 Yimeng Chen1
Yimeng Chen1 Ying Xia
Ying Xia- 1Department of Urology, The Third Affiliated Hospital of Soochow University, Changzhou, China
- 2Shanghai Key Laboratory of Acupuncture Mechanism and Acupoint Function, Fudan University, Shanghai, China
Hypoxia and ischemia cause inflammatory injury and critically participate in the pathogenesis of various diseases in various organs. However, the protective strategies against hypoxic and ischemic insults are very limited in clinical settings up to date. It is of utmost importance to improve our understanding of hypoxic/ischemic (H/I) inflammation and find novel therapies for better prevention/treatment of H/I injury. Recent studies provide strong evidence that the expression of microRNAs (miRNAs), which regulate gene expression and affect H/I inflammation through post-transcriptional mechanisms, are differentially altered in response to H/I stress, while δ-opioid receptors (DOR) play a protective role against H/I insults in different organs, including both H/I-sensitive organs (e.g., brain, kidney, and heart) and H/I-insensitive organs (e.g., liver and muscle). Indeed, many studies have demonstrated the crucial role of the DOR-mediated cyto-protection against H/I injury by several molecular pathways, including NLRP3 inflammasome modulated by miRNAs. In this review, we summarize our recent studies along with those of others worldwide, and compare the effects of DOR on H/I expression of miRNAs in H/I-sensitive and -insensitive organs. The alternation in miRNA expression profiles upon DOR activation and the potential impact on inflammatory injury in different organs under normoxic and hypoxic conditions are discussed at molecular and cellular levels. More in-depth investigations into this field may provide novel clues for new protective strategies against H/I inflammation in different types of organs.
Introduction
MicroRNAs (miRNAs) are a class of 21∼25-ribonucleotide single-strand non-coding RNA molecules which is endogenously expressed in eukaryote cells (Guo et al., 2014). It has been well recognized that miRNAs play an important role in the post-transcriptional regulation of genes by targeting mRNA molecules. Through base-pairing with the 3′untranslated regions (3′UTRs), miRNAs modulate direct cleavage and/or translational repression of target mRNAs (Bartel, 2009), thus regulating wide spectrum of processes in physiological and pathological conditions. Their regulatory networks are very complex, partially because miRNA expression varies dramatically in different tissues in response to different cell stimulations.
Hypoxic/ischemic injury is a process in which blood flow cessation and oxygen deprivation occur in the body, organs, or cells. Metabolic waste products are accumulated due to the starvation of oxygen and nutrients, leading to profound tissue dysfunction and damage (Zhao et al., 2018). More specifically, hypoxia is a state of low oxygen supply, resulted from a decrease in partial pressure of oxygen, inadequate oxygen transport, or the inability of the tissues/organs to uptake oxygen from the blood. In contrast, ischemia is defined as inadequate blood flow to the tissues, resulting in the deprivation of oxygen and glucose, as well as other substances in the blood (Chao and Xia, 2010; Yang et al., 2015). In this review, the term of “H/I injury” is used to describe the general influences of hypoxic and/or ischemic insult, because hypoxia and ischemia processes often occur one after another. A wide range of pathophysiological processes have been reported to be associated with H/I injury, including ATP depletion, calcium overload, mitochondrial dysfunction, oxidative stress, and etc. (Eltzschig and Eckle, 2011; Chen et al., 2019). More recently, emerging evidence suggests that H/I inflammation, a quick pathological response to oxygen/blood flow depletion, has a major impact on H/I injury through pro-inflammatory cytokines and their signaling pathways (Chen et al., 2020c; Sikora et al., 2021; Troscher et al., 2021).
Because of energy imbalance, H/I insult induces pro-inflammatory events including microglia activation, cytokines production, and immune cell development via various biological processes such as apoptosis and oxidative reaction (Chen et al., 2020c). Different organs may suffer from H/I inflammation in different ways, thus leading to various H/I-related diseases. H/I neuroinflammation induces brain injury in ischemic stroke and post-stroke epilepsy (Li X. et al., 2020; Troscher et al., 2021; Xue Y. et al., 2021), while H/I inflammation leads to other pathophysiological events in peripheral organs, including heart failure, acute kidney injury, and hepatocellular damages (Nangaku and Eckardt, 2007; Klune and Tsung, 2010; Frohlich et al., 2013; Yang et al., 2015). On the other side, some beneficial reaction may go through the regulation of inflammatory events. For example, long term limb remote ischemic conditioning could decrease blood pressure via inflammation regulation (Gao et al., 2021). Studies have demonstrated the vital roles of miRNAs in the regulation of gene expression under H/I condition, not only in H/I-sensitive organs such as kidney, heart, and brain, but also in the H/I-insensitive organs such as liver and muscle (Nallamshetty et al., 2013; Bertero et al., 2017). H/I inflammation is mediated, at least partially, by the miRNA-induced repression of gene expressions. The alternations of miRNA expression in response to H/I injury differ widely in different organs due to their differential sensitivities to the reduction of oxygen and blood flow. Numerous studies have shown the changes of miRNA expression profiles along with the potential targets and inflammatory pathways upon H/I insult in specific organs (Tables 1, 2). However, few studies have compared the common changes and the differences of miRNA profiles between H/I-sensitive and -insensitive organs.
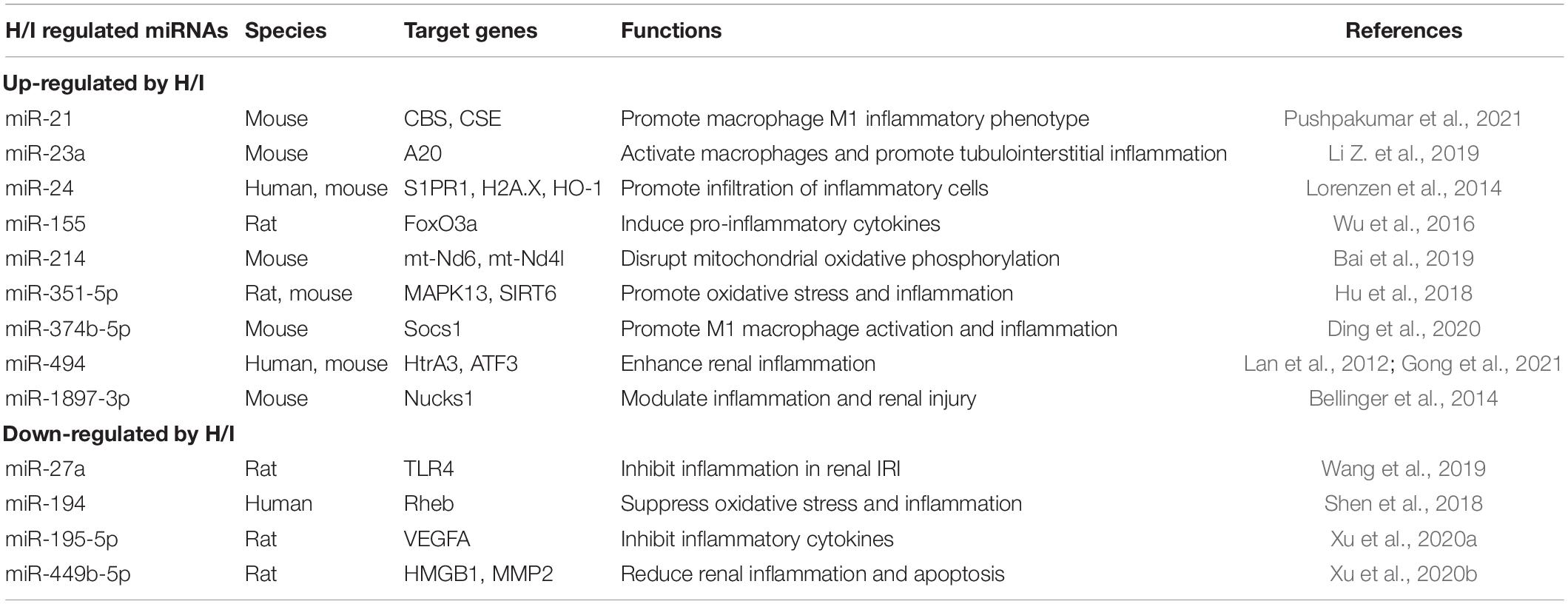
Table 1. H/I-induced changes of miRNAs in the kidney with defined target genes.
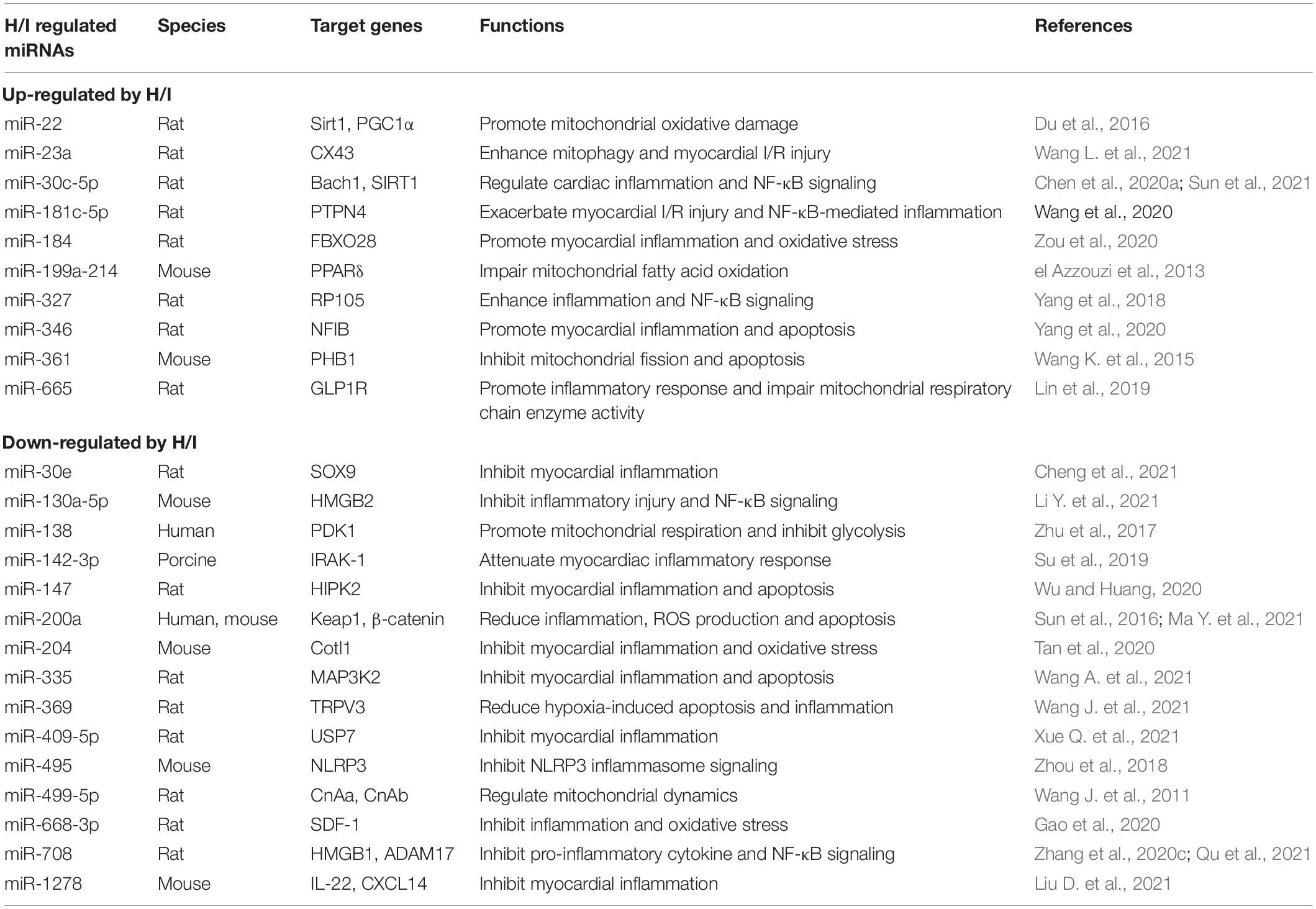
Table 2. H/I-induced changes of miRNAs in the heart with defined target genes.
Opioid receptors belong to the large family of seven-transmembrane G protein-coupled receptors with three major sub-types known as MOR, KOR, and DOR (Xia, 2015). It is well demonstrated that DOR is protective against hypoxic, ischemic, and excitotoxic insults. There exists a differential distribution of DOR in different parts of the brain with a higher density in the cortex, striatum and dorsal root ganglion (Xia, 2015; Tan et al., 2016; Li X. et al., 2020; Gao et al., 2021). DOR is present not only in the nervous system but also in other organs such as the heart, lungs, liver, and gastrointestinal and reproductive tracts (Feng et al., 2012). Our recent studies as well as those of others present strong evidence of the DOR-mediated cyto-protection in different organs, including brain, kidney, heart, and liver (Zhang et al., 2000, 2002, 2006; Ma et al., 2005; Chao et al., 2007, 2008, 2009, 2012; Kang et al., 2009; Chao and Xia, 2010; Feng et al., 2011, 2012; He et al., 2013a; Luo et al., 2019). Moreover, there is accumulating evidence that DOR activation can achieve a protective role against H/I injury by modulating miRNA expression in multiple organs (Yang et al., 2012; He et al., 2013b; Zhi et al., 2016, 2017), especially in the process of neuroinflammation (Chen et al., 2020c). It is therefore possible for DOR signaling, directly or indirectly, to protect organs against H/I inflammation by targeting specific miRNA molecules.
In this review, we highlight some of the recent updates about the effects of H/I and DOR activation on miRNA expression in the current literature, along with our recent work in this field. In particular, we intend to summarize and compare differential regulation of miRNA expression in different organs and discuss potential clinical significances in terms of specific DOR treatment against H/I inflammation.
Effects of Hypoxia/Ischemia on Micrornas in Hypoxia-Sensitive Organs
Kidney
Although blood flow to the kidney accounts for 20% of the cardiac output, the kidney is vulnerable to H/I injury because of the renal vascular anatomy and the high energy consumption of renal tubular epithelial cells (Bracken et al., 2006). H/I-induced inflammatory processes exert a significant role in the development of nephrotic diseases, such as acute kidney injury, glomerulonephritis, and chronic allograft nephropathy (Wang Z. et al., 2020; Nangaku and Eckardt, 2007; Heyman et al., 2008; Chen et al., 2020b). Moreover, the ischemia–reperfusion process during renal transplantation has a profound influence on both short- and long-term recovery outcome of a transplanted kidney (Kosieradzki et al., 2003; Kosieradzki and Rowinski, 2008; Chen et al., 2019). Several studies have suggested that H/I condition, especially a prolonged stress, could strongly influence the miRNA expression profiles in kidneys (Godwin et al., 2010; Shapiro et al., 2011). A number of altered miRNAs have been discovered during H/I kidney injury with identified target genes, which regulate cell inflammatory phenotypes (Table 1). Recent studies in human and rodent models also suggested a crosstalk among different renal cell types after H/I insult. For example, H/I-induced upregulation of miR-21 and miR-374b-5p could promote renal inflammation by activating M1 macrophage (Ding et al., 2020; Pushpakumar et al., 2021). Renal tubular epithelial cells are usually the initial site of renal injury, with activation of fibroblasts or macrophage occurring later by exosome delivery of specific miRNAs (Tan et al., 2016; Li Z. et al., 2019). Pro-inflammatory cytokine production and tubulointerstitial inflammation induced by miRNAs could eventually cause renal fibrosis (Table 1).
Heart
Heart is a H/I-sensitive organ with active metabolism. H/I-induced myocardial injury is closely associated with cardiac disorders such as myocardial infarction and heart failure (Dirksen et al., 2007; Cassavaugh and Lounsbury, 2011; Frohlich et al., 2013). MicroRNAs have been demonstrated to orchestrate many aspects in the development of heart diseases. Recent research on miRNAs regarding myocardial inflammation has drawn much attention from clinicians and scientists. Under H/I condition, the mismatch between energy production and consumption has profound impacts on mitochondrial function and energy metabolism processes of cardiomyocytes (Table 2). Pro-inflammatory miRNAs such as miR-22, miR-199a-214, miR-361, and miR-665 are elevated (el Azzouzi et al., 2013; Wang K. et al., 2015; Du et al., 2016; Lin et al., 2019), whereas anti-inflammatory miRNAs such as miR-138 and miR-499-5p are reduced (Wang J. et al., 2011; Zhu et al., 2017) to impair normal mitochondrial function in myocardial cells. In addition, abnormal expression of miR-30c-5p, miR-181c-5p, miR-327, miR-130a-5p, and miR-708 upon H/I stimulation could regulate myocardial inflammation through NF-κB signaling (Yang et al., 2018; Chen et al., 2020a; Wang S. et al., 2020; Zhang et al., 2020c; Li Y. et al., 2021; Qu et al., 2021; Sun et al., 2021; Table 2). One of the key inflammatory mediators, NLRP3 inflammasome, may also be a direct target of H/I-sensitive miRNAs. A recent study in rodents identified that miR-495 ameliorated cardiac microvascular endothelial cell injury and inflammatory reaction by suppressing the NLRP3 inflammasome signaling pathway (Zhou et al., 2018).
Brain
Hypoxic and/or ischemic injuries are well-documented entities in the pathogenesis of cerebrovascular diseases such as stroke. The effects of H/I on miRNAs in the brain have been widely investigated in patients as well as animal ischemic models with middle cerebral artery occlusion (MCAO). Our previous reviews have summarized the regulation of brain miRNAs in response to hypoxic and ischemic conditions (Yang et al., 2015) and their impact on neuroinflammatory signaling pathways (Chen et al., 2020c). Many studies have elucidated the possible mechanisms and the potential biological processes of H/I-induced miRNA expression changes. The neuroinflammation processes including NLRP3 signaling, mitochondrial impairment, microglia activation, inflammatory cytokines production, and potentially neurodegeneration (Chen et al., 2017, 2020c). In general, up-regulation of pro-inflammatory miRNAs and down-regulation of anti-inflammatory miRNAs are often observed in H/I brains (Table 3). The majority of dysregulated miRNAs in the brain exposed to H/I insult displayed a reduction of miRNA expression, especially after a long-term exposure. Similarly with other H/I-sensitive organs, cerebral miRNAs can directly influence inflammatory cytokines production by modulating target genes. They can also affect NLRP3 inflammasome and NF-κB signaling to modulate neuroinflammation. Moreover, the abnormal expression of cerebral miRNAs, e.g., miR-186-5p, miR-200b, miR-210, miR-449c-5p, miR-302a-3p, miR-424, miR-665-3p, and let-7c-5p, can alter the activation of microglia, the most important inflammatory cell type in the brain, and then indirectly influence neuroinflammation, neuronal death and neurodegeneration (Table 3).
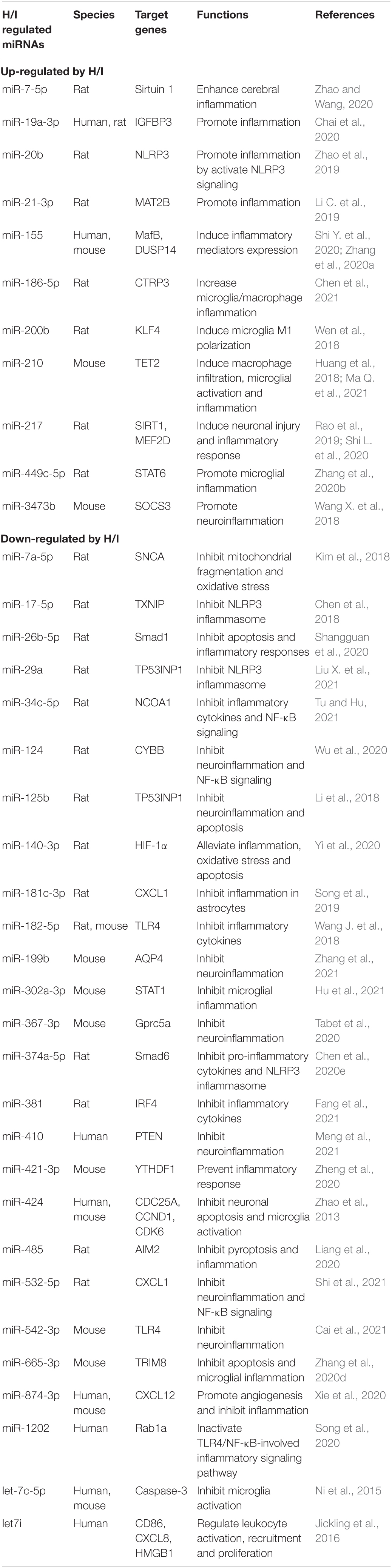
Table 3. H/I-induced changes of miRNAs in the brain with defined target genes.
Effects of Hypoxia/Ischemia on Micrornas in Hypoxia-Insensitive Organs
Liver
Although liver is regarded as one of the hypoxia-insensitive organs, oxygen is also important for liver function maintenance. The insufficiency or deprivation of oxygen and blood flow in hepatic microenvironment due to respiratory/circulatory disorders leads to hepatocellular damage. Indeed, hypoxia activates multiple hypoxia mediators and in turn accelerate or antagonize hepatic damage (Klune and Tsung, 2010; Lu et al., 2016), such as fatty liver disease (Suzuki et al., 2014), hepatocellular carcinoma (HCC) (Liu et al., 2015), and liver-stage malaria (Ng et al., 2014).
Multiple studies have illustrated the H/I injury-induced miRNA changes in liver inflammation, hepatocellular oxidative stress and apoptosis. Meanwhile, altered miRNAs are closely associated with hypoxia-induced HCC pathogenesis by modulating HCC cell angiogenesis, viability and metastasis (Table 4). For example, the hypoxia-sensitive miR-210, which is regarded as the master orchestrator miRNA to H/I insults, is significantly up-regulated in the liver. It mediates hypoxia-induced liver inflammation and HCC cell metastasis through targeting VMP1 (Ying et al., 2011; Song et al., 2014). In addition, hypoxia elevated miR-370 expression and thus promoted inflammation and hepatic histological damage by targeting TGFBR2 (Li et al., 2015). In contrast, cyto-protective miRNAs including miR-24-3p, miR-140-5p, miR-142-3p, miR-146a, and miR-148a were down-regulated under H/I condition, and thus impaired liver function by elevating pro-inflammatory responses (Table 4).
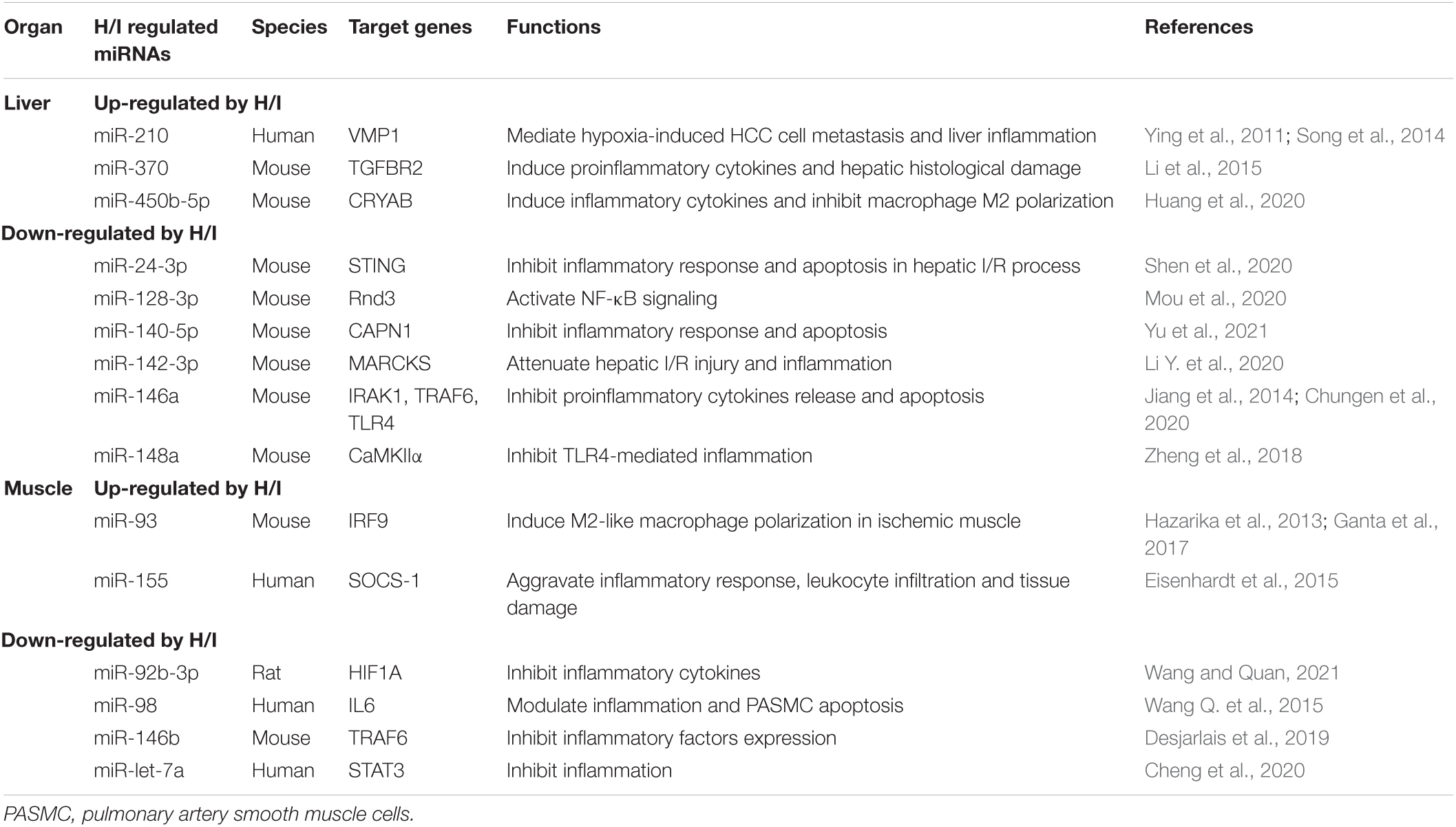
Table 4. H/I-induced changes in miRNAs in H/I-insensitive organs with defined target genes.
Muscle
In comparison to other organs, muscles are relatively tolerant to hypoxic and ischemic stress. Like other H/I-sensitive organs, however, H/I-stimulated muscles also regulate several molecular pathways to better adapt to hypoxic/ischemic environments (Zhu et al., 2022). Recent studies have been devoted to underpinning the mechanisms and pathways of miRNA alternations upon ischemia–reperfusion injury, including macrophage polarization, leukocyte infiltration, and pro-inflammatory cytokine production in ischemic muscle (Table 4). It is reported that miR-93 inhibits IRF9 and induces M2-like polarization in ischemic muscles to enhance angiogenesis, arteriogenesis, and perfusion recovery in peripheral artery disease (Ganta et al., 2017). In in vitro studies, pulmonary artery smooth muscle cells (PASMCs) have been used to investigate H/I-induced muscle miRNA alternations. An H/I down-regulation of miR-98 was observed to modulate inflammation and PASMC apoptosis by directly targeting pro-inflammatory cytokine IL6 (Wang Q. et al., 2015).
The information on H/I muscle miRNAs are still limited at present. Altered miRNAs could significantly affect muscle function and change the status of some muscle diseases by targeting key components in the inflammatory pathways. Therefore, it is needed to further investigate the regulation of muscle miRNAs in H/I condition.
Effects of δ-Opioid Receptor Activation on Microrna Expression Profiles in Different Organs Under Normoxia
δ-Opioid receptor is neuroprotective against H/I injury in the brain (Zhang et al., 2002; Chao et al., 2008, 2009; Kang et al., 2009; He et al., 2013a). The administration of DOR agonists can prolong survival of peripheral organs, such as lung, heart, liver, and kidney (Peart et al., 2005; Patel et al., 2006). As a protective molecule, DOR can tonically regulate miRNA expression even in the normoxic condition. We measured miRNA expression profiles in different organs such as the kidney, heart, brain, and liver after DOR activation with a specific and potent DOR agonist UFP-512 applied in Sprague Dawley rats (Yang et al., 2012; He et al., 2013b; Zhi et al., 2016, 2017). DOR activation can influence the expression of many miRNAs in different organs. As summarized in Table 5, the brain had the most dramatic changes in miRNAs after DOR activation. Conversely, miRNAs in the heart and liver kept relatively stable during the observation (DOR activation for 1, 5, and 10 days), suggesting a differential regulation of miRNAs among organs in response to DOR signals under normoxic condition.

Table 5. DOR-activation induced changes in miRNA expression profiles in normoxic condition.
Among hypoxia-sensitive organs, seven common miRNA changes were observed in the kidney and brain, including miR-21, miR-29b, miR-298, miR-347, miR-351, miR-466b, and miR-511 (Table 5; Yang et al., 2012; He et al., 2013b). MicroRNAs in the kidney displayed the similar expression tendency with those of the brain. For instance, miR-29b was significantly down-regulated in the brain and kidney at 1 day after DOR activation with UFP-512 and maintained at a relatively stable level after 5–10 days DOR activation. Reduced expression level of miR-347 was observed after DOR activation at day 1 in the brain, and at days 5 and 10 in the kidney. Similarly, miR-511 was significantly down-regulated after DOR activation in rat brain and kidney at day 10 (Tables 6, 7). In contrast, there were few common miRNA changes among hypoxia-insensitive organ (liver) and hypoxia-sensitive organs. In our work, only the alternation of miR-107-3p expression was seen in both the heart and liver in normoxic condition upon DOR activation (Table 5). Further investigating their functions and elucidating the mechanistic differences between the H/I sensitive and -insensitive organs will yield valuable information for better understand the DOR-mediated regulation of miRNAs in physiological conditions.
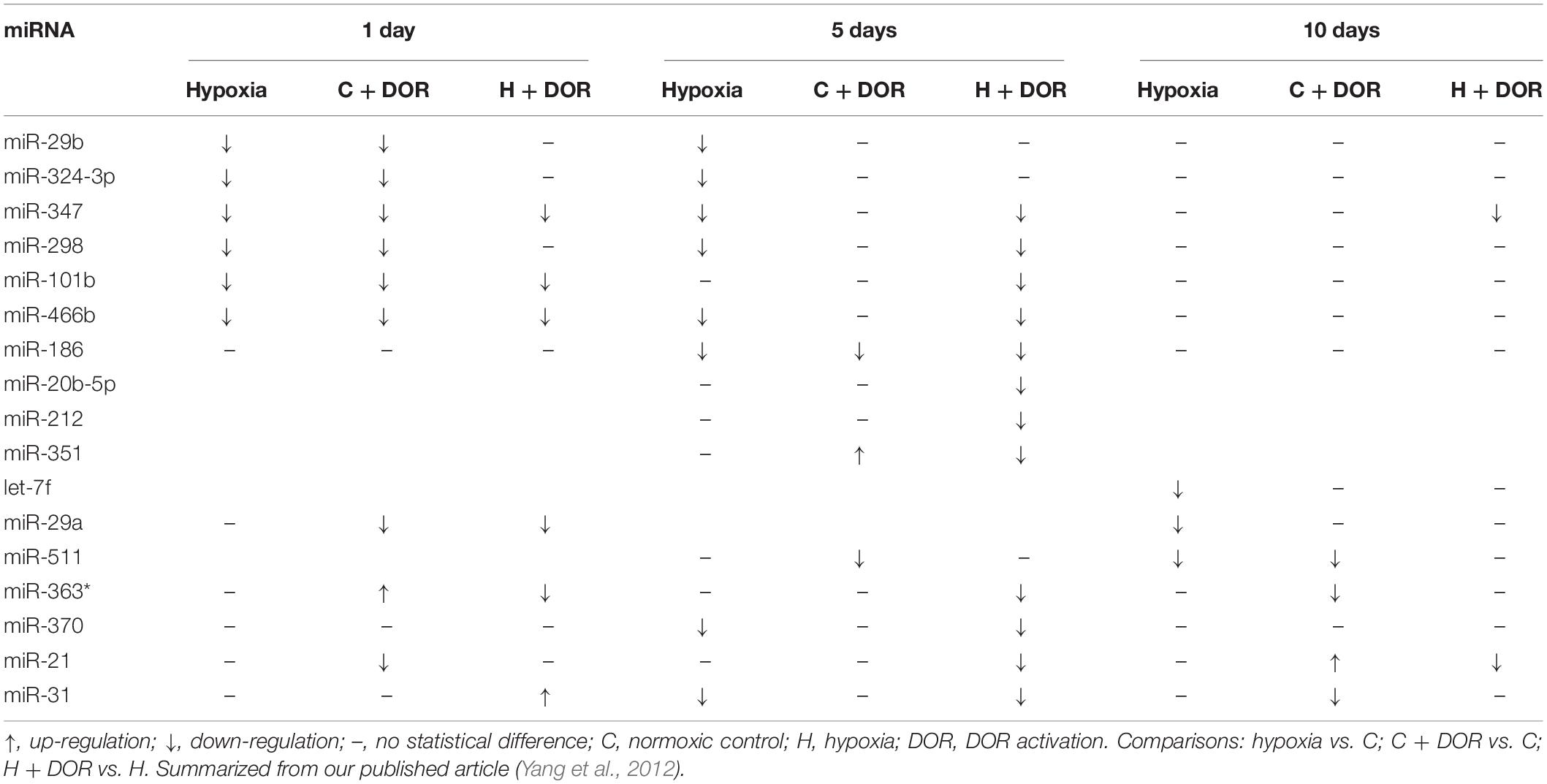
Table 6. Effects of DOR activation on brain miRNAs in prolonged hypoxia.
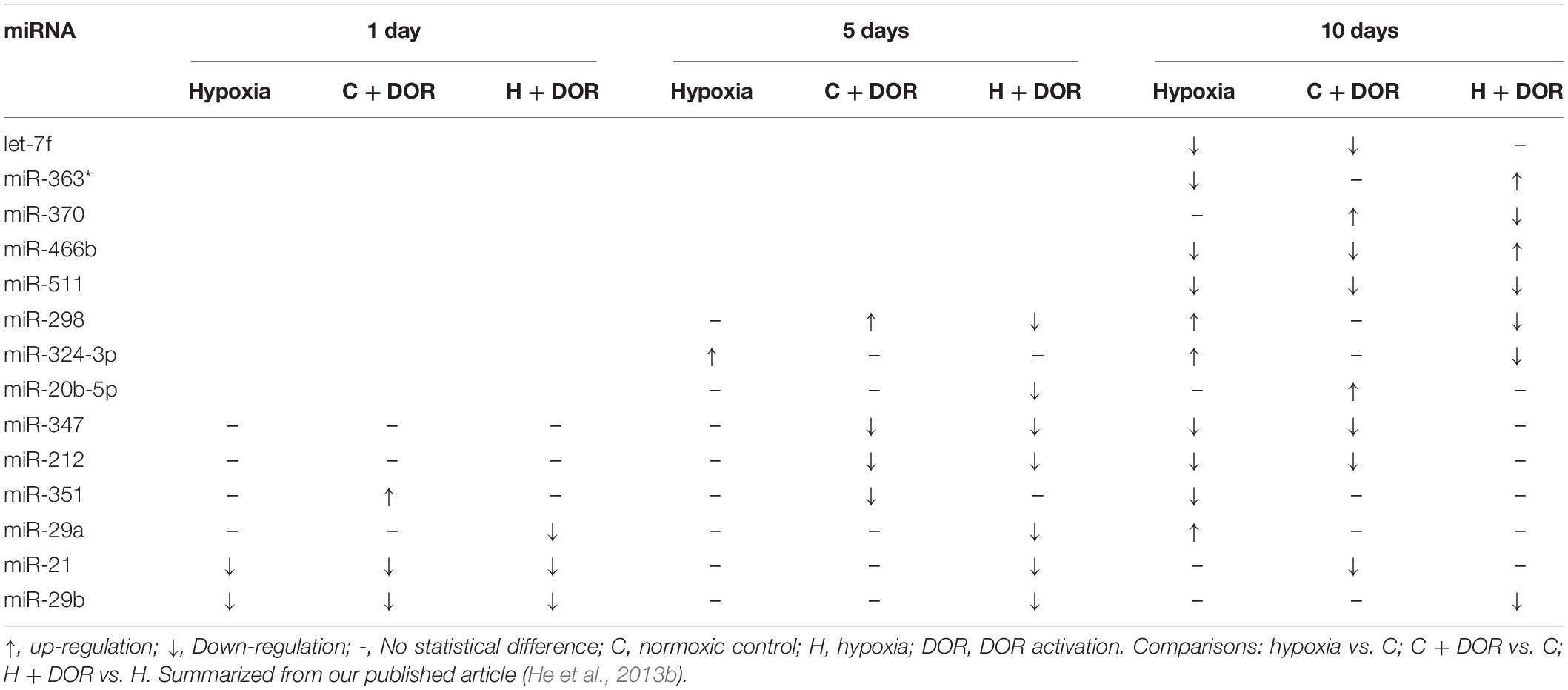
Table 7. Effects of DOR activation on renal miRNAs in prolonged hypoxia.
Effects of δ-Opioid Receptor Activation on Microrna Expression Profiles in Different Organs Under Hypoxia
Since either DOR activation or hypoxic condition has a profound impact on miRNA expression profiles in different organs, it will be interesting to learn the possible effect of DOR activation on hypoxia-induced miRNA changes. Our studies have shown that miRNA expression profiles can be significantly altered when DOR activation was applied to hypoxic organs (Yang et al., 2012; He et al., 2013b; Zhi et al., 2016, 2017). We noticed that the differential alternations in the miRNAs largely depended on the duration of hypoxia, and DOR activation led to diverse outcomes in response to short-term or prolonged hypoxia in different organs.
As one of the hypoxia-sensitive organs, the hypoxic brain showed the most dramatic changes in miRNA expression in response to DOR activation. Some miRNAs were largely altered at the earliest time point (1 day), including miR-347, miR-101b, miR-466b, miR-29a, miR-363*, and miR-31 (Yang et al., 2012). The majority of miRNAs, especially those that mediates cyto-protective function such as anti-neuroinflammation, were down-regulated compared with those in normal condition. Among them, miR-29a alleviated cerebral ischemia/reperfusion injury via down-regulating target gene TP53INP1 and the NF-κB/NLRP3 pathway (Liu X. et al., 2021). In the brain exposed to a mid-term hypoxia (5 days), DOR activation provoked even more miRNA reduction as compared to that in short-term hypoxia (1 day), whereas the expression of miRNAs turned to the baseline level upon DOR activation under prolonged hypoxia (Table 6; Yang et al., 2012). These findings implicated that brain miRNA expression responded quickly after the hypoxia insult, and possibly involved in anti-hypoxic injury by modulating the expression of target genes.
δ-Opioid receptor activation can also modify hypoxia-induced changes of miRNA expression in the kidney. The changes in several miRNAs could be detected upon DOR activation in the kidney exposed to a continuous hypoxia for a prolonged period. For instance, the down-regulation of miR-511 and miR-298 expression was observed upon DOR activation followed by a continuous exposure to hypoxia for 5 to 10 days (He et al., 2013b). In the case of miR-370, miR-20b-5p, and miR-29a/b, although hypoxia alone did not alter their expression levels at certain time points, the administration of UFP-512 to the hypoxic kidney significantly down-regulated the expression of these miRNAs. MiR-21 was reported to induce macrophage M1 inflammatory phenotype, cytokine production, endothelial-mesenchymal transition, and fibrosis in ischemic kidney (Pushpakumar et al., 2021). DOR activation could lower miR-21 expression in prolonged hypoxia (5 days and 10 days). Apparently, DOR activation continuously modulated renal miRNA expression during the whole hypoxic period (Table 7; He et al., 2013b).
Unlike other hypoxia-sensitive organs, the majority of altered miRNAs in the heart was up-regulated under 1 day hypoxia. The administration of UFP-512 to the hypoxic heart induced a further upregulation in terms of the expression of miR-141-3p, miR-376a-3p and miR-134-5p. Moreover, the expression of miR-134-5p and miR-7b was increased throughout the entire time course after DOR activation under hypoxia, suggesting their important roles in the regulation of renal adaptation to hypoxic stress (Table 8; Zhi et al., 2016).
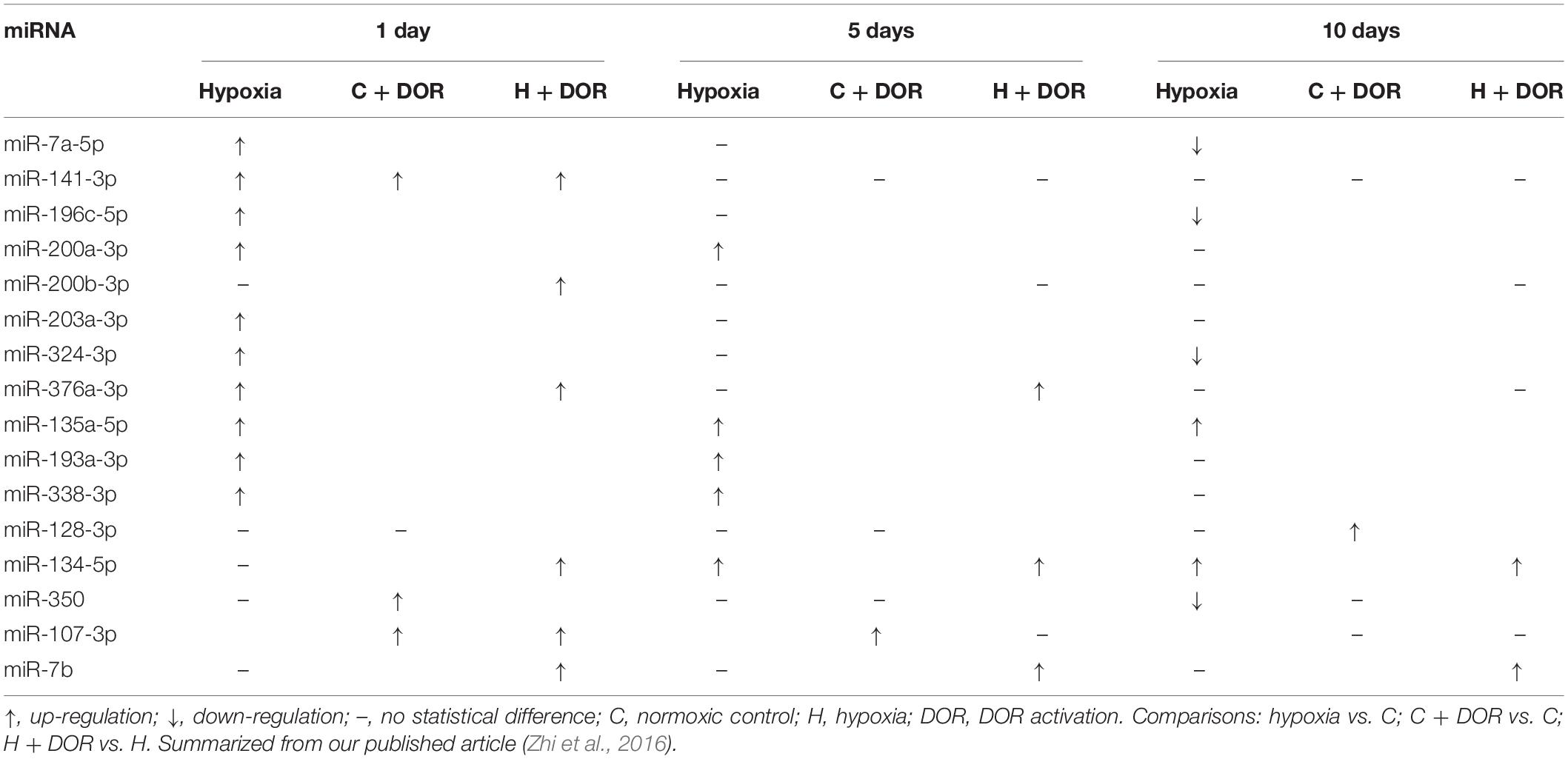
Table 8. Effects of DOR activation on cardiac miRNAs in prolonged hypoxia.
Similar to that in hypoxia-sensitive organs, miRNA expression in hypoxia-insensitive organs can also be influenced by DOR activation in hypoxic condition. The expression of miR-34a-5p, miR-142-5p, miR-145-5p, miR-146a-5p, and miR-204-5p were significantly increased in the liver after DOR activation and hypoxic stress (Zhi et al., 2017). MiR-192-5p was the only miRNA whose expression level started to decrease after DOR activation under hypoxia (Table 9). MiR-146a was reported to ameliorate ischemia/reperfusion injury in vivo and hypoxia/reoxygenation injury in vitro by directly suppressing IRAK1 and TRAF6 in the liver (Jiang et al., 2014). Prolonged hypoxia could down-regulate miR-146a level, whereas DOR activation restored miR-146a expression, which might inhibit pro-inflammatory cytokine release and cellular apoptosis. Therefore, DOR signaling likely functions to upregulate hepatic tolerance to hypoxic stress by differentially modulating the expression of different miRNAs.
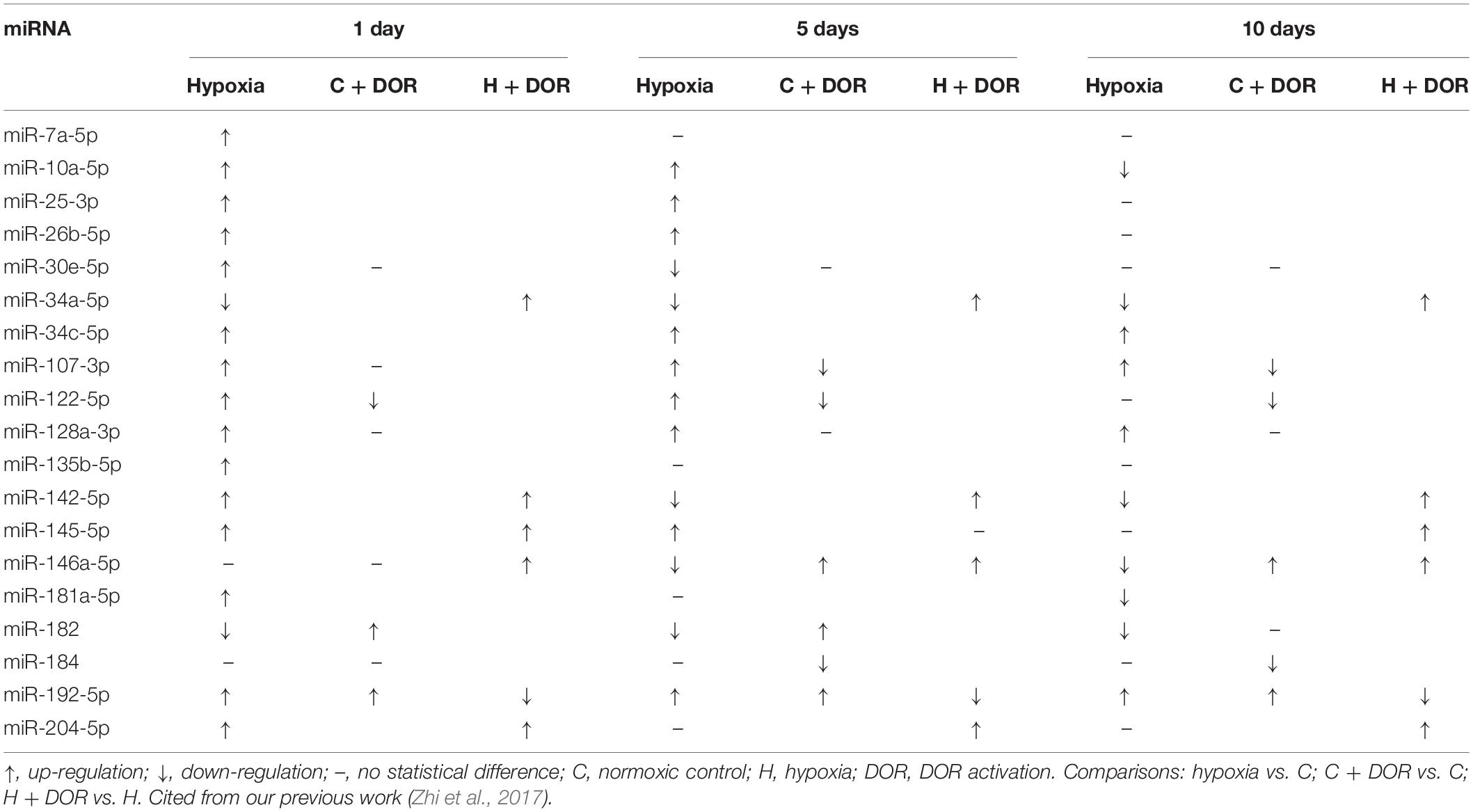
Table 9. Effects of DOR activation on liver miRNAs in prolonged hypoxia.
Collectively, among hypoxia-sensitive organs, the kidney and brain had a common change in the miRNAs, i.e., a significant alternation in nine miRNAs in both organs (Yang et al., 2012; He et al., 2013b), whereas hypoxia-sensitive and -insensitive organs had no common change at all (Table 10).
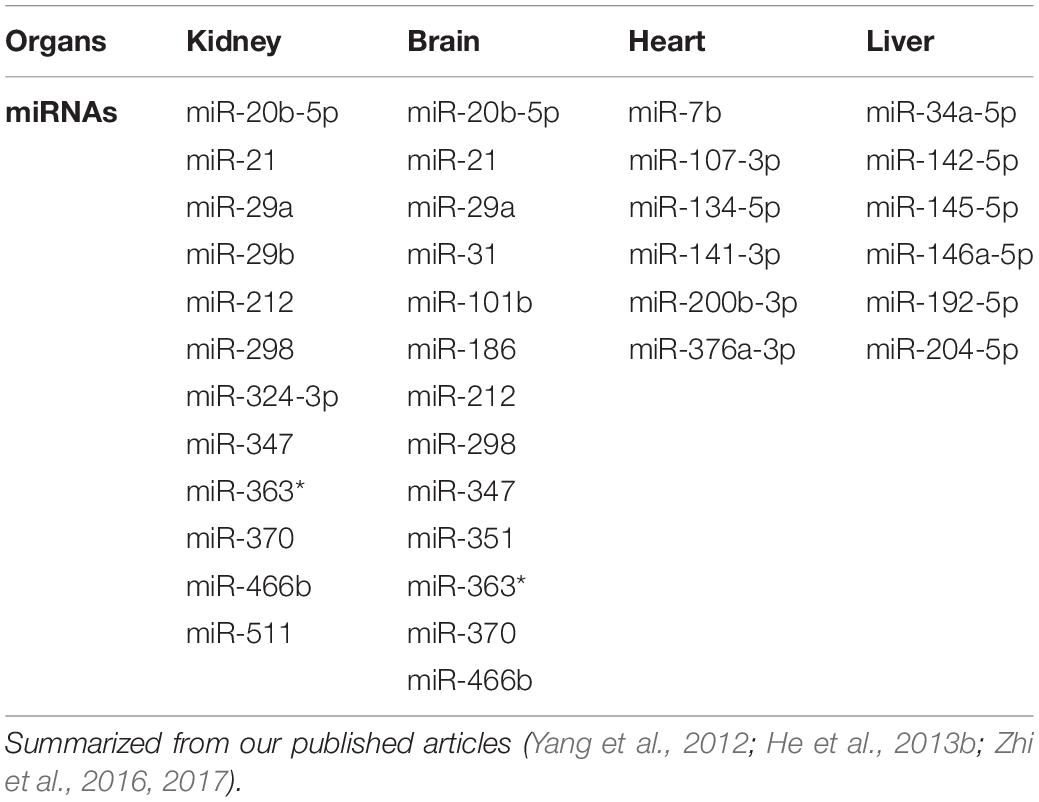
Table 10. DOR-activation modifies hypoxia-induced changes in miRNA expression.
It seems that hypoxia comprehensively modifies miRNA profiles with a major difference among organs, while DOR signaling is able to modulate such regulation in most of these organs (Feng et al., 2012; He et al., 2013a; Nallamshetty et al., 2013; Yang et al., 2015). DOR activation may induce cyto-protection against H/I insult in both hypoxia-sensitive and -insensitive organs, at least partially, through modulating miRNA expression.
Conclusion and Pharmacological Perspectives
Hypoxic/ischemic-induced inflammatory injury to different organs is a frequently encountered clinical problem and the common cause of various diseases with limited therapeutic options. Because the role of miRNAs in controlling H/I inflammation, recent studies on miRNA expression under H/I condition have drawn much attention from clinicians and scientists worldwide. Many experiments have been conducted in cell models, animal models, and patients to investigate the potential targets and signaling pathways of miRNAs involved in H/I pathology. Some of the miRNAs are regarded as injury factors under hypoxic condition by promoting cellular inflammation, mitochondrial dysfunction, oxidative stress and apoptosis, while others play protective roles against H/I insult by inhibiting pro-inflammatory cytokines release and NLRP3 inflammasome (Tables 1–4). Our summary of increased or decreased miRNAs in response to DOR activation (Tables 5, 10) provides a guide for future clarifications of their functions in controlling H/I inflammation.
Hypoxic/ischemic stress comprehensively alters miRNA expression in H/I-sensitive and -insensitive organs, largely depending on the duration of hypoxia. MicroRNA expression in H/I-sensitive organs such as brain and kidney often respond quickly to relatively short-term hypoxia (1 day). After prolonged hypoxia (10 days), the expression of some miRNAs turned back to the baseline level. There is evidence suggesting that a mild/moderate H/I stress may induce pro-inflammatory cytokine release and causes a quick inflammatory response, whereas a severe/prolonged stress eventually causes cell apoptosis and necrosis (Krock et al., 2011; Hadjipanayi and Schilling, 2013). The differential cellular signaling is partially mediated by the miRNA-induced repression of gene expression.
Although various miRNAs can exhibit very different responses even in the same organ under similar H/I condition, there are several H/I sensitive miRNAs that display common changes among different organs. These miRNAs were named “hypoxamiRs,” including miR-21, miR-210, and miR-199a (Nallamshetty et al., 2013; Greco et al., 2014; Azzouzi et al., 2015; Bertero et al., 2017). Accumulating evidence shows that the expression of miR-21 can be directly induced by hypoxia stimulation in several cell types due to the consensus hypoxia-response-element (HRE) sequence in its promoter region (Kulshreshtha et al., 2007; Parikh et al., 2012). Similarly, the expression of miR-210 could be up-regulated by either HIF-dependent mechanism via HRE binding (Lee et al., 2009; Pocock, 2011), or HIF-independent transcriptional regulations (Bertero et al., 2017). One of the most important functions mediated by these “hypoxamiRs” is cellular inflammation (Tables 1–4).
Aging is a key factor affecting H/I inflammation and miRNA expression. In contrast to the immature and young organs, the aged ones are more sensitive to H/I stress. The oxygen-deprived conditions (hypoxia and ischemia) lead to oxidative stress, cellular damage and protein modifications (Adav and Sze, 2020). H/I inflammation and abnormal miRNA expression have been proposed as risk factors for aging and neurodegenerative diseases. Some circulating inflamma-miRs, e.g., miR-21-5p and miR-126-3p, are even thought as potential biomarkers of cognitive impairment AD patients (Giuliani et al., 2021). MicroRNAs implicated in pathological aging such as miR-92a-5p and miR-532-5p are also regarded as potential biomarkers and putative molecular effectors of cognitive frailty (Carini et al., 2021). Serum miR-214 as well as salivary miR-874 and miR-145-3p might serve as auxiliary biomarkers for PD (Chen et al., 2020d; Li L. et al., 2021). In aged rats, transcription factor HIF could protect against ischemic brain injury by reducing inflammatory responses via the Akt signaling pathway (Du et al., 2020). All these facts suggest that an elevated level of inflammation exists in various aging-related chronic diseases, and modulating miRNA expression is a promising avenue for the prevention and treatment of aging and chronic diseases.
The miRNA expression can be modulated by sequence-specific miRNA mimics (or agomirs) and inhibitors (or antagomirs), both in vitro and in vivo. A recent study showed that miR-363-3p treatment attenuates brain ischemia-induced long-term cognitive deficits in rats (Panta et al., 2020), suggesting the potential applications of miRNA mimics. The process of miRNA biogenesis involves transcription, pre-miRNA splicing, exporting, and stability of mature miRNAs. Therefore, the miRNA expression can be modulated by specific molecular signals.
The information in this review suggests that DOR activation is effective in protecting organs against H/I injury, with the capability of modulating miRNAome in both normoxic and hypoxic conditions. Although the direct regulatory mode of DOR on miRNA expression is still unknown, DOR signaling may affect miRNA biogenesis by modulating some key transcriptional factors. For instance, ERK is found to suppress pre-miRNA export from the nucleus to cytoplasm through phosphorylation of Exportin-5, resulting in a global reduction of pre-miRNA loading and miRNA synthesis (Xie et al., 2020; Zhang et al., 2020d). Since ERK activity could be upregulated by DOR activation (Ma et al., 2005; Cai et al., 2021), DOR may regulate miRNA expressions via this signaling pathway.
Different kinds of DOR agonists have been developed in the past. Many of them displayed analgesic, antidepressant, anxiolytic and other opioid effects. For instance, Deltorphin I is an opioid peptide with relatively high affinity and selectivity to DOR and produce centrally mediated analgesic effects in animals (Thomas et al., 1997). Another commonly used one is delta opioid peptide DADLE ([D-Ala2, D-Leu5]-Enkephalin), which was used for pre-conditioning and post-conditioning to induce neuroprotection against hippocampal injury resulted from transient forebrain ischemia in rats (Wang S. et al., 2011). There are also other non-peptide DOR agonists used for antidepressant, anxiolytic or anti-inflammatory properties, including SNC-80 (Bilsky et al., 1995; Perrine et al., 2006), AZD2327 (Hudzik et al., 2014), and ADL5859 (Nozaki et al., 2012). In addition, AR-M 1000390 (Wei et al., 2000) and DPI-3290 (Ananthan, 2006) were also used for DOR agonists. However, UFP-512 that we commonly used in our studies might yield more reliable data for DOR activation because of its specificity and potent binding affinity (Balboni et al., 2002; He et al., 2013b; Xia, 2015). Future development of more specific DOR agonists with lower side-effects may facilitate the application of DOR for the treatment of H/I inflammation.
In summary, it is possible to develop a new protective strategy against H/I injury by activating DOR signaling and targeting certain miRNAs to suppress H/I inflammation in both hypoxia/ischemia-sensitive and -insensitive organs. However, controversies and ambiguity still persist in the literature, especially regarding the up-regulation versus down-regulation of miRNAs under different conditions and their downstream targets in the different organs. Moreover, the molecular mechanisms involved in the DOR-mediated regulation of miRNAs are largely unknown at present. The controversies may be partially attributed to the differences in the models, species, and experimental approaches among different studies. Nevertheless, the solution of the above-mentioned fundamental issues depends on more reliable and in-depth studies in future.
Author Contributions
YX initiated the project and made the outline. YC searched the literature and prepare the manuscript. YH, SZ, and XH participated in writing the manuscript. YX and DX supervised the project and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (81873361), Science and Technology Commission of Shanghai Municipality (18401970100), Natural Science Foundation of Jiangsu Province (BK20200180 and BK20211064), and Changzhou Science and Technology Program (CJ20200089 and CJ20200106).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adav, S. S., and Sze, S. K. (2020). Hypoxia-Induced Degenerative Protein Modifications Associated with Aging and Age-Associated Disorders. Aging Dis. 11, 341–364. doi: 10.14336/AD.2019.0604
Ananthan, S. (2006). Opioid ligands with mixed mu/delta opioid receptor interactions: an emerging approach to novel analgesics. AAPS J. 8, E118–E125. doi: 10.1208/aapsj080114
Azzouzi, H. E., Leptidis, S., Doevendans, P. A., and De Windt, L. J. (2015). HypoxamiRs: regulators of cardiac hypoxia and energy metabolism. Trends Endocrinol. Metab. 26, 502–508. doi: 10.1016/j.tem.2015.06.008
Bai, M., Chen, H., Ding, D., Song, R., Lin, J., Zhang, Y., et al. (2019). MicroRNA-214 promotes chronic kidney disease by disrupting mitochondrial oxidative phosphorylation. Kidney Int. 95, 1389–1404. doi: 10.1016/j.kint.2018.12.028
Balboni, G., Salvadori, S., Guerrini, R., Negri, L., Giannini, E., Jinsmaa, Y., et al. (2002). Potent delta-opioid receptor agonists containing the Dmt-Tic pharmacophore. J. Med. Chem. 45, 5556–5563. doi: 10.1021/jm020336e
Bartel, D. P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136, 215–233. doi: 10.1016/j.cell.2009.01.002
Bellinger, M. A., Bean, J. S., Rader, M. A., Heinz-Taheny, K. M., Nunes, J. S., Haas, J. V., et al. (2014). Concordant changes of plasma and kidney microRNA in the early stages of acute kidney injury: time course in a mouse model of bilateral renal ischemia-reperfusion. PLoS One 9:e93297. doi: 10.1371/journal.pone.0093297
Bertero, T., Rezzonico, R., Pottier, N., and Mari, B. (2017). Impact of MicroRNAs in the Cellular Response to Hypoxia. Int. Rev. Cell Mol. Biol. 333, 91–158. doi: 10.1016/bs.ircmb.2017.03.006
Bilsky, E. J., Calderon, S. N., Wang, T., Bernstein, R. N., Davis, P., Hruby, V. J., et al. (1995). SNC 80, a selective, nonpeptidic and systemically active opioid delta agonist. J. Pharmacol. Exp. Ther. 273, 359–366.
Bracken, C. P., Fedele, A. O., Linke, S., Balrak, W., Lisy, K., Whitelaw, M. L., et al. (2006). Cell-specific regulation of hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha stabilization and transactivation in a graded oxygen environment. J. Biol. Chem. 281, 22575–22585. doi: 10.1074/jbc.M600288200
Cai, G., Cai, G., Zhou, H., Zhuang, Z., Liu, K., Pei, S., et al. (2021). Mesenchymal stem cell-derived exosome miR-542-3p suppresses inflammation and prevents cerebral infarction. Stem Cell Res. Ther. 12:2. doi: 10.1186/s13287-020-02030-w
Carini, G., Musazzi, L., Bolzetta, F., Cester, A., Fiorentini, C., Ieraci, A., et al. (2021). The Potential Role of miRNAs in Cognitive Frailty. Front. Aging Neurosci. 13:763110. doi: 10.3389/fnagi.2021.763110
Cassavaugh, J., and Lounsbury, K. M. (2011). Hypoxia-mediated biological control. J. Cell. Biochem. 112, 735–744. doi: 10.1002/jcb.22956
Chai, Z., Gong, J., Zheng, P., and Zheng, J. (2020). Inhibition of miR-19a-3p decreases cerebral ischemia/reperfusion injury by targeting IGFBP3 in vivo and in vitro. Biol. Res. 53:17. doi: 10.1186/s40659-020-00280-9
Chao, D., Balboni, G., Lazarus, L. H., Salvadori, S., and Xia, Y. (2009). Na+ mechanism of delta-opioid receptor induced protection from anoxic K+ leakage in the cortex. Cell. Mol. Life Sci. 66, 1105–1115. doi: 10.1007/s00018-009-8759-5
Chao, D., Bazzy-Asaad, A., Balboni, G., Salvadori, S., and Xia, Y. (2008). Activation of DOR attenuates anoxic K+ derangement via inhibition of Na+ entry in mouse cortex. Cereb. Cortex 18, 2217–2227. doi: 10.1093/cercor/bhm247
Chao, D., Donnelly, D. F., Feng, Y., Bazzy-Asaad, A., and Xia, Y. (2007). Cortical delta-opioid receptors potentiate K+ homeostasis during anoxia and oxygen-glucose deprivation. J. Cereb. Blood Flow Metab. 27, 356–368. doi: 10.1038/sj.jcbfm.9600352
Chao, D., He, X., Yang, Y., Bazzy-Asaad, A., Lazarus, L. H., Balboni, G., et al. (2012). DOR activation inhibits anoxic/ischemic Na+ influx through Na+ channels via PKC mechanisms in the cortex. Exp. Neurol. 236, 228–239. doi: 10.1016/j.expneurol.2012.05.006
Chao, D., and Xia, Y. (2010). Ionic storm in hypoxic/ischemic stress: can opioid receptors subside it? Prog. Neurobiol. 90, 439–470. doi: 10.1016/j.pneurobio.2009.12.007
Chen, D., Dixon, B. J., Doycheva, D. M., Li, B., Zhang, Y., Hu, Q., et al. (2018). IRE1alpha inhibition decreased TXNIP/NLRP3 inflammasome activation through miR-17-5p after neonatal hypoxic-ischemic brain injury in rats. J. Neuroinflammation 15:32. doi: 10.1186/s12974-018-1077-9
Chen, J., Zhang, M., Zhang, S., Wu, J., and Xue, S. (2020a). Rno-microRNA-30c-5p promotes myocardial ischemia reperfusion injury in rats through activating NF-kappaB pathway and targeting SIRT1. BMC Cardiovasc. Disord. 20:240. doi: 10.1186/s12872-020-01520-2
Chen, Y., Han, X., Sun, Y., He, X., and Xue, D. (2020b). A circulating exosomal microRNA panel as a novel biomarker for monitoring post-transplant renal graft function. J. Cell. Mol. Med. 24, 12154–12163.
Chen, Y., He, X., Wang, S., and Xia, Y. (2020c). delta-Opioid Receptors, microRNAs, and Neuroinflammation in Cerebral Ischemia/Hypoxia. Front. Immunol. 11:421. doi: 10.3389/fimmu.2020.00421
Chen, Y., Zheng, J., Su, L., Chen, F., Zhu, R., Chen, X., et al. (2020d). Increased Salivary microRNAs That Regulate DJ-1 Gene Expression as Potential Markers for Parkinson’s Disease. Front. Aging Neurosci. 12:210. doi: 10.3389/fnagi.2020.00210
Chen, Z., Hu, Y., Lu, R., Ge, M., and Zhang, L. (2020e). MicroRNA-374a-5p inhibits neuroinflammation in neonatal hypoxic-ischemic encephalopathy via regulating NLRP3 inflammasome targeted Smad6. Life Sci. 252:117664. doi: 10.1016/j.lfs.2020.117664
Chen, Y., Gao, C., Sun, Q., Pan, H., Huang, P., Ding, J., et al. (2017). MicroRNA-4639 is a regulator of DJ-1 expression and a potential early diagnostic marker for Parkinson’s disease. Front. Aging Neurosci. 9:232. doi: 10.3389/fnagi.2017.00232
Chen, Y., Liu, W., Chen, M., Sun, Q., Chen, H., and Li, Y. (2021). Up-regulating lncRNA OIP5-AS1 protects neuron injury against cerebral hypoxia-ischemia induced inflammation and oxidative stress in microglia/macrophage through activating CTRP3 via sponging miR-186-5p. Int. Immunopharmacol. 92:107339. doi: 10.1016/j.intimp.2020.107339
Chen, Y., Shi, J., Xia, T. C., Xu, R., He, X., and Xia, Y. (2019). Preservation Solutions for Kidney Transplantation: history, Advances and Mechanisms. Cell Transplant. 28, 1472–1489. doi: 10.1177/0963689719872699
Cheng, G., He, L., and Zhang, Y. (2020). LincRNA-Cox2 promotes pulmonary arterial hypertension by regulating the let-7a-mediated STAT3 signaling pathway. Mol. Cell. Biochem. 475, 239–247. doi: 10.1007/s11010-020-03877-6
Cheng, N., Li, L., Wu, Y., Wang, M., Yang, M., Wei, S., et al. (2021). microRNA-30e up-regulation alleviates myocardial ischemia-reperfusion injury and promotes ventricular remodeling via SOX9 repression. Mol. Immunol. 130, 96–103. doi: 10.1016/j.molimm.2020.11.009
Chungen, Y., Dongfang, Z., and Guoyuan, X. (2020). MicroRNA-146a Protects Against Ischemia/Reperfusion Liver Injury Through Inhibition of Toll-like Receptor 4 Signaling Pathway in Rats. Transplant. Proc. 52, 1007–1013. doi: 10.1016/j.transproceed.2020.01.046
Desjarlais, M., Dussault, S., Rivard, F., Harel, S., Sanchez, V., Hussain, S. N. A., et al. (2019). Forced expression of microRNA-146b reduces TRAF6-dependent inflammation and improves ischemia-induced neovascularization in hypercholesterolemic conditions. Atherosclerosis 289, 73–84. doi: 10.1016/j.atherosclerosis.2019.08.010
Ding, C., Zheng, J., Wang, B., Li, Y., Xiang, H., Dou, M., et al. (2020). Exosomal MicroRNA-374b-5p From Tubular Epithelial Cells Promoted M1 Macrophages Activation and Worsened Renal Ischemia/Reperfusion Injury. Front. Cell Dev. Biol. 8:587693. doi: 10.3389/fcell.2020.587693
Dirksen, M. T., Laarman, G. J., Simoons, M. L., and Duncker, D. J. (2007). Reperfusion injury in humans: a review of clinical trials on reperfusion injury inhibitory strategies. Cardiovasc. Res. 74, 343–355. doi: 10.1016/j.cardiores.2007.01.014
Du, J. K., Cong, B. H., Yu, Q., Wang, H., Wang, L., Wang, C. N., et al. (2016). Upregulation of microRNA-22 contributes to myocardial ischemia-reperfusion injury by interfering with the mitochondrial function. Free Radic. Biol. Med. 96, 406–417. doi: 10.1016/j.freeradbiomed.2016.05.006
Du, X., Yang, J., Liu, C., Wang, S., Zhang, C., Zhao, H., et al. (2020). Hypoxia-Inducible Factor 1alpha and 2alpha Have Beneficial Effects in Remote Ischemic Preconditioning Against Stroke by Modulating Inflammatory Responses in Aged Rats. Front. Aging Neurosci. 12:54. doi: 10.3389/fnagi.2020.00054
Eisenhardt, S. U., Weiss, J. B., Smolka, C., Maxeiner, J., Pankratz, F., Bemtgen, X., et al. (2015). MicroRNA-155 aggravates ischemia-reperfusion injury by modulation of inflammatory cell recruitment and the respiratory oxidative burst. Basic Res. Cardiol. 110:32. doi: 10.1007/s00395-015-0490-9
el Azzouzi, H., Leptidis, S., Dirkx, E., Hoeks, J., van Bree, B., Brand, K., et al. (2013). The hypoxia-inducible microRNA cluster miR-199a approximately 214 targets myocardial PPARdelta and impairs mitochondrial fatty acid oxidation. Cell Metab. 18, 341–354. doi: 10.1016/j.cmet.2013.08.009
Eltzschig, H. K., and Eckle, T. (2011). Ischemia and reperfusion–from mechanism to translation. Nat. Med. 17, 1391–1401. doi: 10.1038/nm.2507
Fang, H., Li, H. F., Yan, J. Y., Yang, M., and Zhang, J. P. (2021). Dexmedetomidine-up-regulated microRNA-381 exerts anti-inflammatory effects in rats with cerebral ischaemic injury via the transcriptional factor IRF4. J. Cell. Mol. Med. 25, 2098–2109. doi: 10.1111/jcmm.16153
Feng, Y., He, X., Yang, Y., Chao, D., Lazarus, L. H., and Xia, Y. (2012). Current research on opioid receptor function. Curr. Drug Targets 13, 230–246.
Feng, Y., He, X., Yang, Y., Chen, J., Yin, K., and Xia, Y. (2011). Effect of delta-opioid receptor over-expression on cortical expression of GABAA receptor alpha1-subunit in hypoxia. Chin. J. Physiol. 54, 118–123.
Frohlich, G. M., Meier, P., White, S. K., Yellon, D. M., and Hausenloy, D. J. (2013). Myocardial reperfusion injury: looking beyond primary PCI. Eur. Heart J. 34, 1714–1722. doi: 10.1093/eurheartj/eht090
Ganta, V. C., Choi, M. H., Kutateladze, A., Fox, T. E., Farber, C. R., and Annex, B. H. (2017). A MicroRNA93-Interferon Regulatory Factor-9-Immunoresponsive Gene-1-Itaconic Acid Pathway Modulates M2-Like Macrophage Polarization to Revascularize Ischemic Muscle. Circulation 135, 2403–2425. doi: 10.1161/CIRCULATIONAHA.116.025490
Gao, Y., Ren, C., Li, X., Yu, W., Li, S., Li, H., et al. (2021). Ischemic Conditioning Ameliorated Hypertension and Vascular Remodeling of Spontaneously Hypertensive Rat via Inflammatory Regulation. Aging Dis. 12, 116–131. doi: 10.14336/AD.2020.0320
Gao, Z., Gao, Q., and Lv, X. (2020). MicroRNA-668-3p Protects Against Oxygen-Glucose Deprivation in a Rat H9c2 Cardiomyocyte Model of Ischemia-Reperfusion Injury by Targeting the Stromal Cell-Derived Factor-1 (SDF-1)/CXCR4 Signaling Pathway. Med. Sci. Monit. 26:e919601. doi: 10.12659/MSM.919601
Giuliani, A., Gaetani, S., Sorgentoni, G., Agarbati, S., Laggetta, M., Matacchione, G., et al. (2021). Circulating Inflamma-miRs as Potential Biomarkers of Cognitive Impairment in Patients Affected by Alzheimer’s Disease. Front. Aging Neurosci. 13:647015. doi: 10.3389/fnagi.2021.647015
Godwin, J. G., Ge, X., Stephan, K., Jurisch, A., Tullius, S. G., and Iacomini, J. (2010). Identification of a microRNA signature of renal ischemia reperfusion injury. Proc. Natl. Acad. Sci. U. S. A. 107, 14339–14344. doi: 10.1073/pnas.0912701107
Gong, Q., Shen, Z. M., Sheng, Z., Jiang, S., and Ge, S. L. (2021). Hsa-miR-494-3p attenuates gene HtrA3 transcription to increase inflammatory response in hypoxia/reoxygenation HK2 Cells. Sci. Rep. 11:1665. doi: 10.1038/s41598-021-81113-x
Greco, S., Gaetano, C., and Martelli, F. (2014). HypoxamiR regulation and function in ischemic cardiovascular diseases. Antioxid. Redox Signal. 21, 1202–1219. doi: 10.1089/ars.2013.5403
Guo, L., Zhao, Y., Yang, S., Zhang, H., and Chen, F. (2014). Integrative analysis of miRNA-mRNA and miRNA-miRNA interactions. Biomed. Res. Int. 2014:907420. doi: 10.1155/2014/907420
Hadjipanayi, E., and Schilling, A. F. (2013). Hypoxia-based strategies for angiogenic induction: the dawn of a new era for ischemia therapy and tissue regeneration. Organogenesis 9, 261–272. doi: 10.4161/org.25970
Hazarika, S., Farber, C. R., Dokun, A. O., Pitsillides, A. N., Wang, T., Lye, R. J., et al. (2013). MicroRNA-93 controls perfusion recovery after hindlimb ischemia by modulating expression of multiple genes in the cell cycle pathway. Circulation 127, 1818–1828. doi: 10.1161/CIRCULATIONAHA.112.000860
He, X., Sandhu, H. K., Yang, Y., Hua, F., Belser, N., Kim, D. H., et al. (2013a). Neuroprotection against hypoxia/ischemia: delta-opioid receptor-mediated cellular/molecular events. Cell. Mol. Life Sci. 70, 2291–2303. doi: 10.1007/s00018-012-1167-2
He, X., Yang, Y., Zhi, F., Moore, M. L., Kang, X., Chao, D., et al. (2013b). delta-Opioid receptor activation modified microRNA expression in the rat kidney under prolonged hypoxia. PLoS One 8:e61080. doi: 10.1371/journal.pone.0061080
Heyman, S. N., Khamaisi, M., Rosen, S., and Rosenberger, C. (2008). Renal parenchymal hypoxia, hypoxia response and the progression of chronic kidney disease. Am. J. Nephrol. 28, 998–1006. doi: 10.1159/000146075
Hu, C., Li, C., Ma, Q., Wang, R., He, Y., Wang, H., et al. (2021). Inhibition of Long Noncoding RNA SNHG15 Ameliorates Hypoxia/Ischemia-Induced Neuronal Damage by Regulating miR-302a-3p/STAT1/NF-kappaB Axis. Yonsei Med. J. 62, 325–337. doi: 10.3349/ymj.2021.62.4.325
Hu, Y., Tao, X., Han, X., Xu, L., Yin, L., Sun, H., et al. (2018). MicroRNA-351-5p aggravates intestinal ischaemia/reperfusion injury through the targeting of MAPK13 and Sirtuin-6. Br. J. Pharmacol. 175, 3594–3609. doi: 10.1111/bph.14428
Huang, L., Ma, Q., Li, Y., Li, B., and Zhang, L. (2018). Inhibition of microRNA-210 suppresses pro-inflammatory response and reduces acute brain injury of ischemic stroke in mice. Exp. Neurol. 300, 41–50. doi: 10.1016/j.expneurol.2017.10.024
Huang, Z., Mou, T., Luo, Y., Pu, X., Pu, J., Wan, L., et al. (2020). Inhibition of miR-450b-5p ameliorates hepatic ischemia/reperfusion injury via targeting CRYAB. Cell Death Dis. 11:455. doi: 10.1038/s41419-020-2648-0
Hudzik, T. J., Pietras, M. R., Caccese, R., Bui, K. H., Yocca, F., Paronis, C. A., et al. (2014). Effects of the delta opioid agonist AZD2327 upon operant behaviors and assessment of its potential for abuse. Pharmacol. Biochem. Behav. 124, 48–57. doi: 10.1016/j.pbb.2014.05.009
Jiang, W., Kong, L., Ni, Q., Lu, Y., Ding, W., Liu, G., et al. (2014). miR-146a ameliorates liver ischemia/reperfusion injury by suppressing IRAK1 and TRAF6. PLoS One 9:e101530. doi: 10.1371/journal.pone.0101530
Jickling, G. C., Ander, B. P., Shroff, N., Orantia, M., Stamova, B., Dykstra-Aiello, C., et al. (2016). Leukocyte response is regulated by microRNA let7i in patients with acute ischemic stroke. Neurology 87, 2198–2205. doi: 10.1212/WNL.0000000000003354
Kang, X., Chao, D., Gu, Q., Ding, G., Wang, Y., Balboni, G., et al. (2009). delta-Opioid receptors protect from anoxic disruption of Na+ homeostasis via Na+ channel regulation. Cell. Mol. Life Sci. 66, 3505–3516. doi: 10.1007/s00018-009-0136-x
Kim, T., Mehta, S. L., Morris-Blanco, K. C., Chokkalla, A. K., Chelluboina, B., Lopez, M., et al. (2018). The microRNA miR-7a-5p ameliorates ischemic brain damage by repressing alpha-synuclein. Sci. Signal. 11:560. doi: 10.1126/scisignal.aat4285
Klune, J. R., and Tsung, A. (2010). Molecular biology of liver ischemia/reperfusion injury: established mechanisms and recent advancements. Surg. Clin. North Am. 90, 665–677. doi: 10.1016/j.suc.2010.04.003
Kosieradzki, M., Kuczynska, J., Piwowarska, J., Wegrowicz-Rebandel, I., Kwiatkowski, A., Lisik, W., et al. (2003). Prognostic significance of free radicals: mediated injury occurring in the kidney donor. Transplantation 75, 1221–1227. doi: 10.1097/01.TP.0000065282.46425.87
Kosieradzki, M., and Rowinski, W. (2008). Ischemia/reperfusion injury in kidney transplantation: mechanisms and prevention. Transplant. Proc. 40, 3279–3288. doi: 10.1016/j.transproceed.2008.10.004
Krock, B. L., Skuli, N., and Simon, M. C. (2011). Hypoxia-induced angiogenesis: good and evil. Genes Cancer 2, 1117–1133. doi: 10.1177/1947601911423654
Kulshreshtha, R., Ferracin, M., Wojcik, S. E., Garzon, R., Alder, H., Agosto-Perez, F. J., et al. (2007). A microRNA signature of hypoxia. Mol. Cell. Biol. 27, 1859–1867. doi: 10.1128/MCB.01395-06
Lan, Y. F., Chen, H. H., Lai, P. F., Cheng, C. F., Huang, Y. T., Lee, Y. C., et al. (2012). MicroRNA-494 reduces ATF3 expression and promotes AKI. J. Am. Soc. Nephrol. 23, 2012–2023. doi: 10.1681/ASN.2012050438
Lee, I., Ajay, S. S., Yook, J. I., Kim, H. S., Hong, S. H., Kim, N. H., et al. (2009). New class of microRNA targets containing simultaneous 5’-UTR and 3’-UTR interaction sites. Genome Res. 19, 1175–1183. doi: 10.1101/gr.089367.108
Li, C., Fei, K., Tian, F., Gao, C., and Yang, S. (2019). Adipose-derived mesenchymal stem cells attenuate ischemic brain injuries in rats by modulating miR-21-3p/MAT2B signaling transduction. Croat. Med. J. 60, 439–448.
Li, Z., Lv, L., Tang, T., Wang, B., Feng, Y., Zhou, L., et al. (2019). HIF-1alpha inducing exosomal microRNA-23a expression mediates the cross-talk between tubular epithelial cells and macrophages in tubulointerstitial inflammation. Kidney Int. 95, 388–404. doi: 10.1016/j.kint.2018.09.013
Li, L., Li, G., Yu, C., Shen, Z., Xu, C., Feng, Z., et al. (2015). A role of microRNA-370 in hepatic ischaemia-reperfusion injury by targeting transforming growth factor-beta receptor II. Liver Int. 35, 1124–1132. doi: 10.1111/liv.12441
Li, L., Ren, J., Pan, C., Li, Y., Xu, J., Dong, H., et al. (2021). Serum miR-214 Serves as a Biomarker for Prodromal Parkinson’s Disease. Front. Aging Neurosci. 13:700959. doi: 10.3389/fnagi.2021.700959
Li, Y., Zhang, H., Li, Z., Yan, X., Li, Y., and Liu, S. (2021). microRNA-130a-5p suppresses myocardial ischemia reperfusion injury by downregulating the HMGB2/NF-kappaB axis. BMC Cardiovasc. Disord. 21:121. doi: 10.1186/s12872-020-01742-4
Li, X., Liao, Y., Dong, Y., Li, S., Wang, F., Wu, R., et al. (2020). Mib2 Deficiency Inhibits Microglial Activation and Alleviates Ischemia-Induced Brain Injury. Aging Dis. 11, 523–535. doi: 10.14336/AD.2019.0807
Li, Y., Gao, M., Xu, L. N., Yin, L. H., Qi, Y., and Peng, J. Y. (2020). MicroRNA-142-3p attenuates hepatic ischemia/reperfusion injury via targeting of myristoylated alanine-rich C-kinase substrate. Pharmacol. Res. 156:104783. doi: 10.1016/j.phrs.2020.104783
Li, X. Q., Yu, Q., Tan, W. F., Zhang, Z. L., and Ma, H. (2018). MicroRNA-125b mimic inhibits ischemia reperfusion-induced neuroinflammation and aberrant p53 apoptotic signalling activation through targeting TP53INP1. Brain Behav. Immun. 74, 154–165. doi: 10.1016/j.bbi.2018.09.002
Liang, J., Wang, Q., Li, J. Q., Guo, T., and Yu, D. (2020). Long non-coding RNA MEG3 promotes cerebral ischemia-reperfusion injury through increasing pyroptosis by targeting miR-485/AIM2 axis. Exp. Neurol. 325:113139. doi: 10.1016/j.expneurol.2019.113139
Lin, B., Feng, D. G., and Xu, J. (2019). microRNA-665 silencing improves cardiac function in rats with heart failure through activation of the cAMP signaling pathway. J. Cell. Physiol. 234, 13169–13181. doi: 10.1002/jcp.27987
Liu, D., Qiao, C., and Luo, H. (2021). MicroRNA-1278 ameliorates the inflammation of cardiomyocytes during myocardial ischemia by targeting both IL-22 and CXCL14. Life Sci. 269:118817. doi: 10.1016/j.lfs.2020.118817
Liu, X., Lv, X., Liu, Z., Zhang, M., and Leng, Y. (2021). MircoRNA-29a in Astrocyte-derived Extracellular Vesicles Suppresses Brain Ischemia Reperfusion Injury via TP53INP1 and the NF-kappaB/NLRP3 Axis. Cell. Mol. Neurobiol. doi: 10.1007/s10571-021-01040-3 [Epub Online ahead of print]
Liu, K., Sun, B., Zhao, X., Wang, X., Li, Y., Qiu, Z., et al. (2015). Hypoxia promotes vasculogenic mimicry formation by the Twist1-Bmi1 connection in hepatocellular carcinoma. Int. J. Mol. Med. 36, 783–791. doi: 10.3892/ijmm.2015.2293
Lorenzen, J. M., Kaucsar, T., Schauerte, C., Schmitt, R., Rong, S., Hubner, A., et al. (2014). MicroRNA-24 antagonism prevents renal ischemia reperfusion injury. J. Am. Soc. Nephrol. 25, 2717–2729. doi: 10.1681/ASN.2013121329
Lu, L., Zhou, H., Ni, M., Wang, X., Busuttil, R., Kupiec-Weglinski, J., et al. (2016). Innate Immune Regulations and Liver Ischemia-Reperfusion Injury. Transplantation 100, 2601–2610. doi: 10.1097/TP.0000000000001411
Luo, F., Xu, R., Song, G., Lu, H., He, X., and Xia, Y. (2019). The delta-Opioid Receptor Differentially Regulates MAPKs and Anti-inflammatory Cytokines in Rat Kidney Epithelial Cells Under Hypoxia. Front. Physiol. 10:1572. doi: 10.3389/fphys.2019.01572
Ma, M. C., Qian, H., Ghassemi, F., Zhao, P., and Xia, Y. (2005). Oxygen-sensitive {delta}-opioid receptor-regulated survival and death signals: novel insights into neuronal preconditioning and protection. J. Biol. Chem. 280, 16208–16218. doi: 10.1074/jbc.M408055200
Ma, Q., Dasgupta, C., Shen, G., Li, Y., and Zhang, L. (2021). MicroRNA-210 downregulates TET2 and contributes to inflammatory response in neonatal hypoxic-ischemic brain injury. J. Neuroinflammation 18:6. doi: 10.1186/s12974-020-02068-w
Ma, Y., Pan, C., Tang, X., Zhang, M., Shi, H., Wang, T., et al. (2021). MicroRNA-200a represses myocardial infarction-related cell death and inflammation by targeting the Keap1/Nrf2 and beta-catenin pathways. Hellenic J. Cardiol. 62, 139–148. doi: 10.1016/j.hjc.2020.10.006
Meng, Q., Yang, P., and Lu, Y. (2021). MicroRNA-410 serves as a candidate biomarker in hypoxic-ischemic encephalopathy newborns and provides neuroprotection in oxygen-glucose deprivation-injured PC12 and SH-SY5Y cells. Brain Behav. 11:e2293. doi: 10.1002/brb3.2293
Mou, T., Luo, Y., Huang, Z., Zheng, D., Pu, X., Shen, A., et al. (2020). Inhibition of microRNA-128-3p alleviates liver ischaemia-reperfusion injury in mice through repressing the Rnd3/NF-kappaB axis. Innate Immun. 26, 528–536. doi: 10.1177/1753425920928449
Nallamshetty, S., Chan, S. Y., and Loscalzo, J. (2013). Hypoxia: a master regulator of microRNA biogenesis and activity. Free Radic. Biol. Med. 64, 20–30. doi: 10.1016/j.freeradbiomed.2013.05.022
Nangaku, M., and Eckardt, K. U. (2007). Hypoxia and the HIF system in kidney disease. J. Mol. Med. 85, 1325–1330. doi: 10.1007/s00109-007-0278-y
Ng, S., March, S., Galstian, A., Hanson, K., Carvalho, T., Mota, M. M., et al. (2014). Hypoxia promotes liver-stage malaria infection in primary human hepatocytes in vitro. Dis. Model Mech. 7, 215–224. doi: 10.1242/dmm.013490
Ni, J., Wang, X., Chen, S., Liu, H., Wang, Y., Xu, X., et al. (2015). MicroRNA let-7c-5p protects against cerebral ischemia injury via mechanisms involving the inhibition of microglia activation. Brain Behav. Immun. 49, 75–85. doi: 10.1016/j.bbi.2015.04.014
Nozaki, C., Le Bourdonnec, B., Reiss, D., Windh, R. T., Little, P. J., Dolle, R. E., et al. (2012). delta-Opioid mechanisms for ADL5747 and ADL5859 effects in mice: analgesia, locomotion, and receptor internalization. J. Pharmacol. Exp. Ther. 342, 799–807. doi: 10.1124/jpet.111.188987
Panta, A., Montgomery, K., Nicolas, M., Mani, K. K., Sampath, D., and Sohrabji, F. (2020). Mir363-3p Treatment Attenuates Long-Term Cognitive Deficits Precipitated by an Ischemic Stroke in Middle-Aged Female Rats. Front. Aging Neurosci. 12:586362. doi: 10.3389/fnagi.2020.586362
Parikh, V. N., Jin, R. C., Rabello, S., Gulbahce, N., White, K., Hale, A., et al. (2012). MicroRNA-21 integrates pathogenic signaling to control pulmonary hypertension: results of a network bioinformatics approach. Circulation 125, 1520–1532. doi: 10.1161/CIRCULATIONAHA.111.060269
Patel, H. H., Head, B. P., Petersen, H. N., Niesman, I. R., Huang, D., Gross, G. J., et al. (2006). Protection of adult rat cardiac myocytes from ischemic cell death: role of caveolar microdomains and delta-opioid receptors. Am. J. Physiol. Heart Circ. Physiol. 291, H344–H350. doi: 10.1152/ajpheart.01100.2005
Peart, J. N., Gross, E. R., and Gross, G. J. (2005). Opioid-induced preconditioning: recent advances and future perspectives. Vascul. Pharmacol. 42, 211–218. doi: 10.1016/j.vph.2005.02.003
Perrine, S. A., Hoshaw, B. A., and Unterwald, E. M. (2006). Delta opioid receptor ligands modulate anxiety-like behaviors in the rat. Br. J. Pharmacol. 147, 864–872. doi: 10.1038/sj.bjp.0706686
Pocock, R. (2011). Invited review: decoding the microRNA response to hypoxia. Pflugers Arch. 461, 307–315. doi: 10.1007/s00424-010-0910-5
Pushpakumar, S., Kundu, S., Weber, G., and Sen, U. (2021). Exogenous hydrogen sulfide and miR-21 antagonism attenuates macrophage-mediated inflammation in ischemia reperfusion injury of the aged kidney. Geroscience 43, 1349–1367. doi: 10.1007/s11357-020-00299-6
Qu, Y., Zhang, J., Zhang, J., and Xiao, W. (2021). MiR-708-3p Alleviates Inflammation and Myocardial Injury After Myocardial Infarction by Suppressing ADAM17 Expression. Inflammation 44, 1083–1095. doi: 10.1007/s10753-020-01404-9
Rao, G., Zhang, W., and Song, S. (2019). MicroRNA217 inhibition relieves cerebral ischemia/reperfusion injury by targeting SIRT1. Mol. Med. Rep. 20, 1221–1229. doi: 10.3892/mmr.2019.10317
Shangguan, Y., Han, J., and Su, H. (2020). GAS5 knockdown ameliorates apoptosis and inflammatory response by modulating miR-26b-5p/Smad1 axis in cerebral ischaemia/reperfusion injury. Behav. Brain Res. 379:112370. doi: 10.1016/j.bbr.2019.112370
Shapiro, M. D., Bagley, J., Latz, J., Godwin, J. G., Ge, X., Tullius, S. G., et al. (2011). MicroRNA expression data reveals a signature of kidney damage following ischemia reperfusion injury. PLoS One 6:e23011. doi: 10.1371/journal.pone.0023011
Shen, A., Zheng, D., Luo, Y., Mou, T., Chen, Q., Huang, Z., et al. (2020). MicroRNA-24-3p alleviates hepatic ischemia and reperfusion injury in mice through the repression of STING signaling. Biochem. Biophys. Res. Commun. 522, 47–52. doi: 10.1016/j.bbrc.2019.10.182
Shen, Y., Zhao, Y., Wang, L., Zhang, W., Liu, C., and Yin, A. (2018). MicroRNA-194 overexpression protects against hypoxia/reperfusion-induced HK-2 cell injury through direct targeting Rheb. J. Cell. Biochem. doi: 10.1002/jcb.28114 [Epub Online ahead of print]
Shi, L., Tian, Z., Fu, Q., Li, H., Zhang, L., Tian, L., et al. (2020). miR-217-regulated MEF2D-HDAC5/ND6 signaling pathway participates in the oxidative stress and inflammatory response after cerebral ischemia. Brain Res. 1739:146835. doi: 10.1016/j.brainres.2020.146835
Shi, Y., Li, K., Xu, K., and Liu, Q. H. (2020). MiR-155-5p accelerates cerebral ischemia-reperfusion injury via targeting DUSP14 by regulating NF-kappaB and MAPKs signaling pathways. Eur. Rev. Med. Pharmacol. Sci. 24, 1408–1419. doi: 10.26355/eurrev_202002_20198
Shi, Y., Yi, Z., Zhao, P., Xu, Y., and Pan, P. (2021). MicroRNA-532-5p protects against cerebral ischemia-reperfusion injury by directly targeting CXCL1. Aging 13, 11528–11541. doi: 10.18632/aging.202846
Sikora, E., Bielak-Zmijewska, A., Dudkowska, M., Krzystyniak, A., Mosieniak, G., Wesierska, M., et al. (2021). Cellular Senescence in Brain Aging. Front. Aging Neurosci. 13:646924. doi: 10.3389/fnagi.2021.646924
Song, G., Jia, H., Xu, H., Liu, W., Zhu, H., Li, S., et al. (2014). Studying the association of microRNA-210 level with chronic hepatitis B progression. J. Viral Hepat. 21, 272–280. doi: 10.1111/jvh.12138
Song, H., Zhang, X., Chen, R., Miao, J., Wang, L., Cui, L., et al. (2019). Cortical Neuron-Derived Exosomal MicroRNA-181c-3p Inhibits Neuroinflammation by Downregulating CXCL1 in Astrocytes of a Rat Model with Ischemic Brain Injury. Neuroimmunomodulation 26, 217–233. doi: 10.1159/000502694
Song, S., Pan, Y., Li, H., and Zhen, H. (2020). MiR-1202 Exerts Neuroprotective Effects on OGD/R Induced Inflammation in HM Cell by Negatively Regulating Rab1a Involved in TLR4/NF-kappaB Signaling Pathway. Neurochem. Res. 45, 1120–1129. doi: 10.1007/s11064-020-02991-7
Su, Q., Lv, X., Ye, Z., Sun, Y., Kong, B., Qin, Z., et al. (2019). The mechanism of miR-142-3p in coronary microembolization-induced myocardiac injury via regulating target gene IRAK-1. Cell Death Dis. 10:61. doi: 10.1038/s41419-019-1341-7
Sun, M., Guo, M., Ma, G., Zhang, N., Pan, F., Fan, X., et al. (2021). MicroRNA-30c-5p protects against myocardial ischemia/reperfusion injury via regulation of Bach1/Nrf2. Toxicol. Appl. Pharmacol. 426:115637. doi: 10.1016/j.taap.2021.115637
Sun, X., Zuo, H., Liu, C., and Yang, Y. (2016). Overexpression of miR-200a protects cardiomyocytes against hypoxia-induced apoptosis by modulating the kelch-like ECH-associated protein 1-nuclear factor erythroid 2-related factor 2 signaling axis. Int. J. Mol. Med. 38, 1303–1311. doi: 10.3892/ijmm.2016.2719
Suzuki, T., Shinjo, S., Arai, T., Kanai, M., and Goda, N. (2014). Hypoxia and fatty liver. World J. Gastroenterol. 20, 15087–15097. doi: 10.3748/wjg.v20.i41.15087
Tabet, F., Lee, S., Zhu, W., Levin, M. G., Toth, C. L., Cuesta Torres, L. F., et al. (2020). microRNA-367-3p regulation of GPRC5A is suppressed in ischemic stroke. J. Cereb. Blood Flow Metab. 40, 1300–1315. doi: 10.1177/0271678X19858637
Tan, D., Chen, X., Bai, T., Zhang, J., and Li, Z. (2020). Sevoflurane up-regulates microRNA-204 to ameliorate myocardial ischemia/reperfusion injury in mice by suppressing Cotl1. Life Sci. 259:118162. doi: 10.1016/j.lfs.2020.118162
Tan, R., Zhou, D., and Liu, Y. (2016). Signaling Crosstalk between Tubular Epithelial Cells and Interstitial Fibroblasts after Kidney Injury. Kidney Dis. 2, 136–144. doi: 10.1159/000446336
Thomas, S. A., Abbruscato, T. J., Hau, V. S., Gillespie, T. J., Zsigo, J., Hruby, V. J., et al. (1997). Structure-activity relationships of a series of [D-Ala2]deltorphin I and II analogues; in vitro blood-brain barrier permeability and stability. J. Pharmacol. Exp. Ther. 281, 817–825.
Troscher, A. R., Gruber, J., Wagner, J. N., Bohm, V., Wahl, A. S., and von Oertzen, T. J. (2021). Inflammation Mediated Epileptogenesis as Possible Mechanism Underlying Ischemic Post-stroke Epilepsy. Front. Aging Neurosci. 13:781174. doi: 10.3389/fnagi.2021.781174
Tu, Y., and Hu, Y. (2021). MiRNA-34c-5p protects against cerebral ischemia/reperfusion injury: involvement of anti-apoptotic and anti-inflammatory activities. Metab. Brain Dis. 36, 1341–1351. doi: 10.1007/s11011-021-00724-5
Wang, A., Dai, L., Yang, L., Wang, Y., Hao, X., Liu, Z., et al. (2021). Upregulation of miR-335 reduces myocardial injury following myocardial infarction via targeting MAP3K2. Eur. Rev. Med. Pharmacol. Sci. 25, 344–352. doi: 10.26355/eurrev_202101_24401
Wang, J., Chen, X., and Huang, W. (2021). MicroRNA-369 attenuates hypoxia-induced cardiomyocyte apoptosis and inflammation via targeting TRPV3. Braz. J. Med. Biol. Res. 54:e10550. doi: 10.1590/1414-431X202010550
Wang, L., Li, Q., Diao, J., Lin, L., and Wei, J. (2021). MiR-23a Is Involved in Myocardial Ischemia/Reperfusion Injury by Directly Targeting CX43 and Regulating Mitophagy. Inflammation 44, 1581–1591. doi: 10.1007/s10753-021-01443-w
Wang, F., and Quan, Q. (2021). Anti-inflammatory role and mechanism of microRNA-92b-3p in the progression of hypoxic pulmonary hypertension. Life Sci. doi: 10.1016/j.lfs.2021.119725 [Epub Online ahead of print]
Wang, J., Jiao, J., Li, Q., Long, B., Wang, K., Liu, J., et al. (2011). miR-499 regulates mitochondrial dynamics by targeting calcineurin and dynamin-related protein-1. Nat. Med. 17, 71–78. doi: 10.1038/nm.2282
Wang, S., Duan, Y., Su, D., Li, W., Tan, J., Yang, D., et al. (2011). Delta opioid peptide [D-Ala2, D-Leu5] enkephalin (DADLE) triggers postconditioning against transient forebrain ischemia. Eur. J. Pharmacol. 658, 140–144. doi: 10.1016/j.ejphar.2011.02.006
Wang, J., Xu, Z., Chen, X., Li, Y., Chen, C., Wang, C., et al. (2018). MicroRNA-182-5p attenuates cerebral ischemia-reperfusion injury by targeting Toll-like receptor 4. Biochem. Biophys. Res. Commun. 505, 677–684. doi: 10.1016/j.bbrc.2018.09.165
Wang, X., Chen, S., Ni, J., Cheng, J., Jia, J., and Zhen, X. (2018). miRNA-3473b contributes to neuroinflammation following cerebral ischemia. Cell Death Dis. 9:11. doi: 10.1038/s41419-017-0014-7
Wang, K., Liu, C., Zhang, X., Feng, C., Zhou, L., Zhao, Y., et al. (2015). miR-361-regulated prohibitin inhibits mitochondrial fission and apoptosis and protects heart from ischemia injury. Cell Death Differ. 22, 1058–1068. doi: 10.1038/cdd.2014.200
Wang, Q., Shu, C., Su, J., and Li, X. (2015). A crosstalk triggered by hypoxia and maintained by MCP-1/miR-98/IL-6/p38 regulatory loop between human aortic smooth muscle cells and macrophages leads to aortic smooth muscle cells apoptosis via Stat1 activation. Int. J. Clin. Exp. Pathol. 8, 2670–2679.
Wang, S., Ge, L., Zhang, D., Wang, L., Liu, H., Ye, X., et al. (2020). MiR-181c-5p Promotes Inflammatory Response during Hypoxia/Reoxygenation Injury by Downregulating Protein Tyrosine Phosphatase Nonreceptor Type 4 in H9C2 Cardiomyocytes. Oxid. Med. Cell. Longev. 2020:7913418. doi: 10.1155/2020/7913418
Wang, Y., Wang, D., and Jin, Z. (2019). miR27a suppresses TLR4induced renal ischemiareperfusion injury. Mol. Med. Rep. 20, 967–976. doi: 10.3892/mmr.2019.10333
Wang, Z., Zhou, C., Sun, Y., Chen, Y., and Xue, D. (2020). Let-7c-5p is involved in chronic kidney disease by targeting TGF-β signaling. Biomed Res. Int. 2020:6960941. doi: 10.1155/2020/6960941
Wei, Z., Brown, W., Takasaki, B., Plobeck, N., Delorme, D., Zhou, F., et al. (2000). N, N-Diethyl-4-(phenylpiperidin-4-ylidenemethyl)benzamide: a novel, exceptionally selective, potent delta opioid receptor agonist with oral bioavailability and its analogues. J. Med. Chem. 43, 3895–3905. doi: 10.1021/jm000229p
Wen, M., Ye, J., Han, Y., Huang, L., Yang, H., Jiang, W., et al. (2018). Hypertonic saline regulates microglial M2 polarization via miR-200b/KLF4 in cerebral edema treatment. Biochem. Biophys. Res. Commun. 499, 345–353. doi: 10.1016/j.bbrc.2018.03.161
Wu, C., and Huang, C. (2020). MicroRNA-147 inhibits myocardial inflammation and apoptosis following myocardial infarction via targeting HIPK2. Eur. Rev. Med. Pharmacol. Sci. 24, 6279–6287. doi: 10.26355/eurrev_202006_21526
Wu, H., Huang, T., Ying, L., Han, C., Li, D., Xu, Y., et al. (2016). MiR-155 is Involved in Renal Ischemia-Reperfusion Injury via Direct Targeting of FoxO3a and Regulating Renal Tubular Cell Pyroptosis. Cell. Physiol. Biochem. 40, 1692–1705. doi: 10.1159/000453218
Wu, Y., Yao, J., and Feng, K. (2020). miR-124-5p/NOX2 Axis Modulates the ROS Production and the Inflammatory Microenvironment to Protect Against the Cerebral I/R Injury. Neurochem. Res. 45, 404–417. doi: 10.1007/s11064-019-02931-0
Xie, K., Cai, Y., Yang, P., Du, F., and Wu, K. (2020). Upregulating microRNA-874-3p inhibits CXCL12 expression to promote angiogenesis and suppress inflammatory response in ischemic stroke. Am. J. Physiol. Cell Physiol. 319, C579–C588. doi: 10.1152/ajpcell.00001.2020
Xu, Y., Jiang, W., Zhong, L., Li, H., Bai, L., Chen, X., et al. (2020a). miR-195-5p alleviates acute kidney injury through repression of inflammation and oxidative stress by targeting vascular endothelial growth factor A. Aging 12, 10235–10245. doi: 10.18632/aging.103160
Xu, Y., Niu, Y., Li, H., and Pan, G. (2020b). Downregulation of lncRNA TUG1 attenuates inflammation and apoptosis of renal tubular epithelial cell induced by ischemia-reperfusion by sponging miR-449b-5p via targeting HMGB1 and MMP2. Inflammation 43, 1362–1374. doi: 10.1007/s10753-020-01214-z
Xue, Q., Yang, D., Zhang, J., Gan, P., Lin, C., Lu, Y., et al. (2021). USP7, negatively regulated by miR-409-5p, aggravates hypoxia-induced cardiomyocyte injury. APMIS 129, 152–162. doi: 10.1111/apm.13100
Xue, Y., Nie, D., Wang, L., Qiu, H., Ma, L., Dong, M., et al. (2021). Microglial Polarization: novel Therapeutic Strategy against Ischemic Stroke. Aging Dis. 12, 466–479. doi: 10.14336/AD.2020.0701
Yang, B., Dong, R., and Zhao, H. (2020). Inhibition of microRNA-346 inhibits myocardial inflammation and apoptosis after myocardial infarction via targeting NFIB. Eur. Rev. Med. Pharmacol. Sci. 24, 11752–11760. doi: 10.26355/eurrev_202011_23827
Yang, Y., Sandhu, H. K., Zhi, F., Hua, F., Wu, M., and Xia, Y. (2015). Effects of hypoxia and ischemia on microRNAs in the brain. Curr. Med. Chem. 22, 1292–1301.
Yang, Y., Yang, J., Liu, X., Ding, J., Li, S., Guo, X., et al. (2018). Down-Regulation of miR-327 Alleviates Ischemia/Reperfusion-Induced Myocardial Damage by Targeting RP105. Cell. Physiol. Biochem. 49, 1049–1063. doi: 10.1159/000493288
Yang, Y., Zhi, F., He, X., Moore, M. L., Kang, X., Chao, D., et al. (2012). delta-opioid receptor activation and microRNA expression of the rat cortex in hypoxia. PLoS One 7:e51524. doi: 10.1371/journal.pone.0051524
Yi, M., Li, Y., Wang, D., Zhang, Q., Yang, L., and Yang, C. (2020). KCNQ1OT1 Exacerbates Ischemia-Reperfusion Injury Through Targeted Inhibition of miR-140-3P. Inflammation 43, 1832–1845. doi: 10.1007/s10753-020-01257-2
Ying, Q., Liang, L., Guo, W., Zha, R., Tian, Q., Huang, S., et al. (2011). Hypoxia-inducible microRNA-210 augments the metastatic potential of tumor cells by targeting vacuole membrane protein 1 in hepatocellular carcinoma. Hepatology 54, 2064–2075. doi: 10.1002/hep.24614
Yu, Q., Chen, S., Tang, H., Yang, H., Zhang, J., Shi, X., et al. (2021). miR1405p alleviates mouse liver ischemia/reperfusion injury by targeting CAPN1. Mol. Med. Rep. 24:675. doi: 10.3892/mmr.2021.12314
Zhang, G., Li, T., Chang, X., and Xing, J. (2021). Long Noncoding RNA SNHG14 Promotes Ischemic Brain Injury via Regulating miR-199b/AQP4 Axis. Neurochem. Res. 46, 1280–1290. doi: 10.1007/s11064-021-03265-6
Zhang, J., Gibney, G. T., Zhao, P., and Xia, Y. (2002). Neuroprotective role of delta-opioid receptors in cortical neurons. Am. J. Physiol. Cell Physiol. 282, C1225–C1234. doi: 10.1152/ajpcell.00226.2001
Zhang, J., Haddad, G. G., and Xia, Y. (2000). delta-, but not mu- and kappa-, opioid receptor activation protects neocortical neurons from glutamate-induced excitotoxic injury. Brain Res. 885, 143–153.
Zhang, J., Qian, H., Zhao, P., Hong, S., and Xia, Y. (2006). Rapid hypoxia preconditioning protects cortical neurons from glutamate toxicity through delta-opioid receptor. Stroke 37, 1094–1099. doi: 10.1161/01.STR.0000206444.29930.18
Zhang, L., Liu, C., Huang, C., Xu, X., and Teng, J. (2020a). miR-155 Knockdown Protects against Cerebral Ischemia and Reperfusion Injury by Targeting MafB. Biomed Res. Int. 2020:6458204. doi: 10.1155/2020/6458204
Zhang, S., Sun, W., Liang, Z., Yin, X., Ji, Z., Chen, X., et al. (2020b). LncRNA SNHG4 Attenuates Inflammatory Responses by Sponging miR-449c-5p and Up-Regulating STAT6 in Microglial During Cerebral Ischemia-Reperfusion Injury. Drug Des. Devel. Ther. 14, 3683–3695. doi: 10.2147/DDDT.S245445
Zhang, S., Wang, Y., Wang, P., and Xuan, J. (2020c). miR-708 affords protective efficacy in anoxia/reoxygenation-stimulated cardiomyocytes by blocking the TLR4 signaling via targeting HMGB1. Mol. Cell. Probes 54:101653. doi: 10.1016/j.mcp.2020.101653
Zhang, X., Feng, Y., Li, J., Zheng, L., Shao, Y., Zhu, F., et al. (2020d). MicroRNA-665-3p attenuates oxygen-glucose deprivation-evoked microglial cell apoptosis and inflammatory response by inhibiting NF-kappaB signaling via targeting TRIM8. Int. Immunopharmacol. 85:106650. doi: 10.1016/j.intimp.2020.106650
Zhao, H., Alam, A., Soo, A. P., George, A. J. T., and Ma, D. (2018). Ischemia-Reperfusion Injury Reduces Long Term Renal Graft Survival: mechanism and Beyond. EBioMedicine 28, 31–42. doi: 10.1016/j.ebiom.2018.01.025
Zhao, H., Wang, J., Gao, L., Wang, R., Liu, X., Gao, Z., et al. (2013). MiRNA-424 protects against permanent focal cerebral ischemia injury in mice involving suppressing microglia activation. Stroke 44, 1706–1713. doi: 10.1161/STROKEAHA.111.000504
Zhao, J., and Wang, B. (2020). MiR-7-5p Enhances Cerebral Ischemia-Reperfusion Injury by Degrading sirt1 mRNA. J. Cardiovasc. Pharmacol. 76, 227–236. doi: 10.1097/FJC.0000000000000852
Zhao, J., Wang, H., Dong, L., Sun, S., and Li, L. (2019). miRNA-20b inhibits cerebral ischemia-induced inflammation through targeting NLRP3. Int. J. Mol. Med. 43, 1167–1178. doi: 10.3892/ijmm.2018.4043
Zheng, D., Li, Z., Wei, X., Liu, R., Shen, A., He, D., et al. (2018). Role of miR-148a in Mitigating Hepatic Ischemia-Reperfusion Injury by Repressing the TLR4 Signaling Pathway via Targeting CaMKIIalpha in Vivo and in Vitro. Cell. Physiol. Biochem. 49, 2060–2072. doi: 10.1159/000493716
Zheng, L., Tang, X., Lu, M., Sun, S., Xie, S., Cai, J., et al. (2020). microRNA-421-3p prevents inflammatory response in cerebral ischemia/reperfusion injury through targeting m6A Reader YTHDF1 to inhibit p65 mRNA translation. Int. Immunopharmacol. 88:106937. doi: 10.1016/j.intimp.2020.106937
Zhi, F., Shao, N., Xue, L., Xu, Y., Kang, X., Yang, Y., et al. (2017). Characteristic MicroRNA Expression Induced by delta-Opioid Receptor Activation in the Rat Liver Under Prolonged Hypoxia. Cell. Physiol. Biochem. 44, 2296–2309. doi: 10.1159/000486067
Zhi, F., Xue, L., Shao, N., Deng, D., Kang, X., Chao, D., et al. (2016). delta-Opioid Receptor Activation and MicroRNA Expression in the Rat Heart Under Prolonged Hypoxia. Cell. Physiol. Biochem. 39, 1118–1128. doi: 10.1159/000447815
Zhou, T., Xiang, D., Li, S., Yang, L., Gao, L., and Feng, C. (2018). MicroRNA-495 Ameliorates Cardiac Microvascular Endothelial Cell Injury and Inflammatory Reaction by Suppressing the NLRP3 Inflammasome Signaling Pathway. Cell. Physiol. Biochem. 49, 798–815. doi: 10.1159/000493042
Zhu, H., Xue, H., Jin, Q., Guo, J., and Chen, Y. (2017). MiR-138 protects cardiac cells against hypoxia through modulation of glucose metabolism by targetting pyruvate dehydrogenase kinase 1. Biosci. Rep. 37:BSR20170296. doi: 10.1042/BSR20170296
Zhu, P., Hamlish, N. X., Thakkar, A. V., Steffeck, A. W. T., Rendleman, E. J., Khan, N. H., et al. (2022). BMAL1 drives muscle repair through control of hypoxic NAD(+) regeneration in satellite cells. Genes Dev. 36, 149–166. doi: 10.1101/gad.349066.121
Keywords: MicroRNAs, hypoxic/ischemic inflammation, δ-opioid receptor (DOR), organs’ differential responses, NLRP3 inflammasome
Citation: Chen Y, He Y, Zhao S, He X, Xue D and Xia Y (2022) Hypoxic/Ischemic Inflammation, MicroRNAs and δ-Opioid Receptors: Hypoxia/Ischemia-Sensitive Versus-Insensitive Organs. Front. Aging Neurosci. 14:847374. doi: 10.3389/fnagi.2022.847374
Received: 02 January 2022; Accepted: 21 March 2022;
Published: 09 May 2022.
Edited by:
Yu-Min Kuo, National Cheng Kung University, TaiwanReviewed by:
Rongfang Que, Southern Medical University, ChinaZhouteng Tao, Shanghai Institute of Materia Medica (CAS), China
Copyright © 2022 Chen, He, Zhao, He, Xue and Xia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dong Xue, eHVlZG9uZ2R4QDE2My5jb20=; Ying Xia, eTU1NzM4MDg4QGdtYWlsLmNvbQ==