 Luis Carlos Chitiva1,2†
Luis Carlos Chitiva1,2† Hair Santiago Lozano-Puentes1,3†
Hair Santiago Lozano-Puentes1,3† Ximena Londoño4Tiago F. Leão2
Ximena Londoño4Tiago F. Leão2 Mónica P. Cala5Eduardo Ruiz-Sanchez6Lucía Ana Díaz-Ariza3Juliet A. Prieto-Rodríguez1Ian Castro-Gamboa2*
Mónica P. Cala5Eduardo Ruiz-Sanchez6Lucía Ana Díaz-Ariza3Juliet A. Prieto-Rodríguez1Ian Castro-Gamboa2* Geison M. Costa1*
Geison M. Costa1*- 1Department of Chemistry, Faculty of Sciences, Pontificia Universidad Javeriana, Bogotá, Colombia
- 2Institute of Chemistry, São Paulo State University (UNESP), Araraquara, Brazil
- 3Department of Biology, Faculty of Sciences, Pontificia Universidad Javeriana, Bogotá, Colombia
- 4Faculty of Agricultural Sciences, Universidad Nacional de Colombia, Palmira, Colombia
- 5Metabolomics Core Facility-MetCore, Universidad de los Andes, Bogotá, Colombia
- 6Department of Botany and Zoology, Universidad de Guadalajara, Jalisco, México
Bamboo species have traditionally been used as building material and potential source of bioactive substances, as they produce a wide variety of phenolic compounds, including flavonoids and cinnamic acid derivatives that are considered biologically active. However, the effects of growth conditions such as location, altitude, climate, and soil on the metabolome of these species still need to be fully understood. This study aimed to evaluate variations in chemical composition induced by altitudinal gradient (0–3000 m) by utilizing an untargeted metabolomics approach and mapping chemical space using molecular networking analysis. We analyzed 111 samples from 12 bamboo species collected from different altitudinal ranges using liquid chromatography coupled with quadrupole time-of-flight mass spectrometry (LC-QTOF-MS). We used multivariate and univariate statistical analyses to identify the metabolites that showed significant differences in the altitude environments. Additionally, we used the Global Natural Products Social Molecular Networking (GNPS) web platform to perform chemical mapping by comparing the metabolome among the studied species and the reference spectra from its database. The results showed 89 differential metabolites between the altitudinal ranges investigated, wherein high altitude environments significantly increased the profile of flavonoids. While, low altitude environments significantly boosted the profile of cinnamic acid derivatives, particularly caffeoylquinic acids (CQAs). MolNetEnhancer networks confirmed the same differential molecular families already found, revealing metabolic diversity. Overall, this study provides the first report of variations induced by altitude in the chemical profile of bamboo species. The findings may possess fascinating active biological properties, thus offering an alternative use for bamboo.
1 Introduction
Bamboo has gained immense value in recent times due to its versatile applications in construction, food, cosmetics, and medicine (Liese et al., 2015; Ming et al., 2017; Chongtham and Bisht, 2020). Bamboo is a rich source of active compounds such as flavonoids, phenolic acid derivatives, alkaloids, terpenes, and essential oils (Coffie et al., 2014; Gomez et al., 2021; Gagliano et al., 2022; Indira et al., 2022; Okido et al., 2022; Cheng et al., 2023) that are characterized by their antioxidant (Panche et al., 2016; Speisky et al., 2022), antimicrobial (Xie et al., 2015), antiviral (Badshah et al., 2021), and anti-inflammatory (Maleki et al., 2019) properties, among others. While most chemical and biological studies have focused on the Asian continent (Clark et al., 2015; Gagliano et al., 2022; Tamang et al., 2022), there is a pressing need to deepen research on Neotropical bamboos to identify alternative uses and create additional value for these species.
The metabolic profile of plants can be affected by several environmental factors, such as temperature, light, ultraviolet radiation levels, precipitation, humidity, nutrients, and altitude (Khalil et al., 2020; Kumari et al., 2022). Additionally, genetic factors, including the presence of genes that control metabolite biosynthesis and the participation of enzymes in different biosynthetic pathways, have been shown to contribute to this variation (Dhami and Mishra, 2015; Sampaio et al., 2016; Pant et al., 2021). However, few studies have investigated changes in the chemical composition of bamboo species due to environmental or genetic effects.
Recent studies have revealed that certain changes can significantly impact the biological potential of bamboo. Specifically, research has shown that seasonal and altitudinal variation in Sasa argenteastriatus (Pleioblastus argenteostriatus (Regel) Nakai) and S. quelpaertensis Nakai leaves is positively correlated with an increase in the content of phenolic and flavonoid compounds, with chlorogenic acid, isoorientin, and vitexin being the most notable compounds exhibiting significant changes (Ni et al., 2012; Ko et al., 2018). Another study on Indocalamus latifolius (Keng) McClure evaluated the impact of altitude on the chemical composition of flavonoids, phenols, and triterpenes, demonstrating that an increase in altitude led to the accumulation of metabolites and a subsequent increase in antioxidant potential (Ni et al., 2013). Based on these findings, we anticipate observing a similar correlation between the increase in phenolic compound content and the species in our study, providing valuable insights for improving crop production and obtaining biologically active metabolites.
Metabolomics is a valuable tool for evaluating metabolic changes in various biological matrices caused by environmental or genetic factors. To analyze large amounts of metabolites in a biological sample, different analytical platforms are currently used. When combined with multivariate analysis, these platforms can identify differentially expressed metabolites, helping to elucidate possible metabolic pathways affected (Verpoorte et al., 2010; Shen et al., 2023). To integrate other platforms and complement the global analysis of the metabolome in bamboo species, we used Global Natural Products Social Molecular Networking (GNPS), a novel platform that facilitates the creation of molecular networks to analyze mass spectrometry (MS/MS) data sets, providing a comprehensive visualization of the chemical space (Wang et al., 2016; Aron et al., 2020; Ramabulana et al., 2021). To our knowledge, this is the first report that applies the untargeted metabolomics approach and molecular networking analysis to the study of the chemical composition of bamboo species under the influence of altitude. This study aimed to evaluate the variations in chemical composition under the influence of an altitudinal gradient (0–3000 m) by utilizing an untargeted metabolomics approach and the mapping of the chemical diversity using molecular networking analysis of the global metabolome in bamboo species.
2 Materials and methods
2.1 Study design
A completely random sampling method was implemented for the metabolomic study by selecting different collection sites in Colombian locations [Cundinamarca (CU), Nariño (NA), Putumayo (PU), and Quindío (QU)] situated at different altitudes, ranging from (0–3000 m). The geographical distributions of the collected species are shown in Figure 1. All samples were collected from natural bamboo in the period 2020 to 2022 (more details are provided in Supplementary Table S1).

FIGURE 1. Geographical distribution map showing the bamboo species studied. Each mark corresponds to the place of collection of each specimen.
2.2 Plant material collection and identification
We collected a total of 111 leaf samples from the upper branches of 12 bamboo species, which were distributed across 40 different altitudes and collected under the same conditions. Harvested bamboo leaves were immediately air-dried at room temperature and then ground into powder for extraction. Based on altitude, we divided the 111 leaf samples into two groups: the low altitude group (∼0–1500 m; n = 40) and high altitude group (∼1500–3000 m; n = 71), as indicated in Table 1. At least three individuals (n = 3) were collected at each collection site, and voucher specimens were deposited in the (HPUJ) Herbarium of the Pontificia Universidad Javeriana.

TABLE 1. Classification of bamboo species according to altitudinal range.
2.3 Metabolite extraction and sample preparation
To extract the plant material (leaves), 300 mg of dried and ground samples were mixed with 10 mL of a solvent containing chloroform, methanol, and water (in a ratio of 5:2.5:2.5 v/v/v). The resulting mixture was then vortexed for 1 min, sonicated for 20 min, and centrifuged at 5000 rpm and 20°C for 10 min. The liquid supernatant was filtered using PTFE syringe filters with a pore size of 0.22 µm (Thermo Scientific, Rockwood, TN), and stored at −80°C until used for metabolomics analysis.
2.4 Untargeted metabolomics using LC-QTOF-MS
Untargeted metabolomics was conducted using an Agilent Infinity 1260 HPLC system coupled to an Agilent 6545 quadrupole time-of-flight (QTOF) mass spectrometer equipped with electrospray ion source (Waldbronn, Germany). 5 μL of the extracts were injected on a C18 column (Kinetex 100 × 2.1 mm, 2.6 µm) at 30°C. The mobile phase employed in LC analyses was composed of 0.1% formic acid in Milli-Q water (v/v) (Solvent A) and acetonitrile (Solvent B) with a constant flow of 400 μL/min. The gradient was set as follows: 3% for 1 min; 3%–97% B in 15 min; 97% B for 2 min; the column was re-equilibrated for 6 min at the initial conditions. Full-scan MS1 and MS/MS spectra were acquired. Data mass spectra were acquired in negative ionization mode (ESI−), in a mass range of m/z 80–1700 Da in data-dependent acquisition (DDA) mode. The QTOF instrument was operated in the 4 GHz (high resolution) mode. The parameters used for data acquisition were set as follows: nitrogen used as nebulizer gas with pressure at 52 psi, a capillary voltage of 3000 V, ion source temperature of 250°C, dry gas flow at 12 L/min, and acquisition rate of one spectrum per second. MS/MS fragmentation was performed using a collision-induced dissociation energy of 20 eV. Throughout the analysis, two reference masses were used for mass correction: m/z 112.9856 [C2O2F3(NH4)] and m/z 1033.9881(C18H18O6N3P3F24).
2.5 Quality control samples
To evaluate system performance and reproducibility in sample analysis, multiple QC samples were created by pooling and mixing equal volumes of each extracted sample. To assess the instrument’s robustness, pooled QC samples were injected before the sample analysis until system equilibration was achieved and after every ten randomized sample injections.
2.6 Metabolomics data processing
The LC-QTOF-MS raw data sets were processed using Agilent MassHunter Workstation Profinder software (B.10.0, Agilent Technologies) to extract molecular features for deconvolution, alignment, and integration. The data were manually inspected to eliminate noise and unrelated ions, and a presence filter was applied. For statistical analysis, features that were present in 100% of the samples for each altitudinal group and had a coefficient of variation (CV) of less than 20% in the QC were selected.
2.7 Statistical analysis
Univariate and multivariate analyses were conducted using MatLab (R2019b, MathWorks, Inc., Natick) and SIMCA 14.0 software (Umetrics, Umeå, Sweden), respectively. The multivariate analysis generated PCA and OPLS-DA models, which were validated using cross-validation less than 0.05 and evaluated based on R2X (change in X explained by the model), R2Y (the total of Y explained), and Q2 (sum parameter in cross-validation). The significantly differential metabolites were identified by calculating the variable importance in the projection (VIP) greater than 1, with jackknife confident interval (JK) not including zero combined with FC > 2.0 or FC < 0.5. The annotated metabolites and their peak area were organized in a table (.csv), which was uploaded to MetaboAnalyst 5.0 software for statistical, functional, and integrative analysis of metabolomics data (https://www.metaboanalyst.ca/). The software was used for visualization using heatmap clustering (Pang et al., 2022), normalized by Pareto scaling. Additionally, univariate analysis was performed to determine the p-value features.
2.8 Metabolite identification
Differential metabolite annotation was conducted by considering the precision of the mass (maximum error of mass 10 ppm), isotopic pattern distribution, and adduct formation, using different public online databases such as METLIN (https://metlin.scripps.edu/), KEGG (https://genome.jp/kegg), HMDB (https://hmdb.ca/), PubChem (https://pubchem.ncbi.nlm.nih.gov/) and ChEBI (https://www.ebi.ac.uk/chebi/) through the CEU Mass Mediator (https://ceumass.eps.uspceu.es/) tool. The identity of the metabolites was confirmed through MS/MS analysis, which included the use of MS-DIAL 4.80 (https://prime.psc.riken.jp/compms/msdial/main.html), MS-FINDER 3.52 (https://prime.psc.riken.jp/compms/msfinder/main.html), CFM-ID 4.0 (https://cfmid.wishartlab.com/) for in silico mass spectral fragmentation, GNPS web platform (https://gnps.ucsd.edu/ProteoSAFe/static/gnps-splash.jsp) and manual interpretation with the Agilent Mass Hunter Qualitative Analysis software (version 10.0). The metabolites were identified according to the metabolomics standards initiative (Schymanski et al., 2014).
2.9 Global natural products social molecular networking (GNPS) web platform workflow description
A molecular network was created using the online workflow (https://ccms-ucsd.github.io/GNPSDocumentation/) on the GNPS website (https://gnps.ucsd.edu/). The precursor ion mass tolerance was set to 0.02 Da and an MS/MS fragment ion tolerance of 0.02 Da. A network was then created where edges were filtered to have a cosine score above 0.6 and more than four matched peaks. Further, edges between two nodes were kept in the network if and only if each of the nodes appeared in each other’s respective top 10 most similar nodes. Finally, the maximum size of a molecular family was set to 0, and the lowest-scoring edges were removed from molecular families until the molecular family size was below this threshold. The spectra in the network were then searched against GNPS spectral libraries. All matches kept between network spectra and library spectra were required to have a score above 0.6 and at least four matched peaks (Wang et al., 2016). To enhance chemical structural information within the molecular network, information from in silico structure annotations from GNPS Library Search, Network Annotation Propagation, Dereplicator were incorporated into the network using the GNPS MolNetEnhancer workflow (https://ccms-ucsd.github.io/GNPSDocumentation/molnetenhancer/). Chemical class annotations were performed using the ClassyFire chemical ontology (Djoumbou Feunang et al., 2016; Mohimani et al., 2017; Da Silva et al., 2018; Ernst et al., 2019). The attribute table of the generated nodes was visualized in the Cytoscape software to analyze the molecular network. The data used for the analysis of molecular networks were deposited in the MassIVE Public GNPS database (http://massive.ucsd.edu) with the accession number MSV000090298. The workflow used in this study is summarized in Figure 2.
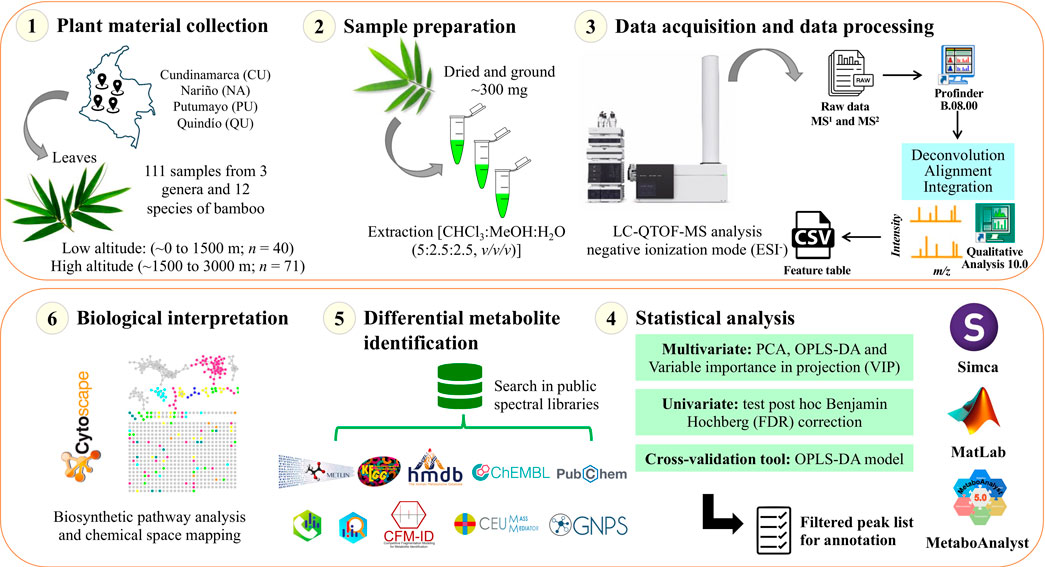
FIGURE 2. Schematic of the experiment analytical workflow used in this study.
3 Results
3.1 Chemical variation of bamboo species presented between low and high altitudes
The quality and stability of the instrument were assessed by conducting a principal component analysis (PCA) on the quality control samples. Supplementary Figure S1 shows a clustering of the QC samples, which indicates the analytical platform’s reliability and the data’s validity. To account for altitudinal variation, we performed PCA and orthogonal partial least squares discriminant analysis (OPLS-DA) models on samples classified into two altitude ranges. Based on this classification, we developed an unsupervised PCA model to observe trends between the two altitude groups. As depicted in Figure 3A, the PCA model showed an initial exploration of the data set, evidencing certain trends between the variables of the altitude groups. Additionally, we constructed a supervised OPLS-DA model to differentiate between low and high altitude groups, with appropriate quality parameters, demonstrating complete separation of the groups. (Figure 3B). This allowed us to identify the significant variables that contributed to the separation between the groups, yielding a total of 89 significant variables with values (false discovery rate, FDR) < 0.05, VIP >1, and FC > 2.0 (or <0.5), as identified according to the metabolomics standards initiative (Schymanski et al., 2014). To validate the OPLS-DA model, we conducted permutation analysis, plotting R2 and Q2 of 200 permutation tests, as shown in Figure 3C.
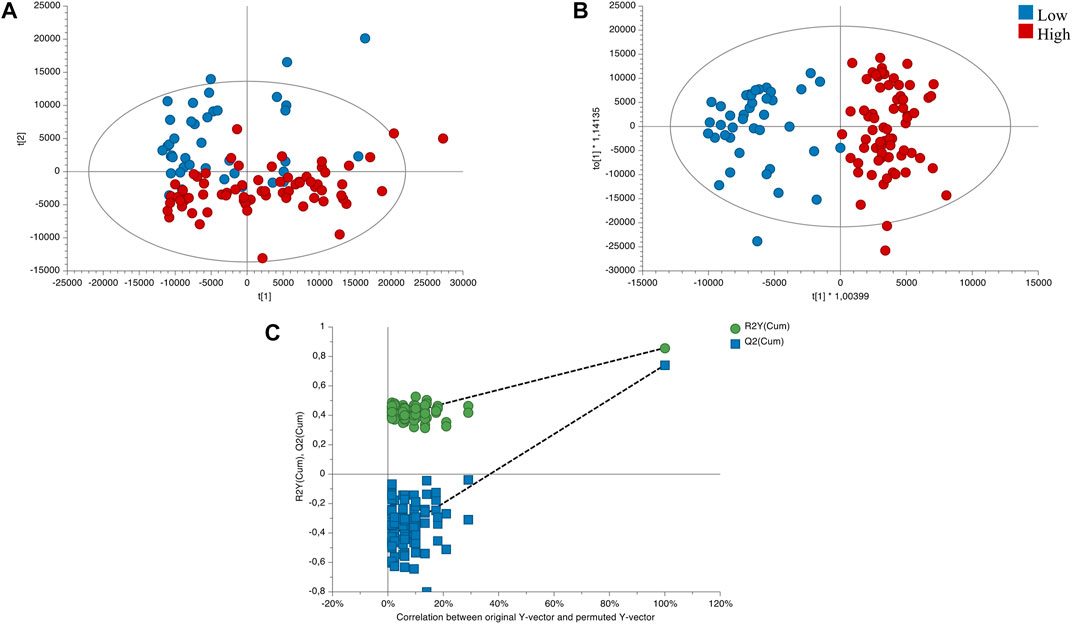
FIGURE 3. Score plots of the influence of direction compared low/high altitudes. (A) PCA score plot of total samples (R2X(cum): 0.813; Q2(cum): 0.54). (B) OPLS-DA score plot of the low vs. high altitudes sample (R2X(cum): 0.494; R2Y(cum):0.858; Q2(cum):0.742; CV Anova: 9.61549e-27). (C) Cross-validation plot of the OPLS-DA model with 200 permutation test. Low altitude (∼0–1500 m; n = 40) and high altitude (∼1500–3000 m; n = 71).
Table 2 presents the differential metabolites (89) that belong to various families, such as flavonoids (48%), fatty acids (19%), cinnamic acid derivatives (11%), unknowns (6%), peptides (3%), glycosylated lignans (2%), alkaloids (2%), carboxylic acids (1%), phenols (1%), carbohydrates (1%), steroids (1%), glycosylated stilbenes (1%) and prenolipids (1%). The largest group of differentially expressed metabolites comprises flavonoids which are the most frequently reported metabolites for these species. Many metabolites in this group showed an upward trend at higher altitudes, potentially contributing to the medicinal potential of these species.
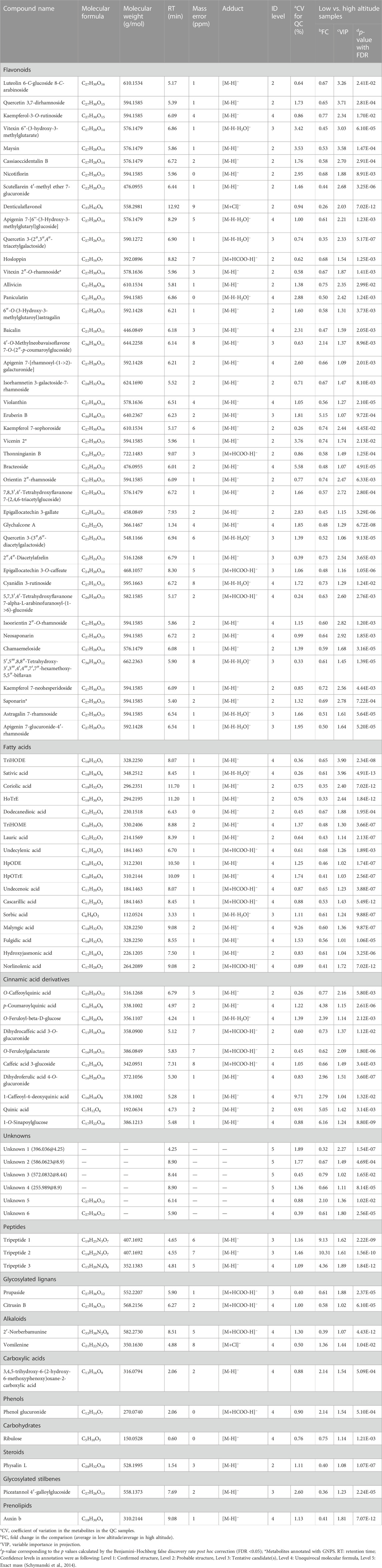
TABLE 2. Significantly differential metabolites found the altitudinal variation.
3.2 Molecular networks and heatmap analyses reveal the chemical variation under the influence of altitudinal variation
To explore the leading chemical classes, we created a molecular network using the GNPS platform, which enabled us to visualize the chemical space of the metabolome in these species. Figure 4A illustrates the molecular network analyzed in the MolNetEnhancer platform, revealing the classes of MF present in the metabolome. We identified seven groups of molecular families, including phenylpropanoids and polyketides, lipids and lipid-like molecules, organic acid derivatives, lignans, neolignans and related compounds, alkaloids, and derivatives, organoheterocyclic compounds, and oxygenated organic compounds. However, due to the complexity of the metabolome in these species and the limitations of the spectral libraries, many of the nodes did not match any spectral reference. Therefore, it is crucial to continue exploring the chemical composition of these species, which is still limited in the literature. Notably, when we created a molecular network of the flavonoid cluster and compared the low vs. high altitude groups of the annotated compounds, we found that vitexin 2″-O-rhamnoside (FC = 1.494; p-value = 1.41E-03), saponarin (FC = 1.458; p-value = 7.22E-04), and vicenin 2 (FC = 1.360; p-value = 2.13E-02) (Supplementary Figures S2A–C) exhibited positive correlations at higher altitudes, confirming the statistical analysis performed previously (Figure 4B).
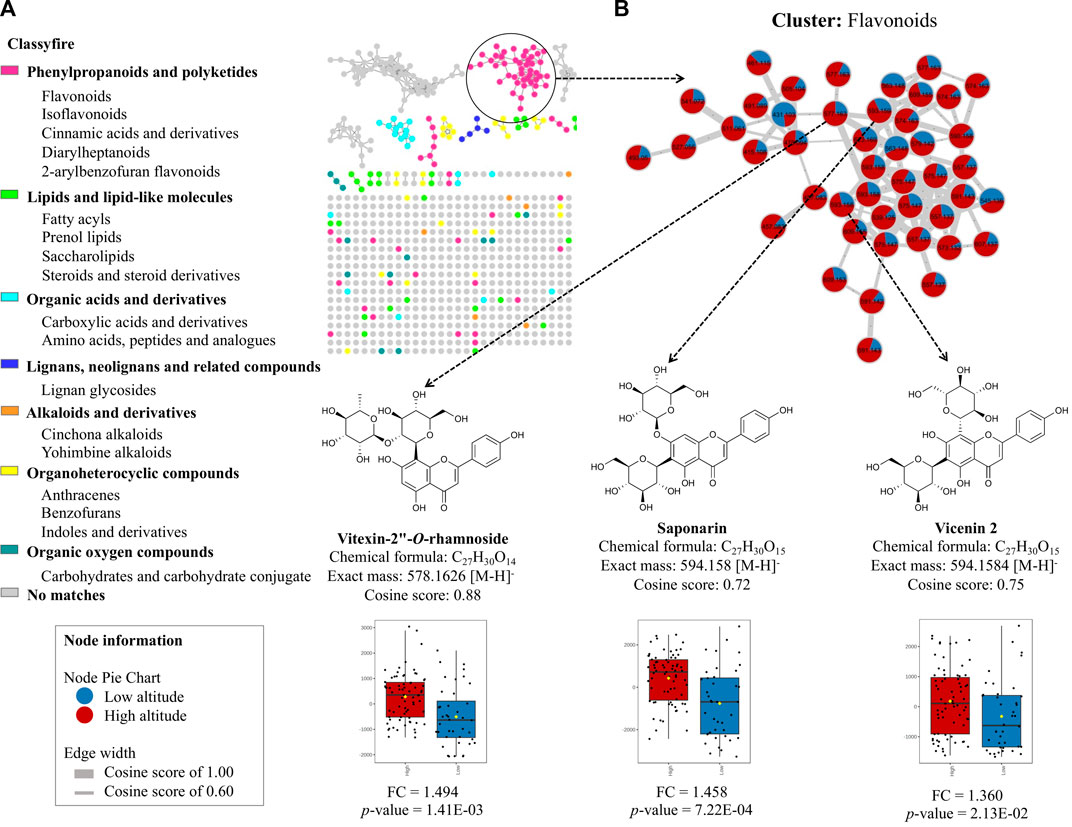
FIGURE 4. Molecular network analysis obtained from GNPS platform online. (A) Identification of major classes of chemical constituents of bamboo species using MolNetEnhancer technique and GNPS molecular networking. Color of the node is set according to the chemical class using “Classyfire”. (B) Molecular network of the flavonoid cluster comparing low altitude (blue) and high altitude (red).
By conducting hierarchical clustering analysis and generating a heatmap based on the two most relevant groups of annotated metabolites that exhibited significant changes with altitude (flavonoids and cinnamic acid derivatives), we identified two distinct groups that were associated with altitude. We found that each group exhibited different patterns in terms of chemical composition and levels of presence and abundance. Figure 5 depicts the clustering of the two altitudinal ranges evaluated, with the first cluster corresponding to the high altitude samples that exhibited a high accumulation of flavonoids, while the low altitude samples exhibited a high accumulation of cinnamic acid derivatives.
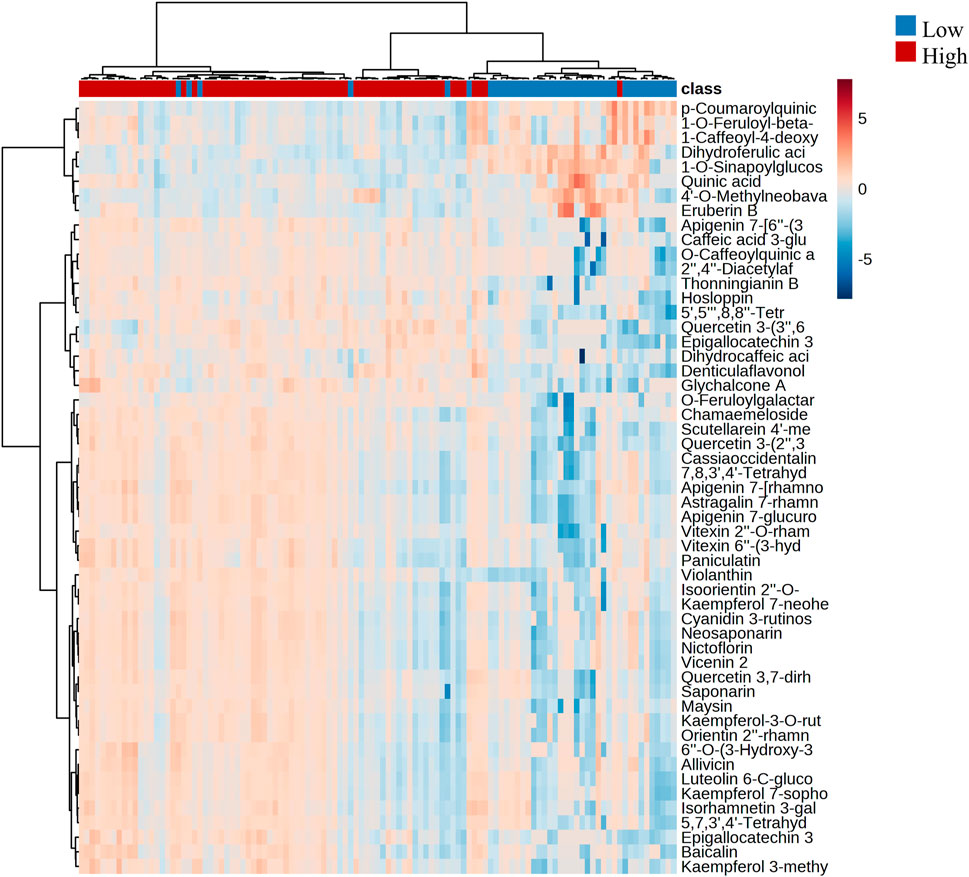
FIGURE 5. Hierarchical clustering with heatmap illustrating the differences in the metabolite abundance between low and high altitudes. The x-axis shows the clustering of all the samples, and the y-axis shows the clustering of the annotated flavonoids and cinnamic acid derivatives.
3.3 Changes in the concentration and analysis of biosynthesis pathways of the most relevant metabolites in bamboo species
The box plot shows the differential metabolites for the two altitude groups. In Figure 6 we present the fold change of the main metabolites that exhibited a significant change. We found that cinnamic acid derivatives and flavonoids showed a significant change with the low and high altitude groups, respectively. Our study also highlighted the significance of quinic acid (QA) is a major differential metabolite, which acts as a crucial precursor in the biosynthetic pathway of phenylpropanoids and is essential in the production of a diverse array of phenolic compounds.
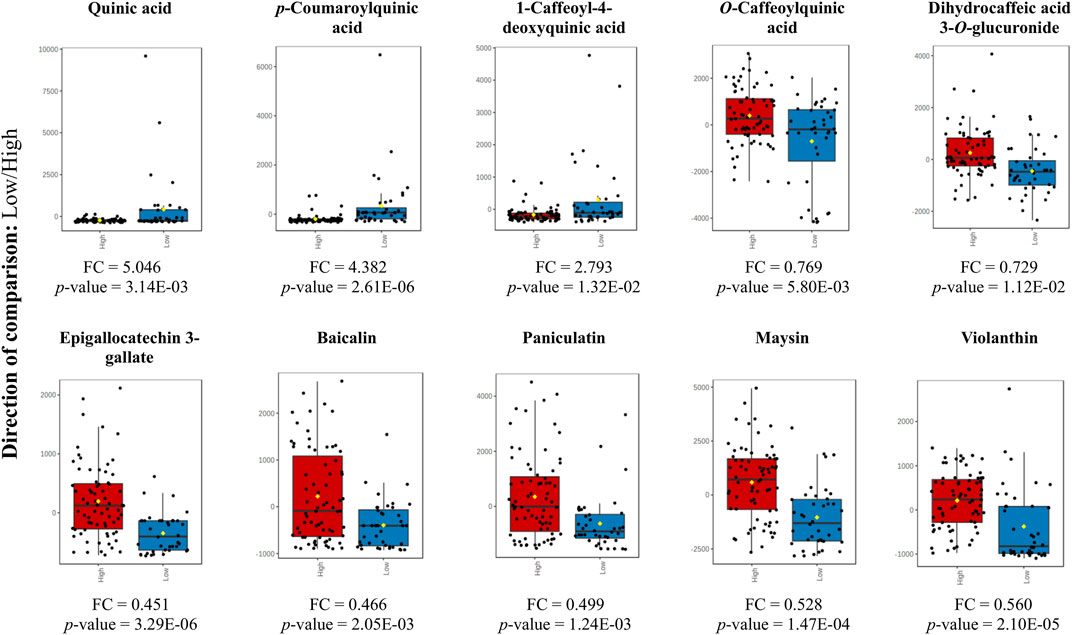
FIGURE 6. Box plot for the altered metabolites corresponding to cinnamic acid derivatives and flavonoids (p-value <0.05) at low altitude (blue) and high altitude (red). Y-axes are represented as relative units. The data were normalized with respect to the total spectral area. Bar charts show normalized values (mean ± one standard deviation). Boxes range from the 5% and 95% percentiles are indicated as error bars; individual data points are indicated by circles. The medians are indicated by horizontal lines within each box.
3.4 Molecular networking for chemical space mapping of the metabolome of bamboo species by MolNetEnhancer
To complement the study on the chemical composition of the global metabolome of bamboo species, a molecular network was constructed using the GNPS platform to compare other types of flavonoids that were not statistically significant among the twelve species studied. Four flavonoid C-glycosides (isorhamnetin 7-rhamnoside, isovitexin, isoschaftoside, and rhoifolin) and one flavonoid O-glycoside (cyanidin 3-O-sophoroside) were annotated (Supplementary Figures S2D–H). It was observed that the Guadua angustifolia species was found to have an abundant profile of flavonoid C-glucosides, specifically of the compound isoschaftoside, which is functionally related to apigenin (Figure 7). In terms of flavonoid variation between species, isovitexin and isorhamnetin 7-rhamnoside were found in most species.
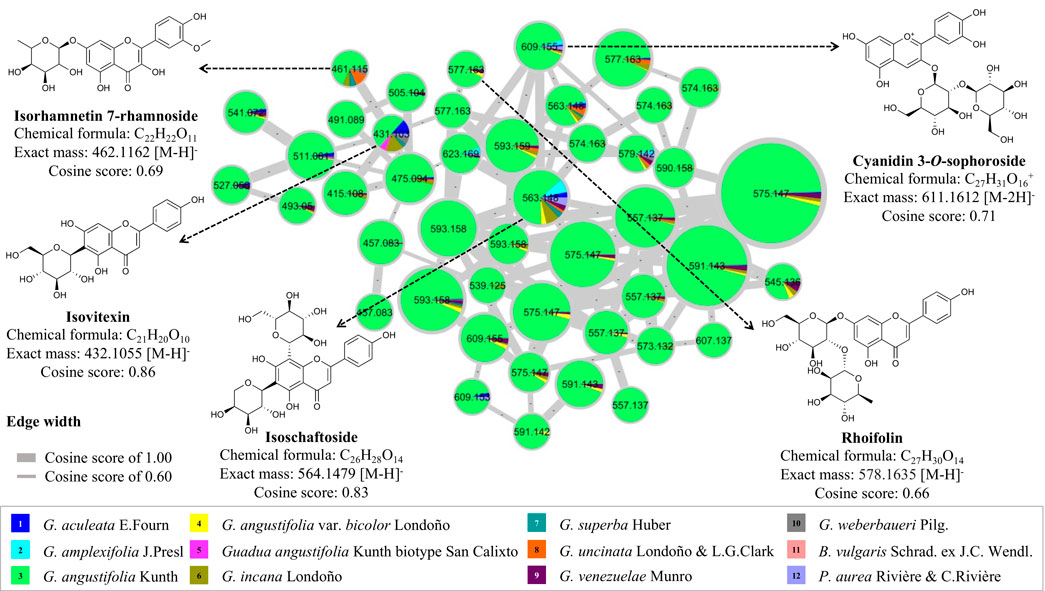
FIGURE 7. Molecular networking and dereplication of flavonoids comparing twelve bamboo species.
4 Discussion
Our findings indicate that flavonoids significantly increased at high altitudes, while cinnamic acid derivatives exhibited an increasing trend at low altitudes. Notably, flavonoids from G. angustifolia showed a positive correlation with the high altitude group, as did cinnamic acid derivatives from G. aculeata, G. amplexifolia, G. angustifolia var. bicolor, G. angustifolia biotype San Calixto, G. incana, G. superba, G. uncinata, G. venezuelae, B. vulgaris, and P. aurea with low altitude (Figure 8). However, further research is needed to determine the exact nature of this correlation. The effect of altitudinal variation on the chemical composition of the leaves of bamboo species is poorly documented. Previous studies have suggested that plants grown at high altitudes tend to have higher levels of flavonoids compared to those grown at low altitudes (Wang et al., 2020; Zhou et al., 2021). This phenomenon can be attributed to an increase of in ultraviolet radiation, illumination time and the delay of the phenophase of the plant, along with elevation, which is a protective mechanism that plants use against unfavorable environmental conditions (Ni et al., 2013). In a recent study, Zhou et al. (2021) investigated the influence of an altitudinal gradient on the variation of flavonoids in Agriophyllum squarrosum and found that these metabolites were enriched at high altitudes. The study also demonstrated a strong positive correlation between the contents of flavonoids, such as quercetin, tricine, and rutin, and environmental variables, such as latitude, longitude, and precipitation gradients. These findings corroborate our results and provide valuable information on the variation in the chemical profile of bamboo under various abiotic factors such as altitude, temperature, light, and soil, etc.
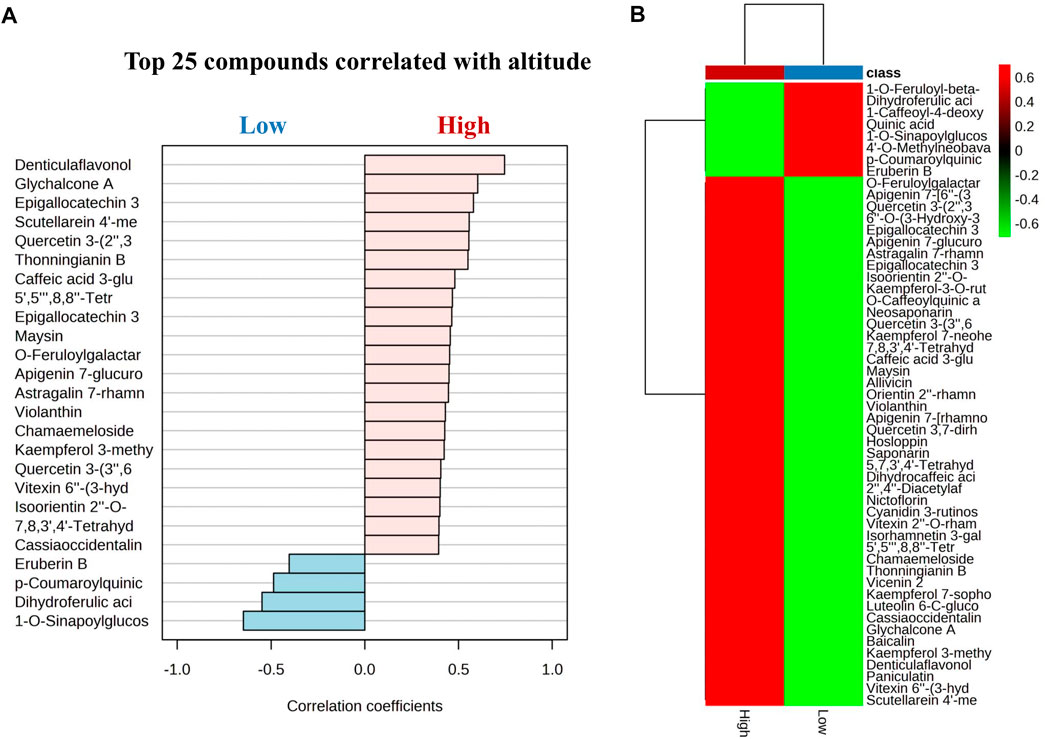
FIGURE 8. (A) Pearson’s correlations using pattern search of the top 25 metabolites between low and high altitude. (B) Cluster heatmap based on correlation between flavonoids and acid cinnamic derivatives and the altitude variable. The analysis shows a positive (red) and negative (green) correlation with a p-value = 0.05 for all the metabolites.
The metabolic differences expressed in bamboo species between high and low altitudes provide valuable information about the ideal growing conditions to promote the production of phenolic compounds with important biological properties in medicine. Moreover, the bamboo leaves have added value as are agro-industrial residues generated in the construction industry. Our results suggest that the accumulation of flavonoids at high altitudes is due to the adaptability of bamboo species to environmental conditions. These giant grasses biosynthesize flavonoid-type phenolic compounds that are used as defence and potential antioxidants (Falcone Ferreyra et al., 2012; Panche et al., 2016; Wang et al., 2018). Notably, the phenylpropanoid biosynthesis pathway, which is the starting point for producing many essential compounds such as flavonoids, coumarins, lignans, and hydroxycinnamic acid conjugates (Fraser and Chapple, 2011), is possibly the pathway that has been altered considering the results of this study. Flavonoid production occurs mainly through a diverse biosynthetic pathway involving the shikimate pathway and polyketide pathways (Mouradov and Spangenberg, 2014; Liu et al., 2021). Interestingly, the phenylpropanoid biosynthetic pathway showed a significant change in the presence of QA, with a fold change of 5.05, VIP of 1.42, and a p-value with FDR 3.14E-03. QA is a metabolite that is closely related to the biosynthesis pathway of caffeoylquinic acids (CQAs) and are specialized bioactive metabolites that are derived from the phenylpropanoid biosynthesis pathway. Consequently, QA is a critical intermediate in the biosynthesis of many flavonoids through cinnamic acid, which is a necessary precursor (Alcázar Magaña et al., 2021; Liu et al., 2021). Furthermore, the study revealed interesting chemical diversity in these species, with a predominance of groups of metabolites that are flavonoids and cinnamic acid derivatives. The mapping of the chemical space (including information about known reference spectra) allowed for the visualization of a large part of the global chemical composition of these species. However, the GNPS platform had a low annotation rate, resulting in many no-matches.
To complement the study of changes in chemical composition under the influence of altitude, the molecular network was used to analyze the specific cluster for flavonoids, comparing low and high altitudes. The study showed that the flavonoids such as vitexin 2″-O-rhamnoside, saponarin, and vicenin 2 had a positive tendency to increase at high altitudes. This finding suggests a close relationship between the concentration of phenolic compounds (flavonoids and cinnamic acid derivatives) in bamboo species. The heatmap shows a clear difference in the content of flavonoids and cinnamic acid derivatives between the groups compared under the effect of the altitudinal gradient, which could serve as marker compounds for chemical classification. The metabolites exposed to the variable altitude showed a significant difference, indicating that environmental factors have previously influenced the genetic and chemical diversity of plants (Pacheco-Hernández et al., 2021).
The metabolome of various bamboo species was compared, it was found that G. angustifolia had a rich profile of C-glycoside flavonoids. Similar metabolites have been reported for other bamboo species such as P. nigra var. henonis (Zhang et al., 2008; Ibrahim et al., 2021) P. pubescens (Tanaka et al., 2014) and B. vulgaris (Akhtar and Patowary, 2022). Some flavonoids were found to be shared among other species, such as G. aculeata., G. angustifolia, G. angustifolia biotype San Calixto, G. incana, G. uncinata, and G. venezuelae. Out of the twelve species analyzed, B. vulgaris and P. aurea were found to be the most studied species at the chemical level in Asia. However, considering the limited information available in the literature and the results obtained from the metabolomic analysis, this study presents an opportunity to explore the metabolome of these species further, especially those belonging to the genus Guadua and distributed in the Neotropical region.
5 Conclusion
This study employed an untargeted metabolomics approach and molecular networking analysis to assess changes in the chemical composition of bamboo species due to variations in altitude. The study revealed that high altitude had a significant influence on the increase of flavonoid profiles, while low altitude led to an increase in cinnamic acid derivatives profiles. The molecular network analysis further demonstrated the diverse chemical composition of these species, including flavonoid glycosides, cinnamic acid derivatives, lignans, alkaloids, carbohydrates, and fatty acids. Conducting metabolomic studies on bamboo can provide a detailed understanding of its chemical composition and metabolic conservation, aiding in identifying patterns and trends in relation to environmental and cultivation factors. This information can help to enhance the production and quality of bamboo, identify bioactive compounds for natural health products, and improve sustainability, positively impacting the industry and the economy.
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author contributions
LC and HL-P writing of the original manuscript; LC, HL-P, LD-A, and GC experimental design and activities; XL taxonomy, classification, and description of bamboo; XL supply of bamboo samples of Quindío; LC, HL-P, and MC data analysis; LC, HL-P, TL, and IC-G molecular networking analysis; XL, TL, MC, ER-S, LD-A, JP-R, IC-G, and GC writing, review and editing of the manuscript. GC for research supervision. All authors contributed to the article and approved the submitted version.
Acknowledgments
The authors would like to acknowledge Pontificia Universidad Javeriana, al Ministerio de Ciencia, Tecnología e Innovación, al Ministerio de Educación Nacional, al Ministerio de Industria, Comercio y Turismo e ICETEX, 2a Convocatoria Ecosistema Científico—Colombia Científica 792-2017, Programa “Generación de alternativas terapéuticas en cáncer a partir de plantas a través de procesos de investigación y desarrollo traslacional, articulados en sistemas de valor sostenibles ambiental y económicamente” (Contrato no. FP44842-221-2018) and the producers linked to the Federación Nacional de Cafeteros and Fundación Suyusama and El Paraiso del Bambú y la Guadua for providing the raw material from different Guadua species. We would also thank the Ministerio de Ambiente y Desarrollo Sostenible for allowing the use of genetic resources and products derived (Contract number 212/2018; Resolution 210/2020).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2023.1192088/full#supplementary-material
References
Akhtar, J., and Patowary, L. (2022). Bambusa vulgaris: A comprehensive review of its traditional uses, phytochemicals and pharmacological activities. Sci. Phytochemistry 1, 11–21.
Alcázar Magaña, A., Kamimura, N., Soumyanath, A., Stevens, J. F., and Maier, C. S. (2021). Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 107, 1399. doi:10.1111/TPJ.15390
Aron, A. T., Gentry, E. C., McPhail, K. L., Nothias, L. F., Nothias-Esposito, M., Bouslimani, A., et al. (2020). Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 15, 1954–1991. doi:10.1038/s41596-020-0317-5
Badshah, S. L., Faisal, S., Muhammad, A., Poulson, B. G., Emwas, A. H., and Jaremko, M. (2021). Antiviral activities of flavonoids. Biomed. Pharmacother. 140, 111596. doi:10.1016/J.BIOPHA.2021.111596
Cheng, Y., Wan, S., Yao, L., Lin, D., Wu, T., Chen, Y., et al. (2023). Bamboo leaf: A review of traditional medicinal property, phytochemistry, pharmacology, and purification technology. J. Ethnopharmacol. 306, 116166. doi:10.1016/j.jep.2023.116166
Chongtham, N., and Bisht, M. S. (2020). Bamboo shoot: Superfood for nutrition, health and medicine. Boca Raton; London: CRC Press.
Clark, L. G., Londoño, X., and Ruiz-Sanchez, E. (2015). “Bamboo taxonomy and habitat,” in Springer (Springer International Publishing Switzerland), 1–30. doi:10.1007/978-3-319-14133-6_1
Coffie, G., Antwi-Boasiako, C., and Darkwa, N. (2014). Phytochemical constituents of the leaves of three bamboo (Poaceae) species in Ghana. J. Pharmacogn. Phytochem. 2, 34–38.
Da Silva, R. R., Wang, M., Nothias, L. F., van der Hooft, J. J. J., Caraballo-Rodríguez, A. M., Fox, E., et al. (2018). Propagating annotations of molecular networks using in silico fragmentation. PLoS Comput. Biol. 14, e1006089. doi:10.1371/journal.pcbi.1006089
Dhami, N., and Mishra, A. D. (2015). Phytochemical variation: How to resolve the quality controversies of herbal medicinal products? J. Herb. Med. 5, 118–127. doi:10.1016/j.hermed.2015.04.002
Djoumbou Feunang, Y., Eisner, R., Knox, C., Chepelev, L., Hastings, J., Owen, G., et al. (2016). ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminform 8, 1–20. doi:10.1186/s13321-016-0174-y
Ernst, M., KangBinCaraballo-Rodríguez, K. A. M., Nothias, L. F., Wandy, J., Chen, C., et al. (2019). MolNetEnhancer: Enhanced molecular networks by integrating metabolome mining and annotation tools. Metabolites 9. doi:10.3390/METABO9070144
Falcone Ferreyra, M. L., Rius, S. P., and Casati, P. (2012). Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 3, 222. doi:10.3389/FPLS.2012.00222/BIBTEX
Fraser, C. M., and Chapple, C. (2011). The phenylpropanoid pathway in arabidopsis. American Society of Plant Biologists. doi:10.1199/TAB.0152
Gagliano, J., Anselmo-Moreira, F., Sala-Carvalho, W. R., and Furlan, C. M. (2022). What is known about the medicinal potential of bamboo? Adv. Traditional Med. 22, 467–495. doi:10.1007/s13596-020-00536-5
Gomez, J. P., Velez, J. P. A., Pinzon, M. A., Arango, J. A. M., and Muriel, A. P. (2021). Chemical characterization and antiradical properties of pyroligneous acid from a preserved Bamboo, Guadua angustifolia Kunth. Braz. Archives Biol. Technol. 64, 1–13. doi:10.1590/1678-4324-2021190730
Ibrahim, A., Abdelhameed, R., Habib, E., Ahmed, S., and Badr, J. (2021). Biological activities of different species of the genus Phyllostachys. Rec. Pharm. Biomed. Sci. 5, 64–73. doi:10.21608/rpbs.2021.61559.1092
Indira, A., Santosh, O., Koul, A., and Nirmala, C. (2022). Comparative assessment of the antioxidant potential of bamboo leaves, along with some locally and commercially consumed beverages in India. Adv. Bamboo Sci. 1, 100007. doi:10.1016/j.bamboo.2022.100007
Khalil, N., El-Jalel, L., Yousif, M., and Gonaid, M. (2020). Altitude impact on the chemical profile and biological activities of Satureja thymbra L. essential oil. BMC Complement. Med. Ther. 20, 186. doi:10.1186/S12906-020-02982-9
Ko, H. C., Lee, J. Y., Jang, M. G., Song, H., and Kim, S. J. (2018). Seasonal variations in the phenolic compounds and antioxidant activity of Sasa quelpaertensis. Ind. Crops Prod. 122, 506–512. doi:10.1016/j.indcrop.2018.06.031
Kumari, A., Lakshmi, G. A., Krishna, G. K., Patni, B., Prakash, S., Bhattacharyya, M., et al. (2022). Climate change and its impact on crops: A comprehensive investigation for sustainable agriculture. Agronomy 12, 3008. doi:10.3390/agronomy12123008
Liese, W., Welling, J., and Tang, T. K. H. (2015). “Utilization of bamboo,” in Bamboo: The plant and its uses (Hamburg; Germany: Springer International Publishing), 299–346. doi:10.1007/978-3-319-14133-6_10
Liu, W., Feng, Y., Yu, S., Fan, Z., Li, X., Li, J., et al. (2021). The flavonoid biosynthesis network in plants. Int. J. Mol. Sci. 22. doi:10.3390/IJMS222312824
Maleki, S. J., Crespo, J. F., and Cabanillas, B. (2019). Anti-inflammatory effects of flavonoids. Food Chem. 299, 125124. doi:10.1016/J.FOODCHEM.2019.125124
Ming, C., Jye, W., and Ahmad, H. (2017). Mechanical properties of bamboo and bamboo composites: A review. J. Adv. Res. Mater. Sci. 35, 7–26.
Mohimani, H., Gurevich, A., Mikheenko, A., Garg, N., Nothias, L. F., Ninomiya, A., et al. (2017). Dereplication of peptidic natural products through database search of mass spectra. Nat. Chem. Biol. 13, 30–37. doi:10.1038/nchembio.2219
Mouradov, A., and Spangenberg, G. (2014). Flavonoids: A metabolic network mediating plants adaptation to their real estate. Front. Plant Sci. 5, 1–16. doi:10.3389/FPLS.2014.00620
Ni, Q., Wang, Z., Xu, G., Gao, Q., Yang, D., Morimatsu, F., et al. (2013). Altitudinal variation of antioxidant components and capability in Indocalamus latifolius (Keng) McClure leaf. J. Nutr. Sci. Vitaminol. (Tokyo) 59, 336–342. doi:10.3177/jnsv.59.336
Ni, Q., Xu, G., Wang, Z., Gao, Q., Wang, S., and Zhang, Y. (2012). Seasonal variations of the antioxidant composition in ground bamboo Sasa argenteastriatus leaves. Int. J. Mol. Sci. 13, 2249–2262. doi:10.3390/ijms13022249
Okido, T., Ino, M., Kamada, I., Kihara, J., and Ueno, M. (2022). Biological activity of Kuma bamboo grass (Sasa veitchii) extract against the fungal causal agent (Pyricularia oryzae) of blast disease. Adv. Bamboo Sci. 1, 100004. doi:10.1016/j.bamboo.2022.100004
Pacheco-Hernández, Y., Villa-Ruano, N., Lozoya-Gloria, E., Barrales-Cortés, C. A., Jiménez-Montejo, F. E., and Cruz-López, M. D. C. (2021). Influence of environmental factors on the genetic and chemical diversity of Brickellia veronicifolia populations growing in fragmented shrublands from Mexico. Plants 10, 1–21. doi:10.3390/plants10020325
Panche, A. N., Diwan, A. D., and Chandra, S. R. (2016). Flavonoids: An overview. J. Nutr. Sci. 5, 1–15. doi:10.1017/jns.2016.41
Pang, Z., Zhou, G., Ewald, J., Chang, L., Hacariz, O., Basu, N., et al. (2022). Using MetaboAnalyst 5.0 for LC–HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 17, 1735–1761. doi:10.1038/s41596-022-00710-w
Pant, P., Pandey, S., and Dall’Acqua, S. (2021). The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers. 18, e2100345. doi:10.1002/CBDV.202100345
Ramabulana, A. T., Petras, D., Madala, N. E., and Tugizimana, F. (2021). Metabolomics and molecular networking to characterize the chemical space of four Momordica plant species. Metabolites 11, 763. doi:10.3390/METABO11110763/S1
Sampaio, B. L., Edrada-Ebel, R., and Da Costa, F. B. (2016). Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 6, 1–11. doi:10.1038/srep29265
Schymanski, E. L., Jeon, J., Gulde, R., Fenner, K., Ruff, M., Singer, H. P., et al. (2014). Identifying small molecules via high resolution mass spectrometry: Communicating confidence. Environ. Sci. Technol. 48, 2097–2098. doi:10.1021/es5002105
Shen, S., Zhan, C., Yang, C., Fernie, A. R., and Luo, J. (2023). Metabolomics-centered mining of plant metabolic diversity and function: Past decade and future perspectives. Mol. Plant 16, 43–63. doi:10.1016/j.molp.2022.09.007
Speisky, H., Shahidi, F., de Camargo, A. C., and Fuentes, J. (2022). Revisiting the oxidation of flavonoids: Loss, conservation or enhancement of their antioxidant properties. Antioxidants 11, 133. doi:10.3390/antiox11010133
Tamang, M., Nandy, S., Srinet, R., Das, A. K., and Padalia, H. (2022). Bamboo mapping using earth observation data: A systematic review. J. Indian Soc. Remote Sens. 50, 2055–2072. doi:10.1007/S12524-022-01600-0
Tanaka, A., Zhu, Q., Tan, H., Horiba, H., Ohnuki, K., Mori, Y., et al. (2014). Biological activities and phytochemical profiles of extracts from different parts of bamboo (Phyllostachys pubescens). Molecules 19, 8238–8260. doi:10.3390/molecules19068238
Verpoorte, R., Choi, Y. H., and Kim, H. K. (2010). Metabolomics: what’s new? Flavour Fragr. J. 25, 128–131. doi:10.1002/ffj.1982
Wang, L., Yuan, S., Nie, Y., Zhao, J., Cao, X., Dai, Y., et al. (2020). Dietary flavonoids and the altitudinal preference of wild giant pandas in Foping National Nature Reserve, China. Glob. Ecol. Conserv. 22, e00981. doi:10.1016/j.gecco.2020.e00981
Wang, M., Carver, J. J., Phelan, V. V., Sanchez, L. M., Garg, N., Peng, Y., et al. (2016). Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat. Biotechnol. 34, 828–837. doi:10.1038/nbt.3597
Wang, T. Y., Li, Q., and Bi, K. S. (2018). Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 13, 12–23. doi:10.1016/j.ajps.2017.08.004
Xie, Y., Yang, W., Tang, F., Chen, X., and Ren, L. (2015). Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 22, 132–149. doi:10.2174/0929867321666140916113443
Zhang, Y., Jiao, J., Liu, C., Wu, X., and Zhang, Y. (2008). Isolation and purification of four flavone C-glycosides from antioxidant of bamboo leaves by macroporous resin column chromatography and preparative high-performance liquid chromatography. Food Chem. 107, 1326–1336. doi:10.1016/j.foodchem.2007.09.037
Keywords: bamboo, Guadua, altitudinal variation, flavonoids, cinnamic acid derivatives, untargeted metabolomics, GNPS, natural products
Citation: Chitiva LC, Lozano-Puentes HS, Londoño X, Leão TF, Cala MP, Ruiz-Sanchez E, Díaz-Ariza LA, Prieto-Rodríguez JA, Castro-Gamboa I and Costa GM (2023) Untargeted metabolomics approach and molecular networking analysis reveal changes in chemical composition under the influence of altitudinal variation in bamboo species. Front. Mol. Biosci. 10:1192088. doi: 10.3389/fmolb.2023.1192088
Received: 22 March 2023; Accepted: 10 May 2023;
Published: 24 May 2023.
Edited by:
Guillermo Moyna, Universidad de la República, UruguayReviewed by:
Andrés Perez Parada, Universidad de la República, UruguayYing Liu, Canadian Food Inspection Agency (CFIA), Canada
Wilton Ricardo Sala-Carvalho, University of São Paulo, Brazil
Copyright © 2023 Chitiva, Lozano-Puentes, Londoño, Leão, Cala, Ruiz-Sanchez, Díaz-Ariza, Prieto-Rodríguez, Castro-Gamboa and Costa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ian Castro-Gamboa, aWFuLmNhc3Ryb0B1bmVzcC5icg==; Geison M. Costa, bW9kZXN0aWNvc3RhLmdAamF2ZXJpYW5hLmVkdS5jbw==
†These authors have contributed equally to this work and share first authorship