
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Aging Neurosci. , 07 September 2022
Sec. Parkinson’s Disease and Aging-related Movement Disorders
Volume 14 - 2022 | https://doi.org/10.3389/fnagi.2022.960479
This article is part of the Research Topic Insights in Parkinson’s Disease and Aging Related Movement Disorders View all 11 articles
 Xi Chen1,2*
Xi Chen1,2* Zhaofei Yang3
Zhaofei Yang3 Yaping Shao3
Yaping Shao3 Kunhyok Kim3
Kunhyok Kim3 Yuanyuan Wang3Ying Wang3Haifeng Wu3Xiaolan Xu1,2
Yuanyuan Wang3Ying Wang3Haifeng Wu3Xiaolan Xu1,2 Weidong Le1,2,3*
Weidong Le1,2,3*Background: The classical motor symptoms of Parkinson’s disease (PD) are tightly linked to the gradual loss of dopamine within the striatum. Concomitantly, medium spiny neurons (MSNs) also experience morphological changes, such as reduced dendritic complexity and spine density, which may be potentially associated with motor dysfunction as well. Thus, MSNs may serve as the emerging targets for PD therapy besides the midbrain dopaminergic neurons.
Results: To comprehensively examine pathological alterations of MSNs longitudinally, we established a THCre/Pitx3fl/fl (Pitx3cKO) mouse model that developed canonical PD features, including a significant loss of SNc DAergic neurons and motor deficits. During aging, the targeted neurotransmitter, MSNs morphology and DNA methylation profile were significantly altered upon Pitx3 deficiency. Specifically, dopamine, GABA and glutamate decreased in the model at the early stage. While nuclear, soma and dendritic atrophy, as well as nuclear invaginations increased in the aged MSNs of Pitx3cko mice. Furthermore, more nuclear DNA damages were characterized in MSNs during aging, and Pitx3 deficiency aggravated this phenomenon, together with alterations of DNA methylation profiling associated with lipoprotein and nucleus pathway at the late stage.
Conclusion: The early perturbations of the neurotransmitters within MSNs may potentially contribute to the alterations of metabolism, morphology and epigenetics within the striatum at the late stage, which may provide new perspectives on the diagnosis and pathogenesis of PD.
The striatum is the largest integrative component of the basal ganglia and plays an essential role in modulating complex behaviors, such as facilitation or inhibition of actions and reward learning (Du and Graves, 2019; Prager and Plotkin, 2019). It receives the glutamatergic afferents from the cerebral cortex and thalamus as well as DAergic afferents from the SNc. These massive neurochemical inputs from corticostriatal, thalamostriatal, and nigrostriatal projections are largely processed by striatal MSNs, together with the interneurons in a topographically organized manner (Bariselli et al., 2019; Filipovic et al., 2019; Chen et al., 2020). MSNs use γ-aminobutyric acid (GABA) as a neurotransmitter and constitute 90–95% of the striatal neuronal population (He et al., 2021). Despite high homogeneity, MSNs can be divided into two distinct subpopulations based on their output projection pathways and neurochemical content (Gerfen et al., 1990; Gong et al., 2003; He et al., 2021). Meanwhile, the MSNs are the only striatal neurons with dendritic spines that are highly specialized structures of neuronal connectivity for the regulation of synaptic strength (Bolam et al., 2000; Parisiadou et al., 2014).
In PD patients, the striatum undergoes progressive DA depletion (Wang et al., 2021), consequently leading to cardinal motor symptoms, such as resting tremor, bradykinesia, postural instability, and rigidity. Meanwhile, MSNs as the predominant striatal neuron population, also experience the morphologic alterations, i.e., the dendritic alterations have been observed in postmortem studies of PD brains (McNeill et al., 1988; Stephens et al., 2005; Zaja-Milatovic et al., 2005). Previously, the toxic PD animal models showed the reduced dendritic length and spine density of MSNs, resembling the findings identified in PD patients (Day et al., 2006; Villalba et al., 2009; Zhang et al., 2013; Suarez et al., 2014; Toy et al., 2014). However, these observations may be due in part to the toxic effects and independent of the PD-induced pathology. Later on, Suarez et al. (2018) represented that the MSNs in Pitx3 knockout mice do appear similar morphologic abnormalities to that of toxic PD animal models, excluding the toxic effects and suggesting a close association between PD and MSNs. Pitx3 is a transcription factor mainly expressed in midbrain DAergic neurons and plays an essential role in DAergic neuronal development (Smidt et al., 1997, 2004). Later on, our studies showed that Pitx3 is also involved in maintaining the normal physiological functions in the postnatal DAergic neurons (Wang et al., 2021). During aging, the vulnerability of SNc DAergic neurons increased with an early decline in glial cell line-derived neurotrophic factor (GDNF) and aldehyde dehydrogenase 1a1 (Aldh1a1) levels (Wang et al., 2021). Since the deficiency of Pitx3 triggered a profound loss of SNc DAergic neurons even at the embryonic stage (Hwang et al., 2003; Filali and Lalonde, 2016), few DAergic innervations project onto the striatum and thereby most MSNs are exposed to little DA throughout life, rather than to the progressive depletion of DA during aging. To overcome this drawback and comprehensively examine the pathological alterations of MSNs longitudinally, we generated a THCre/Pitx3fl/fl (Pitx3cKO), a conditional knockout mouse model, where a progressive reduction of striatal DA occurs in the fully developed MSNs. Meanwhile, the mice showed a significant loss of SNc DAergic neurons and movement abnormalities. Thus, the utilization of the Pitx3cKO model may offer great potential for systematically examining the neurochemistry, morphology and epigenetics of MSNs during aging. Our preliminary studies revealed that multiple neurotransmitters were deducted first in the young Pitx3cKO mice. The early disruption of neurochemistry may thereby contribute to the remodeling of MSNs late—reduced dendritic complexity of MSNs, and shrinkage of soma and nuclear size, together with altered epigenetic profiling in Pitx3cKO mice.
The heterozygous mice Pitx3Flox/wt with C57BL/6J background were generated by ViewSolid Biotech Co., Ltd. (Beijing, China) and the TH-Cre driver mice with C57BL/6J background were generated by Shanghai Model Organisms (Shanghai, China); both mouse lines are available upon request. The RioTag mice were purchased from JAX lab (#011029) (Shigeoka et al., 2016). To achieve the conditional knockout Pitx3 mouse model in the DA neuronal system, Pitx3cKO mice were produced by breeding mice carrying an Cre recombinase under the TH promoter with the homozygous mice Pitx3Flox/Flox. All experimental mice were maintained under specific-pathogen-free (SPF) conditions (temperature, 22°C ± 2°C; air exchange, per 20 min; 12 h/12 h light–dark cycle) with free access to food and water. Animal care and procedures were carried out in accordance with the Laboratory Animal Care Guidelines approved by the Institutional Animal Care Committee at Dalian Medical University. The protocol was approved by the Institutional Animal Care Committee at Dalian Medical University.
For Pitx3Flox/wt mice, CRISPR technology was applied to cut the DNA of the intron of the Pitx3 gene, providing the homologous template donor. The sequences of floxp were inserted at both ends of the specific exons (exon 2 and exon 3) through homologous recombination. When mated with tissue-specific expression Cre mice, the specific exon 2 and exon 3 of Pitx3 were deleted, thereby achieving the purpose of conditional knockout of the Pitx3 gene. For TH-Cre knock-in mouse driver line, the Cre recombinase was cloned following an IRES sequence. An frt-flanked neomycin selection cassette was added, and the construct was cloned in the 3′ untranslated end of the TH gene (as described by Althini et al., 2003). The coding sequence of TH is not affected, nor are the expression levels, so both the TH and Cre recombinase proteins are produced in Th-expressing cells of this mouse line.
The open field test was performed in a quiet testing room. To measure the locomotor activity, mice were placed into an Activity Monitor instrument (25 cm × 25 cm × 30 cm, Med Associates Inc., St. Albans, United States) equipped with computer-controlled photocells. Locomotor activity was automatically recorded for 6 min, and different elements of open field test were calculated by the Med system.
Rotarod motor skill learning test was performed as described previously (Wu et al., 2019). Mice were placed onto a rotating rod with auto acceleration from 0 to 40 rpm in 5 min (Model 755, IITC Life Science). The length of time the mouse stayed on the rotating rod was recorded across 10 trials. Such experiments were performed on six continuous days.
Mouse brains were collected at indicated time points. The brains were rapidly isolated and postfixed in ice-cold 4% paraformaldehyde and subsequently dehydrated for 24 h in 15% and 30% sucrose at 4°C, as described previously (Dong et al., 2020). Sections were incubated for 1 h in blocking solution (5% normal goat serum, 0.2% Triton-X 100, and 0.05% NaN3 in PBS). The primary antibodies were used as follows: anti-TH (1:1,000, AB152; Millipore, United States), anti-TH (1:2,000, T1299; Sigma-Aldrich, United States), anti-TH (1:1,000, TYH, Aves Labs, United States), anti-NeuN (1:1,000, MAB377; Millipore, United States), anti-LaminB1 (1:1,000, 12987-1-AP; Proteintech, United States), anti-Darpp32 (1:1,000, 2306; CST, United States), anti-phospho-Histone H2AX (1:1,000, 2577; CST, United States), anti-GFP (1:1,000, ab6662; Abcam, United States), anti-nuclear pore complex (1:1,000, ab24609; Abcam, United States), anti-TOM20 (1:1,000, 42406; CST, United States), anti-Ctip2 (1:500, ab18465; Abcam, United States), anti-HA (1:500, ab9110; Abcam, United States) and anti-Pitx3 (provided by Dr. Marten P. Smidt’s lab at the University of Amsterdam, Netherlands). The section images were visualized and photographed directly with a confocal microscope (A1 confocal, Nikon Instruments [Shanghai] Co., Ltd.) and a DP80 CCD brightfield microscope (Olympus, Japan). The outlines of the SNc and VTA were delimited according to anatomical landmarks (Fu et al., 2012).
For neuron counting, a series of coronal sections (40 μm per section, every third section from Bregma –2.70 to –3.88 mm) were selected and stained with anti-TH and anti-NeuN antibodies for quantification. Usually, 10–11 sections were collected per animal. The entire midbrain regions were scanned under a 10X objective (A1 confocal, Nikon Instruments [Shanghai] Co., Ltd.). We multiplied the total calculated from 10 to 11 sections by 3 to obtain the final number of TH+ or NeuN+ neurons (Dong et al., 2020; Wang et al., 2021). The IFC intensity of the striatum was analyzed using ImageJ software. Typically, the data were collected from 2 to 3 slices per animal.
The stereotaxic AAV injections (AAV-hSyn1-eGFP, GeneCopoeia) were conducted on 6- and 12-month-old Pitx3cWT and Pitx3cKO mice. Before surgery, mice were deeply anesthetized by intraperitoneal injection of ketamine (100 mg/kg)/xylazine (10 mg/kg) solution. To achieve sparse labeling, 1.1 × 1012 viral particles with a total volume of 500 nl were injected into dorsal striatum (coordinates used, AP: 0.98 mm, ML: ± 2.2 mm from bregma, DV: –3.0 mm from exposed dura mater). Virus solution was injected at an infusion rate of 100 nl/min and the needle was withdrawn 10 min after the end of injection. Following virus injection, the scalp was sutured, and the mice were returned to their home cages. The virus-injected mice were used for experiment at least 4 weeks after the virus infusion.
Based on the previous study (Lu and Yang, 2017), the AAV-infused mouse brains were sectioned at 60 μm of thickness. The sections were stained with GFP antibody (1:1000, ab6662; Abcam, Cambridge, UK) and Ctip2 antibody (1:500, ab18465, Abcam, Cambridge, UK). Afterward, the stained sections were imaged using a laser scanning confocal microscope (A1 confocal, Nikon Instruments [Shanghai]Co., Ltd.) under 40× objective lens. The MSNs were identified based on the positive staining of Ctip2. Neuronal structure reconstruction with neuTube (Feng et al., 2015) and Sholl analysis were performed with ImageJ (Longair et al., 2011).
First, we added 400 μl solution 1 (methanol/water, 1:1, v/v, with 0.1% formic acid) with succinic acid as interior label into the striatum tissue, and the mixture were homogenized. The homogenate was ultrasounded in the ice bath for 10 min, and then was incubated in the ice bath (–20°C) for 30 min. The samples were centrifuged at 12,000 rpm × 10 min under 4°C and 300 μl supernatant was preserved. The pellet was incubated with 200 μl solution 1, vortex for 30 s, and then was ultrasounded in the ice bath for 5 min. The samples were centrifuged at 12,000 rpm × 10 min under 4°C and 200 μl supernatant was preserved. We combined 200 μl supernatant with 300 μl preserved supernatant above and obtained 500 μl supernatant totally. All these 500 μl supernatant was transferred into a glass vial to conduct vacuum-drying. After dissolving and centrifuging, the obtained supernatant was used to identify dopamine, GABA and glutamate using LC-MS system.
Genomic DNA was extracted from striatum tissues per mouse at indicated timepoints (QIAamp DNA Micro kit Qiagen, German). MethylRAD library preparation and sequencing were conducted according to the protocol described by Wang et al. (2015). PE sequencing was performed on the Illumina HiSeq X-Ten platform. After QC and filtering of the original reads and removal of the sequences with linkers, low-quality sequences (more than five bases with a quality lower than 10), and those with Ns (unidentified bases), the high-quality clean reads containing the methylated CG/CWG sites were mapped to the reference sequence (signatures with CG/CWG sites) of the mouse genome GENCODE V38 by the SOAP program (version 2.21). Sites covered by at least three reads were regarded as reliable DNA methylation sites. Then, the number of methylated sites and the depth of signature coverage of each sample were calculated. The methylation levels of a site (CG/CWG) could be reflected by the sequencing depth of the methylated signature. The unit of the quantitative value of site methylation was RPM (reads per million), which means that the quantitative value of the methylation level of a site was equal to the coverage at that site in number of reads/the number of high-quality reads in the library multiplied by 1,000,000. Furthermore, the distributions of the methylated CG/CWG sites on different elements of the genome, especially on the different regions of genes, were evaluated by SnpEff software (version: 4.1g) (Cingolani et al., 2012) and bed tools software (v2.25.0) (Quinlan and Hall, 2010). Then, the DNA methylation levels of the genes were evaluated by summing the methylation levels of sites that were localized in the gene region. The differential DNA methylation levels of the sites and genes were identified by using the R package DESeq (Anders and Huber, 2010). Finally, the genes with different methylation levels in different samples were further analyzed based on Gene Ontology (GO) enrichment by DAVID and Kyoto Encyclopedia of Genes and Genomes (KEGG)1 enrichment.
Graph Pad Prism 8 and R were used for statistical analysis. The data were collected and processed randomly. No statistical methods were used to predetermine sample size, but our sample sizes are similar to those reported in previous publications. The statistical significance was determined using Student’s t- test, 2way ANOVA with Sidak’s multiple comparisons test, conditional logistic regression, and multiple t-test with Benjamini and Hochberg test.
Our Pitx3cKO mouse model utilized a Cre-mediated recombination system driven by the TH promoter, resulting in the removal of the second and third coding exons of Pitx3 (Supplementary Figure 1). After multiple generations of breeding, homozygous Pitx3-floxed mice harboring (Pitx3cKO) or not harboring (Pitx3cWT) the Cre gene were achieved, and the genotypes were characterized by PCR analysis (Supplementary Figure 1). The expression of Pitx3 was hardly detected in 2-month-old Pitx3cKO mice by IFC staining (Supplementary Figure 1), indicating the success of Pitx3 deletion. Unlike ak and traditional Pitx3 knockout mice, Pitx3cKO mice rarely showed eye defects. Additionally, to confirm the TH-driven Cre expressions, we crossed the RiboTag mice with TH-Cre line (Supplementary Figure 2).
Also, the number of DAergic neurons (Figure 1) and the dendritic complexity of MSN cells (Supplementary Figure 3) were comparable between Pitx3cKO and Pitx3cWT mice at the age of 6 months. These results indicated that the development of retina cells, DAergic neurons and MSNs was not disrupted. However, a significant loss of DAergic neurons was characterized in 12-month-old Pitx3cKO mice, where about 20% of SNc DAergic neurons died (Figure 1). Moreover, such deficit was exaggerated at 18 months with around 31.5% neuronal loss in Pitx3cKO mice compared to Pitx3cWT mice (Figure 1). Interestingly, VTA DAergic neurons were less affected by Pitx3-deficiency and remained intact during aging (Figure 1), resembling the neuropathological phenotype observed in ak and traditional knockout mice. Also, during aging, there is a natural decline in the number of DAergic neurons of 18-month-old Pitx3cWT mice compared to 12-month-old ones (Supplementary Figure 4). Taken collectively, these results demonstrated the importance of the Pitx3 gene in adult neuronal survival (Wang et al., 2021).
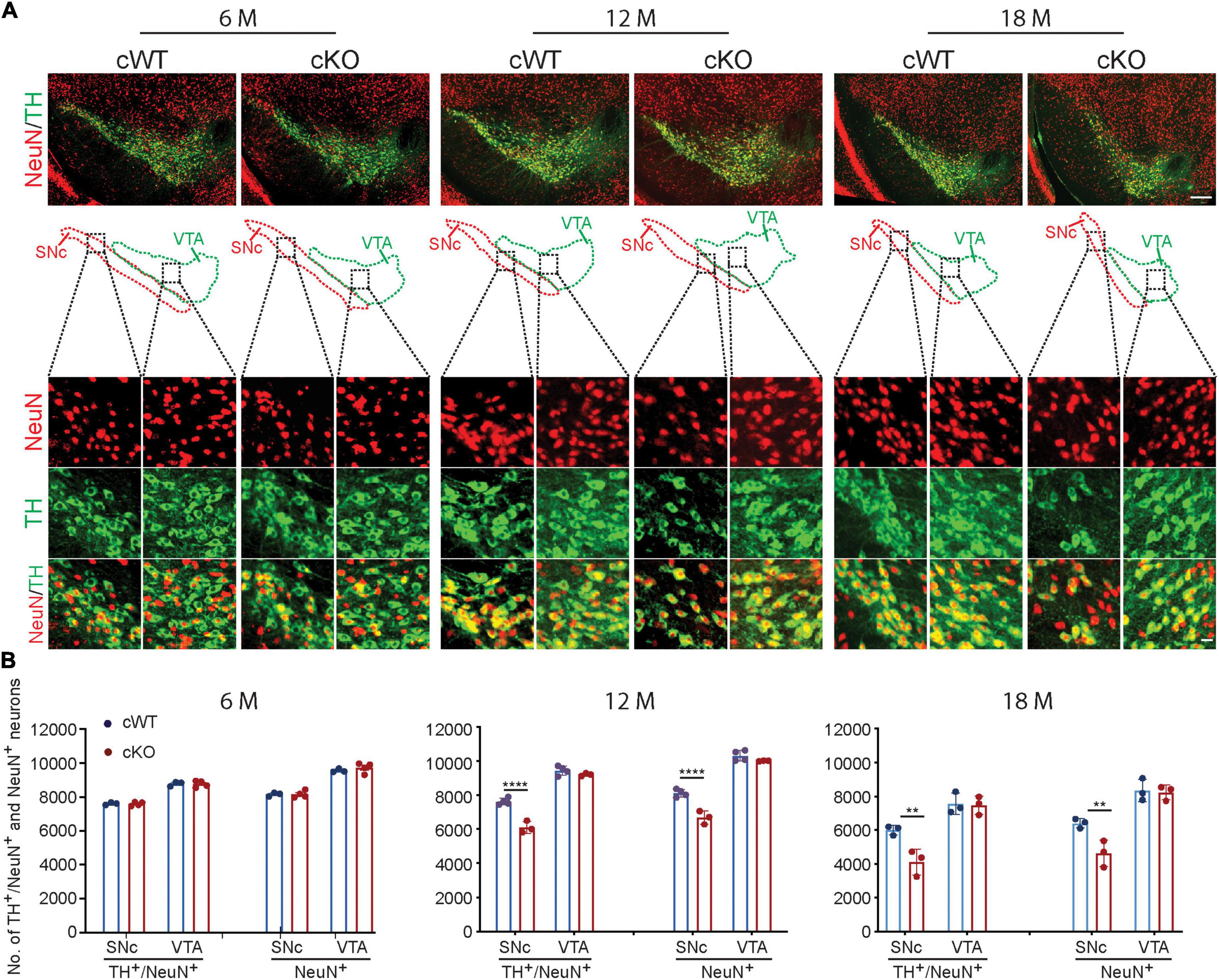
Figure 1. Neurodegeneration in 12- and 18-month-old Pitx3cKO mice. (A) IFC co-staining of TH and NeuN in the ventral midbrain sections from 6-, 12-, and 18-month-old Pitx3cWT and Pitx3cKO mice. SNc and VTA were outlined, respectively (scale bar: 200 μm; high-magnification, 20 μm). (B) Quantification of TH+/NeuN+ and NeuN+ neurons in the SNc and VTA from 6-, 12-, and 18-month-old Pitx3cWT and Pitx3cKO mice (N = 3–4 mice per genotype; all males except for two females in 6-month-old Pitx3cKO and 12-month-old Pitx3cWT). 2way ANOVA analysis with Sidak’s multiple comparisons test, ****p < 0.0001 (12 months for TH+/NeuN+ co-staining), ****p < 0.0001 (12 months for NeuN+ staining), **p = 0.0041 (18 months for TH+/NeuN+ co-staining), **p = 0.0064 (18 months for NeuN+ staining).
Besides neuronal loss, our findings further elucidated that the striatal TH expression levels were decreased by 46% in 18-month-old Pitx3cKO mice (Figures 2A,B), and the reduction of striatal DA levels in Pitx3cKO mice was identified as early as 6 months of age (Figure 2C). Specifically, a 36% reduction of DA levels was identified in 12-month-old Pitx3cKO mice, compared to the age-matched Pitx3cWT mice (Figure 2C). Besides dopamine, we also analyzed the contents of GABA and glutamate within the striatum of mice at the age of 6 and 12 months, respectively. GABA as the primary neurotransmitter of MSNs was substantially decreased in 6-month-old Pitx3cKO, at which point glutamate also showed a significant reduction in the model (Figure 2C), but there were no significant changes in these two neurotransmitters between the two genotypes at the late stage, suggesting that the striatal GABA and glutamate displayed age-dependent alterations in our model.
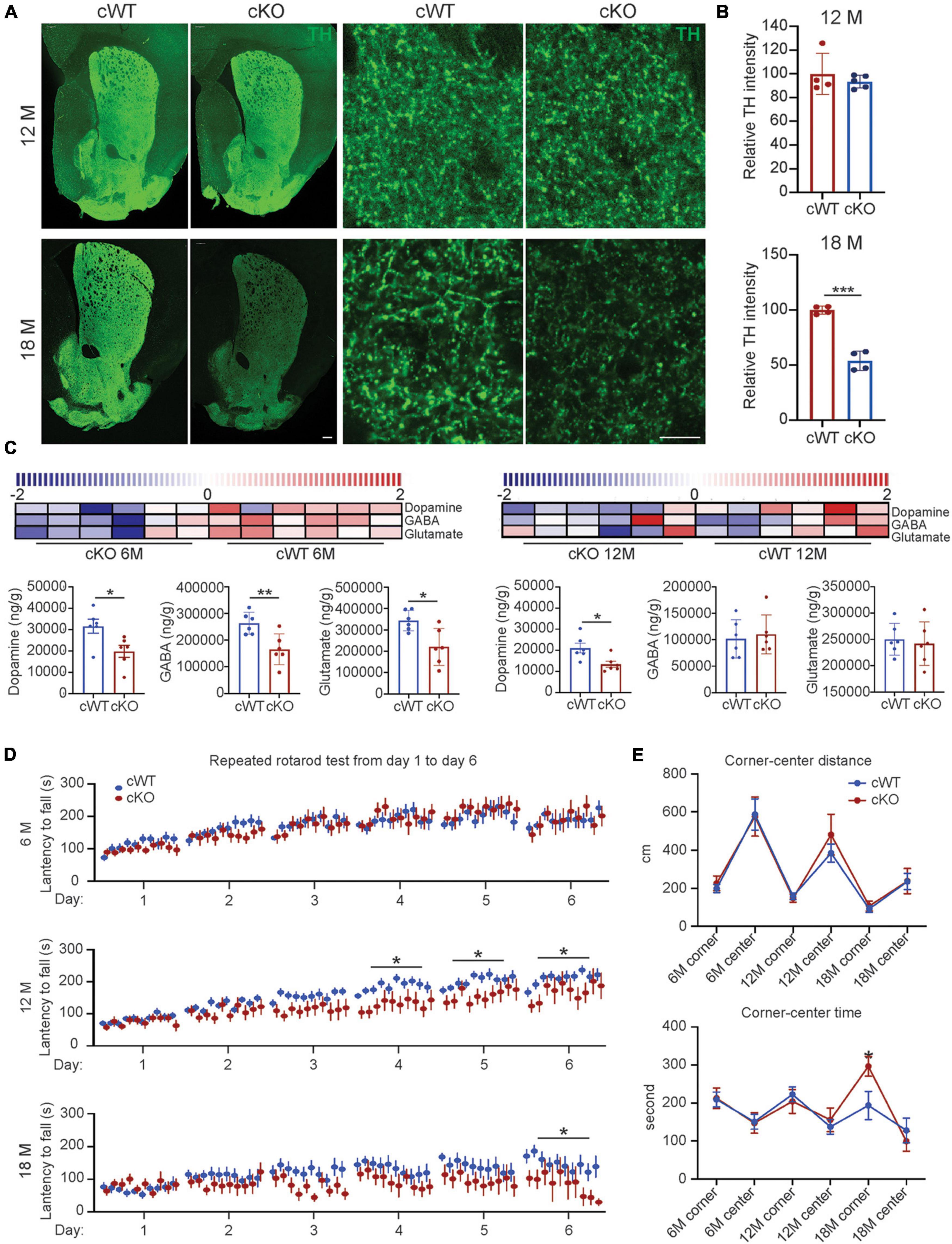
Figure 2. Striatal pathology triggered movement abnormalities in Pitx3cKO mice. (A) IFC staining of TH in the striatal sections from 12- to 18-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 200 μm; high-magnification, 10 μm). (B) Quantification of relative TH intensity in the striatum from 12- to 18-month-old Pitx3cWT and Pitx3cKO mice (N = 4 mice per genotype; all males except for two females in 6-month-old Pitx3cKO and 12-month-old Pitx3cWT). Unpaired t-test, ***p = 0.0006 (18 months). (C) Levels of neurotransmitter in 6- (N = 6 mice per genotype; all males) and 12-month-old Pitx3cKO and Pitx3cWT mice (N = 6 mice per genotype; all males). The scaled intensity of three metabolites is relatively depicted according to the color key shown on the above. Red indicates high intensity levels; blue, low intensity levels. Unpaired t-test, *p = 0.026 (Dopamine, 6M), **p = 0.0083 (GABA, 6M), *p = 0.012 (Glutamate, 6M), *p = 0.0202 (Dopamine, 12M). (D) The latency to fall from rotarod was recorded from Pitx3cWT and Pitx3cKO mice at 6 (N = 11–13 mice per genotype; all males), 12 (N = 11–15 per genotype; all males) and 18 months of age (N = 9–11 mice per genotype; all males). 2way ANOVA analysis with Sidak’s multiple comparisons test at 12 and 18 months, *p = 0.0171 (day 4, 12 months), *p = 0.0202 (day 5, 12 months), *p = 0.0376 (day 6, 12 months), *p = 0.0326 (day 6, 18 months). (E) Center-corner preference analyses for Pitx3cWT and Pitx3cKO mice at 6 (N = 11–13 mice per genotype; all males), 12 (N = 12–14 per genotype; all males), and 18 (N = 9–10 mice per genotype; all males) months of age. 2way ANOVA analysis with Sidak’s multiple comparisons test, *p = 0.05 (time in corner at 18 months).
The remarkable striatal pathology may contribute to motor behavioral abnormalities (Valentin et al., 2016). We applied a well-adopted repeated accelerating rotarod test (Yin et al., 2009) to evaluate motor skill learning of mice. 6-month-old Pitx3cKO mice performed equally well with age-matched Pitx3cWT mice during the 6-day trials, while 12-month-old Pitx3cKO mice showed markedly fewer improvements after the first 3 days’ training (Figure 2D). Surprisingly, a severely disrupted motor learning phenotype was observed in 18-month-old Pitx3cWT mice, and Pitx3-deficiency can further affect the last day’s training at this advanced stage (Figure 2D). Furthermore, we monitored the voluntary movement of Pitx3cKO mice in an open-field test at the age of 6, 12, and 18 months. Overall, multiple elements of locomotor activity were strongly age-dependent rather than genotype-dependent, such as distance, rearing and walking speed (Supplementary Figure 5). Additionally, in center-corner behavioral tests, 18-month-old Pitx3cKO mice prefer to stay in the corner for a longer time, compared to age-matched Pitx3cWT mice (Figure 2E), suggesting that anxiety levels may be increased. However, the distance traveled in the central zone did not vary between the two genotypes and further investigation may be required, such as an elevated plus-maze test (Rodgers and Dalvi, 1997).
To examine individual MSN morphology, we stereotactically injected AAV1 vectors into the striatum of 6- and 12-month-old Pitx3cWT and Pitx3cKO mice, which express a green fluorescent protein (GFP) under the control of synapsin1 promoter. Due to the low viral titer managed, only a few MSNs with GFP signals could be identified in each hemisphere. We subsequently analyze the dendritic complexity by using 3D reconstruction of individual MSN dendritic trees (Figure 3A and Supplementary Figure 3). The results revealed that a profound reduction in the cumulative length of all dendrites was observed in the MSNs of 12-month-old Pitx3cKO mice compared to Pitx3cWT mice (Figures 3A–E). Additionally, dendritic atrophy was age-dependent, since we did not detect any apparent alterations in MSN dendritic length in Pitx3cKO mice at the age of 6 months (Supplementary Figure 3).
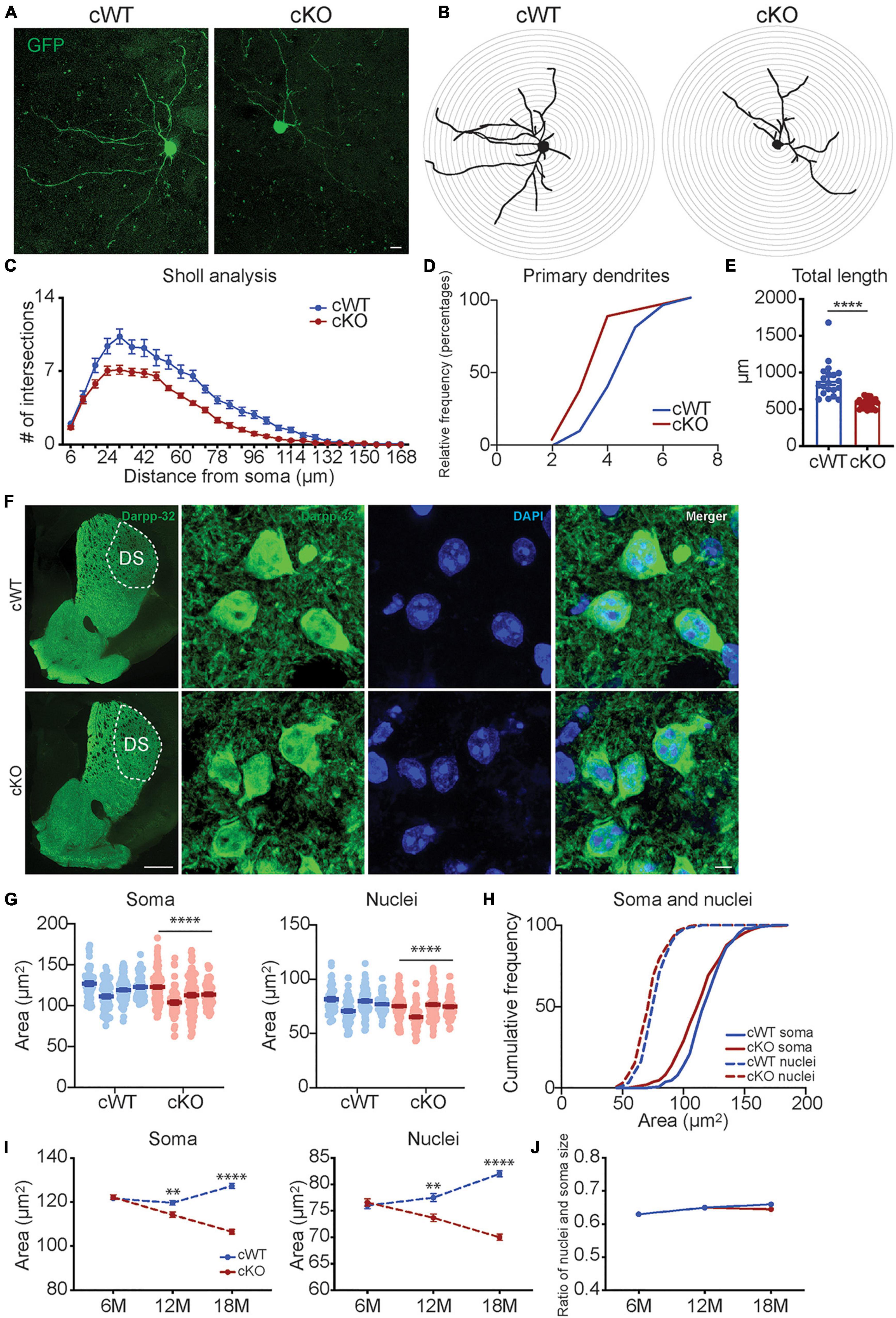
Figure 3. Analyses of neuronal morphology in Pitx3cKO mice during aging. (A) The GFP-labeled individual MSN in 12-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 10 μm). (B,C) Sholl analysis of dendritic complexity of GFP-labeled MSNs in 12-month-old Pitx3cWT and Pitx3cKO mice (N = 3 mice per genotype; 5–8 neurons per mouse were counted). Benjamin- Hochberg multiple comparison test of dendritic complexity at 18, 24, 30, 36, 42, 54, 60, 66, 72, 78, 84, 90, 96, 102, 108, and 114 μm from soma, q ≤ 0.05. (D) Analyses of primary dendrites (N = 3 mice per genotype; 5–8 neurons per mouse were counted; all males). (E) Dendritic length of GFP-labeled MSNs in 12-month-old Pitx3cWT and Pitx3cKO mice (N = 3 mice per genotype; 5–8 neurons per mouse were counted) unpaired t-test, ****p < 0.0001. (F) Co-staining of Darpp32 and DAPI in MSNs of 12-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 500 μm; high-magnification, 5 μm). (G) The soma and nucleus size of MSNs in 12-month-old Pitx3cWT and Pitx3cKO mice (N = 4 mice per genotype; all males). Conditional logistic regression test, ****p < 0.0001. (H) Cumulative frequency of the soma and nuclear size distribution in MSNs of 12-month-old Pitx3cWT and Pitx3cKO mice. (I) The soma and nucleus size of MSNs in Pitx3cWT and Pitx3cKO mice at 6 (N = 4 mice per genotype; all males), 12 (N = 4 mice per genotype; all males) and 18 months (N = 3 mice per genotype; all males) of age. 2way ANOVA analysis with Sidak’s multiple comparisons test, **p = 0.0075 (soma, 12 months), ****p < 0.0001 (soma, 18 months), **p = 0.0022 (nuclei, 12 months), ****p < 0.0001 (nuclei, 18 months). (J) The nuclear size and soma size ratio (N/C ratio) of MSNs in Pitx3cWT and Pitx3cKO mice. DS, dorsal striatum.
We further explored the soma morphology of MSNs in 6-, 12-, and 18-month-old Pitx3cWT and Pitx3cKO mice and found a marked reduction of the soma and nuclear size in Pitx3cKO during aging (Figures 3F–I). Our longitudinal data demonstrated that the soma and nuclear size of MSNs were steadily increased in Pitx3cWT mice from 6 to 18 months of age (Figure 3I). In contrast, the decreased sizes of soma and nucleus in Pitx3cKO mice were observed from 6 to 18 months of age (Figure 3I). Despite the alterations in the soma and nuclear size, the nucleus to soma ratio (N/C ratio) remained unchanged (Figure 3J). Besides nuclear size, the nuclear shape was altered as well during aging (Figures 4A,B). The increased nuclear invaginations were characterized in 18-month-old Pitx3cKO mice by immunofluorescent staining, where the nuclear envelope marker LaminB and MSN-specific nuclear marker Ctip2 were used (Figures 4A,B). Additionally, the nuclear pore structures were also identified in the enfolded nuclear envelope (Figure 4C), showing the presence of type II nuclear invagination in the MSNs, i.e., both outer and inner nuclear membranes were enfolded. Furthermore, we identified clusters of mitochondria near the mouth of nuclear invagination within striatal neurons (Figure 4D), which may provide extra ATP and/or calcium buffering capacity to protect against the further deformation of nuclear structures (Drozdz and Vaux, 2017). Nuclear invagination was reported to be closely associated with the expressions of γH2AX, a marker for DNA double-strand breaks and damage (Valdiglesias et al., 2013). We thereby examined the DNA stability in the MSNs of Pitx3cWT and Pitx3cKO mice at 12 and 18 months of age. Our results showed a substantial increase in the percentage of MSNs with 10 or more γH2AX-positive foci in the nuclei of 18-month-old Pitx3cKO mice compared to Pitx3cWT (Figures 4E,F), corresponding to the observed alterations of nuclear shape. Together, these data suggest that the early neurotransmitters’ disruptions may be involved in regulating nuclear and soma morphology during aging.
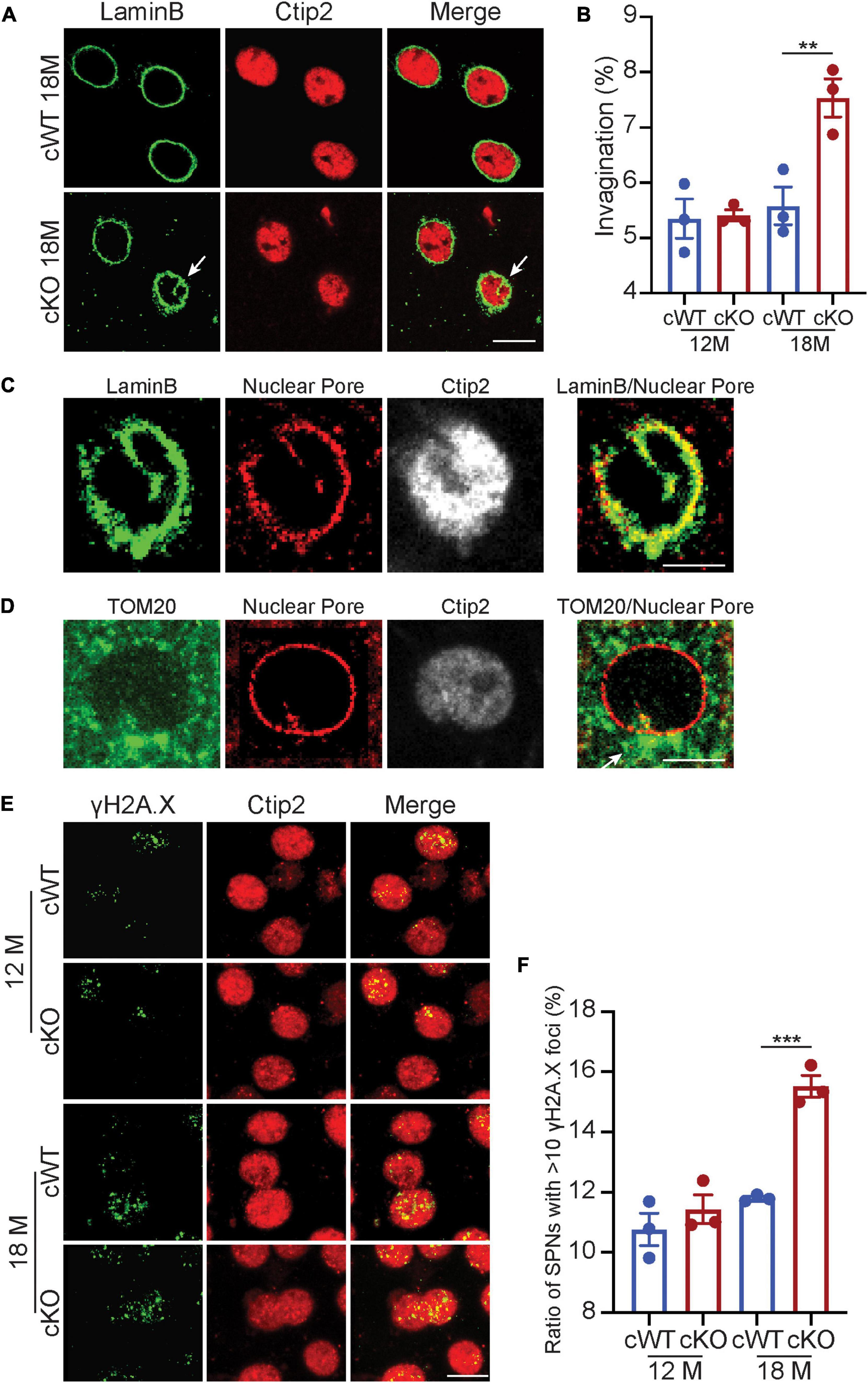
Figure 4. Nuclear invaginations increase accompanied with genomic instability in Pitx3cKO mice. (A) Co-staining of LaminB and Ctip2 in MSNs of 18-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 10 μm) white arrow points to nuclear invagination. (B) Ratio of MSN nuclei containing ≥ 1 invagination in Pitx3cWT and Pitx3cKO mice at 12 (N = 3 mice per genotype; all males) and 18 (N = 3 mice per genotype; all males) month of age. Unpaired t-test, **p = 0.0041. (C) Co-staining of Nuclear Pore and LaminB (scale bar: 5 μm). (D) Costaining of Nuclear Pore and TOM20 (scale bar: 5 μm). White arrow points to a cluster of mitochondria. (E) Co- staining of γH2A.X and Ctip2 in the striatal sections of 12- and 18-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 10 μm). (F) The ratios of MSNs with 10 or more γH2A.X-positive foci in the nuclei (N = 3 mice per genotype). Unpaired t-test, ***p = 0.0005.
To further investigate the role of epigenetics on the whole genome within striatal cells, we used MethylRAD sequencing to analyze the DNA methylation at CG and CWG (W for A or T) sites of Pitx3cWT and Pitx3cKO mice’ genome at the age of 12 and 18 months. Overall, the total DNA methylation ratios on CG (methylated CG/total CG sites) were greatly decreased in 18-month-old Pitx3cWT and Pitx3cKO (Supplementary Figure 6 and Supplementary Table 1), indicating that aging plays an important role in the global DNA methylation changes. We further examined the distribution patterns of CG methylation sites at the different elements of the genome in all 12 samples (Supplementary Figure 6). The CG methylated sites were concentrated in the introns, followed by the exon and intergenic regions (Supplementary Figure 6). Since CG is more predominant within DNA methylation, we only considered the data from CG sites for the subsequent analyses.
Besides the global genome examination, the DNA methylation levels of individual genes were also evaluated by summing the methylation levels of sites localized in the gene regions. An analysis of differentially methylated genes (DMGs) was conducted for all the samples. We found 182 DMGs (96 hypermethylated and 86 hypomethylated genes) at 12 months and 262 DMGs (154 hypermethylated and 108 hypomethylated genes) at 18 months, respectively (Figure 5A). Further gene-network studies indicated that the DMGs at 12 months are involved in olfaction and mitochondria transportation pathway, whereas the DMGs at 18 months participate in lipoprotein and nucleus pathway, which may be associated with nuclear morphological changes at this advanced stage (Figure 5C). We further identified the DMGs related to normal aging, characterized from 12- to 18-month-old Pitx3cWT mice. These genes are involved in multiple cellular process, including aging, glycolytic process, synapse assembly, regulation of translation and mitochondrion organization, consistent with the previous findings (Figure 5B). After crossing these DMGs with the ones characterized from 12- to 18-month-old Pitx3cKO mice, we totally identified 448 genes, and the alterations of their methylation levels were independent of genotype during aging (Figure 5D). Of them, hypermethylated genes are mainly involved in presynaptic membrane assembly and neural tube development, and hypomethylated genes preferentially participate in xenobiotic glucuronidation and regulation of transcription (Figure 5D). Notably, in the metabolic pathway analysis, retinol metabolism was affected largely, suggesting that retinoic acid within striatal cells could be regulated specifically by the epigenetic way during aging (Figure 5E). Together, these data imply that the DNA methylation modes alters with aging and genotype and may affect multiple cellular process, including retinol metabolism and nuclear pathway, which are potentially associated with the striatal pathology at the advanced stage.
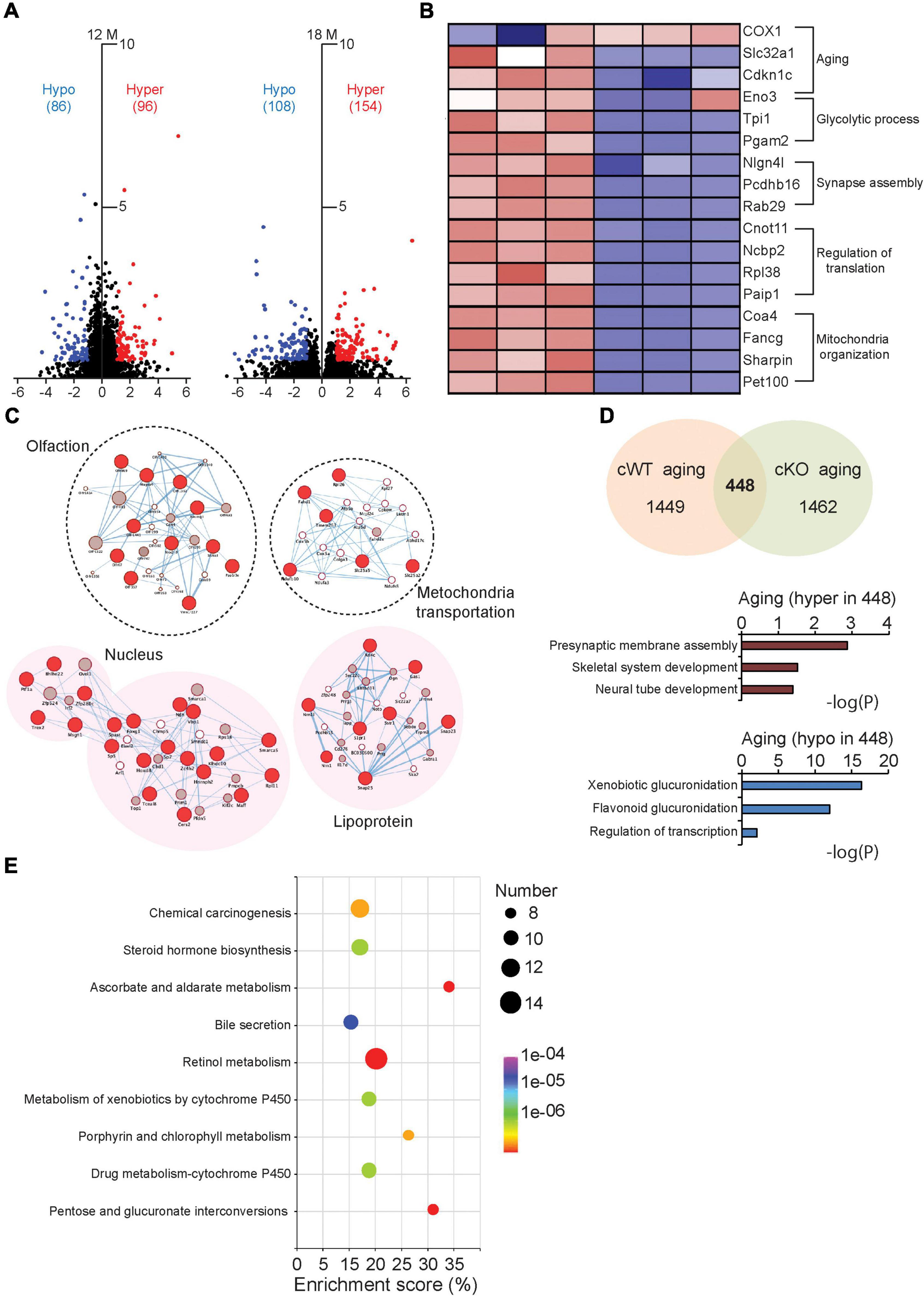
Figure 5. Comparative analysis of the DMGs in Pitx3cWT and Pitx3cKO during aging. (A) The volcano plots of DNA methylation data collected from the striatum of Pitx3cWT and Pitx3cKO mice at 12 (N = 3 mice per genotype; all males) and 18 months of age (N = 3 mice per genotype; all males). (B) Supervised clustering for the DMGs data collected from the 12- and 18-month-old Pitx3cWT mice. (C) Integrated map of GO terms enriched among the DMGs of Pitx3cWT and Pitx3cKO mice at 12 (background with white color) and 18 months of age (background with pink color). Red circles represented DMGs identified from our data. (D,E) 448 DMGs were identified during aging independent of genotype. They were analyzed with GO terms (D) and metabolic pathway (E), respectively.
Here we demonstrated that Pitx3cKO mice showing PD-related features, such as reduced DA and DAergic neuronal degeneration. Meanwhile, besides DA, the homeostasis of GABA and glutamate was impaired at the early stage in the model, potentially contributing to the striatal pathologies at the late stage. Furthermore, we novelly characterized nuclear atrophy and nuclear invagination increase in MSNs during aging, and these aberrant nuclear phenotypes may be associated with epigenetic alterations at the advanced stage.
In PD, the deficit of midbrain DAergic neurons produces the reduction of DA in the basal ganglia (Weintraub et al., 2022). Our Pitx3cKO mice showed a significant reduction of DA at 6 months, and the deficit aggravated at the late stage. Thus, the model provided great potential for studying the age-dependent striatal pathologies under progressive DA depletion. First, we examined the levels of two main neurotransmitters, GABA and glutamate, since they may be involved in remodeling the plasticity of MSNs, synergistically with DA. Interestingly, GABA and glutamate were both decreased at the early stage of our model, compared to controls. Previously the remarkable alterations of GABAergic neurotransmission within the basal ganglia circuit were reported in PD (Jamwal and Kumar, 2019). Moreover, in MPTP mice, the decreased levels of GABA in the striatum have been characterized, where about 75–80% of SNc were lost (Singh et al., 2017). However, in our studies, the significantly altered GABA levels were only characterized in the young Pitx3cKO mice, i.e., there were no changes in GABA levels between the two genotypes at the advanced stage, indicating that an adaptive system may respond to restoring the GABA levels in our model during aging. On the other hand, like GABA, the perturbation of glutamate homeostasis also altered in our model in an age-dependent way, i.e., the notably changed glutamate levels in Pitx3cKO mice were only identified in the early stage, but not in the late stage. However, our glutamate results were inconsistent with the previous studies that the enhancement of glutamate content was associated with the robust MSNs hyperactivity in the PD-related animal models (Calabresi et al., 1993; Singh et al., 2015; Tozzi et al., 2021). Noticeably, these models were majorly administrated with 6-OHDA, MPTP, or α-syn-PFF, rather than genetic models. One of the outstanding features of these models is the occurrence of the severe loss of DAergic neurons, usually reaching 70–80%, whereas, in genetic models, the death rate of DAergic neurons typically reached 40–50% (Konnova and Swanberg, 2018). Thus, the differences in DAergic neuron loss among the animal models may affect the glutamate release in the striatum. Additionally, when determining neurotransmitter levels, we used whole striatum tissue extracts for HPLC analysis, which may compromise the subtle changes occurring at the extracellular/synaptic levels. In the future, the use of microdialysis will be another better choice. Taken together, the neurotransmitter levels were altered at 6 months, while the MSNs of Pitx3cKO mice remained integrity at that time. During aging, the atrophy of dendritic complexity, soma and nuclei was identified in the MSNs of 12-month-old Pitx3cKO mice, concomitated with the significant loss of SNc neurons and movement abnormalities. We thereby suggested that the early perturbation of neurotransmitters may progressively trigger the striatal pathologies.
MSNs in our model have shorter dendritic lengths and lower maximal branch order of the dendrites at 12 months. This phenotype is similar to what is observed in patients and the animal models of PD (McNeill et al., 1988; Toy et al., 2014). Unlike ak and traditional Pitx3 knockout mice, MSNs develop and mature with abundant DA afferents in our genetic model. Thus, the observed defects of tree complexity exclude the cause of developmental defects. Besides dendritic shortage, we identified nuclear morphological alterations in MSNs. Irregular shapes of nuclei have been reported in the neurons of PD patients with LRRK2-related G2019S (Liu et al., 2012; Shani et al., 2019), transgenic mice carrying R1441C mutations (Tsika et al., 2014) and LRRK2 knockout mice (Chen et al., 2020). As an extension of these findings, our present studies demonstrated that MSNs showed reduced nuclear size and increased nuclear invagination during aging. The decreased size of the nucleus likely reflects lower biosynthetic activities of DNA repair/synthesis, transcription, and translation in cells (Koda et al., 2006), which potentially contributed to the striatal neuronal dysfunction at the advanced stage. On the other hand, in our mouse model, increased nuclear invagination was characterized at the advanced stage, reflecting that the higher neuronal excitability might occur over there (Chen et al., 2020). Comparably it was reported that the depolarizing current significantly evoked more action potentials in MSNs of Pitx3 knockout mice (Suarez et al., 2018). Thus, our results re-emphasize that increased neural activity facilitates nuclear invagination formation. Together, not only the dendritic complexity but also the nuclear morphology alters in MSNs upon PD-like stress during aging. Especially, the aberrant nuclear phenotype may bring out severe effects on genomics, and further impact the downstream cellular progresses.
Age-related remodeling of DNA methylation comprises events of both hypo- and hypermethylation (Miranda-Morales et al., 2017; Yang et al., 2022). These epigenetic alterations mediated heritable changes in the gene activity and contributed to genomic instability. Our data showed that increased γH2AX was observed in the MSNs of 18-month-old Pitx3cKO mice, indicating that DNA damage might be persistent and accumulative during the aging process. To further elucidate the whole-genome epigenetic changes during aging, we performed DNA methylation studies between Pitx3cKO and Pitx3cWT mice at 12 and 18 months. Our data showed that the total DNA methylation ratios were greatly decreased in 18-month-old mice compared to 12-month-old ones, consistent with previous studies that DNA hypomethylation occurs globally over time (Miranda-Morales et al., 2017). For methylation onto individual genes, the previous studies reported that the MAPT gene was hypomethylated in the putamen of PD patients’ post-mortem brains (Coupland et al., 2014). Whereas in our longitudinal epigenetic data, many DMGs involved in the nucleus pathway were characterized in Pitx3cKO mice at the advanced stage, potentially associating with their nuclear morphological alterations. However, what factor contributed to the changes in DNA methylation profile? Recent studies indicated that DA could modify histone H3 glutamine 5 (H3Q5dop) to regulate cocaine-induced transcriptional plasticity in the midbrain (Lepack et al., 2020). The non-neurotransmission roles of neurotransmitters in the epigenetic process have attracted more attention recently. In our model, the progressive reduction of DA and temporally altered neurotransmitters have been identified, thus the perturbation of neurochemicals might affect the epigenetic changes and further remodel the neuronal plasticity. More detailed biochemical and molecular mechanisms of how neurotransmitters regulate neuronal plasticity are also needed in the future.
Previously, we reported another Pitx3 conditional knockout model, Pitx3fl/fl/DATCreERT2 mice (Wang et al., 2021). Similar to the DAT model, our model has progressive DAergic neuron degeneration and DA reduction during aging, greatly different from ak and Pitx3–/– mice that showed about 60% SNc neuron loss and 80% striatal DA reduction as early as the age of 2 months. One of the reasons why our model facilitates the development of DAergic neurons may be that Pitx3 in SNc is expressed earlier than TH during development (Maxwell et al., 2005). Thus, prior to Cre recombination, some Pitx3 proteins are already present and exert the biological function, i.e., a serials of downstream development events have been triggered. Furthermore, Pitx3 as a transcription factor is considered to modulate TH expressions (Cazorla et al., 2000). Therefore, decreased Pitx3 expressions may reduce the efficiency of the TH-driven Cre-loxp system. Taken together, these contributing factors may aid in the development of DAergic neurons in our model. However, whether the development of DAergic neurons is intrinsically affected in our model needs to be further investigated by examining the expression levels of development-related molecules, such as DAT, Vmat2, Aadc, and so on.
The striatum is the key player in facilitating voluntary movement. In PD, the striatal neurons undergo the progressive depletion of DA, resulting in impaired physiological function and contributing to the motor symptom of PD. To further explore the striatal pathologies during aging, we generated the Pitx3cKO mice, where a progressive reduction of striatal DA was identified. In this model, the levels of GABA and glutamate decreased at the early stage besides DA. Such early disturbance of neurochemical homeostasis may contribute to the longitudinal plasticity remodeling of neurons, including morphology and movement abnormalities as well as aberrant epigenetic modifications. Our studies may expand the overview of PD treatments and provide a new potential window for therapeutic strategies.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://submit.ncbi.nlm.nih.gov/subs/bioproject/SUB11723472/overview.
This animal study was reviewed and approved by the Institutional Animal Care Committee at Dalian Medical University.
XC and WL designed the experiments, wrote, and edited the manuscript. ZY contributed to stereotactic injection. XC and YS contributed to metabolic analyses. XC and YiW contributed to imaging experiments and data analysis. KK, YuW, and HW contributed to behavior test and data analysis. XC and XX contributed to sample preparation and data analysis for DNA methylation. All authors read and approved the final manuscript.
The youth program of National Natural Science Foundation of China (81901405 to XC), the Key Research and Development Program of Sichuan (2021YFS0382 to XC), and the Key Project of the Medical Science Department, University of Electronic Science and Technology of China (ZYGX2020ZB035 to WL).
We are cordially thankful to Marten P. Smidt (University of Amsterdam) for providing Pitx3 antibody, and to all members in our laboratory for caring, help, and advice.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2022.960479/full#supplementary-material
Supplementary Figure 1 | Conditional knockout of Pitx3 in DA neurons. (A) The basic strategy for the generation of THCre/Pitx3fl/fl knockout mice. (B) PCR detection of Cre transgene (upper) and Pitx3 floxed allele (lower). (C) IFC staining for Pitx3 expression in DA neurons was performed using an antibody against Pitx3 (green) together with TH (red) in 2-month-old Pitx3cWT and Pitx3cKO mice (scale bar: 100 μm; high-magnification, 5 μm).
Supplementary Figure 2 | Specific labeling of DAergic neurons using RiboTag mice (Scale bar: 100 μm).
Supplementary Figure 3 | Analyses of dendritic complexity in 6-month-old Pitx3cKO mice. (A) The GFP-labeled individual MSN (scale bar: 10 μm). (B,C) Sholl analysis of dendritic complexity of GFP-labeled MSNs (N = 3 mice per genotype; 5–8 neurons per mouse were counted; all males). (D) Analyses of primary dendrites (N = 3 mice per genotype; 5–8 neurons per mouse were counted; all males). (E) Dendritic length of GFP-labeled MSNs (N = 3 mice per genotype; 5–8 neurons per mouse were counted; all males).
Supplementary Figure 4 | TH+ neurons in 12 and 18-month-old Pitx3cWT mice (scale bar: 200 μm).
Supplementary Figure 5 | Open field test for Pitx3cWT and Pitx3cKO mice at 6 (N = 11–13 mice per genotype; all males), 12 (N = 12–14 mice per genotype; all males), and 18 months of age (N = 9–10 mice per genotype; all males).
Supplementary Figure 6 | Overview of the DNA methylation ratio and distribution of CG sites in Pitx3cWT and Pitx3cKO mice. (A) The genomic DNA methylation ratio at CG and CWG sites in 12- and 18-month-old Pitx3cWT and Pitx3cKO. (B) The distribution of CG sites on different functional genic components. Upstream, downstream, exon, and intron indicate the regions 2,000 bp upstream of the transcription start site, the regions 2,000 bp downstream of the transcription terminal site, the whole exons of genes, and the whole introns of genes, respectively. 3′UTR and 5′UTR indicate the regions at the 3′ end and 5′ end of a mature transcript that are not translated into a protein. Intergenic indicates the intergenic regions.
Supplementary Table 1 | DNA methylation ratios in 12- and 18-month-old Pitx3cWT and Pitx3cKO mice.
PD, Parkinson’s disease; MSNs, spiny projection neurons; DA, dopamine; TH, tyrosine hydroxylase; SNc, substantia nigra; GABA, γ-aminobutyric acid; ak, aphakia; VTA, ventral tegmental area; GFP, green fluorescent protein; DMGs, differentially methylated genes.
Althini, S., Bengtsson, H., Usoskin, D., Söderström, S., Kylberg, A., Lindqvist, E., et al. (2003). Normal nigrostriatal innervation but dopamine dysfunction in mice carrying hypomorphic tyrosine hydroxylase alleles. J. Neurosci. Res. 15, 444–453. doi: 10.1002/jnr.10606
Anders, S., and Huber, W. (2010). Differential expression analysis for sequence count data. Genome Biol. 11:R106. doi: 10.1186/gb-2010-11-10-r106
Bariselli, S., Fobbs, W. C., Creed, M. C., and Kravitz, A. V. (2019). A competitive model for striatal action selection. Brain Res. 1713, 70–79. doi: 10.1016/j.brainres.2018.10.009
Bolam, J. P., Hanley, J. J., Booth, P. A., and Bevan, M. D. (2000). Synaptic organisation of the basal ganglia. J. Anat. 196, 527–542. doi: 10.1046/j.1469-7580.2000.19640527.x
Calabresi, P., Mercuri, N. B., Sancesario, G., and Bernardi, G. (1993). Electrophysiology of dopaminedenervated striatal neurons. Implications for Parkinson’s disease. Brain 116, 433–52.
Cazorla, P., Smidt, M. P., O’Malley, K. L., and Burbach, J. P. (2000). A response element for the homeodomain transcription factor Ptx3 in the tyrosine hydroxylase gene promoter. J. Neurochem. 74, 1829–1837. doi: 10.1046/j.1471-4159.2000.0741829.x
Chen, X., Xie, C., Tian, W., Sun, L., Zheng, W., Hawes, S., et al. (2020). Parkinson’s disease-related Leucine-rich repeat kinase 2 modulates nuclear morphology and genomic stability in striatal projection neurons during aging. Mol. Neurodegener. 15:12. doi: 10.1186/s13024-020-00360-0
Cingolani, P., Platts, A., Wang le, L., Coon, M., Nguyen, T., Wang, L., et al. (2012). A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 6, 80–92. doi: 10.4161/fly.19695
Coupland, K. G., Mellick, G. D., Silburn, P. A., Mather, K., Armstrong, N. J., Sachdev, P. S., et al. (2014). DNA methylation of the MAPT gene in Parkinson’s disease cohorts and modulation by vitamin E in vitro. Mov. Disord. 29, 1606–1614. doi: 10.1002/mds.25784
Day, M., Wang, Z., Ding, J., An, X., Ingham, C. A., Shering, A. F., et al. (2006). Selective elimination of glutamatergic synapses on stratopallidal neurons in Parkinson disease models. Nat. Neurosci. 9, 251–259. doi: 10.1038/nn1632
Dong, J., Liu, X., Wang, Y., Cai, H., and Le, W. (2020). Nurr1(Cd11bcre) conditional knockout mice display inflammatory injury to nigrostriatal dopaminergic neurons. Glia 68, 2057–2069. doi: 10.1002/glia.23826
Drozdz, M. M., and Vaux, D. J. (2017). Shared mechanisms in physiological and pathological nucleoplasmic reticulum formation. Nucleus 8, 34–45. doi: 10.1080/19491034.2016.1252893
Du, Y., and Graves, S. M. (2019). Spiny Projection Neuron Dynamics in Toxin and Transgenic Models of Parkinson’s Disease. Front. Neural Circuits 13:17. doi: 10.3389/fncir.2019.00017
Feng, L., Zhao, T., and Kim, J. (2015). neuTube 1.0: A New Design for Efficient Neuron Reconstruction Software Based on the SWC Format. eNeuro 2:ENEURO.0049–14.2014. doi: 10.1523/ENEURO.0049-14.2014
Filali, M., and Lalonde, R. (2016). Neurobehavioral Anomalies in the Pitx3/ak Murine Model of Parkinson’s Disease and MPTP. Behav. Genet. 46, 228–241. doi: 10.1007/s10519-015-9753-3
Filipovic, M., Ketzef, M., Reig, R., Aertsen, A., Silberberg, G., and Kumar, A. (2019). Direct pathway neurons in mouse dorsolateral striatum in vivo receive stronger synaptic input than indirect pathway neurons. J. Neurophysiol. 122, 2294–2303. doi: 10.1152/jn.00481.2019
Fu, Y., Yuan, Y., Halliday, G., Rusznak, Z., Watson, C., and Paxinos, G. (2012). A cytoarchitectonic and chemoarchitectonic analysis of the dopamine cell groups in the substantia nigra, ventral tegmental area, and retrorubral field in the mouse. Brain Struct. Funct. 217, 591–612. doi: 10.1007/s00429-011-0349-2
Gerfen, C. R., Engber, T. M., Mahan, L. C., Susel, Z., Chase, T. N., and Monsma, F. J. Jr., et al. (1990). D1 and D2 dopamine receptor-regulated gene expression of stratonigral and striatopallidal neurons. Science 250, 1429–1432. doi: 10.1126/science.2147780
Gong, S., Zheng, C., Doughty, M. L., Losos, K., Didkovsky, N., Schambra, U. B., et al. (2003). A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature 425, 917–925. doi: 10.1038/nature02033
He, J., Kleyman, M., Chen, J., Alikaya, A., Rothenhoefer, K. M., Ozturk, B. E., et al. (2021). Transcriptional and anatomical diversity of medium spiny neurons in the primate striatum. Curr. Biol. 31, 5473–5486.e6. doi: 10.1016/j.cub.2021.10.015
Hwang, D. Y., Ardayfio, P., Kang, U. J., Semina, E. V., and Kim, K. S. (2003). Selective loss of dopaminergic neurons in the substantia nigra of Pitx3-deficient aphakia mice. Brain Res. Mol. Brain Res. 114, 123–131. doi: 10.1016/S0169-328X(03)00162-1
Jamwal, S., and Kumar, P. (2019). Insight Into the Emerging Role of Striatal Neurotransmitters in the Pathophysiology of Parkinson’s Disease and Huntington’s Disease: A Review. Curr. Neuropharmacol. 17, 165–175. doi: 10.2174/1570159X16666180302115032
Koda, M., Takemura, G., Okada, H., Kanoh, M., Maruyama, R., Esaki, M., et al. (2006). Nuclear hypertrophy reflects increased biosynthetic activities in myocytes of human hypertrophic hearts. Circ. J. 70, 710–718. doi: 10.1253/circj.70.710
Konnova, E. A., and Swanberg, M. (2018). “Chapter 5: Animal models of Parkinson’s disease,” in Parkinson’s disease: Pathogenesis and clinical aspects [Internet], ed. T. B. Stoker and J. C. Greenland (Brisbane, AU: Codon Publications).
Lepack, A. E., Werner, C. T., Stewart, A. F., Fulton, S. L., Zhong, P., Farrelly, L. A., et al. (2020). Dopaminylation of histone H3 in ventral tegmental area regulates cocaine seeking. Science 368, 197–201. doi: 10.1126/science.aaw8806
Liu, G. H., Qu, J., Suzuki, K., Nivet, E., Li, M., Montserrat, N., et al. (2012). Progressive degeneration of human neural stem cells caused by pathogenic LRRK2. Nature 491, 603–607. doi: 10.1038/nature11557
Longair, M. H., Baker, D. A., and Armstrong, J. D. (2011). Simple Neurite Tracer: Open source software for reconstruction, visualization and analysis of neuronal processes. Bioinformatics 27, 2453–2454. doi: 10.1093/bioinformatics/btr390
Lu, X. H., and Yang, X. W. (2017). Genetically-directed Sparse Neuronal Labeling in BAC Transgenic Mice through Mononucleotide Repeat Frameshift. Sci. Rep. 7:43915. doi: 10.1038/srep43915
Maxwell, S. L., Ho, H. Y., Kuehner, E., Zhao, S., and Li, M. (2005). Pitx3 regulates tyrosine hydroxylase expression in the substantia nigra and identifies a subgroup of mesencephalic dopaminergic progenitor neurons during mouse development. Dev. Biol. 282, 467–479. doi: 10.1016/j.ydbio.2005.03.028
McNeill, T. H., Brown, S. A., Rafols, J. A., and Shoulson, I. (1988). Atrophy of medium spiny I striatal dendrites in advanced Parkinson’s disease. Brain Res. 455, 148–152. doi: 10.1016/0006-8993(88)90124-2
Miranda-Morales, E., Meier, K., Sandoval-Carrillo, A., Salas-Pacheco, J., Vazquez-Cardenas, P., and Arias-Carrion, O. (2017). Implications of DNA Methylation in Parkinson’s Disease. Front. Mol. Neurosci. 10:225. doi: 10.3389/fnmol.2017.00225
Parisiadou, L., Yu, J., Sgobio, C., Xie, C., Liu, G., Sun, L., et al. (2014). LRRK2 regulates synaptogenesis and dopamine receptor activation through modulation of PKA activity. Nat. Neurosci. 17, 367–376. doi: 10.1038/nn.3636
Prager, E. M., and Plotkin, J. L. (2019). Compartmental function and modulation of the striatum. J. Neurosci. Res. 97, 1503–1514.
Quinlan, A. R., and Hall, I. M. (2010). BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842. doi: 10.1093/bioinformatics/btq033
Rodgers, R. J., and Dalvi, A. (1997). Anxiety, defence and the elevated plus-maze. Neurosci. Biobehav. Rev. 21, 801–810.
Shani, V., Safory, H., Szargel, R., Wang, N., Cohen, T., Elghani, F. A., et al. (2019). Physiological and pathological roles of LRRK2 in the nuclear envelope integrity. Hum. Mol. Genet. 28, 3982–3996. doi: 10.1093/hmg/ddz245
Shigeoka, T., Jung, H., Jung, J., Turner-Bridger, B., Ohk, J., Lin, J. Q., et al. (2016). Dynamic axonal translation in developing and mature visual circuits. Cell 166, 181–192.
Singh, A., Liang, L., Kaneoke, Y., Cao, X., and Papa, S. M. (2015). Dopamine regulates distinctively the activity patterns of striatal ouput neurons in advanced parkinsonian primates. J. Neurophysiol. 113, 1533–1544.
Singh, S., Jamwal, S., and Kumar, P. (2017). Neuroprotective potential of Quercetin in combination with piperine against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity. Neural Regen. Res. 12, 1137–1144. doi: 10.4103/1673-5374.211194
Smidt, M. P., Smits, S. M., and Burbach, J. P. (2004). Homeobox gene Pitx3 and its role in the development of dopamine neurons of the substantia nigra. Cell Tissue Res. 318, 35–43.
Smidt, M. P., van Schaick, H. S., Lanctot, C., Tremblay, J. J., Cox, J. J., van der Kleij, A. A., et al. (1997). A homeodomain gene Ptx3 has highly restricted brain expression in mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci. U.S.A. 94, 13305–13310. doi: 10.1073/pnas.94.24.13305
Stephens, B., Mueller, A. J., Shering, A. F., Hood, S. H., Taggart, P., Arbuthnott, G. W., et al. (2005). Evidence of a breakdown of corticostriatal connections in Parkinson’s disease. Neuroscience 132, 741–754.
Suarez, L. M., Alberquilla, S., Garcia-Montes, J. R., and Moratalla, R. (2018). Differential Synaptic Remodeling by Dopamine in Direct and Indirect Striatal Projection Neurons in Pitx3(–/–) Mice, a Genetic Model of Parkinson’s Disease. J. Neurosci. 38, 3619–3630. doi: 10.1523/JNEUROSCI.3184-17.2018
Suarez, L. M., Solis, O., Carames, J. M., Taravini, I. R., Solis, J. M., Murer, M. G., et al. (2014). L-DOPA treatment selectively restores spine density in dopamine receptor D2-expressing projection neurons in dyskinetic mice. Biol. Psychiatry 75, 711–722. doi: 10.1016/j.biopsych.2013.05.006
Toy, W. A., Petzinger, G. M., Leyshon, B. J., Akopian, G. K., Walsh, J. P., Hoffman, M. V., et al. (2014). Treadmill exercise reverses dendritic spine loss in direct and indirect striatal medium spiny neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model of Parkinson’s disease. Neurobiol. Dis. 63, 201–209. doi: 10.1016/j.nbd.2013.11.017
Tozzi, A., Sciaccaluga, M., Loffredo, V., Megaro, A., Ledonne, A., Cardinale, A., et al. (2021). Dopaminedependent early synaptic and motor dysfunctions induced by α-synuclein in the nigrostriatal circuit. Brain 144, 3477–3791.
Tsika, E., Kannan, M., Foo, C. S., Dikeman, D., Glauser, L., Gellhaar, S., et al. (2014). Conditional expression of Parkinson’s disease-related R1441C LRRK2 in midbrain dopaminergic neurons of mice causes nuclear abnormalities without neurodegeneration. Neurobiol. Dis. 71, 345–358. doi: 10.1016/j.nbd.2014.08.027
Valdiglesias, V., Giunta, S., Fenech, M., Neri, M., and Bonassi, S. (2013). gammaH2AX as a marker of DNA double strand breaks and genomic instability in human population studies. Mutat. Res. 753, 24–40.
Valentin, V. V., Maddox, W. T., and Ashby, F. G. (2016). Dopamine dependence in aggregate feedback learning: A computational cognitive neuroscience approach. Brain Cogn. 109, 1–18. doi: 10.1016/j.bandc.2016.06.002
Villalba, R. M., Lee, H., and Smith, Y. (2009). Dopaminergic denervation and spine loss in the striatum of MPTP-treated monkeys. Exp. Neurol. 215, 220–227.
Wang, S., Lv, J., Zhang, L., Dou, J., Sun, Y., Li, X., et al. (2015). MethylRAD: A simple and scalable method for genome-wide DNA methylation profiling using methylation-dependent restriction enzymes. Open Biol. 5:150130. doi: 10.1098/rsob.150130
Wang, Y., Chen, X., Wang, Y., Li, S., Cai, H., and Le, W. (2021). The essential role of transcription factor Pitx3 in preventing mesodiencephalic dopaminergic neurodegeneration and maintaining neuronal subtype identities during aging. Cell Death Dis. 12:1008. doi: 10.1038/s41419-021-04319-x
Weintraub, D., Aarsland, D., Chaudhuri, K. R., Dobkin, R. D., Leentjens, A. F., Rodriguez-Violante, M., et al. (2022). The neuropsychiatry of Parkinson’s disease: Advances and challenges. Lancet Neurol. 21, 89–102.
Wu, J., Kung, J., Dong, J., Chang, L., Xie, C., Habib, A., et al. (2019). Distinct Connectivity and Functionality of Aldehyde Dehydrogenase 1a1-Positive Nigrostriatal Dopaminergic Neurons in Motor Learning. Cell Rep. 28, 1167–1181.e7. doi: 10.1016/j.celrep.2019.06.095
Yang, J., Qu, J., and Ma, H. (2022). Recent developments in understanding brain aging: Sex differences, mechanisms, and implications in diseases. Aging Neur. Dis. 2:3. doi: 10.1016/B978-0-444-53630-3.00006-3
Yin, H. H., Mulcare, S. P., Hilario, M. R., Clouse, E., Holloway, T., Davis, M. I., et al. (2009). Dynamic reorganization of striatal circuits during the acquisition and consolidation of a skill. Nat. Neurosci. 12, 333–341. doi: 10.1038/nn.2261
Zaja-Milatovic, S., Milatovic, D., Schantz, A. M., Zhang, J., Montine, K. S., Samii, A., et al. (2005). Dendritic degeneration in neostriatal medium spiny neurons in Parkinson disease. Neurology 64, 545–547.
Zhang, Y., Meredith, G. E., Mendoza-Elias, N., Rademacher, D. J., Tseng, K. Y., and Steece-Collier, K. (2013). Aberrant restoration of spines and their synapses in L-DOPA-induced dyskinesia: Involvement of corticostriatal but not thalamostriatal synapses. J. Neurosci. 33, 11655–11667. doi: 10.1523/JNEUROSCI.0288-13.2013
Keywords: medium spiny neurons, neuronal morphology, DNA methylation, aging, Parkinson’s disease
Citation: Chen X, Yang Z, Shao Y, Kim K, Wang Y, Wang Y, Wu H, Xu X and Le W (2022) Pitx3 deficiency promotes age-dependent alterations in striatal medium spiny neurons. Front. Aging Neurosci. 14:960479. doi: 10.3389/fnagi.2022.960479
Received: 03 June 2022; Accepted: 16 August 2022;
Published: 07 September 2022.
Edited by:
Robert Petersen, Central Michigan University, United StatesReviewed by:
Rahul Srinivasan, Texas A&M Health Science Center, United StatesCopyright © 2022 Chen, Yang, Shao, Kim, Wang, Wang, Wu, Xu and Le. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xi Chen, Y3hkZTIwMThAMTYzLmNvbQ==; Weidong Le, d2RsZUBzaWJzLmFjLmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.