
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Behav. Neurosci., 04 November 2021
Sec. Learning and Memory
Volume 15 - 2021 | https://doi.org/10.3389/fnbeh.2021.768694
This article is part of the Research TopicBehavior-Driven Changes in Gene ExpressionView all 9 articles
 Amanda Anqueira-GonzálezJenny P. Acevedo-GonzalezAirined Montes-Mercado
Amanda Anqueira-GonzálezJenny P. Acevedo-GonzalezAirined Montes-Mercado Claudia Irizarry-HernándezNicolás L. Fuenzalida-Uribe
Claudia Irizarry-HernándezNicolás L. Fuenzalida-Uribe Alfredo Ghezzi*
Alfredo Ghezzi*When presented with the choice, Drosophila melanogaster females will often prefer to lay eggs on food containing a significant amount of alcohol. While, in some cases, this behavioral decision can provide a survival advantage to the developing larvae, it can also lead to developmental and cognitive problems. Alcohol consumption can affect executive functions, episodic memory, and other brain function capacities. However, in the fruit fly, the initial cognitive effects of alcohol consumption have been shown to reverse upon persistent exposure to alcohol. Using an olfactory conditioning assay where an odorant is implemented as a conditioned stimulus and paired with a heat shock as an unconditioned stimulus, a previous study has shown that when exposed to a short acute dose of alcohol, Drosophila larvae can no longer learn this association. Interestingly, upon prolonged chronic alcohol exposure, larvae seem to successfully avoid the conditioned stimulus just as well as control alcohol-naive larvae, suggestive of alcohol-induced neuroadaptations. However, the mechanisms by which Drosophila adapt to the presence of alcohol remains unknown. In this study, we explore the transcriptional correlates of neuroadaptation in Drosophila larvae exposed to chronic alcohol to understand the genetic and cellular components responsible for this adaptation. For this, we employed RNA sequencing technology to evaluate differences in gene expression in the brain of larvae chronically exposed to alcohol. Our results suggest that alcohol-induced neuroadaptations are modulated by a diverse array of synaptic genes within the larval brain through a series of epigenetic modulators.
The fruit fly Drosophila melanogaster has been used for over a hundred years as a model organism to further our understanding of various biological processes. More recently, this versatile model system has also provided insights into the behavioral genetics of alcohol use disorders since its alcohol responses closely resemble mammalian responses (reviewed in Rothenfluh et al., 2014). However, it is important to note that Drosophila melanogaster has an intimate relationship with alcohol. They naturally feed on decaying fruits, which contain varying concentrations of alcohol—a by-product of fermentation. Consumption of food with low to moderate levels of alcohol [(alcohol) < 4%) can result in increased larval fitness, while higher concentrations [(alcohol) > 4%] can cause significant developmental delays and cognitive dysfunction (McKenzie and Parsons, 1972; Robinson et al., 2012b; Schumann et al., 2021). Thus, it is not surprising that adult D. melanogaster females prefer to lay eggs on foods containing low to moderate alcohol concentrations while avoiding concentrations higher than 5% (Kacsoh et al., 2013). Interestingly, when threatened by parasitic wasps or in certain spatial arrangements, female flies will often choose to lay on food that surpasses concentrations of 10% alcohol by volume (Kacsoh et al., 2013; Sumethasorn and Turner, 2016).
Drosophila larvae can adapt to the presence of alcohol in the food and minimize some of its detrimental effects. By employing an olfactory heat shock conditioning assay, a previous study by Robinson et al. (2012a) showed that while Drosophila larvae acutely maintained on ethanol food for 1 h failed to establish memories after conditioning, larvae chronically exposed to alcohol over 6 days were capable of establishing a conditioned memory just as well as larvae that had not been exposed to alcohol (Robinson et al., 2012a). This study demonstrated that acute exposure to alcohol results in a deficiency in learning while chronic exposure does not present this deficiency. Consequently, this result indicates that the constant presence of alcohol induces a series of neuroadaptations that mitigate the effects of alcohol on learning.
Cognitive functions including attention, working memory, processing speed, visuospatial abilities, executive functions, impulsivity, learning, memory, and verbal fluency have all been shown to be impaired by alcohol dependence in humans (Stavro et al., 2013). While most of the cognitive repercussions of alcohol consumption are well-documented, the molecular and cellular mechanisms underlying interactions between alcohol neuroadaptations and cognitive impairments remain largely unknown. By employing the use of next-generation sequencing, here we evaluate larval brain differential gene expression between alcohol-adapted larvae and alcohol naive larvae to identify the genes whose expression changes because of these neuroadaptations.
Wild-type Drosophila melanogaster flies of the Canton S strain (CS) were used in all experiments. Flies were raised in standard cornmeal food at 25°C on a 12-h light/dark schedule. Chronic alcohol exposure was performed following the procedure described by Robinson et al. (2012a). Briefly, standard cornmeal food was supplemented with pure alcohol (Ethyl alcohol no. S73985A, Fisher Scientific) to obtain a 5% alcohol concentration in food (alcohol food). After melting the standard fly food for approximately 30 seconds, 2.65 mL of 95% alcohol was added to 50 mL of fly food just before re-solidification. Food was stirred to homogenize the solution and allowed to cool at room temperature for 24 h. After the 24 h were through, adult CS files were transferred to each bottle and allowed to lay eggs for 24 h. At the end of the 24 h, adult flies were discarded. The alcohol food was supplemented daily by pipetting 0.3 mL with 10% alcohol on the surface of the food until larvae reached the third instar stage, ∼5–6 days after eggs had been laid. For control larvae, the food was supplemented with water when melted instead of alcohol (control food). Control food was not supplemented daily.
Six days after egg-laying, as larvae began to crawl out of the food to proceed with pupae formation, individual third-instar larvae were collected using a needle, transferred to microcentrifuge tubes, and frozen at −80°C for 24–48 h. Freezing was followed by the addition of 200 μL of RNAlater-ICE Frozen Tissue Transition Solution (AM7030, Thermo Fisher Scientific) to larvae. Approximately 30 larval brains were dissected per replicate in RNAlater-ICE, for a total of three replicates (∼90 larval brains) for each group. Total RNA was extracted from larval brains using the EZ1 RNA Tissue mini kit (EZ1 RNA Tissue Mini Kit no. 959034, Qiagen). All samples were treated with DNase I (DNase Set no. 79254, Qiagen). RNA concentration and quality were evaluated using a Qubit 2.0 fluorometer (Life Technologies, United States) and Agilent 2100 Bioanalyzer (Agilent Technologies, United States).
Capture of mRNA (poly-A selection) and library preparation was carried out using the Illumina TruSeq® Stranded mRNA Library Prep kit from 300 ng of total RNA. Libraries were sequenced with an Illumina NextSeq 500/550 High Output Kit v2.5 for 80 cycles following the manufacturer’s protocol (Illumina, Inc.). At least 20 million reads were obtained for each replicate (a pool of 30 brains). The generated sequencing data was stored in FASTQ format and, the quality of the sequencing reads was assessed using FastQC 0.11.9. Adapters present in the sequences were eliminated using Trimmomatic 0.39. Tools from the “new tuxedo suite” were used to process the reads following the protocol by Pertea et al. (2016). Briefly, sequence alignment to the Drosophila dm6 genome assembly was completed using HISAT 2.2.0; SamTools was used to compress, sort, and index transcripts, while merging and quantifying transcripts was done using StringTie 2.1.4. Calculation of differential expression was achieved using Ballgown (Pertea et al., 2016) and visualized using R (Frazee et al., 2015). To remove low abundance genes, we applied a variance filter to remove all transcripts with a variance across samples of <1. No rRNA or ncRNA filtering was performed. For gene ontology (GO) annotation search and enrichment analysis, significant gene categories for differentially expressed genes were identified using DAVID web-accessible version 6.8 using default parameters and official gene symbols as input (Huang da et al., 2009).
RNA-seq analysis revealed significant changes in the expression of mRNA transcripts between control (alcohol-naive) larvae and the alcohol-grown larvae. Specifically, a total of 216 transcripts were found to be differentially expressed with a p-value < 0.05 and a fold change cutoff of 2-fold (log2 > 1). Of these, 113 transcripts were upregulated, while 103 transcripts were downregulated. Differentially expressed transcripts were defined as such using a fold change cutoff of 2-fold (log2 fold change > 1) and a P-value < 0.5. The distribution of these transcripts within these categories is shown in Figure 1.
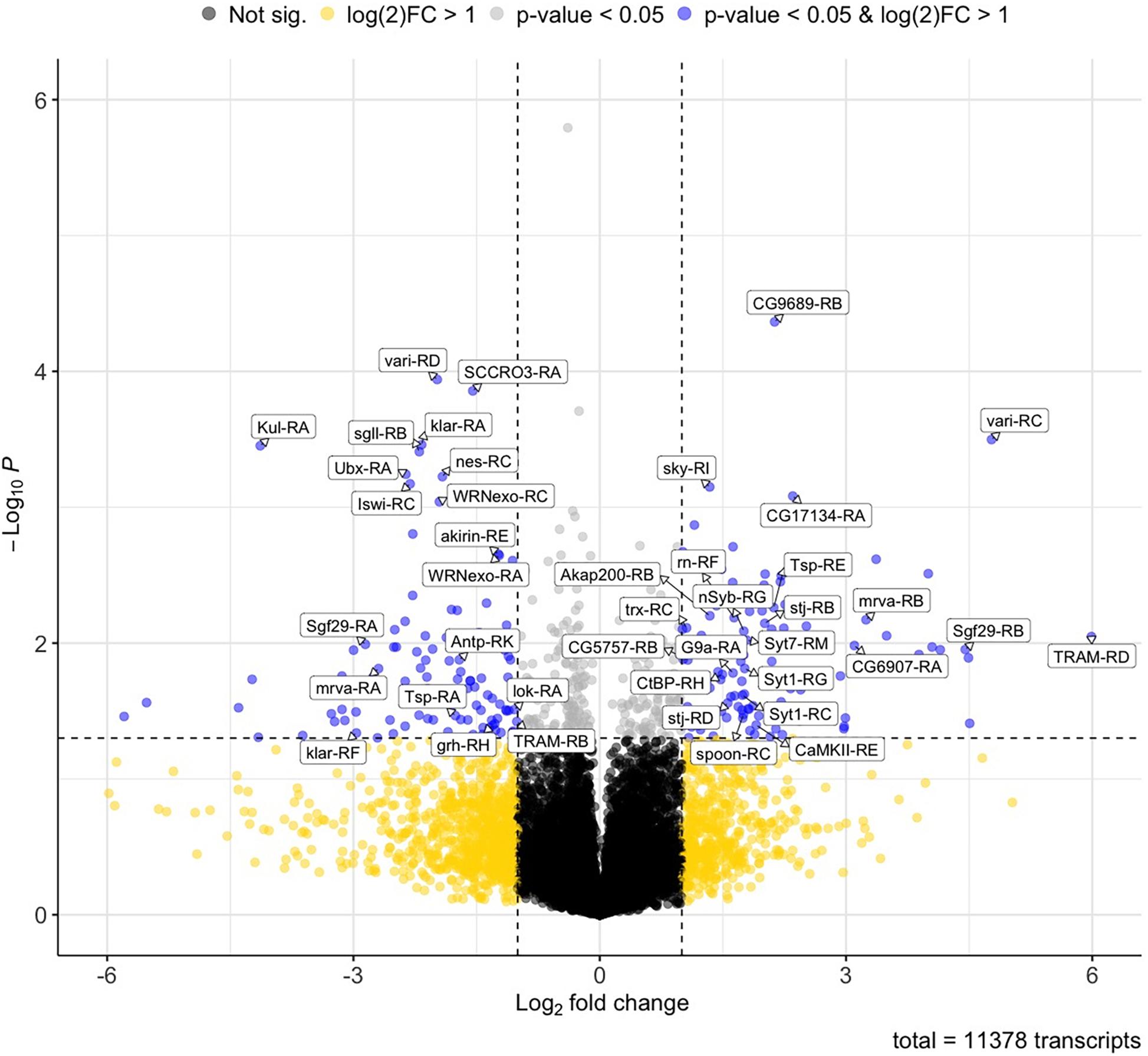
Figure 1. Volcano plot of differentially expressed transcripts induced by chronic alcohol exposure. Volcano plots showing fold change of differential transcript expression [log2 fold change] compared to the inverse of statistical significance [−log10 (P-value)]. Points in black or yellow are transcripts with no significant changes in expression (P > 0.05). Points in gray and blue correspond to transcripts with significant changes in expression (P < 0.05). Points in blue correspond to transcripts with significant changes in expression (P < 0.05) and that display a fold change higher than 2-fold (log2 fold change > 1). Names of genes encoding representative transcripts are labeled in the plot.
A total of 201 genes were represented in the 216 differentially expressed transcripts, with a total of 15 genes that were represented by two distinct differently expressed transcriptional isoforms. These genes are CG18649, corto, Eno, Fbp1, Gyg, klar, mrva, Sgf29, stj, Syt1, TRAM, trol, Tsp, vari, and WRNexo. Furthermore, it was particularly surprising to observe that of the 15 doubly represented genes, 5 genes had transcripts with opposing alcohol-induced effects, where 1 isoform was significantly upregulated by alcohol and the other significantly downregulated by alcohol, suggesting possible molecular switches. These genes are displayed in Table 1. A complete list of all differentially expressed transcripts, the represented genes is presented in Supplementary Table 1. Similarly, a complete list of FPKM counts for all samples tested is presented in Supplementary Table 2.
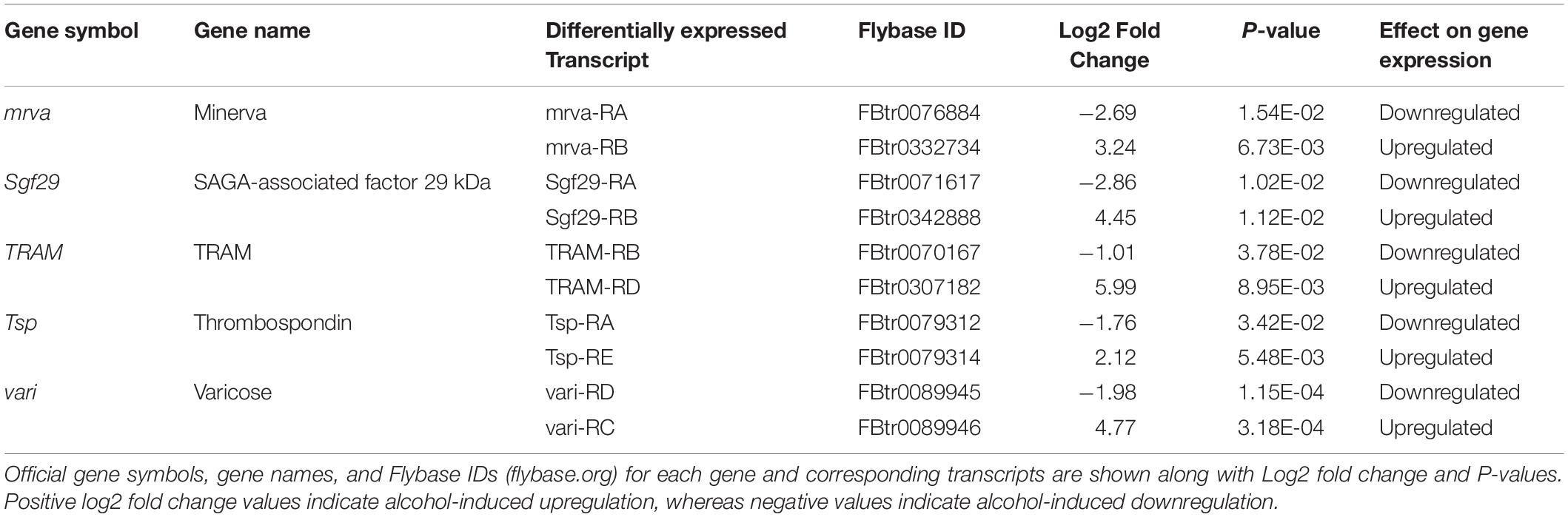
Table 1. Genes with transcripts showing opposing alcohol-induced effects.
It is important to note that none of the differentially expressed transcripts reach a q-value (FDR) < 0.05. Unfortunately, this is a common problem associated with the larger number of multiple corrections used in transcript-level analyses as compared with gene-level analyses. Analysis of individual transcripts results in more than 34,000 multiple corrections, as compared to gene-level analysis, where only there are approximately 16,000 multiple corrections. However, we believe that examination at the transcript level is particularly powerful as it detects changes that affect single transcriptional isoforms instead of whole gene transcription. This is important since most Drosophila genes can produce up to 10 distinct transcripts and 5 different protein products (Brown et al., 2014), and thus important changes can be obscured by opposing transcriptional responses of different transcripts within the same gene. Therefore, we proceeded with a statistical assessment of differential transcript usage of a selected list of implicated targets (Figure 2). This analysis reveals several genes where expression of specific transcriptional isoforms within a gene are significantly changed, while others remain unchanged. This was the case of the genes TRAM, klar, Syt1, Syt7, and trol. In other cases, however, as described before, some transcriptional isoforms were significantly upregulated by alcohol, while other transcriptional isoforms within the same gene were significantly downregulated. As shown in Figure 2, this was the case of the genes Sgf29, vari, and Kul, where alcohol differentially affects distinct transcriptional isoforms.
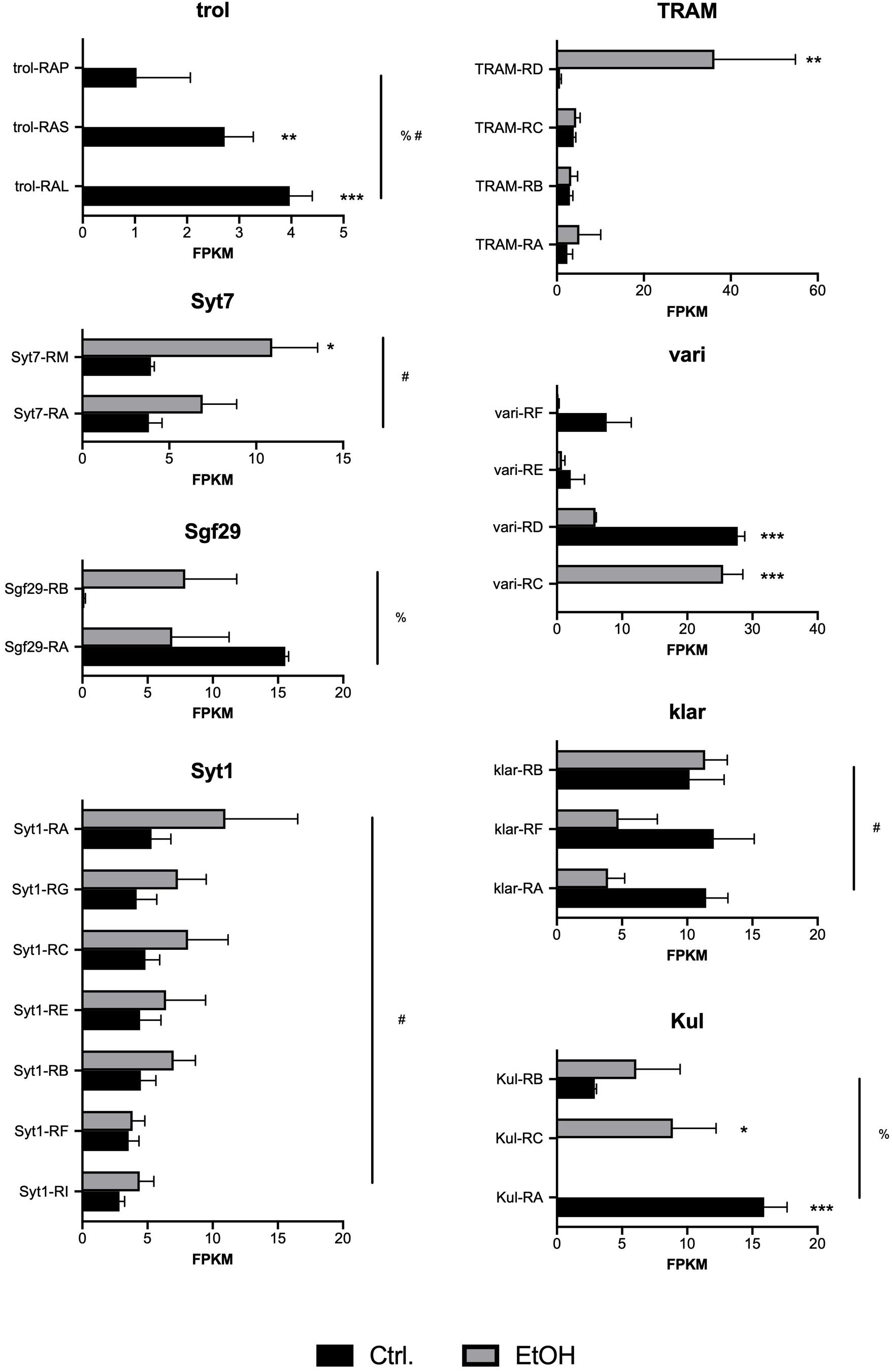
Figure 2. Transcript-specific expression induced by chronic alcohol exposure. Shown are FPKM counts in control (Ctrl, black bars) and alcohol treated (EtOH, gray bars) larvae for independent transcriptional isoforms of eight selected genes: trol, Syt7, Sgf29, Syt1, TRAM, vari, klar, and Kul. Significant differences were detected by Two-way ANOVA followed by Sidak’s multiple comparisons test. Asterisks denote significant differences between control and alcohol treated samples (*p < 0.05, **p < 0.01, ***p < 0.001). The pound sign (#) denotes a significant effect of treatment on all transcripts within the gene (p < 0.05). The percent (%) sign denotes a significant interaction between treatment and transcript identity (p < 0.05).
Finally, to identify potential biological pathways involved in the neuroadaptation to alcohol, we performed GO enrichment analysis of the differentially expressed genes. This analysis was completed using the Database for Annotation, Visualization and Integrated Discovery (DAVID) tool (Huang da et al., 2009). GO term enrichment analysis among the upregulated transcripts revealed important biological processes involved (Table 2). The most significantly enriched GO term was “histone H3 acetylation,” with a fold enrichment of 20.8. The genes associated with this term include CG6907, Sgf29, and trx, which are well-known chromatin remodelers involved in the positive regulation of transcription and biological development.

Table 2. Overrepresented gene ontology (GO) annotations for alcohol-upregulated genes.
Interestingly, within the most significant GO terms enriched amid upregulated genes, we also found several terms associated with the regulation of synaptic transmission. Among these, we find “regulation of calcium ion-dependent exocytosis” (represented by Sec23 and Ank), “calcium ion-regulated exocytosis of neurotransmitter” (represented by Syt1 and Syt7), “chemical synaptic transmission” (represented by CaMKII, sky, and Syt1), and “neurotransmitter secretion” (represented by Syt1, nSyb, and Syt7). This suggests that alcohol may be causing dysregulation of chemical synaptic transmission in neurons across the brain, a phenomenon known to occur when individuals are continuously exposed to alcohol (Kyzar and Pandey, 2015). Other important categories also represented include “long-term memory” (represented by CaMKII, spoon, and G9a) and “sensory perception of pain” (represented by CtBP, CaMKII, Akap200, stj, trx, rn, TRAM, and CG5757). Not surprisingly, several genes are represented in different GO terms, suggesting that the terms belong to interconnected biological processes.
Within the down-regulated transcripts, GO term enrichment analysis also revealed important biological processes (Table 3). This time, the top enriched GO term was “specification of segmental identity, thorax,” with a fold enrichment of 91.63. The genes associated with this term include Antp and Ubx, two well-studied genes involved in biological development. In fact, many other GO terms were also associated with biological development and cell fate specification, suggesting a systematic downregulation of development. On the other hand, we again observed genes important for “sensory perception of pain,” however this time, the genes represented were different (P5CDh1, dom, fok, Dad, UbcE2H, Gp150, CG4975, VhaPPA1-1, Unr, fax, Iswi, Vha100-1) with one exception: TRAM—this gene had one upregulated transcriptional isoform, and one downregulated transcriptional isoform. The fact that this GO-term appears in both the upregulated and downregulated gene cohorts suggests that the genes involved in this process serve as an important regulatory switch.
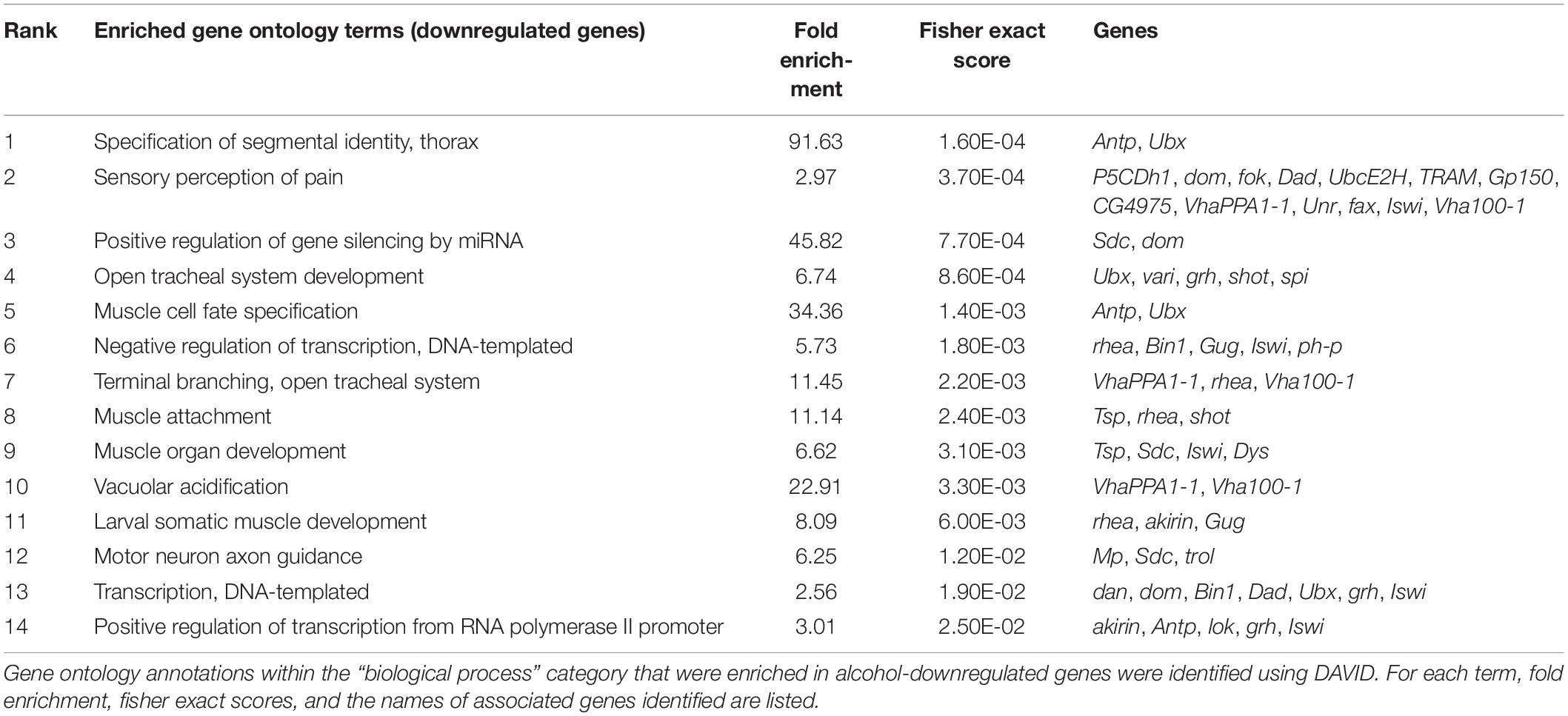
Table 3. Overrepresented gene ontology (GO) annotations for alcohol-downregulated genes.
RNA sequencing analysis of the effects of chronic alcohol exposure on the Drosophila larval brain showed many interesting differentially expressed genes. Our findings indicate that alcohol is altering different brain functions and disrupting several developmental pathways, many of them involved in higher-order processing. We find of particular interest the gene cohorts involved in the regulation of synaptic transmission, learning and memory, as well as those involved in the epigenetic control of developmental processes.
One of the most interesting gene cohorts seen in our data lies within the family of synaptic calcium-binding SNARE proteins, Synaptotagmins. This protein family is incredibly important in the process of vesicle trafficking and neurotransmitter release, both in Drosophila and mammals (Quiñones-Frías and Littleton, 2021). Our results show two Synaptotagmin paralogs, Syt1 and Syt7. Syt1 has been implicated in alcohol-related memory encoding in the Drosophila mushroom bodies (Petruccelli et al., 2020). Interestingly, Syt1 interacts closely with nSyb, a vesicle fusion protein also over-expressed in our data (Wang and Chapman, 2010), strengthening the notion that chronic alcohol affects synaptic integrity. In the case of Syt7, an interaction exists between said isoform and the synaptic protein Drep2 (Andlauer et al., 2014). This protein was present in our results, though not significantly regulated. Drep2 is known to be involved in learning and memory in Drosophila and is also known to colocalize with metabotropic glutamate receptors (Andlauer et al., 2014). Another study conducted in mice revealed Syt7’s presence in glutamatergic hippocampal neurons (Weber et al., 2014). During larval development, Syt7 is found in the neuronal cell bodies, which could explain the interaction with Drep2, a postsynaptic protein (Adolfsen et al., 2004; Andlauer et al., 2014). How these proteins interact with each other during the adult stages of the Drosophila, when Syt7 is localized in the synapse, requires further investigation (Guan et al., 2020). This interaction of the significantly over-expressed gene Syt7 with Drep2 is suggestive of an altered signaling pathway or mechanism as a result of chronic alcohol exposure.
Similarly, we found that many transcripts of genes involved in histone modifications and chromatin remodeling were altered as a result of chronic alcohol exposure. Epigenetic modifications are essential regulators of the transcriptional adaptation to drugs of abuse. In the past years, studies have addressed many epigenetic mechanisms important for modulating neural physiology as a response to alcohol (Ghezzi et al., 2013, 2017; Ramirez-Roman et al., 2018). Our results strengthen this idea and reveal many potential regulatory elements relevant to this process. As previously mentioned, we observed significant upregulation of genes involved in histone acetylation and methylation, including Sgf29, G9a, and trx. The protein Sgf29 is part of the Ada2/Gcn5/Ada3 transcription activator complex. However, little is known about its biological function (Petruk et al., 2006; Suganuma et al., 2008). On the other hand, G9a does not only function as a histone modification protein but is also involved in learning and memory, suggesting a tight link between the epigenetic reprogramming induced by alcohol and the modulation of higher-order processes in the brain (Kramer et al., 2011). Finally, Trx, a chromatin-modifying enzyme involved in gene regulation, is known to antagonize the epigenetic silencing by Polycomb group proteins and is known to contribute to axon guidance, eye development, and germ cell migration. Given that all these proteins are involved in histone modifications or chromatin remodeling, our results strengthen the idea that these mechanisms are of utter importance to cognitive alcohol neuroadaptations.
Another interesting phenomenon induced by chronic alcohol exposure identified in this study is the presence of potential transcriptional switches. For a subset of genes, differential transcript usage was triggered by alcohol, where one specific isoform was upregulated while another was downregulated. One particularly significant example of this phenomenon is observed in the gene vari, where the transcriptional isoform vari-RC is significantly induced by alcohol, while the isoform vari-RD is suppressed (Figure 2). Close inspection of the vari locus reveals that these two major transcript isoforms share seven exons encoding the core PDZ, SH3, HOOK, and GUK domains. However, the longer isoform (vari-RD) also encodes an N-terminal L27 protein-protein interaction domain that is absent from the shorter isoforms (vari-RC). The protein encoded by vari is a membrane-associated guanylate kinase (MAGUK) involved in tracheal system development and septate junction assembly (Wu et al., 2007). Vari proteins are known to interact with several proteins, including Fasciclin 3 (Wells et al., 2013), Semaphorin 1a (Jeong et al., 2017), Neurexin IV, and veli (Bachmann et al., 2008); these proteins are involved in ensheathing glial cells and neurons, axon guidance and synaptic growth at the neuromuscular junction, respectively. The differential expression of two Vari isoforms, which differ by the presence or absence of the L27 domain, is certainly intriguing. L27 domains can potentially stabilize interactions to different proteins by binding to the L27 domain of a corresponding partner. However, its role in alcohol neuroadaptation remains to be resolved.
While our study centers primarily on the transcriptional changes, additional experiments focusing on morphological and electrophysiological changes should offer new insights into the neuroadaptive mechanisms of chronic alcohol exposure. Nonetheless, this study facilitates our understanding of molecular and cellular dynamics of alcohol neuroadaptation in Drosophila larvae. Findings from this study, along with future research, could also lead to a better understanding of the effect of alcohol on learning and memory beyond the fruit fly and gain an insight into how addiction to this substance may contribute to cognitive decline and dependence in humans. Alcohol misuse is, in fact, one of the leading causes of death in the United States, with approximately 95,000 deaths occurring annually. According to the National Institute on Alcohol Abuse and Alcoholism, in 2018, 14.1 million adults of ages 18 and up suffered from alcoholism (NIAAA, 2020).
The data presented in this study are deposited in the NCBI Gene Expression Omnibus (GEO) data repository, accession number GSE185625.
AA-G and AG designed the study and wrote and reviewed the manuscript. AA-G and JA-G performed the alcohol treatments, larval brain extractions, and prepared samples for sequencing. CI-H and NF-U assisted in the larval brain dissections. AA-G, AM-M, and AG performed the bioinformatic analysis. All authors contributed to the article and approved the submitted version.
This study was funded by the NIH Blueprint initiative “Enhancing Neuroscience Diversity through Undergraduate Research Education Experiences (NIH BP-ENDURE)” grant R25 NS080687, the NIH Research Initiative for Scientific Enhancement (NIH-RISE) grant 5R25 GM061151, the NIH Centers of Biomedical Research Excellence: Puerto Rico Center for Neuroplasticity (NIH-COBRE) grant P20 GM103642. This manuscript was also made possible with support from the Sequencing and Genomics Facility of the UPR Río Piedras & MSRC/UPR, funded by NIH/NIGMS grant P20GM103475.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We want to thank all members of the Ghezzi lab for their continuous input during the development of this study. We would like to especially thank Cristina Martinez Benito and the fellow students in the Undergraduate Scientific Writing course of the Department of Biology at the University of Puerto Rico at Río Piedras for their guidance and input in the development of this manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnbeh.2021.768694/full#supplementary-material
Supplementary Table 1 | Complete annotated list of differentially expressed transcripts after chronic alcohol exposure.
Supplementary Table 2 | Complete FPKM counts for each replicate sample.
Adolfsen, B., Saraswati, S., Yoshihara, M., and Littleton, J. T. (2004). Synaptotagmins are trafficked to distinct subcellular domains including the postsynaptic compartment. J. Cell Biol. 166, 249–260. doi: 10.1083/jcb.200312054
Andlauer, T. F., Scholz-Kornehl, S., Tian, R., Kirchner, M., Babikir, H. A., Depner, H., et al. (2014). Drep-2 is a novel synaptic protein important for learning and memory. Elife 3:e03895. doi: 10.7554/eLife.03895
Bachmann, A., Draga, M., Grawe, F., and Knust, E. (2008). On the role of the MAGUK proteins encoded by Drosophila varicose during embryonic and postembryonic development. BMC Dev. Biol. 8:55. doi: 10.1186/1471-213X-8-55
Brown, J. B., Boley, N., Eisman, R., May, G. E., Stoiber, M. H., Duff, M. O., et al. (2014). Diversity and dynamics of the Drosophila transcriptome. Nature 512, 393–399.
Frazee, A. C., Pertea, G., Jaffe, A. E., Langmead, B., Salzberg, S. L., and Leek, J. T. (2015). Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 33, 243–246. doi: 10.1038/nbt.3172
Ghezzi, A., Krishnan, H. R., Lew, L., Prado, F. J., Ong, D. S., and Atkinson, N. S. (2013). Alcohol-induced histone acetylation reveals a gene network involved in alcohol tolerance. PLoS Genet. 9:e1003986. doi: 10.1371/journal.pgen.1003986
Ghezzi, A., Li, X., Lew, L. K., Wijesekera, T. P., and Atkinson, N. S. (2017). Alcohol-induced neuroadaptation is orchestrated by the histone acetyltransferase CBP. Front. Mol. Neurosci. 10:103. doi: 10.3389/fnmol.2017.00103
Guan, Z., Quinones-Frias, M. C., Akbergenova, Y., and Littleton, J. T. (2020). Drosophila Synaptotagmin 7 negatively regulates synaptic vesicle release and replenishment in a dosage-dependent manner. Elife 9:e55443. doi: 10.7554/eLife.55443
Huang da, W., Sherman, B. T., and Lempicki, R. A. (2009). Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57.
Jeong, S., Yang, D. S., Hong, Y. G., Mitchell, S. P., Brown, M. P., and Kolodkin, A. L. (2017). Varicose and cheerio collaborate with pebble to mediate semaphorin-1a reverse signaling in Drosophila. Proc. Natl. Acad. Sci. USA 114, E8254–E8263. doi: 10.1073/pnas.1713010114
Kacsoh, B. Z., Lynch, Z. R., Mortimer, N. T., and Schlenke, T. A. (2013). Fruit flies medicate offspring after seeing parasites. Science 339, 947–950. doi: 10.1126/science.1229625
Kramer, J. M., Kochinke, K., Oortveld, M. A., Marks, H., Kramer, D., de Jong, E. K., et al. (2011). Epigenetic regulation of learning and memory by Drosophila EHMT/G9a. PLoS Biol. 9:e1000569. doi: 10.1371/journal.pbio.1000569
Kyzar, E. J., and Pandey, S. C. (2015). Molecular mechanisms of synaptic remodeling in alcoholism. Neurosci. Lett. 601, 11–19.
McKenzie, J. A., and Parsons, P. A. (1972). Alcohol tolerance: An ecological parameter in the relative success of Drosophila melanogaster and Drosophila simulans. Oecologia 10, 373–388. doi: 10.1007/BF00345738
Pertea, M., Kim, D., Pertea, G. M., Leek, J. T., and Salzberg, S. L. (2016). Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 11, 1650–1667. doi: 10.1038/nprot.2016.095
Petruccelli, E., Brown, T., Waterman, A., Ledru, N., and Kaun, K. R. (2020). Alcohol Causes Lasting Differential Transcription in Drosophila Mushroom Body Neurons. Genetics 215, 103–116. doi: 10.1534/genetics.120.303101
Petruk, S., Sedkov, Y., Riley, K. M., Hodgson, J., Schweisguth, F., Hirose, S., et al. (2006). Transcription of bxd noncoding RNAs promoted by trithorax represses Ubx in cis by transcriptional interference. Cell 127, 1209–1221. doi: 10.1016/j.cell.2006.10.039
Quiñones-Frías, M. C., and Littleton, J. T. (2021). Function of Drosophila Synaptotagmins in membrane trafficking at synapses. Cell Mol. Life Sci. 78, 4335–4364.
Ramirez-Roman, M. E., Billini, C. E., and Ghezzi, A. (2018). Epigenetic mechanisms of alcohol neuroadaptation: insights from drosophila. J. Exp. Neurosci. 12:1179069518779809. doi: 10.1177/1179069518779809
Robinson, B. G., Khurana, S., Pohl, J. B., Li, W. K., Ghezzi, A., Cady, A. M., et al. (2012b). A low concentration of ethanol impairs learning but not motor and sensory behavior in Drosophila larvae. PLoS One 7: e37394.
Robinson, B. G., Khurana, S., Kuperman, A., and Atkinson, N. S. (2012a). Neural adaptation leads to cognitive ethanol dependence. Curr. Biol. 22, 2338–2341. doi: 10.1016/j.cub.2012.10.038
Rothenfluh, A., Troutwine, B. R., Ghezzi, A., and Atkinson, N. S. (2014). “The genetics of alcohol responses of invertebrate model systems,” in Neurobiology of alcohol dependence, eds A. B. C. Noronha, C. Cui, R. A. Harris, and J. C. Crabbe (Amsterdam: Elsevier), 463–491.
Schumann, I., Berger, M., Nowag, N., Schafer, Y., Saumweber, J., Scholz, H., et al. (2021). Ethanol-guided behavior in Drosophila larvae. Sci. Rep. 11:12307. doi: 10.1038/s41598-021-91677-3
Stavro, K., Pelletier, J., and Potvin, S. (2013). Widespread and sustained cognitive deficits in alcoholism: a meta-analysis. Addict. Biol. 18, 203–213.
Suganuma, T., Gutiérrez, J. L., Li, B., Florens, L., Swanson, S. K., Washburn, M. P., et al. (2008). ATAC is a double histone acetyltransferase complex that stimulates nucleosome sliding. Nat. Struct. Mol. Biol. 15, 364–372. doi: 10.1038/nsmb.1397
Sumethasorn, M., and Turner, T. L. (2016). Oviposition preferences for ethanol depend on spatial arrangement and differ dramatically among closely related Drosophila species. Biol. Open 5, 1642–1647. doi: 10.1242/bio.019380
Wang, Z., and Chapman, E. R. (2010). Rat and Drosophila synaptotagmin 4 have opposite effects during SNARE-catalyzed membrane fusion. J. Biol. Chem. 285, 30759–30766. doi: 10.1074/jbc.M110.137745
Weber, J. P., Toft-Bertelsen, T. L., Mohrmann, R., Delgado-Martinez, I., and Sørensen, J. B. (2014). Synaptotagmin-7 is an asynchronous calcium sensor for synaptic transmission in neurons expressing SNAP-23. PLoS One 9:e114033. doi: 10.1371/journal.pone.0114033
Wells, R. E., Barry, J. D., Warrington, S. J., Cuhlmann, S., Evans, P., Huber, W., et al. (2013). Control of tissue morphology by Fasciclin III-mediated intercellular adhesion. Development 140, 3858–3868. doi: 10.1242/dev.096214
Keywords: neuroadaptation, alcohol use disorder, Drosophila melanogaster, cognition, differential gene expression, RNA-seq
Citation: Anqueira-González A, Acevedo-Gonzalez JP, Montes-Mercado A, Irizarry-Hernández C, Fuenzalida-Uribe NL and Ghezzi A (2021) Transcriptional Correlates of Chronic Alcohol Neuroadaptation in Drosophila Larvae. Front. Behav. Neurosci. 15:768694. doi: 10.3389/fnbeh.2021.768694
Received: 01 September 2021; Accepted: 15 October 2021;
Published: 04 November 2021.
Edited by:
Alfonso Martin-Peña, University of Florida, United StatesReviewed by:
Katrin Vogt, University of Konstanz, GermanyCopyright © 2021 Anqueira-González, Acevedo-Gonzalez, Montes-Mercado, Irizarry-Hernández, Fuenzalida-Uribe and Ghezzi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alfredo Ghezzi, YWxmcmVkby5naGV6emlAdXByLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.