
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Neuroanat. , 16 October 2024
Volume 18 - 2024 | https://doi.org/10.3389/fnana.2024.1465421
This article is part of the Research Topic Updates and Discussions About Basal Ganglia and Their Circuits View all 7 articles
 Alessandra Marinho Miranda Lucena1,2
Alessandra Marinho Miranda Lucena1,2 Eudes Euler de Souza Lucena2*Sebastião Pacheco Duque Neto2Leonardo Thiago Duarte Barreto Nobre1,2
Eudes Euler de Souza Lucena2*Sebastião Pacheco Duque Neto2Leonardo Thiago Duarte Barreto Nobre1,2 Hugo Alexandre Oliveira Rocha1Rafael Barros Gomes Câmara1,2
Hugo Alexandre Oliveira Rocha1Rafael Barros Gomes Câmara1,2The objective of this review was to verify the therapeutic effect of polysaccharides derived from algae in neurodegenerative disease models involving the basal ganglia. To achieve this goal, a literature search was conducted in PubMed, Science Direct, Scopus, Web of Science, Embase, and Google Scholar databases. The descriptors “neuroprotective or neural regenerative or immunomodulatory activity or neuroprotection,” “polysaccharide or carbohydrate or carbohydrate polymers,” “marine algae or seaweed,” and “basal ganglia” according to the Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) methodology were used. This methodology involved the steps of searching, pre-selection, and inclusion of articles. A total of 737 records were identified. Following the data analysis, 698 studies were excluded, resulting in a final sample of 8 studies. Species such as Turbinaria decurrens, Gracilaria cornea, Chlorella pyrenoidosa, Arthrospira (Spirulina) platensis, Fucus vesiculosus, and Laminaria japonica have demonstrated significant neuroprotective effects. This review suggests that polysaccharides derived from marine algae possess therapeutic potential for neuroprotection, modulation of inflammation, and amelioration of functional deficits. Their use in neurodegenerative disease models warrants further consideration.
Basal ganglia (BG) or basal nucleus as formally named by International Neuroanatomical Terminology (INT) (FIPAT, 2017) are complex neuroanatomical and hodological subcortical structures involved in both motor and “non-motor” functions. The neuroanatomical topography of the basal nucleus is composed of (1) the striatum (subdivided into the caudate nucleus, putamen, and nucleus accumbens), (2) the globus pallidus lateral, (3) the globus pallidus medial, (4) the subthalamic nucleus, and (5) the substantia nigra (Rocha et al., 2023).
The functional organization of basal nuclei circuitry includes dopaminergic, GABAergic, and glutamatergic intrinsic and extrinsic projections (afferent and efferent), in both direct and indirect pathways (Simonyan, 2019). The three most important dopaminergic mesencephalic ascendent systems are the nigrostriatal, mesolimbic, and mesocortical or mesocorticolimbic pathways. Dysfunction in these pathways can lead to conditions such as Parkinson’s disease (PD) and Huntington’s disease (HD) (Andres and Darbin, 2018; Obeso et al., 2014).
In HD, early pathological events such as an increase in the production and release of inflammatory mediators (e.g., IL-6, IL-8, and TNF-ɑ) can be observed (Jia et al., 2022). In PD, oxidative stress following mitochondrial damage due to environmental risk factors or chronic microglial inflammation mediates the activation of ɑ-synuclein-rich protein aggregates in the form of Lewy bodies, which are also present in the autosomal dominant form of PD (Vázquez-Vélez and Zoghbi, 2021). The overexpression of ɑ-synuclein, in turn, leads to mitochondrial death (Vázquez-Vélez and Zoghbi, 2021). Excitotoxicity, which is also related to oxidative stress, causes several intracellular damages in microglia, leading to the constant expression of pro-inflammatory cytokines, morphological changes, and cell death (Jia et al., 2022).
Neurodegeneration is a multifaceted process that leads to neuronal dysfunction. It involves intricate intracellular and extracellular mechanisms operating through diverse biochemical pathways. When considering effective prevention and therapy for neurodegenerative diseases, it is essential to consider all of these interconnected factors. Several studies have shown that neuroprotective agents from natural sources play an important role in the prevention and treatment of neurodegenerative diseases (NDDs). Among the molecules studied, polysaccharides extracted from plants, fungi, and algae are noteworthy (Dhahri et al., 2021; Wang et al., 2022). Natural polysaccharides that are capable of preventing or interfering with the initial neurodegenerative processes hold promise for preventive interventions.
Due to their multiple biological activities, especially antioxidant and anti-inflammatory properties, and their low risk of toxicity and adverse effects (Xu et al., 2022), polysaccharides have great potential for the prevention and treatment of NDD. Additionally, these molecules could reverse neurodegeneration and improve cognitive function, learning, memory, and motor skills, which could be valuable for therapeutic purposes (Dhahri et al., 2021).
Notably, algae have emerged as a rich source of novel chemical compounds with significant biological effects, having been used for nutritional and medicinal purposes for many years (Silva et al., 2023). The cultivation and consumption of marine macroalgae are expanding widely and proving to be safe and sustainable in several aspects, enabling the biotechnological development of economically responsible therapeutic alternatives (Sugumaran et al., 2022).
Therefore, this study aims to systematically review original articles that investigated the use of polysaccharides from marine algae in the treatment of neural damage and diseases associated with the basal ganglia.
The present study comprises a systematic review of the literature in order to gather and analyze studies concerning the effects of polysaccharides from marine algae on experimental animal models of neuroprotection and neuroregenerative aspects involving the basal ganglia. The approach was developed in accordance with well-defined stages, including search, identification, selection, and eligibility strategies. For database searches, we used specific terminologies, filters, and descriptors relevant to articles published in six databases, namely PubMed, Science Direct, Scopus, Web of Science, Embase, and Google Scholar.
The selected terms were based on the technical vocabulary commonly used to index articles in the field of health sciences. These terms align with the Medical Subject Headings (MeSH) provided by the US National Library of Medicine (NLM) for accurate descriptor-based searching: “neuroprotective or neural regenerative or immunomodulatory activity or anti-inflammatory or neuroprotection” and “polysaccharide or carbohydrate or carbohydrate polymers” and “marine algae or seaweed” and “Basal ganglia” or “Basal nuclei” or “striatum” or “caudate nucleus” or “putamen” or “nucleus accumbens” or “globus pallidus” or “subthalamic nucleus” or “substantia nigra.” The combinations of these keywords were used to search for all studies that analyzed the effects of polysaccharides from marine algae on experimental models of both neurological disorders and NDD. This review was conducted following the recommendations of the Preferred Reporting Items for Systematic reviews and Meta-Analyses (PRISMA) (Page et al., 2021). There were no disagreements regarding the selection of the sample, and there were no temporal limitations. Figure 1 shows the search process.
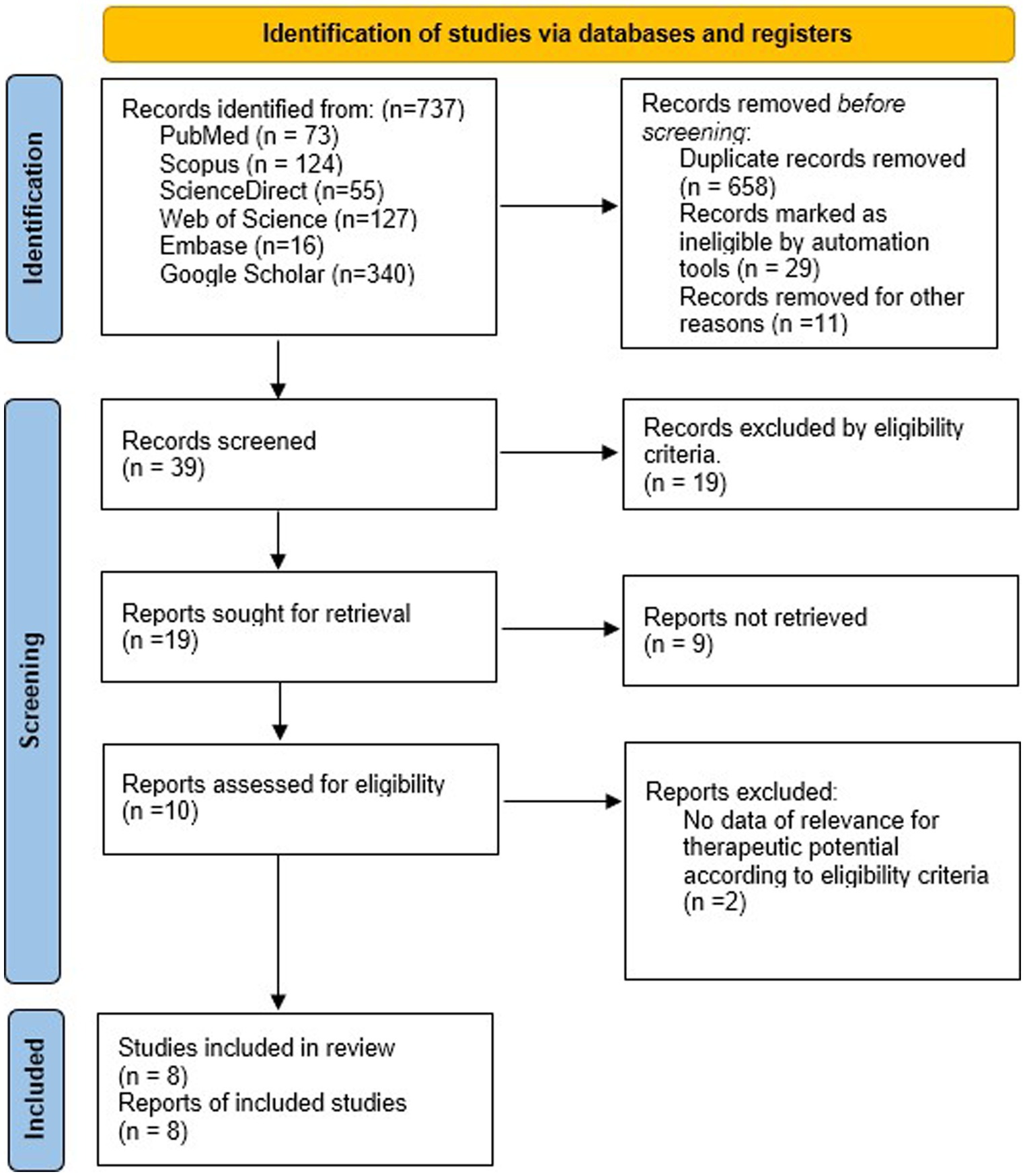
Figure 1. Synthesis of the search process according to the PRISMA model.
In our study, two expert evaluators independently conducted a systematic search for relevant articles. They screened the titles and abstracts and resolved any discrepancies through subsequent consensus meetings. Articles were limited to those published in English. After initial screening, full-length publications were read and compared. We included experimental studies that examined the effects of polysaccharides in vivo, focusing on specific pathological conditions. Additionally, we considered studies that assessed behavioral improvements related to the pathology. The exclusion criteria encompassed non-original articles, studies using cell culture models, analyses combining neural effects with other compounds (without an isolated algae group), and the absence of a comparable control group.
In this systematic review, we have established rigorous eligibility criteria for the inclusion of studies, including experiments conducted on murine or rat models with induced neurological lesions, treatment administered either before or after the occurrence of damage or intervention, comprehensive analysis of the effects on injury and underlying action mechanisms, and the inclusion of animal models exhibiting clinical symptoms relevant to the specific pathologies under investigation. Additionally, a questionnaire was used to assess how well a manuscript complies with the ARRIVE Essential 10 (Percie du Sert et al., 2020). This questionnaire was utilized to describe comparative experiments conducted on living animals, considering the following aspects: study design, sample size, inclusion and exclusion criteria, randomization, blinding, outcomes measures, statistical methods, experimental animals, experimental procedures, and results.
However, the evaluated studies exhibited notable inconsistencies: the inducing agents used to simulate neurodegenerative diseases varied across studies, treatment durations were heterogeneous, and behavioral tests employed diverse methodologies. Neuronal damage simulation utilized different compounds, each acting through distinct mechanisms. Furthermore, these compounds were administered at different doses via different routes, further contributing to the study heterogeneity.
In this study, researchers initially collected 737 records. After rigorous data analysis, 698 studies were excluded based on specific criteria. Ultimately, eight studies fulfilled the inclusion and quality assessment criteria. Figure 1 shows the research flow. The selected articles underwent thorough examination, focusing on authorship, publication year, algae species, research methods, evaluation, and key findings (Table 1).
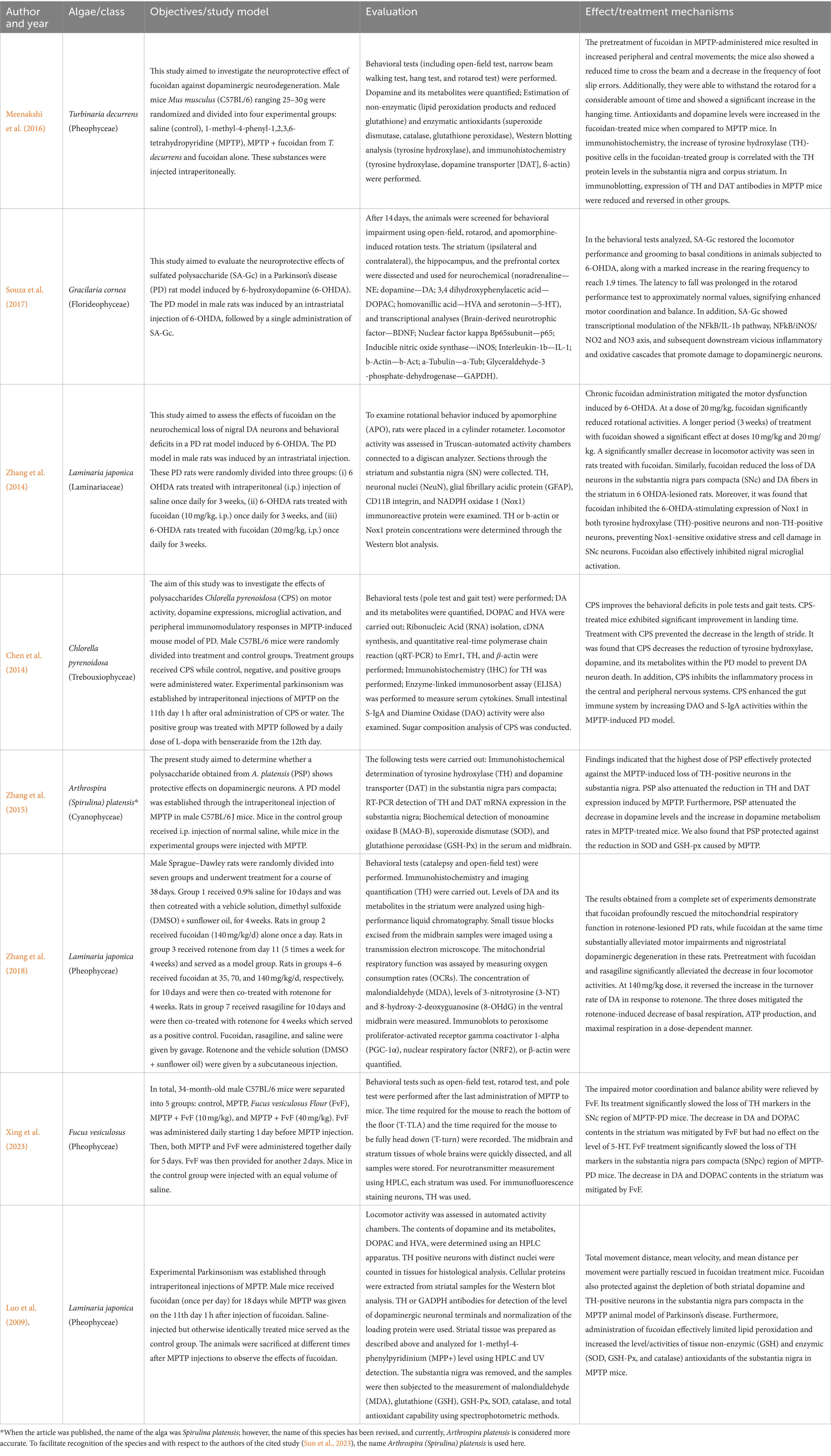
Table 1. Summary of selected studies in this review.
Neuroinflammatory and oxidative stress mechanisms are at the root of the pathogenesis of BG neurodegeneration prior to the onset of its conditions (Singh et al., 2019). Excessive levels of reactive oxygen species (ROS) can modulate the inflammatory response by inducing the expression of pro-inflammatory molecules (Singh et al., 2019). Conversely, the inflammatory response generates ROS through multiple pathways by signaling the pro-inflammatory transcription factor NFκβ (Singh and Singh, 2020). It appears that Nrf2 and the subunit of NFκβ P65 constitute a point of intersection between the antioxidant and inflammatory pathways (George et al., 2022). It is important to highlight that inflammatory molecules can cross the blood–brain barrier and move both toward the peripheral and central nervous systems (Jia et al., 2022).
The 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model replicates key pathobiochemical features of PD, including oxidative stress, mitochondrial dysfunction, excitotoxicity, inflammation, and apoptosis (Beal, 2021). Oxidative stress, driven by ROS, significantly contributes to PD pathogenesis, with mitochondria being the primary source of ROS. Polysaccharides have emerged as potential therapeutic agents against PD and Alzheimer’s disease (AD), acting through mechanisms involving neuroinflammation, oxidative stress, autophagy, apoptosis, and mitochondrial dysfunction. Similarly, HD is primarily related to oxidative stress and neurotoxicity (Wang et al., 2022). These shared biological processes play pivotal roles in the progression and pathogenesis of neurodegenerative diseases, prompting extensive research to identify therapeutic targets (Pradhan et al., 2020; Sun et al., 2023; Mohd Sairazi and Sirajudeen, 2020).
Polysaccharides, such as fucoidan, can act both directly by scavenging radicals, chelating metal ions, and donating electrons to unstable molecules and indirectly by increasing the expression, protein level, and activity of antioxidant enzymes (Dhahri et al., 2021).
According to Meenakshi et al. (2016), fucoidan from Turbinaria decurrens holds promise as a potential therapeutic agent for PD, impacting dopamine (DA) levels and providing antioxidative benefits. In the striatum of PD animals, DA levels and its metabolites were decreased due to dopaminergic neuron impairment. Pretreatment with this fucoidan effectively reversed the abnormally low levels of 3,4-dihydroxyphenylacetic acid (DOPAC), homovanillic acid (HVA), and DA induced by the action of MPTP. Mice demonstrated a reduction in the levels of thiobarbituric acid reactive substance (TBARS), a product of lipid peroxidation, and an increase in glutathione (GSH) levels and glutathione peroxidase (GPx) activity. As a result, these mice exhibited proper muscular coordination and retained their natural activity.
In their research, Zhang et al. (2014) investigated the effects of fucoidan from Laminaria japonica on PD by using a 6-hydroxydopamine (6-OHDA) model. This fucoidan significantly mitigated motor dysfunction and attenuated damage to the dopaminergic system both in the striatum and substantia nigra pars compacta induced by 6-OHDA. The loss of striatal tyrosine hydroxylase (TH-ir) immunoreactive fibers and TH-ir neurons in the central nervous system was associated with behavioral dysfunction, elevated levels of protein carbonylation, microglial activation, and increased expression of Nox1. Fuicodan was capable of suppressing oxidative stress and reducing neuronal damage caused by 6-OHDA by limiting the increase in Nox1. It holds promise as a potential therapeutic agent for PD, acting through mitochondrial support and modulation of antioxidant pathways. This polysaccharide is capable of protecting the dopaminergic pathway, which plays a critical role in motor control. Additionally, it ameliorates the behavioral deficits observed in PD animal models.
In animals exposed to rotenone, a toxin associated with PD pathology, another fuicodan from L. japonica exerted protective effects by enhancing mitochondrial function (Zhang et al., 2018). It reduces oxidative stress within the ventral midbrain in rotenone-treated rats. The study suggests that the upregulation of two key proteins—peroxisome proliferator-activated receptor-γ coactivator 1 alpha (PGC-1α) and nuclear respiratory factor (NRF2)—significantly contributes to the protective effects of Laminaria fucoidan. In the fuicodan treatment group, NRF2 exhibits increased expression, which correlates with enhanced antioxidant activity.
Using an MPTP-induced PD mouse model, fucoidan from Fucus vesiculosus demonstrated both neuroprotective and anti-PD effects (Xing et al., 2023). Using target fishing technology, the researchers have identified ATP5F1a, a component of mitochondrial complex V, as the primary target of this fuicodan, which plays a crucial role in reducing ROS. This protein mainly regulates adenosine triphosphate (ATP) production and is involved in cellular energy metabolism. Consequently, it is essential to further investigate the impact of polysaccharides on neuronal cell metabolism.
In the third study, researchers investigated the effects of fucoidan from L. japonica on MPTP-induced dopaminergic neuron loss (Luo et al., 2009). Notably, this fucoidan contributed to the increase in the levels of GPx, superoxide dismutase (SOD), catalase (CAT), and GSH in the brain homogenate of the substantia nigra. However, this fuicodan did not affect the pharmacokinetics of MPTP, indicating that its protective effect is not the result of an interaction with the neurotoxin. Furthermore, the increase in TH-positive cells, which are associated with dopaminergic neurons, observed in the fucoidan-treated group is correlated with TH protein levels in the substantia nigra. Notably, this increase exceeds the content of DA and DOPAC. One plausible explanation is that dopaminergic terminals are more sensitive to MPTP toxicity, resulting in more severe damage compared to dopaminergic cell bodies.
Research findings indicate that 6-OHDA increased IL-1β transcription in the rat striatum (Souza et al., 2017). However, treatment with a sulfated polysaccharide obtained from the alga G. cornea (SA-Gc) resulted in the modulation of the NFκβ/iNOS/NO2 and NO3 pathways (Souza et al., 2017). SA-Gc promoted the downregulation of pro-inflammatory genes (p65, iNOS, and IL-1β) to basal levels and the upregulation of the brain-derived neurotrophic factor (BDNF) gene. Furthermore, it enhanced GSH production in the striatum, hippocampus, and prefrontal cortex, and caused an improvement in motor functions of 6-OHDA exposed animals. The expression of p65 is associated with anti-inflammatory and antioxidant activity. This protein acts as a crosslink between inflammation and redox homeostasis (George et al., 2022).
Rats treated with SA-Gc exhibited improvements in locomotor disturbances and behavioral changes (Souza et al., 2017). Additionally, an apomorphine-induced rotation test, along with an analysis of striatal monoamine levels, revealed that SA-Gc-treated animals showed significant dose-dependent increases in DA and DOPAC contents. This suggests the integrity of dopaminergic neurons. Furthermore, SA-Gc administration led to weight gain in the hemi-parkinsonian rats, possibly indicating a neuroprotective effect on the maintenance of the dopaminergic pathway.
Polysaccharides isolated from microalgae have also been found to act as protective agents against damage to the central nervous system. Examples of these polysaccharides are those obtained from Chlorella pyrenoidosa (Chen et al., 2014) and Spirulina platensis (Zhang et al., 2015).
The polysaccharide of C. pyrenoidosa (CPS) appears to have anti-inflammatory effects and positively impacts the gut health of animals exposed to MPTP (Chen et al., 2014). Mice exposed to MPTP and treated with CPS exhibited a reduction in the levels of pro-inflammatory cytokines and a decrease in the rate of AD renewal in the striatum. Additionally, the effective reversal of the abnormal drop in serum diaminoxidase activity (DAO) and S-IgA levels in the small intestine was observed. For molecules that do not cross the blood–brain barrier, the mechanism that uses the brain-gut axis pathway appears to be very efficient. This is because SP regulates the gut microbiota and its metabolic products, exerting a neuroprotective effect (Wang et al., 2022).
In a study conducted by Zhang et al. (2015), it was observed that the polysaccharides of Spirulina platensis (PSP) had a neuroprotective effect on dopaminergic neurons in a mouse model of PD induced by MPTP. Researchers have found that the protective effect of PSP was not directly related to the inhibition of monoamine oxidase type B (MAO-B), as it did not significantly affect MAO-B activity. Instead, other mechanisms appeared to have been involved. Otherwise, PSP increased the activities of SOD and GPx in mice exposed to MPTP. These enzymes play a crucial role in antioxidant defense, suggesting that the PSP mechanism is related to the modulation of endogenous antioxidant tools and the consequent protection of dopaminergic neurons.
In summary, the species T. decurrens, G. cornea, L. japonica, C. pyrenoidosa, S. platensis, and F. vesiculosus showed significant neuroprotective effects against different neurotoxicity inducers (MPTP and 6-OHDA) through various routes of treatment administration, including intraperitoneal and intrastriatal injections, as well as oral administration. Various forms of evaluation have been employed, including behavior studies, Western blotting analysis, immunohistochemistry, flow cytometry analysis, transcriptional analysis, biochemical detection of enzymes, spectrophotometric methods, liquid chromatography, and quantification of neurotransmitters.
The mechanisms of action behind the neuroprotective benefits suggest that they can act on different targets: directly targeting ROS (Dhahri et al., 2021); enhancing GSH levels (Meenakshi et al., 2016); attenuating Nox1 expressing levels (Zhang et al., 2014); upregulating PGC-1α and NRF2 proteins (Zhang et al., 2018); improving mitochondrial respiratory function by ATP5F1a protein interaction (Xing et al., 2023); increasing the levels of GPx (Luo et al., 2009), SOD (Luo et al., 2009), CAT (Luo et al., 2009), and GSH (Meenakshi et al., 2016; Luo et al., 2009); modulating the transcription of NFκβ/iNOS/NO2 and NO3 pathways (Souza et al., 2017); modulating the immune response through the brain–gut axis (Chen et al., 2014); and increasing the activity of antioxidant enzymes such as GPx (Meenakshi et al., 2016; Zhang et al., 2015) and SOD (Zhang et al., 2015).
The limited number of studies analyzed showed a diversity of lesion models and evaluation methods. The possible mechanisms of protection for polysaccharides from algae in neural damage and diseases associated with the basal ganglia are speculative at this stage, necessitating further investigation.
The outcomes of this review highlight the potential therapeutic significance of algae-derived polysaccharides. These compounds show promise as neuroprotective agents in areas such as the modulation of inflammation and endogenous antioxidant pathways. They also promoted important amelioration in the morphophysiology of the nervous system under stress. Thus, they may hold value for the treatment of neurodegenerative diseases linked to the basal ganglia. Therefore, the search for neuroprotective polysaccharides in seaweed provides opportunities for researchers to evaluate the potential of these molecules as innovative systems for the prevention and treatment of neurodegenerative diseases. These molecules may also contribute to overcoming the limitations of current treatments.
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
AL: Writing – original draft, Writing – review & editing. ES: Writing – original draft, Writing – review & editing, Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization. SN: Writing – original draft, Writing – review & editing. LN: Writing – original draft, Writing – review & editing. HR: Writing – original draft, Writing – review & editing. RC: Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
BG, Basal ganglia; ROS, Reactive oxygen species; NFκβ, Nuclear factor kappa B; Nrf2, Nuclear factor erythroid 2-related factor 2; HD, Huntington’s disease; IL-6, Interleucin 6; IL-8, Interleucin 8; TNFα, Tumor necrosis factor-alpha; PD, Parkinson’s disease; NDD, Neurodegenerative diseases; MeSH, Medical Subject Headings; NLM, US National Library of Medicine; PRISMA, Preferred Reporting Items for Systematic reviews and Meta-Analyses; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; AD, Alzheimer’s disease; DA, Dopamine; DOPAC, 3,4-dihydroxyphenylacetic acid; HVA, Homovanillic acid; TBARS, Thiobarbituric acid reactive substance; GSH, Glutathione; GPx, Glutathione peroxidase; 6-OHDA, 6-Hydroxydopamine; TH-ir, Striatal tyrosine hydroxylase; Nox1, NADPH oxidase; BDNF, Brain-derived neurotrophic factor; CPS, Polysaccharide of Chlorella pyrenoidosa; PGC-1α, Peroxisome proliferator-activated receptor-γ coactivator 1 alpha; ATP, Adenosine triphosphate; SOD, Superoxide dismutase; CAT, Catalase; TH, Tyrosine hydroxylase; IL-1β, Interleucin 1 beta; SA-Gc, Sulfated polysaccharide from the alga Gracilaria cornea; iNOS, Inducible nitric oxide synthase; DAO, Diaminoxidase; S-IgA, Secretory immunoglobulin A; PSP, Polysaccharides of Spirulina platensis; MAO-B, Monoamine oxidase type B; SNpc, Substantia nigra pars compacta; ARRIVE, Animal Research: Reporting of In Vivo Experiments; GADPH, Glyceraldehyde-3-Phosphate Dehydrogenase; HPLC, High Performance Liquid Chromatography; SP, Sultated polysaccharide; UV, Ultraviolet; NADPH, Nicotinamide Adenine Dinucleotide Phosphate.
Andres, D. S., and Darbin, O. (2018). Complex dynamics in the basal ganglia: health and disease beyond the motor system. J. Neuropsychiatry Clin. Neurosci. 30, 101–114. doi: 10.1176/appi.neuropsych.17020039
Beal, M. (2021). Experimental models of Parkinson's disease. Nat. Rev. Neurosci. 2, 325–332. doi: 10.1038/35072550
Chen, P., Wang, H., Liu, Y., Lin, S., Chou, H. N., and Sheen, L. (2014). Immunomodulatory activities of polysaccharides from Chlorella pyrenoidosa in a mouse model of Parkinson's disease. J. Funct. Foods 11, 103–113. doi: 10.1016/j.jff.2014.08.019
Dhahri, M., Alghrably, M., Mohammed, H. A., Badshah, S. L., Noreen, N., Mouffouk, F., et al. (2021). Natural polysaccharides as preventive and therapeutic horizon for neurodegenerative diseases. Pharmaceutics. 14:1. doi: 10.3390/pharmaceutics14010001
George, M., Tharakan, M., Culberson, J., Reddy, A. P., and Reddy, P. H. (2022). Role of Nrf2 in aging, Alzheimer's and other neurodegenerative diseases. Ageing Res. Rev. 82:101756. doi: 10.1016/j.arr.2022.101756
Jia, Q., Li, S., Li, X.-J., and Yin, P. (2022). Neuroinflammation in Huntington’s disease: from animal models to clinical therapeutics. Front. Immunol. 13:1088124. doi: 10.3389/fimmu.2022.1088124
Luo, D., Zhang, Q., Wang, H., Cui, Y., Sun, Z., Yang, J., et al. (2009). Fucoidan protects against dopaminergic neuron death in vivo and in vitro. Eur. J. Pharmacol. 617, 33–40. doi: 10.1016/j.ejphar.2009.06.015
Meenakshi, S., Umayaparvathi, S., Saravanan, R., Manivasagam, T., and Balasubramanian, T. (2016). Neuroprotective effect of fucoidan from Turbinaria decurrens in MPTP intoxicated Parkinsonic mice. Int. J. Biol. Macromol. 86, 425–433. doi: 10.1016/j.ijbiomac.2015.12.025
Mohd Sairazi, N. S., and Sirajudeen, K. N. S. (2020). Natural products and their bioactive compounds: neuroprotective potentials against neurodegenerative diseases. Evid. Based Complement. Alternat. Med. 2020:6565396. doi: 10.1155/2020/6565396
Obeso, J. A., Rodriguez-Oroz, M. C., Stamelou, M., Bhatia, K. P., and Burn, D. J. (2014). The expanding universe of disorders of the basal ganglia. Lancet 384, 523–531. doi: 10.1016/S0140-6736(13)62418-6
Page, M. J., McKenzie, J. E., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., et al. (2021). The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 372:n71. doi: 10.1136/bmj.n71
Percie du Sert, N., Hurst, V., Ahluwalia, A., Alam, S., Avey, M. T., Baker, M., et al. (2020). The ARRIVE guidelines 2.0: updated guidelines for reporting animal research. BMC Vet. Res. 16:242. doi: 10.1186/s12917-020-02451-y
Pradhan, B., Patra, S., Nayak, R., Behera, C., Dash, S. R., Nayak, S., et al. (2020). Multifunctional role of fucoidan, sulfated polysaccharides in human health and disease: a journey under the sea in pursuit of potent therapeutic agents. Int. J. Biol. Macromol. 164, 4263–4278. doi: 10.1016/j.ijbiomac.2020.09.019
Rocha, G. S., Freire, M. A. M., Britto, A. M., Paiva, K. M., Oliveira, R. F., Fonseca, I. A. T., et al. (2023). Basal ganglia for beginners: the basic concepts you need to know and their role in movement control. Front. Syst. Neurosci. 17:1242929. doi: 10.3389/fnsys.2023.1242929
Silva, J., Alves, C., Soledade, F., Martins, A., Pinteus, S., Gaspar, H., et al. (2023). Marine-derived components: can they be a potential therapeutic approach to Parkinson’s disease? Mar. Drugs 21:451. doi: 10.3390/md21080451
Simonyan, K. (2019). Recent advances in understanding the role of the basal ganglia. F1000Res 8:F1000 Faculty Rev-122. doi: 10.12688/f1000research.16524.1.
Singh, A., Kukreti, R., Saso, L., and Kukreti, S. (2019). Oxidative stress: a key modulator in neurodegenerative diseases. Molecules 24:1583. doi: 10.3390/molecules24081583
Singh, S., and Singh, T. G. (2020). Role of nuclear factor kappa B (NF-κB) signalling in neurodegenerative diseases: an mechanistic approach. Curr. Neuropharmacol. 18, 918–935. doi: 10.2174/1570159X18666200207120949
Souza, R. B., Frota, A. F., Sousa, R. S., Cezario, N. A., Santos, T. B., Souza, L. M., et al. (2017). Neuroprotective effects of Sulphated Agaran from marine alga Gracilaria cornea in rat 6-Hydroxydopamine Parkinson's disease model: Behavioural, neurochemical and transcriptional alterations. Basic Clin. Pharmacol. Toxicol. 120, 159–170. doi: 10.1111/bcpt.12669
Sugumaran, R., Padam, B. S., Yong, W. T. L., Saallah, S., Ahmed, K., and Yusof, N. A. (2022). Retrospective review of global commercial seaweed production—current challenges, biosecurity and mitigation measures and prospects. Int. J. Environ. Res. Public Health 19:7087. doi: 10.3390/ijerph19127087
Sun, X., Liu, H., Tan, Z., Hou, Y., Pang, M., Chen, S., et al. (2023). Remodeling microenvironment for endogenous repair through precise modulation of chondroitin sulfate proteoglycans following spinal cord injury. Small 19:e2205012. doi: 10.1002/smll.202205012
Vázquez-Vélez, G. E., and Zoghbi, H. Y. (2021). Parkinson's disease genetics and pathophysiology. Annu. Rev. Neurosci. 44, 87–108. doi: 10.1146/annurev-neuro-100720-034518
Wang, Y., Chen, R., Yang, Z., Wen, Q., Cao, X., Zhao, N., et al. (2022). Protective effects of polysaccharides in neurodegenerative diseases. Front. Aging Neurosci. 14:917629. doi: 10.3389/fnagi.2022.917629
Xing, M., Li, G., Liu, Y., Yang, L., Zhang, Y., Zhang, Y., et al. (2023). Fucoidan from Fucus vesiculosus prevents the loss of dopaminergic neurons by alleviating mitochondrial dysfunction through targeting ATP5F1a. Carbohydr. Polym. 303:120470. doi: 10.1016/j.carbpol.2022.120470
Xu, X. L., Li, S., Zhang, R., and Le, W. D. (2022). Neuroprotective effects of naturally sourced bioactive polysaccharides: an update. Neural Regen. Res. 17, 1907–19012. doi: 10.4103/1673-5374.335142
Zhang, L., Hao, J., Zheng, Y., Su, R., Liao, Y., Gong, X., et al. (2018). Fucoidan protects dopaminergic neurons by enhancing the mitochondrial function in a rotenone-induced rat model of Parkinson's disease. Aging Dis. 9, 590–604. doi: 10.14336/AD.2017.0831
Zhang, F. L., He, Y., Zheng, Y., Zhang, W. J., Wang, Q., Jia, Y. J., et al. (2014). Therapeutic effects of fucoidan in 6-hydroxydopamine-lesioned rat model of Parkinson's disease: role of NADPH oxidase-1. CNS Neurosci. Ther. 20, 1036–1044. doi: 10.1111/cns.12340
Keywords: polysaccharides, algae, neuroprotection, neurodegenerative, basal ganglia
Citation: Lucena AMM, de Souza Lucena EE, Neto SPD, Nobre LTDB, Rocha HAO and Câmara RBG (2024) Algal polysaccharides: new perspectives for the treatment of basal ganglia neurodegenerative diseases. Front. Neuroanat. 18:1465421. doi: 10.3389/fnana.2024.1465421
Edited by:
José Rodolfo Lopes De Paiva Cavalcanti, University of the State of Rio Grande do Norte, BrazilReviewed by:
Felipe Fiuza, Edmond and Lily Safra International Institute of Neuroscience, BrazilCopyright © 2024 Lucena, de Souza Lucena, Neto, Nobre, Rocha and Câmara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eudes Euler de Souza Lucena, ZXVkZXNldWxlckBob3RtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.