
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Neurosci. , 19 August 2020
Sec. Non-Neuronal Cells
Volume 14 - 2020 | https://doi.org/10.3389/fncel.2020.00239
This article is part of the Research Topic Journey to the Center of the Brain: Cell Physiology and Intercellular Communication In White Matter View all 10 articles
 Olaf Jahn1
Olaf Jahn1 Sophie B. Siems2
Sophie B. Siems2 Kathrin Kusch2
Kathrin Kusch2 Dörte Hesse1Ramona B. Jung2Thomas Liepold1Marina Uecker1
Dörte Hesse1Ramona B. Jung2Thomas Liepold1Marina Uecker1 Ting Sun2
Ting Sun2 Hauke B. Werner2*
Hauke B. Werner2*Myelin membranes are dominated by lipids while the complexity of their protein composition has long been considered to be low. However, numerous additional myelin proteins have been identified since. Here we revisit the proteome of myelin biochemically purified from the brains of healthy c56Bl/6N-mice utilizing complementary proteomic approaches for deep qualitative and quantitative coverage. By gel-free, label-free mass spectrometry, the most abundant myelin proteins PLP, MBP, CNP, and MOG constitute 38, 30, 5, and 1% of the total myelin protein, respectively. The relative abundance of myelin proteins displays a dynamic range of over four orders of magnitude, implying that PLP and MBP have overshadowed less abundant myelin constituents in initial gel-based approaches. By comparisons with published datasets we evaluate to which degree the CNS myelin proteome correlates with the mRNA and protein abundance profiles of myelin and oligodendrocytes. Notably, the myelin proteome displays only minor changes if assessed after a post-mortem delay of 6 h. These data provide the most comprehensive proteome resource of CNS myelin so far and a basis for addressing proteomic heterogeneity of myelin in mouse models and human patients with white matter disorders.
In the central nervous system (CNS) of vertebrates, the velocity of nerve conduction is accelerated by the insulation of axons with multiple layers of myelin membrane provided by oligodendrocytes (Nave and Werner, 2014; Snaidero and Simons, 2017). Compared to other cellular membranes myelin is unusually enriched for lipids, in particular cholesterol, galactolipids and plasmalogens (Norton and Poduslo, 1973a; Schmitt et al., 2015; Poitelon et al., 2020). Indeed, the biogenesis of myelin may involve the coalescence of lipid-rich membrane-microdomains in the oligodendroglial secretory pathway (Lee, 2001; Chrast et al., 2011). Notably, the dominant CNS myelin protein, proteolipid protein (PLP), displays a high affinity to cholesterol-rich membrane-microdomains (Simons et al., 2000; Werner et al., 2013). PLP and other cholesterol-associated myelin proteins may thus enhance the coalescence and intracellular traffic of prospective myelin membranes (Schardt et al., 2009). Indeed, both cholesterol and PLP are rate-limiting for myelination, as demonstrated by the dysmyelination observed in mice lacking oligodendroglial cholesterol synthesis (Saher et al., 2005) or PLP-expression (Yool et al., 2001; Möbius et al., 2008; de Monasterio-Schrader et al., 2013).
As a key stage of myelination, the compaction of adjacent CNS myelin layers requires myelin basic protein (MBP), as evidenced by the complete lack of compact myelin in the CNS of MBP-deficient shiverer-mice (Roach et al., 1985). It is now thought that MBP both displaces filamentous actin and cytoskeleton-associated proteins (Nawaz et al., 2015; Zuchero et al., 2015; Snaidero et al., 2017) and saturates negative charges of the headgroups of phosphatidylinositol-4,5-bisphosphate (PIP2) on the cytoplasmic myelin membrane surfaces (Musse et al., 2008; Nawaz et al., 2009, 2013) thereby pulling together and compacting myelin membranes at the major dense line (Raasakka et al., 2017).
It has been noted already in the early 1970s that PLP and MBP constitute the most abundant CNS myelin proteins. At that time the methods were developed for the enrichment of myelin from nervous tissue by sucrose density gradient centrifugation (Norton and Poduslo, 1973b; Erwig et al., 2019a) separation by one-dimensional (1D)-polyacrylamide gel electrophoresis (SDS-PAGE) and protein staining using Buffalo Black (Morris et al., 1971) Fast Green (Morell et al., 1972) or Coomassie Blue (Magno-Sumbilla and Campagnoni, 1977). Indeed, only few bands were visible that we now know are mainly constituted by PLP, MBP and cyclic nucleotide phosphodiesterase (CNP; Sprinkle et al., 1983). Deficiency of CNP in mice impairs both the ultrastructure of myelin and the long-term preservation of axonal integrity (Lappe-Siefke et al., 2003; Edgar et al., 2009; Patzig et al., 2016; Snaidero et al., 2017).
Evidently, the protein composition of myelin is more complex when considering that various additional myelin proteins have been identified, including myelin associated glycoprotein (MAG; Quarles, 2007; Myllykoski et al., 2018), myelin oligodendrocyte glycoprotein (MOG; Johns and Bernard, 1999; von Büdingen et al., 2015), and claudin 11 (CLDN11; Gow et al., 1999; Denninger et al., 2015). This insight motivated attempts to utilize the emerging mass spectrometric techniques to approach all myelin proteins at once, thereby covering the entire myelin proteome. Indeed, purified myelin is suited for systematic assessment of its molecular constituents (De Monasterio-Schrader et al., 2012; Gopalakrishnan et al., 2013). Most early approaches involved 2D-gels (Taylor et al., 2004; Vanrobaeys et al., 2005; Werner et al., 2007), soon to be complemented by gel-free shotgun-approaches (Vanrobaeys et al., 2005; Roth et al., 2006; Dhaunchak et al., 2010) and hybrid workflows (Ishii et al., 2009). However, first systematic information on the relative abundance of myelin proteins was achieved by label-free quantification involving peptide-separation by liquid chromatography (LC) coupled to detection with quadrupole time-of-flight (QTOF) mass spectrometery (MS) (Jahn et al., 2009) or by chemical peptide labeling with isobaric tags for relative and absolute quantitation (iTRAQ) and subsequent LC-MS-analysis (Manrique-Hoyos et al., 2012). A meta-analysis of the approaches to the myelin proteome published by 2012 is available (De Monasterio-Schrader et al., 2012). Since then, label-free protein quantification by LC-MS has proven useful in the differential analysis of myelin in mouse models including mice lacking PLP, CNP, or MAG (Patzig et al., 2016). For example, this approach allowed identifying cytoskeletal septin filaments to stabilize the ultrastructure of CNS myelin, thereby preventing the formation of pathological myelin outfoldings (Patzig et al., 2016; Erwig et al., 2019b).
The intention of this work was to both establish an updated comprehensive compendium of the proteins associated with CNS myelin and to accurately quantify their relative abundance, as recently achieved for the proteome of myelin in the peripheral nervous system (Siems et al., 2020). To this aim we combined various gel-based and gel-free proteomic techniques. In particular, we used nano-flow ultra-performance liquid chromatography (nanoUPLC) for peptide separation and an ion mobility-enabled QTOF-system for label-free protein quantification by data-independent acquisition (DIA) mass spectrometry in an alternating low and elevated energy mode (MSE). While the MSE-mode allows quantifying myelin proteins with the required dynamic range of over four orders of magnitude, an ion mobility-enhanced version thereof [referred to as ultra-definition (UD)-MSE] covers about twice as many myelin-associated proteins, though at the expense of dynamic range. Our workflow thus facilitates both to reliably quantify the exceptionally abundant PLP, MBP, and CNP and to appreciate the complexity of low-abundant myelin constituents.
Male c57BL/6N wild-type mice at postnatal day 75 (P75) were used for all experiments except for the differential analysis of myelin purified from brains immediately frozen after dissection compared to a post-mortem delay of 6 h at room temperature (Figure 4), for which female c57BL/6N wild-type mice at P56 were used. Mice were bred and kept in the animal facility of the Max Planck Institute of Experimental Medicine and sacrificed by cervical dislocation. For the procedure of sacrificing mice for subsequent preparation of tissue, all regulations given in the German animal protection law (TierSchG §4) are followed. Since sacrificing of rodents is not an experiment on animals according to §7 Abs. 2 Satz 3 TierSchG, no specific authorization or notification is required for the present work.
A myelin-enriched light-weight membrane fraction was biochemically purified from mouse brains by sucrose density centrifugation and osmotic shocks as recently described in detail (Erwig et al., 2019a). Mice were sacrificed by cervical dislocation at the indicated ages as three biological replicates per condition (n = 3). Protein concentration was determined using the DC Protein Assay Kit (Bio-Rad). Initial quality control by gel electrophoresis and silver staining of gels was performed as described (de Monasterio-Schrader et al., 2013; Joseph et al., 2019). Briefly, samples were separated on a 12% SDS-PAGE gel (1 h at 200 V) using the Bio-Rad system, fixated overnight in 10% [v/v] acetic acid and 40% [v/v] ethanol and then washed in 30% ethanol (2 × 20 min) and ddH2O (1 × 20 min). For sensitization, gels were incubated 1 min in 0.012% [v/v] Na2S2O3 and subsequently washed with ddH2O (3 × 20 s). For silver staining, gels were impregnated for 20 min in 0.2% [w/v] AgNO3/0.04% formaldehyde, washed with ddH2O (3 × 20 s) and developed in 3% [w/v] Na2CO3/0.04% [w/v] formaldehyde. The reaction was stopped by exchanging the solution with 5% [v/v] acetic acid.
Gel-electrophoretic separation of myelin proteins with different pre-cast gel systems (Serva) was performed essentially as recently described in detail (Erwig et al., 2019a). Briefly, 1D separations were performed with 5 μg protein load before (pre-wash) or after (post-wash) subjecting myelin to consecutive high-salt and high-pH washing/centrifugation cycles as previously described (Werner et al., 2007; Jahn et al., 2013). Automated tryptic in-gel digestion of proteins in gel bands (Schmidt et al., 2013) and protein identification by LC-MS was performed as described (Ott et al., 2015). For 2D separations, myelin was first delipidated by methanol/chloroform precipitation and 300 μg protein was loaded on a 24 cm immobilized non-linear pH-gradient 3-12 strip (Serva) by active rehydration (Erwig et al., 2019a). Automated tryptic in-gel digestion of proteins in gel spots and protein identification by MALDI-TOF mass spectrometry was performed as described (Jahn et al., 2006; Werner et al., 2007).
In-solution digestion of myelin proteins according to an automated filter-aided sample preparation (FASP) protocol (Erwig et al., 2019a) and LC-MS-analysis by different MSE-type data-independent acquisition (DIA) mass spectrometry approaches was performed as recently established for PNS myelin (Siems et al., 2020). Briefly, protein fractions corresponding to 10 μg myelin protein were dissolved in lysis buffer (1% ASB-14, 7 M urea, 2 M thiourea, 10 mM DTT, 0.1 M Tris pH 8.5) and processed according to a CHAPS-based FASP protocol in centrifugal filter units (30 kDa MWCO, Merck Millipore). After removal of the detergents, protein alkylation with iodoacetamide, and buffer exchange to digestion buffer [50 mM ammonium bicarbonate (ABC), 10% acetonitrile], proteins were digested overnight at 37°C with 400 ng trypsin. Tryptic peptides were recovered by centrifugation and extracted with 40 μl of 50 mM ABC and 40 μl of 1% trifluoroacetic acid (TFA), respectively. Combined flow-through were directly subjected to LC-MS-analysis. For quantification according to the TOP3 approach (Silva et al., 2006), aliquots were spiked with 10 fmol/μl of yeast enolase-1 tryptic digest or Hi3 EColi standard (Waters Corporation), the latter containing a set of quantified synthetic peptides derived from E. coli. Chaperone protein ClpB.
Nanoscale reversed-phase UPLC separation of tryptic peptides was performed with a nanoAcquity UPLC system equipped with a Symmetry C18 5 μm, 180 μm × 20 mm trap column and a HSS T3 C18 1.8 μm, 75 μm × 250 mm analytical column (Waters Corporation) maintained at 45°C. Peptides were separated over 120 min at a flow rate of 300 nl/min with a gradient comprising two linear steps of 3–35% mobile phase B (acetonitrile containing 0.1% formic acid) in 105 min and 35–60% mobile phase B in 15 min, respectively. Mass spectrometric analysis of tryptic peptides was performed using a Synapt G2-S QTOF mass spectrometer equipped with ion mobility option (Waters Corporation). UDMSE analysis was performed in the ion mobility-enhanced data-independent acquisition mode with drift time-specific collision energies as described in detail (Distler et al., 2014a, 2016). Continuum LC-MS data were processed using Waters ProteinLynx Global Server (PLGS) and searched against a custom database compiled by adding the sequence information for yeast enolase 1, E. coli Chaperone protein ClpB and porcine trypsin to the UniProtKB/Swiss-Prot mouse proteome and by appending the reversed sequence of each entry to enable the determination of false discovery rate (FDR). Precursor and fragment ion mass tolerances were automatically determined by PLGS and were typically below 5 ppm for precursor ions and below 10 ppm (root mean square) for fragment ions. Carbamidomethylation of cysteine was specified as fixed and oxidation of methionine as variable modification. One missed trypsin cleavage was allowed. Minimal ion matching requirements were two fragments per peptide, five fragments per protein, and one peptide per protein. The FDR for protein identification was set to 1% threshold.
For post-identification analysis including TOP3 quantification of proteins, the freely available software ISOQuant1 was used (Kuharev et al., 2015). Only peptides with a minimum length of seven amino acids that were identified with scores above or equal to 5.5 in at least two runs were considered. FDR for both peptides and proteins was set to 1% threshold and only proteins reported by at least two peptides (one of which unique) were quantified as parts per million (ppm) abundance values (i.e., the relative amount (w/w) of each protein in respect to the sum over all detected proteins). The Bioconductor R packages “limma” and “q-value” were used to detect significant changes in protein abundance by moderated t-statistics as described (Ambrozkiewicz et al., 2018; Siems et al., 2020). For proteome profiling of wild-type myelin by MSE and UDMSE, three independent experiments were performed, each with three biological replicates and sample processing with duplicate digestion and injection, resulting in a total of 12 LC-MS runs per experiment. Abundance values in ppm are given as averages of the four technical replicates per biological replicate and only proteins quantified in at least two out of three experiments are reported in the proteome resource (Supplementary Table S1). Proteins identified as contaminants from blood (albumin, hemoglobin) or hair cells (keratins) were removed from the list. Proteome profiling comparing wild-type myelin without and with post-mortem delay (Figure 4 and Supplementary Table S2) was performed with three biological replicates and duplicate digestion, resulting in a total of 6 LC-MS runs per condition. Data acquisition was performed in the DRE-UDMSE mode (Siems et al., 2020) i.e., a deflection device was cycled between full (100% for 0.4 s) and reduced (5% for 0.4 s) ion transmission during one 0.8 s full scan, thereby providing a compromise between identification rates and dynamic range.
Published single-cell RNA-sequencing (scRNA-seq) gene expression matrices from datasets GSE60361 (Zeisel et al., 2015), GSE75330 (Marques et al., 2016), and GSE113973 (Falcão et al., 2018), were obtained from Gene Expression Omnibus (GEO) and analyzed using R package Seurat v3.1.0 (Butler et al., 2018; Stuart et al., 2019). Mature oligodendrocyte cell populations were selected from each dataset as specified in the results section and normalized gene counts were used for calculating average expression profiles across single cells. Bulk proteome and transcriptome datasets specified in the results section were used as supplied in the Supplementary Tables to the respective publications.
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al., 2019) partner repository with dataset identifier PXD020007. Pie chart, scatter plots, volcano plot and heatmap were prepared in Microsoft Excel 2013 and GraphPad Prism 8. Area-proportional Venn diagrams were prepared using BioVenn (Hulsen et al., 2008)2. Trans-membrane domains were predicted using TMHMM Server v. 2.0 (Krogh et al., 2001)3 and Phobius (Käll et al., 2007)4.
To purify CNS myelin, we applied an established protocol (Erwig et al., 2019a) to prepare a light-weight membrane fraction from the brains of healthy c57Bl/6N-mice at P75. Aiming to systematically identify myelin-associated proteins we used five complementary approaches as summarized in Figure 1A. As the most straightforward way of preparing myelin for proteomic analysis, we separated proteins by 1D-SDS-PAGE and sectioned the lane into 24 equally sized slices (Figure 1B), which we subjected to automated tryptic in-gel digest followed by LC-MS-analysis, thereby identifying 788 proteins (Supplementary Table S1). When we subjected myelin to an additional washing step of high-pH and high-salt conditions (Figure 1B) to deplete soluble and peripheral membrane proteins before 1D-SDS-PAGE-separation and mass spectrometry, we identified 521 proteins (Supplementary Table S1). To establish a reference map of myelin proteins including proteoforms, we increased the resolving power of protein separation by subjecting myelin to 2D-gel electrophoresis with isoelectric focusing (IEF) in the first and horizontal SDS-PAGE in the second dimension (Figure 1C). We stained the comprised proteins with colloidal Coomassie (CBB250), picked CBB250-labeled gel-plugs (i.e., protein spots) for automated tryptic in-gel digestion and identified the proteins by peptide mass fingerprint (PMF) and MS/MS-fragment ion mass spectra, both acquired on a MALDI-TOF mass spectrometer. We identified 181 non-redundant proteins from 352 spots (Supplementary Table S1). Thereby we expanded our previous myelin protein map (131 non-redundant proteins from 217 spots, Werner et al., 2007), mainly owing to increased resolution in the first dimension by utilizing longer IEF-strips with a wider pH-range (Erwig et al., 2019a). When comparing the proteins identified using the three gel-based approaches we found a total of 930 proteins with a fair overlap (Figure 1D).
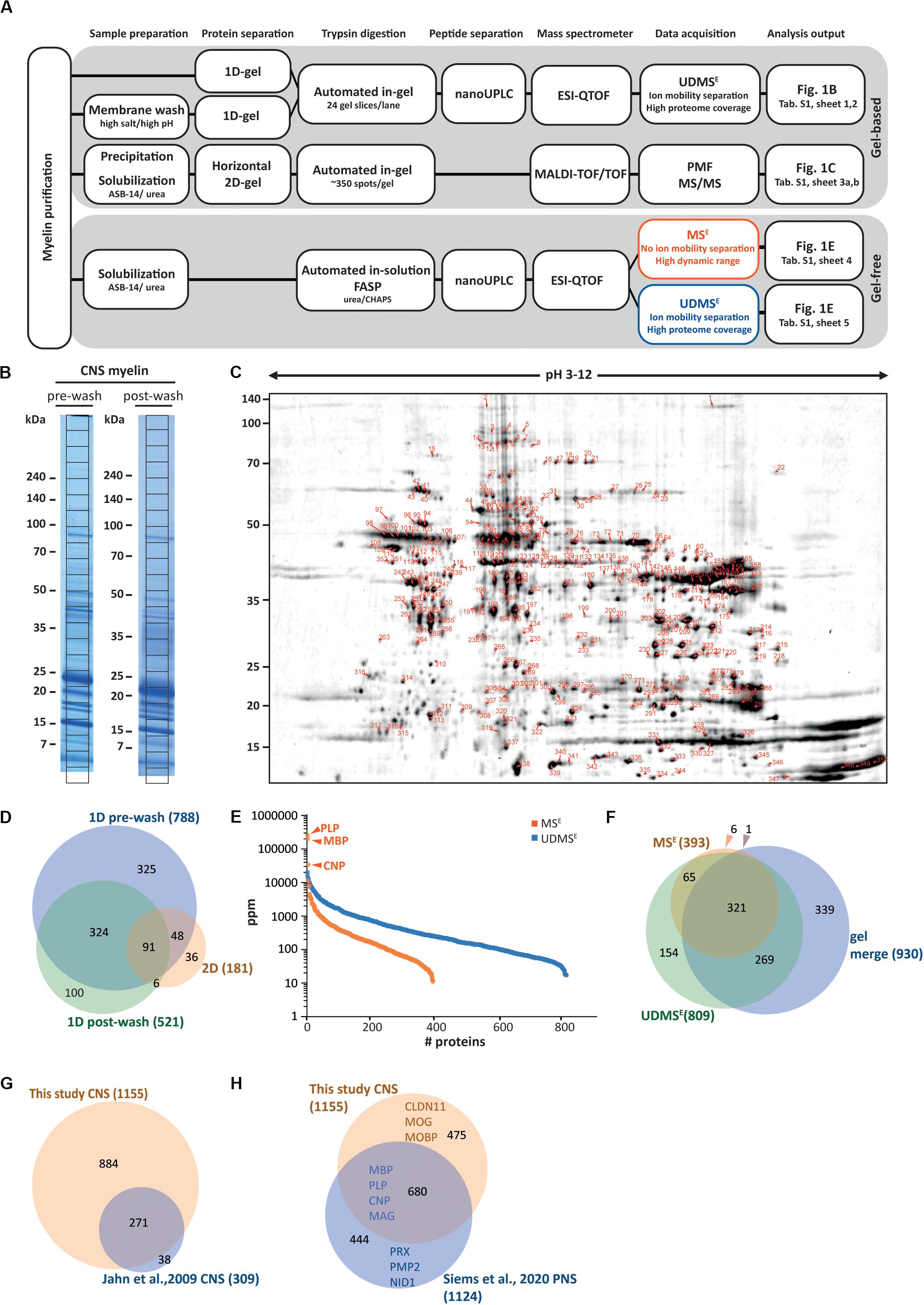
Figure 1. Proteome analysis of CNS myelin. (A) Schematic illustration of the gel-based (top) and gel-free (bottom) proteomic workflow to approach CNS myelin purified from the brains of wild-type c57Bl/6N mice dissected at P75. Note that gel-free proteome analysis enables largely automated sample processing and omits labor-intense gel-electrophoresis, thus reducing hands-on time. (B) One-dimensional gel-separation of CNS myelin. Myelin was separated by SDS-PAGE without (pre-wash) or upon (post-wash) depleting soluble and peripheral membrane proteins by an additional step of high-pH and high-salt conditions. Proteins were visualized with colloidal Coomassie (CBB250). The denoted grid subdivides each lane into 24 equally sized slices, which were excised for automated tryptic digest, peptide separation by nanoUPLC and data acquisition using an ESI-QTOF mass spectrometer, thereby identifying 788 (pre-wash) and 521 (post-wash) proteins, respectively (see Supplementary Table S1). (C) Two-dimensional gel-separation of CNS myelin. Myelin was two-dimensionally separated using a 2D-IEF/SDS-PAGE with isoelectric focusing (IEF) in a 24 cm gel strip with nonlinear pH-gradient (pH 3–12) as the first and 10–15% acrylamide gradient SDS-PAGE (25.5 × 20 cm, gel thickness 0.65 mm) as the second dimension. Proteins were visualized by colloidal Coomassie staining; protein spots were excised, subjected to automated tryptic in-gel digestion and MALDI-TOF mass spectrometry, thereby identifying 181 non-redundant proteins from 352 spots (Supplementary Table S1). (D) Venn diagram comparing the number of proteins identified in CNS myelin by the three gel-based approaches. (E) Number and relative abundance of proteins identified in myelin purified from the brains of wild-type mice using two gel-free data acquisition modes (MSE, UDMSE). Note that MSE (orange) identifies comparatively fewer proteins in purified myelin but provides a dynamic range of more than four orders of magnitude. UDMSE (blue) identifies a larger number of proteins but provides a dynamic range of only about three orders of magnitude. Note that the dynamic range of MSE is required for the quantification of the exceptionally abundant myelin proteins proteolipid protein (PLP), myelin basic protein (MBP) and cyclic nucleotide phosphodiesterase (CNP). Samples were analyzed in three biological replicates with four technical replicates each (duplicate digestion and injection). For datasets see Supplementary Table S1. ppm, parts per million. (F) Venn diagram comparing the number of proteins identified in CNS myelin by MSE, UDMSE and gel-based approaches. (G) Venn diagram of the proteins identified in CNS myelin in this study compared with those identified in a previous approach (Jahn et al., 2009). (H) Venn diagram comparing the proteins identified in CNS myelin in this study with those previously identified in PNS myelin (Siems et al., 2020). Selected marker proteins are denoted.
Considering that contemporary gel-free, label-free proteomic approaches allow the simultaneous identification and quantification of proteins (Neilson et al., 2011; Distler et al., 2014b) we subjected myelin to a workflow of solubilization using ASB-14 and high-urea conditions, automated tryptic in-solution digest by filter-aided sample preparation (FASP), fractionation of peptides by nanoUPLC, and ESI-QTOF mass spectrometry. This workflow was recently established for peripheral myelin (Siems et al., 2020). Importantly, the utilized data-independent acquisition (DIA)-strategy with data acquisition in the MSE-mode allows the simultaneous quantification and identification of all peptides entering the mass spectrometer, and thereby, when signal intensities are correlated with a spike protein of known concentration (TOP3 method; Silva et al., 2006; Ahrné et al., 2013) the reliable quantification of proteins based on peptide intensities. When subjecting myelin to LC-MS-analysis using MSE we quantified 393 proteins (Supplementary Table S1; labeled in orange in Figure 1E) with a false discovery rate (FDR) of <1% and an average sequence coverage of 38.6%. Notably, MSE quantitatively covered myelin proteins with a dynamic range of over four orders of magnitude parts per million (ppm), thereby allowing quantification of the exceptionally abundant PLP and MBP. When using the ultra-definition (UD)-MSE-mode, in which the ion mobility option provides an orthogonal dimension of peptide separation after liquid chromatography and before mass measurement, we identified and quantified 809 proteins (Supplementary Table S1; labeled in blue in Figure 1E) with an average sequence coverage of 35.0%. UDMSE thus identified about twice as many proteins as MSE. However, the larger number of proteins identified by UDMSE went along with a compressed dynamic range of about three orders of magnitude ppm, which is insufficient to reliably quantify the most abundant myelin constituents including PLP, MBP, and CNP. The data acquisition mode-dependent differences in both numbers of quantified proteins and dynamic range are best explained by UDMSE achieving more efficient precursor-fragment ion alignment and precursor fragmentation upon ion mobility separation of peptides (Distler et al., 2014a, 2016) which causes a ceiling effect for the detection of exceptionally intense peptide signals and thus a compressed dynamic range as previously observed for PNS myelin (Siems et al., 2020).
When comparing the proteins identified by MSE, UDMSE and gel-based approaches we found a reasonably high overlap (Figure 1F). Comparison of the 1155 proteins identified in CNS myelin in the present study with those 309 identified >10 years ago with the methodological standards of that time (Jahn et al., 2009) shows a remarkably high overlap as well as an about three-fold increase in the number of identified proteins (Figure 1G). Notwithstanding that a number of the identified proteins will originate from other cellular sources that contaminate purified myelin, we believe that many of them are indeed low-abundant constituents of the non-compact compartments of myelin.
A comparison of the proteins identified in CNS myelin with the recently established PNS myelin proteome (Siems et al., 2020) confirms that numerous proteins are present in both, but also that many proteins were identified exclusively in either CNS or PNS myelin (Figure 1H). Together, the evolving technical standards of in-solution sample preparation and MSE-type DIA mass spectrometry allows to comprehensively identify and quantify proteins in myelin. However, only MSE (but not UDMSE) provides a dynamic range suited to address the relative abundance of the exceptionally abundant PLP, MBP, and CNP. As importantly, the evolution of gel-free methods shifts the major workload in myelin proteome analysis from manual sample handling to data analysis, with much less hands-on time required when compared to gel-based approaches.
As MSE provides the best possible dynamic range (Figure 1E), we evaluated the relative abundance of all 393 proteins identified in myelin by MSE (Figure 2 and Supplementary Table S1). As per this dataset PLP constitutes 38% of the total myelin protein [±1% relative standard deviation (RSD)]. MBP, CNP, and MOG constitute 30% (±1%), 5% (±0.2%), and 1% (±0.03%) of the total myelin protein, respectively (Figure 2). However, the present assessment of CNS myelin by MSE extends well beyond the most abundant myelin constituents, thus quantifying known myelin proteins including the tetraspan-proteins CLDN11, CD81, TSPAN2, PLLP, CD9, CD82, GPM6B, and GJC3, the immunoglobulin-domain containing cell-surface proteins MAG, NFASC, CNTN1, RTN4, CNTN2, CADM4, HEPACAM, JAM3, CD47, and OMG, the enzymes SIRT2, CA2, and ASPA, the cytoskeletal and cytoskeleton-associated proteins TUBB4, SEPT2, SEPT4, SEPT7, SEPT8, TPPP, ANLN, GSN, CFL1, and PADI2 as well as MOBP, BCAS1, NDRG1, opalin, and CRYAB (Figure 2). By MSE, 46 known myelin proteins account for approximately 80% of the total myelin protein (Figure 2). The remaining 27% is constituted by 347 proteins not yet validated as myelin constituents by independent methods.
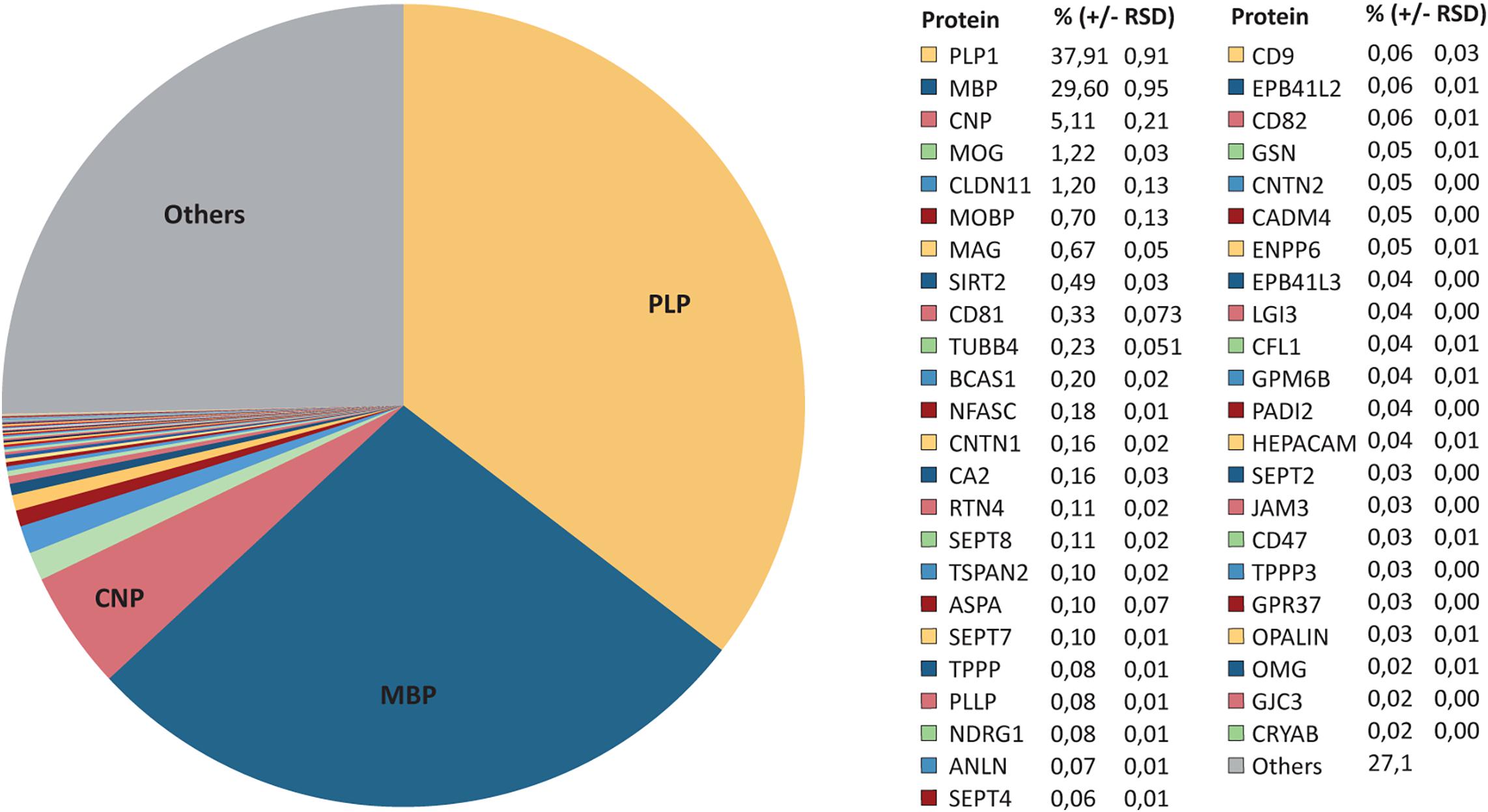
Figure 2. Relative abundance of CNS myelin proteins. Pie chart of the MSE dataset shown in Figure 1E and Supplementary Table S1. The relative abundance of known myelin proteins is given as percent with relative standard deviation (% ±RSD). Note that known myelin proteins constitute approximately 73% of the total myelin protein; proteins so far not independently validated as myelin proteins constitute about 27%.
An increasing number of studies provides mRNA or protein abundance profiles of myelin or oligodendrocytes. To systematically compare the CNS myelin proteome with these profiles, we correlated our MSE-dataset (Figure 2 and Supplementary Table S1) via the gene name entries with related datasets for which quantitative information is publicly available (Figure 3).
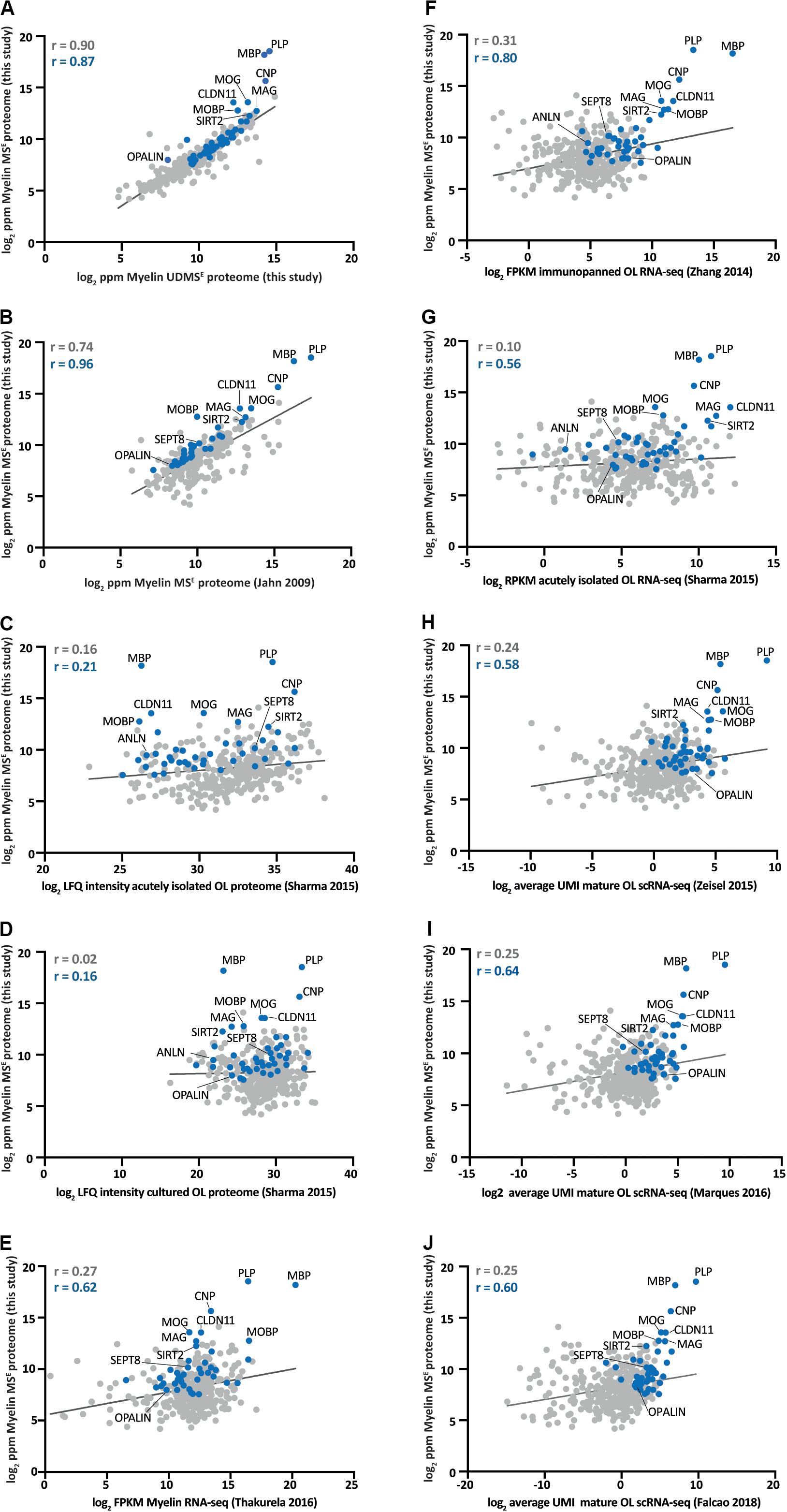
Figure 3. Comparison of the myelin proteome with proteome and transcriptome profiles of myelin and oligodendrocytes. (A) Log2-transformed relative abundance of the proteins identified in myelin in this study by MSE plotted against their log2-transformed relative abundance as quantified by UDMSE. Data points representing known myelin proteins as specified in Figure 2 are labeled in blue; all other data points in gray. The correlation coefficient (r) was calculated for all proteins identified by MSE (displayed in gray) and specifically for the known myelin proteins (given in blue). The regression line is plotted for orientation. ppm, parts per million. (B) Same as (A) but plotted against the myelin proteome as previously assessed by MSE (Jahn et al., 2009). (C) Same as (A) but plotted against the proteome of acutely isolated O4-immunopositive oligodendrocytes (Sharma et al., 2015). LFQ, label-free quantification. (D) Same as (A) but plotted against the proteome of O1-immunopositive primary oligodendrocytes cultured for 4 days in vitro (DIV) (Sharma et al., 2015). (E) Same as (A) but plotted against the RNA-seq-based transcriptome of myelin purified from the brains of mice (Thakurela et al., 2016). FPKM, fragments per kilobase of exon model per million reads mapped. (F) Same as (A) but plotted against the RNA-seq-based transcriptome of oligodendrocytes immunopanned using MOG-specific antibodies (Zhang et al., 2014). (G) Same as (A) but plotted against the RNA-seq-based transcriptome of acutely isolated O4-immunopositive oligodendrocytes (Sharma et al., 2015). RPKM, reads per kilobase per million mapped reads. (H) Same as (A) but plotted against the scRNA-seq-based transcriptome of mature oligodendrocytes in the mouse cortex and hippocampus [mean of all 484 cells in clusters Oligo5 and Oligo6 in Zeisel et al. (2015)]. UMI, unique molecular identifiers. (I) Same as (A) but plotted against the scRNA-seq-based transcriptome of mature oligodendrocytes in 10 regions of the mouse CNS [mean of all 2748 cells in clusters OL1 – OL6 in Marques et al. (2016)]. (J) Same as (A) but plotted against the scRNA-seq-based transcriptome of mature oligodendrocytes in the mouse spinal cord [mean of all 617 cells in clusters MOL2-Ct and MOL5/6-Ct in Falcão et al. (2018)].
We first plotted the present MSE and UDMSE-datasets (Supplementary Table S1) against each other (Figure 3A). Considering that the same starting material has been assessed it is not unexpected that the datasets correlate well, as reflected by a correlation coefficient of 0.90 (Figure 3A). Most visibly diverging from the linear regression line are the most abundant myelin proteins PLP, MBP, CNP, MOG, and CLDN11, reflecting that the dynamic range of UDMSE is compressed in the high ppm-range compared to that of MSE (also see Figure 1E). We then compared the present MSE-dataset with an independent myelin proteome dataset previously established by MSE (Jahn et al., 2009). We calculated a somewhat lower correlation coefficient of 0.74 (Figure 3B), probably owing to the previous use of a predecessor mass spectrometer generation that provided a considerably lower dynamic range. Yet, in conjunction with the high overlap between the proteins identified in the present and the previous study (Jahn et al., 2009) (Figure 1G), myelin proteome analysis emerges as fairly robust across independently purified starting material and different generations of mass spectrometers. We next compared the MSE-dataset to the proteome of acutely isolated O4-immunopositive oligodendrocytes (Figure 3C) as determined by label-free quantification (LFQ) using data-dependent acquisition (DDA) on an orbitrap mass spectrometer and MaxQuant-software (Sharma et al., 2015). The O4-antibody preferentially immunolabels oligodendrocytes at the progenitor (OPC) and pre-myelinating stages (Sommer and Schachner, 1981; Bansal et al., 1989; Goldman and Kuypers, 2015); the correlation coefficient was calculated as 0.16 (Figure 3C). A correlation coefficient of 0.02 (Figure 3D) was found when comparing the MSE-dataset with the LFQ-intensity profile of O1-immunopositive primary oligodendrocytes after 4 days in vitro (DIV) (Sharma et al., 2015). The myelin proteome as determined here is thus more closely related to the proteome of acutely isolated O4-immunopositive oligodendrocytes than to that of O1-immunopositive primary oligodendrocytes 4 DIV.
We then compared the MSE-dataset with various available mRNA-abundance profiles. When comparing the MSE-dataset to the transcriptome of purified CNS myelin as determined by RNA-seq (Thakurela et al., 2016) we calculated a correlation coefficient of 0.27 (Figure 3E). Interestingly, the comparison between the MSE-dataset and the RNA-seq-based transcriptome of oligodendrocytes immunopanned from the cortex using antibodies against MOG (Zhang et al., 2014) revealed a roughly comparable correlation coefficient of 0.31 (Figure 3F). Notably, MOG-immunopositivity labels myelinating oligodendrocytes, implying that the stage of oligodendrocyte differentiation must be considered when judging dataset correlations. It is thus not surprising that a somewhat lower correlation coefficient of 0.10 (Figure 3G) was calculated when comparing the MSE-dataset with the RNA-seq-based transcriptome of acutely isolated O4+- oligodendrocytes (Sharma et al., 2015). Finally, we compared the MSE-dataset to several scRNA-seq-based transcriptome datasets (Zeisel et al., 2015; Marques et al., 2016; Falcão et al., 2018). To this aim we calculated the mean transcript abundance as average count reads per unique molecular identifier (UMI) of the cells in those clusters that reflect mature oligodendrocytes. When comparing the MSE-dataset to mature oligodendrocytes sorted from the mouse cortex and hippocampus [all 484 cells in clusters Oligo5 and Oligo6 in Zeisel et al. (2015)], we find a correlation coefficient of 0.24 (Figure 3H). Importantly, we find a roughly similar correlation coefficient when comparing the MSE-dataset to mature oligodendrocytes sorted from 10 regions of the mouse CNS [all 2748 cells in clusters MOL1–MOL6 in Marques et al. (2016)] (Figure 3I) or to mature oligodendrocytes sorted from the spinal cord of mice [all 617 cells in clusters MOL2-Ct and MOL5/6-Ct in Falcão et al. (2018)] (Figure 3J).
Together, when judging correlations between large datasets evaluating mRNA and protein abundance profiles of oligodendrocytes and myelin, aspects to be considered include the method of sample preparation, the stage of oligodendrocyte differentiation and the methodology of analysis. Yet, roughly similar correlation coefficients were calculated when comparing the myelin proteome with various proteomic and transcriptomic approaches to the molecular profiles of oligodendrocytes.
Autopsy material from human patients and healthy-appearing controls is increasingly evaluated by systematic molecular profiling, as exemplified by the recent snRNA-seq-based assessment of oligodendroglial transcriptional profiles in multiple sclerosis patients (Jäkel et al., 2019). Notably, the use of autopsy material involves a post-mortem delay between the death of a subject and the collection of a biopsy. However, post-mortem delay may affect sample integrity and thus data validity. Considering that proteomic analysis of myelin in mice is usually performed upon freezing of samples immediately after dissection, we asked whether the myelin proteome can also be assessed upon post-mortem delay. We thus purified myelin from the brains of c57Bl/6N-mice to compare the myelin proteome between mice after a post-mortem delay of 6 h at room temperature with that of mice upon sample freezing immediately after dissection. Upon SDS-PAGE-separation and silver staining, no signs of major degradation were evident and the band patterns appeared essentially similar (Figure 4A). We next subjected myelin to routine differential proteome profiling by UDMSE with dynamic range enhancement (DRE-UDMSE) (Supplementary Table S2). Using this data acquisition mode with intermediate features as to identification rates and dynamic range (for methodological details see Siems et al., 2020) we found that known myelin proteins displayed only minor differences as visualized in a volcano plot (red data points in Figure 4B) and a heatmap (Figure 4C). Indeed, no known myelin protein exceeded the threshold of a log2-fold transformed fold-change (FC) of −1/+1, i.e., a 2-fold increased or 0.5-fold diminished relative abundance. Together, the myelin proteome displays only minor changes upon a post-mortem delay of 6 h, implying that proteomic assessment of myelin purified from autopsy samples appears feasible.
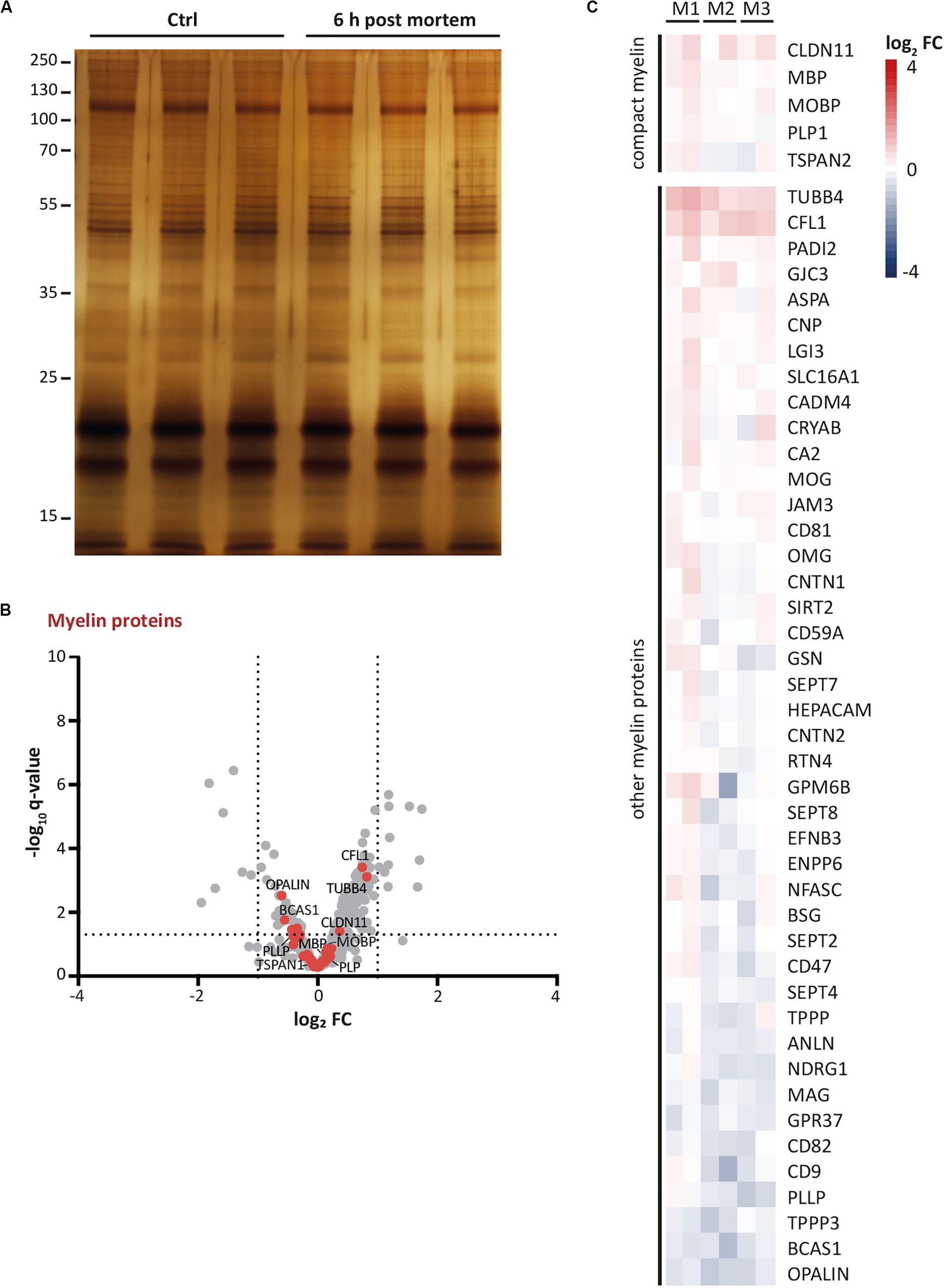
Figure 4. Persistence of myelin proteins upon post-mortem delay. (A) Myelin purified from the brains of mice at P56 was separated by SDS-PAGE (0.5 μg protein load) and proteins were visualized by silver staining. Myelin of brains frozen upon a post-mortem delay of 6 h at room temperature was compared with myelin of brains frozen immediately upon dissection (Ctrl). Note the similar band pattern. Gel shows n = 3 biological replicates per condition. (B) Volcano plot representing differential proteome analysis by DRE-UDMSE to compare myelin purified from brains upon post-mortem delay with myelin of brains immediately frozen upon dissection. For entire dataset see Supplementary Table S1. Data points represent proteins quantified in myelin purified from mouse brains frozen after a post-mortem delay of 6 h at room temperature compared to immediately frozen brains and are plotted as the log2-transformed fold-change (FC) on the x-axis against the –log10-transformed q-value on the y-axis. Vertical stippled lines mark a 2-fold/0.5-fold change (FC) as significance threshold. Horizontal stippled line represents a –log10-transformed q-value of 1.301, reflecting a q-value of 0.05 as significance threshold. Data points highlighted in red represent known myelin proteins as specified in Figure 4C. Note that no known myelin protein exceeds the fold-change significance threshold. (C) Heatmap displaying known myelin proteins as highlighted by the red data points in Figure 4B. Heatmap shows reduced (blue) or increased (red) abundance in myelin purified from brains after post-mortem delay. Each horizontal line corresponds to the fold-change (FC) of a distinct myelin protein compared to its average abundance in control myelin plotted on a log2 color scale. Heatmap displays 6 replicates, i.e., three biological replicates per condition (M1, M2, M3) with two technical replicates each.
Understanding the molecular complexity of the nervous system involves molecular profiling of cells and cellular specializations including myelin. Here we combined various proteomic approaches for comprehensive coverage of the CNS myelin proteome and identified 1155 proteins in myelin biochemically purified from the brains of mice. We note that gel-based methods involving separation at the protein level facilitated a slightly higher identification rate compared to gel-free methods comprising in situ-digestion of the entire proteome, likely because of the pre-fractionation effect inherent to the former. On the other hand, gel-free data acquisition by UDMSE also enabled deep qualitative coverage while necessitating considerably less input material and manual sample handling.
Importantly, the MSE-data acquisition mode covered a dynamic range of over four orders of magnitude of protein abundance. Indeed, compared to a previous approach (Jahn et al., 2009) the technical advancements implemented in the current mass spectrometer generation now allow reliable quantification of myelin proteins spanning from the exceptionally abundant PLP and MBP to low-abundant constituents including oligodendrocyte myelin glycoprotein (OMG) (Wang et al., 2002), oligodendrocytic myelin paranodal and inner loop protein (OPALIN) (Golan et al., 2008; Kippert et al., 2008) and the G-protein coupled receptor GPR37 (Yang et al., 2016). For PLP, MBP and CNP our quantification is in accordance with but specifies prior estimates based on 1D-gel separation and various protein staining techniques, in which they were proposed to constitute 30–45%, 22–35%, and 4–15% of the total myelin protein, respectively (Morell et al., 1972, 1973; Banik and Smith, 1977; Deber and Reynolds, 1991). Notably, it also shifts our previous MSE-based estimates for PLP and MBP (Jahn et al., 2009) toward higher relative abundance, with the lower dynamic range of the mass spectrometers at that time being the most likely reason for the former under-quantification. It is not surprising that PLP, MBP, and CNP have overshadowed less abundant myelin constituents in initial gel-based approaches when considering the exceptional dynamic range of the relative abundance of myelin proteins. Together, the myelin proteome provided here provides an updated comprehensive compendium and re-adjusts the relative abundance of CNS myelin proteins.
Do true myelin proteins exist that escape proteomic identification? As exemplified by myelin and lymphocyte protein (MAL) (Schaeren-Wiemers et al., 2004), the tryptic digest of some myelin proteins may result in peptides incompatible with mass spectrometric detection; their identification would require the use of proteases other than trypsin. We also note that some low-abundant signaling proteins with potent functions in regulating myelination may be assumed to localize to myelin in vivo but were not mass spectrometrically identified, as exemplified by the G-protein coupled receptors GPR17 (Chen et al., 2009) and GPR56/ADGRG1 (Ackerman et al., 2015; Giera et al., 2015) and the Ig-domain containing LINGO1 (Mi et al., 2005). It is currently speculative if these proteins are preferentially expressed in oligodendroglial cell bodies rather than myelin membranes or during the stages of oligodendrocyte differentiation that precede myelination. It is also speculative if enhanced mass spectrometric sensitivity would facilitate their identification in myelin. Indeed, we can not formally exclude that these proteins may be identified if less rigorous criteria were applied (e.g., demanding only one peptide per protein), which may be sufficient for identification but not for the reliable quantification of proteins as aimed at in the present study. Importantly, however, lower stringency may not only identify more true myelin constituents but also false-positive hits. This is a concern, in particular when considering that the myelin-enriched fraction may comprise up to 5% contaminants from other cellular sources (De Monasterio-Schrader et al., 2012). We note that currently no biochemical method is available that allows preventing this limitation. Yet, comparing various datasets yields systematic information, for example on the presence of a transcript in oligodendrocytes as expected for a CNS myelin protein.
Mutations affecting genes that encode classical myelin proteins including PLP, CNP, MAG, TUBB4, and ASPA cause severe neurological disorders including hypomyelinating leukodystrophies (HLD) and spastic paraplegias (SPG) (Kaul et al., 1993; Saugier-Veber et al., 1994; Simons et al., 2013; Lossos et al., 2015; Al-Abdi et al., 2020) (Table 1). However, current sequencing efforts also identify disease-causing genes that encode less well-characterized proteins. Notably, most types of leukodystrophies and spastic paraplegias are caused by mutations affecting genes of which the transcripts are enriched in neurons, astrocytes or microglia rather than oligodendrocytes (Nave and Werner, 2014; van der Knaap et al., 2019). For newly identified disease genes, thus, evaluating mRNA-expression using transcriptome datasets and presence of the protein in myelin using the present myelin proteome resource may serve as a useful entry point into identifying the primarily affected cell type. For example, mutations of the HSPD1 gene cause HLD4 or SPG13 (Hansen et al., 2002; Magen et al., 2008) and mutations of the TMEM63A gene cause HLD19 (Yan et al., 2019) (Table 1). Considering that both transcripts are expressed in oligodendrocytes as per transcriptome datasets and both proteins are comprised in the myelin proteome, the disease mechanisms may involve primary impairment of the biogenesis, maintenance or functions of myelin.
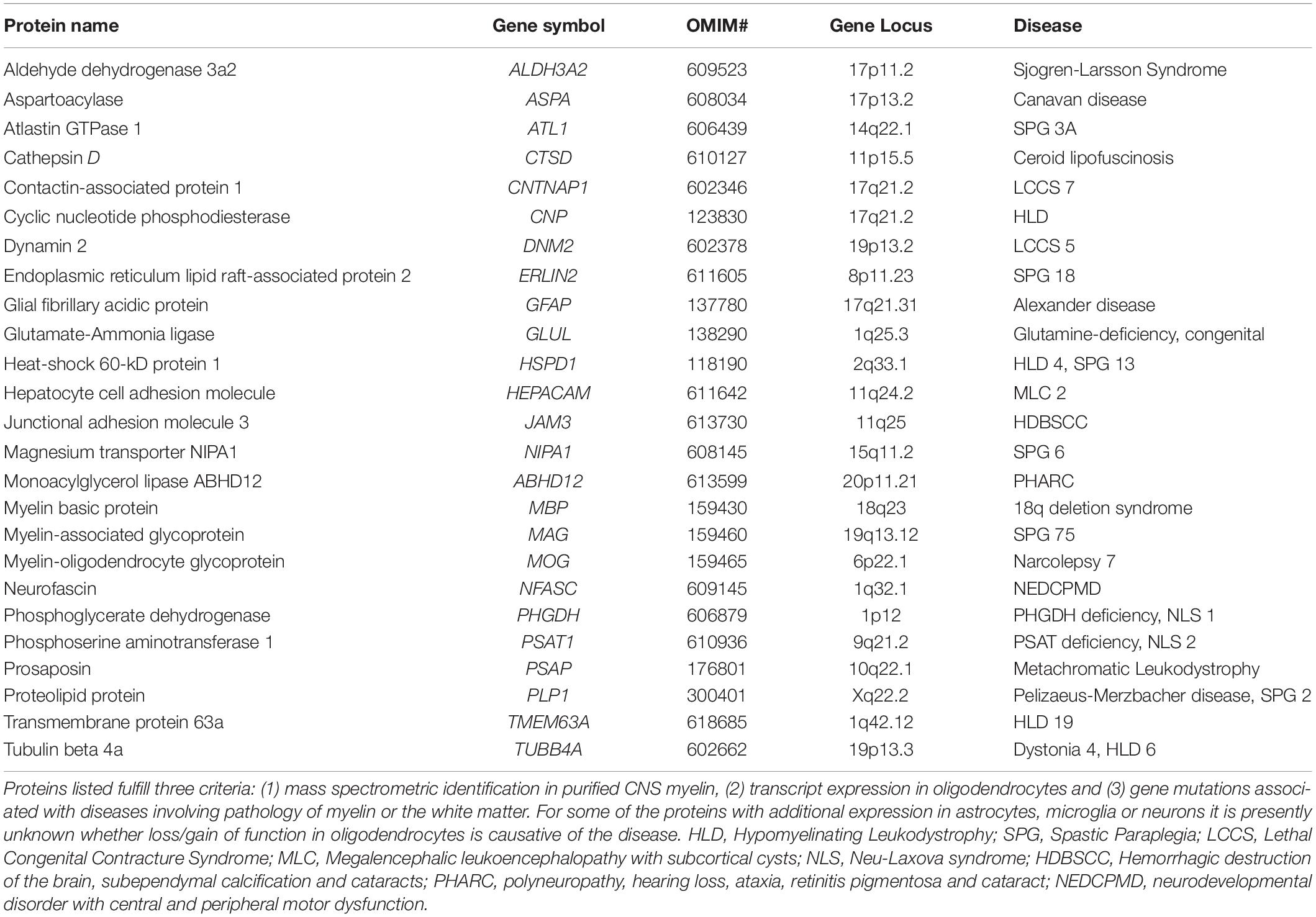
Table 1. Comparison of proteins identified in CNS myelin and disease genes associated with white matter pathology.
Dysfunctions of oligodendrocytes and myelin contribute to the neuropathology in a growing number of neurodegenerative disorders and their respective mouse models, including Rett syndrome (Nguyen et al., 2013), amyotrophic lateral sclerosis (Kang et al., 2013), Down syndrome (Olmos-Serrano et al., 2016), Alzheimer’s disease (Nasrabady et al., 2018) and multiple sclerosis (Factor et al., 2020). Considering that molecular assessments now frequently involve autopsy material, it is motivating that our data imply that myelin proteome analysis appears well possible post-mortem, at least up to a 6 h delay. A systematic understanding of the abundance profiles of all myelin proteins in the healthy brain and in myelin-related disorders may contribute to comprehending myelin-related physiology and pathophysiology. Myelin proteome analysis as pursued here provides a basis for addressing possible proteomic heterogeneity of myelin in dependence of CNS region, age and species, as well as in mouse models and human patients with white matter disorders.
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Perez-Riverol et al., 2019) partner repository with dataset identifier, PXD020007.
Ethical review and approval was not required for the animal study because for the procedure of sacrificing mice for subsequent preparation of tissue, all regulations given in the German animal protection law (TierSchG §4) are followed. Since sacrificing of rodents is not an experiment on animals according to §7 Abs. 2 Satz 3 TierSchG, no specific authorization or notification is required for the present work.
KK, DH, RJ, TL, and MU performed the experiments. SS analyzed the data and performed the statistical analysis. TS wrote the code to interpret single cell resolution transcriptome data. OJ and HW conceived, designed, and directed the study. HW wrote the manuscript with major contributions by SS and OJ. All the authors contributed to revising the manuscript and approved the submitted version.
Our work was supported by the Deutsche Forschungsgemeinschaft (DFG; Grants WE 2720/2-2, WE 2720/4-1, and WE 2720/5-1 to HW).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank G. Castelo-Branco and D. van Bruggen for providing dataset metadata information, S. Tenzer, M. Eichel, and T. Buscham for discussions, L. Piepkorn for support in data analysis, K.-A. Nave for support made possible by a European Research Council Advanced Grant (‘MyeliNano’ to K.-A. Nave), and the International Max Planck Research School for Genome Science (IMPRS-GS) for supporting SS.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fncel.2020.00239/full#supplementary-material
TABLE S1 | Compendium of the mouse CNS myelin proteome. (Sheet 1) 1D gel separation of proteins followed by in-gel digestion and LC-MS analysis (788 proteins). (Sheet 2) 1D gel separation of proteins after membrane wash followed by in-gel digestion and LC-MS analysis (521 proteins). (Sheet 3a) 2D gel separation of proteins followed by in-gel digestion and MALDI-TOF-MS: spot annotation of the master gel shown in Figure 3C (352 gel spots). (Sheet 3b) 2D gel separation of proteins followed by in-gel digestion and MALDI-TOF-MS: non-redundant protein list (181 proteins). (Sheet 4) In-solution digestion of proteins and label-free quantification by MSE (393 proteins). (Sheet 5) In-solution digestion of proteins and label-free quantification by UDMSE (809 proteins). (Sheet 6) Compendium providing information on the approach by which a myelin-associated protein was identified and on predicted transmembrane domains (1155 proteins).
TABLE S2 | Label-free quantification of proteins in CNS myelin purified from mouse brains without (Ctrl) and after 6 h post-mortem delay (PMD6h) by DRE-UDMSE. Related to Figure 4. (Sheet 1) Dataset.
Ackerman, S. D., Garcia, C., Piao, X., Gutmann, D. H., and Monk, K. R. (2015). The adhesion GPCR Gpr56 regulates oligodendrocyte development via interactions with Gα12/13 and RhoA. Nat. Commun. 6:6122. doi: 10.1038/ncomms7122
Ahrné, E., Molzahn, L., Glatter, T., and Schmidt, A. (2013). Critical assessment of proteome-wide label-free absolute abundance estimation strategies. Proteomics 13, 2567–2578. doi: 10.1002/pmic.201300135
Al-Abdi, L., Al Murshedi, F., Elmanzalawy, A., Al Habsi, A., Helaby, R., Ganesh, A., et al. (2020). CNP deficiency causes severe hypomyelinating leukodystrophy in humans. Hum. Genet. 139, 615–622. doi: 10.1007/s00439-020-02144-4
Ambrozkiewicz, M. C., Schwark, M., Kishimoto-Suga, M., Borisova, E., Hori, K., Salazar-Lázaro, A., et al. (2018). Polarity Acquisition in Cortical Neurons Is Driven by Synergistic Action of Sox9-Regulated Wwp1 and Wwp2 E3 Ubiquitin Ligases and Intronic miR-140. Neuron 100:1097-1115.e15. doi: 10.1016/j.neuron.2018.10.008
Banik, N. L., and Smith, M. E. (1977). Protein determinants of myelination in different regions of developing rat central nervous system. Biochem. J. 162, 247–255. doi: 10.1042/bj1620247
Bansal, R., Warrington, A. E., Gard, A. L., Ranscht, B., and Pfeiffer, S. E. (1989). Multiple and novel specificities of monoclonal antibodies O1, O4, and R-mAb used in the analysis of oligodendrocyte development. J. Neurosci. Res. 24, 548–557. doi: 10.1002/jnr.490240413
Butler, A., Hoffman, P., Smibert, P., Papalexi, E., and Satija, R. (2018). Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420. doi: 10.1038/nbt.4096
Chen, Y., Wu, H., Wang, S., Koito, H., Li, J., Ye, F., et al. (2009). The oligodendrocyte-specific G protein-coupled receptor GPR17 is a cell-intrinsic timer of myelination. Nat. Neurosci. 12, 1398–1406. doi: 10.1038/nn.2410
Chrast, R., Saher, G., Nave, K. A., and Verheijen, M. H. G. (2011). Lipid metabolism in myelinating glial cells: Lessons from human inherited disorders and mouse models. J. Lipid Res. 52, 419–434. doi: 10.1194/jlr.R009761
De Monasterio-Schrader, P., Jahn, O., Tenzer, S., Wichert, S. P., Patzig, J., and Werner, H. B. (2012). Systematic approaches to central nervous system myelin. Cell. Mol. Life Sci. 69, 2879–2894. doi: 10.1007/s00018-012-0958-9
de Monasterio-Schrader, P., Patzig, J., Möbius, W., Barrette, B., Wagner, T. L., Kusch, K., et al. (2013). Uncoupling of neuroinflammation from axonal degeneration in mice lacking the myelin protein tetraspanin-2. Glia 61, 1832–1847. doi: 10.1002/glia.22561
Deber, C. M., and Reynolds, S. J. (1991). Central nervous system myelin: structure, function, and pathology. Clin. Biochem. 24, 113–134. doi: 10.1016/0009-9120(91)90421-A
Denninger, A. R., Breglio, A., Maheras, K. J., Leduc, G., Cristiglio, V., Demé, B., et al. (2015). Claudin-11 tight junctions in myelin are a barrier to diffusion and lack strong adhesive properties. Biophys. J. 109, 1387–1397. doi: 10.1016/j.bpj.2015.08.012
Dhaunchak, A. S., Huang, J. K., De Faria, O., Roth, A. D., Pedraza, L., Antel, J. P., et al. (2010). A proteome map of axoglial specializations isolated and purified from human central nervous system. Glia 58, 1949–1960. doi: 10.1002/glia.21064
Distler, U., Kuharev, J., Navarro, P., Levin, Y., Schild, H., and Tenzer, S. (2014a). Drift time-specific collision energies enable deep-coverage data-independent acquisition proteomics. Nat. Methods 11, 167–170. doi: 10.1038/nmeth.2767
Distler, U., Kuharev, J., and Tenzer, S. (2014b). Biomedical applications of ion mobility-enhanced data-independent acquisition-based label-free quantitative proteomics. Expert Rev. Proteomics 11, 675–684. doi: 10.1586/14789450.2014.971114
Distler, U., Kuharev, J., Navarro, P., and Tenzer, S. (2016). Label-free quantification in ion mobility–enhanced data-independent acquisition proteomics. Nat. Protoc. 11, 795–812. doi: 10.1038/nprot.2016.042
Edgar, J. M., McLaughlin, M., Werner, H. B., McCulloch, M. C., Barrie, J. A., Brown, A., et al. (2009). Early ultrastructural defects of axons and axon-glia junctions in mice lacking expression of Cnp1. Glia 57, 1815–1824. doi: 10.1002/glia.20893
Erwig, M. S., Hesse, D., Jung, R. B., Uecker, M., Kusch, K., Tenzer, S., et al. (2019a). ‘Myelin: Methods for Purification and Proteome Analysis’. Methods Mol. Biol. 1936, 37–63. doi: 10.1007/978-1-4939-9072-6_3
Erwig, M. S., Patzig, J., Steyer, A. M., Dibaj, P., Heilmann, M., Heilmann, I., et al. (2019b). Anillin facilitates septin assembly to prevent pathological outfoldings of central nervous system myelin. eLife 8:e43888. doi: 10.7554/eLife.43888
Factor, D. C., Barbeau, A. M., Allan, K. C., Hu, L. R., Madhavan, M., Hoang, A. T., et al. (2020). Cell type-specific intralocus interactions reveal oligodendrocyte mechanisms in MS. Cell 181:382-395.e21. doi: 10.1016/j.cell.2020.03.002
Falcão, A. M., van Bruggen, D., Marques, S., Meijer, M., Jäkel, S., Agirre, E., et al. (2018). Disease-specific oligodendrocyte lineage cells arise in multiple sclerosis. Nat. Med. 24, 1837–1844. doi: 10.1038/s41591-018-0236-y
Giera, S., Deng, Y., Luo, R., Ackerman, S. D., Mogha, A., Monk, K. R., et al. (2015). The adhesion G protein-coupled receptor GPR56 is a cell-autonomous regulator of oligodendrocyte development. Nat. Commun. 6:6121. doi: 10.1038/ncomms7121
Golan, N., Adamsky, K., Kartvelishvily, E., Brockschnieder, D., Möbius, W., Spiegel, I., et al. (2008). Identification of Tmem10/Opalin as an oligodendrocyte enriched gene using expression profiling combined with genetic cell ablation. Glia 56, 1176–1186. doi: 10.1002/glia.20688
Goldman, S. A., and Kuypers, N. J. (2015). How to make an oligodendrocyte. Development 142, 3983–3995. doi: 10.1242/dev.126409
Gopalakrishnan, G., Awasthi, A., Belkaid, W., De Faria, O., Liazoghli, D., Colman, D. R., et al. (2013). Lipidome and proteome map of myelin membranes. J. Neurosci. Res. 91, 321–334. doi: 10.1002/jnr.23157
Gow, A., Southwood, C. M., Li, J. S., Pariali, M., Riordan, G. P., Brodie, S. E., et al. (1999). CNS Myelin and sertoli cell tight junction strands are absent in OSP/claudin-11 null mice. Cell 99, 649–659. doi: 10.1016/S0092-8674(00)81553-6
Hansen, J. J., Dürr, A., Cournu-Rebeix, I., Georgopoulos, C., Ang, D., Nielsen, M. N., et al. (2002). Hereditary spastic paraplegia SPG13 is associated with a mutation in the gene encoding the mitochondrial chaperonin Hsp60. Am. J. Hum. Genet. 70, 1328–1332. doi: 10.1086/339935
Hulsen, T., de Vlieg, J., and Alkema, W. (2008). BioVenn – a web application for the comparison and visualization of biological lists using area-proportional Venn diagrams. BMC Genomics 9:488. doi: 10.1186/1471-2164-9-488
Ishii, A., Dutta, R., Wark, G. M., Hwang, S. II, Han, D. K., Trapp, B. D., et al. (2009). Human myelin proteome and comparative analysis with mouse myelin. Proc. Natl. Acad. Sci. U.S.A. 106, 14605–14610. doi: 10.1073/pnas.0905936106
Jahn, O., Hesse, D., Reinelt, M., and Kratzin, H. D. (2006). Technical innovations for the automated identification of gel-separated proteins by MALDI-TOF mass spectrometry. Anal. Bioanal. Chem. 386, 92–103. doi: 10.1007/s00216-006-0592-1
Jahn, O., Tenzer, S., Bartsch, N., Patzig, J., and Werner, H. B. (2013). “Myelin proteome analysis: methods and implications for the myelin cytoskeleton,” in The Cytoskeleton. Neuromethods, Vol. 79, ed. R. Dermietzel, (Totowa, NJ: Humana Press), doi: 10.1007/978-1-62703-266-7_15
Jahn, O., Tenzer, S., and Werner, H. B. (2009). Myelin proteomics: Molecular anatomy of an insulating sheath. Mol. Neurobiol. 40, 55–72. doi: 10.1007/s12035-009-8071-2
Jäkel, S., Agirre, E., Mendanha Falcão, A., van Bruggen, D., Lee, K. W., Knuesel, I., et al. (2019). Altered human oligodendrocyte heterogeneity in multiple sclerosis. Nature 566, 543–547. doi: 10.1038/s41586-019-0903-2
Johns, T. G., and Bernard, C. C. A. (1999). The structure and function of myelin oligodendrocyte glycoprotein. J. Neurochem. 72, 1–9. doi: 10.1046/j.1471-4159.1999.0720001.x
Joseph, S., Werner, H., and Stegmüller, J. (2019). Gallyas silver impregnation of myelinated nerve fibers. Bio-Protocol 9:e3436. doi: 10.21769/bioprotoc.3436
Käll, L., Krogh, A., and Sonnhammer, E. L. L. (2007). Advantages of combined transmembrane topology and signal peptide prediction-the Phobius web server. Nucleic Acids Res. 35, W429–W432. doi: 10.1093/nar/gkm256
Kang, S. H., Li, Y., Fukaya, M., Lorenzini, I., Cleveland, D. W., Ostrow, L. W., et al. (2013). Degeneration and impaired regeneration of gray matter oligodendrocytes in amyotrophic lateral sclerosis. Nat. Neurosci. 16, 571–579. doi: 10.1038/nn.3357
Kaul, R., Gao, G. P., Balamurugan, K., and Matalon, R. (1993). Cloning of the human aspartoacylase cDNA and a common missense mutation in Canavan disease. Nat. Genet. 5, 118–123. doi: 10.1038/ng1093-118
Kippert, A., Trajkovic, K., Fitzner, D., Opitz, L., and Simons, M. (2008). Identification of tmem10/opalin as a novel marker for oligodendrocytes using gene expression profiling. BMC Neurosci. 9:40. doi: 10.1186/1471-2202-9-40
Krogh, A., Larsson, B., Von Heijne, G., and Sonnhammer, E. L. L. (2001). Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 305, 567–580. doi: 10.1006/jmbi.2000.4315
Kuharev, J., Navarro, P., Distler, U., Jahn, O., and Tenzer, S. (2015). In-depth evaluation of software tools for data-independent acquisition based label-free quantification. Proteomics 15, 3140–3151. doi: 10.1002/pmic.201400396
Lappe-Siefke, C., Goebbels, S., Gravel, M., Nicksch, E., Lee, J., Braun, P. E., et al. (2003). Disruption of Cnp1 uncouples oligodendroglial functions in axonal support and myelination. Nat. Genet. 33, 366–374. doi: 10.1038/ng1095
Lee, A. G. (2001). Myelin: delivery by raft. Curr. Biol. 11, R60–R62. doi: 10.1016/S0960-9822(01)00008-2
Lossos, A., Ponger, P., Newman, J. P., Elazar, N., Mor, N., Eshed-Eisenbach, Y., et al. (2015). Myelin-associated glycoprotein gene mutation causes Pelizaeus-Merzbacher disease-like disorder. Brain 138, 2521–2536. doi: 10.1093/brain/awv204
Magen, D., Georgopoulos, C., Bross, P., Ang, D., Segev, Y., Goldsher, D., et al. (2008). Mitochondrial Hsp60 chaperonopathy causes an autosomal-recessive neurodegenerative disorder linked to brain hypomyelination and leukodystrophy. Am. J. Hum. Genet. 83, 30–42. doi: 10.1016/j.ajhg.2008.05.016
Magno-Sumbilla, C., and Campagnoni, A. T. (1977). Factors affecting the electrophoretic analysis of myelin proteins: Application to changes occurring during brain development. Brain Res. 126, 131–148. doi: 10.1016/0006-8993(77)90220-7
Manrique-Hoyos, N., Jürgens, T., Grønborg, M., Kreutzfeldt, M., Schedensack, M., Kuhlmann, T., et al. (2012). Late motor decline after accomplished remyelination: Impact for progressive multiple sclerosis. Ann. Neurol. 71, 227–244. doi: 10.1002/ana.22681
Marques, S., Zeisel, A., Codeluppi, S., van Bruggen, D., Mendanha Falcao, A., Xiao, L., et al. (2016). Oligodendrocyte heterogeneity in the mouse juvenile and adult central nervous system. Science 352, 1326–1329. doi: 10.1126/science.aaf6463
Mi, S., Miller, R. H., Lee, X., Scott, M. L., Shulag-Morskaya, S., Shao, Z., et al. (2005). LINGO-1 negatively regulates myelination by oligodendrocytes. Nat. Neurosci. 8, 745–751. doi: 10.1038/nn1460
Möbius, W., Patzig, J., Nave, K. A., and Werner, H. B. (2008). Phylogeny of proteolipid proteins: Divergence, constraints, and the evolution of novel functions in myelination and neuroprotection. Neuron Glia Biol. 4, 111–127. doi: 10.1017/S1740925X0900009X
Morell, P., Greenfield, S., Costantino-Ceccarini, E., and Wisniewski, H. (1972). Changes in the protein composition of mouse brain myelin during development. J. Neurochem. 19, 2545–2554. doi: 10.1111/j.1471-4159.1972.tb01313.x
Morell, P., Lipkind, R., and Greenfield, S. (1973). Protein composition of myelin from brain and spinal cord of several species. Brain Res. 58, 510–514. doi: 10.1016/0006-8993(73)90023-1
Morris, S. J., Louis, C. F., and Shooter, E. M. (1971). Separation of myelin proteins on two different polyacrylamide gel systems. Neurobiology 1, 64–67.
Musse, A. A., Gao, W., Homchaudhuri, L., Boggs, J. M., and Harauz, G. (2008). Myelin basic protein as a ‘PI(4,5)P2-modulin’: A new biological function for a major central nervous system protein. Biochemistry 47, 10372–10382. doi: 10.1021/bi801302b
Myllykoski, M., Eichel, M. A., Jung, R. B., Kelm, S., Werner, H. B., and Kursula, P. (2018). High-affinity heterotetramer formation between the large myelin-associated glycoprotein and the dynein light chain DYNLL1. J. Neurochem. 147, 764–783. doi: 10.1111/jnc.14598
Nasrabady, S. E., Rizvi, B., Goldman, J. E., and Brickman, A. M. (2018). White matter changes in Alzheimer’s disease: a focus on myelin and oligodendrocytes. Acta Neuropathol. Commun. 6:22. doi: 10.1186/s40478-018-0515-3
Nave, K.-A., and Werner, H. B. (2014). Myelination of the Nervous System: Mechanisms and Functions. Annu. Rev. Cell Dev. Biol. 30, 503–533. doi: 10.1146/annurev-cellbio-100913-013101
Nawaz, S., Kippert, A., Saab, A. S., Werner, H. B., Lang, T., Nave, K. A., et al. (2009). Phosphatidylinositol 4,5-bisphosphate-dependent interaction of myelin basic protein with the plasma membrane in oligodendroglial cells and its rapid perturbation by elevated calcium. J. Neurosci. 29, 4794–4807. doi: 10.1523/JNEUROSCI.3955-08.2009
Nawaz, S., Sánchez, P., Schmitt, S., Snaidero, N., Mitkovski, M., Velte, C., et al. (2015). Actin filament turnover drives leading edge growth during myelin sheath formation in the central nervous system. Dev. Cell. 34, 139–151. doi: 10.1016/j.devcel.2015.05.013
Nawaz, S., Schweitzer, J., Jahn, O., and Werner, H. B. (2013). Molecular evolution of myelin basic protein, an abundant structural myelin component. Glia 61, 1364–1377. doi: 10.1002/glia.22520
Neilson, K. A., Ali, N. A., Muralidharan, S., Mirzaei, M., Mariani, M., Assadourian, G., et al. (2011). Less label, more free: Approaches in label-free quantitative mass spectrometry. Proteomics 11, 535–553. doi: 10.1002/pmic.201000553
Nguyen, M. V. C., Felice, C. A., Du, F., Covey, M. V., Robinson, J. K., Mandel, G., et al. (2013). Oligodendrocyte lineage cells contribute unique features to rett syndrome neuropathology. J. Neurosci. 33, 18764–18774. doi: 10.1523/JNEUROSCI.2657-13.2013
Norton, W. T., and Poduslo, S. E. (1973a). Myelination in rat brain: changes in myelin composition during brain maturation. J. Neurochem. 21, 759–773. doi: 10.1111/j.1471-4159.1973.tb07520.x
Norton, W. T., and Poduslo, S. E. (1973b). Myelination in rat brain: method of myelin isolation. J. Neurochem. 21, 749–757. doi: 10.1111/j.1471-4159.1973.tb07519.x
Olmos-Serrano, J. L., Kang, H. J., Tyler, W. A., Silbereis, J. C., Cheng, F., Zhu, Y., et al. (2016). Down syndrome developmental brain transcriptome reveals defective oligodendrocyte differentiation and myelination. Neuron 89, 1208–1222. doi: 10.1016/j.neuron.2016.01.042
Ott, C., Martens, H., Hassouna, I., Oliveira, B., Erck, C., Zafeiriou, M. P., et al. (2015). Widespread expression of erythropoietin receptor in brain and its induction by injury. Mol. Med. 21, 803–815. doi: 10.2119/molmed.2015.00192
Patzig, J., Erwig, M. S., Tenzer, S., Kusch, K., Dibaj, P., Möbius, W., et al. (2016). Septin/anillin filaments scaffold central nervous system myelin to accelerate nerve conduction. eLife 5:e17119. doi: 10.7554/eLife.17119.001
Perez-Riverol, Y., Csordas, A., Bai, J., Bernal-Llinares, M., Hewapathirana, S., Kundu, D. J., et al. (2019). The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 47, D442–D450. doi: 10.1093/nar/gky1106
Poitelon, Y., Kopec, A. M., and Belin, S. (2020). Myelin fat facts: an overview of lipids and fatty acid metabolism. Cells 9:812. doi: 10.3390/cells9040812
Quarles, R. H. (2007). Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 100, 1431–1448. doi: 10.1111/j.1471-4159.2006.04319.x
Raasakka, A., Ruskamo, S., Kowal, J., Barker, R., Baumann, A., Martel, A., et al. (2017). Membrane association landscape of myelin basic protein portrays formation of the myelin major dense line. Sci. Rep. 7:4974. doi: 10.1038/s41598-017-05364-3
Roach, A., Takahashi, N., Pravtcheva, D., Ruddle, F., and Hood, L. (1985). Chromosomal mapping of mouse myelin basic protein gene and structure and transcription of the partially deleted gene in shiverer mutant mice. Cell 42, 149–155. doi: 10.1016/S0092-8674(85)80110-0
Roth, A. D., Ivanova, A., and Colman, D. R. (2006). New observations on the compact myelin proteome. Neuron Glia Biol. 2, 15–21. doi: 10.1017/S1740925X06000068
Saher, G., Brügger, B., Lappe-Siefke, C., Möbius, W., Tozawa, R. I., Wehr, M. C., et al. (2005). High cholesterol level is essential for myelin membrane growth. Nat. Neurosci. 8, 468–475. doi: 10.1038/nn1426
Saugier-Veber, P., Munnich, A., Bonneau, D., Rozet, J. M., le Merrer, M., Gil, R., et al. (1994). X-linked spastic paraplegia and Pelizaeus-Merzbacher disease are allelic disorders at the proteolipid protein locus. Nat. Genet. 6, 257–262. doi: 10.1038/ng0394-257
Schaeren-Wiemers, N., Bonnet, A., Erb, M., Erne, B., Bartsch, U., Kern, F., et al. (2004). The raft-associated protein MAL is required for maintenance of proper axon–glia interactions in the central nervous system. J. Cell Biol. 166, 731–742. doi: 10.1083/jcb.200406092
Schardt, A., Brinkmann, B. G., Mitkovski, M., Sereda, M. W., Werner, H. B., and Nave, K.-A. (2009). The SNARE protein SNAP-29 interacts with the GTPase Rab3A: implications for membrane trafficking in myelinating glia. J. Neurosci. Res. 87, 3465–3479. doi: 10.1002/jnr.22005
Schmidt, C., Hesse, D., Raabe, M., Urlaub, H., and Jahn, O. (2013). An automated in-gel digestion/iTRAQ-labeling workflow for robust quantification of gel-separated proteins. Proteomics 13, 1417–1422. doi: 10.1002/pmic.201200366
Schmitt, S., Cantuti Castelvetri, L., and Simons, M. (2015). Metabolism and functions of lipids in myelin. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 1851, 999–1005. doi: 10.1016/j.bbalip.2014.12.016
Sharma, K., Schmitt, S., Bergner, C. G., Tyanova, S., Kannaiyan, N., Manrique-Hoyos, N., et al. (2015). Cell type– and brain region–resolved mouse brain proteome. Nat. Neurosci. 18, 1819–1831. doi: 10.1038/nn.4160
Siems, S. B., Jahn, O., Eichel, M. A., Kannaiyan, N., Wu, L. M. N., Sherman, D. L., et al. (2020). Proteome profile of peripheral myelin in healthy mice and in a neuropathy model. eLife 9:e51406. doi: 10.7554/eLife.51406
Silva, J. C., Gorenstein, M. V., Li, G.-Z., Vissers, J. P. C., and Geromanos, S. J. (2006). Absolute Quantification of Proteins by LCMS E. Mol. Cell. Proteomics 5, 144–156. doi: 10.1074/mcp.M500230-MCP200
Simons, C., Wolf, N. I., McNeil, N., Caldovic, L., Devaney, J. M., Takanohashi, A., et al. (2013). A de novo mutation in the β-tubulin gene TUBB4A results in the leukoencephalopathy hypomyelination with atrophy of the basal ganglia and cerebellum. Am. J. Hum. Genet. 92, 767–773. doi: 10.1016/j.ajhg.2013.03.018
Simons, M., Krämer, E. M., Thiele, C., Stoffel, W., and Trotter, J. (2000). Assembly of myelin by association of proteolipid protein with cholesterol- and galactosylceramide-rich membrane domains. J. Cell Biol. 151, 143–154. doi: 10.1083/jcb.151.1.143
Snaidero, N., and Simons, M. (2017). The logistics of myelin biogenesis in the central nervous system. Glia 65, 1021–1031. doi: 10.1002/glia.23116
Snaidero, N., Velte, C., Myllykoski, M., Raasakka, A., Ignatev, A., Werner, H. B., et al. (2017). Antagonistic Functions of MBP and CNP establish cytosolic channels in CNS myelin. Cell Rep. 18, 314–323. doi: 10.1016/j.celrep.2016.12.053
Sommer, I., and Schachner, M. (1981). Monoclonal antibodies (O1 to O4) to oligodendrocyte cell surfaces: An immunocytological study in the central nervous system. Dev. Biol. 83, 311–327. doi: 10.1016/0012-1606(81)90477-2
Sprinkle, T. J., Sheedlo, H. J., Buxton, T. B., and Rissing, J. P. (1983). Immunochemical Identification of 2’, 3’-Cyclic Nucleotide 3’-Phosphodiesterase in Central and Peripheral Nervous System Myelin, the Wolfgram Protein Fraction, and Bovine Oligodendrocytes. J. Neurochem. 41, 1664–1671. doi: 10.1111/j.1471-4159.1983.tb00878.x
Stuart, T., Butler, A., Hoffman, P., Hafemeister, C., Papalexi, E., Mauck, W. M., et al. (2019). Comprehensive integration of single-cell data. Cell 177:1888-1902.e21. doi: 10.1016/j.cell.2019.05.031
Taylor, C. M., Marta, C. B., Claycomb, R. J., Han, D. K., Rasband, M. N., Coetzee, T., et al. (2004). Proteomic mapping provides powerful insights into functional myelin biology. Proc. Natl. Acad. Sci. U.S.A. 101, 4643–4648. doi: 10.1073/pnas.0400922101
Thakurela, S., Garding, A., Jung, R. B., Müller, C., Goebbels, S., White, R., et al. (2016). The transcriptome of mouse central nervous system myelin. Sci. Rep. 6:25828. doi: 10.1038/srep25828
van der Knaap, M. S., Schiffmann, R., Mochel, F., and Wolf, N. I. (2019). Diagnosis, prognosis, and treatment of leukodystrophies. Lancet Neurol. 18, 962–972. doi: 10.1016/S1474-4422(19)30143-7
Vanrobaeys, F., Van Coster, R., Dhondt, G., Devreese, B., and Van Beeumen, J. (2005). Profiling of myelin proteins by 2D-gel electrophoresis and multidimensional liquid chromatography coupled to MALDI TOF-TOF mass spectrometry. J. Proteome Res. 4, 2283–2293. doi: 10.1021/pr050205c
von Büdingen, H. C., Mei, F., Greenfield, A., Jahn, S., Shen, Y. A. A., Reid, H. H., et al. (2015). The myelin oligodendrocyte glycoprotein directly binds nerve growth factor to modulate central axon circuitry. J. Cell Biol. 210, 891–898. doi: 10.1083/jcb.201504106
Wang, K. C., Koprivica, V., Kim, J. A., Sivasankaran, R., Guo, Y., Neve, R. L., et al. (2002). Oligodendrocyte-myelin glycoprotein is a Nogo receptor ligand that inhibits neurite outgrowth. Nature 417, 941–944. doi: 10.1038/nature00867
Werner, H. B., Krämer-Albers, E. M., Strenzke, N., Saher, G., Tenzer, S., Ohno-Iwashita, Y., et al. (2013). A critical role for the cholesterol-associated proteolipids PLP and M6B in myelination of the central nervous system. Glia 61, 567–586. doi: 10.1002/glia.22456
Werner, H. B., Kuhlmann, K., Shen, S., Uecker, M., Schardt, A., Dimova, K., et al. (2007). Proteolipid Protein Is Required for Transport of Sirtuin 2 into CNS Myelin. J. Neurosci. 27, 7717–7730. doi: 10.1523/JNEUROSCI.1254-07.2007
Yan, H., Helman, G., Murthy, S. E., Ji, H., Crawford, J., Kubisiak, T., et al. (2019). Heterozygous Variants in the Mechanosensitive Ion Channel TMEM63A Result in Transient Hypomyelination during Infancy. Am. J. Hum. Genet. 105, 996–1004. doi: 10.1016/j.ajhg.2019.09.011
Yang, H. J., Vainshtein, A., Maik-Rachline, G., and Peles, E. (2016). G protein-coupled receptor 37 is a negative regulator of oligodendrocyte differentiation and myelination. Nat. Commun. 7:10884. doi: 10.1038/ncomms10884
Yool, D. A., Klugmann, M., McLaughlin, M., Vouyiouklis, D. A., Dimou, L., Barrie, J. A., et al. (2001). Myelin proteolipid proteins promote the interaction of oligodendrocytes and axons. J. Neurosci. Res. 63, 151–164. doi: 10.1002/1097-4547(20010115)63:2<151::AID-JNR1007>3.0.CO;2-Y
Zeisel, A., Muñoz-Manchado, A. B., Codeluppi, S., Lönnerberg, P., Manno, G. L., Juréus, A., et al. (2015). Cell types in the mouse cortex and hippocampus revealed by single-cell RNA-seq. Science 347, 1138–1142. doi: 10.1126/science.aaa1934
Zhang, Y., Chen, K., Sloan, S. A., Bennett, M. L., Scholze, A. R., O’Keeffe, S., et al. (2014). An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 34, 11929–11947. doi: 10.1523/JNEUROSCI.1860-14.2014
Keywords: oligodendrocyte, myelin proteome, central nervous system (CNS), demyelination, post-mortem delay, autopsy, label-free proteomics, transcriptome
Citation: Jahn O, Siems SB, Kusch K, Hesse D, Jung RB, Liepold T, Uecker M, Sun T and Werner HB (2020) The CNS Myelin Proteome: Deep Profile and Persistence After Post-mortem Delay. Front. Cell. Neurosci. 14:239. doi: 10.3389/fncel.2020.00239
Received: 14 May 2020; Accepted: 07 July 2020;
Published: 19 August 2020.
Edited by:
Nicola B. Hamilton-Whitaker, King’s College London, United KingdomReviewed by:
Zsolt Illes, University of Southern Denmark, DenmarkCopyright © 2020 Jahn, Siems, Kusch, Hesse, Jung, Liepold, Uecker, Sun and Werner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hauke B. Werner, SGF1a2VAZW0ubXBnLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.