 Sara La Manna
Sara La Manna Ilaria De Benedictis
Ilaria De Benedictis Daniela Marasco
Daniela Marasco
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mol. Biosci., 03 January 2022
Sec. Structural Biology
Volume 8 - 2021 | https://doi.org/10.3389/fmolb.2021.792546
This article is part of the Research TopicTailored Modulation of Interactions between Biomolecules: Fundamentals and ApplicationsView all 5 articles
The JAK-STAT pathway is a crucial cellular signaling cascade, including an intricate network of Protein–protein interactions (PPIs) responsible for its regulation. It mediates the activities of several cytokines, interferons, and growth factors and transduces extracellular signals into transcriptional programs to regulate cell growth and differentiation. It is essential for the development and function of both innate and adaptive immunities, and its aberrant deregulation was highlighted in neuroinflammatory diseases and in crucial mechanisms for tumor cell recognition and tumor-induced immune escape. For its involvement in a multitude of biological processes, it can be considered a valuable target for the development of drugs even if a specific focus on possible side effects associated with its inhibition is required. Herein, we review the possibilities to target JAK–STAT by focusing on its natural inhibitors as the suppressor of cytokine signaling (SOCS) proteins. This protein family is a crucial checkpoint inhibitor in immune homeostasis and a valuable target in immunotherapeutic approaches to cancer and immune deficiency disorders.
Janus tyrosine kinases (JAKs) play crucial roles in the transduction of signals triggered by membrane receptors of interleukins during inflammatory response (Groner and von Manstein, 2017; Bousoik and Montazeri Aliabadi, 2018). They are associated with important downstream proteins including signal transducers and activators of transcription (STATs), which in turn regulate the expression of a variety of proteins involved in the induction/prevention of apoptosis (Loh et al., 2019). STATs are able to induce transcription and epithelial/mesenchymal transition (EMT), affect gene expression, generate a pro-tumorigenic microenvironment, and promote cellular self-renewal and differentiation and are involved in the formation of pre-metastatic niches (Groner and von Manstein, 2017; Piranlioglu et al., 2019). Through the JAK-STAT pathway, cytokines, interferons (IFNs), and growth factors are regulated, and extracellular signals are converted into programs of cell growth and differentiation (Yan et al., 2018). It is essential in innate and adaptive immune responses, and its aberrant activation was highlighted in neuroinflammatory diseases, tumor cell recognition, and tumor-driven immune escape (Owen et al., 2019). In addition, the involvement in proliferation and survival provides important roles in resistance to targeted drug treatments (Nascimento et al., 2017).
Aberrant activation of JAK-STAT is encountered in many immune-mediated diseases (Jamilloux et al., 2019), including rheumatoid arthritis (RA) (Malemud, 2018), systemic lupus erythematosus (SLE) (Goropevšek et al., 2017), psoriatic arthritis (PsA) (Fiocco et al., 2014), psoriasis (Kwatra et al., 2012), inflammatory bowel disease (IBD) (Salas et al., 2020), Crohn’s disease, ulcerative colitis (Rogler, 2020), discoid lupus erythematosus (DLE), and dermatomyositis (DM) (Kahn et al., 2018). The hyperactivation of JAK-STAT has been highlighted in the aforementioned diseases, but more recently, it has also been reported in patients with pulmonary hypertension (PH): in their isolated pulmonary arteries, JAK2 and STAT3 mRNA transcript levels and protein expressions were higher than those in health subjects (Roger et al., 2021). In cancer, the hyperactivation of this pathway can be either due to a JAK mutation, as mostly observed in some hematological malignancies (Vainchenker and Constantinescu, 2013), or due to the intrinsic nature of cancer, as in some solid tumors (Qureshy et al., 2020). The activation of the JAK-STAT pathway is strictly correlated to the expression of pro-inflammatory genes, and it is linked to the formation of atheromatous plaques. During early stages of atherogenesis, in vascular smooth muscle cells (VSMCs) and mice models of atherosclerosis, the leukocyte recruitment, the migration and proliferation of VSMCs, and the formation of lipid-laden macrophages occur combined with the activation of the pathway (Gharavi et al., 2007; Boengler et al., 2008; Lim et al., 2008; Ortiz-Muñoz et al., 2009; Taleb et al., 2009; Qin et al., 2014), which, in turn, attenuates nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) expression and activity (Manea S. A et al., 2010; Manea A et al., 2010; Fenyo et al., 2011).
The plethora of involved diseases suggests that the employment of drugs that are able to downregulate JAK-STAT could have beneficial effects (Favoino et al., 2021). The JAK family is made up of four kinases: JAK1-3 and TYK2. While JAK1, 2, and TYK2 are ubiquitously expressed in mammals, JAK3 is expressed mainly in hematopoietic and lymphoid tissues (Ghoreschi et al., 2009). JAKs share a common modular structure with seven JAK homology (JH) regions including the catalytically active kinase domain (JH1) and pseudokinase domain (JH2) without catalytic activity. JAK mechanisms are tuned by their phosphorylation states through transphosphorylation events. JAKs, in turn, phosphorylate tyrosines of type I/II cytokine receptors and STATs, inducing their dimerization and translocation into the nucleus for transcription of pro-inflammatory genes (Rawlings et al., 2004). STATs consist of seven DNA-binding proteins: STAT1-6 (5A, 5B variants) regulate the effector responses, STAT1, 4 are responsible for the antiviral type 1 (Th1) response, STAT6 is responsible for the anti-helminth type 2 (Th2), and STAT3 is responsible for the antibacterial/fungal type 3 (Th17) (O’Shea et al., 2015). Conversely, STAT5 induces the expression of the T-regulatory cell (Treg) transcription factor, Foxp3, which is implicated in their differentiation (Burchill et al., 2007). In the present review, different from many others concerning the inhibition of the JAK-STAT pathway with small-molecule ATP-competitive inhibitors, we focused on the structural approach to design modulators of the entire pathway as mimetics of natural inhibitors as SOCS proteins are natural inhibitors of the kinase activity of JAKs.
Suppressors of cytokine signaling (SOCS) proteins negatively regulate JAK-STAT and consist of eight members: SOCS1-7 and cytokine-inducible SH2-containing protein (CIS) (Alexander and Hilton, 2004; Croker et al., 2008; Yoshimura et al., 2007). While CIS and SOCS1,3 are well characterized as negative feedback regulators of this pathway, the biological functions of other members are still under investigation (Linossi et al., 2013). SOCS proteins have a modular organization of regions: 1) an N-terminal region of variable length, 2) a central Src homology 2 (SH2) domain, and 3) a well-conserved C-terminal domain named SOCS box (Yoshimura et al., 1995). Only SOCS1 and 3 bear a kinase inhibitory region (KIR) in the N-terminal part, even with different mechanisms of action (MOAs). Indeed, while SOCS3 binds simultaneously to JAK and the cytokine receptor proteins (Babon et al., 2014), SOCS1 binds to JAK even in its unphosphorylated form (Liau et al., 2018) (as schematically described in Figure 1) and demonstrates the most potent inhibitor independently from the presence of receptors (Nicholson et al., 1999).
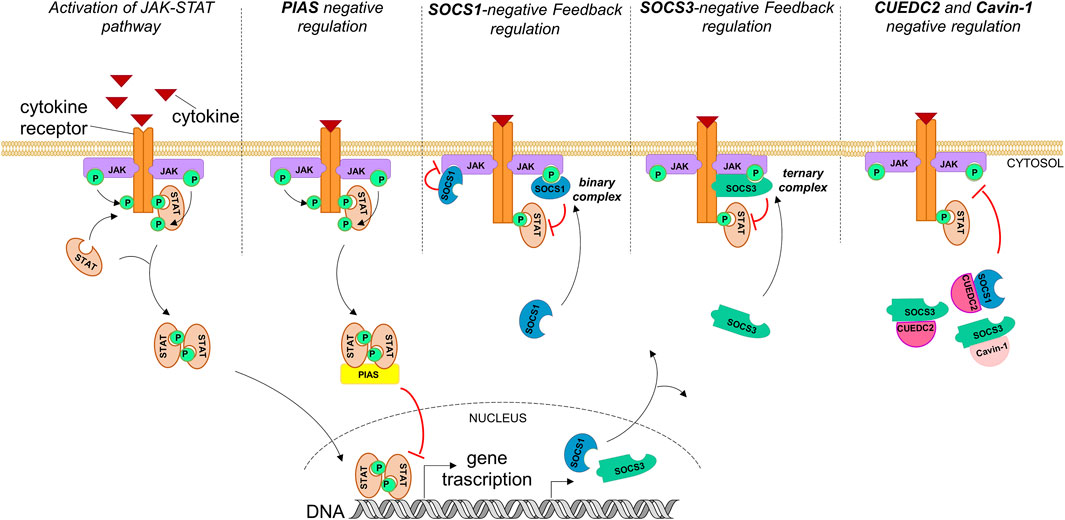
FIGURE 1. Endogenous activation of the JAK-STAT pathway and its regulation through natural inhibitors. PIASs act mainly by suppressing STAT-regulated gene expression and inhibiting STAT binding to DNA. Hyperactivation of the JAK-STAT pathway also activates its own negative feedback regulators: SOCS proteins. SOCS1 and SOCS3 inhibit signaling by different mechanisms: SOCS1 forms a binary complex with JAKs (phosphorylated or unphosphorylated) and inhibits their catalytic activity; SOCS3 inhibits JAK activity through the formation of a ternary complex, binding simultaneously to the cytokine receptor and JAKs. Cavin-1 and CUEDC2 inhibit the JAK-STAT pathway by interacting with SOCS1,3 via an SH2 domain PEST sequence.
SOCS proteins are induced individually by cytokines/growth factors, generating a negative feedback loop with a strictly regulatory function in cellular development and homeostasis, and often demonstrate tumor suppressor and anti-inflammatory effects (Trengove and Ward, 2013) since the repression of SOCS1, 3 leads to the progression of inflammatory diseases (Xin et al., 2020). Hyperglycemia contributes to renal damage through hyperactivation of JAK-STAT in diabetic nephropathy; it has been demonstrated that the overexpression of SOCS proteins reverts the glucose-induced activation. The intrarenal delivery of adenovirus expressing both SOCS1 and SOCS3 in diabetic rats ameliorated renal function and reduced lesions associated with diabetes, as well as the activation of STAT1 and STAT3 and the expression of pro-inflammatory and profibrotic proteins in the diabetic kidney (Ortiz-Muñoz et al., 2010).
In infections caused by herpes simplex virus, cytomegalovirus (Alston and Dix, 2019), chronic hepatitis B virus (HBV), and hepatitis C virus (HCV) (Xie et al., 2021), overexpression of SOCS1 and 3 was observed: this behavior is likely due to an escape mechanism to SOCS-dependent immune regulation. In this context, the employment of inhibitors of SOCSs could provide therapeutic effects (Akhtar and Benveniste, 2011).
SOCS1 strictly regulates IFN signaling (Alexander et al., 1999; Fenner et al., 2006), and interleukins (IL)-2 (Davey et al., 2005), -12, -23 (Eyles et al., 2002), -6 (Tamiya et al., 2011) and their overexpression repress pro-inflammatory genes such as intercellular cell adhesion molecule-1 (ICAM-1), C-X-C motif chemokine ligands (CXCL) 9,10, and C-C motif chemokine ligands (CCL) 2, 5 (Madonna et al., 2008). Its deficiency leads to inflammation (Ihle et al., 1995; Ilangumaran et al., 2004), whereas its haploinsufficiency induces autoimmune diseases (Hadjadj et al., 2020). The ROS-dependent apoptosis of immune cells induces SOCS1 expression, and conversely, SOCS1 overexpression represses apoptosis caused by oxidants (Jung et al., 2016; Schuett et al., 2019). In atherosclerotic models, the inhibition of JAK2, STAT1, and STAT3 prevented lesion formation (Torella et al., 2007; Miklossy et al., 2013), and many efforts based on the therapeutic role of SOCS1,3 effectively deregulate pathological JAK-STAT hyperactivation in cardiovascular disease studies (Recio et al., 2015; Lopez-Sanz et al., 2018).
There is a direct interaction between p53 and SOCS1 that involves the SH2 domain of SOCS1 and the p53 N-terminal transactivation domain 2 (TAD2). Recent investigations outlined that SOCS1 can be phosphorylated on Tyr80 in its extended SH2 domain (ESS) by members of the SRC family of non-receptor tyrosine kinases (SFKs) (Saint-Germain et al., 2019). The phosphorylation modification makes SOCS1 able to dimerize. Both phosphorylation and dimerization regulate the p53-SOCS1 tumor suppressor axis: SOCS1 bridges p53 and ATM at DNA damage foci, leading to p53 phosphorylation and a subsequent increase in its transcriptional activity (Calabrese et al., 2009).
During early phases of atherosclerosis, STAT3 activation represses SOCS3 levels that, in turn, increases resistin, cytokine, and reactive oxygen species (ROS) production depending on NADPH oxidase (by increased NOX1 expression) (Gan et al., 2013; Barrett et al., 2019), while the overexpression of SOCS1 represses ROS generation by inhibiting NOX expression caused by the upstream inactivation of JAK2, STAT1, and phosphoinositide 3-kinase (PI3K) (Hernández-Vargas et al., 2005; Ortiz-Muñoz et al., 2009; Recio et al., 2015). Physiological levels of SOCS3 are unable to limit acute inflammation (Orr et al., 2007), but the presence of its recombinant form suppresses it (Jo et al., 2005). Indeed, in malignant fibrous histiocytoma (Shouda et al., 2006), colorectal (Chu et al., 2017), ovarian cancer cell lines (Shang et al., 2017), and solid tumors based on the IL-6/STAT3 axis, SOCS3 overexpression demonstrated beneficial effects: in T47D and in MCF7 BC (breast cancer) cells, it represses cell proliferation through reduction in STAT3 expression and STAT5 phosphorylation (Barclay et al., 2009), and in mouse xenograft models, it represses tumor growth and metastasis formation (Hill et al., 2010), while downregulated SOCS3 expression provides enhanced risks of recurrent disease in BC patients (Ying et al., 2010). On the other hand, STAT3 hyperactivation [as in cholangiocarcinoma (CCA)] determines an epigenetic silencing of the SOCS3 gene and cellular resistance to apoptosis and induction of EMT (Isomoto, 2009). In this context, a SOCS3 mimetic could have great therapeutic importance since, in addition to being a potential anti–IL-6R compound, it could revert harmful effects of STAT3 activation for the progression of carcinoma (Zhou et al., 2015).
Recently, several pieces of evidence emerged of additional SOCS-interacting proteins required for optimal inhibition of cytokine signaling: cavin-1, an essential component of caveolae (Cheng and Nichols, 2016), and CUEDC2 [coupling of ubiquitin to endoplasmic reticulum degradation (CUE) domain-containing protein 2], which is important for cell cycle control (Gao et al., 2011; Zhang et al., 2013). Their MOAs are schematically reported in Figure 1. In detail, the interaction of SOCS3 with cavin-1 requires its SH2 domain and is independent of its ability to bind Tyr-phosphorylated targets. The structural required element for the interaction is a PEST sequence present in almost all SH2 domains (Wang et al., 2001); the SOCS3 PEST sequence revealed that it is able to control its turnover without affecting the inhibition of JAK-STAT (Babon et al., 2006). Conversely, CUEDC2 can suppress the phosphorylation and activation of the JAK1-STAT3 axis but requires the presence of SOCS3 for a similar direct interaction assessed in vitro (Zhang et al., 2012). However, at present, no structures of SOCS3 or its SH2 domain with either full length cavin-1 (Williams et al., 2018) or CUEDC2 are available (Zhang et al., 2012). The lack of structural information prompted several authors to employ an overlapping peptide array approach with the aim to identify and optimize high-affinity SOCS-derived sequences as antagonists of the interaction (Brown et al., 2013). The identification of CUEDC2 and cavin-1 as regulators of SOCS function and stability could represent another potent approach for the modulation of the JAK-STAT pathway, since both cavin-1 and CUEDC2 enhance SOCS3 function (and SOCS1 in the case of CUEDC2).
An important class of protein regulators of STAT proteins that function often in conjunction with SOCS is represented by the protein inhibitor of activated STAT (PIAS) (Niu et al., 2018). In RA pathology, SOCSs and PIASs are dysfunctional (Malemud, 2016), and experimental overexpression of SOCS3 in RA synovial tissue represses the inflammation (Shouda et al., 2001; Mahony et al., 2016; Silvagni et al., 2021). Different from SOCSs, they are expressed constitutively and are involved in apoptosis during normal homeostasis, as well as cell survival and tissue renewal (Mohan et al., 2020; Xin et al., 2020). They belong to a protein family of five components: PIAS1, PIASxα, PIASxβ, PIAS3, and PIAS4 (PIASγ) (Heppler and Frank, 2017), with a modular structure having a serine/threonine-rich domain located at the C terminus, a Zn-binding RING finger–like domain at the central portion, and a conserved SAP domain (scaffold attachment factor A/B, Acinus, PIAS) close to the N terminus (Kipp et al., 2000). PIAS proteins inhibit STAT transcriptional activity through three major mechanisms: 1) the direct interaction with STAT and blocking of STAT–DNA interactions (Prigge and Schmidt, 2006), 2) the recruitment of transcriptional cofactors to STAT target genes (Heppler and Frank, 2017), and 3) the induction of SUMOylation of the protein (Ungureanu et al., 2003; Yuan et al., 2015) (Figure 1).
Currently, the employment of inhibitors of JAK proteins as small molecules is clinically accepted in proliferative neoplasia and in autoimmune diseases, where they modulate immune responses, T-cell differentiation and B-cell activation (Villarino et al., 2015). Conversely, they are under clinical investigation in chronic inflammatory skin diseases where they can be also be applied topically with innovative formulation systems (Welsch et al., 2017). Usually, JAK inhibitors (JAKis) (Table 1) target the ATP-binding site of the kinase domain but simultaneously block different pathways: these features confer to these agents a “broad-spectrum” MOA (Qureshy et al., 2020) as well as immunosuppressive drugs (Flanagan et al., 2010). Preclinical studies in solid tumor cell lines show that JAK inhibitors decrease STAT activation, cell proliferation and survival, and tumor growth in in vivo models (Schwartz et al., 2017). The first U.S. Food and Drug Administration (FDA)–approved JAKi, ruxolitinib, is a JAK1,2 inhibitor that was registered for myeloproliferative disorders (Verstovsek et al., 2010); during preclinical studies, it demonstrated the ability to sensitize cell lines and murine models to chemotherapy, immunotherapy, and oncolytic viral therapy and provides good results in combination with other drugs as capecitabine for advanced/metastatic pancreatic cancer (Hurwitz et al., 2018) and BC (O'Shaughnessy et al., 2018) or afatinib in non–small-cell lung carcinoma (NSCLC) (Park et al., 2019). In multiple myeloma (MM) patients’ cells and in RPMI-8226 and U266 MM cell lines, JAK1 and JAK2 resulted in overexpression, and upon treatment with ruxolitinib and bortezomib, these cells presented 50% of cells in late apoptosis, a reduction in antiapoptotic gene expression, and a higher number of cells in the SubG0 phase. In addition, the combination of ruxolitinib, bortezomib, and lenalidomide provided 72% of cell death similar to the results obtained upon the treatment of a combination of bortezomib, lenalidomide, and dexamethasone that is actually employed in clinical practice (de Oliveira et al., 2017). Tofacitinib, selective for JAK2, 3, was approved by the FDA for the therapy of RA and ulcerative colitis (Dhillon, 2017), while in IBD, it demonstrated efficacy in early phase trials (Harris and Cummings, 2021).

TABLE 1. Most common JAKis: their preferential targets and diseases where they are on market/in clinical trials.
One of the most important issues in BC is the onset of chemoresistance. A recent study reported the effects of several JAKis on the kinase activity of chemotherapy-resistant MCF7 (MCF7Res) and chemotherapy-sensitive MCF7 cells. In detail, the treatment with tofacitinib in combination with doxorubicin induced a conversion from chemoresistant to chemosensitive cells, leading to apoptosis (Nascimento et al., 2017). In Table 1, most common JAKis and their preferential targets and diseases for which they are on the market/in clinical trials are reported.
While JAKis are in advanced stages of clinical development, inhibitors of STATs are in early development since these proteins are less easily druggable with respect to enzymes such as JAKs. STAT3 and STAT5 demonstrated to be involved in many cellular processes including the regulation of growth and survival (de Araujo et al., 2020); consequently, they also exert a primary role in oncological processes. Efforts in the inhibition of STATs include small molecules, antisense and decoy oligonucleotides, and oligopeptides (Gadina et al., 2018; Yang et al., 2019).
Phosphorylated STAT3 (pSTAT3) is strictly involved in cellular plasticity processes that imply the coexistence of cancer stem cells (CSCs) and the EMT. The administration of a STAT3 inhibitor, napabucasin, revealed its ability to improve chemosensitivity of resistant cells acting at the level of interconversion from differentiated to stem-like states (Shih and Mei, 2021).
A series of non-competitive inhibitors of STAT3 were identified by structure-based high-throughput virtual screening; it includes STA-21, Stattic (Stat three-inhibitory compound), S3I-201/NSC74859, BP-1–102, OPB-31121, TTI-101 (C188-9), and their analogs. Most of them demonstrated the ability to inhibit cell growth of several cancer lines for the suppression of the binding of STAT3 to its pY-peptide ligands and thus blocking IL-6–STAT3 activation, nuclear translocation, and transcriptional gene activation. Preclinical studies of these small STAT3 inhibitors showed promising results (Bharadwaj et al., 2020): when tested in several mouse models of fibrosis, including bleomycin-induced lung and skin fibrosis (scleroderma) and in combination with cucurbitacin-B, some of them revealed their ability to reduce carbon tetrachloride–induced liver fibrosis that is accompanied by reduced levels of pSTAT3 (Sallam et al., 2018).
One controversial issue concerns the selectivity of JAKis: first-generation inhibitors target different JAKs, but their efficacy is often associated with adverse events. Indeed, herpes zoster infections were more frequent in patients treated with tofacitinib than in those receiving biological therapeutics such as antibodies siltuximab (anti–IL-6), tocilizumab (anti–IL-6R), secukinumab (anti–IL-17), and mepolizumab (anti–IL-5) (Dymock et al., 2014). In addition, in several cases, thrombocytopenia and neutropenia were encountered (Lee et al., 2014). Nevertheless, similar studies in RA patients did not point out increased risk of hematological cancers or solid tumors after long-term treatment with ruxolitinib (Durham et al., 2019). Actually, many pharmaceutical efforts toward selective JAKis are underway as filgotinib is selective for JAK1 (Mease et al., 2018). However, the exclusive search for selectivity could lead to a loss of inhibitory effects and points out the necessity of novel therapeutic strategies including alternative ways targeting JAK-STAT with minimum adverse reactions (Ou et al., 2021).
Protein–protein interactions (PPIs) tune biological processes and are deeply involved in the development and progression of diseases (Milroy et al., 2014). Proteomic investigations pointed out that hundreds of thousands of PPIs occur (Stumpf et al., 2008); hence, considerable pharmaceutical efforts are focused on their modulation for therapeutic intervention (Barnard et al., 2015; Scott et al., 2016; Feng et al., 2017; Stefan et al., 2018; Hymel et al., 2021). In this context, small-molecule approaches often fail mainly because such a compound for its size cannot establish a suitable number of favorable interactions, thus hampering this traditional pharmaceutical approach. Conversely, proteomimetics represent more efficient and promising agents toward PPIs. In them, a scaffold reproducing the spatial and angular projections of “hot spot” side chains at the protein interfaces (Gigante et al., 2020) mimicking secondary (e.g., α-helix and β-strand) and tertiary (e.g., coiled-coil) structure (de Araujo et al., 2014; Lingard et al., 2014; Di Natale et al., 2020a) is developed. In this context, peptides and peptidomimetics are elective compounds (Scognamiglio et al., 2013; Carotenuto et al., 2014; La Manna et al., 2018a): they act as proteomimetics able to selectively recognize their target protein in biophysical assays and to mimic, totally or partially, the cellular function of native proteins (Lonardo et al., 2010; Causa et al., 2013; Sawyer et al., 2017). A prominent strategy, mostly lying on the knowledge of the structure of protein complex, consists in the identification of isolated protein fragments mainly responsible for the formation of the protein complex and successively on their conformational stabilization through covalent restraints to improve the affinity and stabilities of unstructured peptides for target proteins (Russo et al., 2015; Soler et al., 2017; Adedeji Olulana et al., 2021; La Manna et al., 2021). In this scenario, several structure-based inhibitors demonstrated discrete success. Annexin A1 (AnxA1) protein has a potent anti-inflammatory effect, and its mimetic peptide, Ac2-26, was able to ameliorate inflammation and wrinkle formation in TNF-α/IFN-γ-stimulated human keratinocytes (HaCaT) and fibroblasts (Detroit 551) and decrease the expression of pro-inflammatory chemokines through the inhibition of several cellular pathways including JAK-STAT. It also induced the synthesis of collagen by generating pro-collagen and the suppression of collagen degradation by inhibiting the expression of collagenases matrix metalloproteinase (MMP)-1 and MMP-8 (Kim et al., 2020). Similarly, the N-terminal domain (NTD) of STAT3 is considered to be mainly involved in STAT3-interactomes, and structural data available allowed for the rational design of analogs of the helix 2 of STAT3 able to directly and selectively bind STAT3 but not STAT1 and to induce apoptosis in BC but not in normal breast cells or STAT3-deficient fibroblasts (Timofeeva et al., 2007). By applying the same approach to STAT1, its NTD was employed to design inhibitors of STAT1 transactivation able to block cytokine-induced STAT1-dependent proliferation in metanephric progenitors, tubulogenesis, and several antiapoptotic factors such as myeloid cell leukemia-(Mcl)-1 and heat shock protein (Hsp)-27 in renal tumor cells with constitutively active STAT1 (Wang et al., 2010).
SOCS proteins have crucial regulatory functions in immune homeostasis, and the reproduction of their biological roles is conceived as a valuable strategy in immunotherapeutic approaches in cancer and immune deficiency disorders (Johnson et al., 2020). In this scenario, new SOCS1, 3 proteomimetics endowed with anti-inflammatory/antioxidant properties have been designed and tested by us and other groups, paving the way to the employment of peptide-based compounds as novel therapeutics that, recently, are under growing magnifying light (Viparelli et al., 2008; Russo et al., 2016; La Manna et al., 2017a; Madonna et al., 2017; La Manna et al., 2018a; La Manna et al., 2018b; La Manna et al., 2020a).
SOCS1 is a direct inhibitor of JAKs; from the crystal structure of the complex SOCS1-JAK1, SOCS1 employs both SH2 and KIR domains for the protein interaction, exhibiting poor structural variations once bound to the kinase. Acting as a pseudosubstrate of kinase (Giordanetto and Kroemer, 2003), only KIR appears more ordered with respect to the unbound state, even if it does not assume any canonical secondary structures (Liau et al., 2018). Within it, the fragment His54-Arg59 forms polar and hydrophobic bonds with residues located in JAK1 catalytic domain. Between the side chains of His885 and Pro1044 of JAK1, His54 of SOCS1 is inserted, while Phe55 acts as the P+1 residue of the pseudosubstrate. These residues, along with Phe58, form the greatest number of hydrophobic contacts with the activation loop and the α-G helix of kinase (Liau et al., 2018).
On the basis of previous mutational studies, several research groups carried out numerous studies where they demonstrated that the linear peptide spanning KIR domain (52–67) inhibits/reduces 1) the activation of STAT by cytokines Th1 and 17 in leukocytes; 2) the activation and migration of vascular cells and macrophages in vitro (Ahmed et al., 2015); 3) the expression of cytokines with pro-inflammatory properties in atherosclerotic plaques (Recio et al., 2014); 4) the renal inflammation, oxidative stress, and fibrosis (Recio et al., 2017; Lopez-Sanz et al., 2018; Opazo-Ríos et al., 2020); 5) the chronic intraocular inflammatory disease (as uveitis) (He et al., 2016; Recio et al., 2017; Ahmed et al., 2018) [very recently, in equine recurrent uveitis (ERU), that is, the only spontaneous model of human recurrent uveitis (Larkin et al., 2020)]; 6) the inflammation in the abdominal aortic aneurysm (AAA), to downstream target genes, and to prevent elastase formation induced by AAA (Bernal et al., 2020); and 7) the severity of skin lesions, autoantibody production, and kidney disease in lupus-associated pathologies (Sharma et al., 2021).
In detail, a preclinical study reported the vasculo-protective action of the SOCS1 mimetic KIR peptide in a mouse model of elastase-induced AAA (Bernal et al., 2020). KIR peptide suppressed STAT1,3 activation in aorta, downregulated cytokines and metalloproteinases, altered the expression of cell differentiation markers, and reduced the incidence of AAA in aortic dilation and elastin degradation. Similar effects were encountered in atherosclerotic plaques of diabetic mice where the presence of KIR peptide significantly reduced the size of both early and aged lesions. In addition, the treatment with this SOCS1 mimetic reduced the accumulation of lipids, macrophages, and T lymphocytes and, conversely, determined an increase in the collagen and smooth muscle cell content. The KIR peptide demonstrated its efficacy in an experimental diabetes model with concomitant renal and macrovascular disease (streptozotocin-induced diabetic apolipoprotein E–deficient mouse). Indeed, this compound provided reno- and athero-protection in diabetic mice; in their kidney and aorta, the levels of superoxide anion, DNA oxidation marker, and Nox subunits were significantly lower, while antioxidant enzymes were highly expressed. These results agreed with a concomitant reduction of renal damage (decreased values of albuminuria, creatinine, and tubular injury), atherosclerosis (lesion size), and inflammation (leukocytes and chemokines) (Ortiz-Muñoz et al., 2009). The topical administration of KIR in diabetic retinopathy (DR) reduced glial activation and neural apoptosis induced by diabetes, as well as retinal levels of pro-inflammatory cytokines. These results occurred along with a significant amelioration of electroretinogram parameters, indicating a direct impact on the retina since it also prevented the disruption of the blood–retinal barrier, which was promising for the treatment of early stages of DR (Hernández et al., 2019). One decade ago, through an alanine scan of the KIR sequence of SOCS1, 52–67 fragments, a truncated linear peptide (62–61), named new KIR (Figure 2), demonstrated the ability to bind to JAK2 (Table 2). Subsequently, by carrying out a screening of “combinatorial focused libraries” in positional scanning (PS) format (Marasco et al., 2008; Lonardo et al., 2010; Ponticelli et al., 2008) of new KIR sequence, a new lead compound, named PS5, carrying the mutations His54/Cys(Acm), Phe55/Arg, and Arg56/Gln was investigated (Figure 2). It demonstrated the ability to bind to the JAK2 catalytic site more efficiently than KIR, as evident by the comparison of KD values reported in Table 2, and to drastically reduce STAT1,3 activation induced by IFN-γ and IL-6 and downstream genes (Doti et al., 2012; Madonna et al., 2013; La Manna et al., 2017b). In detail, PS5 effects were analyzed in keratinocytes, and explants of human skin were activated by IFN-γ that have a crucial role in type-1 skin disorders. The inhibition of the phosphorylation of JAK2, IFN-γRα, and STAT1 and the reductions in the gene expression of IRF-1 transcription factor, ICAM-1, HLA-DR, Cxcl10, and Ccl2 were observed. Consistently, the adhesiveness and migration of T lymphocytes to autologous keratinocytes were reduced (Madonna et al., 2013). Since both cell migration and proliferation concur to vascular remodeling during the formation of atherosclerotic plaques (Hu et al., 2019), PS5 reduced the migration and proliferation (“wound healing”) of VSMCs. In addition, for both VSMCs and RAW 264.7 cells, PS5 demonstrated antioxidant properties since its presence caused a reduction in intracellular O2•− production and the gene expressions of Nox1 and Nox4 and simultaneously increased the expression of superoxide dismutase 1 (Sod1) and catalase (Cat). PS5 also displayed antioxidant and athero-protective properties in a mouse model of atherosclerosis, corroborating the therapeutic perspective for its in vivo application. Indeed, PS5 administration to atherosclerotic mice induced a reduction in 1) the size and extension of atheroma plaques, 2) the intraplaque lipid content, and 3) the accumulation of monocytes/macrophages (La Manna et al., 2020b).
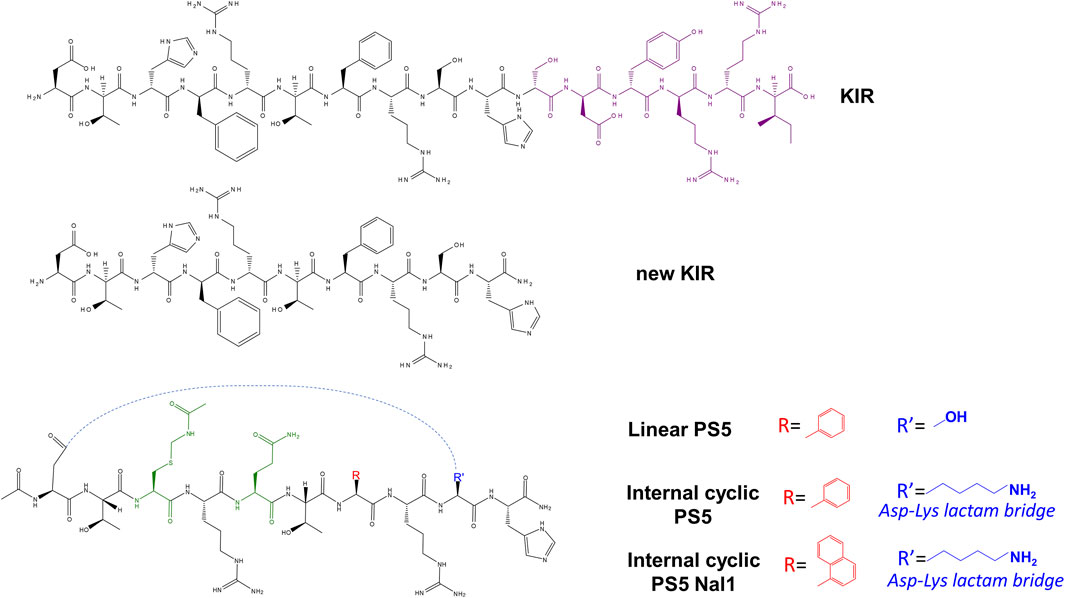
FIGURE 2. Chemical structures of KIR-based SOCS1 proteomimetics. Green represents non-native residues in PS5 upon combinatorial modifications (Cys(Acm) and Gln), blue represents the residue is alternatively Ser60 in PS5 and Lys in internal cycles PS5 for the formation of lactam bridge (with Asp52), and red represents the aromatic residue Nal1 or Phe56.

TABLE 2. SOCS1 proteomimetics: name, sequence, length, and dissociation constants values (µM) toward JAK2 catalytic domain.
In attempts to improve PS5 drug-like features, since no structural details were obtainable for its intrinsic flexibility, cyclic analog compounds were conceived by introducing 1) a disulfide bridge at the extremities (La Manna et al., 2017b), or 2) a lactam bond bridging the side chains of aspartic acid (naturally occurring at position 52) and a lysine residue introduced to substitute Ser60, named internal cyclic PS5 (Figure 2). To further enhance the proteolytic stability and the rigidity of new proteomimetics, a non-natural L-1-naphthylalanine in place of Phe58 was introduced— internal cyclic PS5 Nal1 compound (Figure 2) (La Manna et al., 2021).
Docking, molecular dynamics (MD), and in vitro binding assays suggested that even if van der Waals and electrostatic interactions mainly drive the recognition between JAK2 and internal PS5 cycles, the substitution of Phe/Nal1 allow a greater affinity toward protein target, conferring to aromatic interactions a crucial role for the formation of the complex as also confirmed by structural investigations, circular dichroism (CD), and nuclear magnetic resonance (NMR). In cellular contexts, such as VSMCs and macrophage cell line RAW 264.7, both PS5 cyclic analogs reduce STAT1 cellular migration at a longer extent with respect to Phe-internal cycle PS5, and this could have an impact on ongoing studies of their effects in plaque formation (Charo and Taub, 2011; Demina et al., 2021) as well as for antioxidant properties (La Manna et al., 2020b), confirmed by PCR analysis, which demonstrated a drastic suppression of NADPH oxidase genes and increase in Sod1 and Cat.
Conversely, a small synthetic peptide spanning the phosphorylation activation loop of JAK2, named pJAK2, demonstrated the ability to act as an antagonist of SOCSs. Indeed, in keratinocytes infected with HSV-1, it prevented mice from lethal doses of vaccinia, influenza A, and encephalomyocarditis virus infections (Subramaniam et al., 2004) (Huang et al., 2020). Moreover, during methicillin-resistant Staphylococcus aureus (MRSA) skin infection, SOCS1-deficient mice displayed reduced lesion size and bacterial loads and increased abscess thickness when compared to wild-type mice. The treatment of normal mice with pJAK2 increases phagocytosis and bacterial killing, as well as the levels of INFs, restoring skin host defense in hyperglycemic mice (Klopfenstein et al., 2021).
SOCS3, by forming a ternary complex, interacts simultaneously with the JAK2 kinase domain and glycoprotein 130 (gp130) phosphotyrosine peptide via two adjacent binding interfaces (Kershaw et al., 2013): on JAK2 Gly1071, Gln1072, and Met1073 (GQM motif) interaction with Leu22, Lys23, and Thr24 of KIR-SOCS3 and Val34, Val35, and Val38 of ESS-SOCS3 that is the most perturbed region upon complex formation (Babon et al., 2006). Additional contacts involve a “hinge” region between ESS and helix A (HA) of SH2, where the stretch 46–52, called CONG, bears three adjacent aromatic residues, FYW. In attempts to obtain SOCS3 proteomimetics, for the first time to the best of our knowledge, we analyzed, in three different studies, the ability of different linear peptides with variable lengths, reported in Table 3, all centered on the KIR domain, to recognize JAK2 and to mimic SOCS3 cellular effects. Initially, we analyzed KIR and ESS regions isolated and in combination: the entire KIRESS region exhibited good affinity for JAK2 and an efficient suppression of both IL-22 signaling in keratinocytes. In detail, the KIRESS region prevents STAT3 and Erk activation induced by IL-22 as well as the expression of STAT3-dependent downstream genes, dampening the proliferative and migratory potential of keratinocytes. In addition, in athymic nude mice bearing squamous cell carcinoma (SCC) xenograft, KIRESS concomitantly reduced the tumor growth and STAT3 levels (Madonna et al., 2017). KIRESS effects were also investigated in IL-6 signaling in mouse xenografts of MDA-MB-231-luci tumors as models of human triple negative breast cancer (TNBC) (La Manna et al., 2018b). The uptake of KIRESS, in 4T1 tumor-bearing BALB/c mice, was more efficient in primary tumor as well as in the bone marrow and spleen compared to non–tumor-bearing mice, allowing an easy access for SOCS3 mimetic to disseminated tumor cells in these organs. Indeed, NOD-SCID animals treated with KIRESS exhibited reduced tumor growth and the complete elimination of pulmonary metastasis as assessed by optical imaging in live mice and ex vivo images and histological examination of lungs. The specificity of its MOA was assessed through the evaluation of the SOCS3-pSTAT3-NF-kB pathway by Western blotting experiments. In them, KIRESS significantly reduced the phosphorylation of STAT3 and NF-kB (p65), as well as the expression of inflammatory cytokines including IL-6, leptin, and RANTES in a way similar to the whole SOCS3 protein allowing also its accumulation as consequence of the inhibition of the pSTAT3-NF-kB pathway (La Manna et al., 2018b). More recently, KIRESS peptide demonstrated its efficacy in pathological neovascularization, that is, the major cause of vision loss in the elderly neovascular age-related macular degeneration (nAMD) (Wang et al., 2021). Indeed, the SOCS3 pathway is strictly linked to neovascularization. In a mouse model with a laser-induced choroidal neovascularization (CNV), SOCS3 was significantly induced in myeloid lineage cells; its overexpression inhibited CNV, reduced myeloid lineage–derived macrophage/microglia recruitment onsite, and attenuated pro-inflammatory factor expression; similarly, the presence of KIRESS peptide reduced in vivo CNV.

TABLE 3. SOCS3 proteomimetics: name, sequence, length, and dissociation constants values (µM) toward JAK2 catalytic domain.
To explore the contribution derived from other SOCS3 regions, more recently the CONG region, was investigated in a chimeric peptide, named KIRCONG chim, containing non-contiguous fragments: a smaller region of KIR (25–33) and CONG (46–52) connected by β-alanines as spacers (Table 3). This proteomimetic showed a micromolar value of KD and a good α-helical conformation, as indicated by NMR and MD studies. Its anti-inflammatory properties were investigated in VSMC and RAW 264.7 macrophages. The presence of KIRCONG chim suppressed STAT3 phosphorylation and its translocation into the nuclei, as well as the expression of genes as Cxcl10 and Ccl5 chemokines and Nox2, a superoxide-generating enzyme. The observed anti-inflammatory and antioxidant properties of KIRCONG chim corroborate the potential application of SOCS3 mimetics in inflammatory diseases, including atherosclerosis, even if its low aqueous solubility and high molecular weights hamper their direct use as drugs. Ongoing structure−activity relationship (SAR) investigations through the introduction of ionizable moieties and of conformational constraints confer more suitable drug-like properties (unpublished results).
The development and clinical and preclinical application of small-molecule JAKis and cytokine-targeted biologics have demonstrated that the inhibition of the JAK-STAT pathway can be an efficacious way to treat inflammatory and autoimmune diseases. The identification of molecular mechanisms by which JAK-STAT signaling is regulated may also reveal opportunities to control signaling in new routes. A recent and interesting application regards the actual pandemic COVID-19 in which the SARS-CoV-2 infection triggers inflammation via the JAK-STAT pathway toward a cytokine storm (Di Natale et al., 2020b). JAKis like ruxolitinib, baricitinib, and tofacitinib were employed to reduce inflammation; several regulatory authorities have supported their use, and numerous clinical trials are in progress to prove their safety and efficacy (Satarker et al., 2021). While the therapeutic efficacy of these inhibitors is unquestionable, an important issue still to be answered is related to their best dosage and safety aspects to meet the requirements of the regulatory authorities. An advancement of JAKis in the drug market is expected on the basis of encouraging preclinical and clinical results. In detail, they are expected to find a role in the emerging therapeutic procedures as high-dose induction, low-dose maintenance, or combination therapies (Satarker et al., 2021). Noticeably, the onset of adverse effects and resistance to some of their treatments under chronic inflammatory conditions outline the need to develop more targeted therapeutics with novel MOAs to overcome these limitations. Indeed, a series of warnings on JAKis are hampered to reach the trial’s primary objectives, in terms of clearances and labeling restrictions (Reddy and Cohen, 2020).
In the last decades, the inhibition of PPIs represents a major challenge in chemical biology, and many drug discovery processes are focused on it. Often peptide-based proteomimetics reproduce the biophysical binding selectivity of large proteins in cellular contexts. Furthermore, there is great interest in developing selective protein kinase inhibitors by targeting different sites with respect to catalytic ones; these involve PPIs that, in general, cannot be addressed with small molecules. The great similarity of ATP-binding sites among kinases partially hampers the identification of selective ATP-competitive inhibitors (Rettenmaier et al., 2014). Conversely, the experimental 3D structure of protein complexes can act as unique templates to design novel and selective modulators of cellular pathways by mimicking protein interfaces. In the case of proteomimetics within the JAK-STAT pathway, they can also provide the required selectivity with respect to the inflammatory mediator that triggers inflammatory response (interleukins, interferons, etc.).
Several investigations of peptides targeting SOCSs or their interactors, conceived as mimetics or antagonists, are crucial starting points for the development of drugs for diseases in which these proteins play a role. Their investigations through biophysical and functional perspectives along with their structural and chemical modifications appear mandatory for the development of novel therapeutic agents.
DM contributed to conceptualization and writing—original draft preparation; SLM and IDB reviewed and edited the manuscript; and DM and SLM contributed to visualization.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
SLM was supported by AIRC fellowship for Italy.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adedeji Olulana, A. F., Soler, M. A., Lotteri, M., Vondracek, H., Casalis, L., Marasco, D., et al. (2021). Computational Evolution of Beta-2-Microglobulin Binding Peptides for Nanopatterned Surface Sensors. Ijms 22, 812. doi:10.3390/ijms22020812
Ahmed, C. M. I., Larkin, J., and Johnson, H. M. (2015). SOCS1 Mimetics and Antagonists: A Complementary Approach to Positive and Negative Regulation of Immune Function. Front. Immunol. 6, 183. doi:10.3389/fimmu.2015.00183
Ahmed, C. M., Massengill, M. T., Brown, E. E., Ildefonso, C. J., Johnson, H. M., and Lewin, A. S. (2018). A Cell Penetrating Peptide from SOCS-1 Prevents Ocular Damage in Experimental Autoimmune Uveitis. Exp. Eye Res. 177, 12–22. doi:10.1016/j.exer.2018.07.020
Akhtar, L. N., and Benveniste, E. N. (2011). Viral Exploitation of Host SOCS Protein Functions. J. Virol. 85, 1912–1921. doi:10.1128/jvi.01857-10
Alexander, W. S., and Hilton, D. J. (2004). The Role of Suppressors of Cytokine Signaling (SOCS) Proteins in Regulation of the Immune Response. Annu. Rev. Immunol. 22, 503–529. doi:10.1146/annurev.immunol.22.091003.090312
Alexander, W. S., Starr, R., Fenner, J. E., Scott, C. L., Handman, E., Sprigg, N. S., et al. (1999). SOCS1 Is a Critical Inhibitor of Interferon γ Signaling and Prevents the Potentially Fatal Neonatal Actions of This Cytokine. Cell 98, 597–608. doi:10.1016/s0092-8674(00)80047-1
Alston, C. I., and Dix, R. D. (2019). SOCS and Herpesviruses, with Emphasis on Cytomegalovirus Retinitis. Front. Immunol. 10, 732. doi:10.3389/fimmu.2019.00732
Babon, J. J., McManus, E. J., Yao, S., DeSouza, D. P., Mielke, L. A., Sprigg, N. S., et al. (2006). The Structure of SOCS3 Reveals the Basis of the Extended SH2 Domain Function and Identifies an Unstructured Insertion that Regulates Stability. Mol. Cell 22, 205–216. doi:10.1016/j.molcel.2006.03.024
Babon, J. J., Varghese, L. N., and Nicola, N. A. (2014). Inhibition of IL-6 Family Cytokines by SOCS3. Semin. Immunol. 26, 13–19. doi:10.1016/j.smim.2013.12.004
Barclay, J. L., Anderson, S. T., Waters, M. J., and Curlewis, J. D. (2009). SOCS3 as a Tumor Suppressor in Breast Cancer Cells, and its Regulation by PRL. Int. J. Cancer 124, 1756–1766. doi:10.1002/ijc.24172
Barnard, A., Long, K., Martin, H. L., Miles, J. A., Edwards, T. A., Tomlinson, D. C., et al. (2015). Selective and Potent Proteomimetic Inhibitors of Intracellular Protein-Protein Interactions. Angew. Chem. Int. Ed. 54, 2960–2965. doi:10.1002/anie.201410810
Barrett, T. J., Schlegel, M., Zhou, F., Gorenchtein, M., Bolstorff, J., Moore, K. J., et al. (2019). Platelet Regulation of Myeloid Suppressor of Cytokine Signaling 3 Accelerates Atherosclerosis. Sci. Transl Med. 11, 1. doi:10.1126/scitranslmed.aax0481
Bernal, S., Lopez-Sanz, L., Jimenez-Castilla, L., Prieto, I., Melgar, A., La Manna, S., et al. (2020). Protective Effect of Suppressor of Cytokine Signalling 1-based Therapy in Experimental Abdominal Aortic Aneurysm. Br. J. Pharmacol. 178, 564–581. doi:10.1111/bph.15330
Bharadwaj, U., Kasembeli, M. M., Robinson, P., and Tweardy, D. J. (2020). Targeting Janus Kinases and Signal Transducer and Activator of Transcription 3 to Treat Inflammation, Fibrosis, and Cancer: Rationale, Progress, and Caution. Pharmacol. Rev. 72, 486–526. doi:10.1124/pr.119.018440
Boengler, K., Hilfikerkleiner, D., Drexler, H., Heusch, G., and Schulz, R. (2008). The Myocardial JAK/STAT Pathway: from protection to Failure. Pharmacol. Ther. 120, 172–185. doi:10.1016/j.pharmthera.2008.08.002
Bousoik, E., and Montazeri Aliabadi, H. (2018). "Do We Know Jack" about JAK? A Closer Look at JAK/STAT Signaling Pathway. Front. Oncol. 8, 287. doi:10.3389/fonc.2018.00287
Brown, K. M., Day, J. P., Huston, E., Zimmermann, B., Hampel, K., Christian, F., et al. (2013). Phosphodiesterase-8A Binds to and Regulates Raf-1 Kinase. Proc. Natl. Acad. Sci. 110, E1533–E1542. doi:10.1073/pnas.1303004110
Burchill, M. A., Yang, J., Vogtenhuber, C., Blazar, B. R., and Farrar, M. A. (2007). IL-2 Receptor β-Dependent STAT5 Activation Is Required for the Development of Foxp3+ Regulatory T Cells. J. Immunol. 178, 280–290. doi:10.4049/jimmunol.178.1.280
Calabrese, V., Mallette, F. A., Deschênes-Simard, X., Ramanathan, S., Gagnon, J., Moores, A., et al. (2009). SOCS1 Links Cytokine Signaling to P53 and Senescence. Mol. Cell 36, 754–767. doi:10.1016/j.molcel.2009.09.044
Carotenuto, A., Auriemma, L., Merlino, F., Yousif, A. M., Marasco, D., Limatola, A., et al. (2014). Lead Optimization of P5U and Urantide: Discovery of Novel Potent Ligands at the Urotensin-II Receptor. J. Med. Chem. 57, 5965–5974. doi:10.1021/jm500218x
Causa, F., Della Moglie, R., Iaccino, E., Mimmi, S., Marasco, D., Scognamiglio, P. L., et al. (2013). Evolutionary Screening and Adsorption Behavior of Engineered M13 Bacteriophage and Derived Dodecapeptide for Selective Decoration of Gold Interfaces. J. Colloid Interf. Sci. 389, 220–229. doi:10.1016/j.jcis.2012.08.046
Charo, I. F., and Taub, R. (2011). Anti-inflammatory Therapeutics for the Treatment of Atherosclerosis. Nat. Rev. Drug Discov. 10, 365–376. doi:10.1038/nrd3444
Cheng, J. P. X., and Nichols, B. J. (2016). Caveolae: One Function or Many? Trends Cell Biol. 26, 177–189. doi:10.1016/j.tcb.2015.10.010
Chu, Q., Shen, D., He, L., Wang, H., Liu, C., and Zhang, W. (2017). Prognostic Significance of SOCS3 and its Biological Function in Colorectal Cancer. Gene 627, 114–122. doi:10.1016/j.gene.2017.06.013
Croker, B. A., Kiu, H., and Nicholson, S. E. (2008). SOCS Regulation of the JAK/STAT Signalling Pathway. Semin. Cell Dev. Biol. 19, 414–422. doi:10.1016/j.semcdb.2008.07.010
Davey, G. M., Starr, R., Cornish, A. L., Burghardt, J. T., Alexander, W. S., Carbone, F. R., et al. (2005). SOCS-1 Regulates IL-15-driven Homeostatic Proliferation of Antigen-Naive CD8 T Cells, Limiting Their Autoimmune Potential. J. Exp. Med. 202, 1099–1108. doi:10.1084/jem.20050003
de Araujo, A. D., Hoang, H. N., Kok, W. M., Diness, F., Gupta, P., Hill, T. A., et al. (2014). Comparative α-helicity of Cyclic Pentapeptides in Water. Angew. Chem. Int. Ed. Engl. 53, 6965–6969. doi:10.1002/anie.201310245
de Araujo, E. D., Keserű, G. M., Gunning, P. T., and Moriggl, R. (2020). Targeting STAT3 and STAT5 in Cancer. Multidisciplinary Digital Publishing Institute. doi:10.3390/cancers12082002
de Oliveira, M. B., Fook-Alves, V. L., Eugenio, A. I. P., Fernando, R. C., Sanson, L. F. G., de Carvalho, M. F., et al. (2017). Anti-myeloma Effects of Ruxolitinib Combined with Bortezomib and Lenalidomide: A Rationale for JAK/STAT Pathway Inhibition in Myeloma Patients. Cancer Lett. 403, 206–215. doi:10.1016/j.canlet.2017.06.016
Demina, E. P., Smutova, V., Pan, X., Fougerat, A., Guo, T., Zou, C., et al. (2021). Neuraminidases 1 and 3 Trigger Atherosclerosis by Desialylating Low-Density Lipoproteins and Increasing Their Uptake by Macrophages. J. Am. Heart Assoc. 10, e018756. doi:10.1161/JAHA.120.018756
Dhillon, S. (2017). Tofacitinib: A Review in Rheumatoid Arthritis. Drugs 77, 1987–2001. doi:10.1007/s40265-017-0835-9
Di Natale, C., La Manna, S., Avitabile, C., Florio, D., Morelli, G., Netti, P. A., et al. (2020). Engineered β-hairpin Scaffolds from Human Prion Protein Regions: Structural and Functional Investigations of Aggregates. Bioorg. Chem. 96, 103594. doi:10.1016/j.bioorg.2020.103594
Di Natale, C., La Manna, S., De Benedictis, I., Brandi, P., and Marasco, D. (2020). Perspectives in Peptide-Based Vaccination Strategies for Syndrome Coronavirus 2 Pandemic. Front. Pharmacol. 11, 578382. doi:10.3389/fphar.2020.578382
Doti, N., Scognamiglio, P. L., Madonna, S., Scarponi, C., Ruvo, M., Perretta, G., et al. (2012). New Mimetic Peptides of the Kinase-Inhibitory Region (KIR) of SOCS1 through Focused Peptide Libraries. Biochem. J. 443, 231–240. doi:10.1042/bj20111647
Durham, G. A., Williams, J. J. L., Nasim, M. T., and Palmer, T. M. (2019). Targeting SOCS Proteins to Control JAK-STAT Signalling in Disease. Trends Pharmacol. Sci. 40, 298–308. doi:10.1016/j.tips.2019.03.001
Dymock, B. W., Yang, E. G., Chu-Farseeva, Y., and Yao, L. (2014). Selective JAK Inhibitors. Future Med. Chem. 6, 1439–1471. doi:10.4155/fmc.14.92
Eyles, J. L., Metcalf, D., Grusby, M. J., Hilton, D. J., and Starr, R. (2002). Negative Regulation of Interleukin-12 Signaling by Suppressor of Cytokine Signaling-1. J. Biol. Chem. 277, 43735–43740. doi:10.1074/jbc.m208586200
Favoino, E., Prete, M., Catacchio, G., Ruscitti, P., Navarini, L., Giacomelli, R., et al. (2021). Working and Safety Profiles of JAK/STAT Signaling Inhibitors. Are These Small Molecules Also Smart? Autoimmun. Rev. 20, 102750. doi:10.1016/j.autrev.2021.102750
Feng, Y., Wang, Q., and Wang, T. (2017). Drug Target Protein-Protein Interaction Networks: a Systematic Perspective. Biomed. Research International 2017, 1–13. doi:10.1155/2017/1289259
Fenner, J. E., Starr, R., Cornish, A. L., Zhang, J.-G., Metcalf, D., Schreiber, R. D., et al. (2006). Suppressor of Cytokine Signaling 1 Regulates the Immune Response to Infection by a Unique Inhibition of Type I Interferon Activity. Nat. Immunol. 7, 33–39. doi:10.1038/ni1287
Fenyo, I. M., Florea, I. C., Raicu, M., and Manea, A. (2011). Tyrphostin AG490 Reduces NAPDH Oxidase Activity and Expression in the Aorta of Hypercholesterolemic Apolipoprotein E-Deficient Mice. Vasc. Pharmacol. 54, 100–106. doi:10.1016/j.vph.2011.03.006
Fiocco, U., Accordi, B., Martini, V., Oliviero, F., Facco, M., Cabrelle, A., et al. (2014). JAK/STAT/PKCδ Molecular Pathways in Synovial Fluid T Lymphocytes Reflect the In Vivo T Helper-17 Expansion in Psoriatic Arthritis. Immunol. Res. 58, 61–69. doi:10.1007/s12026-013-8481-0
Flanagan, M. E., Blumenkopf, T. A., Brissette, W. H., Brown, M. F., Casavant, J. M., Shang-Poa, C., et al. (2010). Discovery of CP-690,550: a Potent and Selective Janus Kinase (JAK) Inhibitor for the Treatment of Autoimmune Diseases and Organ Transplant Rejection. J. Med. Chem. 53, 8468–8484. doi:10.1021/jm1004286
Gadina, M., Johnson, C., Schwartz, D., Bonelli, M., Hasni, S., Kanno, Y., et al. (2018). Translational and Clinical Advances in JAK-STAT Biology: The Present and Future of Jakinibs. J. Leukoc. Biol. 104, 499–514. doi:10.1002/jlb.5ri0218-084r
Gan, A. M., Pirvulescu, M. M., Stan, D., Simion, V., Calin, M., Manduteanu, I., et al. (2013). Monocytes and Smooth Muscle Cells Cross-Talk Activates STAT3 and Induces Resistin and Reactive Oxygen Species and Production. J. Cel. Biochem. 114, 2273–2283. doi:10.1002/jcb.24571
Gao, Y.-F., Li, T., Chang, Y., Wang, Y.-B., Zhang, W.-N., Li, W.-H., et al. (2011). Cdk1-phosphorylated CUEDC2 Promotes Spindle Checkpoint Inactivation and Chromosomal Instability. Nat. Cell Biol 13, 924–933. doi:10.1038/ncb2287
Gharavi, N. M., Alva, J. A., Mouillesseaux, K. P., Lai, C., Yeh, M., Yeung, W., et al. (2007). Role of the Jak/STAT Pathway in the Regulation of Interleukin-8 Transcription by Oxidized Phospholipids In Vitro and in Atherosclerosis In Vivo. J. Biol. Chem. 282, 31460–31468. doi:10.1074/jbc.m704267200
Ghoreschi, K., Laurence, A., and O’Shea, J. J. (2009). Janus Kinases in Immune Cell Signaling. Immunological Rev. 228, 273–287. doi:10.1111/j.1600-065x.2008.00754.x
Gigante, A., Sijbesma, E., Sánchez‐Murcia, P. A., Hu, X., Bier, D., Bäcker, S., et al. (2020). A Supramolecular Stabilizer of the 14‐3‐3ζ/ERα Protein‐Protein Interaction with a Synergistic Mode of Action. Angew. Chem. Int. Ed. 59, 5284–5287. doi:10.1002/anie.201914517
Giordanetto, F., and Kroemer, R. T. (2003). A Three-Dimensional Model of Suppressor of Cytokine Signalling 1 (SOCS-1). Protein Eng. 16, 115–124. doi:10.1093/proeng/gzg015
Goropevšek, A., Holcar, M., and Avčin, T. (2017). The Role of STAT Signaling Pathways in the Pathogenesis of Systemic Lupus Erythematosus. Clin. Rev. Allergy Immunol. 52, 164–181.
Groner, B., and von Manstein, V. (2017). Jak Stat Signaling and Cancer: Opportunities, Benefits and Side Effects of Targeted Inhibition. Mol. Cell Endocrinol. 451, 1–14. doi:10.1016/j.mce.2017.05.033
Hadjadj, J., Castro, C. N., Tusseau, M., Stolzenberg, M.-C., Mazerolles, F., Aladjidi, N., et al. (2020). Early-onset Autoimmunity Associated with SOCS1 Haploinsufficiency. Nat. Commun. 11, 5341. doi:10.1038/s41467-020-18925-4
Harris, C., and Cummings, J. R. F. (2021). JAK1 Inhibition and Inflammatory Bowel Disease. Rheumatology 60, ii45–ii51. doi:10.1093/rheumatology/keaa896
He, C., Yu, C. R., Mattapallil, M. J., Sun, L., Larkin Iii, J., and Egwuagu, C. E. (2016). SOCS1 Mimetic Peptide Suppresses Chronic Intraocular Inflammatory Disease (Uveitis). Mediators Inflamm. 2016, 2939370. doi:10.1155/2016/2939370
Heppler, L. N., and Frank, D. A. (2017). Targeting Oncogenic Transcription Factors: Therapeutic Implications of Endogenous STAT Inhibitors. Trends Cancer 3, 816–827. doi:10.1016/j.trecan.2017.10.004
Hernández, C., Bogdanov, P., Gómez-Guerrero, C., Sampedro, J., Solà-Adell, C., Espejo, C., et al. (2019). SOCS1-Derived Peptide Administered by Eye Drops Prevents Retinal Neuroinflammation and Vascular Leakage in Experimental Diabetes. Int. J. Mol. Sci. 20, 1. doi:10.3390/ijms20153615
Hernández-Vargas, P., López-Franco, O., Sanjuán, G., Rupérez, M., Ortiz-Muñoz, G., Suzuki, Y., et al. (2005). Suppressors of Cytokine Signaling Regulate Angiotensin II-Activated Janus Kinase-Signal Transducers and Activators of Transcription Pathway in Renal Cells. Jasn 16, 1673–1683. doi:10.1681/asn.2004050374
Hill, G. R., Kuns, R. D., Raffelt, N. C., Don, A. L. J., Olver, S. D., Markey, K. A., et al. (2010). SOCS3 Regulates Graft-Versus-Host Disease. Blood 116, 287–296. doi:10.1182/blood-2009-12-259598
Hu, D., Yin, C., Luo, S., Habenicht, A. J. R., and Mohanta, S. K. (2019). Vascular Smooth Muscle Cells Contribute to Atherosclerosis Immunity. Front. Immunol. 10, 1101. doi:10.3389/fimmu.2019.01101
Huang, S., Liu, K., Cheng, A., Wang, M., Cui, M., Huang, J., et al. (2020). SOCS Proteins Participate in the Regulation of Innate Immune Response Caused by Viruses. Front. Immunol. 11, 558341. doi:10.3389/fimmu.2020.558341
Hurwitz, H., Van Cutsem, E., Bendell, J., Hidalgo, M., Li, C.-P., Salvo, M. G., et al. (2018). Ruxolitinib + Capecitabine in Advanced/metastatic Pancreatic Cancer after Disease Progression/intolerance to First-Line Therapy: JANUS 1 and 2 Randomized Phase III Studies. Invest. New Drugs 36, 683–695. doi:10.1007/s10637-018-0580-2
Hymel, D., Tsuji, K., Grant, R. A., Chingle, R. M., Kunciw, D. L., Yaffe, M. B., et al. (2021). Design and Synthesis of a New Orthogonally Protected Glutamic Acid Analog and its Use in the Preparation of High Affinity polo-like Kinase 1 polo-box Domain - Binding Peptide Macrocycles. Org. Biomol. Chem. 19, 7843–7854. doi:10.1039/d1ob01120k
Ihle, J. N., Witthuhn, B. A., Quelle, F. W., Yamamoto, K., and Silvennoinen, O. (1995). Signaling through the Hematopoietic Cytokine Receptors. Annu. Rev. Immunol. 13, 369–398. doi:10.1146/annurev.iy.13.040195.002101
Ilangumaran, S., Ramanathan, S., and Rottapel, R. (2004). Regulation of the Immune System by SOCS Family Adaptor Proteins. Semin. Immunol. 16, 351–365. doi:10.1016/j.smim.2004.08.015
Isomoto, H. (2009). Epigenetic Alterations in Cholangiocarcinoma-Sustained IL-6/STAT3 Signaling in Cholangio- Carcinoma Due to SOCS3 Epigenetic Silencing. Digestion 79 (Suppl. 1), 2–8. doi:10.1159/000167859
Jamilloux, Y., El Jammal, T., Vuitton, L., Gerfaud-Valentin, M., Kerever, S., and Sève, P. (2019). JAK Inhibitors for the Treatment of Autoimmune and Inflammatory Diseases. Autoimmun. Rev. 18, 102390. doi:10.1016/j.autrev.2019.102390
Jo, D., Liu, D., Yao, S., Collins, R. D., and Hawiger, J. (2005). Intracellular Protein Therapy with SOCS3 Inhibits Inflammation and Apoptosis. Nat. Med. 11, 892–898. doi:10.1038/nm1269
Johnson, H. M., Lewin, A. S., and Ahmed, C. M. (2020). SOCS, Intrinsic Virulence Factors, and Treatment of COVID-19. Front. Immunol. 11, 582102. doi:10.3389/fimmu.2020.582102
Jung, S.-H., Kim, S.-M., and Lee, C.-E. (2016). Mechanism of Suppressors of Cytokine Signaling 1 Inhibition of Epithelial-Mesenchymal Transition Signaling through ROS Regulation in colon Cancer Cells: Suppression of Src Leading to Thioredoxin Up-Regulation. Oncotarget 7, 62559–62571. doi:10.18632/oncotarget.11537
Kahn, J. S., Deverapalli, S. C., and Rosmarin, D. M. (2018). JAK-STAT Signaling Pathway Inhibition: a Role for Treatment of Discoid Lupus Erythematosus and Dermatomyositis. Int. J. Dermatol. 57, 1007–1014. doi:10.1111/ijd.14064
Kershaw, N. J., Murphy, J. M., Liau, N. P. D., Varghese, L. N., Laktyushin, A., Whitlock, E. L., et al. (2013). SOCS3 Binds Specific Receptor-JAK Complexes to Control Cytokine Signaling by Direct Kinase Inhibition. Nat. Struct. Mol. Biol. 20, 469–476. doi:10.1038/nsmb.2519
Kim, K.-K., Siddiqui, Z., Patel, M., Sarkar, B., and Kumar, V. A. (2020). A Self-Assembled Peptide Hydrogel for Cytokine Sequestration. J. Mater. Chem. B 8, 945–950. doi:10.1039/c9tb02250c
Kipp, M., Göhring, F., Ostendorp, T., van Drunen, C. M., van Driel, R., Przybylski, M., et al. (2000). SAF-box, a Conserved Protein Domain that Specifically Recognizes Scaffold Attachment Region DNA. Mol. Cell Biol 20, 7480–7489. doi:10.1128/mcb.20.20.7480-7489.2000
Klopfenstein, N., Brandt, S. L., Castellanos, S., Gunzer, M., Blackman, A., and Serezani, C. H. (2021). SOCS-1 Inhibition of Type I Interferon Restrains Staphylococcus aureus Skin Host Defense. Plos Pathog. 17, e1009387. doi:10.1371/journal.ppat.1009387
Kwatra, S. G., Dabade, T. S., Gustafson, C. J., and Feldman, S. R. (2012). JAK Inhibitors in Psoriasis: a Promising New Treatment Modality. J. Drugs Dermatol. 11, 913–918.
La Manna, S., Lopez-Sanz, L., Bernal, S., Jimenez-Castilla, L., Prieto, I., Morelli, G., et al. (2020). Antioxidant Effects of PS5, a Peptidomimetic of Suppressor of Cytokine Signaling 1, in Experimental Atherosclerosis. Antioxidants (Basel) 9, 1. doi:10.3390/antiox9080754
La Manna, S., Di Natale, C., Florio, D., and Marasco, D. (2018). Peptides as Therapeutic Agents for Inflammatory-Related Diseases. Ijms 19, 2714. doi:10.3390/ijms19092714
La Manna, S., Lee, E., Ouzounova, M., Di Natale, C., Novellino, E., Merlino, A., et al. (2018). Mimetics of Suppressor of Cytokine Signaling 3: Novel Potential Therapeutics in Triple Breast Cancer. Int. J. Cancer 143, 2177–2186. doi:10.1002/ijc.31594
La Manna, S., Lopez-Sanz, L., Bernal, S., Fortuna, S., Mercurio, F. A., Leone, M., et al. (2021). Cyclic Mimetics of Kinase-Inhibitory Region of Suppressors of Cytokine Signaling 1: Progress toward Novel Anti-inflammatory Therapeutics. Eur. J. Med. Chem. 221, 113547. doi:10.1016/j.ejmech.2021.113547
La Manna, S., Lopez-Sanz, L., Leone, M., Brandi, P., Scognamiglio, P. L., Morelli, G., et al. (2017). Structure-activity Studies of Peptidomimetics Based on Kinase-Inhibitory Region of Suppressors of Cytokine Signaling 1. Biopolymers. doi:10.1002/bip.23082
La Manna, S., Lopez-Sanz, L., Mercurio, F. A., Fortuna, S., Leone, M., Gomez-Guerrero, C., et al. (2020). Chimeric Peptidomimetics of SOCS 3 Able to Interact with JAK2 as Anti-inflammatory Compounds. ACS Med. Chem. Lett. 11, 615–623. doi:10.1021/acsmedchemlett.9b00664
La Manna, S., Scognamiglio, P. L., Di Natale, C., Leone, M., Mercurio, F. A., Malfitano, A. M., et al. (2017). Characterization of Linear Mimetic Peptides of Interleukin-22 from Dissection of Protein Interfaces. Biochimie 138, 106–115. doi:10.1016/j.biochi.2017.05.002
Larkin, J., Polk, T., Sharma, J., Bae, S., Barr, O., Jones, A., et al. (2020). Mitigation of Equine Recurrent Uveitis through Topical Suppressor of Cytokine Signaling-1 (SOCS1) Mimetic Peptide: Open Label Safety and Efficacy Pilot Study. Am Assoc Immnol.
Lee, E. B., Fleischmann, R., Hall, S., Wilkinson, B., Bradley, J. D., Gruben, D., et al. (2014). Tofacitinib versus Methotrexate in Rheumatoid Arthritis. N. Engl. J. Med. 370, 2377–2386. doi:10.1056/nejmoa1310476
Liau, N. P. D., Laktyushin, A., Lucet, I. S., Murphy, J. M., Yao, S., Whitlock, E., et al. (2018). The Molecular Basis of JAK/STAT Inhibition by SOCS1. Nat. Commun. 9, 1558. doi:10.1038/s41467-018-04013-1
Lim, W.-S., Timmins, J. M., Seimon, T. A., Sadler, A., Kolodgie, F. D., Virmani, R., et al. (2008). Signal Transducer and Activator of Transcription-1 Is Critical for Apoptosis in Macrophages Subjected to Endoplasmic Reticulum Stress In Vitro and in Advanced Atherosclerotic Lesions In Vivo. Circulation 117, 940–951. doi:10.1161/circulationaha.107.711275
Lingard, H., Han, J. T., Thompson, A. L., Leung, I. K. H., Scott, R. T. W., Thompson, S., et al. (2014). Diphenylacetylene-Linked Peptide Strands Induce Bidirectional β-Sheet Formation. Angew. Chem. Int. Ed. 53, 3650–3653. doi:10.1002/anie.201309353
Linossi, E. M., Babon, J. J., Hilton, D. J., and Nicholson, S. E. (2013). Suppression of Cytokine Signaling: the SOCS Perspective. Cytokine Growth Factor. Rev. 24, 241–248. doi:10.1016/j.cytogfr.2013.03.005
Loh, C.-Y., Arya, A., Naema, A. F., Wong, W. F., Sethi, G., and Looi, C. Y. (2019). Signal Transducer and Activator of Transcription (STATs) Proteins in Cancer and Inflammation: Functions and Therapeutic Implication. Front. Oncol. 9, 48. doi:10.3389/fonc.2019.00048
Lonardo, E., Parish, C. L., Ponticelli, S., Marasco, D., Ribeiro, D., Ruvo, M., et al. (2010). A Small Synthetic Cripto Blocking Peptide Improves Neural Induction, Dopaminergic Differentiation, and Functional Integration of Mouse Embryonic Stem Cells in a Rat Model of Parkinson's Disease. Stem cells 28, 1326–1337. doi:10.1002/stem.458
Lopez-Sanz, L., Bernal, S., Recio, C., Lazaro, I., Oguiza, A., Melgar, A., et al. (2018). SOCS1-targeted Therapy Ameliorates Renal and Vascular Oxidative Stress in Diabetes via STAT1 and PI3K Inhibition. Lab. Invest. 98, 1276–1290. doi:10.1038/s41374-018-0043-6
Madonna, S., Scarponi, C., De Pità, O., and Albanesi, C. (2008). Suppressor of Cytokine Signaling 1 Inhibits IFN‐γ Inflammatory Signaling in Human Keratinocytes by Sustaining ERK1/2 Activation. FASEB j. 22, 3287–3297. doi:10.1096/fj.08-106831
Madonna, S., Scarponi, C., Doti, N., Carbone, T., Cavani, A., Scognamiglio, P. L., et al. (2013). Therapeutical Potential of a Peptide Mimicking the SOCS1 Kinase Inhibitory Region in Skin Immune Responses. Eur. J. Immunol. 43, 1883–1895. doi:10.1002/eji.201343370
Madonna, S., Scarponi, C., Morelli, M., Sestito, R., Scognamiglio, P. L., Marasco, D., et al. (2017). SOCS3 Inhibits the Pathological Effects of IL-22 in Non-melanoma Skin Tumor-Derived Keratinocytes. Oncotarget 8, 24652–24667. doi:10.18632/oncotarget.15629
Mahony, R., Ahmed, S., Diskin, C., and Stevenson, N. J. (2016). SOCS3 Revisited: a Broad Regulator of Disease, Now Ready for Therapeutic Use? Cell. Mol. Life Sci. 73, 3323–3336. doi:10.1007/s00018-016-2234-x
Malemud, C. J. (2018). The Role of the JAK/STAT Signal Pathway in Rheumatoid Arthritis. Ther. Adv. Musculoskelet. 10, 117–127. doi:10.1177/1759720x18776224
Malemud, C. (2016). Suppressor of Cytokine Signaling and Rheumatoid Arthritis. Integr. Mol. Med. 3, 17–20.
Manea, A., Tanase, L. I., Raicu, M., and Simionescu, M. (2010). Jak/STAT Signaling Pathway Regulates Nox1 and Nox4-Based NADPH Oxidase in Human Aortic Smooth Muscle Cells. Atvb 30, 105–112. doi:10.1161/atvbaha.109.193896
Manea, S.-A., Manea, A., and Heltianu, C. (2010). Inhibition of JAK/STAT Signaling Pathway Prevents High-Glucose-Induced Increase in Endothelin-1 Synthesis in Human Endothelial Cells. Cell Tissue Res 340, 71–79. doi:10.1007/s00441-010-0936-1
Marasco, D., Perretta, G., Sabatella, M., and Ruvo, M. (2008). Past and Future Perspectives of Synthetic Peptide Libraries. Cpps 9, 447–467. doi:10.2174/138920308785915209
Mease, P., Coates, L. C., Helliwell, P. S., Stanislavchuk, M., Rychlewska-Hanczewska, A., Dudek, A., et al. (2018). Efficacy and Safety of Filgotinib, a Selective Janus Kinase 1 Inhibitor, in Patients with Active Psoriatic Arthritis (EQUATOR): Results from a Randomised, Placebo-Controlled, Phase 2 Trial. The Lancet 392, 2367–2377. doi:10.1016/s0140-6736(18)32483-8
Miklossy, G., Hilliard, T. S., and Turkson, J. (2013). Therapeutic Modulators of STAT Signalling for Human Diseases. Nat. Rev. Drug Discov. 12, 611–629. doi:10.1038/nrd4088
Milroy, L.-G., Grossmann, T. N., Hennig, S., Brunsveld, L., and Ottmann, C. (2014). Modulators of Protein-Protein Interactions. Chem. Rev. 114, 4695–4748. doi:10.1021/cr400698c
Mohan, C. D., Rangappa, S., Preetham, H. D., Nayaka, S. C., Gupta, V. K., Basappa, S., et al. (2020). Targeting STAT3 Signaling Pathway in Cancer by Agents Derived from Mother Nature, Seminars in Cancer Biology. Elsevier, 1.
Nascimento, A. S., Peres, L. L., Fari, A. V. S., Milani, R., Silva, R. A., da Costa Fernandes, C. J., et al. (2017). Phosphoproteome Profiling Reveals Critical Role of JAK-STAT Signaling in Maintaining Chemoresistance in Breast Cancer. Oncotarget 8, 114756–114768. doi:10.18632/oncotarget.21801
Nicholson, S. E., Willson, T. A., Farley, A., Starr, R., Zhang, J. G., Baca, M., et al. (1999). Mutational Analyses of the SOCS Proteins Suggest a Dual Domain Requirement but Distinct Mechanisms for Inhibition of LIF and IL-6 Signal Transduction. Embo J. 18, 375–385. doi:10.1093/emboj/18.2.375
Niu, G.-J., Xu, J.-D., Yuan, W.-J., Sun, J.-J., Yang, M.-C., He, Z.-H., et al. (2018). Protein Inhibitor of Activated STAT (PIAS) Negatively Regulates the JAK/STAT Pathway by Inhibiting STAT Phosphorylation and Translocation. Front. Immunol. 9, 2392. doi:10.3389/fimmu.2018.02392
O'Shaughnessy, J., DeMichele, A., Ma, C. X., Richards, P., Yardley, D. A., Wright, G. S., et al. (2018). A Randomized, Double-Blind, Phase 2 Study of Ruxolitinib or Placebo in Combination with Capecitabine in Patients with Advanced HER2-Negative Breast Cancer and Elevated C-Reactive Protein, a Marker of Systemic Inflammation. Breast Cancer Res. Treat. 170, 547–557. doi:10.1007/s10549-018-4770-6
O'Shea, J. J., Schwartz, D. M., Villarino, A. V., Gadina, M., McInnes, I. B., and Laurence, A. (2015). The JAK-STAT Pathway: Impact on Human Disease and Therapeutic Intervention. Annu. Rev. Med. 66, 311–328. doi:10.1146/annurev-med-051113-024537
Opazo-Ríos, L., Sanchez Matus, Y., Rodrigues-Díez, R. R., Carpio, D., Droguett, A., Egido, J., et al. (2020). Anti-inflammatory, Antioxidant and Renoprotective Effects of SOCS1 Mimetic Peptide in the BTBR Ob/ob Mouse Model of Type 2 Diabetes. BMJ Open Diabetes Res. Care 8, 1. doi:10.1136/bmjdrc-2020-001242
Orr, S. J., Morgan, N. M., Elliott, J., Burrows, J. F., Scott, C. J., McVicar, D. W., et al. (2007). CD33 Responses Are Blocked by SOCS3 through Accelerated Proteasomal-Mediated Turnover. Blood 109, 1061–1068. doi:10.1182/blood-2006-05-023556
Ortiz-Muñoz, G., Martin-Ventura, J. L., Hernandez-Vargas, P., Mallavia, B., Lopez-Parra, V., Lopez-Franco, O., et al. (2009). Suppressors of Cytokine Signaling Modulate JAK/STAT-mediated Cell Responses during Atherosclerosis. Arterioscler Thromb. Vasc. Biol. 29, 525–531. doi:10.1161/ATVBAHA.108.173781
Ortiz-Muñoz, G., Lopez-Parra, V., Lopez-Franco, O., Fernandez-Vizarra, P., Mallavia, B., Flores, C., et al. (2010). Suppressors of Cytokine Signaling Abrogate Diabetic Nephropathy. Jasn 21, 763–772. doi:10.1681/asn.2009060625
Ou, A., Ott, M., Fang, D., and Heimberger, A. B. (2021). The Role and Therapeutic Targeting of JAK/STAT Signaling in Glioblastoma. Cancers (Basel) 13, 1. doi:10.3390/cancers13030437
Owen, K. L., Brockwell, N. K., and Parker, B. S. (2019). JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers (Basel) 11, 1. doi:10.3390/cancers11122002
Park, J. S., Hong, M. H., Chun, Y. J., Kim, H. R., and Cho, B. C. (2019). A Phase Ib Study of the Combination of Afatinib and Ruxolitinib in EGFR Mutant NSCLC with Progression on EGFR-TKIs. Lung Cancer 134, 46–51. doi:10.1016/j.lungcan.2019.05.030
Piranlioglu, R., Lee, E., Ouzounova, M., Bollag, R. J., Vinyard, A. H., Arbab, A. S., et al. (2019). Primary Tumor-Induced Immunity Eradicates Disseminated Tumor Cells in Syngeneic Mouse Model. Nat. Commun. 10, 1–13. doi:10.1038/s41467-019-09015-1
Ponticelli, S., Marasco, D., Tarallo, V., Albuquerque, R. J. C., Mitola, S., Takeda, A., et al. (2008). Modulation of Angiogenesis by a Tetrameric Tripeptide that Antagonizes Vascular Endothelial Growth Factor Receptor 1. J. Biol. Chem. 283, 34250–34259. doi:10.1074/jbc.m806607200
Prigge, J. R., and Schmidt, E. E. (2006). Interaction of Protein Inhibitor of Activated STAT (PIAS) Proteins with the TATA-Binding Protein, TBP. J. Biol. Chem. 281, 12260–12269. doi:10.1074/jbc.m510835200
Qin, L., Huang, Q., Zhang, H., Liu, R., Tellides, G., Min, W., et al. (2014). SOCS1 Prevents Graft Arteriosclerosis by Preserving Endothelial Cell Function. J. Am. Coll. Cardiol. 63, 21–29. doi:10.1016/j.jacc.2013.08.694
Qureshy, Z., Johnson, D. E., and Grandis, J. R. (2020). Targeting the JAK/STAT Pathway in Solid Tumors. J. Cancer Metastasis Treat. 6, 1. doi:10.20517/2394-4722.2020.58
Rawlings, J. S., Rosler, K. M., and Harrison, D. A. (2004). The JAK/STAT Signaling Pathway. J. Cell Sci 117, 1281–1283. doi:10.1242/jcs.00963
Recio, C., Lazaro, I., Oguiza, A., Lopez-Sanz, L., Bernal, S., Blanco, J., et al. (2017). Suppressor of Cytokine Signaling-1 Peptidomimetic Limits Progression of Diabetic Nephropathy. Jasn 28, 575–585. doi:10.1681/asn.2016020237
Recio, C., Oguiza, A., Lazaro, I., Mallavia, B., Egido, J., and Gomez-Guerrero, C. (2014). Suppressor of Cytokine Signaling 1-derived Peptide Inhibits Janus Kinase/signal Transducers and Activators of Transcription Pathway and Improves Inflammation and Atherosclerosis in Diabetic Mice. Atvb 34, 1953–1960. doi:10.1161/atvbaha.114.304144
Recio, C., Oguiza, A., Mallavia, B., Lazaro, I., Ortiz-Muñoz, G., Lopez-Franco, O., et al. (2015). Gene Delivery of Suppressors of Cytokine Signaling (SOCS) Inhibits Inflammation and Atherosclerosis Development in Mice. Basic Res. Cardiol. 110, 8. doi:10.1007/s00395-014-0458-1
Reddy, V., and Cohen, S. (2020). Jak Inhibitors: What Is New? Curr. Rheumatol. Rep. 22, 50–10. doi:10.1007/s11926-020-00931-6
Rettenmaier, T. J., Sadowsky, J. D., Thomsen, N. D., Chen, S. C., Doak, A. K., Arkin, M. R., et al. (2014). A Small-Molecule Mimic of a Peptide Docking Motif Inhibits the Protein Kinase PDK1. Proc. Natl. Acad. Sci. USA 111, 18590–18595. doi:10.1073/pnas.1415365112
Roger, I., Milara, J., Montero, P., and Cortijo, J. (2021). The Role of JAK/STAT Molecular Pathway in Vascular Remodeling Associated with Pulmonary Hypertension. Int. J. Mol. Sci. 22, 1. doi:10.3390/ijms22094980
Rogler, G. (2020). Efficacy of JAK Inhibitors in Crohn's Disease. J. Crohns Colitis 14, S746–S754. doi:10.1093/ecco-jcc/jjz186
Russo, A., Aiello, C., Grieco, P., and Marasco, D. (2016). Targeting "Undruggable" Proteins: Design of Synthetic Cyclopeptides. Cmc 23, 748–762. doi:10.2174/0929867323666160112122540
Russo, A., Scognamiglio, P. L., Hong Enriquez, R. P., Santambrogio, C., Grandori, R., Marasco, D., et al. (2015). In Silico generation of Peptides by Replica Exchange Monte Carlo: Docking-Based Optimization of Maltose-Binding-Protein Ligands. PLoS One 10, e0133571. doi:10.1371/journal.pone.0133571
Saint-Germain, E., Mignacca, L., Huot, G., Acevedo, M., Moineau-Vallée, K., Calabrese, V., et al. (2019). Phosphorylation of SOCS1 Inhibits the SOCS1-P53 Tumor Suppressor Axis. Cancer Res. 79, 3306–3319. doi:10.1158/0008-5472.can-18-1503
Salas, A., Hernandez-Rocha, C., Duijvestein, M., Faubion, W., McGovern, D., Vermeire, S., et al. (2020). JAK-STAT Pathway Targeting for the Treatment of Inflammatory Bowel Disease. Nat. Rev. Gastroenterol. Hepatol. 17, 323–337. doi:10.1038/s41575-020-0273-0
Sallam, A. M., Esmat, A., and Abdel-Naim, A. B. (2018). Cucurbitacin-B Attenuates CCl4 -induced Hepatic Fibrosis in Mice through Inhibition of STAT-3. Chem. Biol. Drug Des. 91, 933–941. doi:10.1111/cbdd.13160
Satarker, S., Tom, A. A., Shaji, R. A., Alosious, A., Luvis, M., and Nampoothiri, M. (2021). JAK-STAT Pathway Inhibition and Their Implications in COVID-19 Therapy. Postgrad. Med. 133, 489–507. doi:10.1080/00325481.2020.1855921
Sawyer, N., Watkins, A. M., and Arora, P. S. (2017). Protein Domain Mimics as Modulators of Protein-Protein Interactions. Acc. Chem. Res. 50, 1313–1322. doi:10.1021/acs.accounts.7b00130
Schuett, J., Kreutz, J., Grote, K., Vlacil, A. K., Schuett, H., Oberoi, R., et al. (2019). Suppressor of Cytokine Signaling 1 Is Involved in Gene Regulation Which Controls the Survival of Ly6Clow Monocytes in Mice. Cell Physiol Biochem 52, 336–353. doi:10.33594/000000024
Schwartz, D. M., Kanno, Y., Villarino, A., Ward, M., Gadina, M., and O'Shea, J. J. (2017). JAK Inhibition as a Therapeutic Strategy for Immune and Inflammatory Diseases. Nat. Rev. Drug Discov. 16, 843–862. doi:10.1038/nrd.2017.201
Scognamiglio, P., Di Natale, C., Perretta, G., and Marasco, D. (2013). From Peptides to Small Molecules: an Intriguing but Intricated Way to New Drugs. Cmc 20, 3803–3817. doi:10.2174/09298673113209990184
Scott, D. E., Bayly, A. R., Abell, C., and Skidmore, J. (2016). Small Molecules, Big Targets: Drug Discovery Faces the Protein-Protein Interaction challenge. Nat. Rev. Drug Discov. 15, 533–550. doi:10.1038/nrd.2016.29
Shang, A.-Q., Wu, J., Bi, F., Zhang, Y.-J., Xu, L.-R., Li, L.-L., et al. (2017). Relationship between HER2 and JAK/STAT-SOCS3 Signaling Pathway and Clinicopathological Features and Prognosis of Ovarian Cancer. Cancer Biol. Ther. 18, 314–322. doi:10.1080/15384047.2017.1310343
Sharma, J., Collins, T., Lam, B., Cain, A., Roach, T. R., Cornaby, C., et al. (2021). SOCS1 Mimetics Attenuate Autoimmune Pathology in a Murine Model of Lupus. Am Assoc Immnol. doi:10.1038/s41598-021-86017-4
Shih, P.-C., and Mei, K.-C. (2021). Role of STAT3 Signaling Transduction Pathways in Cancer Stem Cell-Associated Chemoresistance. Drug Discov. Today 26, 1450–1458. doi:10.1016/j.drudis.2020.11.032
Shouda, T., Hiraoka, K., Komiya, S., Hamada, T., Zenmyo, M., Iwasaki, H., et al. (2006). Suppression of IL-6 Production and Proliferation by Blocking STAT3 Activation in Malignant Soft Tissue Tumor Cells. Cancer Lett. 231, 176–184. doi:10.1016/j.canlet.2005.01.042
Shouda, T., Yoshida, T., Hanada, T., Wakioka, T., Oishi, M., Miyoshi, K., et al. (2001). Induction of the Cytokine Signal Regulator SOCS3/CIS3 as a Therapeutic Strategy for Treating Inflammatory Arthritis. J. Clin. Invest. 108, 1781–1788. doi:10.1172/jci13568
Silvagni, E., Missiroli, S., Perrone, M., Patergnani, S., Boncompagni, C., Bortoluzzi, A., et al. (2021). From Bed to Bench and Back: TNF-α, IL-23/IL-17A, and JAK-dependent Inflammation in the Pathogenesis of Psoriatic Synovitis. Front. Pharmacol. 12, 672515. doi:10.3389/fphar.2021.672515
Soler, M. A., Rodriguez, A., Russo, A., Adedeji, A. F., Dongmo Foumthuim, C. J., Cantarutti, C., et al. (2017). Computational Design of Cyclic Peptides for the Customized Oriented Immobilization of Globular Proteins. Phys. Chem. Chem. Phys. 19, 2740–2748. doi:10.1039/c6cp07807a
Stefan, E., Troppmair, J., and Bister, K. (2018). Targeting the Architecture of Deregulated Protein Complexes in Cancer. Adv. Protein Chem. Struct. Biol. 111, 101–132. doi:10.1016/bs.apcsb.2017.07.001
Stumpf, M. P. H., Thorne, T., de Silva, E., Stewart, R., An, H. J., Lappe, M., et al. (2008). Estimating the Size of the Human Interactome. Proc. Natl. Acad. Sci. 105, 6959–6964. doi:10.1073/pnas.0708078105
Subramaniam, S., Mujtaba, M. R. E., and Haider, S. M. I. (2004). Characterization of a Peptide Inhibitor of. J. Immunol. 172, 7510–7518.
Taleb, S., Romain, M., Ramkhelawon, B., Uyttenhove, C., Pasterkamp, G., Herbin, O., et al. (2009). Loss of SOCS3 Expression in T Cells Reveals a Regulatory Role for Interleukin-17 in Atherosclerosis. J. Exp. Med. 206, 2067–2077. doi:10.1084/jem.20090545
Tamiya, T., Kashiwagi, I., Takahashi, R., Yasukawa, H., and Yoshimura, A. (2011). Suppressors of Cytokine Signaling (SOCS) Proteins and JAK/STAT Pathways. Atvb 31, 980–985. doi:10.1161/atvbaha.110.207464
Timofeeva, O. A., Gaponenko, V., Lockett, S. J., Tarasov, S. G., Jiang, S., Michejda, C. J., et al. (2007). Rationally Designed Inhibitors Identify STAT3 N-Domain as a Promising Anticancer Drug Target. ACS Chem. Biol. 2, 799–809. doi:10.1021/cb700186x
Torella, D., Curcio, A., Gasparri, C., Galuppo, V., Serio, D. D., Surace, F. C., et al. (2007). Fludarabine Prevents Smooth Muscle Proliferation In Vitro and Neointimal Hyperplasia In Vivo through Specific Inhibition of STAT-1 Activation. Am. J. Physiology-Heart Circulatory Physiol. 292, H2935–H2943. doi:10.1152/ajpheart.00887.2006
Trengove, M. C., and Ward, A. C. (2013). SOCS Proteins in Development and Disease. Am. J. Clin. Exp. Immunol. 2, 1–29.
Ungureanu, D., Vanhatupa, S., Kotaja, N., Yang, J., Aittomäki, S., Jänne, O. A., et al. (2003). PIAS Proteins Promote SUMO-1 Conjugation to STAT1. Blood 102, 3311–3313. doi:10.1182/blood-2002-12-3816
Vainchenker, W., and Constantinescu, S. N. (2013). JAK/STAT Signaling in Hematological Malignancies. Oncogene 32, 2601–2613. doi:10.1038/onc.2012.347
Verstovsek, S., Kantarjian, H., Mesa, R. A., Pardanani, A. D., Cortes-Franco, J., Thomas, D. A., et al. (2010). Safety and Efficacy of INCB018424, a JAK1 and JAK2 Inhibitor, in Myelofibrosis. N. Engl. J. Med. 363, 1117–1127. doi:10.1056/nejmoa1002028
Villarino, A. V., Kanno, Y., Ferdinand, J. R., and O’Shea, J. J. (2015). Mechanisms of Jak/STAT Signaling in Immunity and Disease. J.I. 194, 21–27. doi:10.4049/jimmunol.1401867
Viparelli, F., Cassese, A., Doti, N., Paturzo, F., Marasco, D., Dathan, N. A., et al. (2008). Targeting of PED/PEA-15 Molecular Interaction with Phospholipase D1 Enhances Insulin Sensitivity in Skeletal Muscle Cells. J. Biol. Chem. 283, 21769–21778. doi:10.1074/jbc.m803771200
Wang, B., Lemay, S., Tsai, S., and Veillette, A. (2001). SH2 Domain-Mediated Interaction of Inhibitory Protein Tyrosine Kinase Csk with Protein Tyrosine Phosphatase-HSCF. Mol. Cell Biol 21, 1077–1088. doi:10.1128/mcb.21.4.1077-1088.2001
Wang, H., Yang, Y., Sharma, N., Tarasova, N. I., Timofeeva, O. A., Winkler-Pickett, R. T., et al. (2010). STAT1 Activation Regulates Proliferation and Differentiation of Renal Progenitors. Cell Signal. 22, 1717–1726. doi:10.1016/j.cellsig.2010.06.012
Wang, T., Zhou, P., Xie, X., Tomita, Y., Cho, S., Tsirukis, D., et al. (2021). Myeloid Lineage Contributes to Pathological Choroidal Neovascularization Formation via SOCS3. EBioMedicine 73, 103632. doi:10.1016/j.ebiom.2021.103632
Welsch, K., Holstein, J., Laurence, A., and Ghoreschi, K. (2017). Targeting JAK/STAT Signalling in Inflammatory Skin Diseases with Small Molecule Inhibitors. Eur. J. Immunol. 47, 1096–1107. doi:10.1002/eji.201646680
Williams, J. J. L., Alotaiq, N., Mullen, W., Burchmore, R., Liu, L., Baillie, G. S., et al. (2018). Interaction of Suppressor of Cytokine Signalling 3 with Cavin-1 Links SOCS3 Function and Cavin-1 Stability. Nat. Commun. 9, 1–17. doi:10.1038/s41467-017-02585-y
Xie, J., Wang, M., Cheng, A., Jia, R., Zhu, D., Liu, M., et al. (2021). The Role of SOCS Proteins in the Development of Virus-Induced Hepatocellular Carcinoma. Virol. J. 18, 1–10. doi:10.1186/s12985-021-01544-w
Xin, P., Xu, X., Deng, C., Liu, S., Wang, Y., Zhou, X., et al. (2020). The Role of JAK/STAT Signaling Pathway and its Inhibitors in Diseases. Int. Immunopharmacology 80, 106210. doi:10.1016/j.intimp.2020.106210
Yan, Z., Gibson, S. A., Buckley, J. A., Qin, H., and Benveniste, E. N. (2018). Role of the JAK/STAT Signaling Pathway in Regulation of Innate Immunity in Neuroinflammatory Diseases. Clin. Immunol. 189, 4–13. doi:10.1016/j.clim.2016.09.014
Yang, L., Lin, S., Xu, L., Lin, J., Zhao, C., and Huang, X. (2019). Novel Activators and Small-Molecule Inhibitors of STAT3 in Cancer. Cytokine Growth Factor. Rev. 49, 10–22. doi:10.1016/j.cytogfr.2019.10.005
Ying, M., Li, D., Yang, L., Wang, M., Wang, N., Chen, Y., et al. (2010). Loss of SOCS3 Expression Is Associated with an Increased Risk of Recurrent Disease in Breast Carcinoma. J. Cancer Res. Clin. Oncol. 136, 1617–1626. doi:10.1007/s00432-010-0819-6
Yoshimura, A., Naka, T., and Kubo, M. (2007). SOCS Proteins, Cytokine Signalling and Immune Regulation. Nat. Rev. Immunol. 7, 454–465. doi:10.1038/nri2093
Yoshimura, A., Ohkubo, T., Kiguchi, T., Jenkins, N. A., Gilbert, D. J., Copeland, N. G., et al. (1995). A Novel Cytokine-Inducible Gene CIS Encodes an SH2-Containing Protein that Binds to Tyrosine-Phosphorylated Interleukin 3 and Erythropoietin Receptors. EMBO J. 14, 2816–2826. doi:10.1002/j.1460-2075.1995.tb07281.x
Yuan, J., Zhang, F., and Niu, R. (2015). Multiple Regulation Pathways and Pivotal Biological Functions of STAT3 in Cancer. Sci. Rep. 5, 1–10. doi:10.1038/srep17663
Zhang, W.-N., Wang, L., Wang, Q., Luo, X., Fang, D.-F., Chen, Y., et al. (2012). CUEDC2 (CUE Domain-Containing 2) and SOCS3 (Suppressors of Cytokine Signaling 3) Cooperate to Negatively Regulate Janus Kinase 1/signal Transducers and Activators of Transcription 3 Signaling. J. Biol. Chem. 287, 382–392. doi:10.1074/jbc.m111.276832
Zhang, W.-N., Zhou, J., Zhou, T., Li, A.-L., Wang, N., Xu, J.-J., et al. (2013). Phosphorylation-triggered CUEDC2 Degradation Promotes UV-Induced G1 Arrest through APC/CCdh1 Regulation. Proc. Natl. Acad. Sci. 110, 11017–11022. doi:10.1073/pnas.1221009110
Keywords: JAK–STAT pathway, SOCS proteins, inflammation, proteomimetics, feedback regulators
Citation: La Manna S, De Benedictis I and Marasco D (2022) Proteomimetics of Natural Regulators of JAK–STAT Pathway: Novel Therapeutic Perspectives. Front. Mol. Biosci. 8:792546. doi: 10.3389/fmolb.2021.792546
Received: 10 October 2021; Accepted: 29 November 2021;
Published: 03 January 2022.
Edited by:
Adriana Erica Miele, Université Claude Bernard Lyon 1, FranceReviewed by:
Margherita Eufemi, Sapienza University of Rome, ItalyCopyright © 2022 La Manna, De Benedictis and Marasco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniela Marasco, ZGFuaWVsYS5tYXJhc2NvQHVuaW5hLml0
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.