
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Aging Neurosci., 28 July 2023
Sec. Alzheimer's Disease and Related Dementias
Volume 15 - 2023 | https://doi.org/10.3389/fnagi.2023.1204134
This article is part of the Research TopicComputational Modeling and Machine Learning Methods in Neurodevelopment and Neurodegeneration: from Basic Research to Clinical ApplicationsView all 11 articles
 Anita Monteverdi1*
Anita Monteverdi1* Fulvia Palesi2
Fulvia Palesi2 Michael Schirner3,4,5,6,7
Michael Schirner3,4,5,6,7 Francesca Argentino2
Francesca Argentino2 Mariateresa Merante2
Mariateresa Merante2 Alberto Redolfi8
Alberto Redolfi8 Francesca Conca9Laura Mazzocchi10
Francesca Conca9Laura Mazzocchi10 Stefano F. Cappa9,11
Stefano F. Cappa9,11 Matteo Cotta Ramusino12
Matteo Cotta Ramusino12 Alfredo Costa2,12
Alfredo Costa2,12 Anna Pichiecchio2,10
Anna Pichiecchio2,10 Lisa M. Farina9
Lisa M. Farina9 Viktor Jirsa13
Viktor Jirsa13 Petra Ritter3,4,5,6,7
Petra Ritter3,4,5,6,7 Claudia A. M. Gandini Wheeler-Kingshott1,2,14
Claudia A. M. Gandini Wheeler-Kingshott1,2,14 Egidio D’Angelo1,2*
Egidio D’Angelo1,2*Introduction: Neural circuit alterations lay at the core of brain physiopathology, and yet are hard to unveil in living subjects. The Virtual Brain (TVB) modeling, by exploiting structural and functional magnetic resonance imaging (MRI), yields mesoscopic parameters of connectivity and synaptic transmission.
Methods: We used TVB to simulate brain networks, which are key for human brain function, in Alzheimer’s disease (AD) and frontotemporal dementia (FTD) patients, whose connectivity and synaptic parameters remain largely unknown; we then compared them to healthy controls, to reveal novel in vivo pathological hallmarks.
Results: The pattern of simulated parameter differed between AD and FTD, shedding light on disease-specific alterations in brain networks. Individual subjects displayed subtle differences in network parameter patterns that significantly correlated with their individual neuropsychological, clinical, and pharmacological profiles.
Discussion: These TVB simulations, by informing about a new personalized set of networks parameters, open new perspectives for understanding dementias mechanisms and design personalized therapeutic approaches.
The advent of advanced in human in vivo recordings of brain signals from, e.g., magnetic resonance imaging (MRI), has led to the identification of brain networks that subtend specific functions (Smitha et al., 2017). The structural, metabolic and/or functional alteration of such networks eventually leads to the clinical manifestation of neurological diseases. In parallel, mathematical modeling of cellular and microcircuit functions are emerging, providing tools to link the micro- to the meso- and the macro-scale properties of brain signals (D’Angelo and Jirsa, 2022).
Neurodegenerative dementias include several neuropathological forms, primarily Alzheimer’s disease (AD) and frontotemporal dementia (FTD). AD is associated with the accumulation of amyloid-β plaques and neurofibrillary tangles, which are widely recognized as typical biomarkers confirming the disease diagnosis. Most AD cases present the typical amnesic form, which reflects the accumulation of protein aggregates in medial temporal lobe structures and evolves in multidomain dementia. Dysfunctions outside the mesial temporal regions characterize atypical AD variants, which present predominant visual, language, executive, behavioral, or motor dysfunction (Graff-Radford et al., 2021). FTD is a heterogeneous neurodegenerative disorder, clinically characterized by behavioral abnormalities, language deficit and motor symptoms. Focal frontal and temporal atrophy are the main macroscopic evidence of FTD pathological changes and distinct atrophy patterns can be associated with different variants (Leyton and Hodges, 2010). Post-mortem histology and in vivo functional MRI (fMRI) studies have suggested a differential engagement of various brain networks in these diseases. However, a comprehensive assessment of functional connectivity (FC) changes in multiple networks in vivo to compare dementias subtypes has been rarely performed (Castellazzi et al., 2014), in favor of investigating specific networks, in particular the default mode network (DMN) specifically in AD (Hohenfeld et al., 2018). Increasing evidence underlines the need to expand the investigation beyond the DMN, considering that widespread increases and decreases in structural, functional and metabolic connectivity have been observed in different brain areas of AD patients (Arnemann et al., 2018; Stefanovski et al., 2021). Moreover, the development of in vivo imaging biomarkers of brain function becomes necessary to achieve efficient tailored diagnosis and personalized treatment, especially in less frequent and more heterogeneous conditions, such as atypical forms of AD or FTD variants (Graff-Radford et al., 2021).
Advanced recording techniques, such as MRI and/or high-density electroencephalography (hd-EEG), are mostly used to study structural and functional brain networks properties and their changes in pathological conditions, but they provide little information about cellular properties such as spatio-temporal dynamics of cellular communication, neuronal firing integrity or synaptic transmission. Proton magnetic resonance spectroscopy (MRS) provides a non-invasive technique to investigate the biochemical properties of the brain and detect metabolic alterations in dementia; aside the fact that acquiring MRS data would prolong the scan time for patients, who are already difficult to image, there is the consideration that most of the studies report extremely heterogeneous results, making clinical application of MRS in AD still limited (Maul et al., 2020). On the other hand, recent studies have addressed FC in FDG-PET data, highlighting the presence of specific metabolic patterns in neurodegenerative dementias, which requires individual subjects’ analyses pipelines as appropriate for clinical settings (Titov et al., 2017).
Therefore, very little is known about the cellular and synaptic changes typical of different diseases, and even more so about whether changes that have cascaded from cells to networks are specific to individual patients.
Recent advances in multiscale brain modeling offer promising tools to study the whole brain temporal dynamics, integrating macroscopic information from structural and functional MRI with mathematical mesoscale representations of the underlying ensemble properties of cells and microcircuits. In particular, The Virtual Brain (TVB) modeling workflow allows the non-invasive investigation of brain features, such as network connectivity strength and excitatory/inhibitory (E/I) balance (Stefanovski et al., 2021; D’Angelo and Jirsa, 2022), which are relevant to brain disease and can be determined for each patient. The E/I balance, in turn, can be extracted at whole brain level or for specific brain networks from parameters measuring excitatory coupling, inhibitory coupling, and recurrent excitation inside network nodes (Deco et al., 2014). Importantly, all neurological conditions involve changes at multiple scales and can gain from the use of TVB for understanding the impact of cellular and microcircuit properties alterations on brain function. The promise for clinical use of TVB has been already suggested in epilepsy surgery (Jirsa et al., 2017), stroke (Falcon et al., 2016), brain tumors (Aerts et al., 2018), Multiple Sclerosis (Marti-Juan et al., 2022), and neurodegenerative conditions like dementia (Zimmermann et al., 2018; Stefanovski et al., 2019; Monteverdi et al., 2022; Triebkorn et al., 2022). Interestingly, the central position of an E/I imbalance in the cascade of pathophysiological events in AD is increasingly recognized (Maestú et al., 2021). However, very little is known on how such network neurophysiology acts in concert with structural and FC alterations to determine cognitive decline. Retrieving E/I information, even if summarized in mesoscale network parameters, is extremely important, as it will provide new insights in neurodegenerative mechanisms of disease that will eventually impact on finding effective treatments.
In this work, we applied TVB to enable the non-invasive investigation of connectivity strength and E/I balance in a heterogeneous cohort of dementia patients, including typical and atypical AD and FTD variants. We explored the relationship between neurophysiological parameters provided by TVB in multiple brain networks and neuropsychological scores recorded during patient examinations. TVB parameters differentiated AD from FTD and proved to be sensitive to profiles of cognitive performance and ongoing pharmacological treatment. In aggregate, this study shows how TVB analysis can be used to provide personalized fingerprints of dementia patients, opening new perspectives for differential diagnosis and for tailoring pharmacological and interventional workflows.
Twenty-three patients affected by neurodegenerative diseases were recruited at the IRCCS Mondino Foundation. The study was approved by the Local Ethical Committee and carried out in accordance with the Declaration of Helsinki. Written informed consent was obtained from all subjects. The protocol was approved by the Local Ethical Committee of the IRCCS Mondino Foundation. Patients underwent a complete diagnostic workup including clinical and neuropsychological assessment (see section below) MRI, and, when available, cerebrospinal fluid (CSF) biomarkers (amyloid-β and τ protein) assessment following the harmonized protocol of the RIN network [Italian Network of the Institutes (IRCCS) of Neuroscience and Neurorehabilitation] (Nigri et al., 2022). Subjects were classified into two main groups: 16 AD patients (13 females, 70 ± 8 years) and 7 FTD patients (1 female, 69 ± 5 years), further classified into distinct phenotypes. In particular, AD patients were additionally classified into: typical AD (10 subjects; Dubois et al., 2014); AD logopenic variant (2 subjects; Dubois et al., 2014); AD frontal variant (ADfv, 1 subject; Dubois et al., 2014); AD posterior cortical atrophy (ADpca, 1 subject; Dubois et al., 2014). One patient was classified as having corticobasal syndrome (CBS, 1 subject; Hassan et al., 2011), and one with dementia with Lewy bodies (DLB, 1 subject; McKeith et al., 2017). On the other hand, FTD patients were classified into: behavioral FTD (FTDbv, 5 subjects; Rascovsky et al., 2011); Primary Progressive Aphasia non-fluent variant (PPAnf, 1 subject; Gorno-Tempini et al., 2011), and Primary Progressive Aphasia semantic variant (PPAsv, 1 subject; Gorno-Tempini et al., 2011). Pharmacological therapy was also recorded.
Ten healthy controls (HC, 6 females, 67 ± 3 years) were enrolled on a voluntary basis as reference group. All HC underwent clinical assessment to exclude any cognitive impairment. For all subjects, exclusion criteria were: age >80 years, a diagnosis of significant medical, neurological and psychiatric disorder, pharmacologically treated delirium or hallucinations and secondary causes of cognitive decline (e.g., vascular metabolic, endocrine, toxic, and iatrogenic). Supplementary Table 1 shows demographic, clinical, and neuropsychological data.
All subjects underwent a neuropsychological examination based on a standardized battery of tests to assess their global cognitive status (Mini-Mental State Examination, MMSE) and different cognitive domains: memory (verbal: Rey’s Auditory Verbal Learning Test, RAVLT; visuo-spatial: Rey–Osterrieth complex figure recall), phonemic and semantic fluency, visuo-constructional abilities (Rey–Osterrieth complex figure copy), attention (Trial Making Test part A, TMT-A) and executive functions (Frontal Assessment Battery, FAB; Trial Making Test part B and B-A; Stroop color-word test interference, time and errors; Raven’s Colored Progressive Matrices, CPM47).
Raw scores were corrected for the effect of age, education, and sex according to the reference norms for the Italian population. Accordingly, corrected scores were classified into five Equivalent Scores (ES), from 0 to 4, with an ES of 0 reflecting a pathological performance, based on percentiles (Capitani and Laiacona, 1997). Domain scores, calculated by averaging the ES of the single tests, were obtained for memory, language-fluency, visuo-constructional abilities, attention, and executive functions, respectively.
All subjects underwent MRI examination using a 3T Siemens Skyra scanner with a 32-channel head coil. The MRI protocol was harmonized within the RIN network including both diffusion weighted imaging (DWI) and resting-state fMRI (rs-fMRI) (Nigri et al., 2022). For DWI data a two-shell standard single-shot echo-planar imaging sequence (EPI) [voxel size = 2.5 mm × 2.5 mm × 2.5 mm, TR/TE = 8,400/93 ms, two shells with 30 isotropically distributed diffusion-weighted directions, diffusion weightings of 1,000 and 2,000 s/mm2, 7 non-diffusion weighted b = 0 s/mm2 images (b0 images) interleaved with diffusion-weighted volumes] was implemented, and 3 non-diffusion weighted images with the reversed phase-encoding acquisition were additionally acquired for distortion correction. For the rs-fMRI data, GE-EPI sequence (voxel size = 3 mm × 3 mm × 3 mm, TR/TE = 2,400/30 ms, 200 volumes) was set. For anatomical reference, the protocol included a whole brain high-resolution 3D sagittal T1-weighted (3DT1) scan (TR/TE = 2,300/2.96 ms, TI = 900 ms, flip angle = 9°, voxel size = 1 mm × 1 mm × 1 mm).
Preprocessing of diffusion and fMRI data was performed according to Monteverdi et al. (2022). Briefly, DWI data were denoised, and corrected for motion and eddy currents distortions (FMRIB Software Library and FSL)1 (Andersson and Sotiropoulos, 2016), then white matter, gray matter (GM), subcortical GM and CSF were segmented from the co-registered 3DT1 volume (MRtrix3)2 (Patenaude et al., 2011). 30 million streamlines whole-brain anatomically constrained tractography (Smith et al., 2012) was performed within MRtrix3, estimating fibers orientation distribution with multi-shell multi-tissue constrained spherical deconvolution (CSD) and using probabilistic streamline tractography (Tournier et al., 2012). fMRI preprocessing was carried out combining SPM123, FSL and MRtrix3 commands in a custom MATLABR2019b script. Marchenko–Pastur principal component analysis (MP-PCA) denoising (Ades-Aron et al., 2020) was firstly performed, followed by slice-timing correction, realignment, co-registration to the 3DT1 volume, polynomial detrending, nuisance regression of 24 motion parameters (Friston et al., 1996) and CSF temporal signal (Muschelli et al., 2014), and temporal band-pass filtering (0.008–0.09 Hz).
An ad hoc anatomical atlas in MNI (Montreal Neurological Institute) space was created combining 93 cerebral (AAL) (including cortical/subcortical structures) and 33 cerebellar (SUIT) labels (Diedrichsen et al., 2009). We then performed a mapping between our ad hoc atlas and the Buckner and Yeo (Buckner et al., 2011; Thomas Yeo et al., 2011) cerebral and cerebellar functional atlases to select the gray matter anatomical nodes of six networks known to support specific functions: (i) integrative networks: DMN, frontoparietal network (FPN), limbic network (LN), and attention network (AN); (ii) motor and sensory networks: visual network (VN) and somatomotor network (SMN) (Figure 1). For each subject, the gray matter parcellation of our combined anatomical atlas was applied to the whole-brain tractography to extract a whole-brain structural connectivity (SC) matrix, with the normalized number of streamlines as edges and cortical/subcortical/cerebellar areas as nodes. The subset of nodes defining each network and their connections were extracted from whole-brain SC obtaining specific network SC matrices, used as input to TVB (as detailed below). In addition, both static and dynamic experimental FC (expFC and expFCD, respectively) were reconstructed from rs-fMRI data for each of the six brain networks, to capture not only synchronous fluctuations of BOLD signals but also their spatiotemporal-dynamics during resting-state (Hansen et al., 2015). The expFC matrix was created by extracting the time-course of BOLD signals for each node and computing the Pearson’s correlation coefficient (PCC) of the time-course of pairs of atlas-defined brain regions. Matrix elements were converted with a Fisher’s z transformation and thresholded at 0.1206 (Palesi et al., 2020). FCD is the dynamic representation of FC over the time and reflects time-variant changes of resting state recordings. To obtain expFCD, expFC was computed over a sliding window of 40 s (expFCsw), shifted incrementally by 1 repetition time, which for our data it means to have 178 expFCsw (Battaglia et al., 2020). Then, each expFCsw was vectorized by considering the upper triangular entries and the vectorized expFCsw were correlated with each other generating the expFCD. Thus, expFCD was calculated as a time-versus-time matrix, containing the Pearson correlation between each expFCsw and all expFCsw, centered at all other time points along the total acquisition window, quantifying, therefore, time-evolving dynamics.
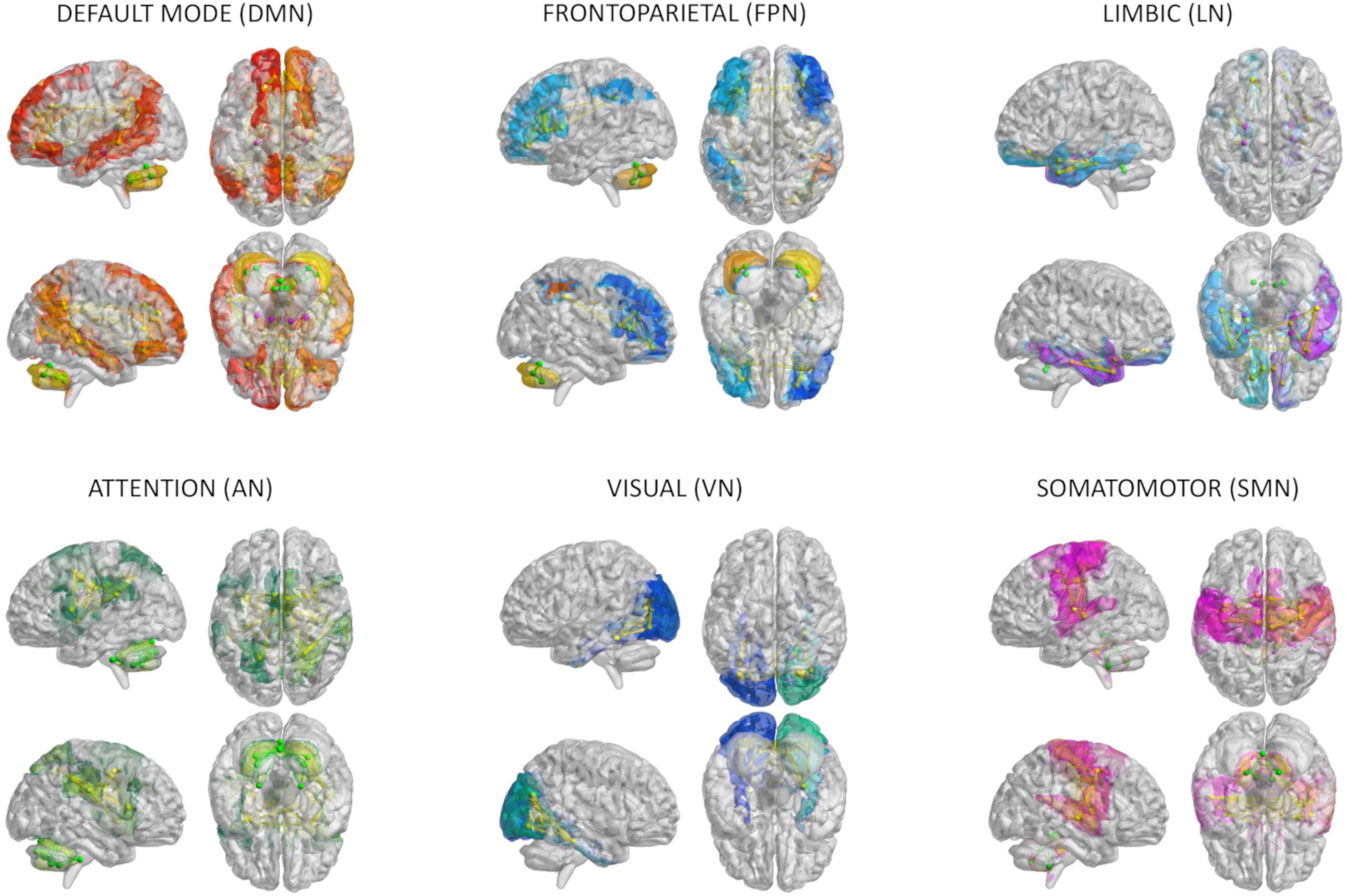
Figure 1. Brain networks. The six networks considered for modeling brain dynamics with The Virtual Brain (TVB): default-mode (DMN), frontoparietal (FPN), limbic (LN), attention (AN), visual (VN), and somatomotor (SMN) network. These networks were defined according to Buckner and Yeo atlases and extracted from whole-brain structural connectivity matrices of each subject, choosing a subset of nodes and connections from the whole brain parcellation. Nodes and edges considered for each network are differently colored.
The TVB workflow [reported in Monteverdi et al. (2022) for the whole brain] was applied to each one of the six selected brain networks (Figure 2). The Wong-Wang neural mass model (Deco et al., 2014; Supplementary Figure 1), implemented with an optimized C code (Schirner et al., 2022), was chosen to simulate local microcircuits activity, resulting from two populations of interconnected excitatory and inhibitory neurons coupled through NMDA and GABA receptor types. In our TVB simulations, this neural mass model was associated to each node of the network, while the SC matrix was used for the nodes interconnection. A set of parameters had to be tuned globally for each network: the global coupling (G), which is a scaling factor that represents the connections strength, and three synaptic parameters, i.e., the excitatory (NMDA) synapses (JNMDA), the inhibitory (GABA) synapses (Ji), and the recurrent excitation (w+). The neural activity simulated with TVB was fed into the Balloon-Windkessel hemodynamic model (Stephan et al., 2007) to reconstruct resting-state BOLD fMRI time-courses over 8 min length and compute simulated FC (simFC) and FCD (simFCD). Parameters were adjusted iteratively using expFC and expFCD of each network as targets to optimize model fitness and the validity of the result was assessed by iterating the optimization using different initial conditions (Supplementary Figure 2; Good et al., 2022). For the simFC vs. expFC comparison, model parameters were tuned until the PCC between experimental and simulated data reached the highest value. For the simFCD vs. expFCD comparison, differences between experimental and simulated FCD were assessed using the Kolmogorov–Smirnov (KS) distance: lower KS values corresponded to a lower distance of frame-by-frame FCD properties, meaning that model and experimental matrices were closest to each other. Thus, to achieve the optimal TVB simulation it was necessary to find both the highest PCC and the lowest KS values. To this aim, an overall cost function was defined as (1 − PCC) + KS and lowest cost function values implied the best fit both to static and dynamic functional data (Kong et al., 2021).
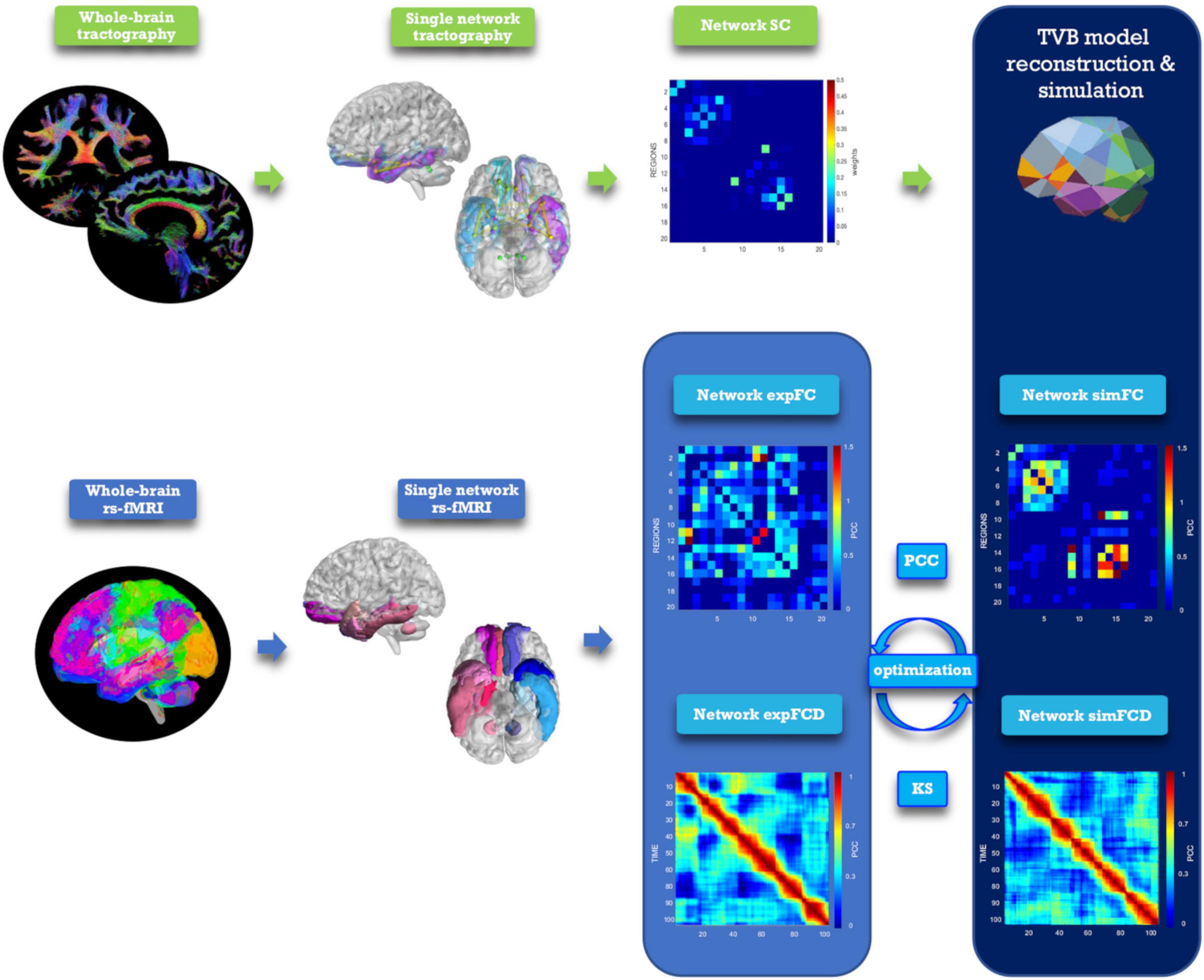
Figure 2. Analysis and modeling workflow. Schematic representation of MRI processing steps integrated in the modeling workflow. From top left, clockwise: diffusion-weighted images after preprocessing and tractography, extraction of a network, structural connectivity (SC) matrix reconstruction for the selected network, TVB simulation performed for the network, reconstruction of simulated static and dynamic (simFC and simFCD) functional connectivity matrices of the same network, optimization of the simulation using model inversion with the experimental FC and FCD (expFC and expFCD), derived from BOLD signals of nodes belonging to the network, as target. Optimal TVB simulation implies both the highest Pearson correlation coefficient (PCC) for static functional data and the lowest Kolmogorov–Smirnov (KS) distance for dynamic functional data.
Statistical tests were performed using SPSS software version 21. Optimal TVB parameters derived for each subject and for each network were tested for normality (Shapiro–Wilk) and then two control tests were performed to assess: (i) whether different networks presented a different E/I balance within the same clinical group (i.e., evaluation of the inter-network E/I balance); and (ii) whether inter-networks E/I balance changed in healthy vs. pathological subjects. Two statistical tests were used: (i) univariate general linear model followed by bias-corrected accelerated Bootstrap (Pek et al., 2018) to correct for age and gender differences in the groups and take into account non-Gaussian data distributions; and (ii) multivariate general linear model between the mean difference (i.e., the difference between the mean value) of TVB parameters in each network compared to the other networks in different clinical groups. Then, a multiple regression analysis was performed to investigate the relationship between individual scores of the 5 cognitive domains (memory, language-fluency, visuo-constructional abilities, attention, and executive functions) and the optimal TVB parameters. Neuropsychological scores in each cognitive domain were considered as dependent variables while model parameters derived for each network were used as predictors in a backward approach. The regression algorithm automatically removed one or more predictors to identify which of them significantly (p < 0.05) explained neuropsychological scores variance.
Meaningful TVB parameters were given as an input to clustering analysis. To avoid overfitting in the study design, the clustering algorithm first performed a feature selection reducing the number of TVB parameters (i) through a semi-supervised approach using LASSO regression model with TVB parameters as independent variables and the diagnostic class as dependent variable; (ii) via PCC between the survived TVB parameters and the diagnostic class; (iii) through Variant Inflation Factors to find out just three meaningful but not correlated features. Then the number of clusters was derived using Gap statistics and the K-means algorithm was applied to label each subject into one cluster defining a personalized fingerprint (Redolfi et al., 2020).
All codes used for this study are open source. The optimized TVB C code can be found at https://github.com/BrainModes/fast_tvb. The dataset will be made available at 10.5281/zenodo.811392.
Model optimization was performed in each of the six brain networks considered in this work. Global coupling (G) and mesoscopic network parameters (Ji, JNMDA, and w+) were adjusted iteratively to fit the experimental data. The reliability of the procedure was assessed by an extensive exploration of the parameter space and by iterating the optimization using different initial conditions (Supplementary Figure 2; Good et al., 2022). Model optimization yielded subject-specific sets of model parameters describing connectivity and E/I balance in each network. TVB parameters revealed differences between networks of healthy and pathological subjects (Supplementary Figure 3 and Supplementary Table 2) that will be further analyzed and explained below.
The mean difference of each network compared to the others was computed in different clinical groups for all the TVB parameters (i.e., G, Ji, JNMDA, and w+). Significant mean difference changes were found both for the TVB parameters in several networks (Figure 3) with network changes summarized in Figure 4. In particular, both in AD and FTD, the connectivity strength (G) decreased in LN and increased in DMN compared to HC; in FTD, G of FPN was lower with respect to other networks. Considering mesoscale synaptic parameters, both FTD and AD showed lower excitatory coupling (JNMDA) in SMN compared to HC; in FTD, JNMDA was lower in VN and higher in FPN; in AD, JNMDA in DMN was higher with respect to other networks. Both in AD and FTD, recurrent excitation (w+) increased in SMN compared to HC; in FTD, w+ was lower in FPN; in AD w+ was lower in DMN with respect to other networks. In FTD, inhibitory coupling (Ji) was lower in FPN and higher in DMN; in AD, AN showed higher Ji and LN lower Ji with respect to other networks.
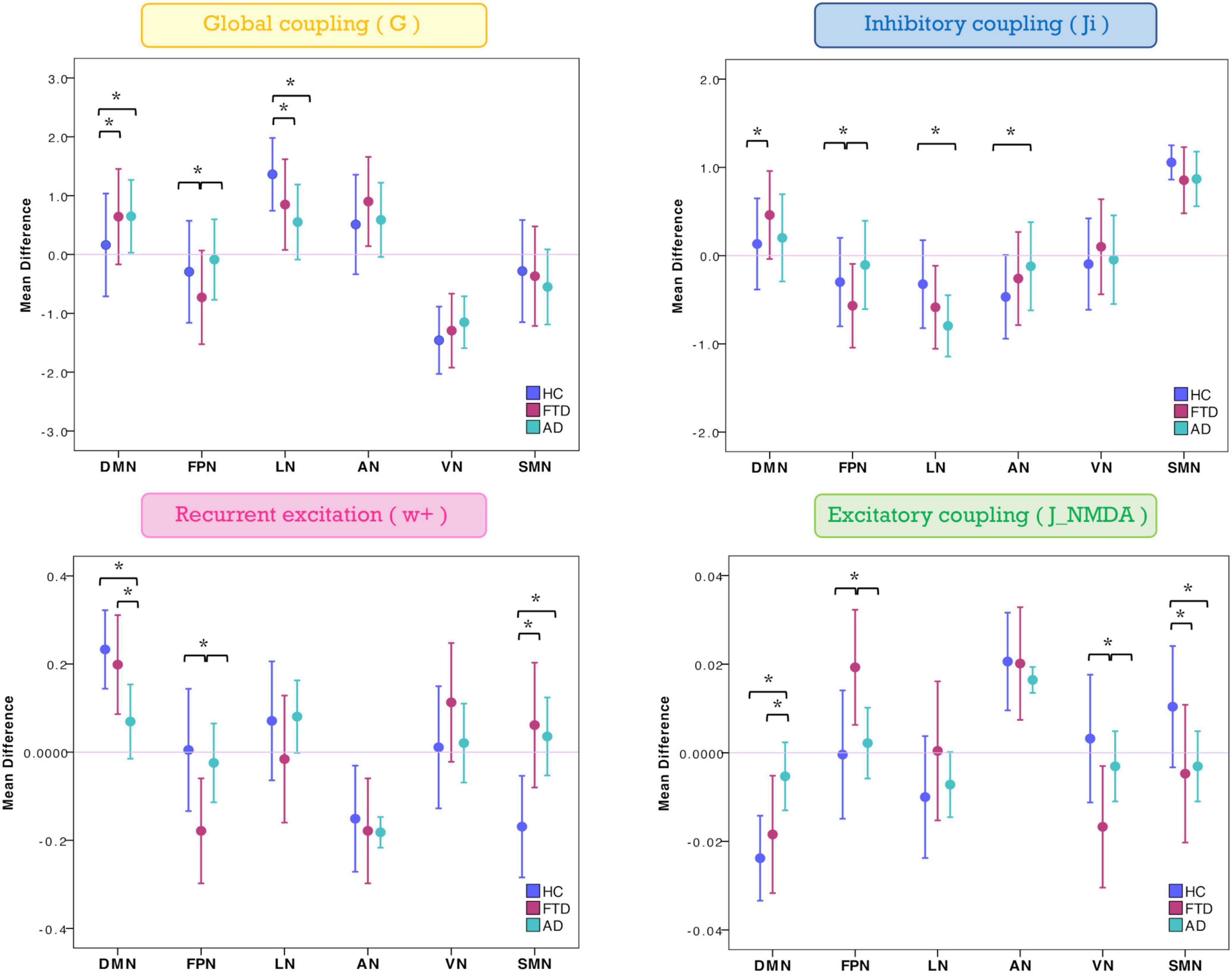
Figure 3. Changes of inter-network relationship. Mean difference of TVB parameters in each given network (DMN, FPN, LN, AN, VN, and SMN) against the others. Positive/negative values indicate a higher/lower TVB parameter mean in a network (on the x-axis) with respect to the TVB parameter mean in the others (line at mean difference 0). Asterisks indicate significant differences (p < 0.05) between clinical groups (HC, FTD, and AD).
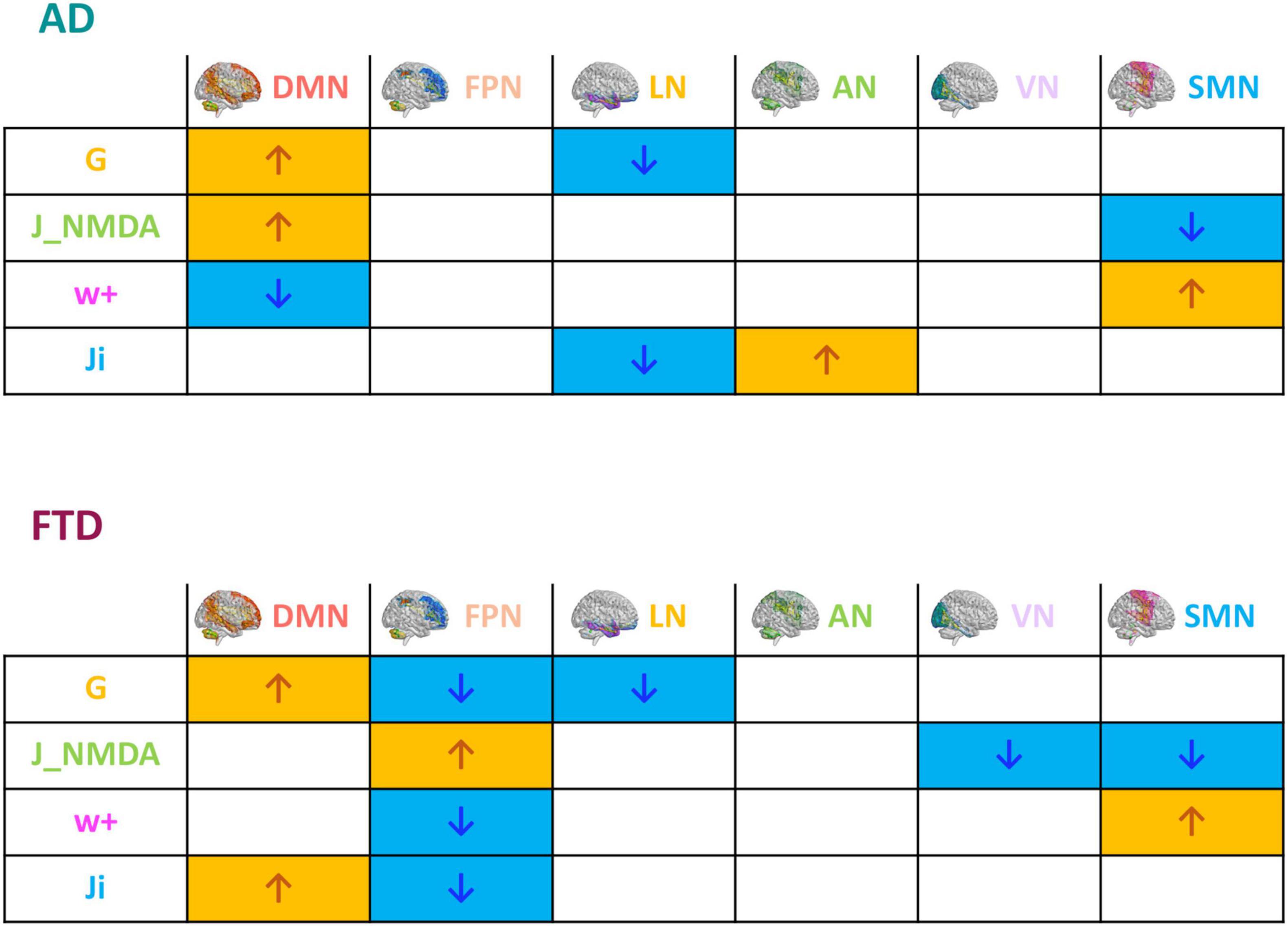
Figure 4. Pathological impact on inter-network relationships. Inter-network (DMN, FPN, LN, AN, VN, and SMN) relationship patterns related to neurodegeneration are summarized in the tables. The increase (yellow) or decrease (blue) of network TVB parameters (G, global coupling; J_NMDA, excitatory coupling; w+, recurrent synaptic excitation; Ji, inhibitory coupling) is indicated with colored arrows.
To assess the significance of the observed mean difference changes in TVB parameters, these were used in backward regression to explain the variation of scores associated to different neuropsychological domains assessed in patients. Network-specific levels of global coupling, excitatory coupling, inhibitory coupling, and recurrent excitation (predictors) significantly (p < 0.05) explained a percentage of variance in the cognitive domains, in which the network is involved (Table 1). The explained variance ranged from ∼20 to ∼45%. Therefore, the mean difference changes in TVB parameters were relevant to explain the neuropsychological performance of patients.
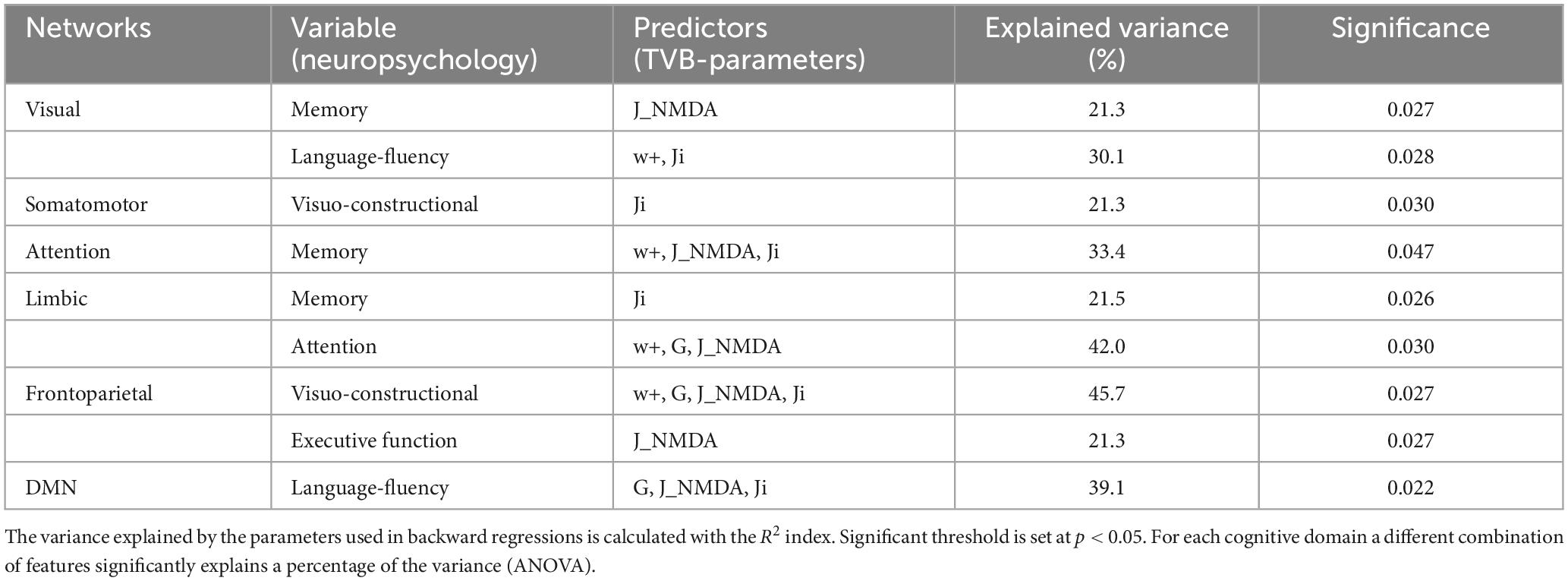
Table 1. Backward regressions results.
The TVB parameters that significantly explained the neuropsychological performance were considered for patients’ labeling using machine learning strategies. From the nineteen parameters identified with backward regression (Table 1) the LASSO algorithm allowed to reduce them to six. Then, G of FPN was excluded, presenting PCC <0.1, and after Variant Inflation Factors three independent and not correlated variables were considered as the most informative features to perform patient’s labeling: Ji of AN, G of the LN and G of the DMN. Gap statistics identified that seven homogeneous classes would be appropriate and the K-means assigned each subject to one of the seven clusters. Each of the identified clusters was characterized by a specific composition of TVB network features (Figure 5A and Supplementary Figure 4). Considering the biophysical meaning of each parameter, they could be described as follows:
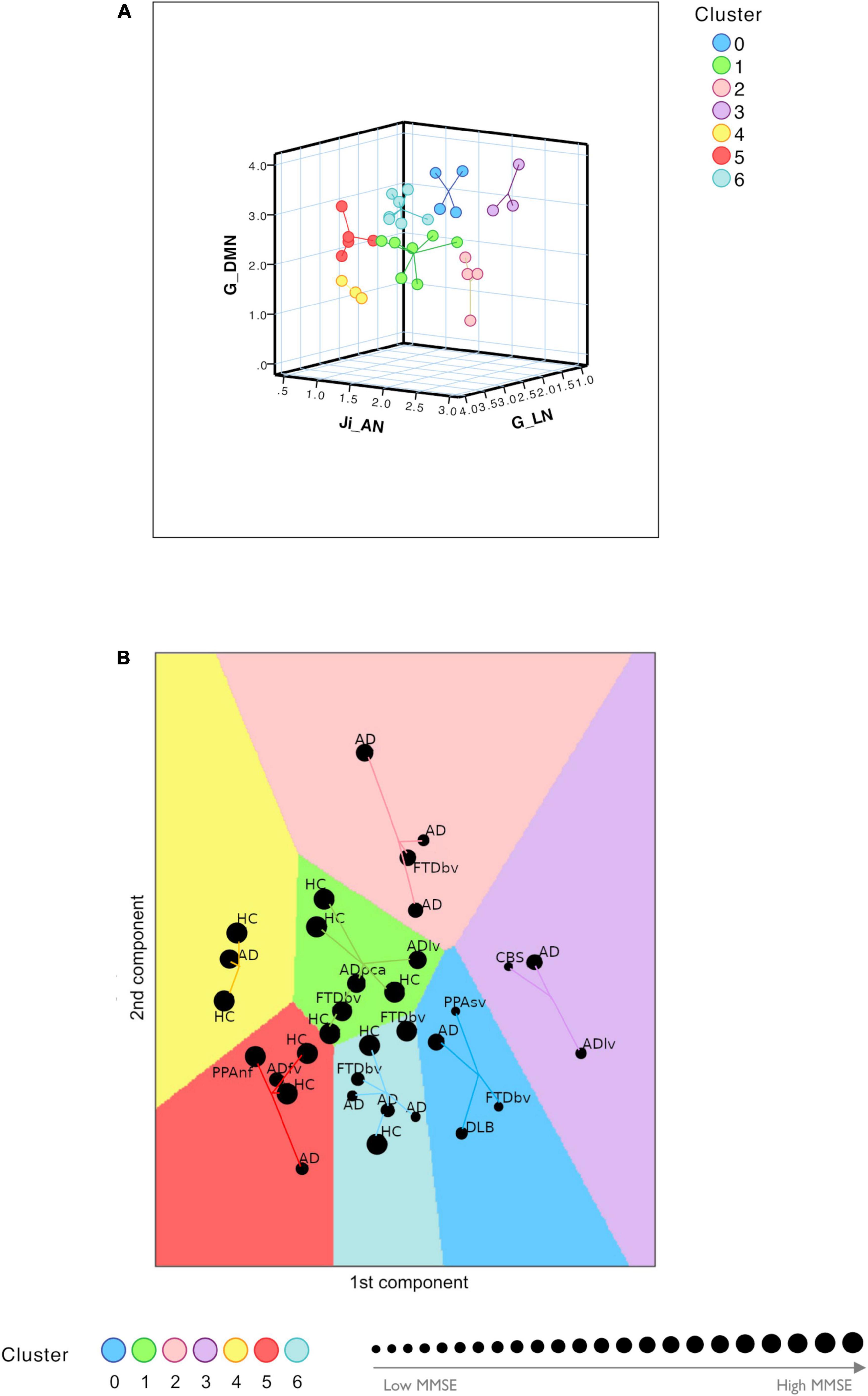
Figure 5. Clustering analysis. (A) Visual representation of the seven clusters (in different colors) identified with K-means analysis using the most meaningful TVB biophysical parameters as input variables. Cognitive network properties (Ji in AN, G in LN, and G in DMN) were considered as the most informative features to perform patient labeling and each of the identified clusters was characterized by a combination of low and high TVB-derived optimal parameters. Each dot represents a subject and lines connect subjects to their own cluster centroid. (B) Each subject was assigned to one of the seven clusters (HC, healthy control; AD, typical Alzheimer’s disease; ADlv, AD logopenic variant; ADfv, AD frontal variant; ADpca, AD posterior cortical atrophy; CBS, corticobasal syndrome; DLB, dementia with Lewy bodies; FTD, frontotemporal dementia; FTDbv, FTD behavioral; PPAnf, primary progressive aphasia non-fluent variant; PPAsv, primary progressive aphasia semantic variant) identifying a personalized fingerprint based on cognitive network properties. Each dot represents a subject and lines connect subjects to their own cluster centroid. The dot dimension corresponds to the MMSE value.
1. Cluster 0 and cluster 3 were mainly characterized by low connectivity strength of LN, high connectivity strength of DMN and hyperinhibition in AN
2. Cluster 1 and cluster 4 were mainly characterized by high connectivity strength of LN, low connectivity strength of DMN and low inhibition in AN
3. Cluster 5 and cluster 6 were mainly characterized by high connectivity strength of LN, high connectivity strength of DMN and low inhibition in AN
4. Cluster 2 was mainly characterized by low connectivity of LN, low connectivity strength of DMN and hyperinhibition in AN.
Clusters 0 and 3 were associated with the lowest mean MMSE values (20.39 ± 5.21 and 18.57 ± 8.28, respectively) while clusters 1 and 4 were associated with the highest mean values (29.08 ± 1.14 and 29.33 ± 1.16, respectively) (Figure 5B and Table 2). No HC was classified into clusters 0 or 3. Moreover, different disease phenotypes were distributed amongst the clusters (Figure 5B): typical AD subjects spread through clusters supporting a heterogeneous distribution of connectivity values in the LN and DMN networks and inhibition of the AN, but no AD patient was found in cluster 1 and the single AD patient belonging to cluster 4 presented a high MMSE score; on the other hand, cluster 0 contained the DLB phenotype, cluster 1 both the non-amnesic variants of AD (ADlv and ADpca), cluster 3 the logopenic variant and the CBS characterized by low MMSE values and cluster 5 contained the frontal variant. Considering the FTD group, FTDbv were heterogeneous and distributed amongst different clusters, but no FTDbv were found in cluster 3. On the other hand, cluster 0 contained PPAsv and cluster 5 PPAnf. Finally, pharmacological assessment of subjects belonging to different groups indicated that the majority of subjects following an antidepressant or anxiolytic treatment belonged to cluster 0 or 1 (Table 2). In particular, subjects belonging to cluster 0 were following an antidepressant therapy mainly with selective serotonin reuptake inhibitors (SSRIs), with the exception of one patient, treated with vortioxetine. Patients belonging to cluster 1, instead, were taking antidepressant drugs different from SSRIs, such as tricyclic antidepressants (e.g., amitriptyline) and serotonin-norepinephrine reuptake inhibitors (e.g., duloxetine), apart from one HC belonging to this group who was found to be on a SSRIs treatment.

Table 2. Mini-mental state examination (mean, SDs) and ongoing pharmacological treatment.
In this work we have generated virtual brain models of dementia patients and simulated neural dynamics of brain networks. The main result is the emergence of specific patterns of alteration in DMN, FPN, and LN, which allow to differentiate AD from FTD. Inter-subject differences, matching the individual neuropsychological profiles and pharmacological treatment, suggest that this approach can generate personalized fingerprints of the disease that could be used to set up future stratification and interventional strategies.
In a first analysis, we compared AD and FTD for their average network model parameters. Model parameters markedly differentiated the mechanisms underlying brain networks dynamics in AD and FTD, with the most typical changes being concentrated in the DMN and LN of AD and in the FPN of FTD.
In both pathologies, G increased in DMN and decreased in LN, while it decreased in FPN in FTD only. It is worth noting that, in these simulations, G represents the overall strength of connections between nodes inside a specific brain network. Moreover, G derives from dynamic TVB analysis and not from functional analysis on fMRI data (Deco et al., 2012), providing new insights into brain connectivity that do not necessarily compare to previously reported connectivity alterations.
In late onset AD there is meta-analytic evidence for a progressive decline of DMN FC, in particular in the posterior component (precuneus, posterior cingulate cortex) (Jones et al., 2016). Increased FC between the posterior DMN and high connectivity hubs, mainly located in the frontal lobes, has been reported in the prodromal stages (Jones et al., 2016). The present observation of increased G in DMN reflects hyper synchronicity, a state in which complexity is reduced along with mutual information transfer among the nodes (Borst and Theunissen, 1999). This concept, deriving from dynamic system theory, is clearly at odd with the common belief that stronger connectivity might represent compensation, leading to the conclusion that a phase-locked hypersynchronous network can perform very limited computations (Deco et al., 2012; Castellazzi et al., 2014). Consistent with this hypothesis is the finding of diffused increase of spectral power in the EEG delta band of AD patients (Babiloni et al., 2015).
Decreased FC inside LN and from LN nodes to neighboring regions has been associated with deterioration of memory and emotional functions (Cai et al., 2017). In FTDbv, a functional disconnection between frontal and limbic areas and an increased FC between DMN regions have been proposed as the probable correlates of apathy and stereotypic behavior (Zhou et al., 2010; Reyes et al., 2018). The decreased G within LN and FPN may be also very detrimental, leading to a reduction of computational states (Deco et al., 2012; Zimmermann et al., 2018).
Another typical pattern differentiating AD from FTD emerged from synaptic parameters. Akin with neuropathology, the major AD changes were detected in DMN, while FTD changes mainly occurred in FPN. DMN showed increased excitatory coupling (JNMDA) and reduced recurrent excitation (w+) in AD, while it showed increased inhibitory coupling (Ji) in FTD. FPN showed no changes at all in AD but it showed a complex set of changes in FTD, including increased JNMDA, reduced w+ and reduced Ji. LN showed reduced Ji in AD. Therefore, the E/I balance, which remarkably impacts on brain dynamics (Deco et al., 2014), was altered in different brain networks, further differentiating AD and FTD.
We can just speculate about the meaning of these changes since information on synaptic parameters in AD and FTD pathologies is sparse. The increased JNMDA in DMN may support the hyperexcitability supposed to explain cognitive impairment in AD (Palop and Mucke, 2016). Local hyperexcitability in the DMN was observed in previous studies, despite a net decrease in inhibitory and excitatory synaptic proteins (Lauterborn et al., 2021; Tok et al., 2021). The reduced Ji of the LN may support the limbic disinhibition reported in AD, which has been associated with a loss of GABAergic receptors (Jiménez-Balado and Eich, 2021). The reduced Ji of the FPN is consistent with the reduction of GABA concentration reported in FTD, which has been associated with behavioral disinhibition (Murley et al., 2021). Our simulations also predict overinhibition in the DMN of FTD, which provides a further differentiation with AD, where inhibition is not changed while excitation is enhanced. DMN has recently been suggested to take part in FTD pathophysiology (Pini et al., 2022). Therefore, the patterns of synaptic changes captured by our study prompts for further experimental and model analysis of synaptic alterations in microcircuits of the AD and FTD brain.
Both in AD and FTD, the SMN showed reduced JNMDA and increased w+. Although the impairment of GABAergic and glutamatergic systems in the motor and sensory networks still needs to be clarified, it should be noted that motor dysfunctions are known to occur in both AD and FTD (Burrell et al., 2011; Lorenzi et al., 2020). In AD, a reduced motor cortex excitability has been reported in mild cognitive impairment (Ferreri et al., 2021), suggesting that these parameters may change along the evolution of the disease. In FTD, motor circuit abnormalities have been suggested to depend on altered glutamatergic transmission (Benussi et al., 2020). Interestingly, in FTD abnormalities of oculomotor functions have been reported (Russell et al., 2021), which might be linked not only to SMN impairment, but also to a more extended involvement of VN, as supported by our results.
Model parameters for individual subjects were correlated with behavioral observations. Global coupling and synaptic parameters of each network significantly contributed to explain neuropsychological scores in specific cognitive domains: LN, AN, and VN with memory; DMN and VN with language-fluency; LN with attention; SMN and FPN with visuo-constructional performance; FPN with executive functions. This evidence is in line with several reports on the importance of motor regions in visuo-constructional performance (Chen et al., 2016), the contribution of AN and limbic areas in memory (Epelbaum et al., 2018), the relevance of frontoparietal areas for executive and visuo-constructional control (Melrose et al., 2013; Dixon et al., 2018), the role of DMN integration for semantic fluency (Jockwitz et al., 2017), and the involvement of visual structures in memory and language-fluency (Kucewicz et al., 2019; Vonk et al., 2019).
Thus, the relationship between neurophysiological parameters in brain networks and neuropsychological scores, which has not been investigated before, provides new cues for understanding the physiopathology of AD and FTD.
The most meaningful model biomarkers for patient’s labeling were G in DMN, G in LN, Ji in AN, consistent with known salient aspects of dementia affecting the ability of daydreaming (DMN), emotional control (LN) and attention (AN). Subjects were found to be distributed between seven different clusters revealing correspondence with their cognitive status (assessed with MMSE) and pharmacological treatment.
Patients with different MMSE scores tended to populate different clusters (see Figure 5), broadly separating patients from HC (MMSE >30), highlighting the importance of DMN, LN, and AN connectivity strength and E/I balance to ensure healthy cognitive function. Interestingly, high G between DMN nodes is associated with a worse performance, being hence disruptive and not compensatory. This analysis suggests that the heterogeneity of subject-specific TVB parameters is able to identify AD “subtypes” (Pini et al., 2021; Rauchmann et al., 2021) and FTD variants. Indeed, subjects belonging to atypical forms of AD and FTD variants were assigned to different clusters, capturing specific aspects of these pathologies and mostly mapping clinical severity assessed with MMSE. A finer grained analysis based on clinical phenotypes is not currently possible, given the limited sample size.
Patients’ labeling based on TVB parameters correlated with pharmacological treatment. Most subjects belonging to clusters 0 and 1 were on antidepressant or anxiolytic treatment (cf. Table 2), which may influence the connectivity strength and the E/I balance of cognitive networks. The effect of SSRIs on LN and DMN FC is increasingly recognized (Van Wingen et al., 2014; Li et al., 2021), while the effect of antidepressant treatment with molecules different from SSRIs, such as vortioxetine, tricyclic molecules or SNRIs (Pérez et al., 2018), as well as the influence of antidepressants on GABA and glutamate levels needs further assessment (Spurny et al., 2021). Considering that patients treated with SSRIs belong to cluster 0 while patients treated with other antidepressant classes belong to cluster 1, our results pose a very intriguing question: is there an opposite impact on cognitive networks exerted by antidepressants with different mechanisms of action or does the cognitive networks profile determine pharmacological treatment response? Future work should study TVB parameters longitudinally pre-post treatment to answer this important question with major potential clinical impact.
It should be noted that, in our cohort, patients were not treated with NMDA receptor antagonists (like memantine) (Robinson and Keating, 2006) or acetylcholinesterase inhibitors (like galantamine, rivastigmine, and donepezil) (Marucci et al., 2021), which are also known to act on AD pathophysiology. NMDA receptors are main triggers of synaptic plasticity, also affected by excitotoxicity and cholinergic receptors that, in turns, act on learning (Waxman and Lynch, 2005; Hasselmo, 2006). Since in the Wong–Wang neural mass model JNMDA is mostly related to slow synaptic mechanisms driven by NMDA receptors (Deco et al., 2014) and receptor density can be remapped onto TVB through parameterization (Deco et al., 2021), an assessment of these receptor-dependent properties could be an important development in future studies.
The small sample size can be seen as a potential limitation in the present study. However, the main aim of this investigation was to assess the ability of TVB to provide a personalized fingerprint of patients, potentially beyond known diagnosis. TVB modeling provides a set of physiological features at single subject level, otherwise not available from standard signal/image acquisition and analysis. Thus, the small sample size does not impact on the TVB ability of uncovering subject-specific features of FC, and E/I profile. The high correlation of TVB parameters with both cognitive performance and pharmacological treatment reveals indeed its exquisite sensitivity to single-subject profiles and opens a broad range of prospective for clinical applications. On the other hand, the application of TVB to a larger cohort of patients bears the potential of improving disease classification of disease subtypes, critical for treatment stratification and for establishing intervention workflows.
The present study demonstrates that brain networks can be characterized in terms of a meaningful set of mesoscale parameters at the single-subject level in humans in vivo. The identification of network abnormalities in patients may be used to design neuromodulation, neuropharmacological, and neuropsychological paradigms capable of regulating circuit function and plasticity (Lin and Wang, 2018), while the high correlation of TVB parameters with both cognitive performance and pharmacological treatment reveals an exquisite sensitivity to single-subject features. As a corollary, it should be remembered that the small sample size does not impact significantly on the TVB capacity of uncovering subject-specific connectivity strength, and E/I profile. At present, it is unclear whether network properties in this study are influenced by therapy suggesting that future studies should systematically address this issue. In aggregate, TVB parameters are shedding light on the changes occurring inside the brain networks of AD and FTD patients opening new perspectives for understanding disease mechanisms and for designing personalized neuromodulation, neuropharmacological and neuropsychological paradigms.
All codes used for this study are open source. The optimized TVB C code can be found at https://github.com/BrainModes/fast_tvb. The dataset is available at https://zenodo.org/record/8113922.
The studies involving human participants were reviewed and approved by the Local Ethical Committee of the IRCCS Mondino Foundation. The patients/participants provided their written informed consent to participate in this study.
MC and AC: patients’ recruitment and clinical assessment. LF, AP, and LM: MRI recordings. FC: neuropsychological testing. AR, AM, FP, and MS: data analysis. AM, FP, MM, FA, ED’A, and CG: TVB modeling and simulation. CG and FP: MRI theory and protocol design. MS, VJ, and PR: TVB support. SC, AC, and MC: neurological support. AM, ED’A, and FP: manuscript writing. ED’A, CG, and FP: work coordination and manuscript finalization. All authors had contributed to manuscript discussion and approved the final version of the manuscript.
The work performed at the IRCCS Mondino Foundation was supported by the Italian Ministry of Health (RC2022-RC2024). The work performed at the University of Pavia was supported by H2020 Research and Innovation Action Grants Human Brain Project 785907 and 945539 (SGA2 and SGA3) to ED’A, FP, and PR. Moreover, the project was supported by the MNL Project “Local Neuronal Microcircuits” of the Centro Fermi (Rome, Italy), #NEXTGENERATIONEU (NGEU) and funded by the Ministry of University and Research (MUR), the National Recovery and Resilience Plan (NRRP), project IR00011-EBRAINS-Italy to ED’A; Horizon2020 [Research and Innovation Action Grants Human Brain Project 945539 (SGA3)], BRC (#BRC704/CAP/CGW), MRC (#MR/S026088/1), Ataxia UK to CW-K; PR acknowledges Digital Europe Grant TEF-Health #101100700; H2020 Research and Innovation Action Grant Human Brain Project (ICEI 800858, EOSC VirtualBrainCloud 82642, AISN 101057655); H2020 Research Infrastructures Grant (EBRAINS-PREP 101079717, EBRAIN-Health 101058516); H2020 European Innovation Council (PHRASE 101058240); H2020 European Research Council Grant (ERC BrainModes 683049); JPND ERA PerMed PatternCog 2522FSB904; Berlin Institute of Health and Foundation Charité; Johanna Quandt Excellence Initiative; and German Research Foundation (SFB 1436, project ID 425899996; SFB 1315, project ID 327654276; SFB 936, project ID 178316478; and SFB-TRR 295, project ID 424778381).
This manuscript has been released as a Pre-Print at BioRxiv (Monteverdi et al., 2023).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2023.1204134/full#supplementary-material
Ades-Aron, B., Lemberskiy, G., Veraart, J., Golfinos, J., Fieremans, E., Novikov, D. S., et al. (2020). Improved task-based functional MRI language mapping in patients with brain tumors through marchenko-pastur principal component analysis denoising. Radiology 298, 365–373. doi: 10.1148/RADIOL.2020200822
Aerts, H., Schirner, M., Jeurissen, B., Van Roost, D., Achten, E., Ritter, P., et al. (2018). Modeling brain dynamics in brain tumor patients using the virtual brain. eNeuro 5:ENEURO.0083-18.2018. doi: 10.1523/ENEURO.0083-18.2018
Andersson, J. L. R., and Sotiropoulos, S. N. (2016). An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. Neuroimage 125, 1063–1078. doi: 10.1016/j.neuroimage.2015.10.019
Arnemann, K. L., Stöber, F., Narayan, S., Rabinovici, G. D., and Jagust, W. J. (2018). Metabolic brain networks in aging and preclinical Alzheimer’s disease. NeuroImage Clin. 17, 987–999. doi: 10.1016/j.nicl.2017.12.037
Babiloni, C., Del Percio, C., Boccardi, M., Lizio, R., Lopez, S., Filippo, C., et al. (2015). Occipital sources of resting state alpha rhythms subjects with amnesic mild cognitive impairment and Alzheimer’s disease. Neurobiol. Aging 36, 556–570. doi: 10.1016/j.neurobiolaging.2014.09.011
Battaglia, D., Boudou, T., Hansen, E. C. A., Lombardo, D., Chettouf, S., Daffertshofer, A., et al. (2020). Dynamic functional connectivity between order and randomness and its evolution across the human adult lifespan. Neuroimage 222:117156. doi: 10.1016/j.neuroimage.2020.117156
Benussi, A., Dell’Era, V., Cantoni, V., Cotelli, M. S., Cosseddu, M., Spallazzi, M., et al. (2020). TMS for staging and predicting functional decline in frontotemporal dementia. Brain Stimul. 13, 386–392. doi: 10.1016/j.brs.2019.11.009
Borst, A., and Theunissen, F. (1999). Information theory and neural networks. North Holl. Math. Libr. 51, 307–340. doi: 10.1016/S0924-6509(08)70042-4
Buckner, R. L., Krienen, F. M., Castellanos, A., Diaz, J. C., and Thomas Yeo, B. T. (2011). The organization of the human cerebellum estimated by intrinsic functional connectivity. J. Neurophysiol. 106, 2322–2345. doi: 10.1152/jn.00339.2011
Burrell, J. R., Kiernan, M. C., Vucic, S., and Hodges, J. R. (2011). Motor neuron dysfunction in frontotemporal dementia. Brain 134, 2582–2594. doi: 10.1093/brain/awr195
Cai, S., Chong, T., Peng, Y., Shen, W., Li, J., von Deneen, K. M., et al. (2017). Altered functional brain networks in amnestic mild cognitive impairment: A resting-state fMRI study. Brain Imaging Behav. 11, 619–631. doi: 10.1007/s11682-016-9539-0
Capitani, E., and Laiacona, M. (1997). Composite neuropsychological batteries and demographic correction: Standardization based on equivalent scores, with a review of published data. J. Clin. Exp. Neuropsychol. 19, 795–809. doi: 10.1080/01688639708403761
Castellazzi, G., Palesi, F., Casali, S., Vitali, P., Wheeler-Kingshott, C. A. M., Sinforiani, E., et al. (2014). A comprehensive assessment of resting state networks: Bidirectional modification of functional integrity in cerebro-cerebellar networks in dementia. Front. Neurosci. 8:223. doi: 10.3389/fnins.2014.00223
Chen, H., Pan, X., Lau, J. K. L., Bickerton, W. L., Pradeep, B., Taheri, M., et al. (2016). Lesion-symptom mapping of a complex figure copy task: A large-scale PCA study of the BCoS trial. NeuroImage Clin. 11, 622–634. doi: 10.1016/j.nicl.2016.04.007
D’Angelo, E. D., and Jirsa, V. (2022). The quest for multiscale brain modeling. Trends Neurosci. 45, 777–790. doi: 10.1016/j.tins.2022.06.007
Deco, G., Jirsa, V., and Friston, K. J. (2012). “The dynamical and structural basis of brain activity,” in Principles of brain dynamics: Global state interactions, eds M. I. Rabinovich, K. J. Friston, and P. Varona (Cambridge, MA: MIT Press), doi: 10.7551/mitpress/9108.003.0003
Deco, G., Kringelbach, M. L., Arnatkeviciute, A., Oldham, S., Sabaroedin, K., Rogasch, N. C., et al. (2021). Dynamical consequences of regional heterogeneity in the brain’s transcriptional landscape. Sci. Adv. 7:eabf4752. doi: 10.1126/sciadv.abf4752
Deco, G., Ponce-Alvarez, A., Hagmann, P., Romani, G. L., Mantini, D., and Corbetta, M. (2014). How local excitation-inhibition ratio impacts the whole brain dynamics. J. Neurosci. 34, 7886–7898. doi: 10.1523/JNEUROSCI.5068-13.2014
Diedrichsen, J., Balsters, J. H., Flavell, J., Cussans, E., and Ramnani, N. (2009). A probabilistic MR atlas of the human cerebellum. Neuroimage 46, 39–46. doi: 10.1016/j.neuroimage.2009.01.045
Dixon, M. L., De La Vega, A., Mills, C., Andrews-Hanna, J., Spreng, R. N., Cole, M. W., et al. (2018). Heterogeneity within the frontoparietal control network and its relationship to the default and dorsal attention networks. Proc. Natl. Acad. Sci. U.S.A. 115, E1598–E1607. doi: 10.1073/pnas.1715766115
Dubois, B., Feldman, H. H., Jacova, C., Hampel, H., Molinuevo, J. L., Blennow, K., et al. (2014). Advancing research diagnostic criteria for Alzheimer’s disease: The IWG-2 criteria. Lancet Neurol. 13, 614–629. doi: 10.1016/S1474-4422(14)70090-0
Epelbaum, S., Bouteloup, V., Mangin, J. F., La Corte, V., Migliaccio, R., Bertin, H., et al. (2018). Neural correlates of episodic memory in the Memento cohort. Alzheimers Dement. 4, 224–233. doi: 10.1016/j.trci.2018.03.010
Falcon, M. I., Riley, J. D., Jirsa, V., McIntosh, A. R., Chen, E. E., and Solodkin, A. (2016). Functional mechanisms of recovery after chronic stroke: Modeling with the virtual brain. eNeuro 3:ENEURO.0158-15.2016. doi: 10.1523/ENEURO.0158-15.2016
Ferreri, F., Guerra, A., Vollero, L., Ponzo, D., Määtta, S., Könönen, M., et al. (2021). TMS-EEG biomarkers of amnestic mild cognitive impairment due to Alzheimer’s disease: A proof-of-concept six years prospective study. Front. Aging Neurosci. 13:737281. doi: 10.3389/fnagi.2021.737281
Friston, K. J., Williams, S., Howard, R., Frackowiak, R. S. J., and Turner, R. (1996). Movement-related effects in fMRI time-series. Magn. Reson. Med. 35, 346–355. doi: 10.1002/mrm.1910350312
Good, T., Schirner, M., Shen, K., Ritter, P., Mukherjee, P., Levine, B., et al. (2022). Personalized connectome-based modeling in patients with semi-acute phase TBI: Relationship to acute neuroimaging and 6 month follow-up. eNeuro 9:ENEURO.0075-21.2022. doi: 10.1523/ENEURO.0075-21.2022
Gorno-Tempini, M. L., Hillis, A. E., Weintraub, S., Kertesz, A., Mendez, M., Cappa, S. F., et al. (2011). Classification of primary progressive aphasia and its variants. Neurology 76, 1006–1014. doi: 10.1212/WNL.0b013e31821103e6
Graff-Radford, J., Yong, K. X. X., Apostolova, L. G., Bouwman, F. H., Carrillo, M., Dickerson, B. C., et al. (2021). New insights into atypical Alzheimer’s disease in the era of biomarkers. Lancet Neurol. 20, 222–234. doi: 10.1016/S1474-4422(20)30440-3
Hansen, E. C. A., Battaglia, D., Spiegler, A., Deco, G., and Jirsa, V. K. (2015). Functional connectivity dynamics: Modeling the switching behavior of the resting state. Neuroimage 105, 525–535. doi: 10.1016/j.neuroimage.2014.11.001
Hassan, A., Whitwell, J. L., and Josephs, K. A. (2011). The corticobasal syndrome-Alzheimer’s disease conundrum. Expert. Rev. Neurother. 11, 1569–1578. doi: 10.1586/ern.11.153
Hasselmo, M. E. (2006). The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol. 16, 710–715. doi: 10.1016/j.conb.2006.09.002.The
Hohenfeld, C., Werner, C. J., and Reetz, K. (2018). Resting-state connectivity in neurodegenerative disorders: Is there potential for an imaging biomarker? NeuroImage Clin. 18, 849–870. doi: 10.1016/j.nicl.2018.03.013
Jiménez-Balado, J., and Eich, T. S. (2021). GABAergic dysfunction, neural network hyperactivity and memory impairments in human aging and Alzheimer’s disease. Semin. Cell Dev. Biol. 116, 146–159. doi: 10.1016/j.semcdb.2021.01.005
Jirsa, V. K., Proix, T., Perdikis, D., Woodman, M. M., Wang, H., Bernard, C., et al. (2017). The Virtual Epileptic Patient: Individualized whole-brain models of epilepsy spread. Neuroimage 145, 377–388. doi: 10.1016/j.neuroimage.2016.04.049
Jockwitz, C., Caspers, S., Lux, S., Jütten, K., Schleicher, A., Eickhoff, S. B., et al. (2017). Age- and function-related regional changes in cortical folding of the default mode network in older adults. Brain Struct. Funct. 222, 83–99. doi: 10.1007/s00429-016-1202-4
Jones, D. T., Knopman, D. S., Gunter, J. L., Graff-Radford, J., Vemuri, P., Boeve, B. F., et al. (2016). Cascading network failure across the Alzheimer’s disease spectrum. Brain 139, 547–562. doi: 10.1093/brain/awv338
Kong, X., Kong, R., Orban, C., Wang, P., Zhang, S., Anderson, K., et al. (2021). Sensory-motor cortices shape functional connectivity dynamics in the human brain. Nat. Commun. 12:6373. doi: 10.1038/s41467-021-26704-y
Kucewicz, M. T., Saboo, K., Berry, B. M., Kremen, V., Miller, L. R., Khadjevand, F., et al. (2019). Human verbal memory encoding is hierarchically distributed in a continuous processing stream. eNeuro 6:ENEURO.0214-18.2018. doi: 10.1523/ENEURO.0214-18.2018
Lauterborn, J. C., Scaduto, P., Cox, C. D., Schulmann, A., Lynch, G., Gall, C. M., et al. (2021). Increased excitatory to inhibitory synaptic ratio in parietal cortex samples from individuals with Alzheimer’s disease. Nat. Commun. 12:2603. doi: 10.1038/s41467-021-22742-8
Leyton, C. E., and Hodges, J. R. (2010). Frontotemporal dementias: Recent advances and current controversies. Ann. Indian Acad. Neurol. 13, S74–S80. doi: 10.4103/0972-2327.74249
Li, L., Su, Y. A., Wu, Y. K., Castellanos, F. X., Li, K., Li, J. T., et al. (2021). Eight-week antidepressant treatment reduces functional connectivity in first-episode drug-naïve patients with major depressive disorder. Hum. Brain Mapp. 42, 2593–2605. doi: 10.1002/hbm.25391
Lin, Y. C., and Wang, Y. P. (2018). Status of noninvasive brain stimulation in the therapy of Alzheimer’s disease. Chin. Med. J. 131, 2899–2903. doi: 10.4103/0366-6999.247217
Lorenzi, R. M., Palesi, F., Castellazzi, G., Vitali, P., Anzalone, N., Bernini, S., et al. (2020). Unsuspected involvement of spinal cord in Alzheimer disease. Front. Cell. Neurosci. 14:6. doi: 10.3389/fncel.2020.00006
Maestú, F., de Haan, W., Busche, M. A., and DeFelipe, J. (2021). Neuronal excitation/inhibition imbalance: Core element of a translational perspective on Alzheimer pathophysiology. Ageing Res. Rev. 69:101372. doi: 10.1016/j.arr.2021.101372
Marti-Juan, G., Sastre-Garriga, J., Vidal-Jordana, A., Llufriu, S., Martinez-Heras, E., Groppa, S., et al. (2022). Using the virtual brain to study the relationship between structural and functional connectivity in people with multiple sclerosis: A multicentre study. Mult. Scler. J. 28, 262–264.
Marucci, G., Buccioni, M., Ben, D. D., Lambertucci, C., Volpini, R., and Amenta, F. (2021). Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 190:108352. doi: 10.1016/j.neuropharm.2020.108352
Maul, S., Giegling, I., and Rujescu, D. (2020). Proton magnetic resonance spectroscopy in common dementias–current status and perspectives. Front. Psychiatry 11:769. doi: 10.3389/fpsyt.2020.00769
McKeith, I. G., Boeve, B. F., Dickson, D. W., Halliday, G., Aarsland, D., Attems, J., et al. (2017). Diagnosis and management of dementia with Lewy bodies: Fourth consensus report of the DLB Consortium. Neurology 89, 88–100.
Melrose, R. J., Harwood, D., Khoo, T., Mandelkern, M., and Sultzer, D. (2013). Association between cerebral metabolism and Rey-Osterrieth Complex Figure Test performance in ALzheimer’s disease. J. Clin. Exp. Neuropsychol. 35, 246–258. doi: 10.1080/13803395.2012.763113.Association
Monteverdi, A., Palesi, F., Costa, A., Vitali, P., Pichiecchio, A., Cotta Ramusino, M., et al. (2022). Subject-specific features of excitation/inhibition profiles in neurodegenerative diseases. Front. Aging Neurosci. 14:868342. doi: 10.3389/fnagi.2022.868342
Monteverdi, A., Palesi, F., Schirner, M., Argentino, F., Merante, M., Redolfi, A., et al. (2023). Virtual brain simulations reveal network-specific parameters in neurodegenerative dementias. bioRxiv [Preprint]. doi: 10.1101/2023.03.10.532087
Murley, A. G., Rouse, M. A., Simon Jones, P., Ye, R., Hezemans, F. H., O’Callaghan, C., et al. (2021). GABA and glutamate deficits from frontotemporal lobar degeneration are associated with disinhibition. Brain 143, 3449–3462. doi: 10.1093/BRAIN/AWAA305
Muschelli, J., Beth Nebel, M., Caffo, B. S., Barber, A. D., Pekar, J. J., and Mostofsky, S. H. (2014). Reduction of motion-related artifacts in resting state fMRI using aCompCor. Neuroimage 96, 22–35. doi: 10.1016/j.neuroimage.2014.03.028
Nigri, A., Ferraro, S., Gandini Wheeler-Kingshott, C. A. M., Tosetti, M., Redolfi, A., Forloni, G., et al. (2022). Quantitative MRI harmonization to maximize clinical impact: The RIN–neuroimaging network. Front. Neurol. 13:855125. doi: 10.3389/fneur.2022.855125
Palesi, F., Lorenzi, R. M., Casellato, C., Ritter, P., Jirsa, V., Gandini Wheeler-Kingshott, C. A. M., et al. (2020). The importance of cerebellar connectivity on simulated brain dynamics. Front. Cell. Neurosci. 14:240. doi: 10.3389/fncel.2020.00240
Palop, J. J., and Mucke, L. (2016). Network abnormalities and interneuron dysfunction in Alzheimer disease. Nat. Rev. Neurosci. 17, 777–792. doi: 10.1038/nrn.2016.141
Patenaude, B., Smith, S. M., Kennedy, D. N., and Jenkinson, M. (2011). A Bayesian model of shape and appearance for subcortical brain segmentation. Neuroimage 56, 907–922. doi: 10.1016/j.neuroimage.2011.02.046
Pek, J., Wong, O., and Wong, A. C. M. (2018). How to address non-normality: A taxonomy of approaches, reviewed, and illustrated. Front. Psychol. 9:2104. doi: 10.3389/fpsyg.2018.02104
Pérez, P. D., Ma, Z., Hamilton, C., Sánchez, C., Mørk, A., Pehrson, A. L., et al. (2018). Acute effects of vortioxetine and duloxetine on resting-state functional connectivity in the awake rat. Neuropharmacology 128, 379–387. doi: 10.1016/j.neuropharm.2017.10.038
Pini, L., Pizzini, F. B., Boscolo-Galazzo, I., Ferrari, C., Galluzzi, S., Cotelli, M., et al. (2022). Brain network modulation in Alzheimer’s and frontotemporal dementia with transcranial electrical stimulation. Neurobiol. Aging 111, 24–34. doi: 10.1016/j.neurobiolaging.2021.11.005
Pini, L., Wennberg, A. M., Salvalaggio, A., Vallesi, A., Pievani, M., and Corbetta, M. (2021). Breakdown of specific functional brain networks in clinical variants of Alzheimer’s disease. Ageing Res. Rev. 72:101482. doi: 10.1016/j.arr.2021.101482
Rascovsky, K., Hodges, J. R., Knopman, D., Mendez, M. F., Kramer, J. H., Neuhaus, J., et al. (2011). Sensitivity of revised diagnostic criteria for the behavioural variant of frontotemporal dementia. Brain 134, 2456–2477. doi: 10.1093/brain/awr179
Rauchmann, B. S., Ersoezlue, E., Stoecklein, S., Keeser, D., Brosseron, F., Buerger, K., et al. (2021). Resting-state network alterations differ between Alzheimer’s disease atrophy subtypes. Cereb. Cortex 31, 4901–4915. doi: 10.1093/cercor/bhab130
Redolfi, A., De Francesco, S., Palesi, F., Galluzzi, S., Muscio, C., Castellazzi, G., et al. (2020). Medical informatics platform (MIP): A pilot study across clinical Italian cohorts. Front. Neurol. 11:1021. doi: 10.3389/fneur.2020.01021
Reyes, P., Ortega-Merchan, M. P., Rueda, A., Uriza, F., Santamaria-García, H., Rojas-Serrano, N., et al. (2018). Functional connectivity changes in behavioral, semantic, and nonfluent variants of frontotemporal dementia. Behav. Neurol. 2018:9684129. doi: 10.1155/2018/9684129
Robinson, D. M., and Keating, G. M. (2006). Memantine: a review of its use in Alzheimer’s disease. Drugs 66, 1515–1534. doi: 10.2165/00003495-200666110-00015
Russell, L. L., Greaves, C. V., Convery, R. S., Bocchetta, M., Warren, J. D., Kaski, D., et al. (2021). Eye movements in frontotemporal dementia: Abnormalities of fixation, saccades and anti-saccades. Alzheimers Dement. 7:e12218. doi: 10.1002/trc2.12218
Schirner, M., Domide, L., Perdikis, D., Triebkorn, P., Stefanovski, L., Pai, R., et al. (2022). Brain simulation as a cloud service: The Virtual Brain on EBRAINS. Neuroimage 251:118973. doi: 10.1016/j.neuroimage.2022.118973
Smith, R. E., Tournier, J. D., Calamante, F., and Connelly, A. (2012). Anatomically-constrained tractography: Improved diffusion MRI streamlines tractography through effective use of anatomical information. Neuroimage 62, 1924–1938. doi: 10.1016/j.neuroimage.2012.06.005
Smitha, K. A., Akhil Raja, K., Arun, K. M., Rajesh, P. G., Thomas, B., Kapilamoorthy, T. R., et al. (2017). Resting state fMRI: A review on methods in resting state connectivity analysis and resting state networks. Neuroradiol. J. 30, 305–317. doi: 10.1177/1971400917697342
Spurny, B., Vanicek, T., Seiger, R., Reed, M. B., Klöbl, M., Ritter, V., et al. (2021). Effects of SSRI treatment on GABA and glutamate levels in an associative relearning paradigm. Neuroimage 232:117913. doi: 10.1016/j.neuroimage.2021.117913
Stefanovski, L., Meier, J. M., Pai, R. K., Triebkorn, P., Lett, T., Martin, L., et al. (2021). Bridging scales in Alzheimer’s disease: Biological framework for brain simulation with the virtual brain. Front. Neuroinform. 15:630172. doi: 10.3389/fninf.2021.630172
Stefanovski, L., Triebkorn, P., Spiegler, A., Diaz-Cortes, M. A., Solodkin, A., Jirsa, V., et al. (2019). Linking molecular pathways and large-scale computational modeling to assess candidate disease mechanisms and pharmacodynamics in Alzheimer’s disease. Front. Comput. Neurosci. 13:54. doi: 10.3389/fncom.2019.00054
Stephan, K. E., Weiskopf, N., Drysdale, P. M., Robinson, P. A., and Friston, K. J. (2007). Comparing hemodynamic models with DCM. Neuroimage 38, 387–401. doi: 10.1016/j.neuroimage.2007.07.040
Thomas Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., et al. (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 106, 1125–1165. doi: 10.1152/jn.00338.2011
Titov, D., Diehl-Schmid, J., Shi, K., Perneczky, R., Zou, N., Grimmer, T., et al. (2017). Metabolic connectivity for differential diagnosis of dementing disorders. J. Cereb. Blood Flow Metab. 37, 252–262. doi: 10.1177/0271678X15622465
Tok, S., Ahnaou, A., and Drinkenburg, W. (2021). Functional neurophysiological biomarkers of early-stage Alzheimer’s disease: A perspective of network hyperexcitability in disease progression. J. Alzheimer’s Dis. 88, 809–836. doi: 10.3233/jad-210397
Tournier, J. D., Calamante, F., and Connelly, A. (2012). MRtrix: Diffusion tractography in crossing fiber regions. Int. J. Imaging Syst. Technol. 22, 53–66. doi: 10.1002/ima.22005
Triebkorn, P., Stefanovski, L., Dhindsa, K., Diaz-Cortes, M., Bey, P., Bülau, K., et al. (2022). Brain simulation augments machine-learning–based classification of dementia. Alzheimers Dement. 8:e12303. doi: 10.1002/trc2.12303
Van Wingen, G. A., Tendolkar, I., Urner, M., van Marle, H. J., Denys, D., Verkes, R. J., et al. (2014). Short-term antidepressant administration reduces default mode and task-positive network connectivity in healthy individuals during rest. Neuroimage 88, 47–53. doi: 10.1016/j.neuroimage.2013.11.022
Vonk, J. M. J., Rizvi, B., Lao, P. J., Budge, M., Manly, J. J., Mayeux, R., et al. (2019). Letter and category fluency performance correlates with distinct patterns of cortical thickness in older adults. Cereb. Cortex 29, 2694–2700. doi: 10.1093/cercor/bhy138
Waxman, E. A., and Lynch, D. R. (2005). N-methyl-D-aspartate receptor subtypes: Multiple roles in excitotoxicity and neurological disease. Neuroscientist 11, 37–49. doi: 10.1177/1073858404269012
Zhou, J., Greicius, M. D., Gennatas, E. D., Growdon, M. E., Jang, J. Y., Rabinovici, G. D., et al. (2010). Divergent network connectivity changes in behavioural variant frontotemporal dementia and Alzheimer’s disease. Brain 133, 1352–1367. doi: 10.1093/brain/awq075
Keywords: virtual brain modeling, brain dynamics, excitatory/inhibitory balance, Alzheimer’s disease, frontotemporal dementia, resting-state networks
Citation: Monteverdi A, Palesi F, Schirner M, Argentino F, Merante M, Redolfi A, Conca F, Mazzocchi L, Cappa SF, Cotta Ramusino M, Costa A, Pichiecchio A, Farina LM, Jirsa V, Ritter P, Gandini Wheeler-Kingshott CAM and D’Angelo E (2023) Virtual brain simulations reveal network-specific parameters in neurodegenerative dementias. Front. Aging Neurosci. 15:1204134. doi: 10.3389/fnagi.2023.1204134
Received: 11 April 2023; Accepted: 10 July 2023;
Published: 28 July 2023.
Edited by:
Pablo Martinez-Cañada, University of Granada, SpainReviewed by:
Basabdatta Sen Bhattacharya, Birla Institute of Technology and Science, IndiaCopyright © 2023 Monteverdi, Palesi, Schirner, Argentino, Merante, Redolfi, Conca, Mazzocchi, Cappa, Cotta Ramusino, Costa, Pichiecchio, Farina, Jirsa, Ritter, Gandini Wheeler-Kingshott and D’Angelo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anita Monteverdi, YW5pdGEubW9udGV2ZXJkaTAxQHVuaXZlcnNpdGFkaXBhdmlhLml0; Egidio D’Angelo, ZGFuZ2Vsb0B1bmlwdi5pdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.