 Le Zhang
Le Zhang Yingshuang Zhang
Yingshuang Zhang Dongsheng Fan
Dongsheng Fan
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Aging Neurosci. , 18 May 2022
Sec. Parkinson’s Disease and Aging-related Movement Disorders
Volume 14 - 2022 | https://doi.org/10.3389/fnagi.2022.890816
This article is part of the Research Topic Peripheral Immune System and Neurodegenerative Disease View all 10 articles
Parkinson's disease (PD) is one of the common neurodegenerative diseases that is characterized by selective degeneration of dopaminergic neurons in the substantia nigra, and misfolding of α-synuclein into aggregates is thought to contribute to its pathology. Studies have shown that immune-inflammatory responses are involved in the development of PD and play an important role in α-synuclein scavenge. Natural killer (NK) cells are first responders in immune cells and can directly promote immune defense mechanisms by cytotoxicity and by secreting cytokines. Recent discoveries suggest that NK cells are increasingly recognized in the pathological features of PD. However, the mechanisms underlying it have not been fully understood. In this review, we systematically retrieved and evaluated published evidence about the functions of NK cells in PD. We find alterations in the number of NK cells and cytotoxicity during the progression of PD, and it seems that NK cells play a neuroprotective role in PD pathogenesis, which may further reveal novel targets for the management and treatment of PD.
Parkinson's disease (PD) is one of the fastest-growing neurological diseases in terms of prevalence and mortality and is the second most common degenerative disease in the elderly with an increasing burden on healthcare systems (Poewe et al., 2017; Brakedal et al., 2021; Feigin et al., 2021). It is principally characterized by motor features, such as postural instability, bradykinesia, tremor, and rigidity. In addition, non-motor symptoms, such as hyposmia, anxiety, depression, and cognitive dysfunction, are also found to be increasingly prevalent over the course of PD development, which are gradually being paid more attention (Schapira et al., 2017). Evidence has demonstrated that progressive and selective loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc) is the main cause of motor symptoms, but the clinical diagnosis of the pathological changes that had already occurred before the manifestation of PD poses a challenge to the early diagnosis and treatment of PD (Gruden et al., 2011; Poewe et al., 2017).
At present, it is generally believed that the abnormal accumulation of α-synuclein (α-syn) is one of the important etiological factors leading to the degeneration of dopaminergic neurons, which is closely related to the formation of the Lewy body (Vekrellis et al., 2011; Poewe et al., 2017). In addition, extensive neuroinflammation characterized by the innate and adaptive immune response is found to contribute to the extensive degeneration of dopaminergic neurons (Tan et al., 2020; Harms et al., 2021). Remarkably, the neurotoxicity of α-syn, neuroinflammation, and neurodegeneration can trigger a series of pathophysiological mechanisms both locally and systemically in PD (Wong and Krainc, 2017; Tan et al., 2020; Harms et al., 2021).
Recent advances in the field of neuroinflammation demonstrate that complex interactions between CNS-resident cells and peripheral immune cells are involved in the PD pathogenesis (Iba et al., 2020; Tan et al., 2020; Harms et al., 2021). However, the mechanism involved in the recruitment of peripheral cells into the CNS and whether it is a passive migration or active participation are still unclear.
Natural killer (NK) cells are members of the innate lymphoid cells (ILCs) family, and represent 5-20% of the peripheral lymphocytes in humans (Abel et al., 2018; Zitti and Bryceson, 2018). A variety of inhibitory or activating receptors are expressed on NK cells, which regulate cell functions by binding to specific ligands and play crucial roles in cellular signal transduction. When the cells downregulate the expression of major histocompatibility complex class I (MHC-I) molecules or upregulate the expression of NK cell-activating receptor ligands abnormally, they may become the immunological attack targets for NK cells. The mechanism of cytotoxicity of NK cells can be basically classified into three types: direct release of the lytic molecules such as perforin and granzyme, activation of the extrinsic apoptotic pathway through Fas ligand (FasL) and TNF-related apoptosis-inducing ligand (TRAIL), and antibody-dependent cell-mediated cytotoxicity (ADCC) effects (Abel et al., 2018; Prager and Watzl, 2019). In addition, NK cells can produce and respond to different cytokines and play a crucial role in immunomodulation (Zitti and Bryceson, 2018).
It has been reported that NK cells are capable of interacting with α-syn in the animal models of PD and modulating neuroinflammation in neurodegenerative diseases (Poli et al., 2013; Earls et al., 2020; Garofalo et al., 2020). The existence of a link between PD and NK cells had been originally suspected due to two main reasons One is that NK cell activity was found to be related to neurological diseases, particularly demyelinating diseases in the previous studies, and researchers speculated whether similar pathological changes or immune interactions might also be present in patients with PD (Bokor et al., 1993). On the other hand, epidemiological studies provided evidence for the fact that patients with PD were more likely to have a lower incidence of cancer, probably because increased activity of natural killer cells provides defense against the infiltration of tumor cells (Mihara et al., 2008). Based on these findings, several basic and clinical studies focused on the pathological role of NK cells and performed the beneficial exploration and the attempt in this context. However, research in this field is still in its infancy, and some of the findings might be controversial. Therefore, we conducted a systematic review of the relevant literature to investigate the alterations of NK cells in PD and discuss how NK cells can influence the disease onset and progression.
This systematic review was reported according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines.
The search of PubMed, Embase, and Web of Science databases (from inception up to January 2022) was performed by two authors (ZL and ZYS) independently. The search strategy involved selecting the most relevant Medical Subject Headings (MeSH) and title/abstract text keywords used in combination or alone: “Parkinson's disease,” “NK cell,” “natural killer cell,” “alpha-synucleinopathy,” and “dopaminergic nerve cell” (detailed search strategy is described in Supplementary Table 1).
To prove the eligibility of the studies, the following inclusion and exclusion criteria were formulated:
Inclusion criteria were as follows: (1) articles published in English, (2) original articles regarding animals and humans (peripheral lymphocyte analysis) and in vitro studies (cell cultivation) of natural killer cells where relevant, and (3) the research theme focused on “Parkinson's disease.” Exclusion criteria were as follows: (1) case report, case series, letter, poster, commentary, proceedings, laboratory science studies, review, and systematic review, (2) repeatedly published literature, unpublished data, or findings reported in abstract form only, and (3) literature of poor quality.
Data extraction from eligible articles was performed by two investigators (ZL and ZYS) independently. The following clinical information was extracted from the study: publication, region of the population, animal models of PD, number of patients and healthy controls, gender statistics, age at diagnosis, disease duration, disease severity, and the main results.
Quality assessment was carried out by two authors (ZL and ZYS) independently. Modified Collaborative Approach to Meta-Analysis and Review of Animal Data from Experimental Studies (CAMARADES) and Newcastle-Ottawa Scale (NOS) were used as quality assessment tools for the animal experimental studies and clinical studies separately. Ten items are included in the modified CAMARADES tool: sample size calculation, random allocation to treatment or control, allocation concealment, blinded assessment of outcome, appropriate animal model, use of anesthetic without significant intrinsic neuroprotective activity, statement of control of temperature, compliance with animal welfare regulations, peer-reviewed publication, and statement of potential conflict of interests. Seven items are included in the NOS tool: adequate case definition, representativeness of the cases, selection of controls, definition of controls, comparability of cases and controls on the basis of the design or analysis (study controls for age or any additional factors), ascertainment of exposure, and same method of ascertainment for cases and controls and non-response rate. The total scores of the two scales are 10 and 9, respectively. The final scores of each study are described in Supplementary Tables 2, 3. Any discrepancies were resolved through discussion. If an agreement could not be reached, the disagreements were resolved with the help of a third author.
A total of 408 records were generated from the database search after removing the duplicates. After screening for title and abstract, 388 records were excluded. Full-text of the remaining 20 articles was assessed, and finally, a total of 12 studies were eligible for the systematic review after full-text screening (Figure 1).
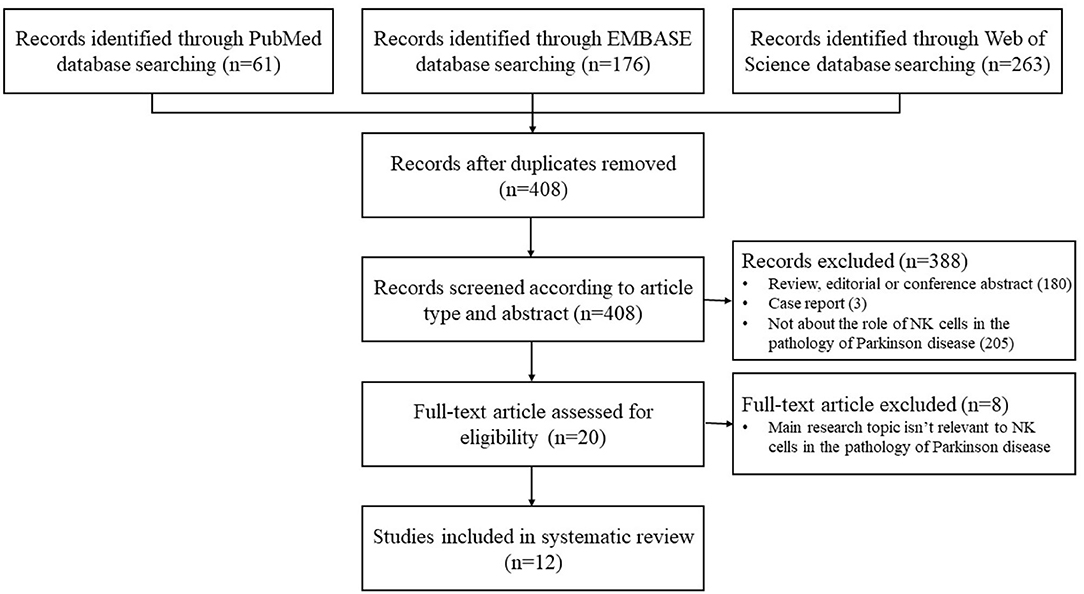
Figure 1. Flowchart of the study selection.
We identified three studies on animals according to the inclusion criteria, and the main findings are summarized in Table 1. Two studies used a preformed fibril (PFF) α-syn injection model in C57BL/6J mice and transgenic mice that overexpressed human A53T α-syn mutant protein separately, and one study used a 6-hydroxydopamine (6-OHDA) injection model in Wistar rats. These two models are commonly used in the study of PD. For the PFF α-syn model, PFF α-syn acts as a template and induces the endogenous α-syn to accumulate into misfolded phosphorylated pathological aggregates in the substantia nigra (Earls et al., 2020). In the 6-OHDA model, 6-OHDA (a neurotoxin) is absorbed by dopaminergic neurons rich in monoamine oxidase in the substantia after injection, which can further convert into free radicals and cause damage to neurons; 6-OHDA does not induce the formation of α-syn aggregates or Lewy-like inclusions in PD (Grembecka et al., 2021).
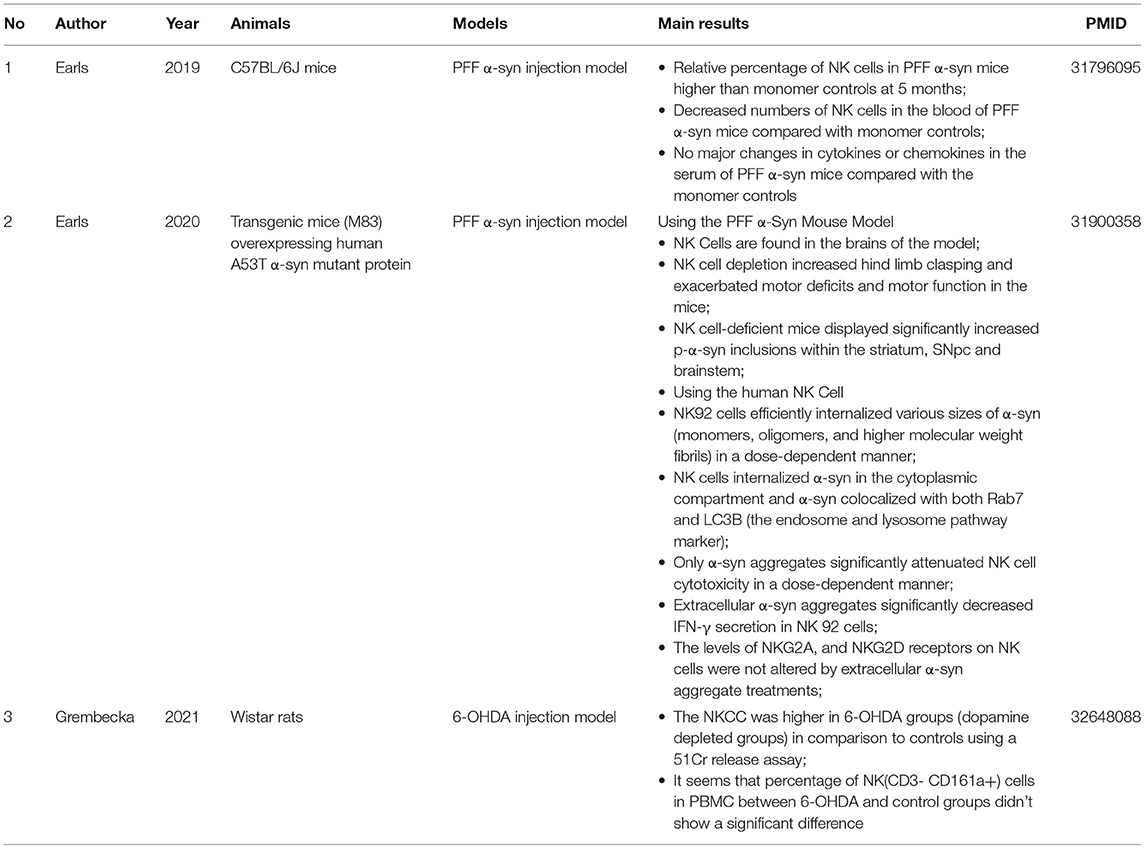
Table 1. Main findings of animal studies.
The first study (Earls et al., 2019) showed that intrastriatal injection of PFF α-syn could induce Lewy body–like pathology, and the percentage of NK cells in the mononuclear immune cells at 5 months post injection (p.i.) was relatively increased in the CNS parenchyma, which was confirmed by the flow cytometry analysis. In addition, characterization of the leukocytes in the peripheral blood showed that the number of NK cells decreased, but the percentage of the NK cell population did not show significant changes.
In the second study, Earls et al. confirmed the presence of NK cells in the SNs of both postmortem PD patients and M83 Tg mice based on immunohistochemistry (IHC) markers (Earls et al., 2020). More importantly, obvious clinical motor deficits and an increased number of p-α-syn inclusions within the striatum, substantia nigra pars compacta (SNpc), and brainstem were observed in PFF α-syn–injected M83 Tg mice after systemic NK cell depletion, which indicated the protective role of NK cells in PD. Moreover, researchers carried out cell-based assays and found the bidirectional effects between extracellular α-syn and NK cells: NK92 cells and primary human NK cells could internalize the extracellular α-syn and scavenge it through the endosome and lysosome pathway, while extracellular α-syn aggregates attenuated NK cell cytotoxicity (NKCC) and IFN-γ secretion without causing any significant changes in NKG2A and NKG2D receptors on the cell surface. It is an interesting phenomenon that NK cells are not aberrantly activated.
For the third study (Grembecka et al., 2021), we extracted information regarding the comparison between the PD model and the blank controls. It seems that the number of NK (CD3-CD161a+) cells in PBMC did not show any significant difference between the 6-OHDA groups and the controls (without p-values recorded). Furthermore, they observed elevated peripheral NKCC levels after 6-OHDA microinjection into the SNpc, which means NK cells might be actively involved in the progression of PD.
Nine clinical studies explored the relationship between NK cells and sporadic Parkinson's disease according to the inclusion criteria, and the basic information and important findings of these studies are summarized in Tables 2, 3. Most of the included studies provided basic information regarding the subject number, gender, age at visit/onset of patients, disease duration, and H&Y/UPDRS-III (part III of the Unified Parkinson's Disease Rating Scale) assessment. NK cells are mainly characterized based on the cell number, cell cytotoxicity, and changes in the cell surface receptors.
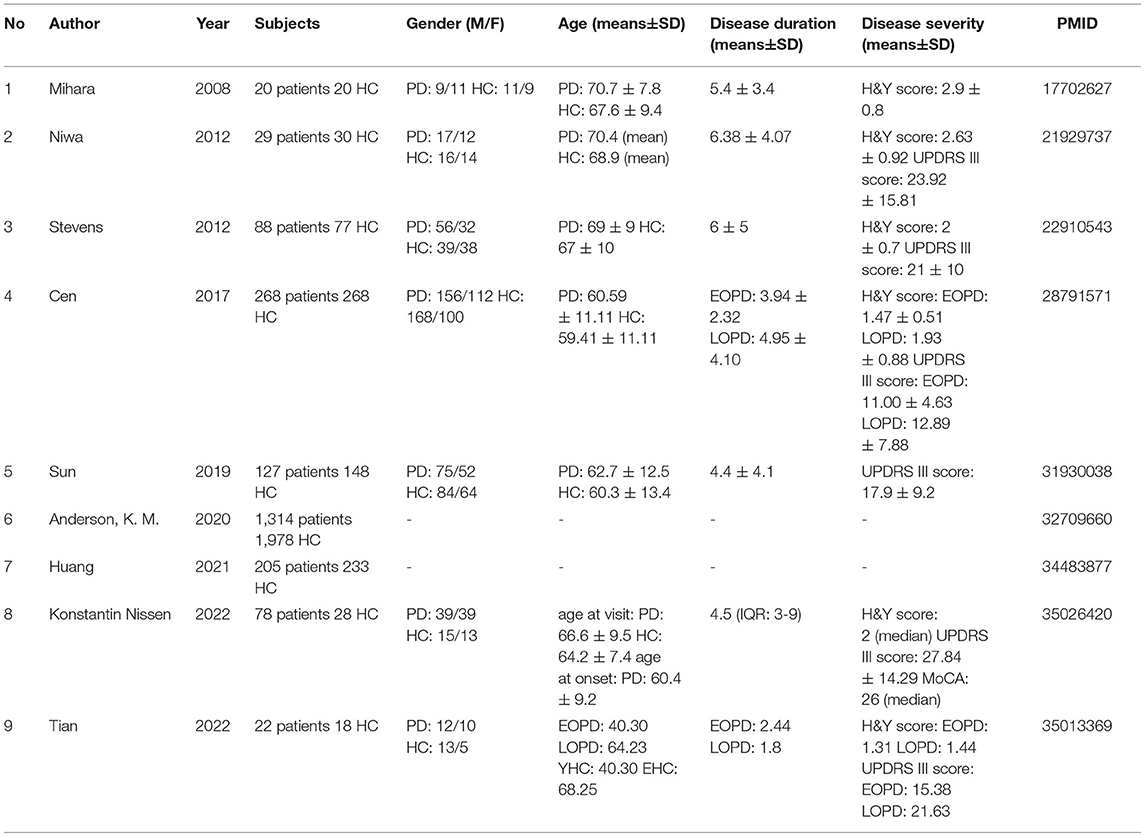
Table 2. Basic information of clinical studies.
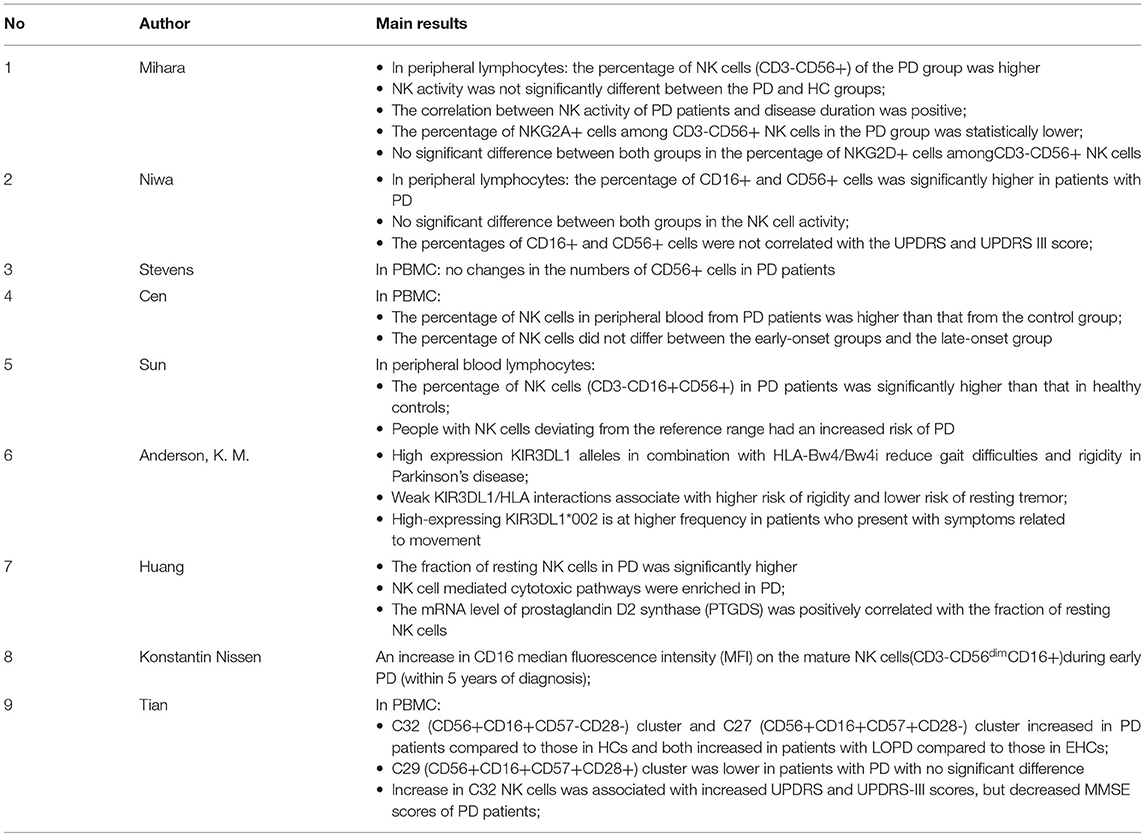
Table 3. Main findings of clinical studies.
Seven studies analyzed NK cell count in the peripheral blood. One of them found no differences between the patients and controls (Stevens et al., 2012), five of them reported a higher percentage of NK cells in the patients with PD (Mihara et al., 2008; Niwa et al., 2012; Cen et al., 2017; Sun et al., 2019; Huang et al., 2021), and in one study, Tian et al. divided NK cells into three clusters and reported that only C32 (CD56+, CD16+, CD57–, and CD28–) and C27 (CD56+, CD16+, CD57+, and CD28–) clusters increased (Tian et al., 2022). Furthermore, we noticed that the markers used to identify NK cells from the PBMC were quite different. CD16, a typical NK cell marker and also known as FCγRIII, is an activation receptor that mediates ADCC effects. CD56, known as nerve cell-associated adhesion molecule (NCAM), is considered as a functional indicator of human NK cells and separates the cells into precursor and mature populations based on its expression. In some studies, cells labeled by CD16 or CD56 were used as markers, while in others, those cells that expressed both CD16 and CD56 were recognized as NK cells. Mihara et al. and Niwa et al. reported the occurrence of the inhibitory NKG2A cells and the activating NKG2D cells, which indicated that the percentage of NKG2A+ cells among CD3-CD56+ NK cells was lower and the percentage of NKG2D+ cells in the peripheral lymphocytes was higher in the PD group (p < 0.05) (Mihara et al., 2008; Niwa et al., 2012).
Experiments and bioinformatics analysis were used to analyze the cytotoxicity in NK cells. Only two studies measured NKCC activity against the K562 target cells through the LDH cytotoxic assay and the calcein acetoxymethyl ester release assay, and both of them found no significant differences between the patients with PD and the controls (Mihara et al., 2008; Niwa et al., 2012). Additionally, the former study provided information that NKCC increased as the disease progressed in PD (Mihara et al., 2008). On the other hand, from the perspective of bioinformatics analysis, Huang et al. carried out the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis and revealed that NK cell-mediated cytotoxic pathways were enriched in PD based on the blood expression profile (Huang et al., 2021).
Since aging is a common predisposing factor for Parkinson's disease, stratification by age at disease onset needs to be considered (Pagano et al., 2016). Two studies analyzed the subset of NK cells in the early-onset Parkinson's disease (EOPD) and late-onset Parkinson's disease (LOPD). Cen et al. observed that the percentage of NK cells in peripheral blood did not differ significantly between the two groups (p = 0.067) (Cen et al., 2017). Tian et al. showed a more detailed comparison between early- or late-onset patients and the age-matched controls. For the less toxic NK cells, CD32 (CD56+, CD16+, CD57–, and CD28–) clusters were significantly higher in the EOPD/LOPD group, while CD27 (CD56+, CD16+, CD57+, and CD28–) clusters were significantly higher only in the LOPD group. Cytotoxic CD29 (CD56+, CD16+, CD57+, and CD28+) clusters were lower in the LOPD group when compared to the control group, with no statistical difference between the groups. Furthermore, it was found that C27 cell clusters increased as the disease was prolonged in LOPD (Tian et al., 2022).
In addition, the disease severity may also be associated with the NK cells. Two studies explored the relationship between NK cells and disease progression measured by UDPRS and Mini-Mental State Examination (MMSE) scales. According to Niwa et al., the percentage of CD16+ cells and CD56+ cells was not correlated with the UPDRS (p > 0.1) and UPDRS-III scores (p > 0.1) in the 29 patients diagnosed with sporadic PD (Niwa et al., 2012). However, an increase in C32 clusters was found to correspond to increased UPDRS and UPDRS-III scores but decreased MMSE scores in the patients with PD (Tian et al., 2022).
In addition, other markers of NK cells were found to have an impact on the clinical manifestations. Using the bioinformatics tools, Anderson et al. analyzed the interaction between polymorphic killer immunoglobulin-like receptors (KIR) and human leukocyte antigen (HLA) class I ligands in patients with PD (Anderson et al., 2020). KIR3DL1, a member of the KIR family, conducts inhibitory signals depending on two immunoreceptor tyrosine-based inhibition motifs (ITIM) and binds to HLA-B molecules containing the Bw4 phenotype. The researchers found that high expression of KIR3DL1 alleles in combination with HLA-Bw4/Bw4i reduced gait difficulties and rigidity in patients, while weak KIR3DL1/HLA interactions were found to be associated with a higher risk of rigidity. Thus, they speculated that high expression levels of inhibitory KIR to reduce NK cell-mediated inflammation may reduce the severity of symptoms and have a protective effect in PD.
This is the first systematic review to evaluate the influence of NK cells in the development of Parkinson's disease based on the data from both foundation medicine and clinical medicine, and several issues should be considered critically and seriously.
Evidence showed that NK cells were found in the brains of patients with PD (Earls et al., 2020), and the percentage of NK cells increased in the CNS parenchyma prior to the dopaminergic neuronal degeneration (Earls et al., 2019), which indicated infiltration of NK cells in the CNS during PD progression. From the point of ethics and experimental convenience, NK cells in the peripheral blood were appropriate and easy to obtain and could also give us information about pathogenesis. However, analysis of the cell counts and cell cytotoxicity had produced some contradictory results in both clinical studies and animal studies.
Most groups reported increased NK cells (Mihara et al., 2008; Niwa et al., 2012; Cen et al., 2017; Sun et al., 2019; Huang et al., 2021; Tian et al., 2022) in patients, while some researchers showed that the number of NK cells did not change significantly when compared to the corresponding controls (Stevens et al., 2012; Earls et al., 2019; Grembecka et al., 2021). We speculated that cohort/model differences, PD subtype, disease duration, or time delay between the clinical measurements of the disease and the pathological changes are the possible influencing factors, and the mechanisms underlying altered immune profiles still need to be explored.
Moreover, peripheral blood mononuclear cells (PBMCs) were shown as a basic reference in most studies. PBMCs are cell groups that include blood cells with a round nucleus (i.e., lymphocytes, monocytes, NK cells, or dendritic cells) (Sen et al., 2017). Among the PBMCs, components other than NK cells may also be altered in the pathological process of PD. For example, changes in monocyte subpopulations were observed in the early and late PD (Harms et al., 2021; Tian et al., 2022). Therefore, it might be better to measure the absolute value of the NK cells for comparison rather than considering only the percentage of cells.
As for the NKCC, Huang et al. confirmed enriched NK cell-mediated cytotoxic pathways in the peripheral blood using Gene Set Enrichment Analysis (GSEA), supporting a disease-associated cytotoxic response of NK cells in PD (Huang et al., 2021). Also, NK cells may be actively involved in the acute injury of dopaminergic neurons, since increased peripheral NKCC was found in rats injected with 6-OHDA for several weeks (Grembecka et al., 2021). However, some studies revealed no significant changes in NKCC targeting the K562 cells lacking ligands of the inhibitory NKG2A receptors (Mihara et al., 2008; Niwa et al., 2012). Given the fact that alterations in the expression levels of NKG2A receptors on NK cells were inconsistent between different studies (Mihara et al., 2008; Niwa et al., 2012), a possible approach involving the measurement of NKCC activity would be an influencing factor in the final results.
Notably, researchers for the first time observed that extracellular α-syn aggregates are internalized and degraded in the NK cells, and a more intriguing finding is that NKCC decreased obviously in a dose-dependent manner after the interaction between α-syn aggregates and NK cells (Earls et al., 2020). It seemed that the internalization of α-syn modulated the cytotoxicity of NK cells in a non-activated way. Recent studies have demonstrated that microglia can ingest and degrade extracellular α-syn with high efficiency, but consequently, α-syn fibrils are likely to induce apoptosis of microglia by increased reactive oxygen species (ROS) production and mitochondrial network disintegration (Choi et al., 2020; Scheiblich et al., 2021). In this case, whether α-syn exhibits toxicity in the NK cells and how α-syn aggregates affect the NKCC after the phagocytosis are still unclear.
In addition, researchers also observed increased CD16 expression on mature NK cells and speculated that an enhanced ADCC effect was primed in patients with PD (Konstantin Nissen et al., 2022). Since information about antibodies and target cells is unknown, cytotoxicity through ADCC is not measured in an intuitive way, and hence more number of experiments are needed to prove this hypothesis. Therefore, some doubts remain about the measurement of NKCC. To further elucidate these findings, mechanisms and further validations are encouraged to identify the cytotoxicity of NK cells more accurately and comprehensively in the dynamic development of PD in patients.
Previous studies demonstrated that older age at onset was associated with a more severe motor phenotype and higher H&Y stage and UPDRS-III score, and similar results were observed by Cen et al. and Tian et el. (Szewczyk-Krolikowski et al., 2014; Pagano et al., 2016). Possibly, it can be explained by a greater dopaminergic dysfunction in patients with older age at onset, but the mechanisms are still unclear. Evidence suggests that genetic variation plays an important role in determining the age of onset of PD (Nalls et al., 2015). GWAS has implicated that SNCA, TMEM175, and GBA are associated with PD age at onset and are also implicated in α-syn aggregation pathways (Blauwendraat et al., 2019), which means that accumulation and clearance of α-syn are likely to be involved in the pathogenesis of age at onset. Besides, differences in the immune status exist in the patients with early- and late-onset PD (Tian et al., 2022). As for NK cells, functions alter with the differential expression of molecules on the cell surface in the relevant development stages and can be affected by various immune responses (Yu et al., 2013; Abel et al., 2018). It is possible that NK cells are related to age at onset in patients with PD.
Tian et al. analyzed peripheral mature NK cells marked by CD16 and CD56, and further distinguished cell maturity by expression of CD57 (a marker of terminal maturation) and CD28 (critical for co-stimulation of the cells to proliferate and produce IFN-γ) (Tian et al., 2022). Cytotoxicity of cells marked by CD57+CD28+ (CD29 clusters) is higher than those marked by CD57+CD28- (CD27 clusters) and CD57-CD28- (CD32 clusters). Interestingly, the researchers found higher CD27 and CD32 cell clusters and lower CD29 clusters in LOPD, while only higher CD32 clusters were noticed in EOPD, which suggested that age at onset might be associated with the development and toxicity alterations of NK cells in PD. Furthermore, considering that aging is one of the main risk factors for PD (Pagano et al., 2016; Poewe et al., 2017), it is crucial to control the variables such as age-matched healthy controls in the study design involving comparison between EOPD and LOPD.
NK cells are essential effectors in innate immunity, since NK cells can infiltrate the CNS during PD progression (Earls et al., 2020). However, whether there are pathophysiological reflections on the clinical motor and non-motor symptoms is an important issue for us to understand the role of NK cells in PD. Tian et al. observed less toxic NK cells increased with more severe motor and cognitive dysfunctions based on the evaluation of UPDRS-III and MMSE scales (Tian et al., 2022). Anderson et al. found that high expression of inhibitory KIR on NK cells might protect against severe motor symptoms in patients (Anderson et al., 2020). Additionally, in the PFF α-syn model, increased motor symptoms, exacerbated motor deficits, and increased p-α-syn inclusions were observed due to NK cell depletion (Earls et al., 2020). Therefore, we speculated that NK cells carry out killing and phagocytosis in a less toxic or non-hyperactivated way, which tend to play a protective role in the progression of PD.
However, previous studies have demonstrated that NK cells likely act as a double-edged sword in central nervous system disorders (Poli et al., 2013, 2018). On the one hand, they can induce the activation of neural cell death (Garofalo et al., 2020), while on the other hand, they have the ability to suppress CNS inflammation (Hao et al., 2010). In addition, immune dysfunction is quite complex in the development of PD. Pro-inflammatory or anti-inflammatory cytokines and cells, such as microglia, monocytes, T cells, B cells, and so on, are undoubtedly important participants in the pathological process and correlate with disease severity and disability in patients with PD (reviewed in detail in: Qin et al., 2016; Chen et al., 2018; Sabatino et al., 2019; Harms et al., 2021). However, only a few studies have explored their interactions with NK cells in PD till now, so the detrimental effect of NK cells is still uncertain and needs further investigation.
Drugs are the influential factors in the study design. Basu et al. reviewed that dopamine and its receptor agonists altered the function of NK cells, for example, administration of dopamine increased the killing ability of specific NK cells (Basu and Dasgupta, 2000). Thus, data collection may be better in newly diagnosed PD patients to reduce the effects of dopamine replacement therapies. Additionally, neurotransmitters and neuroendocrine factors, such as glucocorticoids, serotonin, and epinephrine, have been reported to affect the activities of NK cells through a complex communication between the immune system and the nervous system (Capellino et al., 2020).
While we included the available studies on the relevant topic and systematically summarized the main results, there are some limitations in the article. First, heterogeneity of the inclusion criteria and experimental methods hindered a meta-analysis of the data, so we chose to report a descriptive analysis of our findings, rather than the quantitative conclusions. Second, a small sample size was found in most of the included studies, which reduced the reliability of the corresponding results. Third, the quality of some included studies was affected by several factors, such as lack of information on the basic characteristics of the patients, the absolute count of the measurement, and so on. Furthermore, publication bias toward studies with positive clinical outcomes cannot be ruled out.
In the future, we hope that more promising translational therapeutic targets for PD will emerge with the conducting of extensive research on the essential pathological and physiological changes. It is known that NK cells can launch a rapid attack the next time they encounter the same antigen through antigen-mediated immune memory, and they can then present strong killing functions when perceiving a similar inflammatory environment. In addition, studies have shown that CNS-derived antigens can trigger neuroinflammation responses in PD. Therefore, there is a possibility that memory NK cells exist in the PD progression, and it is quite interesting to be explored in the future. In addition, NK cells have been reported to be associated with cognitive decline due to some complex mechanisms. Since non-motor symptoms, such as cognitive deficits, also have a certain impact on the life quality of patients with PD, looking for connections between NK cells and the non-motor symptoms is worthwhile in the basic experiments and clinical practice.
Overall, we reviewed the pathologic role of NK cells in Parkinson's disease systematically and summarized the characteristics of NK cell numbers, NKCC, and other surface molecules on the NK cells in patients with PD and animal models. Further, we analyzed the relationship between NK cells and the clinical manifestations in PD and speculated a probable neuroprotective role of NK cells. We hope that we have provided the necessary impetus for continued studies on the functions of NK cells in PD, and more potential biomarkers or targets will spring up during the diagnosis and treatment of PD in the future.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
LZ, YZ, and DF: conceptualization. LZ and YZ: methodology and data curation. LZ: writing—original draft preparation. YZ and DF: writing—review and editing. All authors have read and agreed to the published version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnagi.2022.890816/full#supplementary-material
Abel, A. M., Yang, C., Thakar, M. S., and Malarkannan, S. (2018). Natural killer cells: development, maturation, and clinical utilization. Front. Immunol. 9, 1869. doi: 10.3389/fimmu.2018.01869
Anderson, K. M., Augusto, D. G., Dandekar, R., Shams, H., Zhao, C., Yusufali, T., et al. (2020). Killer cell immunoglobulin-like receptor variants are associated with protection from symptoms associated with more severe course in parkinson disease. J. Immunol. 205, 1323–1330. doi: 10.4049/jimmunol.2000144
Basu, S., and Dasgupta, P. S. (2000). Dopamine, a neurotransmitter, influences the immune system. J. Neuroimmunol. 102, 113–124. doi: 10.1016/S0165-5728(99)00176-9
Blauwendraat, C., Heilbron, K., Vallerga, C. L., Bandres-Ciga, S., Von Coelln, R., Pihlstrøm, L., et al. (2019). Parkinson's disease age at onset genome-wide association study: Defining heritability, genetic loci, and α-synuclein mechanisms. Mov. Disord. 34, 866–875. doi: 10.1002/mds.27659
Bokor, M., Farag,ó, A., Garam, T., Malatinszky, G., and Schnabel, R. (1993). Antibody-dependent cell-mediated cytotoxicity (ADCC) in Parkinson's disease. J. Neurol. Sci. 115, 47–50. doi: 10.1016/0022-510X(93)90065-7
Brakedal, B., Tzoulis, C., and Haugarvoll, K. (2021). A nationwide study of the incidence, prevalence and mortality of Parkinson's disease in the Norwegian population. Eur. J. Neurol. 28, 81–81. doi: 10.1038/s41531-022-00280-4
Capellino, S., Claus, M., and Watzl, C. (2020). Regulation of natural killer cell activity by glucocorticoids, serotonin, dopamine, and epinephrine. Cell Mol. Immunol. 17, 705–711. doi: 10.1038/s41423-020-0477-9
Cen, L., Yang, C., Huang, S., Zhou, M., Tang, X., Li, K., et al. (2017). Peripheral lymphocyte subsets as a marker of Parkinson's disease in a Chinese population. Neurosci. Bull. 33, 493–500. doi: 10.1007/s12264-017-0163-9
Chen, Z., Chen, S., and Liu, J. (2018). The role of T cells in the pathogenesis of Parkinson's disease. Prog. Neurobiol. 169, 1–23. doi: 10.1016/j.pneurobio.2018.08.002
Choi, I., Zhang, Y., Seegobin, S. P., Pruvost, M., Wang, Q., Purtell, K., et al. (2020). Microglia clear neuron-released α-synuclein via selective autophagy and prevent neurodegeneration. Nat. Commun. 11, 1386. doi: 10.1038/s41467-020-15119-w
Earls, R. H., Menees, K. B., Chung, J., Barber, J., Gutekunst, C. A., Hazim, M. G., et al. (2019). Intrastriatal injection of preformed alpha-synuclein fibrils alters central and peripheral immune cell profiles in non-transgenic mice. J. Neuroinflammation 16, 250. doi: 10.1186/s12974-019-1636-8
Earls, R. H., Menees, K. B., Chung, J., Gutekunst, C. A., Lee, H. J., Hazim, M. G., et al. (2020). NK cells clear alpha-synuclein and the depletion of NK cells exacerbates synuclein pathology in a mouse model of alpha-synucleinopathy. Proc. Natl. Acad. Sci. U. S. A. 117, 1762–1771. doi: 10.1073/pnas.1909110117
Feigin, V. L., Vos, T., Alahdab, F., Amit, A. M. L., Bärnighausen, T. W., Beghi, E., et al. (2021). Burden of neurological disorders across the US From 1990-2017: a global burden of disease study. JAMA Neurol. 78, 165–176. doi: 10.1001/jamaneurol.2020.4152
Garofalo, S., Cocozza, G., Porzia, A., Inghilleri, M., Raspa, M., Scavizzi, F., et al. (2020). Natural killer cells modulate motor neuron-immune cell cross talk in models of amyotrophic lateral sclerosis. Nat. Commun. 11, 1773. doi: 10.1038/s41467-020-15644-8
Grembecka, B., Glac, W., Listowska, M., Jerzemowska, G. O. D., Plucinska, K., Majkutewicz, I., et al. (2021). Subthalamic deep brain stimulation affects plasma corticosterone concentration and peripheral immunity changes in rat model of Parkinson's disease. J. Neuroimmune Pharmacol. 16, 454–469. doi: 10.1007/s11481-020-09934-7
Gruden, M. A., Sewell, R. D. E., Yanamandra, K., Davidova, T. V., Kucheryanu, V. G., Bocharov, E. V., et al. (2011). Immunoprotection against toxic biomarkers is retained during Parkinson's disease progression. J. Neuroimmunol. 233, 221–227. doi: 10.1016/j.jneuroim.2010.12.001
Hao, J., Liu, R., Piao, W., Zhou, Q., Vollmer, T. L., Campagnolo, D. I., et al. (2010). Central nervous system (CNS)-resident natural killer cells suppress Th17 responses and CNS autoimmune pathology. J. Exp. Med. 207, 1907–1921. doi: 10.1084/jem.20092749
Harms, A. S., Ferreira, S. A., and Romero-Ramos, M. (2021). Periphery and brain, innate and adaptive immunity in Parkinson's disease. Acta Neuropathol. 141, 527–545. doi: 10.1007/s00401-021-02268-5
Huang, Y., Liu, H., Hu, J., Han, C., Zhong, Z., Luo, W., et al. (2021). Significant difference of immune cell fractions and their correlations with differential expression genes in Parkinson's disease. Front. Aging Neurosci. 13:686066. doi: 10.3389/fnagi.2021.686066
Iba, M., Kim, C., Sallin, M., Kwon, S., Verma, A., Overk, C., et al. (2020). Neuroinflammation is associated with infiltration of T cells in Lewy body disease and α-synuclein transgenic models. J. Neuroinflammation 17, 214. doi: 10.1186/s12974-020-01888-0
Konstantin Nissen, S., Farmen, K., Carstensen, M., Schulte, C., Goldeck, D., Brockmann, K., et al. (2022). Changes in CD163+, CD11b+, and CCR2+ peripheral monocytes relate to Parkinson's disease and cognition. Brain Behav. Immun. 101, 182–193. doi: 10.1016/j.bbi.2022.01.005
Mihara, T., Nakashima, M., Kurolwa, A., Akitake, Y., Ono, K., Hosokawa, M., et al. (2008). Natural killer cells of Parkinson's disease patients are set up for activation: A possible role for innate immunity in the pathogenesis of this disease. Parkinsonism Relat. Disord. 14, 46–51. doi: 10.1016/j.parkreldis.2007.05.013
Nalls, M. A., Escott-Price, V., Williams, N. M., Lubbe, S., Keller, M. F., Morris, H. R., et al. (2015). Genetic risk and age in Parkinson's disease: continuum not stratum. Mov. Disord. 30, 850–854. doi: 10.1002/mds.26192
Niwa, F., Kuriyama, N., Nakagawa, M., and Imanishi, J. (2012). Effects of peripheral lymphocyte subpopulations and the clinical correlation with Parkinson's disease. Geriatr. Gerontol. Int. 12, 102–107. doi: 10.1111/j.1447-0594.2011.00740.x
Pagano, G., Ferrara, N., Brooks, D. J., and Pavese, N. (2016). Age at onset and Parkinson disease phenotype. Neurology 86, 1400–1407. doi: 10.1212/WNL.0000000000002461
Poewe, W., Seppi, K., Tanner, C. M., Halliday, G. M., Brundin, P., Volkmann, J., et al. (2017). Parkinson disease. Nat. Rev. Dis. Primers 3, 17013. doi: 10.1038/nrdp.2017.13
Poli, A., Kmiecik, J., Domingues, O., Hentges, F., Blery, M., Chekenya, M., et al. (2013). NK cells in central nervous system disorders. J. Immunol. 190, 5355–5362. doi: 10.4049/jimmunol.1203401
Poli, A., Michel, T., Patil, N., and Zimmer, J. (2018). Revisiting the Functional Impact of NK Cells. Trends Immunol. 39, 460–472. doi: 10.1016/j.it.2018.01.011
Prager, I., and Watzl, C. (2019). Mechanisms of natural killer cell-mediated cellular cytotoxicity. J. Leukoc. Biol. 105, 1319–1329. doi: 10.1002/JLB.MR0718-269R
Qin, X. Y., Zhang, S. P., Cao, C., Loh, Y. P., and Cheng, Y. (2016). Aberrations in peripheral inflammatory cytokine levels in Parkinson disease: a systematic review and meta-analysis. JAMA Neurol. 73, 1316–1324. doi: 10.1001/jamaneurol.2016.2742
Sabatino, J. J. Jr, Probstel, A. K., and Zamvil, S. S. (2019). B cells in autoimmune and neurodegenerative central nervous system diseases. Nat. Rev. Neurosci. 20, 728–745. doi: 10.1038/s41583-019-0233-2
Schapira, A. H. V., Chaudhuri, K. R., and Jenner, P. (2017). Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 18, 509. doi: 10.1038/nrn.2017.62
Scheiblich, H., Dansokho, C., Mercan, D., Schmidt, S. V., Bousset, L., Wischhof, L., et al. (2021). Microglia jointly degrade fibrillar alpha-synuclein cargo by distribution through tunneling nanotubes. Cell 184, 5089–5106.e21. doi: 10.1016/j.cell.2021.09.007
Sen, P., Kemppainen, E., and Oreši,č, M. (2017). Perspectives on systems modeling of human peripheral blood mononuclear cells. Front. Mol. Biosci. 4, 96. doi: 10.3389/fmolb.2017.00096
Stevens, C. H., Rowe, D., Morel-Kopp, M. C., Orr, C., Russell, T., Ranola, M., et al. (2012). Reduced T helper and B lymphocytes in Parkinson's disease. J. Neuroimmunol. 252, 95–99. doi: 10.1016/j.jneuroim.2012.07.015
Sun, C., Zhao, Z., Yu, W., Mo, M., Song, C., Si, Y., et al. (2019). Abnormal subpopulations of peripheral blood lymphocytes are involved in Parkinson's disease. Ann. Transl. Med. 7, 637. doi: 10.21037/atm.2019.10.105
Szewczyk-Krolikowski, K., Tomlinson, P., Nithi, K., Wade-Martins, R., Talbot, K., Ben-Shlomo, Y., et al. (2014). The influence of age and gender on motor and non-motor features of early Parkinson's disease: initial findings from the Oxford Parkinson Disease Center (OPDC) discovery cohort. Parkinsonism Relat. Disord. 20, 99–105. doi: 10.1016/j.parkreldis.2013.09.025
Tan, E. K., Chao, Y. X., West, A., Chan, L. L., Poewe, W., and Jankovic, J. (2020). Parkinson disease and the immune system - associations, mechanisms and therapeutics. Nat. Rev. Neurol. 16, 303–318. doi: 10.1038/s41582-020-0344-4
Tian, J., Dai, S. B., Jiang, S. S., Yang, W. Y., Yan, Y. Q., Lin, Z. H., et al. (2022). Specific immune status in Parkinson's disease at different ages of onset. NPJ Parkinsons Dis. 8, 5. doi: 10.1038/s41531-021-00271-x
Vekrellis, K., Xilouri, M., Emmanouilidou, E., Rideout, H. J., and Stefanis, L. (2011). Pathological roles of alpha-synuclein in neurological disorders. Lancet Neurol. 10, 1015–1025. doi: 10.1016/S1474-4422(11)70213-7
Wong, Y. C., and Krainc, D. (2017). alpha-synuclein toxicity in neurodegeneration: mechanism and therapeutic strategies. Nat. Med. 23, 1–13. doi: 10.1038/nm.4269
Yu, J., Freud, A. G., and Caligiuri, M. A. (2013). Location and cellular stages of natural killer cell development. Trends Immunol. 34, 573–582. doi: 10.1016/j.it.2013.07.005
Keywords: Parkinson’s disease, α-synuclein, natural killer cells, natural killer cell cytotoxicity, neuroinflammation
Citation: Zhang L, Zhang Y and Fan D (2022) Pathological Role of Natural Killer Cells in Parkinson's Disease: A Systematic Review. Front. Aging Neurosci. 14:890816. doi: 10.3389/fnagi.2022.890816
Received: 06 March 2022; Accepted: 14 April 2022;
Published: 18 May 2022.
Edited by:
Chao Wang, Chongqing Medical University, ChinaReviewed by:
Jiali Pu, Zhejiang University, ChinaCopyright © 2022 Zhang, Zhang and Fan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yingshuang Zhang, emhhbmd5czAzMTdAMTI2LmNvbQ==; Dongsheng Fan, ZHNmYW4yMDEwQGFsaXl1bi5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.