (28 days). However, the optimal timing of MSC transplantation for severe IVH in newborn rat pups was considered to be at early stage of 2 days instead of late at 7 days after induction (Park et al., 2016). Therefore, when extrapolating preclinical data into clinical trials, the narrow therapeutic time window of MSC transplantation for ICH intervention should be determined. Meanwhile, it is also worth noting that stereotaxical delivery of MSCs to the outside of the right putamen of macaca fascicularis monkey could induce the local tissue inflammation and necrosis, although this damage might repair over time (Feng et al., 2014). Nevertheless, a total of four intravenous injections of MSCs (1 × 107 cells/kg) with 2 weeks interval did not affect the general health of monkeys (Wang Y. et al., 2012). These studies suggested that the delivery modes and therapeutic time window of MSCs applied for ICH were key factors associated to prognosis and required further elucidation.
Mesenchymal stem cells have been extensively studied and confirmed as an effective treatment strategy for animal cerebral hemorrhage models, however, less research has been investigated in human cerebral hemorrhage. The first application of MSCs in humans was a case report about two persistent vegetative state patients of hemorrhagic stroke, and then they were followed-up with for a 1 year period (Al Fauzi et al., 2016). Patients were intraventricularly injected with MSCs (20 × 106 cells/2.5 mL) three times at intervals of 1 month using an Ommaya reservoir. The National Institute of Health Stroke Scale (NIHSS) scores of the two patients indicated that the functional status was obviously improved. A placebo-controlled study also found that two deliveries of MSCs (2 × 106 cells/3 mL) 1 month apart could significantly improve the neuro-restoration and clinical prognosis of ICH patients with severe disability (Tsang et al., 2017). In another case report of an 80-year-old SAH patient with a past medical history (i.e., diabetes, hypertension, and cardiac bypass) found that MSC transplantation gradually improved the consciousness of the patient at 3 weeks after SAH, and restored the ability of speaking and self-care (Brunet et al., 2019). Consistently, this procedure might provide a safe, feasible and effective treatment for patients with clinical cerebral hemorrhage in the future, and enhance the confidence in the treatment of patients with cerebral hemorrhage.
Based on previous experimental and clinical benefit, MSC transplantation was also carried out on premature infants to explore its safety and efficacy for severe IVH (Ahn et al., 2018). A total of nine premature infants were enrolled in this clinical trial, and different doses of MSCs were injected into the lateral ventricle (5 × 106 cells/kg; 1 × 107 cells/kg) on average at twelve postnatal days. All patients could tolerate the procedure well, and there were no immediate complications (i.e., allergies or death) which occurred within 6 h after transplantation. In these infant patients, four showed continual regression and improvement, and five received shunt placement due to the progressive ventriculomegaly before discharge. In most MSC transplanted patients, the inflammatory factors such as IL-6, TGF-β1/2, TNF-α, and IL-1β all exhibited a decreasing trend compared to before transplantation, that was consistent with that in animal experiments (Bao et al., 2013). Different from preclinical research, VEGF showed a reduced expression after MSC therapy, which was probably related to the up-regulation of baseline VEGF expression caused by hemorrhage insult (Tang et al., 2007). Meanwhile, Safety evaluation results showed both doses of MSCs did not induce the serious adverse events and dose-limiting toxicity, and the mortality was zero even in grade 4 IVH, which implied this strategy was safe when the dose was controlled within 1 × 107 cells/kg of MSCs. As severe IVH and subsequent posthemorrhagic hydrocephalus could cause long-term neurological disorders in surviving preterm infants, children with neurological sequelae underwent a noteworthy clinical trial in which they were treated with bone marrow-derived mononuclear cells (BMMNCs) that might have the synergic effect of hematopoietic stem cells (HSCs) and MSCs (Liem et al., 2019). Except for no observed any adverse side effect, BMMNC transplantation had made great progress in many aspects of neurodevelopment, such as motor function, fine motor skill, personal social, motor adaptive, and language skills, demonstrating the BMMNCs treatment effectiveness in promoting the social interaction and self-care ability of these children. This improvement in human-specific physiological functions was difficult to evaluate in preclinical animal studies, and the functional ameliorate illustrated that the transplanted stem cells had availably differentiated into region specific cell types and integrated into neural network appropriately.
Because of the mechanical damage of hematoma to brain tissue, clot evacuation was a commonly used intervention in the clinical treatment of ICH. Therefore, the combined therapy of MSC transplantation followed by minimally invasive hematoma removal was supposed as a promising treatment for ICH. Compared with surgical treatment, the combined group significantly decreased the brain edema, and increased the BDNF, NGF, and Endothelial progenitor cells level (Zhang, 2016). In a case report of a female patient with a past medical history of hypertension, the combined treatment also observed improved neurologic function consistent with preclinical studies (Zhang Q. et al., 2015; Zahra et al., 2020). However, there were discrepancies among combined therapy with respect to prognosis in different sources of transplanted MSCs, which might be due to differences in the viability, proliferation potential, and neurogenic efficacy of the tested cell line. A retrospective analysis of up to 5-years follow up had documented that the patients who received human umbilical cord-derived MSCs in the combined therapy group showed better outcome on functional recovery and complications than the bone-marrow MSCs combined therapy group and control group (Chang et al., 2016). Overall, the MSCs transplantation had promise to be safely and effectively applied in clinical ICH treatment, even so, the underlying regulatory mechanism, the optimal implantation time, and larger clinical trials still need to be further considered in future research.
Mesenchymal stem cell transplantation had demonstrated the positive therapeutic effects of facilitating tissue repair and prognosis of ICH in animal research and clinical trials. However, significant research differences still existed in the process of transforming preclinical research to clinical application. The uncertainty contained in the differences of this research might affect the therapeutic mechanisms of transplanted MSCs and the stability of clinical benefits from many aspects. Whether the therapeutic mechanisms of MSC therapy reported in animal studies could effectively translate to the complex pathological environment of ICH patients was also unknown.
Actually, the differences between experimental research and clinical trials had already appeared in the screening of research subjects. Healthy rodents were the most versatile research group in preclinical studies. However, patients with ICH usually had a variety of other diseases, because ICH often occurred in the antiplatelet, anticoagulants, and thrombolytics users or elder peoples (Brunet et al., 2019). In addition, the reported ICH patients also included a strict inclusion and exclusion criteria, which did not exist in the healthy animal research subjects (Ahn et al., 2018). Moreover, the small sample size and case report in clinical studies further increased the publication bias. Most importantly, none of the current animal ICH models could reproduce the actual pathological process of ICH. Taken together, in the process of extrapolating the trial results to clinical application, it was necessary to be further verified from the research objects, model establishment, sample size, and other aspects.
It is well known that the cell source, extraction, and expansion process of MSCs are direct factors associated with the biological characteristics and differentiation fate of implanted cells. In the rodent studies, MSCs derived from different sources (bone marrow, adipose, umbilical cord, placenta, dental pulp, and cord blood) had been used for ICH treatment and displayed an improved prognosis (Choi et al., 2018; Chen et al., 2019; Liu et al., 2019; Zhang Y. et al., 2019; Chen K.H. et al., 2020). Nevertheless, this trend of cell source balance had been tilted in human research. Except for a small amount of MSCs derived from umbilical cord or cord blood (Chang et al., 2016; Ahn et al., 2018), most of the transplanted MSCs were extracted through autologous bone marrow puncture in the clinical studies (Liem et al., 2019; Zahra et al., 2020). Different from this, the bone marrow-MSCs in the rodent studies were mainly derived from allogeneic femur and tibia. In addition, the MSCs transplantation in patients were all homogeneous, and MSCs implanted in rodents were also derived from heterogeneous (Chen K.H. et al., 2020). However, no research had focused on the influence of MSCs source or extraction differences on ICH treatment. Considering the risk and time consumption of autologous MSCs extraction, the immunogenicity of allogeneic MSCs, and the high cost of maintaining cells viability, optimized and practical MSCs product preparation still needed to be clarified in research to ensure the stability and predictability during transformation for clinical treatment.
Unlike animal research, registration was introduced to comprehensively and objectively record all clinical trials which had been conducted. However, many clinical studies did not report the registration information, and only a few studies had documented the registration number of ClinicalTrials.gov (Ahn et al., 2018; Zahra et al., 2020). Although the improved prognosis of MSC treatment had been widely reported, the results of animal studies were mainly histopathological examination and neurological evaluation (Liu et al., 2019), and the clinical studies were more about blood composition testing and neurological function scores (Zhang, 2016; Brunet et al., 2019). In fact, clinical results about MSCs treated ICH were more of a case report for the patients (Liem et al., 2019). The lack of control studies not only made the results occasional, but also greatly compromised the quality of clinical trials. Therefore, any conclusive reports about clinical therapeutic mechanisms or effects might be too early and not rigorous. The clinical application of MSCs for ICH treatment still needed more controlled and statistically significant clinical trials.
The extracellular environment plays an important role in the proliferation, differentiation, and clinical prognosis of transplanted MSCs. In fact, the biomimetic microenvironment not only helps to maintain the advantages of MSCs (e.g., proliferation, differentiation, and immunoregulatory properties), but also helps to retain their phenotype, metabolism, adhesion, and response signal to surrounding factors (Kim H. et al., 2019). The central nervous system refers to a complicated system containing neurons, glial cells, blood vessels, and extracellular matrix, which closely interacts with the transplanted MSCs and determines its fate (Dooves et al., 2016). However, the composition of the brain tissue microenvironment with hemorrhage infiltration becomes more complex and changeable, including mass effect, erythrocyte lysate, thrombin, free radicals, cytotoxic, excitotoxic, inflammatory effects, and nitric oxide (Babu et al., 2012; Gao et al., 2018). Meanwhile, many different stimulations had been recognized as the initiators of the stress-induced premature senescence of MSCs, including mechanical stress, osmotic stress, ionizing radiation, reactive oxygen species, and hypoxia (Zhou et al., 2020). Therefore, the extracellular microenvironment of perihematoma should be strictly evaluated when exploring effective and translatable MSCs therapies for hemorrhage stroke (Figure 1).
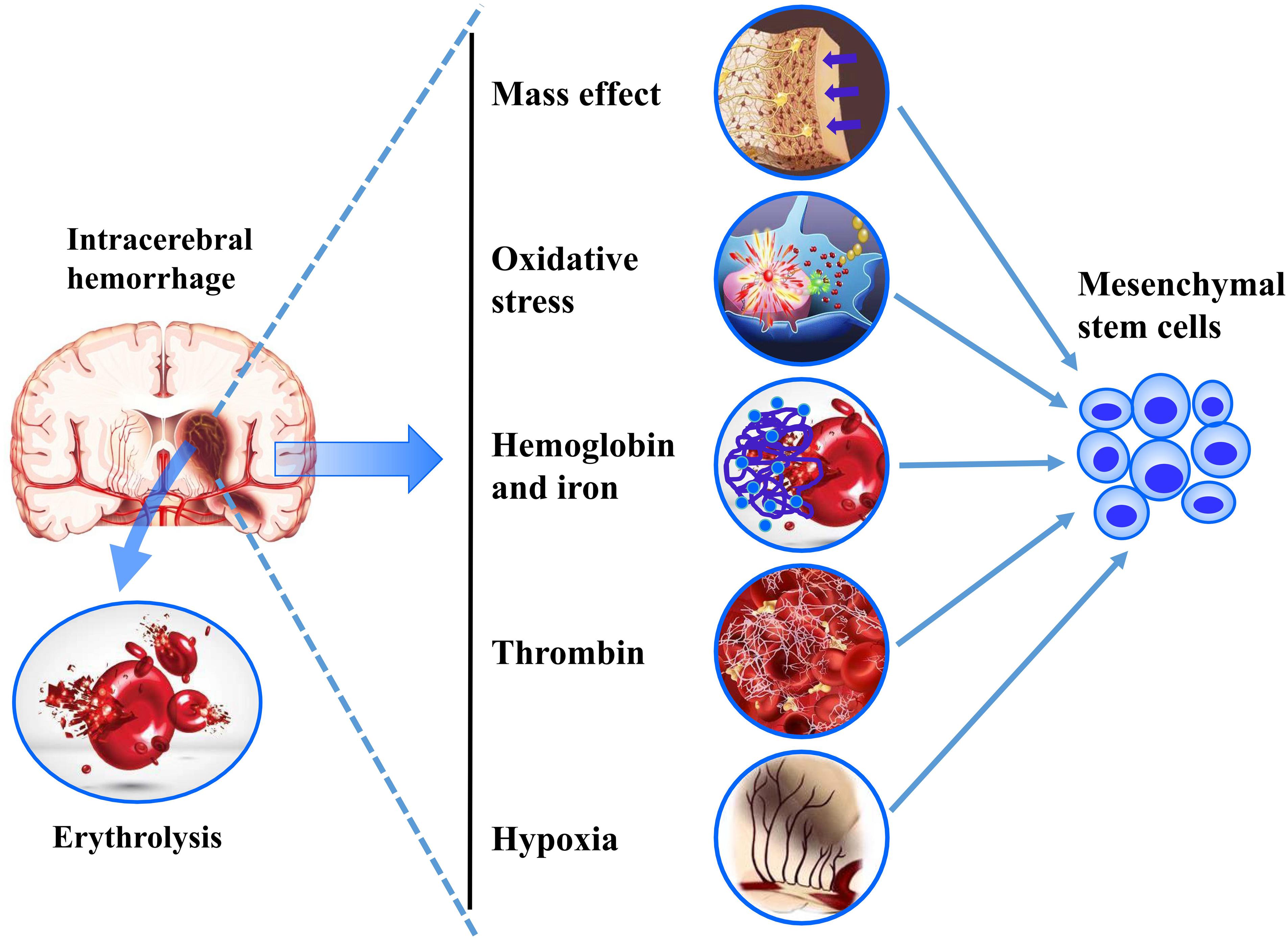
Figure 1. A summarization of the challenges of ICH microenvironments on MSC transplantation.
Mass effect is the main primary brain injury mechanism in ICH, which refers to the mechanical stretching and compression of the surrounding brain tissue by the hematoma (Wilkinson et al., 2018). Moreover, in the limited space of the skull, hematoma can lead to increased intracranial pressure and edema-derived mass effect (Guo et al., 2017). These would all result in a dynamic change and increase of mechanical stress in the intracranial microenvironment. So, the transplanted MSCs would sit in a microenvironment of mechanobiology with multiple complex mechanisms. Our studies had suggested that mechanical pressure enhanced the stretch-activated ion channel Piezo-2 expression in vitro and vivo as early as 8 h after ICH (Guo et al., 2018; Gong et al., 2019). In vitro data from adult Drosophila midgut feeding with indigestible food to generate mechanical loaded showed an increase enteroendocrine cells generation through Piezo protein upregulation, these suggested that the existence of stem cells in the fly midgut could, through mechanical signals, sense to modulate proliferation and differentiation (He et al., 2018). Mechanobiology, as the inductive niche of stem cells, could regulate their developmental processes and self-renewal (Vining and Mooney, 2017).
Thrombin, derived from blood after ICH, is an essential component of the coagulation cascade, which has been observed to induce the brain injury through activating the protease-activated receptors (PAR) (Gao et al., 2014; Mao et al., 2016). Although thrombin is a proven tissue damage mechanism in ICH, thrombin-activated platelet-rich plasma was reported to provide a higher proliferation rate and MSC marker expression in long-term cultured MSCs (Kocaoemer et al., 2007). Thrombin-preconditioned MSCs displayed a five-fold acceleration of MSC-derived extracellular vesicles biogenesis and a two-fold enrichment of their cargo contents, these were also regulated via PAR-mediated intracellular signaling pathways (i.e., ERK1/2, AKT, Rab5, and EEA-1) (Sung et al., 2019). Meanwhile, thrombin also could promote fibronectin secretion of MSCs via PAR-mediated ERK1/2 activation (Chen et al., 2014), and the fibronectin-formed adsorption force could regulate the transmission of the cell traction force and the lineage specifications of MSCs (Lin et al., 2018). Additionally, in the newborn rat model of unilateral carotid artery ligation, thrombin preconditioned human Whartong’s jelly-derived MSC transplantation significantly enhanced the anti-inflammatory, anti-astroglial, anti-apoptotic effects, and neurological recovery (Kim Y.E. et al., 2019). Apparently, thrombin might have multiple undetermined influence mechanisms on the characteristics of transplanted MSCs during hemorrhage stroke.
Intracerebral hemorrhage, except for the primary damage of mass effect, intraparenchymal blood, and its degradation products (hemoglobin, heme, and iron), could continue to insult the brain tissue through inducing the cytotoxic, excitotoxic, oxidative, and inflammatory effects (Babu et al., 2012). Among these, iron overload was considered as an essential factor to increase the lipid peroxidation, lethal reactive oxygen species (ROS) production and neuronal ferroptosis (Li et al., 2017; Wan et al., 2019). Worse still, the iron overload microenvironment also posed a fatal threat to the therapy of stem cells transplantation. It had been demonstrated that ferric ammonium citrate treatment was capable of markedly reducing the proliferation and pluripotency, and inducing apoptosis and senescence in MSCs (Yang et al., 2016). In addition, iron accumulation was found to elevate the ROS level and apoptosis of MSCs, suggesting that the damage mechanism of iron overload in MSCs transplantation may be related to oxidative stress (Lu et al., 2013; Yuan et al., 2019). Notably, interventions to reduce mitochondrial ROS accumulation could significantly reduce the apoptosis of MSCs (Yang et al., 2016; Yu et al., 2018). Besides, the in vitro and in vivo iron overload were found to have the ability to inhibit the osteogenic commitment and differentiation of MSCs in a dose-dependent manner (Balogh et al., 2016). Therefore, these results indicated that the secondary iron overload microenvironment after ICH would affect not only the apoptosis but also the differentiation of transplanted MSCs.
Although ROS is an inevitable product of the defense system and normal cellular metabolism, the imbalanced homeostasis of the oxidation-reduction system can significantly create disruption to the blood-brain barrier, cell death and structural damage. And that ROS aggregation could be induced by the metabolic products of hematoma, inflammatory cells, and excitatory amino acids after ICH (Qu et al., 2016). Meanwhile, ROS had been extensively considered as an important factor of senescence in MSCs (Ko et al., 2012; Zhou et al., 2020). The oxidative stress formed by sub-lethal doses of hydrogen peroxide could considerably reduce proliferation rate, and accelerate telomere attrition, and induce senescence-associated β-galactosidase expression and senescent morphological features (Brandl et al., 2011). The nuclear factor erythroid-2 related factor 2 (Nrf2) is a critical molecule to protect MSCs against oxidative stresses. It had been reported that the MSCs with transient expression of Nrf2 had the ability to resist the hypoxic and oxidative stress induced cell death and apoptosis (Mohammadzadeh et al., 2012). Oxidative stress preconditioning could protect the MSCs vitality through activating the Nrf2 pathway and upregulating its downstream target of the superoxide dismutase, catalase, and HO-1 (Zhang F. et al., 2019). Moreover, many strategies had been explored to avoid cell apoptosis and senescence initiated by oxidative stress, including pioglitazone, vaspin, ganoderic acid D, and vitamin E (Bhatti et al., 2018; Hu et al., 2019; Zhu et al., 2019; Xu et al., 2020). Taken together, before MSCs could be reliably and effectively used as hemorrhage stroke therapy, it is necessary to better understand the multiple microenvironmental regulatory mechanisms driving transplanted MSCs to form repair effect.
The above description highlights the critical function of ICH pathological microenvironment in determining the MSCs fate. The knowledge regarding mass effect and oxidative stress induced MSCs senescence provides a potential mechanism for the failure of MSCs-mediated clinical benefit in ICH. This indicated that effective therapeutic effect of MSCs on ICH required to strict regulation of the impacts of microenvironment on grafted MSCs. Facing the challenges of applying MSCs in the treatment of ICH, several optimizing strategies had been established and showed potential feasibility.
Studies had shown that preconditioning of MSCs could effectively resist the impact of transplantation microenvironment and increase the regenerative potential of cells, although the underlying mechanism was unknown. Hypoxic preconditioning, pharmacological agents, and trophic factors exposure were commonly used to improve culture expansion conditions, enhance survival and proliferation, and avoid the possible changes of MSCs potency (Hu and Li, 2018; Nahhas and Hess, 2018). In the ICH mice model, the transplantation of hypoxic preconditioned MSCs or neural stem cell significantly increased the grafted-cells survival and the behavioral performance (Sun et al., 2015; Wakai et al., 2016). Additionally, apocynin preconditioned MSCs obviously improved the hematoma expansion, neuronal death, brain edema, and therapeutic efficacy in the acute stage of bacterial collagenase induced rat ICH model compared with native MSCs (Min et al., 2018).
Other than pharmacological and molecular preconditioning of the MSCs, the resistance of MSCs to the microenvironment could also be achieved using miRNA. MiRNA based treatments had the characteristics of delayed MSC senescence and multidimensional targets. Given the significant function of miRNA-126 in promoting angiogenesis, the miRNA-126 transfected MSCs were injected into the collagenase-induced ICH rats, leading to decreased brain water content and improved neurological score (Wang et al., 2020). In addition, the CX3CR1 overexpressed MSCs increased the viability and migration ability of transplanted MSCs, and improved the sensory and motor functions of the collagenase induced mice ICH model (Li G. et al., 2019). Moreover, GDNF-transfected MSCs could improve the neurological function of experimental ICH rats through enhancing neurotrophic factor secretion (Deng et al., 2019). Therefore, miRNA-based MSC therapy was expected to purposefully regulate the proliferation and differentiation ability of MSCs in the pathological microenvironment of ICH.
Creating a standardized and locally beneficial microenvironment is another strategy to maintain the repair potential of transplanted MSCs. Iron overloaded and oxidative stress damage after ICH were recognized as not only critical damage factors for ICH, but also important causes of MSC senescence and apoptosis (Wan et al., 2019; Yuan et al., 2019). To improve the iron overload microenvironment after ICH, an injectable core-shell hydrogel was fabricated for ICH in situ therapy. The outer shell hydrogel with quick degradation property was loaded with iron chelator to eliminate iron overload, and the inner core hydrogel loaded with MSCs and growth factors displayed an improved MSCs survival and differentiation (Gong et al., 2020). In addition, surgical evacuation was a commonly used clinical approach to remove hematoma, and the benefits of combined treatment with MSCs transplantation might associate with the improvement of the transplanted cells microenvironment (Zhang Q. et al., 2015; Chang et al., 2016).
The focus of MSC therapy is cell replacement and neural reorganization, however, its embolization and possible tumorigenesis limitations make exosomes a potential alternative. Significantly, exosomes are even suggested to be the main therapeutic mechanism of MSCs treatment (Han et al., 2018). Exosomes are nano-sized extracellular vesicles, which can be isolated from the supernatant of cultured cells in exosome-free medium through centrifugation and other methods (Bedini et al., 2018). Recently, treatment strategies based on exosomes of MSCs have been widely reported in many diseases. The potential mechanism of exosomes treatment in the ICH mainly comprises the anti-apoptotic, neurogenesis, angiogenesis, and anti-inflammation effects through its miRNAs (Cai et al., 2020; Duan et al., 2020).
The autologous blood ICH rats showed that the apoptotic and degenerative neurons in the miR-133b-modified MSC-derived exosomes treatment group was significantly reduced. It was believed that the neuroprotection effect of exosomes on ICH was mainly reflected in the anti-apoptotic effect of miR-133b through mediating RhoA and ERK1/2/CREB signaling pathway (Shen et al., 2018). The MSC-derived miR-206-knockdown exosomes were also confirmed to improve brain edema and neurological deficit in early brain injury of SAH rats, which was probably related to the suppression of neuronal apoptosis via BDNF/TrkB/CREB signaling (Zhao et al., 2019). Except for anti-apoptotic, the effects of neurogenesis and angiogenesis were also observed in the MSCs-derived exosomes treated ICH rats. The exosomes therapy remarkably increased the newly generated endothelial cells around the hematoma, mature neurons in the SVZ, and myelin in the striatum (Han et al., 2018). Moreover, the MSC-derived exosomes therapy was found to significantly alleviate early brain injury of SAH through restraining the HMGB1-TLR4 pathway activation and generating the anti-apoptosis and anti-inflammation effects (Xiong et al., 2020). Collectively, treatments based on exosomes are expected to provide a broader therapeutic strategy for ICH intervention.
Mesenchymal stem cell-based ICH treatments provide a unique opportunity to improve ICH outcome through multimodal therapeutic action. Multi-species preclinical studies have demonstrated promising evidence that MSCs can promote the neural network reconstruction and neurological function recovery through neurogenesis, angiogenesis, anti-inflammation, and anti-apoptosis mechanisms. Meanwhile, human clinical trials have further confirmed the safety and efficacy of MSCs-based therapy after ICH, and the potential to improve patient prognosis. However, additional studies aimed to better understand the pathological microenvironment related to the successful implantation of MSCs after ICH (e.g., mass effect, thrombin, oxidative stress, and hematoma degradation products) will ultimately augment the therapeutic effect and enhance the reliability and stability of clinical transformation. Furthermore, MSC-derived exosomes treatment might represent a promising way to deal with the complex pathological microenvironment of hemorrhagic stroke.
YG searched the literature and wrote the manuscript. BW and SH critically revised the manuscript. All authors have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This study was supported by the National Natural Science Foundation of China (Grant Nos. 11972099 and 31600770), the Chongqing Research Program of Basic Research and Frontier Technology (Grant Nos. cstc2018jcyjAX0836 and cstc2019jcyj-msxmX0753), and the Ph.D. Foundation of Southwest University of Science and Technology (No. 18zx7128).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Ahn, S. Y., Chang, Y. S., Sung, D. K., Sung, S. I., Ahn, J. Y., and Park, W. S. (2017). Pivotal role of brain-derived neurotrophic factor secreted by mesenchymal stem cells in severe intraventricular hemorrhage in newborn rats. Cell Transplant. 26, 145–156. doi: 10.3727/096368916X692861
Ahn, S. Y., Chang, Y. S., Sung, S. I., and Park, W. S. (2018). Mesenchymal stem cells for severe intraventricular hemorrhage in preterm infants: phase i dose-escalation clinical trial. Stem Cells Transl. Med. 7, 847–856. doi: 10.1002/sctm.17-0219
Al Fauzi, A., Suroto, N. S., Bajamal, A. H., and Mh, M. (2016). Intraventricular transplantation of autologous bone marrow mesenchymal stem cells via Ommaya reservoir in persistent vegetative state patients after haemorrhagic stroke: report of two cases & review of the literature. J. Stem Cells Regen. Med. 12, 100–103.
Babu, R., Bagley, J. H., Di, C., Friedman, A. H., and Adamson, C. (2012). Thrombin and hemin as central factors in the mechanisms of intracerebral hemorrhage-induced secondary brain injury and as potential targets for intervention. Neurosurg. Focus. 32:E8. doi: 10.3171/2012.1.FOCUS11366
Baker, E. W., Kinder, H. A., and West, F. D. (2019). Neural stem cell therapy for stroke: a multimechanistic approach to restoring neurological function. Brain Behav. 9:e01214. doi: 10.1002/brb3.1214
Balogh, E., Tolnai, E., Nagy, B. Jr., Nagy, B., Balla, G., Balla, J., et al. (2016). Iron overload inhibits osteogenic commitment and differentiation of mesenchymal stem cells via the induction of ferritin. Biochim. Biophys. Acta 1862, 1640–1649. doi: 10.1016/j.bbadis.2016.06.003
Bao, X. J., Liu, F. Y., Lu, S., Han, Q., Feng, M., Wei, J. J., et al. (2013). Transplantation of Flk-1+ human bone marrow-derived mesenchymal stem cells promotes behavioral recovery and anti-inflammatory and angiogenesis effects in an intracerebral hemorrhage rat model. Int. J. Mol. Med. 31, 1087–1096. doi: 10.3892/ijmm.2013.1290
Bedini, G., Bersano, A., Zanier, E. R., Pischiutta, F., and Parati, E. A. (2018). Mesenchymal stem cell therapy in intracerebral haemorrhagic stroke. Curr. Med. Chem. 25, 2176–2197. doi: 10.2174/0929867325666180111101410
Bhatti, F. U. R., Kim, S. J., Yi, A. K., Hasty, K. A., and Cho, H. (2018). Cytoprotective role of vitamin E in porcine adipose-tissue-derived mesenchymal stem cells against hydrogen-peroxide-induced oxidative stress. Cell Tissue Res. 374, 111–120. doi: 10.1007/s00441-018-2857-3
Brandl, A., Meyer, M., Bechmann, V., Nerlich, M., and Angele, P. (2011). Oxidative stress induces senescence in human mesenchymal stem cells. Exp. Cell Res. 317, 1541–1547. doi: 10.1016/j.yexcr.2011.02.015
Brunet, M. C., Chen, S. H., Khandelwal, P., Hare, J. M., Starke, R. M., Peterson, E. C., et al. (2019). Intravenous stem cell therapy for high-grade aneurysmal subarachnoid hemorrhage: case report and literature review. World Neurosurg. 128, 573–575. doi: 10.1016/j.wneu.2019.04.055
Cai, Y., Liu, W., Lian, L., Xu, Y., Bai, X., Xu, S., et al. (2020). Stroke treatment: is exosome therapy superior to stem cell therapy? Biochimie. 179, 190–204. doi: 10.1016/j.biochi.2020.09.025
Carpenter, A. M., Singh, I. P., Gandhi, C. D., and Prestigiacomo, C. J. (2016). Genetic risk factors for spontaneous intracerebral haemorrhage. Nat. Rev. Neurol. 12, 40–49. doi: 10.1038/nrneurol.2015.226
Chang, Z., Mao, G., Sun, L., Ao, Q., Gu, Y., and Liu, Y. (2016). Cell therapy for cerebral hemorrhage: five year follow-up report. Exp. Ther. Med. 12, 3535–3540. doi: 10.3892/etm.2016.3811
Chen, J., Ma, Y., Wang, Z., Wang, H., Wang, L., and Xiao, F. (2014). Thrombin promotes fibronectin secretion by bone marrow mesenchymal stem cells via the protease-activated receptor mediated signalling pathways. Stem Cell Res. Ther. 5, 36–47.
Chen, K. H., Shao, P. L., Li, Y. C., Chiang, J. Y., Sung, P. H., Chien, H. W., et al. (2020). Human umbilical cord-derived mesenchymal stem cell therapy effectively protected the brain architecture and neurological function in rat after acute traumatic brain injury. Cell Transplant. 29:963689720929313. doi: 10.1177/0963689720929313
Chen, M., Li, X., Zhang, X., He, X., Lai, L., Liu, Y., et al. (2015). The inhibitory effect of mesenchymal stem cell on blood-brain barrier disruption following intracerebral hemorrhage in rats: contribution of TSG-6. J. Neuroinflammation. 12:61. doi: 10.1186/s12974-015-0284-x
Chen, T.-F., Chen, K.-W., Chien, Y., Lai, Y.-H., Hsieh, S.-T., Ma, H.-Y., et al. (2019). Dental pulp stem cell-derived factors alleviate subarachnoid hemorrhage-induced neuroinflammation and ischemic neurological deficits. Int. J. Mol. Sci. 20:3747. doi: 10.3390/ijms20153747
Chen, Y., Wright, N., Guo, Y., Turnbull, I., Kartsonaki, C., Yang, L., et al. (2020). Mortality and recurrent vascular events after first incident stroke: a 9-year community-based study of 0⋅5 million Chinese adults. Lancet Glob. Health. 8, e580–e590. doi: 10.1016/s2214-109x(20)30069-3
Choi, B. Y., Kim, O. J., Min, S. H., Jeong, J. H., Suh, S. W., and Chung, T. N. (2018). Human placenta-derived mesenchymal stem cells reduce mortality and hematoma size in a rat intracerebral hemorrhage model in an acute phase. Stem Cells Int. 2018:1658195. doi: 10.1155/2018/1658195
Deng, L., Gao, X., Fan, G., and Yang, C. (2019). Effects of GDNF-transfected marrow stromal cells on rats with intracerebral hemorrhage. J. Stroke Cerebrovasc. Dis. 28, 2555–2562. doi: 10.1016/j.jstrokecerebrovasdis.2019.06.002
Ding, R., Lin, C., Wei, S., Zhang, N., Tang, L., Lin, Y., et al. (2017). Therapeutic benefits of mesenchymal stromal cells in a rat model of hemoglobin-induced hypertensive intracerebral hemorrhage. Mol. Cells. 40, 133–142. doi: 10.14348/molcells.2017.2251
Dooves, S., van der Knaap, M. S., and Heine, V. M. (2016). Stem cell therapy for white matter disorders: don’t forget the microenvironment! J. Inherit. Metab. Dis. 39, 513–518. doi: 10.1007/s10545-016-9925-1
Duan, S., Wang, F., Cao, J., and Wang, C. (2020). Exosomes derived from MicroRNA-146a-5p-enriched bone marrow mesenchymal stem cells alleviate intracerebral hemorrhage by inhibiting neuronal apoptosis and microglial M1 polarization. Drug Des. Devel. Ther. 14, 3143–3158. doi: 10.2147/DDDT.S255828
Feng, M., Li, Y., Han, Q., Bao, X., Yang, M., Zhu, H., et al. (2014). Preclinical safety evaluation of human mesenchymal stem cell transplantation in cerebrum of nonhuman primates. Int. J. Toxicol. 33, 403–411. doi: 10.1177/1091581814545244
Feng, M., Zhu, H., Zhu, Z., Wei, J., Lu, S., Li, Q., et al. (2011). Serial 18F-FDG PET demonstrates benefit of human mesenchymal stem cells in treatment of intracerebral hematoma: a translational study in a primate model. J. Nucl. Med. 52, 90–97. doi: 10.2967/jnumed.110.080325
Gao, F., Liu, F., Chen, Z., Hua, Y., Keep, R. F., and Xi, G. (2014). Hydrocephalus after intraventricular hemorrhage: the role of thrombin. J. Cereb. Blood Flow Metab. 34, 489–494. doi: 10.1038/jcbfm.2013.225
Gao, L., Xu, W., Li, T., Chen, J., Shao, A., Yan, F., et al. (2018). Stem Cell Therapy: a promising therapeutic method for intracerebral hemorrhage. Cell Transplant. 27, 1809–1824. doi: 10.1177/0963689718773363
Ghanta, S., Kwon, M. Y., Rosas, I. O., Liu, X., and Perrella, M. A. (2018). Mesenchymal stromal cell therapy: does the source matter? Crit. Care Med. 46, 343–345. doi: 10.1097/CCM.0000000000002894
Gong, Y., Gong, Y., Hou, Z., Guo, T., Deng, J., Hao, S., et al. (2019). Establishment of an experimental intracerebral haemorrhage model for mass effect research using a thermo-sensitive hydrogel. Sci. Rep. 9:13838. doi: 10.1038/s41598-019-50188-y
Gong, Y., Wang, Y., Qu, Q., Hou, Z., Guo, T., Xu, Y., et al. (2020). Nanoparticle encapsulated core-shell hydrogel for on-site BMSCs delivery protects from iron overload and enhances functional recovery. J. Control Release 320, 381–391. doi: 10.1016/j.jconrel.2020.01.029
Guo, T., Ren, P., Hao, S., and Wang, B. (2017). The underestimated role of mechanical stimuli in brain diseases and the relate d In vitro models. Curr. Pharm. Des. 23, 2161–2176. doi: 10.2174/1381612822666161027113200
Guo, T., Ren, P., Li, X., Luo, T., Gong, Y., Hao, S., et al. (2018). Neural injuries induced by hydrostatic pressure associated with mass effect after intracerebral hemorrhage. Sci. Rep. 8:9195. doi: 10.1038/s41598-018-27275-7
Han, Y., Li, X., Zhang, Y., Han, Y., Chang, F., and Ding, J. (2019). Mesenchymal stem cells for regenerative medicine. Cells 8:886. doi: 10.3390/cells8080886
Han, Y., Seyfried, D., Meng, Y., Yang, D., Schultz, L., Chopp, M., et al. (2018). Multipotent mesenchymal stromal cell-derived exosomes improve functional recovery after experimental intracerebral hemorrhage in the rat. J. Neurosurg. 131, 290–300. doi: 10.3171/2018.2.JNS171475
He, L., Si, G., Huang, J., Samuel, A. D. T., and Perrimon, N. (2018). Mechanical regulation of stem-cell differentiation by the stretch-activated Piezo channel. Nature 555, 103–106. doi: 10.1038/nature25744
Hu, C., and Li, L. (2018). Preconditioning influences mesenchymal stem cell properties in vitro and in vivo. J. Cell Mol. Med. 22, 1428–1442. doi: 10.1111/jcmm.13492
Hu, Y., Huang, L., Shen, M., Liu, Y., Liu, G., Wu, Y., et al. (2019). Pioglitazone protects compression-mediated apoptosis in nucleus pulposus mesenchymal stem cells by suppressing oxidative stress. Oxid. Med. Cell Longev. 2019:4764071. doi: 10.1155/2019/4764071
Hu, Y., Liu, N., Zhang, P., Pan, C., Zhang, Y., Tang, Y., et al. (2016). Preclinical studies of stem cell transplantation in intracerebral hemorrhage: a systemic review and meta-analysis. Mol. Neurobiol. 53, 5269–5277. doi: 10.1007/s12035-015-9441-6
Keep, R. F., Hua, Y., and Xi, G. (2012). Intracerebral haemorrhage: mechanisms of injury and therapeutic targets. Lancet Neurol. 11, 720–731. doi: 10.1016/s1474-4422(12)70104-7
Kim, H., Bae, C., Kook, Y. M., Koh, W. G., Lee, K., and Park, M. H. (2019). Mesenchymal stem cell 3D encapsulation technologies for biomimetic microenvironment in tissue regeneration. Stem Cell Res. Ther. 10:51. doi: 10.1186/s13287-018-1130-8
Kim, J. M., Lee, S. T., Chu, K., Jung, K. H., Song, E. C., Kim, S. J., et al. (2007). Systemic transplantation of human adipose stem cells attenuated cerebral inflammation and degeneration in a hemorrhagic stroke model. Brain Res. 1183, 43–50. doi: 10.1016/j.brainres.2007.09.005
Kim, K., Park, H. W., Moon, H. E., Kim, J. W., Bae, S., Chang, J. W., et al. (2015). The effect of human umbilical cord blood-derived mesenchymal stem cells in a collagenase-induced intracerebral hemorrhage rat model. Exp. Neurobiol. 24, 146–155. doi: 10.5607/en.2015.24.2.146
Kim, Y. E., Sung, S. I., Chang, Y. S., Ahn, S. Y., Sung, D. K., and Park, W. S. (2019). Thrombin preconditioning enhances therapeutic efficacy of human wharton’s jelly-derived mesenchymal stem cells in severe neonatal hypoxic ischemic encephalopathy. Int. J. Mol. Sci. 20:2477. doi: 10.3390/ijms20102477
Ko, E., Lee, K. Y., and Hwang, D. S. (2012). Human umbilical cord blood-derived mesenchymal stem cells undergo cellular senescence in response to oxidative stress. Stem Cells Dev. 21, 1877–1886. doi: 10.1089/scd.2011.0284
Kocaoemer, A., Kern, S., Kluter, H., and Bieback, K. (2007). Human AB serum and thrombin-activated platelet-rich plasma are suitable alternatives to fetal calf serum for the expansion of mesenchymal stem cells from adipose tissue. Stem Cells 25, 1270–1278. doi: 10.1634/stemcells.2006-0627
Kuramoto, Y., Takagi, T., Tatebayashi, K., Beppu, M., Doe, N., Fujita, M., et al. (2019). Intravenous administration of human adipose-derived stem cells ameliorates motor and cognitive function for intracerebral hemorrhage mouse model. Brain Res. 1711, 58–67. doi: 10.1016/j.brainres.2018.12.042
Lee, H. S., Kim, K. S., Lim, H. S., Choi, M., Kim, H. K., Ahn, H. Y., et al. (2015). Priming wharton’s jelly-derived mesenchymal stromal/stem cells with ROCK inhibitor improves recovery in an intracerebral hemorrhage model. J. Cell Biochem. 116, 310–319. doi: 10.1002/jcb.24969
Li, G., Yu, H., Liu, N., Zhang, P., Tang, Y., Hu, Y., et al. (2019). Overexpression of CX3CR1 in adipose-derived stem cells promotes cell migration and functional recovery after experimental intracerebral hemorrhage. Front. Neurosci. 13:462. doi: 10.3389/fnins.2019.00462
Li, P., Gong, Z., Shultz, L. D., and Ren, G. (2019). Mesenchymal stem cells: from regeneration to cancer. Pharmacol. Ther. 200, 42–54. doi: 10.1016/j.pharmthera.2019.04.005
Li, Q., Han, X., Lan, X., Gao, Y., Wan, J., Durham, F., et al. (2017). Inhibition of neuronal ferroptosis protects hemorrhagic brain. JCI Insight 2:e90777. doi: 10.1172/jci.insight.90777
Liao, W., Zhong, J., Yu, J., Xie, J., Liu, Y., Du, L., et al. (2009). Therapeutic benefit of human umbilical cord derived mesenchymal stromal cells in intracerebral hemorrhage rat: implications of anti-inflammation and angiogenesis. Cell. Physiol. Biochem. 24, 307–316.
Liem, N. T., Huyen, T. L., Huong, L. T., Doan, N. V., Anh, B. V., Anh, N. T. P., et al. (2019). Outcomes of bone marrow mononuclear cell transplantation for neurological sequelae due to intracranial hemorrhage incidence in the neonatal period: report of four cases. Front. Pediatr. 7:543. doi: 10.3389/fped.2019.00543
Lin, M., Mao, S., Wang, J., Xing, J., Wang, Y., Cai, K., et al. (2018). Adsorption force of fibronectin controls transmission of cell traction force and subsequent stem cell fate. Biomaterials 162, 170–182. doi: 10.1016/j.biomaterials.2018.01.036
Liu, W., Li, R., Yin, J., Guo, S., Chen, Y., Fan, H., et al. (2019). Mesenchymal stem cells alleviate the early brain injury of subarachnoid hemorrhage partly by suppression of Notch1-dependent neuroinflammation: involvement of Botch. J. Neuroinflammation 16:8. doi: 10.1186/s12974-019-1396-5
Lu, W., Zhao, M., Rajbhandary, S., Xie, F., Chai, X., Mu, J., et al. (2013). Free iron catalyzes oxidative damage to hematopoietic cells/mesenchymal stem cells in vitro and suppresses hematopoiesis in iron overload patients. Eur. J. Haematol. 91, 249–261. doi: 10.1111/ejh.12159
Ma, X., Qin, J., Song, B., Shi, C., Zhang, R., Liu, X., et al. (2015). Stem cell-based therapies for intracerebral hemorrhage in animal model: a meta-analysis. Neurol. Sci. 36, 1311–1317. doi: 10.1007/s10072-015-2238-6
Mao, S., Xi, G., Keep, R. F., and Hua, Y. (2016). Role of lipocalin-2 in thrombin-induced brain injury. Stroke 47, 1078–1084. doi: 10.1161/STROKEAHA.115.012153
Mello, T. G., Rosado-de-Castro, P. H., Campos, R. M. P., Vasques, J. F., Rangel-Junior, W. S., Mattos, R., et al. (2020). Intravenous human umbilical cord-derived mesenchymal stromal cell administration in models of moderate and severe intracerebral hemorrhage. Stem Cells Dev. 29, 586–598. doi: 10.1089/scd.2019.0176
Min, S., Kim, O. J., Bae, J., and Chung, T. N. (2018). Effect of pretreatment with the NADPH oxidase inhibitor apocynin on the therapeutic efficacy of human placenta-derived mesenchymal stem cells in intracerebral hemorrhage. Int. J. Mol. Sci. 19:3679. doi: 10.3390/ijms19113679
Mohammadzadeh, M., Halabian, R., Gharehbaghian, A., Amirizadeh, N., Jahanian-Najafabadi, A., Roushandeh, A. M., et al. (2012). Nrf-2 overexpression in mesenchymal stem cells reduces oxidative stress-induced apoptosis and cytotoxicity. Cell Stress Chaperones 17, 553–565. doi: 10.1007/s12192-012-0331-9
Murphy, M. B., Moncivais, K., and Caplan, A. I. (2013). Mesenchymal stem cells: environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 45:e54. doi: 10.1038/emm.2013.94
Nahhas, M. I., and Hess, D. C. (2018). Stem cell therapy in cerebrovascular disease. Curr. Treat. Opt. Neurol. 20:49. doi: 10.1007/s11940-018-0532-3
Otero, L., Bonilla, C., Aguayo, C., Zurita, M., and Vaquero, J. (2010). Intralesional administration of allogeneic bone marrow stromal cells reduces functional deficits after intracerebral hemorrhage. Histol. Histopathol. 25, 453–461.
Park, W. S., Sung, S. I., Ahn, S. Y., Sung, D. K., Im, G. H., Yoo, H. S., et al. (2016). Optimal timing of mesenchymal stem cell therapy for neonatal intraventricular hemorrhage. Cell Transplant. 25, 1131–1144. doi: 10.3727/096368915X689640
Qu, J., Chen, W., Hu, R., and Feng, H. (2016). The Injury and therapy of reactive oxygen species in intracerebral hemorrhage looking at mitochondria. Oxid. Med. Cell Longev. 2016:2592935. doi: 10.1155/2016/2592935
Qureshi, A. I., Mendelow, A. D., and Hanley, D. F. (2009). Intracerebral haemorrhage. Lancet 373, 1632–1644.
Seyfried, D., Ding, J., Han, Y., Li, Y., Chen, J., and Chopp, M. (2006). Effects of intravenous administration of human bone marrow stromal cells after intracerebral hemorrhage in rats. J. Neurosurg. 104, 313–318.
Seyfried, D. M., Han, Y., Yang, D., Ding, J., Savant-Bhonsale, S., Shukairy, M. S., et al. (2008). Mannitol enhances delivery of marrow stromal cells to the brain after experimental intracerebral hemorrhage. Brain Res. 1224, 12–19. doi: 10.1016/j.brainres.2008.05.080
Shen, H., Yao, X., Li, H., Li, X., Zhang, T., Sun, Q., et al. (2018). Role of exosomes derived from miR-133b modified MSCs in an experimental rat model of intracerebral hemorrhage. J. Mol. Neurosci. 64, 421–430. doi: 10.1007/s12031-018-1041-2
Sherman, L. S., Romagano, M. P., Williams, S. F., and Rameshwar, P. (2019). Mesenchymal stem cell therapies in brain disease. Semin. Cell Dev. Biol. 95, 111–119. doi: 10.1016/j.semcdb.2019.03.003
Sun, J., Wei, Z. Z., Gu, X., Zhang, J. Y., Zhang, Y., Li, J., et al. (2015). Intranasal delivery of hypoxia-preconditioned bone marrow-derived mesenchymal stem cells enhanced regenerative effects after intracerebral hemorrhagic stroke in mice. Exp. Neurol. 272, 78–87. doi: 10.1016/j.expneurol.2015.03.011
Sun, Z., Wu, K., Gu, L., Huang, L., Zhuge, Q., Yang, S., et al. (2020). IGF-1R stimulation alters microglial polarization via TLR4/NF-kappaB pathway after cerebral hemorrhage in mice. Brain Res. Bull. 164, 221–234. doi: 10.1016/j.brainresbull.2020.08.026
Sung, D. K., Sung, S. I., Ahn, S. Y., Chang, Y. S., and Park, W. S. (2019). Thrombin preconditioning boosts biogenesis of extracellular vesicles from mesenchymal stem cells and enriches their cargo contents via protease-activated receptor-mediated signaling pathways. Int. J. Mol. Sci. 20:2899. doi: 10.3390/ijms20122899
Tang, T., Liu, X. J., Zhang, Z. Q., Zhou, H. J., Luo, J. K., Huang, J. F., et al. (2007). Cerebral angiogenesis after collagenase-induced intracerebral hemorrhage in rats. Brain Res. 1175, 134–142. doi: 10.1016/j.brainres.2007.08.028
Tsang, K. S., Ng, C. P. S., Zhu, X. L., Wong, G. K. C., Lu, G., Ahuja, A. T., et al. (2017). Phase I/II randomized controlled trial of autologous bone marrow-derived mesenchymal stem cell therapy for chronic stroke. World J. Stem Cells. 9, 133–143. doi: 10.4252/wjsc.v9.i8.133
van Asch, C. J. J., Luitse, M. J. A., Rinkel, G. J. E., van der Tweel, I., Algra, A., and Klijn, C. J. M. (2010). Incidence, case fatality, and functional outcome of intracerebral haemorrhage over time, according to age, sex, and ethnic origin: a systematic review and meta-analysis. Lancet Neurol. 9, 167–176. doi: 10.1016/s1474-4422(09)70340-0
Vaquero, J., Otero, L., Bonilla, C., Aguayo, C., Rico, M. A., Rodriguez, A., et al. (2013). Cell therapy with bone marrow stromal cells after intracerebral hemorrhage: impact of platelet-rich plasma scaffolds. Cytotherapy 15, 33–43. doi: 10.1016/j.jcyt.2012.10.005
Vining, K. H., and Mooney, D. J. (2017). Mechanical forces direct stem cell behaviour in development and regeneration. Nat. Rev. Mol. Cell Biol. 18, 728–742. doi: 10.1038/nrm.2017.108
Wakai, T., Narasimhan, P., Sakata, H., Wang, E., Yoshioka, H., Kinouchi, H., et al. (2016). Hypoxic preconditioning enhances neural stem cell transplantation therapy after intracerebral hemorrhage in mice. J. Cereb. Blood Flow Metab. 36, 2134–2145. doi: 10.1177/0271678X15613798
Wan, J., Ren, H., and Wang, J. (2019). Iron toxicity, lipid peroxidation and ferroptosis after intracerebral haemorrhage. Stroke Vasc. Neurol. 4, 93–95. doi: 10.1136/svn-2018-000205
Wang, C., Cao, J., Duan, S., Xu, R., Yu, H., Huo, X., et al. (2020). Effect of MicroRNA-126a-3p on bone marrow mesenchymal stem cells repairing blood-brain barrier and nerve injury after intracerebral hemorrhage. J. Stroke Cerebrovasc. Dis. 29:104748. doi: 10.1016/j.jstrokecerebrovasdis.2020.104748
Wang, C., Fei, Y., Xu, C., Zhao, Y., and Pan, Y. (2015). Bone marrow mesenchymal stem cells ameliorate neurological deficits and blood-brain barrier dysfunction after intracerebral hemorrhage in spontaneously hypertensive rats. Int. J. Clin. Exp. Pathol. 8, 4715–4724.
Wang, S. P., Wang, Z. H., Peng, D. Y., Li, S. M., Wang, H., and Wang, X. H. (2012). Therapeutic effect of mesenchymal stem cells in rats with intracerebral hemorrhage: reduced apoptosis and enhanced neuroprotection. Mol. Med. Rep. 6, 848–854. doi: 10.3892/mmr.2012.997
Wang, Y., Han, Z.-B., Ma, J., Zuo, C., Geng, J., Gong, W., et al. (2012). A toxicity study of multiple-administration human umbilical cord mesenchymal stem cells in cynomolgus monkeys. Stem Cells Dev. 21, 1401–1408. doi: 10.1089/scd.2011.0441
Wang, Y. H., Wu, D. B., Chen, B., Chen, E. Q., and Tang, H. (2018). Progress in mesenchymal stem cell-based therapy for acute liver failure. Stem Cell Res. Ther. 9:227. doi: 10.1186/s13287-018-0972-4
Wilkinson, D. A., Pandey, A. S., Thompson, B. G., Keep, R. F., Hua, Y., and Xi, G. (2018). Injury mechanisms in acute intracerebral hemorrhage. Neuropharmacology 134, 240–248. doi: 10.1016/j.neuropharm.2017.09.033
Williams, A. M., Dennahy, I. S., Bhatti, U. F., Halaweish, I., Xiong, Y., Chang, P., et al. (2019). Mesenchymal stem cell-derived exosomes provide neuroprotection and improve long-term neurologic outcomes in a swine model of traumatic brain injury and hemorrhagic shock. J. Neurotrauma. 36, 54–60. doi: 10.1089/neu.2018.5711
Wu, S., and Anderson, C. S. (2020). A need to re-focus efforts to improve long-term prognosis after stroke in China. Lancet Glob. Health. 8, e468–e469. doi: 10.1016/s2214-109x(20)30086-3
Wu, S., Wu, B., Liu, M., Chen, Z., Wang, W., Anderson, C. S., et al. (2019). Stroke in China: advances and challenges in epidemiology, prevention, and management. Lancet Neurol. 18, 394–405. doi: 10.1016/s1474-4422(18)30500-3
Xie, J., Wang, B., Wang, L., Dong, F., Bai, G., and Liu, Y. (2016). Intracerebral and intravenous transplantation represents a favorable approach for application of human umbilical cord mesenchymal stromal cells in intracerebral hemorrhage rats. Med. Sci. Monit. 22, 3552–3561. doi: 10.12659/msm.900512
Xiong, L., Sun, L., Zhang, Y., Peng, J., Yan, J., and Liu, X. (2020). Exosomes from bone marrow mesenchymal stem cells can alleviate early brain injury after subarachnoid hemorrhage through miRNA129-5p-HMGB1 pathway. Stem Cells Dev. 29, 212–221. doi: 10.1089/scd.2019.0206
Xu, Y., Yuan, H., Luo, Y., Zhao, Y. J., and Xiao, J. H. (2020). Ganoderic acid D protects human amniotic mesenchymal stem cells against oxidative stress-induced senescence through the PERK/NRF2 signaling pathway. Oxid. Med. Cell Longev. 2020:8291413. doi: 10.1155/2020/8291413
Yang, C., Zhou, L., Gao, X., Chen, B., Tu, J., Sun, H., et al. (2011). Neuroprotective effects of bone marrow stem cells overexpressing glial cell line-derived neurotrophic factor on rats with intracerebral hemorrhage and neurons exposed to hypoxia/reoxygenation. Neurosurgery 68, 691–704. doi: 10.1227/NEU.0b013e3182098a8a
Yang, F., Yan, G., Li, Y., Han, Z., Zhang, L., Chen, S., et al. (2016). Astragalus polysaccharide attenuated iron overload-induced dysfunction of mesenchymal stem cells via suppressing mitochondrial ROS. Cell Physiol. Biochem. 39, 1369–1379. doi: 10.1159/000447841
Yang, Y., Ye, Y., Su, X., He, J., Bai, W., and He, X. (2017). MSCs-derived exosomes and neuroinflammation, neurogenesis and therapy of traumatic brain injury. Front. Cell Neurosci. 11:55. doi: 10.3389/fncel.2017.00055
Yu, Z. Y., Ma, D., He, Z. C., Liu, P., Huang, J., Fang, Q., et al. (2018). Heme oxygenase-1 protects bone marrow mesenchymal stem cells from iron overload through decreasing reactive oxygen species and promoting IL-10 generation. Exp. Cell Res. 362, 28–42. doi: 10.1016/j.yexcr.2017.10.029
Yuan, Y., Xu, F., Cao, Y., Xu, L., Yu, C., Yang, F., et al. (2019). Iron accumulation leads to bone loss by inducing mesenchymal stem cell apoptosis through the activation of caspase3. Biol. Trace Elem. Res. 187, 434–441. doi: 10.1007/s12011-018-1388-9
Zahra, K., Turnbull, M. T., Zubair, A. C., Siegel, J. L., Venegas-Borsellino, C. P., Tawk, R. G., et al. (2020). A combined approach to intracerebral hemorrhage: intravenous mesenchymal stem cell therapy with minimally invasive hematoma evacuation. J. Stroke Cerebrovasc. Dis. 29:104931. doi: 10.1016/j.jstrokecerebrovasdis.2020.104931
Zhang, F., Peng, W., Zhang, J., Dong, W., Yuan, D., Zheng, Y., et al. (2019). New strategy of bone marrow mesenchymal stem cells against oxidative stress injury via Nrf2 pathway: oxidative stress preconditioning. J. Cell Biochem. 120, 19902–19914. doi: 10.1002/jcb.29298
Zhang, H., Huang, Z., Xu, Y., and Zhang, S. (2006). Differentiation and neurological benefit of the mesenchymal stem cells transplanted into the rat brain following intracerebral hemorrhage. Neurol. Res. 28, 104–112.
Zhang, Q., Shang, X., Hao, M., Zheng, M., Li, Y., and Liang, Z. (2015). Effects of human umbilical cord mesenchymal stem cell transplantation combined with minimally invasive hematoma aspiration on intracerebral hemorrhage in rats. Am. J. Trans. Res. 7, 2176–2186.
Zhang, R. (2016). Effect of autologous bone mesenchymal stem cell transplantation on neurological function in rehabilitation period of multifocal cerebral hemorrhage. J. Hainan Med. Univ. 22, 142–145.
Zhang, Y., Chopp, M., Meng, Y., Katakowski, M., Xin, H., Mahmood, A., et al. (2015). Effect of exosomes derived from multipluripotent mesenchymal stromal cells on functional recovery and neurovascular plasticity in rats after traumatic brain injury. J. Neurosurg. 122, 856–867. doi: 10.3171/2014.11.JNS14770
Zhang, Y., Deng, H., Hu, Y., Pan, C., Wu, G., Li, Q., et al. (2019). Adipose-derived mesenchymal stem cells stereotactic transplantation alleviate brain edema from intracerebral hemorrhage. J. Cell Biochem. 120, 14372–14382. doi: 10.1002/jcb.28693
Zhao, H., Li, Y., Chen, L., Shen, C., Xiao, Z., Xu, R., et al. (2019). HucMSCs-derived miR-206-knockdown exosomes contribute to neuroprotection in subarachnoid hemorrhage induced early brain injury by targeting BDNF. Neuroscience 417, 11–23. doi: 10.1016/j.neuroscience.2019.07.051
Zheng, H., Zhang, B., Chhatbar, P. Y., Dong, Y., Alawieh, A., Lowe, F., et al. (2018). Mesenchymal stem cell therapy in stroke: a systematic review of literature in pre-clinical and clinical research. Cell Transplant. 27, 1723–1730. doi: 10.1177/0963689718806846
Zhou, H., Zhang, H., Yan, Z., and Xu, R. (2016). Transplantation of human amniotic mesenchymal stem cells promotes neurological recovery in an intracerebral hemorrhage rat model. Biochem. Biophys. Res. Commun. 475, 202–208. doi: 10.1016/j.bbrc.2016.05.075
Zhou, X., Hong, Y., Zhang, H., and Li, X. (2020). Mesenchymal stem cell senescence and rejuvenation: current status and challenges. Front. Cell Dev. Biol. 8:364. doi: 10.3389/fcell.2020.00364
Keywords: intracerebral hemorrhage, mesenchymal stem cells, different species, pathological microenvironment, optimizing strategy
Citation: Gong Y-h, Hao S-l and Wang B-c (2021) Mesenchymal Stem Cells Transplantation in Intracerebral Hemorrhage: Application and Challenges. Front. Cell. Neurosci. 15:653367. doi: 10.3389/fncel.2021.653367
Received: 14 January 2021; Accepted: 01 March 2021;
Published: 24 March 2021.
Edited by:
Haruyuki Kamiya, Hokkaido University, JapanReviewed by:
Zhouping Tang, Huazhong University of Science and Technology, ChinaCopyright © 2021 Gong, Hao and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shi-lei Hao, c2hpbGVpX2hhb0BjcXUuZWR1LmNu; Bo-chu Wang, d2FuZ2JjMjAwMEAxMjYuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.