
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Behav. Neurosci., 26 January 2024
Sec. Learning and Memory
Volume 18 - 2024 | https://doi.org/10.3389/fnbeh.2024.1359729
This article is part of the Research TopicCortico-hippocampal Interactions During Learning and MemoryView all 9 articles
 Verónica Pastor1,2*†
Verónica Pastor1,2*† Cynthia Katche1,3*†
Cynthia Katche1,3*†In the retrosplenial cortex (RSC), the role of cholinergic modulation via α7 nicotinic receptors and their involvement in memory is unknown. In recent years, the RSC has been shown to deteriorate in the early stages of Alzheimer’s disease (AD). Likewise, the cholinergic system has been postulated as one of those responsible for cognitive impairment in patients with AD. Great interest has arisen in the study of α7 nicotinic receptors as more specific targets for the treatment of this disease. For this reason, we aim to study the role of α7 receptors of the RSC in memory processing. We infused a selective α7 receptor antagonist into the anterior part of the RSC (aRSC) to assess its role in different phases of aversive memory processing using an inhibitory avoidance task. We found that α7 nicotinic receptors are involved in memory acquisition and expression, but not in its consolidation. These results identify aRSC α7 nicotinic receptors as key players in aversive memory processing and highlight their significant potential as therapeutic targets for Alzheimer’s disease.
The retrosplenial cortex (RSC) is essential for learning and memory processing (Corcoran et al., 2018; Stacho and Manahan-Vaughan, 2022). In rodents, the retrosplenial cortex comprises the entire posterior cingulum. In humans, it comprises only Broadman’s areas 29 and 30 (Vogt and Peters, 1981). During the last decades, several lesion and neuroimaging studies demonstrated its requirement and activation in memory tasks in humans (Valenstein et al., 1987; Maguire, 2001) and rodents (Sutherland et al., 1988; Vann and Aggleton, 2002, 2004). In particular, our studies in the inhibitory avoidance task highlighted the relevance of the anterior part of the RSC (aRSC) for the consolidation and retrieval of aversive memories (Katche et al., 2013).
Mostly, the RSC has attracted attention in recent years because it is affected in the early stages of Alzheimer’s disease (AD) (Pengas et al., 2010; Aggleton, 2014; Tu et al., 2015) and cognitive impairments associated with aging (Trask and Fournier, 2022). The cholinergic system has been postulated as one of those responsible for cognitive impairment in patients with AD (Hampel et al., 2018; Hoskin et al., 2019; Koukouli and Changeux, 2020). In this sense, the study of pharmacological strategies aimed at modifying cholinergic tone such as acetylcholinesterase inhibitors has dominated preclinical and clinical studies in the field of AD. In fact, in a rodent model of AD, deposits of the ABeta peptide have been described in cholinergic axons innervating the RSC (Robertson et al., 2009).
The RSC is known to receive cholinergic inputs from the basal forebrain (Bigl et al., 1982; Mesulam et al., 1983). However, the role of RSC cholinergic modulation in memory processing is yet to be elucidated. Among ionotropic nicotinic acetylcholine receptors (nAChRs), the α7 subtype is one of the most prominent in the central nervous system. Compared with other nAChR subtypes, α7 nAChR has the highest calcium permeability, which make these receptors key players in plasticity processes, such as long-term potentiation (LTP), and then, in learning and memory (Koukouli and Changeux, 2020; Pastor and Medina, 2023). Indeed, several reports pointed out α7 nAChRs as promising pharmacological targets for cognitive improvement in neurological disorders such as AD (see for references Greenfield et al., 2022). α7 nAChRs are expressed in the RSC, mainly in gabaergic interneurons (Murakami et al., 2013). However, whether their activation is required for memory processing is largely unknown. Here, we studied in the aRSC the effect of the infusion of methyllycaconitine (MLA), a selective antagonist of α7 nAChRs, on different phases of an aversive memory in adult rats. Our results show that α7 nAChRs in the aRSC are differentially involved in the acquisition, consolidation, or retrieval of an aversive memory.
Experiments were conducted in male Wistar rats (UBA, Argentina) weighting 220–250 g. Animals were housed three to a cage and kept at a constant temperature of 23°C, with water and food ad libitum, under a 12-h light/dark cycle (lights on at 7:00 am). Experimental procedures were performed in accordance with the USA National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Animal Care and Use Committees of the University of Buenos Aires (CICUAL).
Animals were anesthetized with a mix of ketamine (80 mg/kg) and xylazine (10 mg/kg) and placed on a stereotaxic frame. The skull was exposed and leveled (flat skull, lambda and bregma at the same elevation). 22-G guide cannulas were bilaterally implanted, aimed to the aRSC: AP −3.9 mm/LL ± 0.5 mm/DV −1.8 mm (from Bregma; Paxinos and Watson, 2007). Cannulas were fixed to the skull with dental acrylic and protected with modified 30-G needles. Immediately after surgery, animals were injected with meloxicam (0.2 mg/kg) as analgesic and gentamicin (2 mg/kg) as antibiotic. Animals were allowed to recover from surgery for 5–7 days before any experimental manipulation.
Infusions were delivered through an injector cannula extending 1 mm beyond the tip of the guide cannula connected to 10 μL Hamilton syringes. The volume infused was 1 μl per side and the infusion rate was 1 μl/min. The injector was left in place for an additional minute after infusion to allow diffusion and to prevent reflux. Dose was as follows: Methyllycaconitine Citrate (MLA; Abcam#ab120072) was dissolved in sterile saline solution and administered bilaterally intra-aRSC (5 μg/μl/hemisphere). The same volume of vehicle (1 μl/hemisphere) was used in control rats. Indicated dose was based on the molecular weight of the salt (MLA) and was determined based on preliminary studies and previous reports (Chan et al., 2007; Pastor et al., 2021, 2023). After completing the behavioral procedures, the positioning of the cannula was confirmed by the aRSC infusion of 1 μl of 4% methylene blue in saline. The extension of the dye was taken as indicative of the diffusion of the drug, as previously published (de Landeta et al., 2022). Only animals with both cannulae in the correct place were included in the analysis.
After recovery from surgery, animals were handled once a day for two days and then trained in the inhibitory avoidance task (IA) as described previously (Bekinschtein et al., 2007). Briefly the apparatus was a 50 cm × 25 cm × 25 cm acrylic box whose floor was a grid made of 1 mm-caliber steel bars. The left end of the grid was covered with a 7 cm-wide, 5 cm-high acrylic platform. Rats were infused into the aRSC with MLA or vehicle 15 min before the training session, immediately after the training session, or 40 min before the test, depending on the experiment. For training, animals were placed on the platform and as they stepped down onto the grid received a single 3-sec, 0.5 mA scrambled footshock. Rats were tested for retention 24 h after training. All animals were tested only once. In the test sessions the footshock was omitted. The time spent on the platform during this session was taken as an indicator of retention.
Locomotor activity was assessed during 5 min in an open field arena (50 cm × 50 cm × 39 cm) which had black walls and floor divided into nine squares by white lines. Rats were infused into the aRSC with MLA or vehicle 15 min before being placed in the open field. The number of line crossings and the number of entries to the central square were measured.
Immediately after the OF test, anxiety-like behavior was assessed in the same group of rats using a maze elevated 65 cm over the floor, which consisted of two open (45 cm length × 10 cm width) and two closed arms (45 cm length × 10 cm width × 50 cm height) opposite to each other and inter-connected by a central platform (10 cm × 10 cm). Each animal was placed in the center, facing an open arm, and left in the maze for 5 min. The percentage of time spent in open arms and the number of entries to open arms were used as indices of anxiety, whereas the total number of entries to open and closed arms were used as an index of general locomotion.
The SOD task was conducted in an independent group of rats to evaluate if the infusion of the α7 nAChR antagonist affected attention and perception (Bekinschtein et al., 2013). The exploration area was triangular in shape with high, homogenous white walls constructed from Perspex to prevent the rat from looking out into the room. All walls were 60 cm wide and 60 cm tall, and the objects were placed 2.50 cm apart along the back wall during the test session. All rats were habituated to the empty apparatus for 10 min in which they were allowed to freely explore it. For the habituation session, the rat was placed in the triangular apparatus, facing the corner of the triangle away from the back wall. Test session began 24 h after the habituation session. MLA and vehicle infusions were made 15 min before the test. All object sets (2 repeated and 1 different) were placed in the back wall of the apparatus before the rat was placed. The rat was placed in the corner of the triangular apparatus (away from the objects). The time spent exploring the three objects during a testing phase was scored for 5 min by an experimenter viewing the rat. Exploration of an object was defined as directing the nose to the object at a distance of < 2 cm and/or touching it with the nose. A SOD index was calculated as follows: [time of exploration of the different object–(time of exploration of similar objects/2)]/total time of object exploration.
The hot plate test was used in an independent group of rats to determine the potential analgesic activity induced by the antagonism of α7 nAChRs in the aRSC. MLA was infused 15 min before placing the rats on the hot plate until a nociceptive response (i.e., paw withdrawal) was observed. An observer blind to the treatment measured the latency of paw withdrawal using a manual stopwatch. The temperature of the hot plate was set and calibrated at 48°C.
Data was analyzed by Student’s t-test, Mann–Whitney or Wilcoxon signed rank test as appropriate. For simplicity, all the results are presented as mean ± SEM and each point represents an individual animal. GraphPad Outliers Grubbs’ test with alpha 0.05 was used to assess the presence of outliers. All experiments were repeated twice to assess reproducibility which was achieved in all cases. Significant differences were set at p < 0.05.
To assess the role of aRSC α7 nAChRs during the acquisition phase of an aversive memory, we infused MLA in the aRSC 15 min before the IA training (Figure 1). We found longer latency to step down compared to controls at 24 h in MLA-infused rats (Mann–Whitney test; U = 36; p = 0.014; nVEH = 15; nMLA = 11), suggesting that aRSC α7 nAChRs are involved in IA memory encoding. Three rats were excluded from the analysis due to cannulae misplacement.
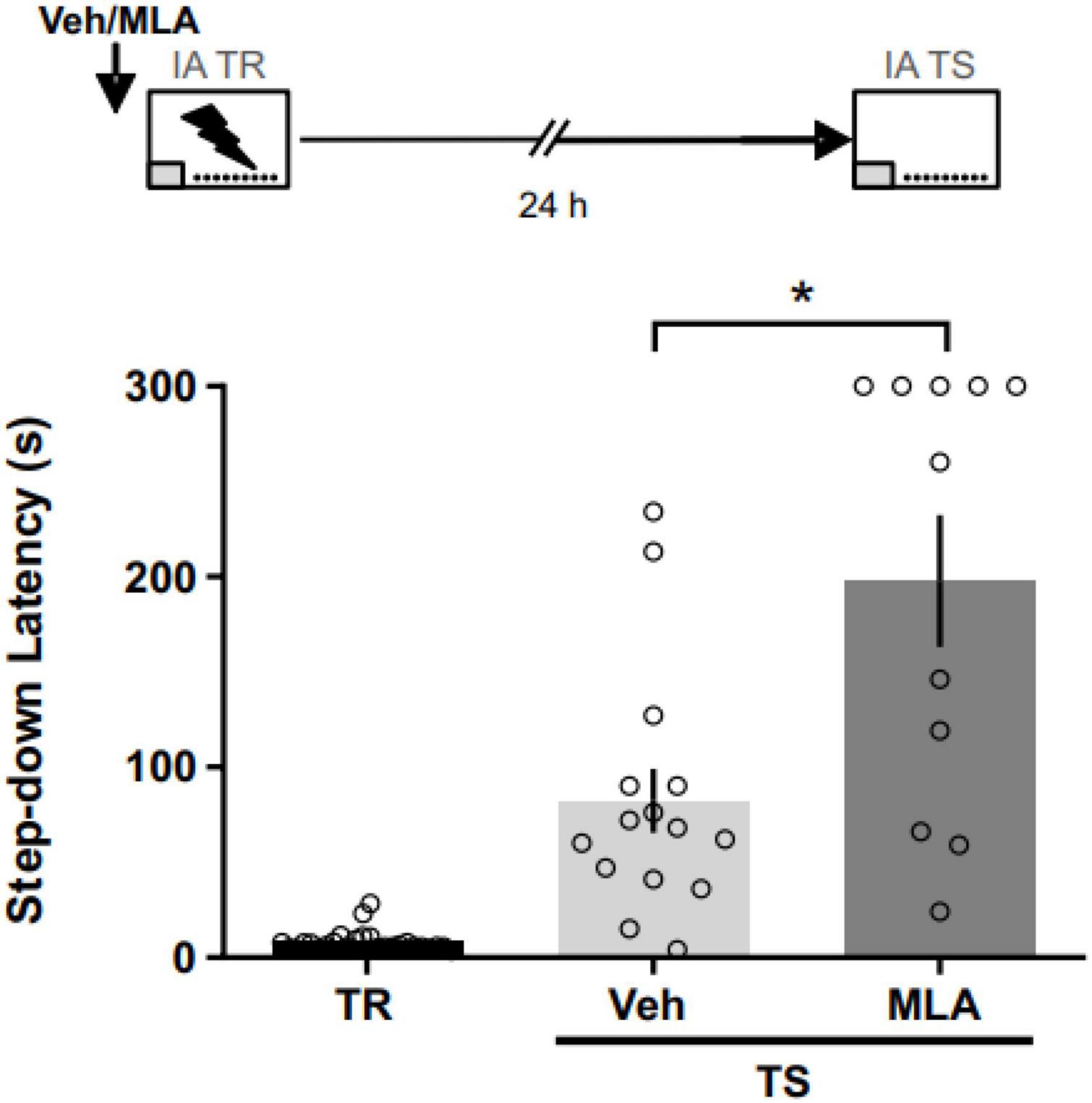
Figure 1. α7 nAChRs antagonism in the aRSC before IA training improved long-term memory expression. Data are expressed as the mean ± SEM of the latency to step down during training (TR) or 24 h test (TS) for rats infused with methyllycaconitine (MLA) or vehicle (Veh) 15 min before TR. Mann–Whitney test; *p < 0.05; n = 11–15.
To assess the role of aRSC α7 nAChRs in the consolidation phase of an aversive memory, we infused MLA in the aRSC immediately after the IA training (Figure 2). We found no differences between MLA-infused rats and vehicle controls at 24 h test (Mann–Whitney test; U = 63; p = 0.687; nVEH = 14; nMLA = 10), suggesting no involvement of aRSC α7 nAChRs in IA memory consolidation. Two rats were excluded from the analysis due to cannulae misplacement.
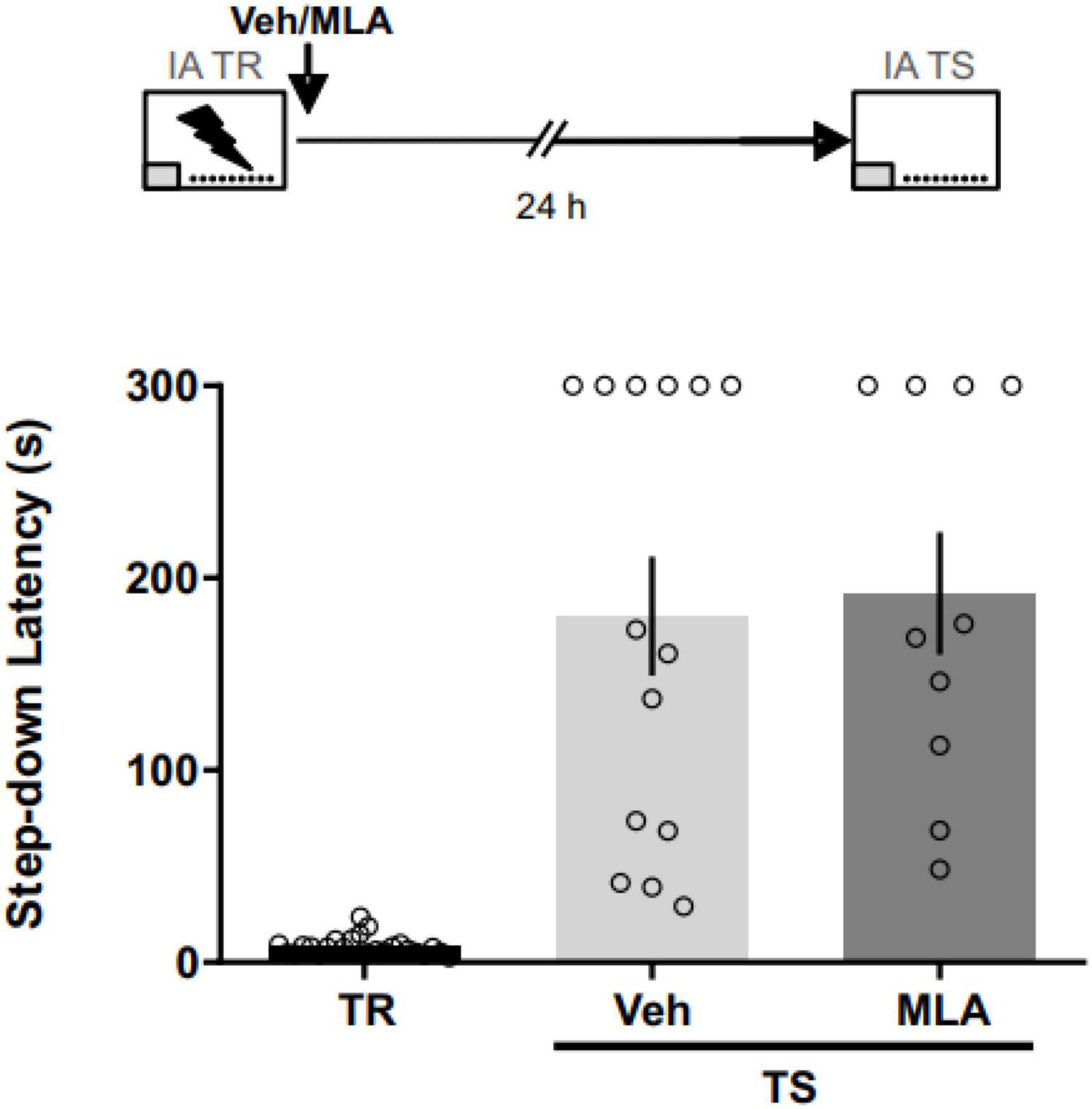
Figure 2. α7 nAChRs antagonism in the aRSC did not affect IA memory consolidation. Data are expressed as the mean ± SEM of the latency to step down during training (TR) or 24 h test (TS). No significant differences were found between rats infused with methyllycaconitine (MLA) or vehicle (Veh) immediately after TR; Mann–Whitney test, n = 10–14.
To assess the role of aRSC α7 nAChRs in IA memory expression, we infused MLA before the 24 h test (Figure 3). We found that MLA shortened latency to step down compared to controls (Figure 3A; Mann–Whitney test; U = 17; p = 0.0030; n = 11 per group), supporting the role of aRSC α7 nAChRs in aversive memory retrieval. Six rats were excluded from the analysis due to cannulae misplacement. In support of our results, in those animals with cannulae misplacement (Figure 3B), MLA did not show any memory blocking effect, evidenced as an increase in the latency to step-down in the test compared to the training (Wilcoxon one-tailed signed rank test; W = 21; p = 0.015; n = 6).
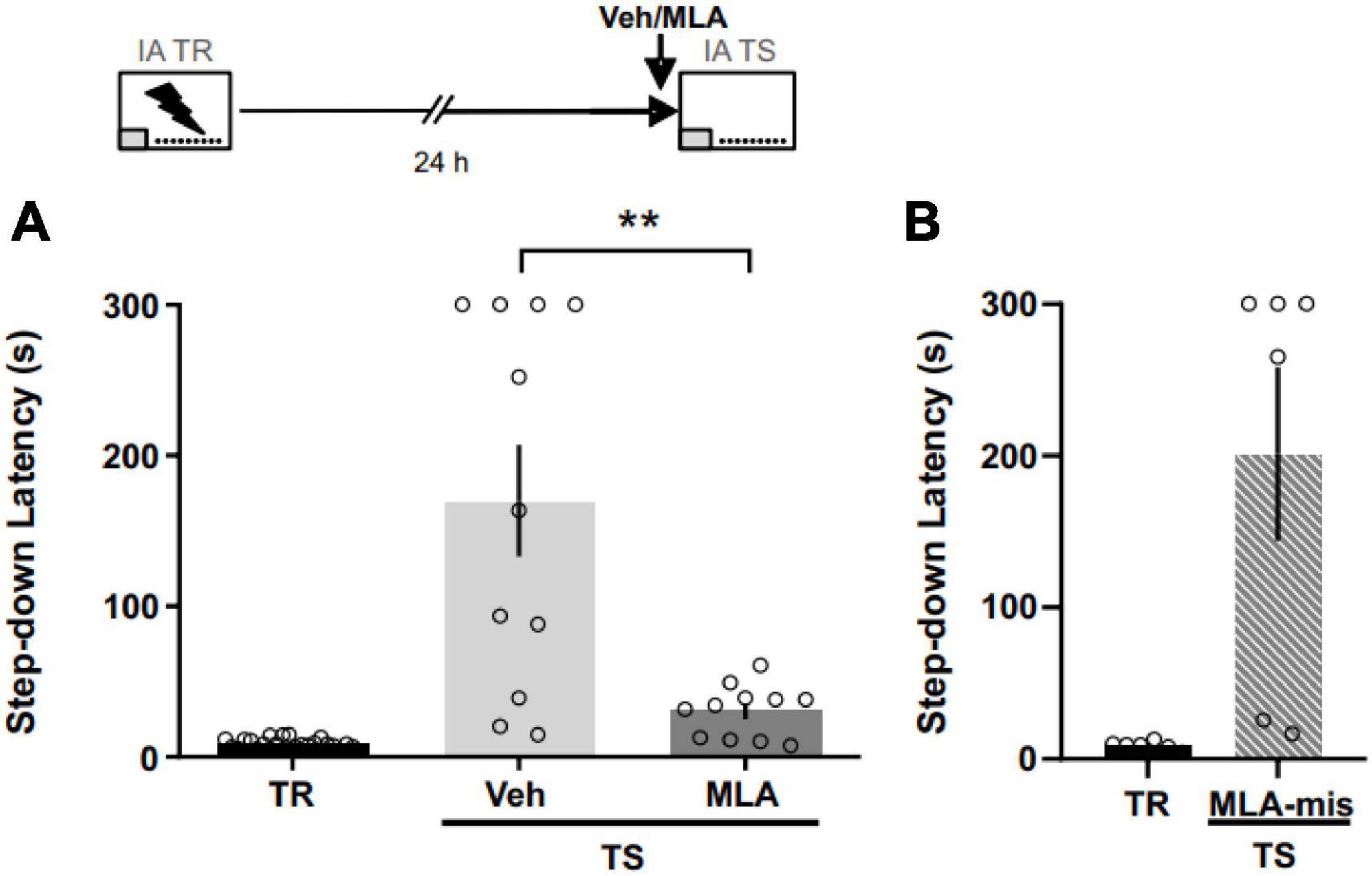
Figure 3. α7 nAChRs antagonism in the aRSC blocked IA memory expression. (A) Data are expressed as the mean ± SEM of the latency to step down during training (TR) or 24 h test (TS) for rats infused with methyllycaconitine (MLA) or vehicle (Veh) 40 min before TS. Mann Whitney test; **p < 0.01; n = 11. (B) Data are expressed as the mean ± SEM of the latency to step down during training (TR) or 24 h test (TS) for rats infused with methyllycaconitine (MLA) with cannulae misplacement.
To investigate if the antagonism of aRSC α7 nAChRs may induce any deficit on locomotor activity which could interfere with our results, we performed an OF test following the infusion of MLA in the aRSC. We found no differences between MLA-infused rats and vehicle controls in the total number of crossings [Figure 4A; Student’s t-test; t(14) = 0.77; p = 0.456] nor in the number of entries to the center zone [Figure 4B; Student’s t-test; t(14) = 0; p > 0.999; n = 8 per group]. To assess the effect of the antagonism of α7 nAChRs on the anxiety-like state in rats infused in the aRSC, we performed an EPM test following MLA administration. We found no differences in the total number of entries to the open arms [Figure 4C; Student’s t-test; t(14) = 0.323; p = 0.751; n = 8 per group] nor in the percentage of time in open arms [Figure 4D; Student’s t-test; t(14) = 1.263; p = 0.227; n = 8 per group] between MLA-infused rats and vehicle controls. Total entries to open + closed arms were quantified to assess locomotor activity during the EPM test and showed no differences between groups [11.5 ± 2.8 for vehicle; 12.4 ± 1.5 for MLA; Student’s t-test: t(14) = 0.276; p = 0.786; n = 8 per group].
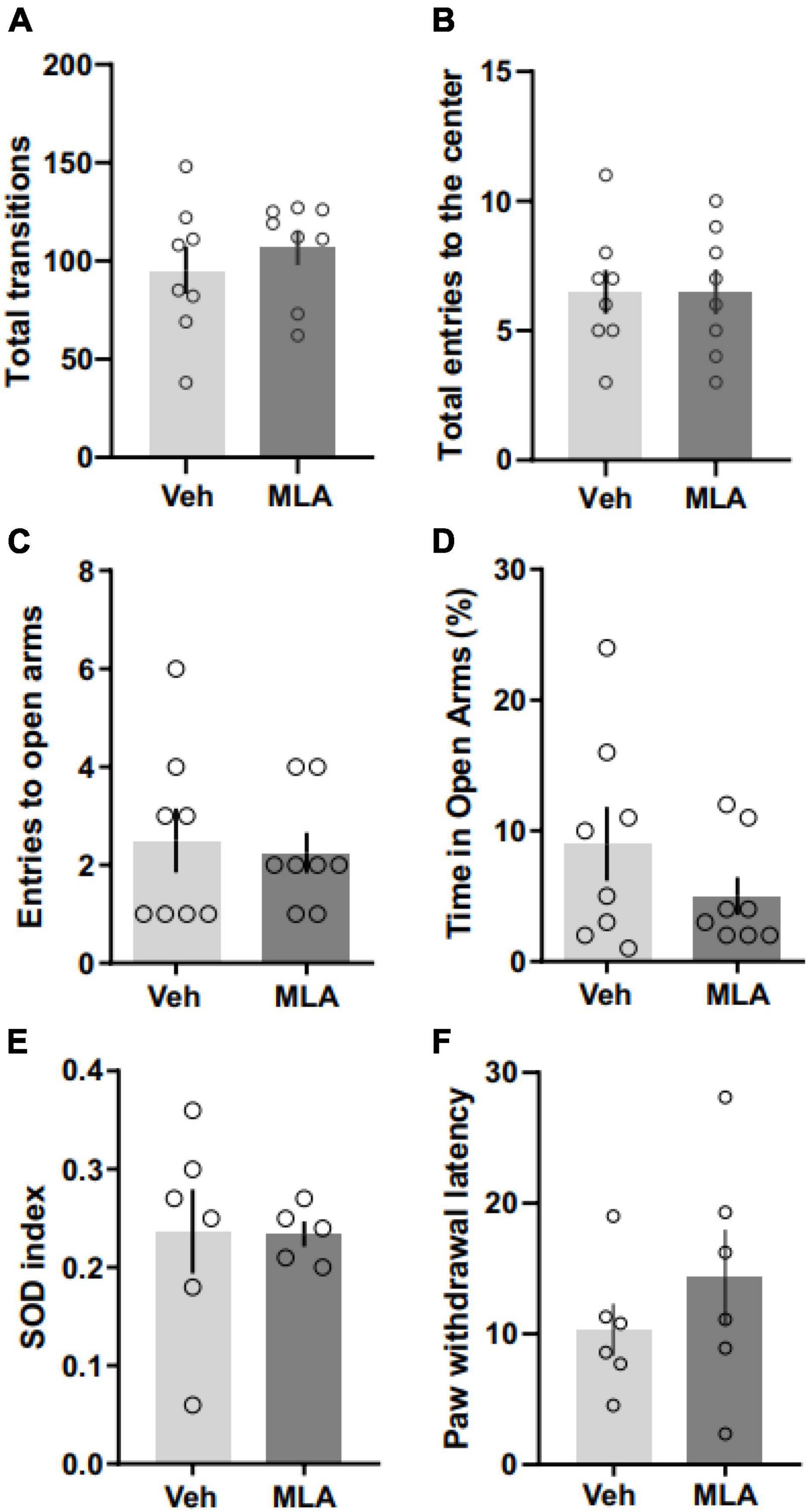
Figure 4. α7 nAChRs antagonism in the aRSC did not affect locomotor activity, anxiety-like behavior, attention, perception or nociception. Data are expressed as the mean + SEM of the total number of transitions (A) or the number of entries to the center zone (B) of an open field; the number of entries to the open arms (C) and the percentage of time spent in open arms (D) of the elevated plus maze; the SOD index (E) or the latency to paw withdrawal in the hot plate test (F). No significant differences were found by Student’s t-test between rats infused with methyllycaconitine (MLA) or vehicle (Veh) 15 min before the behavioral test (n = 5–8).
Considering that our results could be influenced by differences in attention or sensory perception, we decided to assess if the infusion of MLA in the aRSC had any effect on the simultaneous oddity discrimination task or on the hotplate test. We found no differences between MLA and vehicle infused rats in the SOD task (Figure 4E; Mann–Whitney test; U = 12; p = 0.628; n = 5–6) nor in the hotplate test [Figure 4F; Student’s t-test; t(10) = 0.9633; p = 0.358; n = 6], suggesting that the antagonism of α7 nAChRs in the aRSC did not affect sensory perception nor attention.
In this study, we demonstrated for the first time that aRSC α7 nAChRs are critically involved in the processing of information related to fearful experiences. By using a one-trial IA task and local infusion of MLA in the aRSC, we found that α7 nAChRs are differentially involved in the acquisition, consolidation, and expression phases of an aversive memory.
The aRSC is necessary for aversive memory processing (Katche et al., 2013). However, the cellular and molecular mechanisms underlying its function are not fully understood. Different neurotransmitter systems are likely involved in memory processing in the aRSC. Given the importance of the RSC and of the cholinergic system, specifically the α7 nAChR subtype, on AD pathogenesis, we focused on the study of those receptors in memory processing.
We first asked if α7 nAChR activation was necessary for the acquisition of an aversive memory in the aRSC. Our results showed that the antagonism of α7 nAChRs in the aRSC during memory acquisition led to an improvement in memory expression the day after, supporting their involvement in memory encoding. Due to the lack of information about the role of α7 nAChRs in the aRSC, this finding could be interpreted in different ways. Considering that attention is required for memory encoding (Muzzio et al., 2009), aRSC α7 nAChRs role in IA memory encoding may be related to their involvement in attention and not only in memory processing (Ballinger et al., 2016). Thus, we performed a control experiment to assess the effect of infusing MLA in the aRSC before the SOD test. Our results suggest that aRSC α7 nAChRs are not involved in attention processing. Another possibility is that aRSC α7 nAChRs may be modulating how sensory information is perceived when the shock is received during the training session. To assess this possibility, we performed a hot plate test following the infusion of MLA. We did not find any effect suggesting that aRSC α7 nAChRs are not involved in sensory perception.
It is noteworthy that when MLA is infused before training, it likely remains in the brain during and after the training session (Turek et al., 1995). This makes it challenging to determine if MLA effects on memory result from enhancing encoding or affecting early consolidation processes. Therefore, we performed another experiment where we administered MLA immediately after the training session. We found no changes in memory expression, supporting the idea that α7 nAChRs are involved in the encoding but not the consolidation phase of IA memory. This is consistent with previous research showing a differential role of α7 nAChRs in encoding but not consolidation of rewarding memory in the medial prefrontal cortex (Pastor et al., 2021), highlighting that cortical α7 nAChRs have an essential role in the acquisition of different types of memories.
Previous reports have shown that the RSC is involved in the retrieval of aversive memories (Fournier et al., 2021), including those assessed by the IA task (Katche et al., 2013). Thus, we aimed to investigate the potential involvement of the cholinergic system, specifically through α7 nAChRs, in mediating that function. Differently from what we reported for the medial prefrontal cortex, where α7 nAChRs are not involved in aversive memory expression (Pastor et al., 2023), here we found that α7 nAChRs activation in the aRSC is critical for IA memory expression. These findings identify the aRSC as a key brain area where cholinergic modulation through α7 nAChRs influences aversive memory expression.
There is still debate about how the RSC participates in aversive memory processing. Prior studies using fear conditioning protocols showed that the RSC is involved in the retrieval but not in the acquisition of the aversive memory (Fournier et al., 2021). However, an increased activation of the RSC following mild tail shock paired with tactile stimuli was evidenced in mice by assessing the levels of c-fos expression (Radwanska et al., 2010), supporting its role in the acquisition of aversive classical associative learning. Thus, it is likely that different cellular and molecular mechanisms are involved in different phases of aversive memory processing in the RSC. Regarding α7 nAChRs, there is scarce information in the literature about their localization and function in the RSC. It has been described that α7 nAChRs are mainly expressed in RSC GABAergic neurons (Murakami et al., 2013). Thus, the activation of these receptors is likely to modulate local neuronal activity in a complex manner. At a synaptic level, inactivation of α7 nAChRs may facilitate LTP induction as was reported in the hippocampus (Fujii et al., 2000; Miguelez Fernández et al., 2021), thus facilitating memory acquisition. Further research is needed to assess this possibility.
In conclusion, our results suggest a dual effect of α7 nAChRs in the aRSC, highlighting their dynamic involvement in different stages of memory processing. It remains elusive the mechanisms by which α7 nAChRs modulation differentially affects encoding and retrieval of the IA memory and further research is needed to uncover the processes that underlie the observed dual effect. Elucidating these mechanisms will not only enhance our understanding of α7 nAChRs modulation in the aRSC, but also enhance the development of targeted interventions for memory-related disorders.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was approved by the Animal Care and Use Committee of the University Buenos Aires (CICUAL). School of Medicine, University of Buenos Aires. The study was conducted in accordance with the local legislation and institutional requirements.
VP: Conceptualization, Funding acquisition, Investigation, Writing – original draft. CK: Conceptualization, Funding acquisition, Investigation, Writing – original draft.
The authors declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by a grant from the National Agency of Scientific and Technological Promotion of Argentina (ANPCyT, Argentina, grant number 2018-00762), the Young IBRO Maternity/Parenthood Grant to CK, the National Scientific and Technical Research Council (CONICET, PIBAA 2021-00261 to VP, Argentina), and Instituto Tecnológico de Buenos Aires (ITBA, Argentina).
We thank Dr. Jorge Medina for useful comments and discussion on the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aggleton, J. P. (2014). Looking beyond the hippocampus: old and new neurological targets for understanding memory disorders. Proc. Biol. Sci. 281:20140565. doi: 10.1098/RSPB.2014.0565
Ballinger, E. C., Ananth, M., Talmage, D. A., and Role, L. W. (2016). Basal forebrain cholinergic circuits and signaling in cognition and cognitive decline. Neuron 91, 1199–1218. doi: 10.1016/j.neuron.2016.09.006
Bekinschtein, P., Cammarota, M., Igaz, L. M., Bevilaqua, L. R. M., Izquierdo, I., and Medina, J. H. (2007). Persistence of long-term memory storage requires a late protein synthesis- and BDNF- dependent phase in the hippocampus. Neuron 53, 261–277. doi: 10.1016/j.neuron.2006.11.025
Bekinschtein, P., Renner, M. C., Gonzalez, M. C., and Weisstaub, N. (2013). Role of medial prefrontal cortex serotonin 2A receptors in the control of retrieval of recognition memory in rats. J. Neurosci. 33, 15716–15725. doi: 10.1523/JNEUROSCI.2087-13.2013
Bigl, V., Woolf, N. J., and Butcher, L. L. (1982). Cholinergic projections from the basal forebrain to frontal, parietal, temporal, occipital, and cingulate cortices: a combined fluorescent tracer and acetylcholinesterase analysis. Brain Res. Bull. 8, 727–749. doi: 10.1016/0361-9230(82)90101-0
Chan, W. K., Tsun-Hon Wong, P., and Sheu, F. S. (2007). Frontal cortical α7 and α4β2 nicotinic acetylcholine receptors in working and reference memory. Neuropharmacology 52, 1641–1649. doi: 10.1016/j.neuropharm.2007.03.008
Corcoran, K. A., Yamawaki, N., Leaderbrand, K., and Radulovic, J. (2018). Role of retrosplenial cortex in processing stress-related context memories. Behav. Neurosci. 132, 388–395. doi: 10.1037/bne0000223
de Landeta, A. B., Medina, J. H., and Katche, C. (2022). Dopamine D1/D5 receptors in the retrosplenial cortex are necessary to consolidate object recognition memory. Front. Behav. Neurosci. 16:922971. doi: 10.3389/fnbeh.2022.922971
Fournier, D. I., Cheng, H. Y., Tavakkoli, A., Gulledge, A. T., Bucci, D. J., and Todd, T. P. (2021). Retrosplenial cortex inactivation during retrieval, but not encoding, impairs remotely acquired auditory fear conditioning in male rats. Neurobiol. Learn. Mem. 185, 1–25. doi: 10.1016/j.nlm.2021.107517
Fujii, S., Ji, Z., and Sumikawa, K. (2000). Inactivation of α7 ACh receptors and activation of non-α7 ACh receptors both contribute to long term potentiation induction in the hippocampal CA1 region. Neurosci. Lett. 286, 134–138. doi: 10.1016/S0304-3940(00)01076-4
Greenfield, S. A., Cole, G. M., Coen, C. W., Frautschy, S., Singh, R. P., Mekkittikul, M., et al. (2022). A novel process driving Alzheimer’s disease validated in a mouse model: therapeutic potential. Alzheimers Dement. Transl. Res. Clin. Interv. 8, 1–12. doi: 10.1002/trc2.12274
Hampel, H., Mesulam, M. M., Cuello, A. C., Farlow, M. R., Giacobini, E., Grossberg, G. T., et al. (2018). The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 141, 1917–1933. doi: 10.1093/brain/awy132
Hoskin, J. L., Al-Hasan, Y., and Sabbagh, M. N. (2019). Nicotinic acetylcholine receptor agonists for the treatment of Alzheimer’s dementia: an update. Nicotine Tob. Res. 21, 370–376. doi: 10.1093/ntr/nty116
Katche, C., Dorman, G., Slipczuk, L., Cammarota, M., and Medina, J. H. (2013). Functional integrity of the retrosplenial cortex is essential for rapid consolidation and recall of fear memory. Learn. Mem. 20, 170–173. doi: 10.1101/lm.030080.112
Koukouli, F., and Changeux, J. P. (2020). Do nicotinic receptors modulate high-order cognitive processing? Trends Neurosci. 43, 550–564. doi: 10.1016/j.tins.2020.06.001
Maguire, E. A. (2001). The retrosplenial contribution to human navigation: a review of lesion and neuroimaging findings. Scand. J. Psychol. 42, 225–238. doi: 10.1111/1467-9450.00233
Mesulam, M.-M., Mufson, E. J., Levey, A. I., and Wainer, B. H. (1983). Cholinergic innervation of cortex by the basal forebrain: cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (Substantia innominata), and hypothalamus in the rhesus monkey. J. Comp. Neurol. 214, 170–197. doi: 10.1002/cne.902140206
Miguelez Fernández, A. M. M., Molla, H. M., Thomases, D. R., and Tseng, K. Y. (2021). Prefrontal a7nachr signaling differentially modulates afferent drive and trace fear conditioning behavior in adolescent and adult rats. J. Neurosci. 41, 1908–1916. doi: 10.1523/JNEUROSCI.1941-20.2020
Murakami, K., Ishikawa, Y., and Sato, F. (2013). Localization of α7 nicotinic acetylcholine receptor immunoreactivity on GABAergic interneurons in layers I-III of the rat retrosplenial granular cortex. Neuroscience 252, 443–459. doi: 10.1016/j.neuroscience.2013.08.024
Muzzio, I. A., Kentros, C., and Kandel, E. (2009). What is remembered? Role of attention on the encoding and retrieval of hippocampal representations. J. Physiol. 587, 2837–2854. doi: 10.1113/JPHYSIOL.2009.172445
Pastor, V., and Medina, J. H. (2023). α7 nicotinic acetylcholine receptor in memory processing. Eur. J. Neurosci. doi: 10.1111/ejn.15913 [Epub ahead of print].
Pastor, V., Castillo Díaz, F., Sanabria, V. C., Dalto, J. F., Antonelli, M. C., and Medina, J. H. (2021). Prefrontal cortex nicotinic receptor inhibition by methyllycaconitine impaired cocaine-associated memory acquisition and retrieval. Behav. Brain Res. 406:113212. doi: 10.1016/j.bbr.2021.113212
Pastor, V., Dalto, J. F., and Medina, J. H. (2023). α 7 nicotinic acetylcholine receptors in the medial prefrontal cortex control rewarding but not aversive memory expression in a dopamine-sensitive manner. Pharmacol. Biochem. Behav. 228:173594. doi: 10.1016/j.pbb.2023.173594
Paxinos, G., and Watson, C. (2007). The rat brain in stereotaxic coordinates. San Diego, CA: Academic Press.
Pengas, G., Hodges, J. R., Watson, P., and Nestor, P. J. (2010). Focal posterior cingulate atrophy in incipient Alzheimer’s disease. Neurobiol. Aging 31, 25–33. doi: 10.1016/J.NEUROBIOLAGING.2008.03.014
Radwanska, A., Debowska, W., Liguz-Lecznar, M., Brzezicka, A., Kossut, M., and Cybulska-Klosowicz, A. (2010). Involvement of retrosplenial cortex in classical conditioning. Behav. Brain Res. 214, 231–239. doi: 10.1016/j.bbr.2010.05.042
Robertson, R. T., Baratta, J., Yu, J., and LaFerla, F. M. (2009). Amyloid-beta expression in retrosplenial cortex of triple transgenic mice: relationship to cholinergic axonal afferents from medial septum. Neuroscience 164, 1334–1346. doi: 10.1016/J.NEUROSCIENCE.2009.09.024
Stacho, M., and Manahan-Vaughan, D. (2022). Mechanistic flexibility of the retrosplenial cortex enables its contribution to spatial cognition. Trends Neurosci. 45, 284–296. doi: 10.1016/j.tins.2022.01.007
Sutherland, R. J., Wishaw, I. Q., and Kolb, B. (1988). Contributions of cingulate cortex to two forms of spatial learning and memory. J. Neurosci. 8, 1863–1872. doi: 10.1523/jneurosci.08-06-01863.1988
Trask, S., and Fournier, D. I. (2022). Examining a role for the retrosplenial cortex in age-related memory impairment. Neurobiol. Learn. Mem. 189:107601. doi: 10.1016/J.NLM.2022.107601
Tu, S., Wong, S., Hodges, J. R., Irish, M., Piguet, O., and Hornberger, M. (2015). Lost in spatial translation–A novel tool to objectively assess spatial disorientation in Alzheimer’s disease and frontotemporal dementia. Cortex 67, 83–94. doi: 10.1016/J.CORTEX.2015.03.016
Turek, J. W., Kang, C. H., Campbell, J. E., Arneric, S. P., and Sullivan, J. P. (1995). A sensitive technique for the detection of the α7 neuronal nicotinic acetylcholine receptor antagonist, methyllycaconitine, in rat plasma and brain. J. Neurosci. Methods 61, 113–118. doi: 10.1016/0165-0270(95)00032-P
Valenstein, E., Bowers, D., Verfaellie, M., Heilman, K. M., Day, A., and Watson, R. T. (1987). Retrosplenial amnesia. Brain 110(Pt 6), 1631–1646. doi: 10.1093/BRAIN/110.6.1631
Vann, S. D., and Aggleton, J. P. (2002). Extensive cytotoxic lesions of the rat retrosplenial cortex reveal consistent deficits on tasks that tax allocentric spatial memory. Behav. Neurosci. 116, 85–94. doi: 10.1037/0735-7044.116.1.85
Vann, S. D., and Aggleton, J. P. (2004). Testing the importance of the retrosplenial guidance system: effects of different sized retrosplenial cortex lesions on heading direction and spatial working memory. Behav. Brain Res. 155, 97–108. doi: 10.1016/j.bbr.2004.04.005
Keywords: posterior cingulate cortex, inhibitory avoidance, methyllycaconitine, long-term memory, cholinergic system, Alzheimer’s disease, encoding, retrieval
Citation: Pastor V and Katche C (2024) Dual role of α7 nicotinic acetylcholine receptors in the retrosplenial cortex for aversive memory acquisition and retrieval. Front. Behav. Neurosci. 18:1359729. doi: 10.3389/fnbeh.2024.1359729
Received: 21 December 2023; Accepted: 15 January 2024;
Published: 26 January 2024.
Edited by:
Travis P. Todd, University of Vermont, United StatesReviewed by:
John T. Green, University of Vermont, United StatesCopyright © 2024 Pastor and Katche. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Verónica Pastor, dmVycGFzdG9yQGZtZWQudWJhLmFy; Cynthia Katche, Y2thdGNoZUBmbWVkLnViYS5hcg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.