 Mai A. Shafei
Mai A. Shafei Matthew Harris
Matthew Harris Myra E. Conway
Myra E. Conway
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Aging Neurosci. , 02 June 2017
Sec. Alzheimer's Disease and Related Dementias
Volume 9 - 2017 | https://doi.org/10.3389/fnagi.2017.00173
This article is part of the Research Topic Dysregulation of Protein Folding in Neurodegenerative Disorders View all 12 articles
Alzheimer’s disease (AD) is a progressive disease associated with the production and deposition of amyloid β-peptide (Aβ) aggregates and neurofibrillary tangles, which lead to synaptic and neuronal damage. Reduced autophagic flux has been widely associated with the accumulation of autophagic vacuoles (AV), which has been proposed to contribute to aggregate build-up observed in AD. As such, targeting autophagy regulation has received wide review, where an understanding as to how this mechanism can be controlled will be important to neuronal health. The mammalian target of rapamycin complex 1 (mTORC1), which was found to be hyperactive in AD brain, regulates autophagy and is considered to be mechanistically important to aberrant autophagy in AD. Hormones and nutrients such as insulin and leucine, respectively, positively regulate mTORC1 activation and are largely considered to inhibit autophagy. However, in AD brain there is a dysregulation of nutrient metabolism, linked to insulin resistance, where a role for insulin treatment to improve cognition has been proposed. Recent studies have highlighted that mitochondrial proteins such as glutamate dehydrogenase and the human branched chain aminotransferase protein, through metabolism of leucine and glutamate, differentially regulate mTORC1 and autophagy. As the levels of the hBCAT proteins are significantly increased in AD brain relative to aged-matched controls, we discuss how these metabolic pathways offer new potential therapeutic targets. In this review article, we highlight the core regulation of autophagy through mTORC1, focusing on how insulin and leucine will be important to consider in particular with respect to our understanding of nutrient load and AD pathogenesis.
Alzheimer’s disease (AD), similar to other neurodegenerative diseases, is characterized by the accumulation of protein aggregates, namely amyloid β-peptide (Aβ) and Tau tangles, which lead to synaptic and neuronal damage, particularly in the hippocampal and the inferior parietal lobule (IPL) regions of the brain, resulting in memory loss (Braak and Braak, 1991; Borlikova et al., 2013). Autophagy, of which there are three types, microautophagy, chaperone-mediated autophagy and macroautophagy, is important for aggregate clearance and is considered to be dysregulated in neurodegenerative conditions, such as AD. Macroautophagy (referred to as autophagy in this review article) is the major degradation pathway in which constitutive autophagy clears functionally redundant or damaged intracellular structures whilst induced autophagy is initiated in response to environmental factors such as nutrient starvation and oxidative stress, generating recycled amino acids, lipids and other nutrients (Heras-Sandoval et al., 2014). Initially, organelles and proteins to be degraded are surrounded by an isolation membrane (phagophore) which fuses together to form a double membrane vesicle (autophagosomes; Dunn, 1990; Klionsky and Ohsumi, 1999; Figure 1A). Hydrolytic enzymes are acquired by merging of the autophagosome with acidified lysosomes (autolysosome; Appelqvist et al., 2013). Autophagosomes are then trafficked in a dynein-dependent retrograde manner along microtubules to lysosomes, which fuse to form the autolysosome (Seaman, 2012; Small and Petsko, 2015). Maturation or late-stage autophagy is fundamental to clearance and inhibition of maturation, fusion or lysosomal function can interfere with autophagic flux (Yang and Klionsky, 2010). The autophagy pathway is regulated by several signaling cascades; in particular the mammalian target of rapamycin (mTOR) pathway, which controls the initiation stage of autophagy and negatively regulates the biogenesis of lysosomes. Here, we review the role of autophagy in AD and discuss how our current understanding of nutrient load and insulin regulation are involved in its dysregulation through the mTOR pathway.
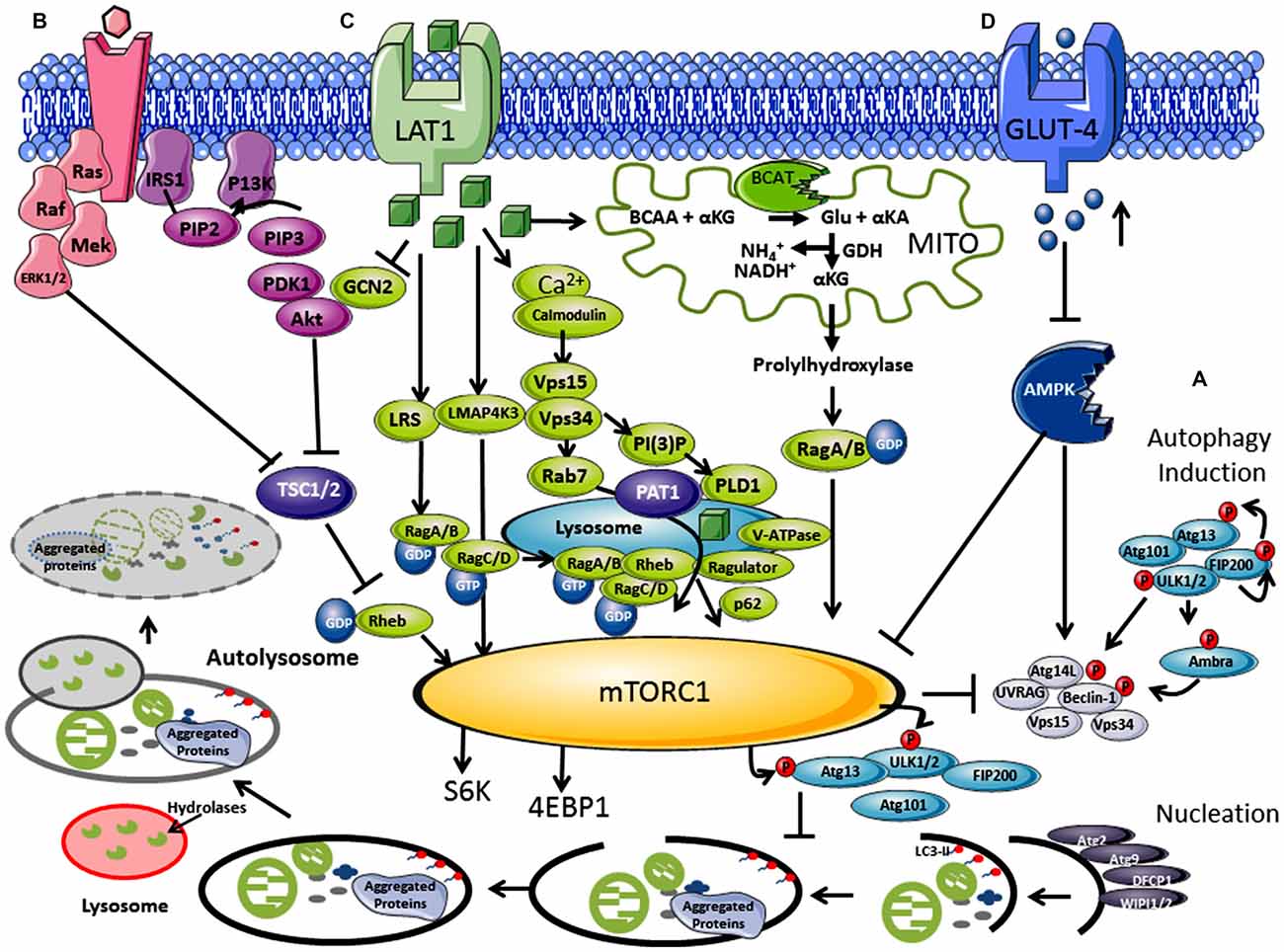
Figure 1. Regulation of autophagy by nutrients and hormones via the mammalian target of rapamycin (mTOR) signaling pathway. (A) Autophagosome induction begins with the activation of the ULK1/2 protein kinase complex that includes the autophagy related proteins (Atgs, where Atg13 is indispensable) and the 200 kDa focal adhesion kinase family-interacting protein (FIP200). This is followed by a nucleation step that is dependent on the class III phosphatidylinositol 3-kinase or human vacuolar protein sorting 34 (hVps34), complexed with BCL-2 interacting moesin-like coiled-coil protein 1 (Beclin 1) and Vps15. Phosphorylation of phosphatidylinositol (PtdIns) by hVps34 signals the recruitment of other autophagy proteins required for elongation. Beclin 1 is involved in two recruitment complexes, the Atg14L and UV radiation resistance-associated gene (UVRAG) complex, which are required for phagophore formation and phagocytosis, respectively. Elongation and closure of the autophagosome requires several Atg proteins, hVps34 and microtubule-associated light chain 3 phosphatidylethanolamine (LC3), which is regulated by GTPase Rab5. The outer membrane of the autophagosome then fuses with a lysosome, exposing the inner single membrane to lysosomal hydrolases whereby the contents are degraded. (B) Hormones and growth factors such as insulin and insulin-like growth factor (IGF-1) trigger mTORC1 activity through a cascade of events resulting in recruitment and activation of Akt which induces phosphorylation and degradation of tuberous sclerosis complex protein 2 (TSC2). TSC2 degradation permits GTP-bound Rheb to directly interact and activate mTORC1. The Ras-ERK pathway also activates mTORC1 through inhibitions of TSC1 and TSC2. (C) Amino acids, in particular leucine, regulate the mTORC1 through the Rag complex (Ras-related GTPase), which recruits mTOR to the lysosomes with Rheb and hVps34. (D) Inhibition of the amp-activated protein kinase (AMPK) signaling pathway is triggered by high glucose levels, resulting in the decrease of AMP:ATP ratio that inhibits TSC1 and TSC2, activating mTORC1 and inhibiting autophagy.
In post-mortem AD brain, autophagic vacuoles (AV) were found to have accumulated and the number of dystrophic neurites containing these AV were considerably greater relative to matched controls (Cataldo et al., 1997; Nixon et al., 2005; Nixon, 2007). High levels of Aβ and γ-secretase subunits found in AVs indicated that amyloid precursor protein (APP) processing can occur, where impaired clearance could contribute to elevated Aβ levels in the brain (Yu et al., 2005). Several studies have suggested that Aβ deposition occurs later in the disease process (Yang et al., 1998; Cataldo et al., 2000). As autophagosome imbalance is thought to occur as an early event in the pathogenesis of AD, dysregulation of this pathway may be upstream of aggregate accumulation (Perez et al., 2015). Autophagy is a multistep process (Figure 1), where a dysfunction in the formation or clearance of the autophagosome or its regulation could result in aggregate accumulation. What is clear is that the autophagy related proteins (Atg) are fundamentally important as knockout of Atg7 in mice results in neurodegenerative disease, with accumulation of ubiquitinated protein aggregates (Komatsu et al., 2005). Several other aspects of the pathway are also vulnerable and are thus seen as potential therapeutic targets. Proteins important for elongation and closure including LC3 and Beclin 1, were found to be downregulated in the IPL of AD tissue, a deficiency of which would compromise autophagosome formation (Pickford et al., 2008; Rohn et al., 2011). In APP transgenic mice models, depletion of Beclin 1 resulted in the accumulation of intracellular and extracellular Aβ, highlighting the importance of early autophagosome formation in Aβ clearance (Pickford et al., 2008). There are also indicators that end-stage processing at the autophagic/lysosomal stage is disrupted, where in AD brain, the lysosomal protease cathepsin D (intracellular aspartyl protease) was found to be upregulated (Cataldo et al., 1995). Cathepsins have β and γ secretase activity, are capable of cleaving APP, and if inhibited or deleted result in a build-up of Aβ (Mueller-Steiner et al., 2006) and tau aggregates (Hamano et al., 2008). The final stage of vesicular trafficking has also been shown to be perturbed, resulting in inefficient clearance of AVs, reducing autophagic flux (for review see Small and Petsko, 2015). Therefore it is clear that autophagy dysregulation (at all stages) has been implicated in aggregate accumulation or ineffective clearance. However, what is not clear are the mechanistic details underpinning or regulating these alterations in autophagy or how specifically it results in Aβ and tau aggreation and more so if the process begins upstream of autophagy.
The mTOR pathway acts as an environmental sensor, which positively regulates protein synthesis and represses autophagy. mTOR forms complexes with several different core proteins, collectively described as mTORC1 and mTORC2 (not a direct autophagy regulator; Tan and Miyamoto, 2016). Knowledge of external stimuli that regulate the P13K/Akt/mTORC1 axis is important as active mTORC1 plays a role in neuronal synaptic plasticity and in neuronal survival during embryonic development (Morita et al., 2015). Hormones and growth factors such as insulin, insulin-like growth factor (IGF-1) and epidermal growth factor trigger mTORC1 activity through a cascade of events that begins with the receptor-mediated activation of phosphatidylinositol 3-kinase-related kinase protein (P13K) through phosphorylation of the insulin receptor substrate (IRS1 and IRS2; Figure 1B; Um et al., 2006). mTORC1 rather than mTORC2 is regulated by nutrients such as amino acids (in particular but not exclusively, leucine) and glucose (Figure 1C; Gulati et al., 2008). Ultimately activation of the eukaryotic initiation factor 4E (eIF4E), its repressor eIF4E binding protein (4E-BP1), and p70S6K results in increased protein translation and synthesis, but lipid and nucleotide synthesis are also regulated (Goberdhan et al., 2016). Under fed conditions, mTORC1 regulates autophagy through phosphorylation at Ser757 of the ULK1 complex and blocks its interaction with 5′ AMP-activated protein kinase (AMPK), preventing autophagosome initiation (Long and Zierath, 2006). However, reports of inhibition at the maturation step through phosphorylation of UVRAG, extends its influence at several stages of autophagy (Liang et al., 2008). Conversely, low glucose, depletion of amino acids and oxidative stress are all key negative regulators of mTORC1 but stimulators of autophagy, where a balance between protein synthesis and clearance maintains cellular homeostasis (Wang et al., 1998). Sustained activation of p70S6K however, also phosphorylates IRS1 at inhibitory sites, negatively regulating Akt and stimulating autophagy (Shah et al., 2004). This fine line between stimulation and inhibition seems to decide direction and we question if there is scope to also consider a gray area, a period where these signaling metabolons form a synchronized collaboration between transitions. Pathways, which influence and respond to mTORC1 activity, such as the RAS-extracellular signal-regulated kinase (Ras-ERK), AMPK and mitogen-activated protein kinase (MAPK), expose a highway of networks that will be altered should mTORC1 activity change (Mendoza et al., 2011).
Leucine was the first of the amino acids shown to activate mTORC1, blocking autophagy (Hara et al., 1998; Beugnet et al., 2003), but other amino acids such as glutamine, serine and arginine also function as key effectors (Jewell et al., 2015; Carroll et al., 2016). Although the mechanistic details are far less understood than the insulin/IGF pathway, their importance is gaining impetuous as the amino acid profiles or their metabolic enzymes are altered in several disease conditions, including Type 2 diabetes mellitus (T2DM) and AD (Vannini et al., 1982; Wang et al., 2011; Hull et al., 2015). The coupling of amino acid transport and metabolism is intrinsically linked, where activity of the system L (LAT) and system A transporter influence mTORC1 (reviewed in Dodd and Tee, 2012; Goberdhan et al., 2016). In brief, leucine is imported by the solute carrier family 7 member 5 (SLC7A5), which requires glutamine exchange through the Na+-linked system-A transporter (or system ASC (SLC1A5), coupled with the glycoprotein CD98 (Nicklin et al., 2009). Studies indicate that p70S6K was not activated until glutamine was exchanged for leucine, and reactivation of starved cells was dependent on glutamine uptake (Chen et al., 2014). Several factors are involved in amino acid signaling including but not limited to the Rag GTPases, the MAP4K3/GLK pathway, leucyl-tRNA synthetase, the adaptor protein p62 and P13K/hVps34 (Figure 1C; reviewed in Meijer et al., 2015).
Cellular uptake of leucine activates Rag GTPase heterodimers (RagA/B and RagC/D; Sancak et al., 2008; Sancak and Sabatini, 2009), which is dependent on hVps34 expression (Nobukuni et al., 2005). Activated Rag A/B-GTP binds mTORC1, through Raptor, and recruits mTORC1 via the Ragulator complex (MP1, p14 and p18) to the lysosome membrane, where Rheb resides (Kogan et al., 2010). The signaling adaptor p62, which influences cell survival and autophagy, has also been assigned a role in the amino acid induced recruitment of mTORC1 to lysosomes (Duran et al., 2011). Ultimately, Rheb a GTPase, now in close proximity, activates mTORC1-GTP and autophagy is inhibited through ULK1/2 and AMPK phosphorylation, increasing protein synthesis. Through amino acid signaling, a protein complex, called GATOR, and their regulators Sestrin 1/2 and CASTOR 1 modulate the interaction of Rags with mTORC1 (Chantranupong et al., 2014). Interestingly, hVps34, long associated with autophagy, shows increased expression in response to amino acids (Nobukuni et al., 2005). Activation of hVps34 by amino acids induced complex formation with hVps15, which is targeted to early endosomes by Rab7 supporting recruitment of proteins containing FYVE or PX domains (Um et al., 2006). Conversely, as described above during amino acid deprivation, hVps34 in a complex with Beclin 1, UVRAG and hVps15 drives autophagy. Thus, hVps34 expression is a shared protein between mTORC1 and autophagy regulation, the association of which seems to be dictated by nutrient load.
The mitochondrial protein, glutamate dehydrogenase (GDH), has been hypothesized to contribute to mTORC1 and autophagy regulation (Meijer and Codogno, 2008). GDH catalyzes the conversion of glutamate to α-keto glutarate (α-KG) releasing ammonia and NADH. It is thought that α-KG (potentially through propylhydrylase) activates RagB, driving mTORC1 (Durán et al., 2012). What is interesting is that the human branched chain aminotransferase (hBCAT) protein (hBCAT), which catalyzes the transamination of the branched-chain amino acids (BCAAs) and α-KG to glutamate and their respective α-keto acids (Conway and Hutson, 2016), have not been considered in these proposals. The hBCAT proteins are redox sensitive proteins (Conway et al., 2002, 2004, 2008), which form a metabolon with GDH in their reduced form, but when oxidized catalysis is reversibly inactivated (Islam et al., 2007). We hypothesize that in its reduced form hBCAT favors glutamate production through leucine transamination, important for GDH activity and thus mTORC1 activation. However, through either amino acid depletion or through an increase in oxidative stress this unique redox switch changes the function of hBCAT preventing metabolon formation with GDH reducing α-KG. This would inhibit mTORC1 activity and stimulate autophagy. Our work has shown that when hBCAT is overexpressed there is a significant increase in the level of p70S6K and a concomitant reduction in autophagy (unpublished observations). The dynamics and vectorality of these mechanisms are not entirely clear but more than likely will involve a nano-switch, such as that described for hBCAT, which responds to changes in cellular homeostasis.
Under conditions where AD pathology persists, there is a reported loss of mTORC1 regulation, resulting in aggregate accumulation in the cell (Figure 2). Indeed, levels of Akt activation (Griffin et al., 2005), mTORC1 phosphorylated at Ser248 only, together with phosphorylated 4EBP1 (Li et al., 2005), p70S6K (Sun et al., 2014) and eIF4E (Li et al., 2005) were significantly increased in AD brain and correlated with Braak staging and tau pathology, indicating that protein translation is radically disordered. mTORC1 hyper-activation also correlated with cognitive decline in AD individuals (Caccamo et al., 2010; Sun et al., 2014). Neurons immunoreactive to PTEN show reduced expression in AD hippocampus and temporal cortex with a negative correlation to the severity of NFTs and plaques (Griffin et al., 2005). As PTEN attenuates PI3K/Akt signaling, through the dephosphorylation of PIP3, reduced levels can result in the hyper-activation of Akt signaling driving mTORC1 activity. Over activation of the PI3K/Akt/mTORC1 axis would inhibit autophagy and potentially contribute to reduced clearance of Aβ. However, we also need to consider that this would generate sustained p70S6K activation, which should phosphorylate IRS1, causing insulin desensitization and interrupted Akt activation, an apparent contradiction to what was reported in post-mortem AD brain. The most likely explanation is that other pathways, independent of insulin activation of mTORC1, may be responsible, such as (but not limited to) the ERK1/2 pathway, which incidentally was also found to be upregulated in AD brain and cell models (Young et al., 2009; Morales-Corraliza et al., 2016). In cell models, Aβ accumulation exacerbated mTORC1 signaling through phosphorylation of proline-rich Akt substrate of 40 kDa (PRAS40), promoting mTORC1 activity and inhibiting autophagy (Caccamo et al., 2011; Tramutola et al., 2015). Inhibition of mTORC1 using rapamycin alleviated Aβ accumulation and improved cognitive function in AD mice models, supporting this pathway as a future target to regulate neuronal health (Caccamo et al., 2010). However, upstream of mTORC1, we also find that disruptions to glucose and amino acid metabolism linked to insulin resistance and AD pathology add even further complexity.
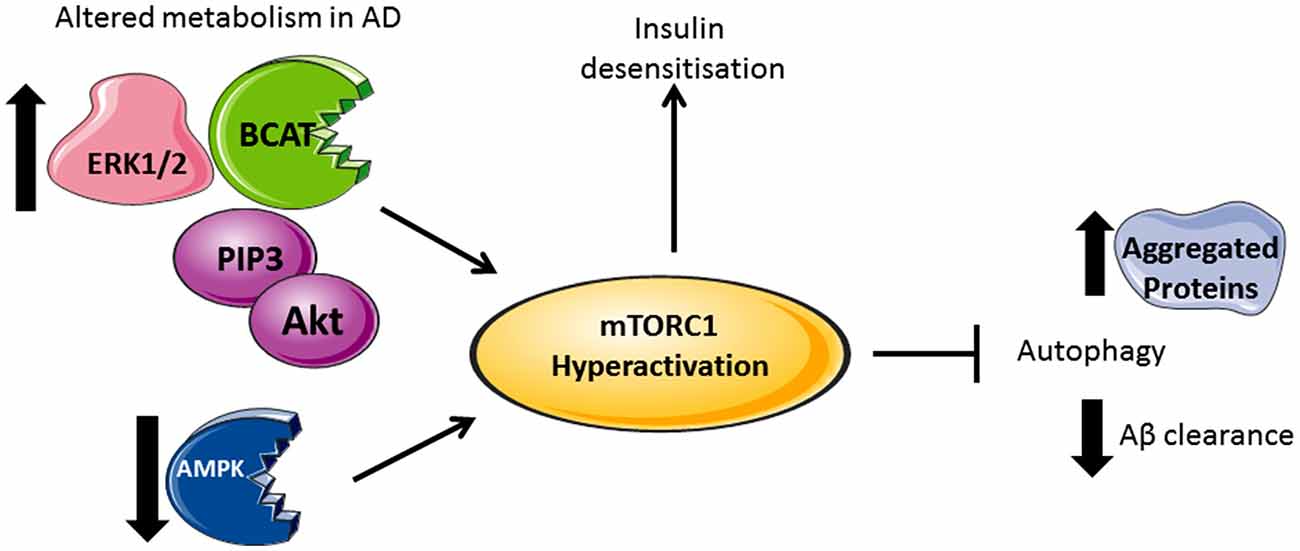
Figure 2. Divergent metabolic regulation in Alzheimer’s disease (AD). In AD brain, levels of Akt, PIP3, ERK1/2, and human branched chain aminotransferase (hBCAT) are increased together with phosphorylation of mTOR at Ser248, 4EBP1, eukaryotic initiation factor 4E (eIF4E) and p70S6K. The decreased expression of PTEN, an inhibitor of PI3K/Akt signaling, further activates mTOR. Over-activation of the PI3K/Akt/mTOR axis inhibits autophagy, supported by reduced levels of Beclin-1 and LC3 in the AD brain, thus reducing aggregate clearance including Aβ. Accumulative Aβ levels, alongside increased p70S6K, cause phosphorylation and inhibition of IRS1 instigating insulin desensitization. mTOR hyperactivation and the associated changes in metabolic proteins correlate with tau pathology and cognitive decline.
Increasing evidence indicates that T2DM doubles the risk of developing AD as well as causing accelerated onset (Biessels et al., 2006; Domínguez et al., 2014; Exalto et al., 2014). Reports of insulin resistance and reduced expression of IGF1 and insulin receptors in AD brain has been linked to mild-cognitive impairment (Steen et al., 2005; Lu et al., 2013; Kim et al., 2015). Insulin resistance is considered to be further perpetuated by levels of Aβ oligomers through increased phosphorylation of the IRS1 inhibitory residue (Ser307; Moloney et al., 2010; O’Neill et al., 2012; Tramutola et al., 2015). Desensitization of neurons to insulin/IGF-1 responses will result in reduced glucose utilization and deficient energy metabolism (Mosconi, 2005). One would anticipate that mTORC1 activation through insulin would subsequently be lost and low cellular glucose should activate the AMPK pathway, further inhibiting mTORC1 driving autophagy. However, in addition to Aβ stimulation of mTORC1, AMPK activity was shown to be diminished in aged brain, and even more pronounced in T2DM, despite reduced intracellular glucose, which may also explain how hyperactivation of mTORC1 persists in the absence of insulin (Kodiha and Stochaj, 2011). Additionally, insulin degrading enzyme (IDE), which regulates extracellular Aβ degradation, showed reduced expression and activity that negatively correlated with Aβ levels in AD (Vekrellis et al., 2000; van der Heide et al., 2006; Zhao et al., 2007). Interestingly, enhanced IDE activity in APP double transgenic mice reduced Aβ levels in the brain preventing plaque formation (Leissring et al., 2003). Together these studies suggest that upregulation of IDE offers therapeutic benefits to target Aβ plaque removal in vivo. Intra-hippocampal administration of insulin in a T2DM rat model attenuated cognitive impairment (McNay et al., 2010) and in a separate study nasal insulin administration in a diabetic mouse model improved diabetic-related decline in cognitive function, offering evidence that overcoming insulin resistance may have therapeutic benefits in AD patients (Wang et al., 2010). A pilot study in humans showed that treatment with intranasal insulin improved delayed memory and preserved general cognition (Craft et al., 2012). However, larger more in-depth studies will decide if this treatment has sustained impact overtime.
Increased blood levels of BCAAs positively correlate with insulin resistance and have been used as signature profiles for T2DM, insulin-resistant states of obesity and Huntington’s disease (Vannini et al., 1982; Mochel et al., 2007, 2011; Wang et al., 2011). For those individuals with T2DM-associated AD increased BCAAs, particularly leucine, could over activate mTORC1 signaling through the various pathways highlighted. Hyperleucinamia in a T2DM mouse model showed that retromer trafficking was impaired, with decreased levels of hVps34 reported, whereby hyperleucinemia may account in part for insulin resistance and driving mTORC1 activation independent of insulin (Morabito et al., 2014). Our group has reported that the hBCAT proteins are significantly upregulated in AD brain relative to age-matched controls and in dementia with lewy bodies and vascular dementia (Hull et al., 2015; Ashby et al., 2017). Here, we proposed that an increase in these proteins was initially to protect neuronal cells through glutamate regulation. In light of our recent work, we now extend their role in brain metabolism as indirect modulators of mTORC1 and autophagy. Here, we suggest that overexpression of hBCAT could contribute to the hyperactivation of mTORC1, disrupting autophagy, potentially through GDH metabolism of glutamate.
On the other hand, supplementation of BCAAs improved glucose homeostasis and insulin resistance in patients with hepatic cirrhosis (Kawaguchi et al., 2008) and increasing dietary leucine intake improved glucose and cholesterol metabolism in mice, indicating that a balance must be met to avoid disequilibrium (Zhang et al., 2007). Contrary to T2DM, levels of BCAAs were reduced in patients recovering from traumatic brain injury and supplementation contributed to improved cognitive function, observed both in humans and rat models of TBI (Vuille-Dit-Bille et al., 2012; Jeter et al., 2013). Although the exact mechanisms controlling this balance between nutrient load and pathology remains elusive these studies highlight the potential for diet to significantly impact these regulatory pathways and should we get the balance correct may be able to delay the onset of AD.
This review has primarily focussed on dysregulated nutrient signaling that impacts autophagy at early endosome formation offering insight into potential pathways that are dysregulated in AD. Clearly, our understanding of how mTORC1 and these signaling networks regulate protein aggregation is far from complete. Importantly, nutrients and growth factors control these pathways and we potentially have an opportunity to regulate brain metabolism through diet. This may be important in neurodegenerative conditions as levels of amino acids are significantly increased in HD and T2DM. However, dysfunctional retromer-dependent trafficking will also be key, in particular with respect to its regulation by nutrient load and cellular stress. It is likely that these pathways operate as metabolons, where clearly a change in function for key metabolic proteins is important for regulation. We speculate that this change in function is triggered by changes in cellular homeostasis, governed by nutrient signals, hypoxia or hormones, and an understanding of which could identify key targets for future neurodegenerative therapeutics. Targeting autophagy and its regulation is therefore of value, where an understanding as to how this mechanism can be controlled will be important to maintain neuronal health.
All authors contributed to the content of the article.
We would like to thank BRACE for supporting this research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Appelqvist, H., Wäster, P., Kågedal, K., and Öllinger, K. (2013). The lysosome: from waste bag to potential therapeutic target. J. Mol. Cell Biol. 5, 214–226. doi: 10.1093/jmcb/mjt022
Ashby, E. L., Kierzkowska, M., Hull, J., Kehoe, P. G., Hutson, S. M., and Conway, M. E. (2017). Altered expression of human mitochondrial branched chain aminotransferase in dementia with lewy bodies and vascular dementia. Neurochem. Res. 42, 306–319. doi: 10.1007/s11064-016-1855-7
Beugnet, A., Tee, A. R., Taylor, P. M., and Proud, C. G. (2003). Regulation of targets of mTOR (mammalian target of rapamycin) signalling by intracellular amino acid availability. Biochem. J. 372, 555–566. doi: 10.1042/BJ20021266
Biessels, G. J., De Leeuw, F. E., Lindeboom, J., Barkhof, F., and Scheltens, P. (2006). Increased cortical atrophy in patients with Alzheimer’s disease and type 2 diabetes mellitus. J. Neurol. Neurosurg. Psychiatry 77, 304–307. doi: 10.1136/jnnp.2005.069583
Borlikova, G. G., Trejo, M., Mably, A. J., Mc Donald, J. M., Sala Frigerio, C., Regan, C. M., et al. (2013). Alzheimer brain-derived amyloid β-protein impairs synaptic remodeling and memory consolidation. Neurobiol. Aging 34, 1315–1327. doi: 10.1016/j.neurobiolaging.2012.10.028
Braak, H., and Braak, E. (1991). Neuropathological staging of Alzheimer-related changes. Acta Neuropathol. 82, 239–259. doi: 10.1007/bf00308809
Caccamo, A., Majumder, S., Richardson, A., Strong, R., and Oddo, S. (2010). Molecular interplay between mammalian target of rapamycin (mTOR), amyloid-beta and tau: effects on cognitive impairments. J. Biol. Chem. 285, 13107–13120. doi: 10.1074/jbc.M110.100420
Caccamo, A., Maldonado, M. A., Majumder, S., Medina, D. X., Holbein, W., Magrí, A., et al. (2011). Naturally secreted amyloid-beta increases mammalian target of rapamycin (mTOR) activity via a PRAS40-mediated mechanism. J. Biol. Chem. 286, 8924–8932. doi: 10.1074/jbc.m110.180638
Carroll, B., Maetzel, D., Maddocks, O. D., Otten, G., Ratcliff, M., Smith, G. R., et al. (2016). Control of TSC2-Rheb signaling axis by arginine regulates mTORC1 activity. Elife 5:e11058. doi: 10.7554/eLife.11058
Cataldo, A. M., Barnett, J. L., Berman, S. A., Li, J., Quarless, S., Bursztajn, S., et al. (1995). Gene expression and cellular content of cathepsin D in Alzheimer’s disease brain: evidence for early up-regulation of the endosomal-lysosomal system. Neuron 14, 671–680. doi: 10.1016/0896-6273(95)90324-0
Cataldo, A. M., Barnett, J. L., Pieroni, C., and Nixon, R. A. (1997). Increased neuronal endocytosis and protease delivery to early endosomes in sporadic Alzheimer’s disease: neuropathologic evidence for a mechanism of increased β-amyloidogenesis. J. Neurosci. 17, 6142–6151.
Cataldo, A. M., Peterhoff, C. M., Troncoso, J. C., Gomez-Isla, T., Hyman, B. T., and Nixon, R. A. (2000). Endocytic pathway abnormalities precede amyloid beta deposition in sporadic Alzheimer’s disease and down syndrome: differential effects of APOE genotype and presenilin mutations. Am. J. Pathol. 157, 277–286. doi: 10.1016/s0002-9440(10)64538-5
Chantranupong, L., Wolfson, R. L., Orozco, J. M., Saxton, R. A., Scaria, S. M., Bar-Peled, L., et al. (2014). The sestrins interact with GATOR2 to negatively regulate the amino-acid-sensing pathway upstream of mTORC1. Cell Rep. 9, 1–8. doi: 10.1016/j.celrep.2014.09.014
Chen, R., Zou, Y., Mao, D., Sun, D., Gao, G., Shi, J., et al. (2014). The general amino acid control pathway regulates mTOR and autophagy during serum/glutamine starvation. J. Cell Biol. 206, 173–182. doi: 10.1083/jcb.201403009
Conway, M. E., and Hutson, S. M. (2016). BCAA metabolism and NH3 homeostasis. Adv. Neurobiol. 13, 99–132. doi: 10.1007/978-3-319-45096-4_5
Conway, M. E., Yennawar, N., Wallin, R., Poole, L. B., and Hutson, S. M. (2002). Identification of a peroxide-sensitive redox switch at the CXXC motif in the human mitochondrial branched chain aminotransferase. Biochemistry 41, 9070–9078. doi: 10.1021/bi020200i
Conway, M. E., Poole, L. B., and Hutson, S. M. (2004). Roles for cysteine residues in the regulatory CXXC motif of human mitochondrial branched chain aminotransferase enzyme. Biochemistry 43, 7356–7364. doi: 10.1021/bi0498050
Conway, M. E., Coles, S. J., Islam, M. M., and Hutson, S. M. (2008). Regulatory control of human cytosolic branched-chain aminotransferase by oxidation and S-glutathionylation and its interactions with redox sensitive neuronal proteins. Biochemistry 47, 5465–5479. doi: 10.1021/bi800303h
Craft, S., Baker, L. D., Montine, T. J., Minoshima, S., Watson, G. S., Claxton, A., et al. (2012). Intranasal insulin therapy for Alzheimer disease and amnestic mild cognitive impairment: a pilot clinical trial. Arch. Neurol. 69, 29–38. doi: 10.1001/archneurol.2011.233
Dodd, K. M., and Tee, A. R. (2012). Leucine and mTORC1: a complex relationship. Am. J. Physiol. Endocrinol. Metab. 302, E1329–E1342. doi: 10.1152/ajpendo.00525.2011
Domínguez, R. O., Pagano, M. A., Marschoff, E. R., González, S. E., Repetto, M. G., and Serra, J. A. (2014). Alzheimer disease and cognitive impairment associated with diabetes mellitus type 2: associations and a hypothesis. Neurologia. 29, 567–572. doi: 10.1016/j.nrl.2013.05.006
Dunn, W. A. (1990). Studies on the mechanisms of autophagy: formation of the autophagic vacuole. J. Cell Biol. 110, 1923–1933. doi: 10.1083/jcb.110.6.1923
Duran, A., Amanchy, R., Linares, J. F., Joshi, J., Abu-Baker, S., Porollo, A., et al. (2011). p62 is a key regulator of nutrient sensing in the mTORC1 pathway. Mol. Cell 44, 134–146. doi: 10.1016/j.molcel.2011.06.038
Durán, R. V., Oppliger, W., Robitaille, A. M., Heiserich, L., Skendaj, R., Gottlieb, E., et al. (2012). Glutaminolysis activates rag-mTORC1 signaling. Mol. Cell 47, 349–358. doi: 10.1016/j.molcel.2012.05.043
Exalto, L. G., van der Flier, W. M., Scheltens, P., Vrenken, H., and Biessels, G. J. (2014). Dysglycemia, brain volume and vascular lesions on MRI in a memory clinic population. J. Diabetes Complicat. 28, 85–90. doi: 10.1016/j.jdiacomp.2012.12.003
Goberdhan, D. C. I., Wilson, C., and Harris, A. L. (2016). Amino acid sensing by mTORC1: intracellular transporters mark the spot. Cell Metab. 23, 580–589. doi: 10.1016/j.cmet.2016.03.013
Griffin, R. J., Moloney, A., Kelliher, M., Johnston, J. A., Ravid, R., Dockery, P., et al. (2005). Activation of Akt/PKB, increased phosphorylation of Akt substrates and loss and altered distribution of Akt and PTEN are features of Alzheimer’s disease pathology. J. Neurochem. 93, 105–117. doi: 10.1111/j.1471-4159.2004.02949.x
Gulati, P., Gaspers, L. D., Dann, S. G., Joaquin, M., Nobukuni, T., Natt, F., et al. (2008). Amino acids activate mTOR complex 1 via Ca2+/CaM signaling to hVps34. Cell Metab. 7, 456–465. doi: 10.1016/j.cmet.2008.03.002
Hamano, T., Gendron, T. F., Causevic, E., Yen, S. H., Lin, W. L., Isidoro, C., et al. (2008). Autophagic-lysosomal perturbation enhances tau aggregation in transfectants with induced wild-type tau expression. Eur. J. Neurosci. 27, 1119–1130. doi: 10.1111/j.1460-9568.2008.06084.x
Hara, K., Yonezawa, K., Weng, Q. P., Kozlowski, M. T., Belham, C., and Avruch, J. (1998). Amino acid sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector mechanism. J. Biol. Chem. 273, 14484–14494. doi: 10.1074/jbc.273.23.14484
Heras-Sandoval, D., Pérez-Rojas, J. M., Hernández-Damián, J., and Pedraza-Chaverri, J. (2014). The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell. Signal. 26, 2694–2701. doi: 10.1016/j.cellsig.2014.08.019
Hull, J., Patel, V., El Hindy, M., Lee, C., Odeleye, E., Hezwani, M., et al. (2015). Regional increase in the expression of the BCAT proteins in Alzheimer’s disease brain: implications in glutamate toxicity. J. Alzheimers Dis. 45, 891–905. doi: 10.3233/JAD-142970
Islam, M. M., Wallin, R., Wynn, R. M., Conway, M., Fujii, H., Mobley, J. A., et al. (2007). A novel branched-chain amino acid metabolon. protein-protein interactions in a supramolecular complex. J. Biol. Chem. 282, 11893–11903. doi: 10.1074/jbc.M700198200
Jeter, C. B., Hergenroeder, G. W., Ward, N. H., Moore, A. N., and Dash, P. K. (2013). Human mild traumatic brain injury decreases circulating branched-chain amino acids and their metabolite levels. J. Neurotrauma 30, 671–679. doi: 10.1089/neu.2012.2491
Jewell, J. L., Kim, Y. C., Russell, R. C., Yu, F. X., Park, H. W., Plouffe, S. W., et al. (2015). Metabolism. differential regulation of mTORC1 by leucine and glutamine. Science 347, 194–198. doi: 10.1126/science.1259472
Kawaguchi, T., Nagao, Y., Matsuoka, H., Ide, T., and Sata, M. (2008). Branched-chain amino acid-enriched supplementation improves insulin resistance in patients with chronic liver disease. Int. J. Mol. Med. 22, 105–112. doi: 10.3892/ijmm.22.1.105
Kim, T. E., Lee, D. H., Kim, Y. J., Mok, J. O., Kim, C. H., Park, J. H., et al. (2015). The relationship between cognitive performance and insulin resistance in non-diabetic patients with mild cognitive impairment. Int. J. Geriatr. Psychiatry 30, 551–557. doi: 10.1002/gps.4181
Klionsky, D. J., and Ohsumi, Y. (1999). Vacuolar import of proteins and organelles from the cytoplasm. Annu. Rev. Cell Dev. Biol. 15, 1–32. doi: 10.1146/annurev.cellbio.15.1.1
Kodiha, M., and Stochaj, U. (2011). AMP kinase: the missing link between type 2 diabetes and neurodegenerative diseases? Trends Mol. Med. 17, 613–614. doi: 10.1016/j.molmed.2011.07.001
Kogan, K., Spear, E. D., Kaiser, C. A., and Fass, D. (2010). Structural conservation of components in the amino acid sensing branch of the TOR pathway in yeast and mammals. J. Mol. Biol. 402, 388–398. doi: 10.1016/j.jmb.2010.07.034
Komatsu, M., Waguri, S., Ueno, T., Iwata, J., Murata, S., Tanida, I., et al. (2005). Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Biol. 169, 425–434. doi: 10.1083/jcb.200412022
Long, Y. C., and Zierath, J. R. (2006). AMP-activated protein kinase signaling in metabolic regulation. J. Clin. Invest. 116, 1776–1783. doi: 10.1172/JCI29044
Liang, C., Lee, J. S., Inn, K. S., Gack, M. U., Li, Q., Roberts, E. A., et al. (2008). Beclin1-binding UVRAG targets the class C Vps complex to coordinate autophagosome maturation and endocytic trafficking. Nat. Cell Biol. 10, 776–787. doi: 10.1038/ncb1740
Leissring, M. A., Farris, W., Chang, A. Y., Walsh, D. M., Wu, X., Sun, X., et al. (2003). Enhanced proteolysis of β-amyloid in APP transgenic mice prevents plaque formation, secondary pathology and premature death. Neuron 40, 1087–1093. doi: 10.1016/s0896-6273(03)00787-6
Lu, J., Xie, G., and Jia, W. (2013). Insulin resistance and the metabolism of branched-chain amino acids. Front. Med. 7, 53–59. doi: 10.1007/s11684-013-0255-5
McNay, E. C., Ong, C. T., McCrimmon, R. J., Cresswell, J., Bogan, J. S., and Sherwin, R. S. (2010). Hippocampal memory processes are modulated by insulin and high-fat-induced insulin resistance. Neurobiol. Learn. Mem. 93, 546–553. doi: 10.1016/j.nlm.2010.02.002
Meijer, A. J., and Codogno, P. (2008). Nutrient sensing: TOR’s ragtime. Nat. Cell Biol. 10, 881–883. doi: 10.1038/ncb0808-881
Meijer, A. J., Lorin, S., Blommaart, E. F., and Codogno, P. (2015). Regulation of autophagy by amino acids and MTOR-dependent signal transduction. Amino Acids 47, 2037–2063. doi: 10.1007/s00726-014-1765-4
Mendoza, M. C., Er, E. E., and Blenis, J. (2011). The Ras-ERK and PI3K-mTOR pathways: cross-talk and compensation. Trends Biochem. Sci. 36, 320–328. doi: 10.1016/j.tibs.2011.03.006
Mochel, F., Benaich, S., Rabier, D., and Durr, A. (2011). Validation of plasma branched chain amino acids as biomarkers in huntington disease. Arch. Neurol. 68, 265–267. doi: 10.1001/archneurol.2010.358
Mochel, F., Charles, P., Seguin, F., Barritault, J., Coussieu, C., Perin, L., et al. (2007). Early energy deficit in huntington disease: identification of a plasma biomarker traceable during disease progression. PLoS One 2:e647. doi: 10.1371/journal.pone.0000647
Moloney, A. M., Griffin, R. J., Timmons, S., O’Connor, R., Ravid, R., and O’Neill, C. (2010). Defects in IGF-1 receptor, insulin receptor and IRS-1/2 in Alzheimer’s disease indicate possible resistance to IGF-1 and insulin signalling. Neurobiol. Aging 31, 224–243. doi: 10.1016/j.neurobiolaging.2008.04.002
Morabito, M. V., Berman, D. E., Schneider, R. T., Zhang, Y., Leibel, R. L., and Small, S. A. (2014). Hyperleucinemia causes hippocampal retromer deficiency linking diabetes to Alzheimer’s disease. Neurobiol. Dis. 65, 188–192. doi: 10.1016/j.nbd.2013.12.017
Morales-Corraliza, J., Wong, H., Mazzella, M. J., Che, S., Lee, S. H., Petkova, E., et al. (2016). Brain-wide insulin resistance, tau phosphorylation changes and hippocampal neprilysin and amyloid-β alterations in a monkey model of type 1 diabetes. J. Neurosci. 36, 4248–4258. doi: 10.1523/JNEUROSCI.4640-14.2016
Morita, M., Gravel, S. P., Hulea, L., Larsson, O., Pollak, M., St-Pierre, J., et al. (2015). mTOR coordinates protein synthesis, mitochondrial activity and proliferation. Cell Cycle 14, 473–480. doi: 10.4161/15384101.2014.991572
Mosconi, L. (2005). Brain glucose metabolism in the early and specific diagnosis of Alzheimer’s disease. FDG-PET studies in MCI and AD. Eur. J. Nucl. Med. Mol. Imaging 32, 486–510. doi: 10.1007/s00259-005-1762-7
Mueller-Steiner, S., Zhou, Y., Arai, H., Roberson, E. D., Sun, B., Chen, J., et al. (2006). Antiamyloidogenic and neuroprotective functions of cathepsin B: implications for Alzheimer’s disease. Neuron 51, 703–714. doi: 10.1016/j.neuron.2006.07.027
Nicklin, P., Bergman, P., Zhang, B., Triantafellow, E., Wang, H., Nyfeler, B., et al. (2009). Bidirectional transport of amino acids regulates mTOR and autophagy. Cell 136, 521–534. doi: 10.1016/j.cell.2008.11.044
Nixon, R. A. (2007). Autophagy, amyloidogenesis and Alzheimer disease. J. Cell Sci. 120, 4081–4091. doi: 10.1242/jcs.019265
Nixon, R. A., Wegiel, J., Kumar, A., Yu, W. H., Peterhoff, C., Cataldo, A., et al. (2005). Extensive involvement of autophagy in Alzheimer disease: an immuno-electron microscopy study. J. Neuropathol. Exp. Neurol. 64, 113–122. doi: 10.1093/jnen/64.2.113
Nobukuni, T., Joaquin, M., Roccio, M., Dann, S. G., Kim, S. Y., Gulati, P., et al. (2005). Amino acids mediate mTOR/raptor signaling through activation of class 3 phosphatidylinositol 3OH-kinase. Proc. Natl. Acad. Sci. U S A 102, 14238–14243. doi: 10.1073/pnas.0506925102
Li, X., Alafuzoff, I., Soininen, H., Winblad, B., and Pei, J. J. (2005). Levels of mTOR and its downstream targets 4E-BP1, eEF2 and eEF2 kinase in relationships with tau in Alzheimer’s disease brain. FEBS J. 272, 4211–4220. doi: 10.1111/j.1742-4658.2005.04833.x
O’Neill, C., Kiely, A. P., Coakley, M. F., Manning, S., and Long-Smith, C. M. (2012). Insulin and IGF-1 signalling: longevity, protein homoeostasis and Alzheimer’s disease. Biochem. Soc. Trans. 40, 721–727. doi: 10.1042/BST20120080
Perez, S. E., He, B., Nadeem, M., Wuu, J., Ginsberg, S. D., Ikonomovic, M. D., et al. (2015). Hippocampal endosomal, lysosomal and autophagic dysregulation in mild cognitive impairment: correlation with aβ and tau pathology. J. Neuropathol. Exp. Neurol. 74, 345–358. doi: 10.1097/NEN.0000000000000179
Pickford, F., Masliah, E., Britschgi, M., Lucin, K., Narasimhan, R., Jaeger, P. A., et al. (2008). The autophagy-related protein beclin 1 shows reduced expression in early Alzheimer disease and regulates amyloid beta accumulation in mice. J. Clin. Invest. 118, 2190–2199. doi: 10.1172/JCI33585
Rohn, T. T., Wirawan, E., Brown, R. J., Harris, J. R., Masliah, E., and Vandenabeele, P. (2011). Depletion of beclin-1 due to proteolytic cleavage by caspases in the Alzheimer’s disease brain. Neurobiol. Dis. 43, 68–78. doi: 10.1016/j.nbd.2010.11.003
Sancak, Y., Peterson, T. R., Shaul, Y. D., Lindquist, R. A., Thoreen, C. C., Bar-Peled, L., et al. (2008). The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320, 1496–1501. doi: 10.1126/science.1157535
Sancak, Y., and Sabatini, D. M. (2009). Rag proteins regulate amino-acid-induced mTORC1 signalling. Biochem. Soc. Trans. 37, 289–290. doi: 10.1042/BST0370289
Seaman, M. N. (2012). The retromer complex - endosomal protein recycling and beyond. J. Cell Sci. 125, 4693–4702. doi: 10.1242/jcs.103440
Shah, O. J., Wang, Z., and Hunter, T. (2004). Inappropriate activation of the TSC/Rheb/mTOR/S6K cassette induces IRS1/2 depletion, insulin resistance and cell survival deficiencies. Curr. Biol. 14, 1650–1656. doi: 10.1016/j.cub.2004.08.026
Small, S. A., and Petsko, G. A. (2015). Retromer in Alzheimer disease, Parkinson disease and other neurological disorders. Nat. Rev. Neurosci. 16, 126–132. doi: 10.1038/nrn3896
Steen, E., Terry, B. M., Rivera, E. J., Cannon, J. L., Neely, T. R., Tavares, R., et al. (2005). Impaired insulin and insulin-like growth factor expression and signaling mechanisms in Alzheimer’s disease—is this type 3 diabetes? J. Alzheimers Dis. 7, 63–80. doi: 10.3233/JAD-2005-7107
Sun, Y. X., Ji, X., Mao, X., Xie, L., Jia, J., Galvan, V., et al. (2014). Differential activation of mTOR complex 1 signaling in human brain with mild to severe Alzheimer’s disease. J. Alzheimer’s Dis. 38, 437–444. doi: 10.3233/JAD-131124
Tan, V. P., and Miyamoto, S. (2016). Nutrient-sensing mTORC1: integration of metabolic and autophagic signals. J. Mol. Cell. Cardiol. 95, 31–41. doi: 10.1016/j.yjmcc.2016.01.005
Tramutola, A., Triplett, J. C., Di Domenico, F., Niedowicz, D. M., Murphy, M. P., Coccia, R., et al. (2015). Alteration of mTOR signaling occurs early in the progression of Alzheimer disease (AD): analysis of brain from subjects with pre-clinical AD, amnestic mild cognitive impairment and late-stage AD. J. Neurochem. 133, 739–749. doi: 10.1111/jnc.13037
Um, S. H., D’Alessio, D., and Thomas, G. (2006). Nutrient overload, insulin resistance and ribosomal protein S6 kinase 1, S6K1. Cell Metab. 3, 393–402. doi: 10.1016/j.cmet.2006.05.003
Wang, T. J., Larson, M. G., Vasan, R. S., Cheng, S., Rhee, E. P., McCabe, E., et al. (2011). Metabolite profiles and the risk of developing diabetes. Nat. Med. 17, 448–453. doi: 10.1038/nm.2307
Wang, X., Campbell, L. E., Miller, C. M., and Proud, C. G. (1998). Amino acid availability regulates p70 S6 kinase and multiple translation factors. Biochem. J. 334, 261–267. doi: 10.1042/bj3340261
Wang, X., Zheng, W., Xie, J. W., Wang, T., Wang, S. L., Teng, W. P., et al. (2010). Insulin deficiency exacerbates cerebral amyloidosis and behavioral deficits in an Alzheimer transgenic mouse model. Mol. Neurodegener. 5:46. doi: 10.1186/1750-1326-5-46
van der Heide, L. P., Ramakers, G. M., and Smidt, M. P. (2006). Insulin signaling in the central nervous system: learning to survive. Prog. Neurobiol. 79, 205–221. doi: 10.1016/j.pneurobio.2006.06.003
Vannini, P., Marchesini, G., Forlani, G., Angiolini, A., Ciavarella, A., Zoli, M., et al. (1982). Branched-chain amino acids and alanine as indices of the metabolic control in type 1 (insulin-dependent) and type 2 (non-insulin-dependent) diabetic patients. Diabetologia 22, 217–219. doi: 10.1007/bf00283757
Vekrellis, K., Ye, Z., Qiu, W. Q., Walsh, D., Hartley, D., Chesneau, V., et al. (2000). Neurons regulate extracellular levels of amyloid beta-protein via proteolysis by insulin-degrading enzyme. J. Neurosci. 20, 1657–1665.
Vuille-Dit-Bille, R. N., Ha-Huy, R., and Stover, J. F. (2012). Changes in plasma phenylalanine, isoleucine, leucine and valine are associated with significant changes in intracranial pressure and jugular venous oxygen saturation in patients with severe traumatic brain injury. Amino Acids 43, 1287–1296. doi: 10.1007/s00726-011-1202-x
Yang, Z., and Klionsky, D. J. (2010). Eaten alive: a history of macroautophagy. Nat. Cell Biol. 12, 814–822. doi: 10.1038/ncb0910-814
Yang, A. J., Chandswangbhuvana, D., Margol, L., and Glabe, C. G. (1998). Loss of endosomal/lysosomal membrane impermeability is an early event in amyloid Abeta1–42 pathogenesis. J. Neurosci. Res. 52, 691–698. doi: 10.1002/(SICI)1097-4547(19980615)52:6<691::AID-JNR8>3.0.CO;2-3
Young, K. F., Pasternak, S. H., and Rylett, R. J. (2009). Oligomeric aggregates of amyloid beta peptide 1–42 activate ERK/MAPK in SH-SY5Y cells via the α7 nicotinic receptor. Neurochem. Int. 55, 796–801. doi: 10.1016/j.neuint.2009.08.002
Yu, W. H., Cuervo, A. M., Kumar, A., Peterhoff, C. M., Schmidt, S. D., Lee, J. H., et al. (2005). Macroautophagy—a novel β-amyloid peptide-generating pathway activated in Alzheimer’s disease. J. Cell Biol. 171, 87–98. doi: 10.1083/jcb.200505082
Zhao, Z., Xiang, Z., Haroutunian, V., Buxbaum, J. D., Stetka, B., and Pasinetti, G. M. (2007). Insulin degrading enzyme activity selectively decreases in the hippocampal formation of cases at high risk to develop Alzheimer’s disease. Neurobiol. Aging 28, 824–830. doi: 10.1016/j.neurobiolaging.2006.05.001
Keywords: autophagy, mTORC1, Alzheimer’s disease, insulin, leucine, BCAT, GDH
Citation: Shafei MA, Harris M and Conway ME (2017) Divergent Metabolic Regulation of Autophagy and mTORC1—Early Events in Alzheimer’s Disease? Front. Aging Neurosci. 9:173. doi: 10.3389/fnagi.2017.00173
Received: 20 January 2017; Accepted: 17 May 2017;
Published: 02 June 2017.
Edited by:
Nibaldo C. Inestrosa, Pontificia Universidad Católica de Chile, ChileReviewed by:
Francisco G. Wandosell, Centro de Biologà-a Molecular Severo Ochoa (CSIC), SpainCopyright © 2017 Shafei, Harris and Conway. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Myra E. Conway, bXlyYS5jb253YXlAdXdlLmFjLnVr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.