 Jin Peng
Jin Peng Jinhui Wu
Jinhui Wu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Aging Neurosci., 31 March 2022
Sec. Alzheimer's Disease and Related Dementias
Volume 14 - 2022 | https://doi.org/10.3389/fnagi.2022.863901
This article is part of the Research TopicPeripheral Immune System and Neurodegenerative DiseaseView all 10 articles
Population aging is an inevitable problem nowadays, and the elderly are going through a lot of geriatric symptoms, especially cognitive impairment. Irisin, an exercise-stimulating cleaved product from transmembrane fibronectin type III domain-containing protein 5 (FNDC5), has been linked with favorable effects on many metabolic diseases. Recently, mounting studies also highlighted the neuroprotective effects of irisin on dementia. The current evidence remains uncertain, and few clinical trials have been undertaken to limit its clinical practice. Therefore, we provided an overview of current scientific knowledge focusing on the preventive mechanisms of irisin on senile cognitive decline and dementia, in terms of the possible connections between irisin and neurogenesis, neuroinflammation, oxidative stress, and dementia-related diseases. This study summarized the recent advances and ongoing studies, aiming to provide a better scope into the effectiveness of irisin on dementia progression, as well as a mediator of muscle brain cross talk to provide theoretical support for exercise therapy for patients with dementia. Whether irisin is a diagnostic or prognostic factor for dementia needs more researches.
The world has entered an aging society. In addition to chronic diseases, the elderly is accompanied by a series of geriatric symptoms. Cognitive impairment is a classic symptom of geriatric syndrome, which occurs from mild cognitive impairment (MCI) to dementia (Sanford, 2017). MCI is an intermediate state between neurotypical cognition and neurodegenerative dementia (Petersen et al., 2018). The prevalence of MCI in the elderly population aged ≥60 years is approximately 6.4–25% and increases with age according to the American Academy of Neurology (AAN) guidelines (Cheng et al., 2017). Dementia is the most serious form of cognitive impairment; diminishes the physical and mental function of older people, quality of life, and disability; and is the fifth leading cause of death (Winblad et al., 2004).
There were some risk factors of MCI and dementia, such as cardiovascular diseases (Schumacher et al., 2013), inflammation (Huh et al., 2014), and stroke (Chen et al., 2019). Alzheimer’s disease (AD) is a classic type of dementia, which is characterized at the neuropathological level by deposits of insoluble amyloid β-peptide (Aβ) in extracellular plaques and aggregated Tau proteins (Hodson, 2018). Developing evidence suggested that decreased brain-derived neurotrophic factor (BDNF) (Amidfar et al., 2020) and damaged synaptic plasticity (Skaper et al., 2017) led to dementia. However, the mechanism remains to be clarified.
Irisin, a myokine containing 112 amino acids, is secreted by skeletal muscle after exercise stimulation, which was first found in 2012 by Boström et al. (2012). It is processed from the type I membrane protein encoded by the FNDC5 gene, then secreted into the blood and circulated to several systems, and passed through the blood-brain barrier (BBB). Irisin consists of an N-terminal fibronectin III (FN III)-like domain attached to a flexible C-terminal tail and a continuous inter-subunit β-sheet dimer (Mahgoub et al., 2018). This structure is stabilized because of the hydrogen bonds and its interactions between the side chains of adjacent subunits, especially between Arg-75 and Glu-79, thus protecting the dimer ends and Trp-90/Trp-90 (Schumacher et al., 2013). Peroxisome proliferator-activated receptor γ (PPARγ) coactivator-1 α (PGC-1α) is the main regulator of FNDC5 in skeletal muscles in rodents and humans (Huh et al., 2014). Endurance exercise activates on PGC-1α to induce cleavage of FNDC5 to irisin. PGC-1α interacts with a wide range of transcription factors, and it is expressed in skeletal muscle, heart, and brain (Lin et al., 2002). It interacted with several pathways such as the p38 mitogen-activated protein kinase (MAPK) pathway stimulated by exercise (Akimoto et al., 2005), 5’ adenosine monophosphate-activated protein kinase (AMPK) pathway (Chen et al., 2019), Sirtuin1 (Sirt1) pathway (Safarpour et al., 2020), and the cyclic adenosine monophosphate (cAMP) response element-binding (CREB) pathway. The cAMP-mediated PGC-1α/CREB signaling bolstered the expression of FNDC5 (Yang et al., 2018). Besides, FNDC5 and irisin expressed in many tissues, such as skeletal muscle, pancreas, brown adipose tissue (BAT), liver, and brain, especially in the hippocampus and hypothalamus, are important for memory and cognition (Dun et al., 2013; Varela-Rodríguez et al., 2016).
Irisin was associated with various metabolic diseases such as diabetes, cardiovascular disease, and obesity (Polyzos et al., 2018). It induced the expression of mitochondrial uncoupling protein 1 (UCP1) (Castillo-Quan, 2012), increasing thermogenesis and converting white adipose tissue (WAT) into BAT. Furthermore, irisin exerted favorable effects on glucose metabolism to maintain glucose homeostasis and improve insulin resistance, of which mechanisms involved β cell regeneration (Natalicchio et al., 2017), reducing gluconeogenesis and promoting glycogen synthesis (Polyzos et al., 2013; Roca-Rivada et al., 2013). Besides, irisin performed a protective function on lipid metabolism involving several pathways such as the AMPK-SREBP2 pathway (Tang et al., 2016). It was also antioxidative, anti-inflammatory, and attenuating apoptosis, functioning to alleviate mitochondrial dysfunction (Mazur-Bialy et al., 2017b; Tu et al., 2020; Zhang et al., 2020). Many studies have reported that irisin had neuroprotective functions in AD (Kim and Song, 2018; Lourenco et al., 2019; de Freitas et al., 2020). Lourenco et al. (2019) elucidated that FNDC5/irisin was decreased in AD brains and CSF and in AD experimental models, but there was no significance in plasma irisin levels. Conti et al. (2019) reported a slight increase in irisin serum levels in patients with AD. Zhang et al. (2021) suggested that serum irisin might be a biomarker of cognitive decline in vascular dementia. Bičíková et al. (2021) reported that movement was a positive modulator of aging and the PPARγ is a critical link between mental function and aging. FNDC5/irisin is stimulated by PGC-1α, indicating irisin might be the mediator of muscle and brain cross talk. Some clinical observations and mechanisms were reported.
We try to summarize the research on the relationship between irisin and cognitive impairment and to understand the mechanisms of direct neuroprotective and indirect risk reduction. This study intended to explore whether irisin is a potential serum predictor of cognitive impairment in the elderly and an underlying mediator of muscle-brain cross talk to support exercise therapy for patients with dementia.
Accumulating evidence is supporting the existence of muscle brain cross talk, a muscle-brain endocrine loop (Pedersen, 2019). Brain sensed exercise indirectly via adiponectin and liver-derived proteins such as fibroblast growth factor 21 (FGF21) and insulin-like growth factor 1 (IGF1), and muscle secreted myokines to regulate the brain function as a loop. The exercise was believed to decrease the risk of dementia (Santos-Lozano et al., 2016), delay the cognitive decline in patients with neurodegenerative disorders and prevent stress, anxiety, and depression (Pedersen and Saltin, 2015). The underlying mechanism might be the muscle brain cross talk. The physical activity enhanced circulating levels of myokines to enable the direct cross talk of muscle and brain, affecting neuronal proliferation and differentiation, synaptic plasticity, memory, and learning (Scisciola et al., 2021).
The exercise was tightly related to the PGC1-α/FNDC5/BDNF pathway. FNDC5 gene expression was elevated following the increased PGC-1α expression induced by exercise both in central and peripheral organs, which stimulated the expression of BDNF in the brain (Boström et al., 2012). Irisin, as a myokine dissected from FNDC5, was also mediated by PGC-1α and passed through the BBB to increase the BDNF expression and enhance learning, memory, and mood (Lourenco et al., 2019). On the one hand, periphery irisin delivered to the brain and overexpressed irisin in the brain increased BDNF. On the other hand, knockdown of FNDC5 reduced the central BDNF expression (Severinsen and Pedersen, 2020). Figure 1 elucidated that irisin acted as a mediator of muscle brain cross talk and the effects of FNDC5/irisin on elderly cognition.

Figure 1. Effects of FNDC5/irisin on elderly dementia and cognition. Exercise promoted PGC-1α, which stimulated fibronectin type III domain-containing protein 5 (FNDC5) to be dissected into irisin. Irisin was shed into a blood vessel, circulating the whole body and passing through blood–brain barrier. Irisin might increase the expression of brain-derived neurotrophic factors, which improved synaptic plasticity, neuronal survival, neuronal differentiation, and neuronal health, thus cognition. FNDC5 was also highly expressed in hippocampus and stimulated by peroxisome proliferator-activated receptor γ (PPARγ) coactivator-1 α (PGC-1α) to be cleaved into irisin. Besides, irisin acts with its anti-inflammation and anti-oxidative effects to defend cognition deficits. Furthermore, FNDC5/irisin might have positive connections to dementia-related diseases, such as coronary artery disease, hypertension, heart failure, stroke, and Parkinson’s disease, and depression to protect against dementia.
Brain-derived neurotrophic factor expresses highly in the brain, and it has considerable effects on synapses (Lu et al., 2014). It, mostly released from microglia and astrocytes, acts to promote synaptic plasticity, neuronal survival, neuronal differentiation, and neuronal health (Binder and Scharfman, 2004; Zuccato and Cattaneo, 2009). It was well-related to neurofunction and cognition. BDNF is bound to tropomyosin-related kinase B (TrKB) receptor to exert considerable effects. Decreased BDNF/TrkB activity resulted in neurodegeneration. Downregulation of BDNF/TrkB caused neuroinflammation, increasing inflammatory cytokines such as IL-1β and IL-6. Then triggered the JAK2/STAT3 pathway, resulting in the upregulation of C/EBPβ/AEP signaling, which led to Aβ precursor protein and Tau protein cleavage, and the Aβ and Tau alterations finally caused cognitive impairment (Wang Z. H. et al., 2019). Many studies reported BDNF levels decreased in AD patients and MCI (Tanila, 2017).
Circulating and central irisin acted on the brain to exert beneficial effects. Irisin bound and modified the function of neurotransmitter receptors in the forebrain, then neurons. The receptor of irisin in the brain was integrin-αV/β5 heterodimers (Jackson et al., 2021). Recombinant irisin stimulated the cAMP/PKA/CREB pathway in human cortical slices (Lourenco et al., 2019). CREB protein is a cellular transcription factor that plays a widely confirmed role in neuronal plasticity and long-term memory formation in the brain (Sen and Stress, 2019). Irisin increased cAMP and phosphorylated CREB (pCREB) in mouse hippocampal slices, which bolstered the expression of BDNF. According to the study by Lourenco et al. (2019), irisin-induced CREB phosphorylation was mediated by PKA. Fahimi et al. (2017) reported that mice after exercises appeared appreciable increase in BDNF mRNA and protein levels, distinctively elevated synaptic load in the dentate gyrus, and increased irisin and TrkB receptor levels in the astrocytes, indicating that irisin might mediate the effects of exercise on brain function and could be a messenger of periphery and central cross talk. Zsuga et al. (2016) proposed that irisin may be a mediator between exercise and reward-related learning and motivation through the irisin-BDNF/TrKB-MEK/ERK-mTOR pathway. The TrKB linked with dopamine 3 (D3) receptor signaling such as PI3/Akt/mTOR pathway was also involved. The two pathways were under the control of BDNF and caused increased dopamine content, neuronal plasticity, and raised neuronal survival (Collo et al., 2014). Moon et al. (2013) described that irisin performed favorable effects on hippocampal neuron proliferation primarily via the STAT3 signaling pathway. Activation of STAT3 has been confirmed to correlate with stimulating hippocampal neurogenesis (Jung et al., 2006).
FNDC5 was highly expressed in the brain especially in the hippocampus (Wrann et al., 2013; Lourenco et al., 2019). Neuronal FNDC5 gene expression was also regulated by PGC-1α. The orphan nuclear receptor estrogen-related receptor alpha (ERRα) was a central metabolic regulator interacting with PGC-1α (Schreiber et al., 2004). Wrann et al. (2013) found that ERRα was up-regulated in the hippocampus upon exercise. Furthermore, FNDC5 regulated BDNF gene expression in a cell-autonomous manner, and BDNF decreased FNDC5 gene expression as a part of a potential feedback loop. Elevated expression of FNDC5 strikingly up-regulated BDNF gene expression. Moreover, peripheral delivery of FNDC5 also increased BDNF expression in the hippocampus, and ERK1/2 was a critical regulator of FNDC5 expression and function on neuronal differentiation (Hosseini Farahabadi et al., 2015; Wrann, 2015). In addition to the direct regulation of FNDC5 to BDNF, irisin was also processed from FNDC5 in the hippocampus. Thus, FNDC5/irisin acted as a messenger of muscle brain cross talk, influencing the neurogenesis in cognitive impairment, in particular through the neuroprotective effects of BDNF.
Emerging evidence suggested the importance of inflammation in the pathogenesis of AD and mild cognitive impairment (Holmes, 2013; Shen et al., 2019). According to a meta-analysis of 170 studies, patients with AD and MCI were accompanied with elevated inflammatory markers in both CSF and periphery, such as C-reactive protein (CRP), interleukin-6 (IL-6), soluble tumor necrosis factor receptor 1 (sTNFR1), soluble tumor necrosis factor receptor 2(sTNFR2), alpha1-antichymotrypsin (α1-ACT), IL-1β, soluble CD40 ligand, IL-10, monocyte chemoattractant protein-1 (MCP-1), transforming growth factor-beta 1(TGF-β1), soluble triggering receptor expressed on myeloid cells 2 (sTREM2), and so on (Shen et al., 2019).
The most common neuroinflammation is postoperative. Disruption of the BBB is the hallmark of neuroinflammation; BBB dysfunction like increased BBB permeability has been regarded as accounting for cognitive impairment (Yang et al., 2017). Surgical trauma induced the innate immune system of the brain through the nuclear factor-κB (NF-κB) pathway, leading to endothelial dysfunction and increased permeability of the BBB (Alam et al., 2018). The neuroinflammation consequences included neuronal apoptosis, damaged hippocampal neurogenesis, and impaired synaptic plasticity connections, resulting in neurodegenerative diseases (Zhang et al., 2016; Feng et al., 2017; Alam et al., 2018).
Another type of neuroinflammation is obesity-related inflammation. Obesity is related to chronic low-grade systemic inflammation (Gregor and Hotamisligil, 2011; Spencer, 2013). Inflammatory cascade was initiated by the stimulation of free fatty acid and lipopolysaccharide (LPS) receptor, toll-like receptor 4 (TLR4) on immune cells (Shu et al., 2012). The downstream factors of the TLR family signaling involve the adapter molecule MyD88, which activated NF-κB and MAPK pathways. Both of them were important for the production of cytokines and chemokines (Trinchieri and Sher, 2007; Lim and Staudt, 2013). Maric et al. (2014) suggested that the hypothalamic mRNA expression of IL-1β, IL-6, and TNF-α significantly increased in high-saturated fat (HSF)-diet rats. Qin et al. (2007) investigated that LPS-induced MAPK and STAT-3 activation, as well as the expression of IL-10, made a difference to the suppressor of cytokine signaling 3 (SOCS3) transcription and expression in macrophages and microglia, which alleviated adaptive and innate immune responses. SOCS3 activated the ERK-MAPK pathway, inhibited the NF-κB pathway, and offended cAMP-mediated signaling (Qin et al., 2007). In addition, neuroinflammation was related to microglia 1 (M1), a pro-inflammatory cell, and the anti-inflammatory microglia 2 (M2) (Sica and Mantovani, 2012). Similarly, astrocytes also have two phenotypes, pro-inflammatory astrocytes 1 (A1) and anti-inflammatory astrocytes 2 (A2) (Kwon and Koh, 2020). As a result, the neuroinflammation is under control of the polarization status of M1/M2 and A1/A2.
Irisin has already been confirmed to have anti-inflammatory effects (Pukajło et al., 2015). FNDC5 has been confirmed to attenuate adipose tissue inflammation through the AMPK pathway to induce macrophage polarization in obese mice (Xiong et al., 2018). Irisin prevented LPS-mediated liver injury by inhibiting apoptosis, nod-like receptor pyrin-3 (NLRP3) inflammasome activation, and NF-κB signaling (Li et al., 2021). Mazur-Bialy (2017) demonstrated that irisin not only promoted the activity and proliferation of macrophages and phagocytosis but also attenuated the respiratory burst of macrophages, which increased immunocompetent activity. Mazur-Bialy et al. (2017a) reported that irisin exerted its anti-inflammatory effects by downregulating the NF-κB pathway, reducing TNF-α, IL-6, and MCP-1 in adipocyte 3T3 L1 cell line, thus attenuating the obesity-related neuroinflammation. Irisin was proved to improve memory and cognition in diabetic mice by reducing the expression of IL-1β and IL-6 in the murine hippocampus (Wang K. et al., 2019). The underlying mechanism was by downregulating the P38, STAT3, and NFκB pathways, which was related to the cytokine cascade. The reactive oxygen species-NLRP3 (ROS-NLRP3) pathway was also involved in the inhibition of irisin on the neuroinflammation (Peng J. et al., 2017). Furthermore, irisin played a pivotal role in the phenotypic switch of adipose tissue macrophages from M1 to M2 to regulate neuroinflammation (Dong et al., 2016). Irisin was also involved in autophagy, which affected Tau proteins in dementia (Pesce et al., 2020). Different pathways involved in how irisin affected autophagy, such as the AMPK/SIRT1/PGC-1α pathway in pancreatic β cells in insulin resistance stage (Li Q. et al., 2019), and MAPK pathways in the hepatic I/R injury model (Bi et al., 2020). Although the mechanisms on how irisin directly influenced central autophagy were scarce, there was a consensus on the link between irisin and AMPK. The indirect effects of irisin in autophagy are reliable, and the direct pathway still needs to explore. Table 1 summarizes the experimental studies suggesting the roles of FNDC5/irisin in inflammation.
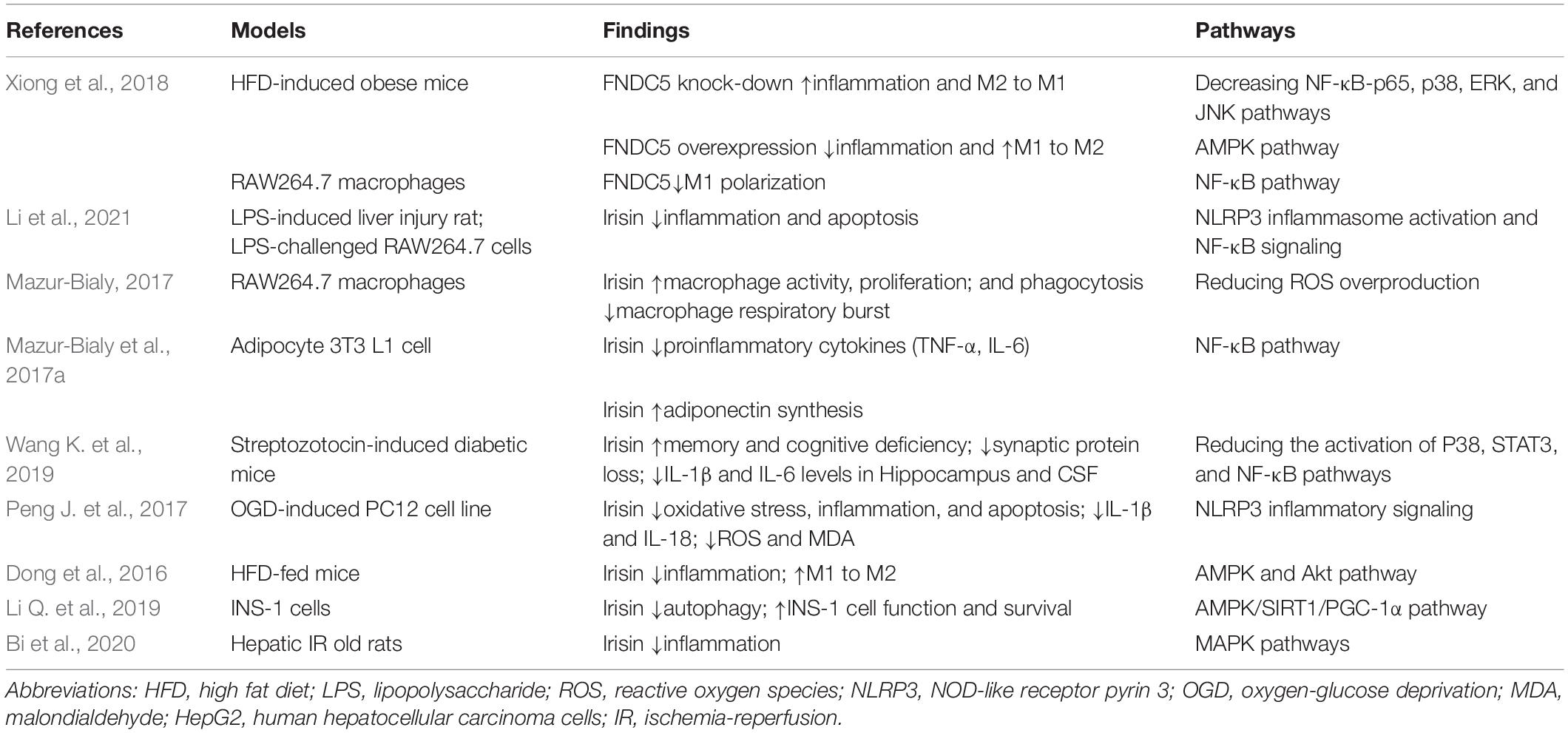
Table 1. Experimental studies suggesting the roles of FNDC5/irisin in inflammation.
Oxidative stress is critical in elderly cognitive impairment and AD (Chen and Zhong, 2014). The mechanisms of oxidative stress in AD included mitochondrial dysfunction, metal accumulation, hyperphosphorylated Tau protein, and inflammation. Mitochondrial dysfunction was mainly associated with ROS production resulting from Aβ (Perez Ortiz and Swerdlow, 2019). Increased Aβ1–40 and Aβ1–42 and decreased ATP synthesis and ATPase activity were reported to promote ROS generation in mitochondria (Sharma et al., 2021). Metal ions, such as Cu, Zn, and Fe, were perceived to play a pivotal role in AD (Faller and Hureau, 2012). Metal ions accumulation was also associated with Aβ for its metal binding sites for Zn2+, Cu2+, and Fe3+. Theoretically, Aβ binds to Cu2+ or Fe3+ resulting in reduced Cu+ and Fe2+, respectively. The binding was accompanied by the production of hydrogen peroxide (H2O2), which reacted with Fe2+ to generate Fe3+ and hydroxyl radicals (OH) (Gaeta and Hider, 2005; Chen and Zhong, 2014). Metal mal-metabolism increased the oxidative stress. Violet et al. (2014) suggested the Tau protein alterations contributed to the impaired safeguarding function of DNA and RNA, promoting the aggregation of nucleic acid oxidative damage in the AD brain. Finally, as mentioned before, the inflammation arose the generation of ROS.
FNDC5/irisin has been confirmed the anti-oxidative effects in many studies. Zhang et al. (2020) suggested that FNDC5 decreased ROS production, MDA level, and NADPH oxidase activity via its subunit p67phox and increased SOD1 and SOD2 expression in doxorubicin-treated hearts. Besides, FNDC5/irisin exerted the anti-oxidative effects via the AKT/GSK3β/FYN/Nrf2 signaling in an mTOR-independent manner. Wang et al. (2020) reported that irisin attenuated oxidative stress via 8-OHdG and reversed Sirt3 and UCP-1 pathways to promote mitochondrial membrane potential (MMP), ATP production, and the catalase to alleviate reactive oxygen radical generation, mitochondrial fusion and fission in the osteoarthritis model. Irisin targeted mitochondria to promote SOD-2 activity and prevented the loss of MMP, decreased the ROS activity, and finally relieved the oxidative stress in the ischemia/reperfusion (I/R) heart (Wang et al., 2018). Besides, in an ischemia/reperfusion (I/R) liver model, irisin was shown to reduce oxidative stress via improving UCP-2 expression, which led to reduced ROS production, restrained mitochondrial fission, and increased mitochondrial DNA copy to improve mitochondrial biogenesis (Bi et al., 2019). The Nrf2/HO-1/HMGB1 signaling participated in the anti-oxidative performance of irisin, increasing the expression of anti-oxidative factors such as SOD-1, glutathione peroxidase (GPx), and catalase-9 (Cat-9) (Mazur-Bialy and Pocheć, 2021). Activation of the AMPK-Sirt1-PGC-1α pathway and Akt/ERK1/2 pathway were involved in the irisin’s anti-oxidative effect (Li et al., 2017; Wu et al., 2020). Table 2 summarizes the experimental studies suggesting the roles of FNDC5/irisin in oxidative stress.
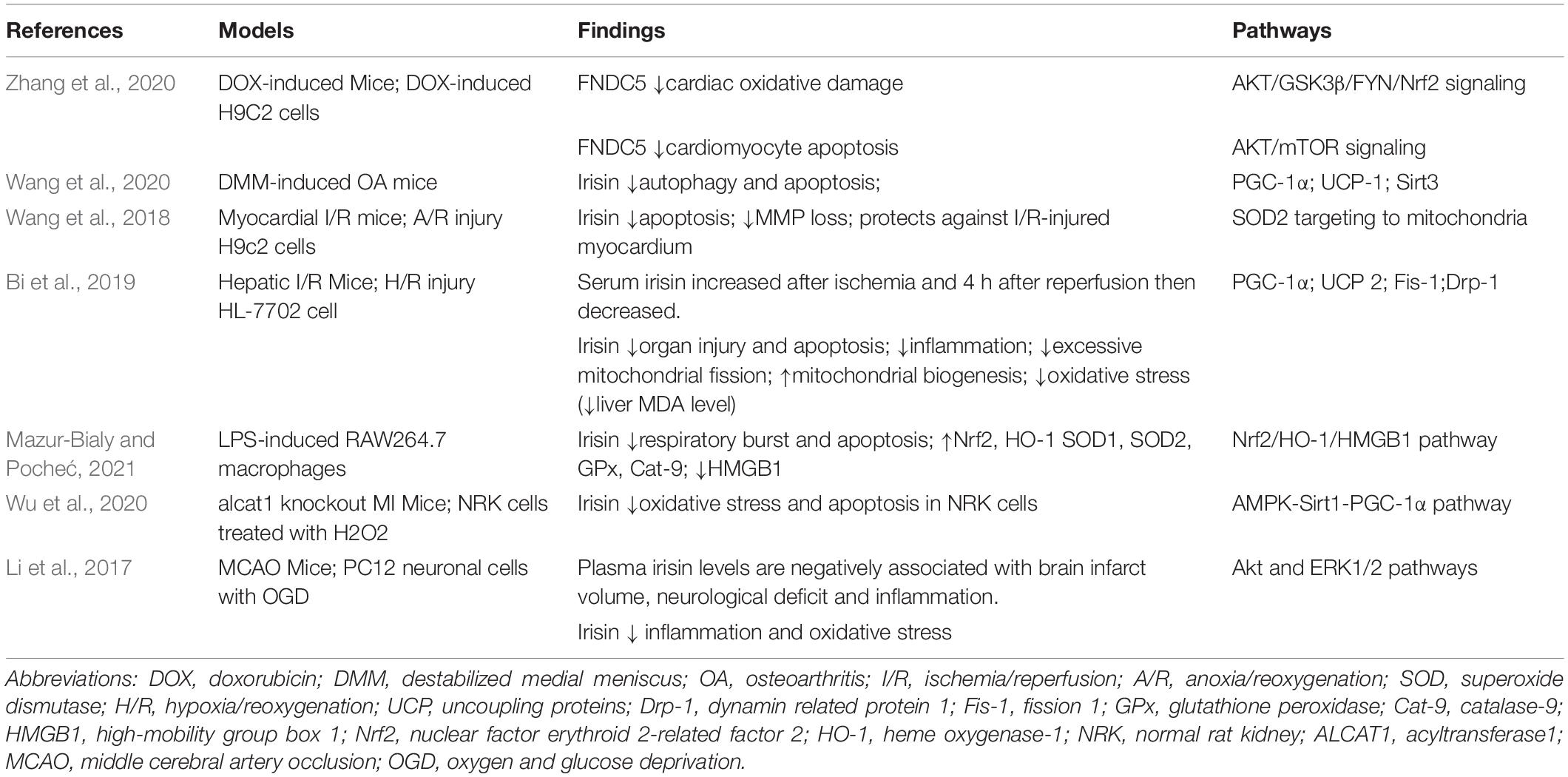
Table 2. Experimental studies suggesting the roles of FNDC5/irisin in oxidative stress.
Coronary artery disease (CAD) was associated with dementia as they shared common risk factors such as aging, obesity, type 2 diabetes (T2DM), and hypercholesterolemia. The prevalence of both dementia and CAD increases with age, with the prevalence of dementia in those with acute myocardial infarction (AMI) increasing from 1.2% in those aged 65–69 years to 14.8% in those aged above 85 years (Fowkes et al., 2016).
Various studies suggested serum irisin levels were decreased in patients with CAD, indicating the positive effects of irisin on CAD (Khorasani et al., 2019; Wang S. et al., 2019; Guo et al., 2020). In a myocardial infarction (MI) mouse model, irisin appeared to suppress cardiomyocyte apoptosis and fibrosis and promote angiogenesis via the ERK signaling, which collectively improved the cardiac function and reduced the infarct size of the post-MI model (Liao et al., 2019). Zhao et al. (2016) found that in histone deacetylases (HDAC)-over-expressed H9c2 cardio-myoblasts that went through hypoxia/reoxygenation-induced injury, irisin treatment increased cardio-myoblast survival and decreased the LDH release to alleviate cytotoxicity. Besides, irisin repressed the cell apoptosis via reducing active-caspase 3 and annexin V signals, mitigating the loss of MMP to protect mitochondrial damage. Furthermore, irisin held back the opening of mitochondrial permeability transition pore, which was critical for myocardial injury.
Hypertension is associated with an increased incidence of vascular dementia (Sharp et al., 2011). Midlife systolic blood pressure (SBP) was suggested to be a significant predictor of cognition that deficits later in life (Launer et al., 1995). In the elderly, dysfunction of cerebral autoregulation led to vulnerable cerebral hemodynamics. Autoregulation protected the brain from hypertension but increased the risk of cerebral hypotension. Inappropriate antihypertensive therapy might further increase the risk of chronic cerebral hypoperfusion and subsequent dementia (Feldstein, 2012). Higher diastolic blood pressure (DBP) and lower SBP were correlated with impaired cognition (Nilsson et al., 2007; Tsivgoulis et al., 2009).
Irisin made a difference in regulating blood pressure through central and peripheral pathways; central irisin increased cardiac output and blood pressure by activating hypothalamic paraventricular nucleus of the hypothalamus (PVN) neurons, while peripheral irisin secreted from skeletal muscle reduced blood pressure via Adenosine triphosphate-sensitive potassium (KATP) channels to dilate vessels (Zhang et al., 2015). Besides, Irisin improved hypertension by protecting endothelial function via the AMPK-Akt-eNOS-NO and Nrf2 signaling pathway, the Nrf2 signaling pathway also participated in alleviating oxidative stress in the hypothalamus (Fu et al., 2016; Huo et al., 2020). Huang et al. (2022) proposed that irisin inhibited the NF-κB signaling pathway to lower blood pressure, along with reduced angiotensin II type 1 receptor (AT1R) expression and function.
A considerable number of patients with heart failure (HF) have cognitive problems (Cannon et al., 2017). Vascular dysfunction and loss of cardiac perfusion pump function can trigger the typical AD feature such as Aβ accumulation and hyperphosphorylated Tau tangles, as HF and AD shares common risk factors like inflammation and oxidative stress (Daniele et al., 2020).
Irisin exerted positive influences on mitochondrial dysfunction, oxidative stress, metabolic imbalance, and energy expenditure in HF (Ho and Wang, 2021). Cohort and experimental studies were conducted to elucidate the correlation between irisin and HF. Several cohorts showed increased serum irisin levels in patients with HF (Shen et al., 2017; Kalkan et al., 2018; Abd El-Mottaleb et al., 2019). Peng Q. et al. (2017) suggested that irisin ameliorated H2O2-induced apoptosis in H9c2 cardio-myoblasts and improved cell viability via miR-19b/PTEN/AKT/mTOR pathway. Li R. et al. (2019) found that irisin-induced protective autophagy and alleviated apoptosis signaling attenuated the myocardial hypertrophy and cardiomyocytes apoptosis. The AMPK-ULK1 pathway might be involved in the underlying mechanisms (Li et al., 2018).
Stroke is a pronounced disease related to cognition impairment and contributes to damaged life quality (Obaid et al., 2020). Stroke is divided into the ischemic and hemorrhagic stroke, the former makes up 85% (Amarenco et al., 2009; Beal, 2010). A total of 23.9% of older stroke survivors developed dementia (Allan et al., 2011). Taking ischemic stroke as an example, brain injury secondary to the stroke was a result of the post-stroke excitotoxicity, oxidative and nitrative stress, inflammation, and apoptosis (Khoshnam et al., 2017). Besides, Goulay et al. (2020) have reported that stroke exacerbated the deposition of Aβ.
Irisin has been reported to perform neuroprotective effects on stroke (Liu et al., 2020). Irisin mitigated brain injury after stroke via inhibiting inflammation and oxidative stress and preventing BBB dysfunction (Peng J. et al., 2017; Guo et al., 2019). Jin et al. (2019) suggested that irisin attenuated the brain injury after the cerebral ischemia/reperfusion (I/R) injury especially in the hippocampus region through the Notch signaling pathway. Irisin promoted the Notch1, Notch intracellular domain (NICD), and Hes1 expression, which were reported to exert effects in AD and other neurodegenerative diseases. Irisin alleviated neuronal apoptosis, accompanied by decreasing the caspase-3 expression, a critical apoptotic effector. Besides, irisin reduced the inflammation, decreasing the TNF-α and IL-1β levels (Berezovska et al., 1998; Alberi et al., 2013). Yu et al. (2020) reported that irisin protected the neurological function in a middle cerebral artery occlusion (MCAO) I/R injury model via suppressing the TLR4 and NF-κB pathways. Others also elucidated the neuroprotective effects of irisin in mice with MCAO and OGD neuronal cells via Akt and ERK1/2 signaling pathways (Li et al., 2017). Irisin relieved the post-ischemic inflammation by downregulating TNF-α and IL-6 expression, suppressed the microglial infiltration, and decreased the MPO-1+ cell numbers, as well as reduced the post-ischemic oxidative stress by decreasing the levels of 4-HNE and MDA. Furthermore, mitochondrial dynamics were involved in the ischemic stroke, and mitochondrial defects are critical for AD (Yan et al., 2013; Anzell et al., 2018). Irisin improved mitochondrial function via AMPK pathway as the AMPK was a guardian of mitochondrial homeostasis (Tang et al., 2016; Herzig and Shaw, 2018; Siteneski et al., 2018; Xin et al., 2020). In summary, irisin exerted neuroprotective effects after stroke to prevent cognitive impairment primarily through its anti-inflammatory and anti-oxidative effects, as well as the beneficial effects on mitochondria.
Parkinson’s disease is the second most frequent senile neurodegenerative disease (Mhyre et al., 2012). Patients with PD often developed cognitive deficits and dementia, especially in elderly patients (Aarsland et al., 2017). PD-dementia is a classic type of dementia.
Irisin played a protective role in PD. In a mouse model of PD, irisin treatment prevented dopaminergic neurons from apoptosis and degeneration (Zarbakhsh et al., 2019). Mahalakshmi et al. (2020) elucidated the benefits of exercise on PD, and irisin was a mediator of exercise-induced BDNF. Raefsky and Mattson (2017) suggested that irisin might protect neuronal mitochondria function in PD via antioxidation, autophagy, and DNA repair regulations.
Depression and dementia often occur at the same time in the elderly (Bennett and Thomas, 2014). Depression is both the risk factor and prodrome of dementia (Gutzmann and Qazi, 2015). The interreaction of depression and dementia is complex.
Irisin improved depressive neuropathology by regulating mitochondria function via PGC-1α signaling and modifying synaptic plasticity via BDNF signaling (Jo and Song, 2021). Hou et al. (2020) proposed that irisin attenuated the postoperative depressive-like behavior and reduced neuron death and cytokines release from astrocytes through inhibiting the surface expression of epidermal growth factor receptors (EGFR) in the mice model. Siteneski et al. (2018) also suggested that central irisin administration manifested antidepression effect, associated with the adjustment of gene expression of PGC-1α, FNDC5, and BDNF in the hippocampus and prefrontal cortex of mice.
There is a long way to intervene and delay the progression of elderly cognitive impairment. Based on the irisin secretion and function to optimize the exercise protocol such as the amount of exercise, the form of exercise, and the duration of exercise, further research is needed. Factors affecting exercise, such as age, frailty, sarcopenia, and fracture, also need to be considered. Although many studies have been reported to support the favorable effects of FNDC5/irisin, there are some limitations. Many studies are based on experimental studies, and direct studies of irisin on central autophagy are scarce. Besides, the difference of plasma irisin levels alterations in patients with dementia was not significant and has not reached a consensus. Interfering factors such as age, gender, race, and disease duration differences cannot be ignored. There are also some controversial results and views. On the one hand, Raschke et al. (2013) argued that the beneficial effect of irisin observed in mice can be translated to humans. Although there are many registered clinical trials to clarify the effects of irisin on the human body, large-scale clinical research and long-term follow-up are required to study the relationship between FNDC5/irisin and cognition. Besides, to carry out the animal experiments and clinical research simultaneously and to conduct comparative analysis are very necessary to elucidate the difference in FNDC5/irisin effects in the mice and human body. On the other hand, the current irisin detection method is still insufficient. ELISA has been widely used in the examination of irisin levels in serum or other specimens in humans and animals. However, Albrecht et al. (2015) argued that ELISA kit for irisin may not be accurate. Besides, ELISA can be influenced by a series of factors such as preservation conditions, temperature, antibody, and operational contingency. As a result of the conflicting opinions, conducting comparative studies on the sensitivity and specificity of current ELISA kits is a research direction. A high-quality meta-analysis or systematic review of the efficacy of ELISA kits for irisin also can be considered. Most of the ELISA kits for irisin were for laboratory research only, not for drug, diagnostic, or other use. Exploring new methods with high sensitivity and specificity as well as diagnostic value in clinical conditions is also the direction of future efforts, such as the application of sensors or nanotechnology.
Cognitive impairment is a worldwide public health problem, which seriously affects the quality of life of the elderly and increases the burden of care. Clarifying the pathological mechanism of dementia and exploring drugs to prevent, treat, and delay the course of dementia have always been the direction of efforts. Physical exercise and lifestyle are believed to defend against cognitive decline in the elderly. Irisin might be a mediator of muscle and brain cross talk mainly through the PGC-1α/FNDC5/BDNF pathway. More information is needed to optimize exercise protocols based on irisin for patients with dementia. Our review discussed the favorable effects of irisin on cognitive impairment, such as the positive effect irisin on neurogenesis and synapse; anti-inflammatory and anti-oxidative effects; and possible connections of irisin on dementia-related diseases such as CAD, hypertension, HF, stroke, PD, and depression. The serum irisin level alterations in dementia have not reached a consensus. Large-scale clinical research and long-term follow-up are required to explore whether serum irisin is a diagnostic or prognostic factor for dementia. The current detection method for irisin is still limited to ELISA. It is also an exploratory direction to find more sensitive, specific, and simple detection methods.
JP participated in literature collection, preparation, and wrote the draft. JW supervised the whole project. Both authors participated in the conception and study design, contributed to the manuscript revision, and approved the submitted version.
This work was supported by China National Key R&D Program (Nos. 2018YFC2002100 and 2018YFC2002103) to JW.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank the participants.
Aarsland, D., Creese, B., Politis, M., Chaudhuri, K. R., Ffytche, D. H., Weintraub, D., et al. (2017). Cognitive decline in Parkinson disease. Nat. Rev. Neurol. 13, 217–231.
Abd El-Mottaleb, N. A., Galal, H. M., El Maghraby, K. M., and Gadallah, A. I. (2019). Serum irisin level in myocardial infarction patients with or without heart failure. Can. J. Physiol. Pharmacol. 97, 932–938. doi: 10.1139/cjpp-2018-0736
Akimoto, T., Pohnert, S. C., Li, P., Zhang, M., Gumbs, C., Rosenberg, P. B., et al. (2005). Exercise stimulates Pgc-1alpha transcription in skeletal muscle through activation of the p38 MAPK pathway. J. Biol. Chem. 280, 19587–19593. doi: 10.1074/jbc.M408862200
Alam, A., Hana, Z., Jin, Z., Suen, K. C., and Ma, D. (2018). Surgery, neuroinflammation and cognitive impairment. EBioMedicine 37, 547–556. doi: 10.1016/j.ebiom.2018.10.021
Alberi, L., Hoey, S. E., Brai, E., Scotti, A. L., and Marathe, S. (2013). Notch signaling in the brain: in good and bad times. Ageing Res. Rev. 12, 801–814. doi: 10.1016/j.arr.2013.03.004
Albrecht, E., Norheim, F., Thiede, B., Holen, T., Ohashi, T., Schering, L., et al. (2015). Irisin - a myth rather than an exercise-inducible myokine. Sci. Rep. 5:8889. doi: 10.1038/srep08889
Allan, L. M., Rowan, E. N., Firbank, M. J., Thomas, A. J., Parry, S. W., Polvikoski, T. M., et al. (2011). Long term incidence of dementia, predictors of mortality and pathological diagnosis in older stroke survivors. Brain 134, 3716–3727. doi: 10.1093/brain/awr273
Amarenco, P., Bogousslavsky, J., Caplan, L. R., Donnan, G. A., and Hennerici, M. G. (2009). Classification of stroke subtypes. Cerebrovasc. Dis. 27, 493–501. doi: 10.1159/000210432
Amidfar, M., de Oliveira, J., Kucharska, E., Budni, J., and Kim, Y. K. (2020). The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 257:118020. doi: 10.1016/j.lfs.2020.118020
Anzell, A. R., Maizy, R., Przyklenk, K., and Sanderson, T. H. (2018). Mitochondrial quality control and disease: insights into ischemia-reperfusion injury. Mol. Neurobiol. 55, 2547–2564. doi: 10.1007/s12035-017-0503-9
Beal, C. C. (2010). Gender and stroke symptoms: a review of the current literature. J. Neurosci. Nurs. 42, 80–87. doi: 10.1097/jnn.0b013e3181ce5c70
Bennett, S., and Thomas, A. J. (2014). Depression and dementia: cause, consequence or coincidence? Maturitas 79, 184–190. doi: 10.1016/j.maturitas.2014.05.009
Berezovska, O., Xia, M. Q., and Hyman, B. T. (1998). Notch is expressed in adult brain, is coexpressed with presenilin-1, and is altered in Alzheimer disease. J. Neuropathol. Exp. Neurol. 57, 738–745. doi: 10.1097/00005072-199808000-00003
Bi, J., Yang, L., Wang, T., Zhang, J., Li, T., Ren, Y., et al. (2020). Irisin improves autophagy of aged hepatocytes via increasing telomerase activity in liver injury. Oxid. Med. Cell Longev. 2020:6946037. doi: 10.1155/2020/6946037
Bi, J., Zhang, J., Ren, Y., Du, Z., Li, Q., Wang, Y., et al. (2019). Irisin alleviates liver ischemia-reperfusion injury by inhibiting excessive mitochondrial fission, promoting mitochondrial biogenesis and decreasing oxidative stress. Redox Biol. 20, 296–306. doi: 10.1016/j.redox.2018.10.019
Bičíková, M., Máčová, L., Jandová, D., Tøískala, Z., and Hill, M. (2021). Movement as a positive modulator of aging. Int. J. Mol. Sci. 22:6278. doi: 10.3390/ijms22126278
Binder, D. K., and Scharfman, H. E. (2004). Brain-derived neurotrophic factor. Growth Fact. 22, 123–131.
Boström, P., Wu, J., Jedrychowski, M. P., Korde, A., Ye, L., Lo, J. C., et al. (2012). A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 481, 463–468. doi: 10.1038/nature10777
Cannon, J. A., Moffitt, P., Perez-Moreno, A. C., Walters, M. R., Broomfield, N. M., McMurray, J. J. V., et al. (2017). Cognitive impairment and heart failure: systematic review and meta-analysis. J. Card. Fail. 23, 464–475. doi: 10.1016/j.cardfail.2017.04.007
Castillo-Quan, J. I. (2012). From white to brown fat through the PGC-1α-dependent myokine irisin: implications for diabetes and obesity. Dis. Model. Mech. 5, 293–295. doi: 10.1242/dmm.009894
Chen, S. Q., Ding, L. N., Zeng, N. X., Liu, H. M., Zheng, S. H., Xu, J. W., et al. (2019). Icariin induces irisin/FNDC5 expression in C2C12 cells via the AMPK pathway. Biomed. Pharmacother. 115:108930. doi: 10.1016/j.biopha.2019.108930
Chen, Z., and Zhong, C. (2014). Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 30, 271–281.
Cheng, Y. W., Chen, T. F., and Chiu, M. J. (2017). From mild cognitive impairment to subjective cognitive decline: conceptual and methodological evolution. Neuropsychiatr. Dis. Treat. 13, 491–498. doi: 10.2147/NDT.S123428
Collo, G., Cavalleri, L., and Spano, P. (2014). Structural plasticity in mesencephalic dopaminergic neurons produced by drugs of abuse: critical role of BDNF and dopamine. Front. Pharmacol. 5:259. doi: 10.3389/fphar.2014.00259
Conti, E., Grana, D., Stefanoni, G., Corsini, A., Botta, M., Magni, P., et al. (2019). Irisin and BDNF serum levels and behavioral disturbances in Alzheimer’s disease. Neurol. Sci. 40, 1145–1150. doi: 10.1007/s10072-019-03781-y
Daniele, G., DiLucia, S., Masci, P. G., and Del Monte, F. (2020). Heart and brain: complex relationships for left ventricular dysfunction. Curr. Cardiol. Rep. 22:72. doi: 10.1007/s11886-020-01318-w
de Freitas, G. B., Lourenco, M. V., and De Felice, F. G. (2020). Protective actions of exercise-related FNDC5/Irisin in memory and Alzheimer’s disease. J. Neurochem. 155, 602–611. doi: 10.1111/jnc.15039
Dong, J., Dong, Y., Dong, Y., Chen, F., Mitch, W. E., and Zhang, L. (2016). Inhibition of myostatin in mice improves insulin sensitivity via irisin-mediated cross talk between muscle and adipose tissues. Int. J. Obes. 40, 434–442. doi: 10.1038/ijo.2015.200
Dun, S. L., Lyu, R. M., Chen, Y. H., Chang, J. K., Luo, J. J., and Dun, N. J. (2013). Irisin-immunoreactivity in neural and non-neural cells of the rodent. Neuroscience 240, 155–162. doi: 10.1016/j.neuroscience.2013.02.050
Fahimi, A., Baktir, M. A., Moghadam, S., Mojabi, F. S., Sumanth, K., McNerney, M. W., et al. (2017). Physical exercise induces structural alterations in the hippocampal astrocytes: exploring the role of BDNF-TrkB signaling. Brain Struct. Funct. 222, 1797–1808. doi: 10.1007/s00429-016-1308-8
Faller, P., and Hureau, C. (2012). A bioinorganic view of Alzheimer’s disease: when misplaced metal ions (re)direct the electrons to the wrong target. Chemistry 18, 15910–15920. doi: 10.1002/chem.201202697
Feldstein, C. A. (2012). Association between chronic blood pressure changes and development of Alzheimer’s disease. J. Alzheimers Dis. 32, 753–763. doi: 10.3233/JAD-2012-120613
Feng, X., Valdearcos, M., Uchida, Y., Lutrin, D., Maze, M., and Koliwad, S. K. (2017). Microglia mediate postoperative hippocampal inflammation and cognitive decline in mice. JCI Insight 2:e91229. doi: 10.1172/jci.insight.91229
Fowkes, R., Byrne, M., Sinclair, H., Tang, E., and Kunadian, V. (2016). Coronary artery disease in patients with dementia. Coron Artery Dis. 27, 511–520. doi: 10.1097/MCA.0000000000000377
Fu, J., Han, Y., Wang, J., Liu, Y., Zheng, S., Zhou, L., et al. (2016). Irisin lowers blood pressure by improvement of endothelial dysfunction via AMPK-Akt-eNOS-NO pathway in the spontaneously hypertensive rat. J. Am. Heart Assoc. 5:e003433. doi: 10.1161/JAHA.116.003433
Gaeta, A., and Hider, R. C. (2005). The crucial role of metal ions in neurodegeneration: the basis for a promising therapeutic strategy. Br. J. Pharmacol. 146, 1041–1059. doi: 10.1038/sj.bjp.0706416
Goulay, R., Mena Romo, L., Hol, E. M., and Dijkhuizen, R. M. (2020). From stroke to dementia: a comprehensive review exposing tight interactions between stroke and amyloid-β formation. Transl. Stroke Res. 11, 601–614. doi: 10.1007/s12975-019-00755-2
Gregor, M. F., and Hotamisligil, G. S. (2011). Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 29, 415–445.
Guo, P., Jin, Z., Wu, H., Li, X., Ke, J., Zhang, Z., et al. (2019). Effects of irisin on the dysfunction of blood-brain barrier in rats after focal cerebral ischemia/reperfusion. Brain Behav. 9:e01425. doi: 10.1002/brb3.1425
Guo, W., Zhang, B., and Wang, X. (2020). Lower irisin levels in coronary artery disease: a meta-analysis. Minerva Endocrinol. 45, 61–69. doi: 10.23736/S0391-1977.17.02663-3
Gutzmann, H., and Qazi, A. (2015). Depression associated with dementia. Z. Gerontol. Geriatr. 48, 305–311.
Herzig, S., and Shaw, R. J. (2018). AMPK: guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 19, 121–135. doi: 10.1038/nrm.2017.95
Ho, M. Y., and Wang, C. Y. (2021). Role of irisin in myocardial infarction, heart failure, and cardiac hypertrophy. Cells 10:2103. doi: 10.3390/cells10082103
Holmes, C. (2013). Review: systemic inflammation and Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 39, 51–68. doi: 10.1111/j.1365-2990.2012.01307.x
Hosseini Farahabadi, S. S., Ghaedi, K., Ghazvini Zadegan, F., Karbalaie, K., Rabiee, F., Nematollahi, M., et al. (2015). ERK1/2 is a key regulator of Fndc5 and PGC1α expression during neural differentiation of mESCs. Neuroscience 297, 252–261. doi: 10.1016/j.neuroscience.2015.03.069
Hou, Z., Zhang, J., Yu, K., and Song, F. (2020). Irisin ameliorates the postoperative depressive-like behavior by reducing the surface expression of epidermal growth factor receptor in mice. Neurochem. Int. 135:104705. doi: 10.1016/j.neuint.2020.104705
Huang, Y., Chen, Q., Jiang, Q., Zhao, Z., Fang, J., and Chen, L. (2022). Irisin lowers blood pressure in Zucker diabetic rats by regulating the functions of renal angiotensin II type 1 receptor via the inhibition of the NF-κB signaling pathway. Peptides 147:170688. doi: 10.1016/j.peptides.2021.170688
Huh, J. Y., Mougios, V., Kabasakalis, A., Fatouros, I., Siopi, A., Douroudos, I. I., et al. (2014). Exercise-induced irisin secretion is independent of age or fitness level and increased irisin may directly modulate muscle metabolism through AMPK activation. J. Clin. Endocrinol. Metab. 99, E2154–E2161. doi: 10.1210/jc.2014-1437
Huo, C. J., Yu, X. J., Sun, Y. J., Li, H. B., Su, Q., Bai, J., et al. (2020). Irisin lowers blood pressure by activating the Nrf2 signaling pathway in the hypothalamic paraventricular nucleus of spontaneously hypertensive rats. Toxicol. Appl. Pharmacol. 394:114953. doi: 10.1016/j.taap.2020.114953
Jackson, T. C., Gorse, K., Herrmann, J. R., and Kochanek, P. M. (2021). Hippocampal and prefrontal cortical brain tissue levels of irisin and GDF15 receptor subunits in children. Mol. Neurobiol. 58, 2145–2157. doi: 10.1007/s12035-020-02250-4
Jin, Z., Guo, P., Li, X., Ke, J., Wang, Y., and Wu, H. (2019). Neuroprotective effects of irisin against cerebral ischemia/reperfusion injury via Notch signaling pathway. Biomed. Pharmacother. 120:109452. doi: 10.1016/j.biopha.2019.109452
Jo, D., and Song, J. (2021). Irisin acts via the PGC-1α and BDNF pathway to improve depression-like behavior. Clin. Nutr. Res. 10, 292–302. doi: 10.7762/cnr.2021.10.4.292
Jung, K. H., Chu, K., Lee, S. T., Kim, S. J., Sinn, D. I., Kim, S. U., et al. (2006). Granulocyte colony-stimulating factor stimulates neurogenesis via vascular endothelial growth factor with STAT activation. Brain Res. 1073-1074, 190–201. doi: 10.1016/j.brainres.2005.12.037
Kalkan, A. K., Cakmak, H. A., Erturk, M., Kalkan, K. E., Uzun, F., Tasbulak, O., et al. (2018). Adropin and irisin in patients with cardiac cachexia. Arq. Bras. Cardiol. 111, 39–47.
Khorasani, Z. M., Bagheri, R. K., Yaghoubi, M. A., Chobkar, S., Aghaee, M. A., Abbaszadegan, M. R., et al. (2019). The association between serum irisin levels and cardiovascular disease in diabetic patients. Diabetes Metab. Syndr. 13, 786–790. doi: 10.1016/j.dsx.2018.11.050
Khoshnam, S. E., Winlow, W., Farzaneh, M., Farbood, Y., and Moghaddam, H. F. (2017). Pathogenic mechanisms following ischemic stroke. Neurol. Sci. 38, 1167–1186. doi: 10.1007/s10072-017-2938-1
Kim, O. Y., and Song, J. (2018). The role of irisin in Alzheimer’s disease. J. Clin. Med. 7:407. doi: 10.3390/jcm7110407
Kwon, H. S., and Koh, S. H. (2020). Neuroinflammation in neurodegenerative disorders: the roles of microglia and astrocytes. Transl. Neurodegener. 9:42. doi: 10.1186/s40035-020-00221-2
Launer, L. J., Masaki, K., Petrovitch, H., Foley, D., and Havlik, R. J. (1995). The association between midlife blood pressure levels and late-life cognitive function. The Honolulu-Asia Aging Study. JAMA 274, 1846–1851. doi: 10.1001/jama.1995.03530230032026
Li, D. J., Li, Y. H., Yuan, H. B., Qu, L. F., and Wang, P. (2017). The novel exercise-induced hormone irisin protects against neuronal injury via activation of the Akt and ERK1/2 signaling pathways and contributes to the neuroprotection of physical exercise in cerebral ischemia. Metabolism 68, 31–42. doi: 10.1016/j.metabol.2016.12.003
Li, Q., Jia, S., Xu, L., Li, B., and Chen, N. (2019). Metformin-induced autophagy and irisin improves INS-1 cell function and survival in high-glucose environment via AMPK/SIRT1/PGC-1α signal pathway. Food Sci. Nutr. 7, 1695–1703. doi: 10.1002/fsn3.1006
Li, Q., Tan, Y., Chen, S., Xiao, X., Zhang, M., Wu, Q., et al. (2021). Irisin alleviates LPS-induced liver injury and inflammation through inhibition of NLRP3 inflammasome and NF-κB signaling. J. Recept. Signal. Transduct. Res. 41, 294–303. doi: 10.1080/10799893.2020.1808675
Li, R., Wang, X., Wu, S., Wu, Y., Chen, H., Xin, J., et al. (2019). Irisin ameliorates angiotensin II-induced cardiomyocyte apoptosis through autophagy. J. Cell Physiol. 234, 17578–17588. doi: 10.1002/jcp.28382
Li, R. L., Wu, S. S., Wu, Y., Wang, X. X., Chen, H. Y., Xin, J. J., et al. (2018). Irisin alleviates pressure overload-induced cardiac hypertrophy by inducing protective autophagy via mTOR-independent activation of the AMPK-ULK1 pathway. J. Mol. Cell Cardiol. 121, 242–255. doi: 10.1016/j.yjmcc.2018.07.250
Liao, Q., Qu, S., Tang, L. X., Li, L. P., He, D. F., Zeng, C. Y., et al. (2019). Irisin exerts a therapeutic effect against myocardial infarction via promoting angiogenesis. Acta Pharmacol. Sin. 40, 1314–1321. doi: 10.1038/s41401-019-0230-z
Lim, K. H., and Staudt, L. M. (2013). Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 5:a011247.
Lin, J., Wu, H., Tarr, P. T., Zhang, C. Y., Wu, Z., Boss, O., et al. (2002). Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature 418, 797–801. doi: 10.1038/nature00904
Liu, Y., Zhu, C., Guo, J., Chen, Y., and Meng, C. (2020). The neuroprotective effect of irisin in ischemic stroke. Front. Aging Neurosci. 12:588958. doi: 10.3389/fnagi.2020.588958
Lourenco, M. V., Frozza, R. L., de Freitas, G. B., Zhang, H., Kincheski, G. C., Ribeiro, F. C., et al. (2019). Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat. Med. 25, 165–175. doi: 10.1038/s41591-018-0275-4
Lu, B., Nagappan, G., and Lu, Y. (2014). BDNF and synaptic plasticity, cognitive function, and dysfunction. Handb. Exp. Pharmacol. 220, 223–250. doi: 10.1007/978-3-642-45106-5_9
Mahalakshmi, B., Maurya, N., Lee, S. D., and Bharath Kumar, V. (2020). Possible neuroprotective mechanisms of physical exercise in neurodegeneration. Int. J. Mol. Sci. 21:5895. doi: 10.3390/ijms21165895
Mahgoub, M. O., D’Souza, C., Al Darmaki, R., Baniyas, M., and Adeghate, E. (2018). An update on the role of irisin in the regulation of endocrine and metabolic functions. Peptides 104, 15–23. doi: 10.1016/j.peptides.2018.03.018
Maric, T., Woodside, B., and Luheshi, G. N. (2014). The effects of dietary saturated fat on basal hypothalamic neuroinflammation in rats. Brain Behav. Immun. 36, 35–45. doi: 10.1016/j.bbi.2013.09.011
Mazur-Bialy, A. I. (2017). Irisin acts as a regulator of macrophages host defense. Life Sci. 176, 21–25. doi: 10.1016/j.lfs.2017.03.011
Mazur-Bialy, A. I., Bilski, J., Pochec, E., and Brzozowski, T. (2017a). New insight into the direct anti-inflammatory activity of a myokine irisin against proinflammatory activation of adipocytes. Implication for exercise in obesity. J. Physiol. Pharmacol. 68, 243–251.
Mazur-Bialy, A. I., Pocheć, E., and Zarawski, M. (2017b). Anti-inflammatory properties of irisin, mediator of physical activity, are connected with TLR4/MyD88 signaling pathway activation. Int. J. Mol. Sci. 18:701. doi: 10.3390/ijms18040701
Mazur-Bialy, A. I., and Pocheć, E. (2021). The time-course of antioxidant irisin activity: role of the Nrf2/HO-1/HMGB1 axis. Antioxidants 10:88. doi: 10.3390/antiox10010088
Mhyre, T. R., Boyd, J. T., Hamill, R. W., and Maguire-Zeiss, K. A. (2012). Parkinson’s disease. Subcell. Biochem. 65, 389–455.
Moon, H. S., Dincer, F., and Mantzoros, C. S. (2013). Pharmacological concentrations of irisin increase cell proliferation without influencing markers of neurite outgrowth and synaptogenesis in mouse H19-7 hippocampal cell lines. Metabolism 62, 1131–1136. doi: 10.1016/j.metabol.2013.04.007
Natalicchio, A., Marrano, N., Biondi, G., Spagnuolo, R., Labarbuta, R., Porreca, I., et al. (2017). The myokine irisin is released in response to saturated fatty acids and promotes pancreatic β-cell survival and insulin secretion. Diabetes 66, 2849–2856. doi: 10.2337/db17-0002
Nilsson, S. E., Read, S., Berg, S., Johansson, B., Melander, A., and Lindblad, U. (2007). Low systolic blood pressure is associated with impaired cognitive function in the oldest old: longitudinal observations in a population-based sample 80 years and older. Aging Clin. Exp. Res. 19, 41–47. doi: 10.1007/BF03325209
Obaid, M., Flach, C., Marshall, I., Wolfe, C. D. A., and Douiri, A. (2020). Long-term outcomes in stroke patients with cognitive impairment: a population-based study. Geriatrics 5:32. doi: 10.3390/geriatrics5020032
Pedersen, B. K. (2019). Physical activity and muscle-brain crosstalk. Nat. Rev. Endocrinol. 15, 383–392. doi: 10.1038/s41574-019-0174-x
Pedersen, B. K., and Saltin, B. (2015). Exercise as medicine - evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 25, (Suppl. 3), 1–72. doi: 10.1111/sms.12581
Peng, J., Deng, X., Huang, W., Yu, J. H., Wang, J. X., Wang, J. P., et al. (2017). Irisin protects against neuronal injury induced by oxygen-glucose deprivation in part depends on the inhibition of ROS-NLRP3 inflammatory signaling pathway. Mol. Immunol. 91, 185–194. doi: 10.1016/j.molimm.2017.09.014
Peng, Q., Wang, X., Wu, K., Liu, K., Wang, S., and Chen, X. (2017). Irisin attenuates H(2)O(2)-induced apoptosis in cardiomyocytes via microRNA-19b/AKT/mTOR signaling pathway. Int. J. Clin. Exp. Pathol. 10, 7707–7717.
Perez Ortiz, J. M., and Swerdlow, R. H. (2019). Mitochondrial dysfunction in Alzheimer’s disease: role in pathogenesis and novel therapeutic opportunities. Br. J. Pharmacol. 176, 3489–3507. doi: 10.1111/bph.14585
Pesce, M., Ballerini, P., Paolucci, T., Puca, I., Farzaei, M. H., and Patruno, A. (2020). Irisin and autophagy: first update. Int. J. Mol. Sci. 21:7587. doi: 10.3390/ijms21207587
Petersen, R. C., Lopez, O., Armstrong, M. J., Getchius, T. S. D., Ganguli, M., Gloss, D., et al. (2018). Practice guideline update summary: mild cognitive impairment: report of the guideline development, dissemination, and implementation subcommittee of the american academy of neurology. Neurology 90, 126–135. doi: 10.1212/wnl.0000000000004826
Polyzos, S. A., Anastasilakis, A. D., Efstathiadou, Z. A., Makras, P., Perakakis, N., Kountouras, J., et al. (2018). Irisin in metabolic diseases. Endocrine 59, 260–274. doi: 10.1007/s12020-017-1476-1
Polyzos, S. A., Kountouras, J., Shields, K., and Mantzoros, C. S. (2013). Irisin: a renaissance in metabolism? Metabolism 62, 1037–1044. doi: 10.1016/j.metabol.2013.04.008
Pukajło, K., Kolackov, K., Łaczmański, Ł, and Daroszewski, J. (2015). [Irisin–a new mediator of energy homeostasis]. Postepy Hig. Med. Dosw. 69, 233–242. doi: 10.5604/17322693.1141097
Qin, H., Roberts, K. L., Niyongere, S. A., Cong, Y., Elson, C. O., and Benveniste, E. N. (2007). Molecular mechanism of lipopolysaccharide-induced SOCS-3 gene expression in macrophages and microglia. J. Immunol. 179, 5966–5976. doi: 10.4049/jimmunol.179.9.5966
Raefsky, S. M., and Mattson, M. P. (2017). Adaptive responses of neuronal mitochondria to bioenergetic challenges: roles in neuroplasticity and disease resistance. Free Radic. Biol. Med. 102, 203–216. doi: 10.1016/j.freeradbiomed.2016.11.045
Raschke, S., Elsen, M., Gassenhuber, H., Sommerfeld, M., Schwahn, U., Brockmann, B., et al. (2013). Evidence against a beneficial effect of irisin in humans. PLoS One 8:e73680. doi: 10.1371/journal.pone.0073680
Roca-Rivada, A., Castelao, C., Senin, L. L., Landrove, M. O., Baltar, J., Belén Crujeiras, A., et al. (2013). FNDC5/irisin is not only a myokine but also an adipokine. PLoS One 8:e60563. doi: 10.1371/journal.pone.0060563
Safarpour, P., Daneshi-Maskooni, M., Vafa, M., Nourbakhsh, M., Janani, L., Maddah, M., et al. (2020). Vitamin D supplementation improves SIRT1, Irisin, and glucose indices in overweight or obese type 2 diabetic patients: a double-blind randomized placebo-controlled clinical trial. BMC Fam. Pract. 21:26. doi: 10.1186/s12875-020-1096-3
Santos-Lozano, A., Pareja-Galeano, H., Sanchis-Gomar, F., Quindós-Rubial, M., Fiuza-Luces, C., Cristi-Montero, C., et al. (2016). Physical activity and Alzheimer disease: a protective association. Mayo Clin. Proc. 91, 999–1020. doi: 10.1016/j.mayocp.2016.04.024
Schreiber, S. N., Emter, R., Hock, M. B., Knutti, D., Cardenas, J., Podvinec, M., et al. (2004). The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc. Natl. Acad. Sci. U.S.A. 101, 6472–6477. doi: 10.1073/pnas.0308686101
Schumacher, M. A., Chinnam, N., Ohashi, T., Shah, R. S., and Erickson, H. P. (2013). The structure of irisin reveals a novel intersubunit β-sheet fibronectin type III (FNIII) dimer: implications for receptor activation. J. Biol. Chem. 288, 33738–33744. doi: 10.1074/jbc.M113.516641
Scisciola, L., Fontanella, R. A., Surina Cataldo, V., Paolisso, G., and Barbieri, M. (2021). Sarcopenia and cognitive function: role of myokines in muscle brain cross-talk. Life 11:173. doi: 10.3390/life11020173
Sen, N., and Stress, E. R. (2019). CREB, and memory: a tangled emerging link in disease. Neuroscientist 25, 420–433. doi: 10.1177/1073858418816611
Severinsen, M. C. K., and Pedersen, B. K. (2020). Muscle-organ crosstalk: the emerging roles of myokines. Endocr. Rev. 41, 594–609. doi: 10.1210/endrev/bnaa016
Sharma, C., Kim, S., Nam, Y., Jung, U. J., and Kim, S. R. (2021). Mitochondrial dysfunction as a driver of cognitive impairment in Alzheimer’s disease. Int. J. Mol. Sci. 22:4850. doi: 10.3390/ijms22094850
Sharp, S. I., Aarsland, D., Day, S., Sønnesyn, H., and Ballard, C. (2011). Hypertension is a potential risk factor for vascular dementia: systematic review. Int. J. Geriatr. Psychiatry 26, 661–669. doi: 10.1002/gps.2572
Shen, S., Gao, R., Bei, Y., Li, J., Zhang, H., Zhou, Y., et al. (2017). Serum irisin predicts mortality risk in acute heart failure patients. Cell Physiol. Biochem. 42, 615–622. doi: 10.1159/000477867
Shen, X. N., Niu, L. D., Wang, Y. J., Cao, X. P., Liu, Q., Tan, L., et al. (2019). Inflammatory markers in Alzheimer’s disease and mild cognitive impairment: a meta-analysis and systematic review of 170 studies. J. Neurol. Neurosurg. Psychiatry 90, 590–598. doi: 10.1136/jnnp-2018-319148
Shu, C. J., Benoist, C., and Mathis, D. (2012). The immune system’s involvement in obesity-driven type 2 diabetes. Semin. Immunol. 24, 436–442. doi: 10.1016/j.smim.2012.12.001
Sica, A., and Mantovani, A. (2012). Macrophage plasticity and polarization: in vivo veritas. J. Clin. Invest. 122, 787–795. doi: 10.1172/JCI59643
Siteneski, A., Cunha, M. P., Lieberknecht, V., Pazini, F. L., Gruhn, K., Brocardo, P. S., et al. (2018). Central irisin administration affords antidepressant-like effect and modulates neuroplasticity-related genes in the hippocampus and prefrontal cortex of mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 84, 294–303. doi: 10.1016/j.pnpbp.2018.03.004
Skaper, S. D., Facci, L., Zusso, M., and Giusti, P. (2017). Synaptic plasticity, dementia and Alzheimer disease. CNS Neurol. Disord. Drug Targets 16, 220–233. doi: 10.2174/1871527316666170113120853
Spencer, S. J. (2013). Perinatal nutrition programs neuroimmune function long-term: mechanisms and implications. Front. Neurosci. 7:144. doi: 10.3389/fnins.2013.00144
Tang, H., Yu, R., Liu, S., Huwatibieke, B., Li, Z., and Zhang, W. (2016). Irisin inhibits hepatic cholesterol synthesis via AMPK-SREBP2 signaling. EBioMedicine 6, 139–148. doi: 10.1016/j.ebiom.2016.02.041
Tanila, H. (2017). The role of BDNF in Alzheimer’s disease. Neurobiol. Dis. 97, 114–118. doi: 10.1016/j.nbd.2016.05.008
Trinchieri, G., and Sher, A. (2007). Cooperation of Toll-like receptor signals in innate immune defence. Nat. Rev. Immunol. 7, 179–190. doi: 10.1038/nri2038
Tsivgoulis, G., Alexandrov, A. V., Wadley, V. G., Unverzagt, F. W., Go, R. C., Moy, C. S., et al. (2009). Association of higher diastolic blood pressure levels with cognitive impairment. Neurology 73, 589–595. doi: 10.1212/WNL.0b013e3181b38969
Tu, T., Peng, J., and Jiang, Y. (2020). FNDC5/irisin: a new protagonist in acute brain injury. Stem Cells Dev. 29, 533–543. doi: 10.1089/scd.2019.0232
Varela-Rodríguez, B. M., Pena-Bello, L., Juiz-Valiña, P., Vidal-Bretal, B., Cordido, F., and Sangiao-Alvarellos, S. (2016). FNDC5 expression and circulating irisin levels are modified by diet and hormonal conditions in hypothalamus, adipose tissue and muscle. Sci. Rep. 6:29898. doi: 10.1038/srep29898
Violet, M., Delattre, L., Tardivel, M., Sultan, A., Chauderlier, A., Caillierez, R., et al. (2014). A major role for Tau in neuronal DNA and RNA protection in vivo under physiological and hyperthermic conditions. Front. Cell Neurosci. 8:84. doi: 10.3389/fncel.2014.00084
Wang, F. S., Kuo, C. W., Ko, J. Y., Chen, Y. S., Wang, S. Y., Ke, H. J., et al. (2020). Irisin mitigates oxidative stress, chondrocyte dysfunction and osteoarthritis development through regulating mitochondrial integrity and autophagy. Antioxidants 9:810. doi: 10.3390/antiox9090810
Wang, K., Song, F., Xu, K., Liu, Z., Han, S., Li, F., et al. (2019). Irisin attenuates neuroinflammation and prevents the memory and cognitive deterioration in streptozotocin-induced diabetic mice. Mediat. Inflamm. 2019:1567179. doi: 10.1155/2019/1567179
Wang, S., Liao, J. W., Yin, H. G., Ji, H. H., Yu, H., Zou, Y. H., et al. (2019). [The cardiovascular protection of irisin and its research progress in sports field]. Sheng Li Xue Bao 71, 478–484.
Wang, Z., Chen, K., Han, Y., Zhu, H., Zhou, X., Tan, T., et al. (2018). Irisin protects heart against ischemia-reperfusion injury through a SOD2-dependent mitochondria mechanism. J. Cardiovasc. Pharmacol. 72, 259–269. doi: 10.1097/FJC.0000000000000608
Wang, Z. H., Xiang, J., Liu, X., Yu, S. P., Manfredsson, F. P., Sandoval, I. M., et al. (2019). Deficiency in BDNF/TrkB neurotrophic activity stimulates δ-secretase by upregulating C/EBPβ in Alzheimer’s disease. Cell Rep. 28, 655.e5–669.e5. doi: 10.1016/j.celrep.2019.06.054
Winblad, B., Palmer, K., Kivipelto, M., Jelic, V., Fratiglioni, L., Wahlund, L. O., et al. (2004). Mild cognitive impairment–beyond controversies, towards a consensus: report of the International Working Group on Mild Cognitive Impairment. J. Intern. Med. 256, 240–246. doi: 10.1111/j.1365-2796.2004.01380.x
Wrann, C. D. (2015). FNDC5/irisin - their role in the nervous system and as a mediator for beneficial effects of exercise on the brain. Brain Plast. 1, 55–61. doi: 10.3233/bpl-150019
Wrann, C. D., White, J. P., Salogiannnis, J., Laznik-Bogoslavski, D., Wu, J., Ma, D., et al. (2013). Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell Metab. 18, 649–659. doi: 10.1016/j.cmet.2013.09.008
Wu, F., Li, Z., Cai, M., Xi, Y., Xu, Z., Zhang, Z., et al. (2020). Aerobic exercise alleviates oxidative stress-induced apoptosis in kidneys of myocardial infarction mice by inhibiting ALCAT1 and activating FNDC5/Irisin signaling pathway. Free Radic. Biol. Med. 158, 171–180. doi: 10.1016/j.freeradbiomed.2020.06.038
Xin, C., Zhang, Z., Gao, G., Ding, L., Yang, C., Wang, C., et al. (2020). Irisin attenuates myocardial ischemia/reperfusion injury and improves mitochondrial function through AMPK pathway in diabetic mice. Front. Pharmacol. 11:565160. doi: 10.3389/fphar.2020.565160
Xiong, X. Q., Geng, Z., Zhou, B., Zhang, F., Han, Y., Zhou, Y. B., et al. (2018). FNDC5 attenuates adipose tissue inflammation and insulin resistance via AMPK-mediated macrophage polarization in obesity. Metabolism 83, 31–41. doi: 10.1016/j.metabol.2018.01.013
Yan, M. H., Wang, X., and Zhu, X. (2013). Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 62, 90–101. doi: 10.1016/j.freeradbiomed.2012.11.014
Yang, S., Gu, C., Mandeville, E. T., Dong, Y., Esposito, E., Zhang, Y., et al. (2017). Anesthesia and surgery impair blood-brain barrier and cognitive function in mice. Front. Immunol. 8:902. doi: 10.3389/fimmu.2017.00902
Yang, X. Y., Tse, M. C. L., Hu, X., Jia, W. H., Du, G. H., and Chan, C. B. (2018). Interaction of CREB and PGC-1α induces fibronectin type III domain-containing protein 5 expression in C2C12 myotubes. Cell Physiol. Biochem. 50, 1574–1584. doi: 10.1159/000494655
Yu, Q., Li, G., Ding, Q., Tao, L., Li, J., Sun, L., et al. (2020). Irisin protects brain against ischemia/reperfusion injury through suppressing TLR4/MyD88 pathway. Cerebrovasc. Dis. 49, 346–354. doi: 10.1159/000505961
Zarbakhsh, S., Safari, M., Aldaghi, M. R., Sameni, H. R., Ghahari, L., Khaleghi Lagmouj, Y., et al. (2019). Irisin protects the substantia nigra dopaminergic neurons in the rat model of Parkinson’s disease. Iran. J. Basic Med. Sci. 22, 722–728. doi: 10.22038/ijbms.2019.33444.7987
Zhang, F., Hou, G., Hou, G., Wang, C., Shi, B., and Zheng, Y. (2021). Serum irisin as a potential biomarker for cognitive decline in vascular dementia. Front. Neurol. 12:755046. doi: 10.3389/fneur.2021.755046
Zhang, S., Dong, H., Zhang, X., Li, N., Sun, J., and Qian, Y. (2016). Cerebral mast cells contribute to postoperative cognitive dysfunction by promoting blood brain barrier disruption. Behav. Brain Res. 298, 158–166. doi: 10.1016/j.bbr.2015.11.003
Zhang, W., Chang, L., Zhang, C., Zhang, R., Li, Z., Chai, B., et al. (2015). Central and peripheral irisin differentially regulate blood pressure. Cardiovasc. Drugs Ther. 29, 121–127. doi: 10.1007/s10557-015-6580-y
Zhang, X., Hu, C., Kong, C. Y., Song, P., Wu, H. M., Xu, S. C., et al. (2020). FNDC5 alleviates oxidative stress and cardiomyocyte apoptosis in doxorubicin-induced cardiotoxicity via activating AKT. Cell Death Differ. 27, 540–555. doi: 10.1038/s41418-019-0372-z
Zhao, Y. T., Wang, H., Zhang, S., Du, J., Zhuang, S., and Zhao, T. C. (2016). Irisin ameliorates hypoxia/reoxygenation-induced injury through modulation of histone deacetylase 4. PLoS One 11:e0166182. doi: 10.1371/journal.pone.0166182
Zsuga, J., Tajti, G., Papp, C., Juhasz, B., and Gesztelyi, R. (2016). FNDC5/irisin, a molecular target for boosting reward-related learning and motivation. Med. Hypotheses. 90, 23–28. doi: 10.1016/j.mehy.2016.02.020
Keywords: dementia, irisin, muscle brain cross talk, BDNF, inflammation, oxidative stress
Citation: Peng J and Wu J (2022) Effects of the FNDC5/Irisin on Elderly Dementia and Cognitive Impairment. Front. Aging Neurosci. 14:863901. doi: 10.3389/fnagi.2022.863901
Received: 27 January 2022; Accepted: 01 March 2022;
Published: 31 March 2022.
Edited by:
Ke Zhang, China Medical University, ChinaReviewed by:
Jiumei Cao, Shanghai Jiao Tong University, ChinaCopyright © 2022 Peng and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jinhui Wu, d3VqaW5odWlAc2N1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.