
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mol. Biosci. , 23 July 2020
Sec. Molecular Recognition
Volume 7 - 2020 | https://doi.org/10.3389/fmolb.2020.00160
This article is part of the Research Topic Prokaryotic Communications: From Macromolecular Interdomain to Intercellular Talks (Recognition) and Beyond View all 21 articles
 José Francisco Sanchez-Herrero1,2,3
José Francisco Sanchez-Herrero1,2,3 Manuel Bernabeu1
Manuel Bernabeu1 Alejandro Prieto1
Alejandro Prieto1 Mário Hüttener1
Mário Hüttener1 Antonio Juárez1,4*
Antonio Juárez1,4*Gene duplications are a feature of bacterial genomes. In the present work we analyze the extent of gene duplications in the genomes of three microorganisms that belong to the Firmicutes phylum and that are etiologic agents of several nosocomial infections: Staphylococcus aureus, Enterococcus faecium, and Enterococcus faecalis. In all three groups, there is an irregular distribution of duplications in the genomes of the strains analyzed. Whereas in some of the strains duplications are scarce, hundreds of duplications are present in others. In all three species, mobile DNA accounts for a large percentage of the duplicated genes: phage DNA in S. aureus, and plasmid DNA in the enterococci. Duplicates also include core genes. In all three species, a reduced group of genes is duplicated in all strains analyzed. Duplication of the deoC and rpmG genes is a hallmark of S. aureus genomes. Duplication of the gene encoding the PTS IIB subunit is detected in all enterococci genomes. In E. faecalis it is remarkable that the genomes of some strains encode duplicates of the prgB and prgU genes. They belong to the prgABCU cluster, which responds to the presence of the peptide pheromone cCF10 by expressing the surface adhesins PrgA, PrgB, and PrgC.
Gene duplication is an event in which one gene gives rise to two genes that cannot be operationally distinguished from each other. The duplicated genes remain in the same genome. Gene duplications are among the oldest and perhaps the most frequent of mutation types (Lynch et al., 2008; Lipinski et al., 2011). A duplicated gene provides a greater chance for natural selection to shape a novel function (Long et al., 2003). Gene duplication occurs both in eukaryotes and prokaryotes, and significantly impact their gene repertoires, generating functional diversity and increasing the genome complexity (Zhang, 2003; Conant and Wolfe, 2008; Serres et al., 2009; Innan and Kondrashov, 2010; Gao et al., 2017). Duplication events are highly relevant from a biological point of view because, whenever cellular growth is restricted, escape from these growth restrictions can occur by duplication events that resolve the selective problem. In turn, novel duplication events may facilitate subsequent genetic change by allowing cells to proliferate, hence increasing the probability for subsequent adaptive mutations to occur either in the amplified genes or in unrelated ones (Andersson and Hughes, 2009).
In the bacterial kingdom, gene duplication has been associated with survival in extreme or fluctuating conditions, including exposure to antimicrobial compounds or growth on poor nutrient sources, and may have a role in the coevolution between host and pathogens (Romero and Palacios, 1997; Riehle et al., 2001; Duvernay et al., 2011; Kondrashov, 2012; Sun et al., 2012; Toussaint et al., 2017). Several examples correlating gene duplication with bacterial adaptation to the environment are available. For instance, when high gene dosage confers selective benefits, bacteria maintain tandem arrays of duplicated genes (as previously reviewed Romero and Palacios, 1997; Andersson and Hughes, 2009). There is a high natural frequency of bacterial gene duplication, which exceeds the rate of spontaneous point mutation by several orders of magnitude (Andersson and Hughes, 2009). Recent studies indicate that more than 20% of cells in a population contain duplications in some genomic region despite the absence of any evident selection for such duplications (Anderson and Roth, 1981; Hooper and Berg, 2003; Treangen and Rocha, 2011; Elliott et al., 2013).
In most studies the presence of gene duplications is restricted to specific genes or genomic regions, and a global view of the impact of gene duplications in the bacterial genomes is missing. Escherichia coli is an example. Previous studies in E. coli had shown that some genes such as flu, which encodes the adhesin Ag43, can be present in several copies in different strains (van der Woude and Henderson, 2008; Elliott et al., 2013; Arun et al., 2016), but until recently the extent of gene duplications in the genomes of the different types of pathogenic E. coli has not been available (Bernabeu et al., 2019). Most pathogenic E. coli strains harbor between 80 and 100 duplicated genes. Despite the high genomic diversity of E. coli, a group of about 25 genes is duplicated in most of the virulent E. coli strains, irrespective of the pathotype to which they belong (Bernabeu et al., 2019). Most of those genes code for proteins of unknown function and, as they are absent from the genomes of commensal strains, their gene products likely play a role in virulence.
In the present report we have undertaken a whole-genome analysis of gene duplications in the genomes of some of the most clinically relevant Gram-positive cocci, namely Staphylococcus aureus, Enterococcus faecium, and Enterococcus faecalis.
S. aureus and E. faecium are the Gram-positive representatives of the ESKAPE group, which includes microorganisms that are frequent causes of life-threatening nosocomial infections and display multiple antibiotic resistance phenotypes (Murray, 2000; Naimi et al., 2001; Torell et al., 2005). Staphylococcal and enterococcal bacteremia are prevalent in hospitalized patients, and are associated with significant morbidity and mortality (Bartash and Nori, 2017). The emergence of S. aureus strains resistant to many antibiotics, including methicillin-resistance (MRSA), poses a serious threat to human health even in countries with well-developed health surveillance systems. Some E. faecium and E. faecalis isolates account for about 15% of hospital acquired infections in Europe and the US (Werner et al., 2008; Zarb et al., 2012). E. faecium infections are nowadays of major concern because of their multidrug resistance phenotypes, including resistance to vancomycin (VRE) and ampicillin. Strains of E. faecalis are commensals of the gut microbiota, but under some circumstances they can be pathogenic. Pathogenic strains of E. faecalis are increasingly recognized as serious clinical threats due to both the acquisition of multiple antibiotic resistance determinants and to their capacity to disseminate resistance and virulence features by horizontal gene transfer (HGT) mechanisms (Kao and Kline, 2019). Coinfection of MRSA with VRE can occur, being VRE able to transfer VR to the staphylococci (Kos et al., 2012; McGuinness et al., 2017; Cong et al., 2020).
The genomic analysis performed in this work highlights the importance of some genes in the physiology of staphylococci and enterococci. Some of the identified duplicates likely play a role in virulence and hence can be considered as targets of antimicrobial therapies designed to combat infections caused by these pathogens.
We retrieved and analyzed data (genomic fasta, genbank format file, and the translated coding sequences) of all S. aureus (n = 473), E. faecium (n = 133), and E. faecalis (n = 40) complete assembled genomes from NCBI Refseq (Supplementary Table S1).
For each of the strains studied, irrespective of the species, we downloaded the data and analyzed the extent of gene duplications within its genome. Once each strain was analyzed, we summarized the results obtained and generated the corresponding analysis at the level of species. For each of the three species analyzed we selected a reference strain and only for these we created visual representations of their duplicates. Then we analyzed the gene duplications shared with other strains of the same species. We also analyzed further specific characteristics of each of the species analyzed for further interpretation of the data.
For the analysis of gene duplications we performed an all-vs.-all BLASTp (Altschul et al., 1990) protein similarity search using the translated coding sequence regions and filtering the results with a similarity cutoff >85%, an alignment length between pairs >85%, bit-score >50, and an e-value <10-10. We discarded auto hits and grouped duplicates accordingly.
We analyzed the putative insertion of phages in the bacterial genomic sequences by using the PhiSpy tool (Akhter et al., 2012) and the genbank format file (gbk) for each of the strains. We used the appropriate training set for each strain according to their species and additional default parameters.
We generated a phylogenetic and clusterization analysis using all strains for each of the species analyzed. We used Mash (Ondov et al., 2016) and Sourmash (Titus and Irber, 2016) as they extend a dimensionality-reduction technique to include a pairwise mutation distance enabling the efficient clustering of massive sequence collections. We employed the genomic FASTA files downloaded for each strain and default parameters. For each species, the order of the strains in the different tables follows its phylogenetic relationship.
We summarized and plotted using R1 the duplicated count of groups and genes identified and the number of proteins encoding for transposases, selected within the functional annotation associated. We also analyzed the correlation between the amount of duplicated genes and some variables of interest: phages inserted and duplicated proteins annotated as transposases, hypothetical proteins or proteins of unknown function.
For the visualization of duplicates in the corresponding genomes we retrieved, for each duplicate, genomic features such as the start and end coordinates and strand harboring the coding sequence [either from the genomic feature format file (gff) or from the genomic features within the fasta sequence header]. By using R package BioCircos (Cui et al., 2016) we created a circular representation of the duplicate coordinates along the main chromosome and plasmid sequences if any.
For the analysis of gene duplications shared with other strains of the same species, we selected a single sequence within each duplicated group and we employed BLASTp with filtering parameters as above (similarity >85%, alignment >85%, bit-score >50, and e-value <10-10).
The analysis of the presence of SCV-associated genes (deoC, sstD, plsY, and eap) was done in several staphylococcal strains both coagulase + (S. aureus) and coagulase - (S. epidermidis, S. carnosus, and S. xylosus). We downloaded from NCBI Refseq the complete assembled genomes for S. epidermidis (n = 29) and any available genome for S. carnosus (n = 10) and S. xylosus (n = 57) (Supplementary Table S2). Homology search was done by using BLASTp and the translated coding sequences of these selected genes (Supplementary Table S3) and Staphylococcus spp. proteomes. In this case, we used less stringent filtering parameters, a similarity cutoff >50%, following protein homology guidelines (Pearson, 2013), and other parameters as above (alignment >85%, bit-score >50, and e-value <10–10).
Methicillin and vancomycin resistance determinants were searched for each strain by using the Comprehensive Antibiotic Resistance Database (CARD) (Alcock et al., 2020). They were clustered in operons as previously reported (Courvalin, 2006; Shore and Coleman, 2013). We also used BLASTp to identify the presence of each determinant (similarity >80%, alignment >85%, bit-score >50, and e-value <10-10) and manually curated the results. The presence of mecA or mecC genes in S. aureus (CARD IDs ARO: 3000617 and 3001209, respectively) was considered to confer the methicillin resistance phenotype (MR) and the presence in Enterococcus of vanA, vanB, and/or vanG genes (CARD IDs ARO: 3000010, 3000013, and 3002909, respectively) was considered to confer the vancomycin resistance phenotype (VRE).
The bioinformatics scripts employed for the analysis were deposited and are available at the github website: https://github.com/molevol-ub/BacterialDuplicates.
We searched first for the presence of duplications in the overall number of 473 S. aureus genomes available at the Refseq NCBI database (Supplementary Table S1) performing an all-vs.-all protein search by BLASTp for each strain (Figure 1A and Supplementary Table S4). We also looked for the presence of methicillin resistance determinants and the putative insertion of phages within each genome. From the 473 genomes analyzed, some contain more than 50 groups of duplicates, with up to 190 duplicates. Duplications range from 6 to 84 groups, with more than 50% of the strains encoding more than 26 groups of duplicates. No clear correlation was identified between the total number of duplicates and the number of duplicated transposases annotated (R2 = 0.332, pval-adj = 1.894e-43). On the other hand, a slight correlation was identified between the total number of duplicates and duplicated hypothetical or proteins of unknown function (R2 = 0.8, pval-adj = 4.735e-167 and R2 = 0.532, pval-adj = 7.439e-80, respectively). We also explored the distribution of duplicates under a cutoff of phages inserted. Those strains with >2 phages inserted contain, on average, more duplicates (pval = 0.024).
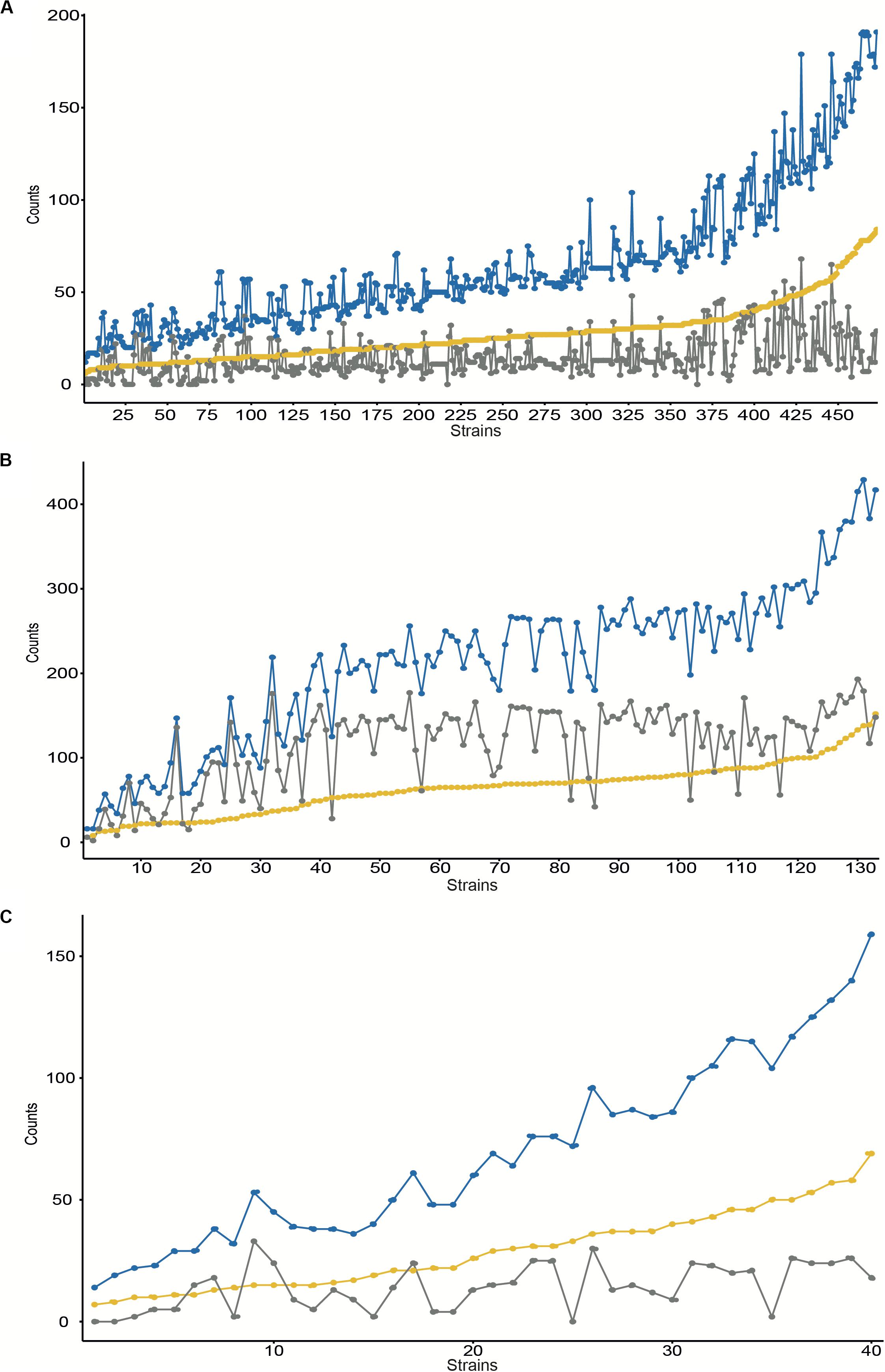
Figure 1. Number of gene duplications identified at the complete assembled genomes for NCBI Refseq entries for S. aureus (A), E. faecalis (B), and E. faecium (C). The Y-axis contains the number of counts for the number of duplicated groups (yellow), the number of gene duplications (blue), and the duplicated transposases (gray). Results are ordered by duplicated groups in increasing order for each strain (axis X).
With regard to E. faecium, we searched for the presence of gene duplications within each strain in all 133 genomes available at the NCBI Refseq database (Supplementary Table S1) by using the same BLAST strategy described above (Figure 1B and Supplementary Table S5). From the 133 genomes analyzed some contain more than 50 groups of duplicates, with up to 429 duplicates. Duplications range from 6 to 152 groups, with more than 50% of the strains encoding more than 66 groups of duplicates. A half correlation was identified between the total number of duplicates and the number of duplicated transposases annotated (R2 = 0.708, pval-adj = 4.828e-37) and with the duplicated hypothetical proteins annotated (R2 = 0.721, pval-adj = 2.0439e-38). We also explored the distribution of duplicates under a cutoff of phages inserted. Those strains with >4 phages inserted contain, on average, more duplicates (p-val = 0.0017).
To complete our survey of duplications, we searched for the presence of duplications in all 40 E. faecalis genomes available (Supplementary Table S1) by using the same strategy described above (Figure 1C and Supplementary Table S6). From the 40 genomes analyzed, some contain more than 50 groups of duplicates, with up to 159 duplicates. Duplications range from 14 to 69 groups, with more than 50% of the strains encoding more than 26 groups of duplicates. No clear correlation was identified between the total number of duplicates and the number of duplicated transposases annotated (R2 = 0.320, pval-adj = 8.4656e-05). A medium correlation was identified with the duplicated hypothetical proteins annotated (R2 = 0.787, pval-adj = 1.435e-14). We also explored the distribution of duplicates under a cutoff of phages inserted. Those strains with >4 phages inserted contain, on average, more duplicates (p-val = 0.0014).
To further study the gene duplications in S. aureus genomes, we decided to analyze them in a well-characterized strain such as S. aureus Newman. It was isolated in 1952 from a human infection (Duthie and Lorenz, 1952) and has been commonly used as a model strain both for studying S. aureus pathogenesis (Richardson et al., 2008; Alonzo et al., 2013) and for the assessment of the therapeutic efficacy of antimicrobial compounds designed to threat S. aureus infections (Thammavongsa et al., 2013; Zhang et al., 2014). Its genome sequence has been available since 2008 (Baba et al., 2008).
We analyzed the extent of gene duplications in strain Newman (GCA_000010465.1), and mapped along the Newman genome those genes that are present in two or more copies (Figure 2 and Supplementary Table S7). A total number of 78 genes are duplicated in that strain. Most of the duplicated genes are located in two main regions (Supplementary Table S7). The insertion phage analysis identified five putative phages within the chromosome of this strain. Four of this phage coordinates match quite well with four prophages previously described in the Newman strain (Bae et al., 2006; Baba et al., 2008). Many of the genes that are duplicated in this strain are in the same coordinates as these phages (Figure 2). Specifically, several duplicates correspond to genes of phages ΦMN4, ΦMN2, and ΦMN1. Some ΦMN4 genes are present in both ΦMN2 and ΦMN1 and hence are present as triplicates. Other ΦMN2 genes are also present in ΦMN1, and are therefore present as duplicates (Supplementary Table S7). A small percentage of the duplicates maps outside the phage genomes (Figure 2).
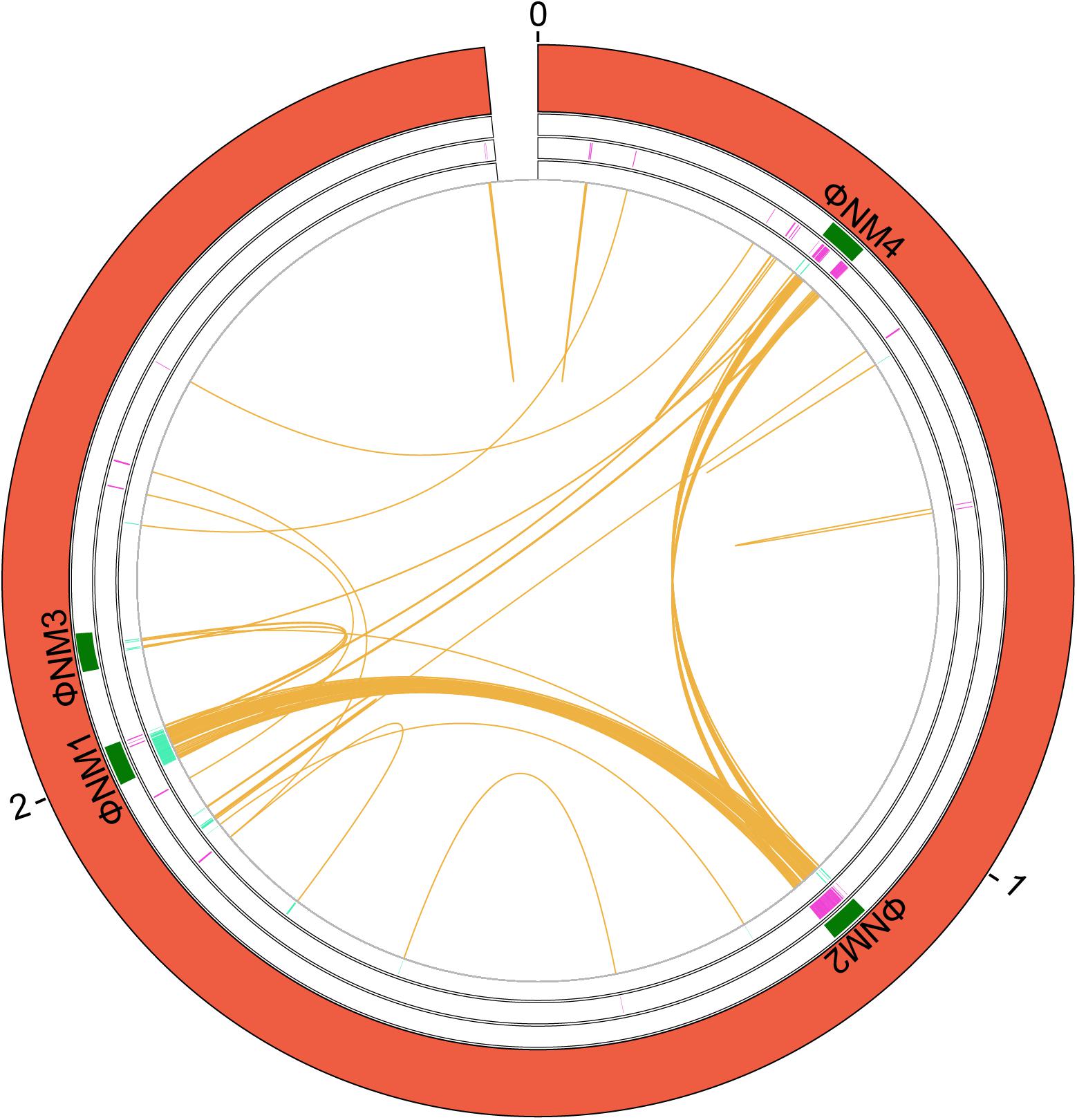
Figure 2. Genes duplicated in the S. aureus strain Newman. A circular map of the chromosome showing the duplications is shown. From the outside in, the outer circle represents the chromosome (red). The second circle includes the position of the identified phages that are inserted in the chromosome (green). Next circle shows the duplicated genes in the (+) strand at each coordinate (purple). The innermost circle shows the duplicated genes in the (−) strand at each coordinate (turquoise). The orange color shows the connection between duplicates. The size is shown in Mb.
With regard to the gene functions of the duplicated genes, 33% correspond to hypothetical proteins, 20% of proteins of unknown function, 19% to phage proteins, and the rest of proteins display miscellaneous functions (Supplementary Table S7).
We addressed next the question as to whether the existing duplicates in this S. aureus strain are strain-specific or, on the contrary, they were generated in some putative ancestor and are also present in many other S. aureus strains. We used the 473 S. aureus genomes to check the shared duplicated genes (see Material and Methods for details). A representative summary of the results obtained is presented in Figure 3. The complete analysis is detailed in Supplementary Table S8.
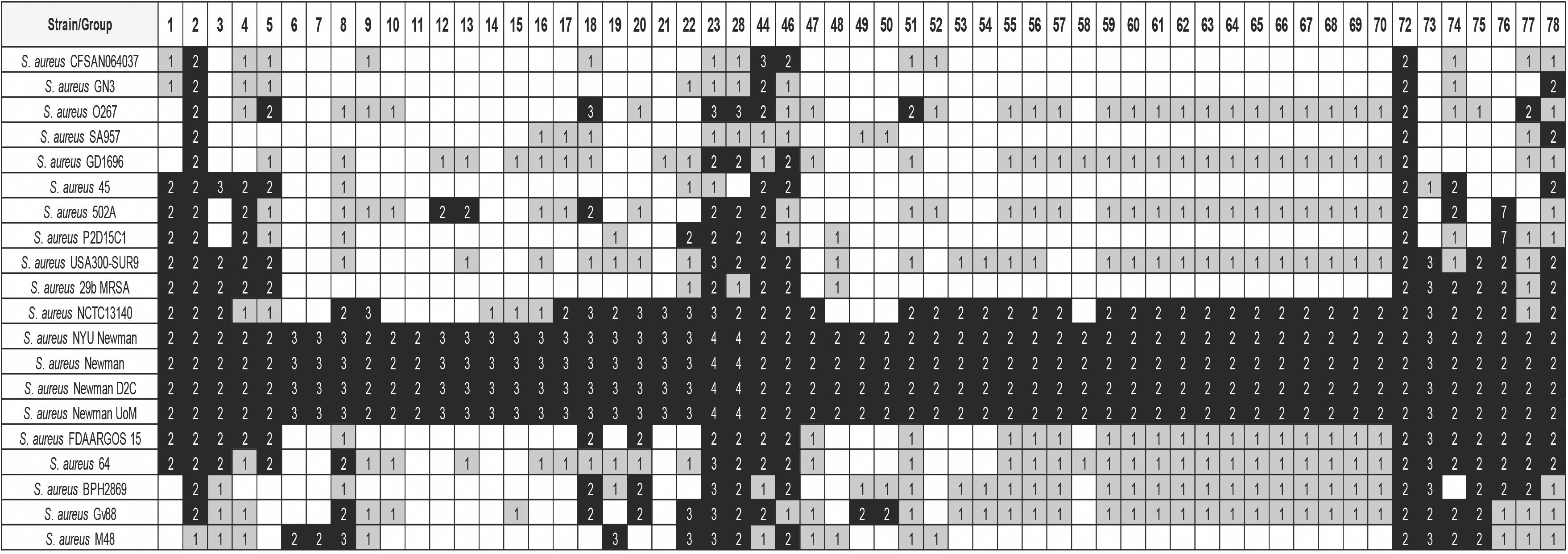
Figure 3. Distribution of some of the detected groups of duplicated genes in strain Newman, in 20 selected S. aureus strains. The white, gray, and black colors indicate, respectively, gene absent; gene present in a single copy; gene present in two or more copies. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship. Analysis of the whole set of 473 genomes is shown in Supplementary Table S8. Details of the duplicated groups are referred in Supplementary Table S7.
As expected, other sequenced Newman strains [strains 412 (GCA_002310435.1), 414 (GCA_002310395.1), and 415 (GCA_900092595.1)] show the same duplication pattern than that obtained with the strain used for the analysis of duplications in the genome [strain 413 (GCA_000010465.1)]. It is also remarkable that strain NCTC13140 [strain 411 (GCA_900474725.1)] shows a gene duplication pattern quite similar to that of the Newman strain. This suggests a close phylogenetic relationship between this strain and the different Newman strains.
The analysis performed also shows that two of the duplicates in strain Newman are also duplicated in most of the strains analyzed (Table 1). These genes are deoC (group 2) and rpmG (group 72). Mutations in deoC (codes for the enzyme deoxyribose phosphate aldolase), prkC (codes for the serine/threonine-protein kinase PrkC), plsY (codes for the enzyme glycerol-3-phosphate acyltransferase), eap (codes for an extracellular adherence protein), and sstD (codes for an iron-binding protein belonging to an ABC uptake transporter) have been shown to result in the generation of small colony variants (SCV) of S. aureus (Chen et al., 2018). We analyzed therefore whether there also existed duplicates of other genes related to the formation of SCVs in all staphylococci (Supplementary Table S2). For this analysis, a lower similarity cutoff (>50%) was used in order to detect duplicates with lower similarity (Supplementary Table S9). Out of deoC, the rest of the genes that have been associated to the generation of SCVs are not duplicated in the genus Staphylococcus. Interestingly, deoC is duplicated in all S. aureus strains analyzed, but not in other catalase negative staphylococci.
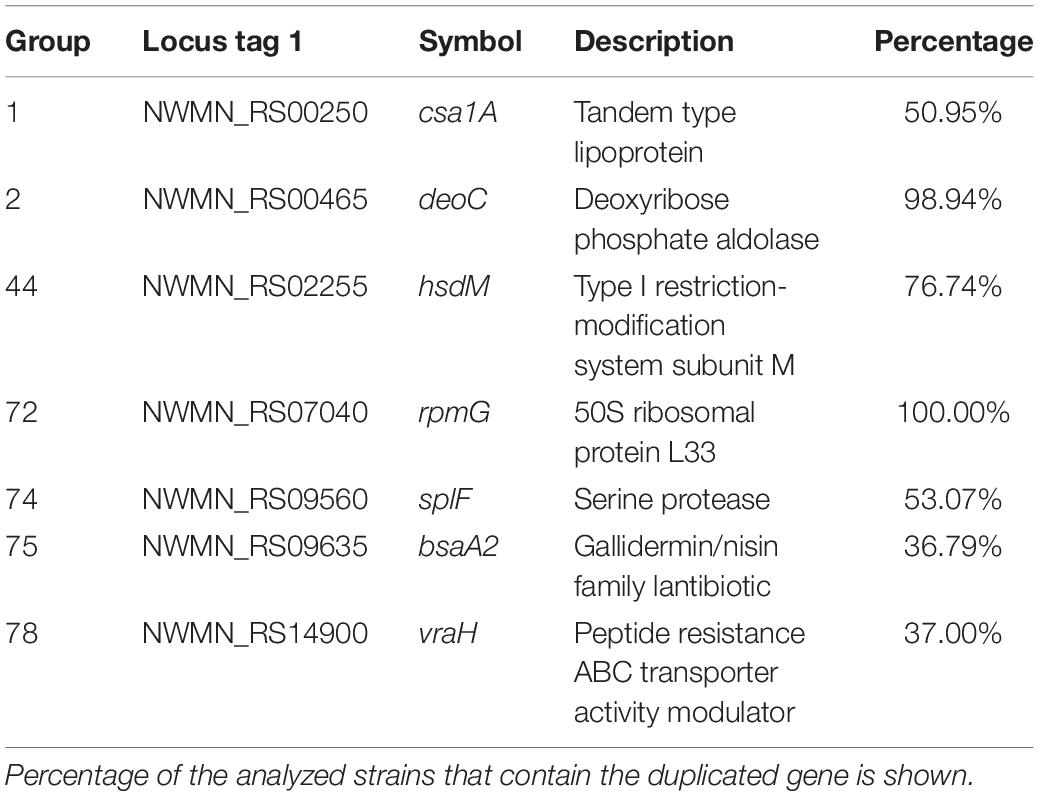
Table 1. Details of the selected duplicated genes of the strain Staphylococcus aureus Newman that are also duplicated in other S. aureus strains.
The rpmG gene codes for the ribosomal protein L33. The existence of duplicates of the genes coding, among other ribosomal proteins, for the ribosomal protein L33 was already reported in some Gram-positive microorganisms (i.e., Bacillus subtilis, B. anthracis, and Lactococcus lactis) as well as in some mycoplasma (Makarova et al., 2001; R Development Core Team, 2008; Kandari et al., 2018).
Out of deoC and rpmG, another set of genes are duplicated in a group of about 250 strains that are phylogenetically related (Figure 3 and Supplementary Table S8). From these genes, (groups 1, 3, 4, 5, 8, 18–23, 28, 44, 46, 73–78; Supplementary Table S7) eight are phage genes (groups: 8, 18–23, 28). From the rest, the function of some is known (Table 1). They code, respectively, for a lipoprotein (group 1, gene csa1A), for the M subunit of a restriction/modification system (group44, gene hsdM), for the SplF serine protease (group 74, gene splF), for a lantibiotic of the gallidemin/nisin family (group 75, gene bsaA2), and for a gene belonging to the vraDEH operon (group 78, gene vraH), associated to the S. aureus resistance to antimicrobial peptides and to cells survival in an infection model (Popella et al., 2016). S. aureus Spl proteases are believed to induce allergic reactions (Stentzel et al., 2017).
We used for the study strain 6E6 (GCA_001518735.1), a vancomycin resistant isolate from the University of Minnesota (Geldart and Kaznessis, 2017), that contains a large number of duplicates (n = 337). We identified and mapped those genes that are present in two or more copies in strain 6E6 (see section “Materials and Methods” for details; Figure 4 and Supplementary Table S10). A total number of 111 gene groups are duplicated in that strain. Some of them (23%) correspond to transposases, which are present in three or more copies (Supplementary Table S10). Several of the duplicates contain at least one of the copies in a plasmid (Figure 4). With regard to the gene functions of the duplicated genes, 41% code for hypothetical proteins, 4.5% code for proteins of unknown function, and the rest for proteins with varying functions (Supplementary Table S10). The high percentage of duplicates that code for proteins of unknown function can be correlated with the HGT origin of these genes.
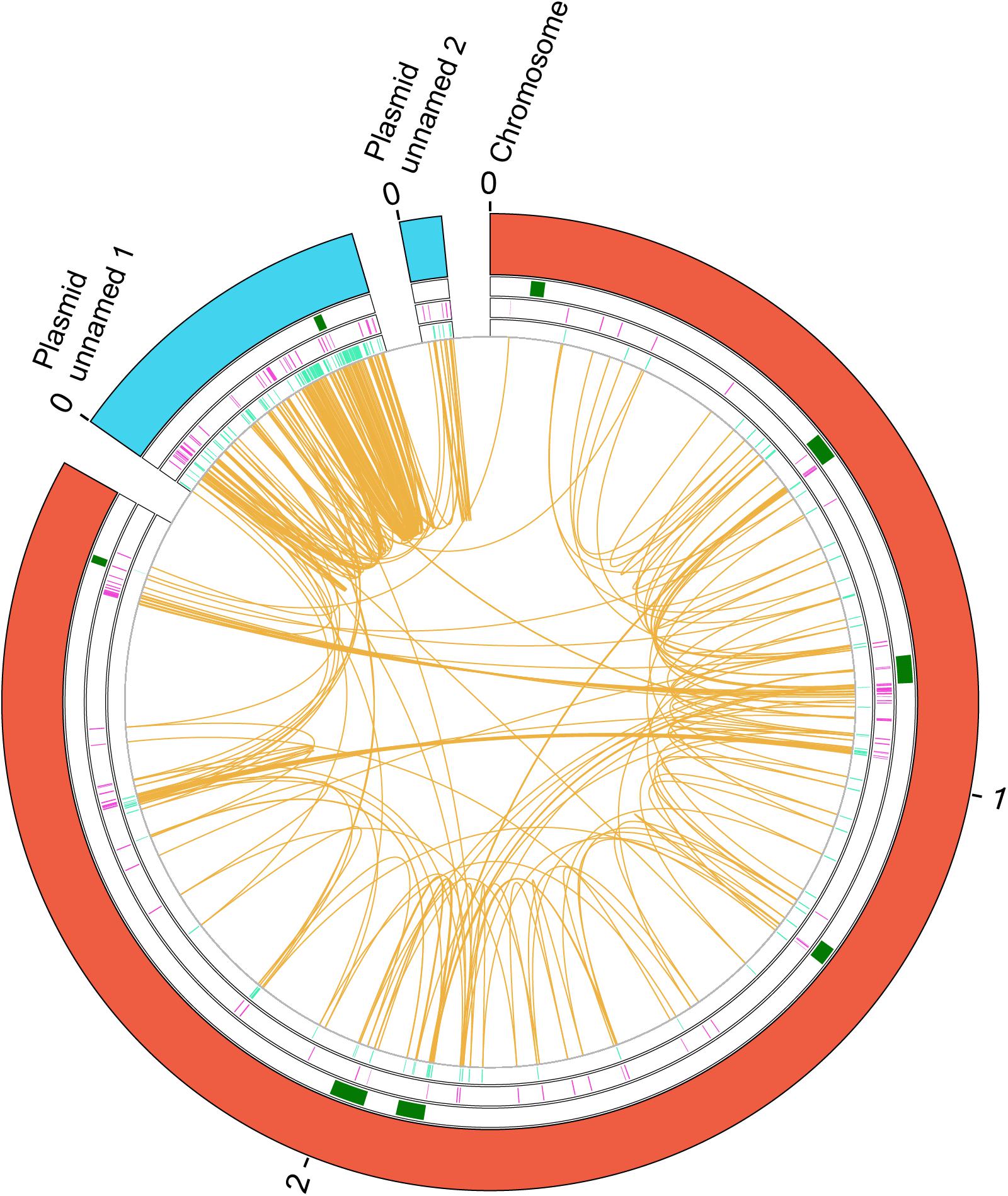
Figure 4. Genes duplicated in the E. faecium 6E6 strain. A circular map of the genome of the E. faecium 6E6 strain showing the duplicates in the chromosome and in two plasmids is shown. From the outside in, the outer circle represents the chromosome (red) and the plasmids (blue). The second circle includes the position of the identified phages that are inserted in the chromosome (green). Next circle shows the duplicated genes in the (+) strand at each coordinate (purple). The innermost circle shows the duplicated genes in the (−) strand at each coordinate (turquoise). The orange color shows the connection between duplicates. The size is shown in Mb.
Once we determined the gene duplications in the E. faecium 6E6 genome, we analyzed whether the existing duplicates are also present in other E. faecium strains. We used the 133 E. faecium genomes to check the shared duplicated genes. Among the genes that are duplicated in strain 6E6 (Supplementary Table S10), a set of 26 genes are also duplicated in most if not all of the strains. A summary of representative data is presented in Figure 5 (see Supplementary Table S11 for the complete analysis). These duplicates can be divided in two groups. The first group (18 genes) includes either transposases or transposon-associated genes. At least one of the corresponding copies maps in a plasmid. The second group (eight genes) comprises four other genes that code either for phage proteins or for hypothetical proteins (groups 105–108), and four genes that code for proteins, which display well-characterized physiological functions (Table 2). They code, respectively, for a GlsB-like protein (group 103), for a LysM-like protein (group 104), and for two proteins of the PTS system: the lactose/cellobiose IIA and IIB subunits (groups 109 and 110, respectively).
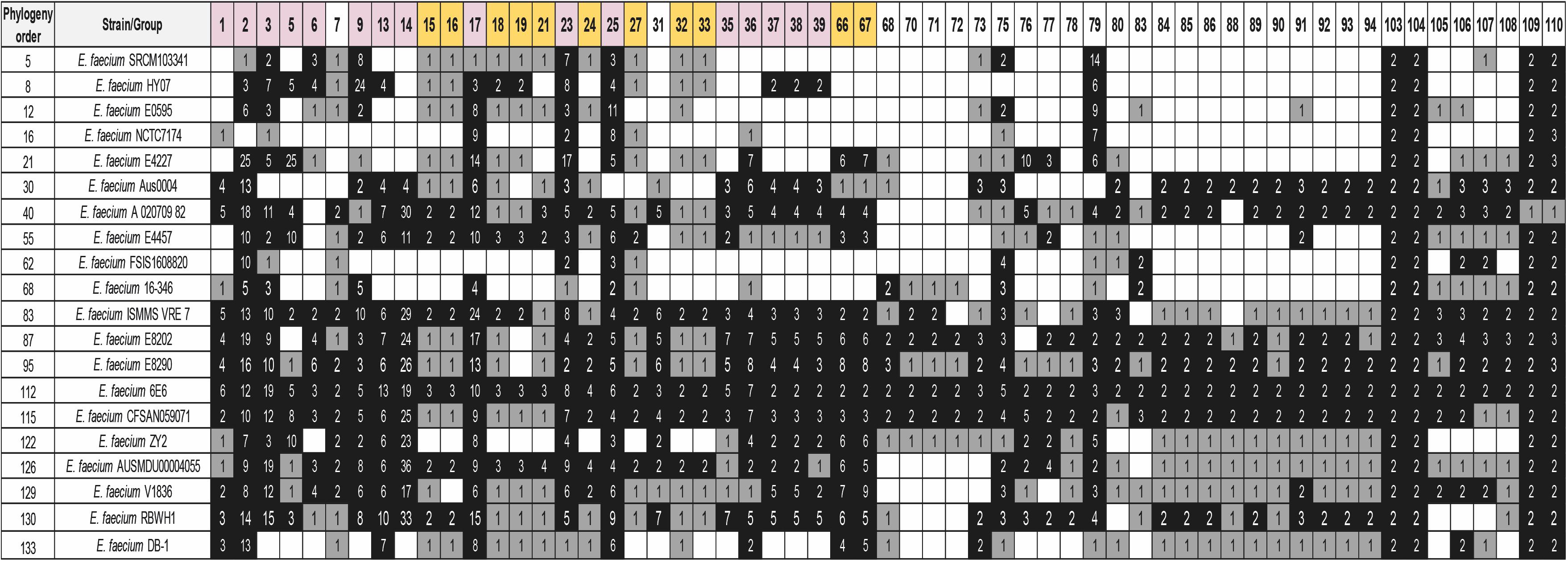
Figure 5. Distribution of some of the detected groups of duplicated genes of strain 6E6 in 20 selected E. faecium strains. BLASTp analysis was used for the study. The white, gray, and black colors indicate, respectively, gene absent; gene present in a single copy; gene present in two or more copies. Purple color corresponds to genes with at least one copy in the chromosome and one copy in a plasmid. The gold color corresponds to plasmid genes. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship. Analysis of the whole set of 133 genomes together with their phylogenetic relationship is shown in the Supplementary Table S11. Details of the duplicated groups are shown in Supplementary Table S10.
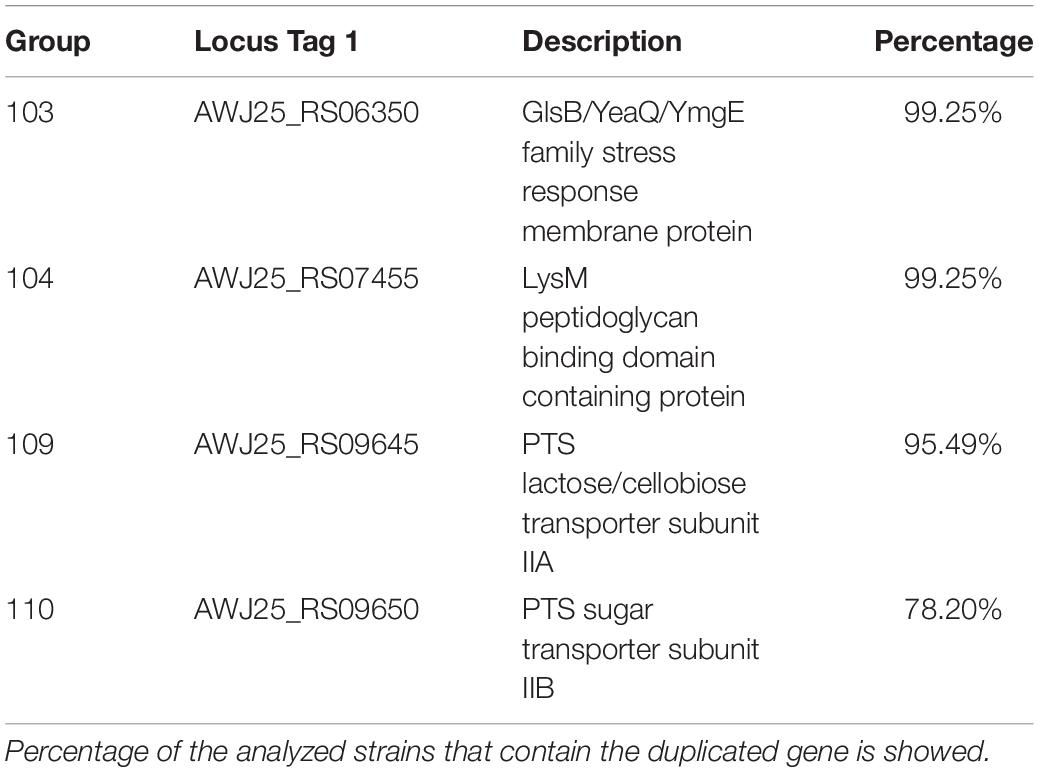
Table 2. Details of the selected duplicated genes of the strain Enterococcus faecium 6E6 that are also duplicated in other E. faecium strains.
We selected for this study as a reference strain E. faecalis V583 (GCA_000007785.1), a VanB-type vancomycin-resistant virulent isolate that is a model strain for E. faecalis studies (Paulsen et al., 2003).
We analyzed the gene duplications in strain V583 (see Materials and Methods for details), and mapped along the V583 genome those genes that are present in two or more copies (Figure 6 and Supplementary Table S12). A total number of 52 gene groups are duplicated in that strain. Some of them (5.7%) correspond to transposases (Supplementary Table S12). As it happened with E. faecium strain 6E6, a large number of duplicates (50.9%) are plasmid genes. With regard to the gene functions of the duplicated genes, 18 duplicated genes code for hypothetical proteins (35%), and the rest code for proteins with different functions (Supplementary Table S12).

Figure 6. Genes duplicated in the E. faecalis V583 strain. A circular map of the genome showing the duplicates in the chromosome and in three plasmids is shown. From the outside in, the outer circle represents the chromosome (red) and plasmids (blue). The second circle shows the position of the identified phages that inserted in the chromosome (green). The next circle shows the duplicated genes in the (+) strand at each coordinate (purple). The innermost circle shows the duplicated genes in the (−) strand at each coordinate (turquoise). The orange color shows the connection between duplicates. The size is shown in Mb.
We also analyzed if the existing duplicates in strain V583 are also present in many other E. faecalis strains. We used the 40 E. faecalis genomes (Supplementary Table S1) to check the shared duplicated genes with strain V583 (see Material and Methods for details) (Figure 7 and Supplementary Table S13). In contrast to E. faecium, only a small group of duplicates (five) is present in almost all the strains analyzed. All of them are chromosomal duplicates. The dgaEF genes (groups 41 and 42) are required for microbial growth on glucose aminoate (Miller et al., 2013). The genes from group 43 code for the subunit IIB of the PTS system. This gene is also duplicated in E. faecium. The genes from group 44 (galE) code for the UDP glucose 4 epimerase that catalyzes the last step of the Leloir pathway for the assimilation of galactose. The genes from Group 52 code for a protein containing a LPXTG cell wall anchor domain (Table 3).
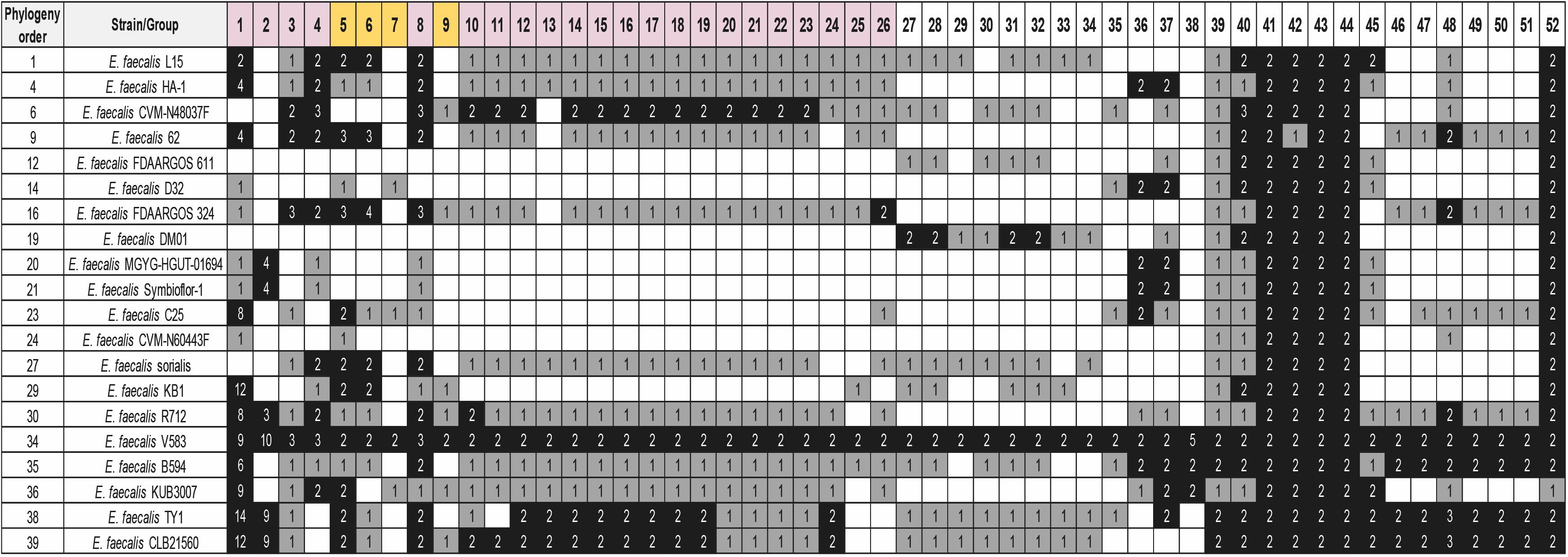
Figure 7. Distribution of the detected groups of duplicated genes of strain V583 in 20 selected E. faecalis strains. BLASTp analysis was used for the study. The white, gray, and black colors indicate, respectively, gene absent; gene present in a single copy; gene present in two or more copies. Purple color corresponds to genes with at least one copy in the chromosome and one copy in a plasmid. The gold color corresponds to genes with all copies in plasmids. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship. Analysis of the whole set of 40 genomes together with their phylogenetic relationship is shown in Supplementary Table S13. Details of the duplicated groups are shown in Supplementary Table S12.
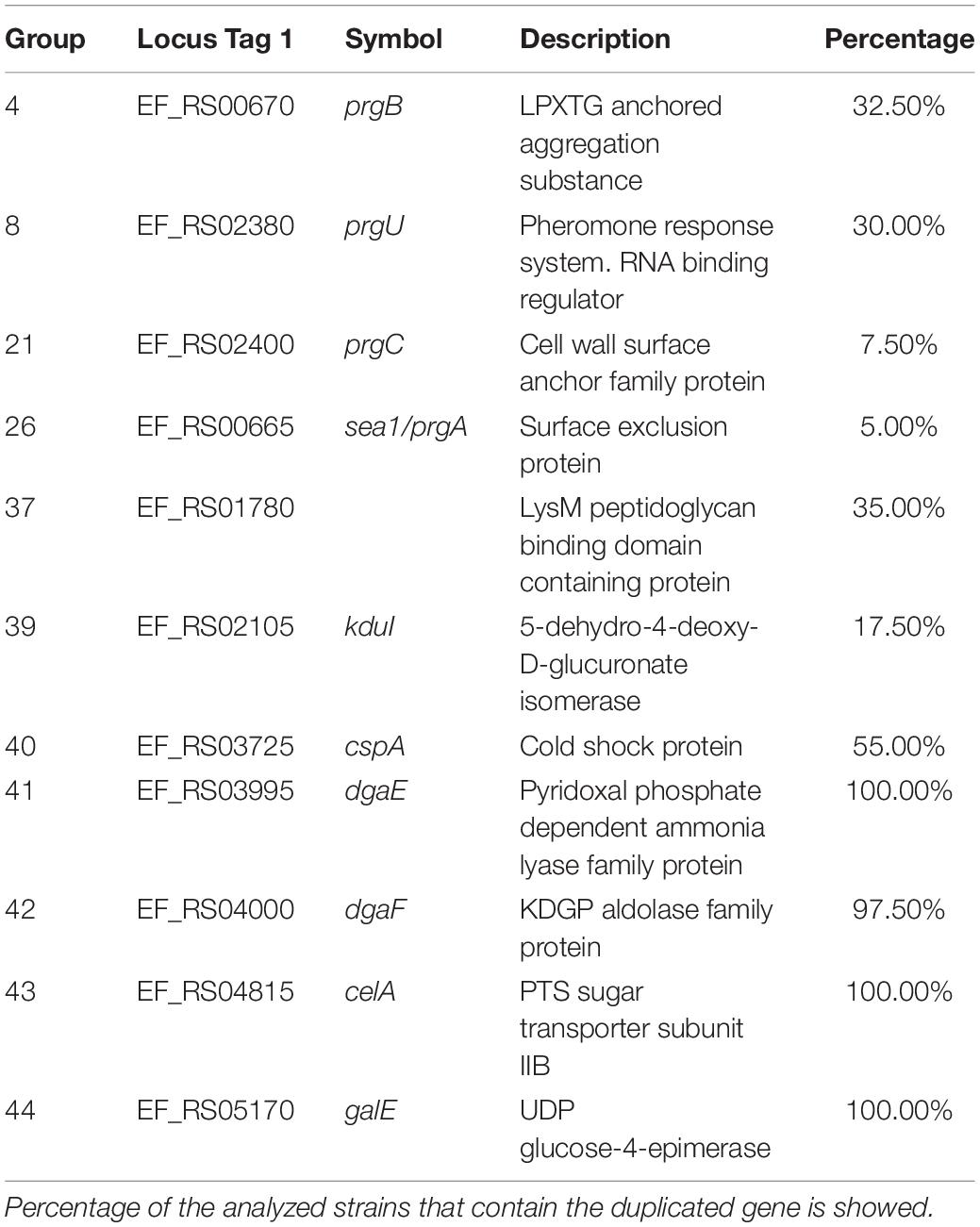
Table 3. Details of the selected duplicated genes of the strain Enterococcus faecalis V583 that are also duplicated in other E. faecalis strains.
Two other groups of duplicates are present in a significant number of the strains analyzed: groups 1–9, encoded in plasmids, and groups 36–40, encoded in the chromosome. From groups 1–9, it is relevant to mention here the prgB and prgU genes (groups 4 and 8, respectively). E. faecalis strains harboring plasmid pCF10 respond to the presence of the peptide pheromone cCF10 by expressing three surface adhesins: PrgA, PrgB, and PrgC. They play a relevant role in host tissue attachment and biofilm formation (Gilmore et al., 2014). Overexpression of PrgB can be highly toxic to E. faecalis cells, and PrgU mitigates toxicity by downregulating PrgB synthesis (Bhatty et al., 2017). It was already reported that strain V583 contains several copies of the prgU gene (Bhatty et al., 2017). We show here that this duplication is present in several other E. faecalis strains.
With respect to groups 36–40, they code, respectively, for a holin, for a protein containing a LysM peptide-binding domain, for a transposase, for a protein that participates in pectin degradation (the kdu gene product) and for the cold shock protein CspA (Table 3).
Strains V583, VE18379, VE14089, and VE18395 that appear to share a set of duplicates, are closely related. Strain VE14089 is plasmid-free V583. Strains VE18379 and VE18395 are derivatives from strain VE14089.
A relevant question to be addressed is whether the identified duplicated genes that code for core functions result either from ancient duplications and are located in fixed points of the chromosome, or have been generated because of some of these genes being flanked by IS elements and jumping to different positions in the chromosome. To assess this, we analyzed the genomic context of both copies of the deoC gene in representative strains of S. aureus, and of the celA gene in representative strains of E. faecium and E. faecalis (Figure 8). The genomic context of the two alleles of the deoC and celA genes is the same in the different strains analyzed. They are not surrounded by IS elements, but by other core genes.
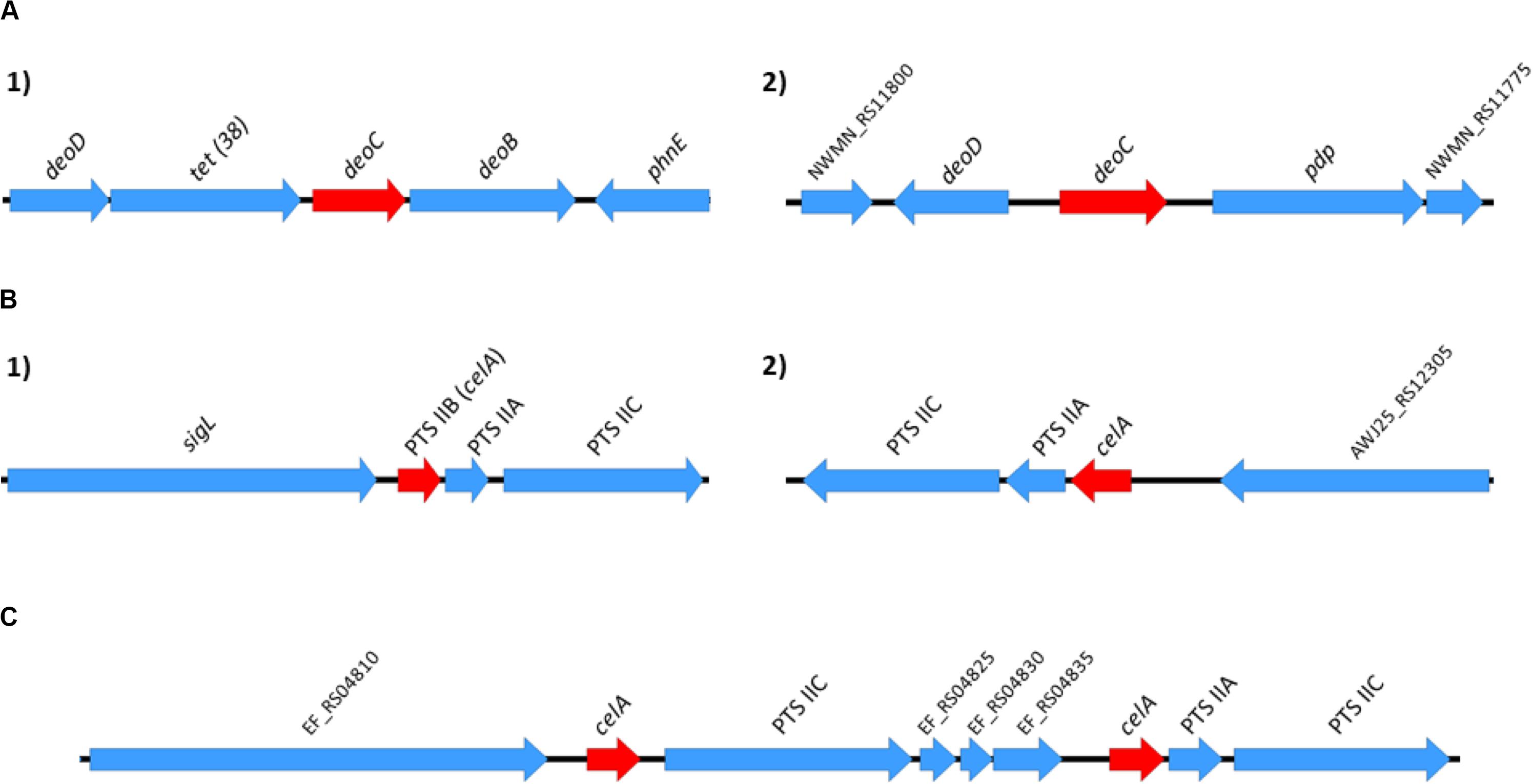
Figure 8. Genomic context of the S. aureus deoC duplicated genes (A) and of the E. faecium and E. faecalis celA duplicated genes (B,C, respectively). The genomic context was analyzed in the following representative strains: S. aureus Newman, S. aureus N315, S. aureus USA300, and S. aureus MW2; E. faecium 6E6, E. faecium Aus0004 and E. faecium DO; E. faecalis V583, E. faecalis OG1RF and E. faecalis Symbioflor. Red arrows in positions 1 and 2 correspond to the alleles of the duplicated genes. The genomic context for the deoC and celA genes was identical in all S. aureus and E. faecium/E. faecalis strains analyzed.
It is apparent that in the Gram-positive cocci studied in this work, mobile DNA elements encode a significant part of the duplicates present in their genomes. The statistical analysis performed correlates duplicates both with phages inserted in the chromosome and with genes encoding either hypothetical or proteins of unknown function (much more common in mobile DNA than in the core genome). In S. aureus, most of the duplicated genes are of phage origin. This is not surprising because of the relevant role that phages have in the biology of this microorganism (Xia and Wolz, 2014; Ingmer et al., 2019). In contrast, plasmids are the predominant mobile elements, which encode a significant part of the duplicates in the enterococci. In E. faecium there exists a significant correlation between duplicates and transposases. This is also shown when the existing duplicates are identified by BLAST analysis. The question to be addressed is the biological significance of the duplication of genes encoded in mobile DNA in these microorganisms.
Although duplicates located in mobile DNA predominate in the microorganisms studied here, others are located in the chromosome. Some of these latter duplicates are widespread among all the strains of the same species analyzed. In S. aureus, two duplicates are present in almost all strains analyzed: the rpmG and the deoC genes. The former codes for the ribosomal protein L33. Duplications of the gene encoding the L33 protein appear to be a hallmark of several Gram-positive genera. As a general rule, one of the rmpG paralogs codes for a protein that contains a Zn-binding motif comprising a two pair of conserved CXXC stretch (CC form), which is absent in the other (C- form). In strain Newman, two copies are C- (NWMN_RS07040, and NWMN_RS08205, respectively), and the third is CC [NWMN_0496.1 (this latter shows less than 85% identity)]. In addition to their role in translation, ribosomes also serve as reservoirs for zinc in the cell (Moore and Helmann, 2005). The zinc-responsive regulator Zur has been shown to repress the C- form (Akanuma et al., 2006; Gabriel and Helmann, 2009). Under zinc-depleted conditions, the Zur mediated repression of the genes encoding the C- forms of the ribosomal proteins is alleviated. These C- forms then replace the corresponding CC forms from the ribosomes, resulting in exoneration of zinc, which can then be used by other metalloproteins. This enables the bacterial cell to survive in zinc limiting environments (Moore et al., 2005; Akanuma et al., 2006).
The deoC gene product is the deoxyribose phosphate aldolase, which enables bacterial cells to grow on deoxyribonucleosides as the carbon source. As commented above, mutations in the S. aureus deoC gene have been associated with the generation of SCVs (Chen et al., 2018). Interestingly, deoC mutations were associated with alterations in the response to extracellular signaling in E. coli (Joloba and Rather, 2003). It can be hypothesized that, as the deoC gene is duplicated in S. aureus but not in other catalase negative cocci, its gene product can play a role in S. aureus virulence.
In addition to these widespread duplicates, another group of duplicates is present in a subset of the S. aureus genomes. The strains containing these duplicates are phylogenetically related. The reported functions for these genes (i.e., vraH, splF) are also related to virulence, and they can be considered as virulence markers of that group of S. aureus strains. A question to be addressed is whether the duplication of these genes confers specific virulence features to S. aureus.
In E. faecium genomes, a significant part of the duplicates (58%) are located (at least one of the copies) in plasmids. Several of these duplicates code either for transposases or for hypothetical proteins. Some of them are shared by most of the strains analyzed. In addition to these genes of plasmid origin, four chromosomal genes are also duplicated in most of the E. faecium strains analyzed. Expression of GlsB proteins has been associated with virulence and bile salt stress (Choudhury et al., 2011; Zhang et al., 2013a). Proteins containing a LysM domain have been shown to be induced under infection conditions of a mammalian host (Cacaci et al., 2018). Although the proteins of the lactose/cellobiose PTS system IIA and IIB have not been hitherto described as relevant elements in E. faecium virulence, the relevance of the PTS system for the ability of E. faecium to colonize the host has been previously reported. Deletion of the pstD gene, which is predicted to encode the enzyme IID subunit of a PTS system, influenced E. faecium virulence (Zhang et al., 2013b). Furthermore, insertional inactivation of the bepA gene, coding for putative a PTS permease, was found to be relevant for E. faecium pathogenesis (Paganelli et al., 2016). The fact that, as shown in this report, other PTS specific components are duplicated in E. faecium strains further highlights the role of the phosphotransferase system in E. faecium physiology and hence, in the ability of virulent strains to colonize their hosts.
As it happens in E. faecium, about half of the E. faecalis duplicates are located (at least one of the copies) in a plasmid. Nevertheless, in contrast to E. faecium, few of the E. faecalis duplicates that are located in plasmids are transposases. A cluster of duplicated plasmid genes (prgABCU) are of special relevance. Its gene products enable E. faecalis to respond to the presence of the peptide pheromone cCF10 by expressing the surface adhesins PrgA, PrgB, and PrgC (Gilmore et al., 2014). It has been suggested that prgU expression controls prgB expression, avoiding that excess of the PrgB protein can be deleterious for the cell. prgB and prgU are present in several copies in strain V583, and there exists a genetic linkage between both genes (Bhatty et al., 2017). We show in this work that gene duplication occurs predominantly with both the prgB and prgU genes, and not with prgA and prgC genes. Our data are hence consistent with the genetic linkage of prgB and prgU (Bhatty et al., 2017). prgU genes are widely distributed on plasmids and chromosomes of E. faecalis and other enterococci, and it has been suggested that the prgB-prgU genetic linkage might have evolved to ensure the controlled synthesis of PrgB-like adhesins (Bhatty et al., 2017). Accordingly, we show here that the genetic linkage of prgB-prgU also involves gene duplication. In accordance with the rule that we suggested previously for E. coli (Bernabeu et al., 2019), the duplication of the regulated gene (prgB) correlates with the duplication of its modulator (prgU).
In E. faecalis there also exists a group of duplicates located in the chromosome that is shared by all the strains analyzed. Three of them code for proteins playing a role in cell metabolism, including the duplication of the gene coding for of the subunit IIB of the lactose/cellobiose PTS system. Different components of the PTS system have also been reported as relevant for E. faecalis colonization (Paulsen et al., 2003), and they have deserved special attention in the last years (Ruiz-Cruz et al., 2015; Sauvageot et al., 2017; Grand et al., 2019).
Both for S. aureus and Enterococcus, the genomic context of the duplicates coding for core genes is similar among strains, and corresponds to other core genes. This fact, together with the widespread distribution of these genes among all strains analyzed suggests that these duplications correspond to ancient events that have been positively selected in the course of evolution.
Genomic analysis is powerful to gain insight into several aspects of the biology of organisms. We show here that the analysis of the pattern of gene duplications in microorganisms can provide relevant information that can be useful for both establishing phylogenetic relationships between strains, and for the identification of genes that can play relevant roles in, among other processes, bacterial virulence.
All datasets generated for this study are included in the article/Supplementary Material.
JS-H, MB, and AJ designed this study. JS-H and MB performed the in silico work. JS-H, MB, AP, MH, and AJ wrote the manuscript. MH and AJ did the final version of the manuscript. All authors analyzed and discussed the results, read and approved the final manuscript.
JS-H was beneficiary of a contract by the Instituto de Salud Carlos III through the Accion Estrategica en Salud 2018 (Co-funded by the European Regional Development Fund/European Social Fund; “A way to make Europe”/“Investing in your future”). MB was the recipient of a FI Fellowship from the Generalitat de Catalunya. This work was supported by grant BIO2016-76412-C2-1-R (AEI/FEDER, UE) from the Ministerio de Economía, Industria y Competitividad, Fundació La Marató TV3, Spain (Project ref. 201818 10), and the CERCA Programme/Generalitat de Catalunya to AJ.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the IGTP High Content Genomics & Bioinformatics Unit Core Facility and staff (Dr. Lauro Sumoy) for their contribution to this publication. We additionally thank the Evolutionary Genomics and Bioinformatics Group and Dr. Julio Rozas particularly at the University Barcelona for providing us with computational resources for this study.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmolb.2020.00160/full#supplementary-material
TABLE S1 | List of the complete assembled genomes corresponding to NCBI Refseq entries for S. aureus, E. faecalis, and E. faecium used for the gene duplication analysis.
TABLE S2 | List of the NCBI Refseq entries for S. epidermidis, S. carnosus, and S. xylosus used for the analysis of the duplications of genes involved in the generation of small colony variants (SCV).
TABLE S3 | Details of the genes involved in the generation of SCVs used for the duplication analysis.
TABLE S4 | Gene duplication data obtained for the S. aureus strains analyzed. Indicated are: (i) Phylogeny order: according to the clusterization analysis using mash and sourmash tools. (ii) Groups: counts of groups of duplicated genes. (iii) Dups: total count of duplicated genes. (iv) Transpo: counts of genes within duplicates annotated as “transposase”. (v) Hypo: counts of gene names within duplications annotated or containing “hypothetical” functions. (vi) Phages: Number of putative phages identified with PhiSpy inserted in the genome. (vii) MRSA: presence of methicillin resistance determinants mecA and/or mecC. The order of the strains follows its phylogenetic relationship.
TABLE S5 | Gene duplication data obtained for the E. faecium strains analyzed. Indicated are: (i) Phylogeny order: according to the clusterization analysis using mash and sourmash tools. (ii) Groups: counts of groups of duplicated genes. (iii) Dups: total count of duplicated genes. (iv) Transpo: counts of genes within duplicates annotated as “transposase”. (v) Hypo: counts of gene names within duplications annotated or containing “hypothetical” functions. (vi) Phages: Number of putative phages identified with PhiSpy inserted in the genome. (vii) VRE: presence of vancomycin resistance determinants vanA (blue), vanB (yellow), vanA/vanB (green), or vanG (red). The order of the strains follows its phylogenetic relationship.
TABLE S6 | Gene duplication data obtained for the E. faecalis strains analyzed. Indicated are: (i) Phylogeny order: according to the clusterization analysis using mash and sourmash tools. (ii) Groups: counts of groups of duplicated genes. (iii) Dups: total count of duplicated genes. (iv) Transpo: counts of genes within duplicates annotated as “transposase”. (v) Hypo: counts of gene names within duplications annotated or containing “hypothetical” functions. (vi) Phages: Number of putative phages identified with PhiSpy inserted in the genome. (vii) VRE: presence of vancomycin resistance determinants vanA (blue) or vanB (yellow). The order of the strains follows its phylogenetic relationship.
TABLE S7 | Locus tag and description of the duplicates identified in S. aureus strain Newman. Red, blue and green colors correspond to genes located in the ΦMN4, ΦMN2, and ΦMN1 phages, respectively.
TABLE S8 | Distribution of the detected duplicated genes of strain Newman in the genomes of the other 472 S. aureus strains analyzed. BLASTp analysis was used for the study. The white, gray, and black colors indicate, respectively: gene absent; gene present in a single copy; gene present in two or more copies. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship.
TABLE S9 | Analysis of the duplications of genes deoC, sstD, plsY, and eap in S. aureus, S. epidermidis, S. carnosus, and S. xylosus.
TABLE S10 | Locus tag and description of the duplicates identified in E. faecium strain 6E6. The purple and gold colors correspond, respectively, to genes with at least one copy in chromosome and one in plasmids, and to genes with all copies in plasmids.
TABLE S11 | Distribution of the detected duplicated genes of strain 6E6 in the genomes of the other 132 E. faecium strains analyzed. BLASTp analysis was used for the study. The white, gray, and black colors indicate, respectively: gene absent; gene present in a single copy; gene present in two or more copies. Purple color corresponds to genes with at least one copy in the chromosome and one copy in a plasmid. The gold color corresponds to genes with all copies in plasmids. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship.
TABLE S12 | Locus tag and description of the duplicates identified in E. faecalis strain V583. The purple and gold colors correspond, respectively, to genes with at least one copy in chromosome and one in plasmids, and to genes with all copies in plasmids.
TABLE S13 | Distribution of the detected duplicated genes of strain V583 in the genomes of the other 39 E. faecalis strains analyzed. BLASTp analysis was used for the study. The white, gray, and black colors indicate, respectively: gene absent; gene present in a single copy; gene present in two or more copies. Purple color corresponds to genes with at least one copy in the chromosome and one copy in a plasmid. The gold color corresponds to genes with all copies in plasmids. The numbers show the copy number of each gene. The order of the strains follows its phylogenetic relationship.
Akanuma, G., Nanamiya, H., Natori, Y., Nomura, N., and Kawamura, F. (2006). Liberation of zinc-containing L31 (RpmE) from ribosomes by its paralogous gene product, YtiA, in Bacillus subtilis. J. Bacteriol. 188, 2715–2720. doi: 10.1128/JB.188.7.2715-2720.2006
Akhter, S., Aziz, R. K., and Edwards, R. A. (2012). PhiSpy: a novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies. Nucleic Acids Res. 40, e126. doi: 10.1093/nar/gks406
Alcock, B. P., Raphenya, A. R., Lau, T. T. Y., Tsang, K. K., Bouchard, M., Edalatmand, A., et al. (2020). CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 48, D517–D525. doi: 10.1093/nar/gkz935
Alonzo, F., Kozhaya, L., Rawlings, S. A., Reyes-Robles, T., DuMont, A. L., Myszka, D. G., et al. (2013). CCR5 is a receptor for Staphylococcus aureus leukotoxin ED. Nature 493, 51–55. doi: 10.1038/nature11724
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Anderson, P., and Roth, J. (1981). Spontaneous tandem genetic duplications in Salmonella Typhimurium arise by unequal recombination between rRNA (rrn) cistrons. Proc. Natl. Acad. Sci. U.S.A. 78, 3113–3117. doi: 10.1073/pnas.78.5.3113
Andersson, D. I., and Hughes, D. (2009). Gene amplification and adaptive evolution in bacteria. Annu. Rev. Genet. 43, 167–195. doi: 10.1146/annurev-genet-102108-134805
Arun, P. V. P. S., Miryala, S. K., Chattopadhyay, S., Thiyyagura, K., Bawa, P., Bhattacharjee, M., et al. (2016). Identification and functional analysis of essential, conserved, housekeeping and duplicated genes. FEBS Lett. 590, 1428–1437. doi: 10.1002/1873-3468.12192
Baba, T., Bae, T., Schneewind, O., Takeuchi, F., and Hiramatsu, K. (2008). Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 190, 300–310. doi: 10.1128/jb.01000-07
Bae, T., Baba, T., Hiramatsu, K., and Schneewind, O. (2006). Prophages of Staphylococcus aureus Newman and their contribution to virulence. Mol. Microbiol. 62, 1035–1047. doi: 10.1111/j.1365-2958.2006.05441.x
Bartash, R., and Nori, P. (2017). Beta-lactam combination therapy for the treatment of Staphylococcus aureus and Enterococcus species bacteremia: a summary and appraisal of the evidence. Int. J. Infect. Dis. 63, 7–12. doi: 10.1016/j.ijid.2017.07.019
Bernabeu, M., Sánchez-Herrero, J. F., Huedo, P., Prieto, A., Hüttener, M., Rozas, J., et al. (2019). Gene duplications in the E. coli genome: common themes among pathotypes. BMC Genomics 20:313. doi: 10.1186/s12864-019-5683-4
Bhatty, M., Camacho, M. I., Gonzalez-Rivera, C., Frank, K. L., Dale, J. L., Manias, D. A., et al. (2017). PrgU: a suppressor of sex pheromone toxicity in Enterococcus faecalis. Mol. Microbiol. 103, 398–412. doi: 10.1111/mmi.13563
Cacaci, M., Giraud, C., Leger, L., Torelli, R., Martini, C., Posteraro, B., et al. (2018). Expression profiling in a mammalian host reveals the strong induction of genes encoding LysM domain-containing proteins in Enterococcus faecium. Sci. Rep. 8:12412. doi: 10.1038/s41598-018-30882-z
Chen, H., Wang, Q., Yin, Y., Li, S., Niu, D.-K., and Wang, H. (2018). Genotypic variations between wild-type and small colony variant of Staphylococcus aureus in prosthetic valve infectious endocarditis: a comparative genomic and transcriptomic analysis. Int. J. Antimicrob. Agents 51, 655–658. doi: 10.1016/j.ijantimicag.2017.12.006
Choudhury, T., Singh, K. V., Sillanpää, J., Nallapareddy, S. R., and Murray, B. E. (2011). Importance of two Enterococcus faecium loci encoding Gls-like proteins for in vitro bile salts stress response and virulence. J. Infect. Dis. 203, 1147–1154. doi: 10.1093/infdis/jiq160
Conant, G. C., and Wolfe, K. H. (2008). Turning a hobby into a job: how duplicated genes find new functions. Nat. Rev. Genet. 9, 938–950. doi: 10.1038/nrg2482
Cong, Y., Yang, S., and Rao, X. (2020). Vancomycin resistant Staphylococcus aureus infections: a review of case updating and clinical features. J. Adv. Res. 21, 169–176. doi: 10.1016/j.jare.2019.10.005
Courvalin, P. (2006). Vancomycin resistance in gram-positive cocci. Clin. Infect. Dis. 42(Suppl. 1), S25–S34. doi: 10.1086/491711
Cui, Y., Chen, X., Luo, H., Fan, Z., Luo, J., He, S., et al. (2016). BioCircos.js: an interactive Circos JavaScript library for biological data visualization on web applications. Bioinformatics 32, 1740–1742. doi: 10.1093/bioinformatics/btw041
Duthie, E. S., and Lorenz, L. L. (1952). Staphylococcal coagulase; mode of action and antigenicity. J. Gen. Microbiol. 6, 95–107.
Duvernay, C., Coulange, L., Dutilh, B., Dubois, V., Quentin, C., and Arpin, C. (2011). Duplication of the chromosomal blaSHV-11 gene in a clinical hypermutable strain of Klebsiella pneumoniae. Microbiology (Reading, Engl.) 157, 496–503. doi: 10.1099/mic.0.043885-0
Elliott, K. T., Cuff, L. E., and Neidle, E. L. (2013). Copy number change: evolving views on gene amplification. Future Microbiol. 8, 887–899. doi: 10.2217/fmb.13.53
Gabriel, S. E., and Helmann, J. D. (2009). Contributions of Zur-controlled ribosomal proteins to growth under zinc starvation conditions. J. Bacteriol. 191, 6116–6122. doi: 10.1128/jb.00802-09
Gao, Y., Zhao, H., Jin, Y., Xu, X., and Han, G.-Z. (2017). Extent and evolution of gene duplication in DNA viruses. Virus Res. 240, 161–165. doi: 10.1016/j.virusres.2017.08.005
Geldart, K., and Kaznessis, Y. N. (2017). Characterization of class iia bacteriocin resistance in enterococcus faecium. Antimicrob. Agents Chemother. 61: e2033-16.
Gilmore, M. S., Clewell, D. B., Ike, Y., Shankar, N., Clewell, D. B., Weaver, K. E., et al. (2014). Extrachromosomal and Mobile Elements in Enterococci: Transmission, Maintenance, and Epidemiology. Boston, MA: Massachusetts Eye and Ear Infirmary.
Grand, M., Aubourg, M., Pikis, A., Thompson, J., Deutscher, J., Hartke, A., et al. (2019). Characterization of the gen locus involved in β-1,6-oligosaccharide utilization by Enterococcus faecalis. Mol. Microbiol. 112, 1744–1756. doi: 10.1111/mmi.14390
Hooper, S. D., and Berg, O. G. (2003). Duplication is more common among laterally transferred genes than among indigenous genes. Genome Biol. 4:R48. doi: 10.1186/gb-2003-4-8-r48
Ingmer, H., Gerlach, D., and Wolz, C. (2019). Temperate phages of Staphylococcus aureus. Microbiol Spectr 7:GPP-3-0058-2018.
Innan, H., and Kondrashov, F. (2010). The evolution of gene duplications: classifying and distinguishing between models. Nat. Rev. Genet. 11, 97–108. doi: 10.1038/nrg2689
Joloba, M. L., and Rather, P. N. (2003). Mutations in deoB and deoC alter an extracellular signaling pathway required for activation of the gab operon in Escherichia coli. FEMS Microbiol. Lett. 228, 151–157. doi: 10.1016/s0378-1097(03)00754-7
Kandari, D., Gopalani, M., Gupta, M., Joshi, H., Bhatnagar, S., and Bhatnagar, R. (2018). Identification, functional characterization, and regulon prediction of the zinc uptake regulator (zur) of Bacillus anthracis – An insight into the zinc homeostasis of the pathogen. Front. Microbiol. 9:3314. doi: 10.3389/fmicb.2018.03314
Kao, P. H. N., and Kline, K. A. (2019). Dr. Jekyll and Mr. Hide: how Enterococcus faecalis subverts the host immune response to cause infection. J. Mol. Biol. 431, 2932–2945. doi: 10.1016/j.jmb.2019.05.030
Kondrashov, F. A. (2012). Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. Biol. Sci. 279, 5048–5057. doi: 10.1098/rspb.2012.1108
Kos, V. N., Desjardins, C. A., Griggs, A., Cerqueira, G., Van Tonder, A., Holden, M. T. G., et al. (2012). Comparative genomics of vancomycin-resistant Staphylococcus aureus strains and their positions within the clade most commonly associated with Methicillin-resistant S. aureus hospital-acquired infection in the United States. MBio 3:e112.
Lipinski, K. J., Farslow, J. C., Fitzpatrick, K. A., Lynch, M., Katju, V., and Bergthorsson, U. (2011). High spontaneous rate of gene duplication in Caenorhabditis elegans. Curr. Biol. 21, 306–310. doi: 10.1016/j.cub.2011.01.026
Long, M., Betrán, E., Thornton, K., and Wang, W. (2003). The origin of new genes: glimpses from the young and old. Nat. Rev. Genet. 4, 865–875. doi: 10.1038/nrg1204
Lynch, M., Sung, W., Morris, K., Coffey, N., Landry, C. R., Dopman, E. B., et al. (2008). A genome-wide view of the spectrum of spontaneous mutations in yeast. Proc. Natl. Acad. Sci. U.S.A. 105, 9272–9277. doi: 10.1073/pnas.0803466105
Makarova, K. S., Ponomarev, V. A., and Koonin, E. V. (2001). Two C or not two C: recurrent disruption of Zn-ribbons, gene duplication, lineage-specific gene loss, and horizontal gene transfer in evolution of bacterial ribosomal proteins. Genome Biol. 2:RESEARCH0033. doi: 10.1186/gb-2001-2-9-research0033
McGuinness, W. A., Malachowa, N., and DeLeo, F. R. (2017). Vancomycin resistance in Staphylococcus aureus. Yale J Biol Med 90, 269–281.
Miller, K. A., Phillips, R. S., Mrázek, J., and Hoover, T. R. (2013). Salmonella utilizes D-glucosaminate via a mannose family phosphotransferase system permease and associated enzymes. J. Bacteriol. 195, 4057–4066. doi: 10.1128/jb.00290-13
Moore, C. M., Gaballa, A., Hui, M., Ye, R. W., and Helmann, J. D. (2005). Genetic and physiological responses of Bacillus subtilis to metal ion stress. Mol. Microbiol. 57, 27–40. doi: 10.1111/j.1365-2958.2005.04642.x
Moore, C. M., and Helmann, J. D. (2005). Metal ion homeostasis in Bacillus subtilis. Curr. Opin. Microbiol. 8, 188–195. doi: 10.1016/j.mib.2005.02.007
Murray, B. E. (2000). Vancomycin-resistant enterococcal infections. N. Engl. J. Med. 342, 710–721. doi: 10.1056/NEJM200003093421007
Naimi, T. S., LeDell, K. H., Boxrud, D. J., Groom, A. V., Steward, C. D., Johnson, S. K., et al. (2001). Epidemiology and clonality of community-acquired methicillin-resistant Staphylococcus aureus in Minnesota, 1996–1998. Clin. Infect. Dis. 33, 990–996. doi: 10.1086/322693
Ondov, B. D., Treangen, T. J., Melsted, P., Mallonee, A. B., Bergman, N. H., Koren, S., et al. (2016). Mash: fast genome and metagenome distance estimation using MinHash. Genome Biol. 17:132. doi: 10.1186/s13059-016-0997-x
Paganelli, F. L., Huebner, J., Singh, K. V., Zhang, X., van Schaik, W., Wobser, D., et al. (2016). Genome-wide screening identifies phosphotransferase system permease BepA to be involved in Enterococcus faecium endocarditis and biofilm formation. J. Infect. Dis. 214, 189–195. doi: 10.1093/infdis/jiw108
Paulsen, I. T., Banerjei, L., Myers, G. S. A., Nelson, K. E., Seshadri, R., Read, T. D., et al. (2003). Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 299, 2071–2074. doi: 10.1126/science.1080613
Pearson, W. R. (2013). An introduction to sequence similarity (“homology”) searching. Curr. Protoc. Bioinformat. Chapter 3:Unit3.1. doi: 10.1002/0471250953.bi0301s42
Popella, P., Krauss, S., Ebner, P., Nega, M., Deibert, J., and Götz, F. (2016). VraH is the third component of the Staphylococcus aureus VraDEH system involved in gallidermin and daptomycin resistance and pathogenicity. Antimicrob. Agents Chemother. 60, 2391–2401. doi: 10.1128/aac.02865-15
R Development Core Team (2008). R Development Core Team R-project.org. Available online at: http://www.R-project.org.
Richardson, A. R., Libby, S. J., and Fang, F. C. (2008). A nitric oxide-inducible lactate dehydrogenase enables Staphylococcus aureus to resist innate immunity. Science 319, 1672–1676. doi: 10.1126/science.1155207
Riehle, M. M., Bennett, A. F., and Long, A. D. (2001). Genetic architecture of thermal adaptation in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 98, 525–530. doi: 10.1073/pnas.021448998
Romero, D., and Palacios, R. (1997). Gene amplification and genomic plasticity in prokaryotes. Annu. Rev. Genet. 31, 91–111. doi: 10.1146/annurev.genet.31.1.91
Ruiz-Cruz, S., Espinosa, M., Goldmann, O., and Bravo, A. (2015). Global regulation of gene expression by the MafR protein of Enterococcus faecalis. Front. Microbiol. 6:1521. doi: 10.3389/fmicb.2015.01521
Sauvageot, N., Mokhtari, A., Joyet, P., Budin-Verneuil, A., Blancato, V. S., Repizo, G. D., et al. (2017). Enterococcus faecalis uses a phosphotransferase system permease and a host colonization-related ABC transporter for maltodextrin uptake. J. Bacteriol. 199:e00878-16.
Serres, M. H., Kerr, A. R. W., McCormack, T. J., and Riley, M. (2009). Evolution by leaps: gene duplication in bacteria. Biol. Direct. 4:46. doi: 10.1186/1745-6150-4-46
Shore, A. C., and Coleman, D. C. (2013). Staphylococcal cassette chromosome mec: recent advances and new insights. Int. J. Med. Microbiol. 303, 350–359. doi: 10.1016/j.ijmm.2013.02.002
Stentzel, S., Teufelberger, A., Nordengrün, M., Kolata, J., Schmidt, F., van Crombruggen, K., et al. (2017). Staphylococcal serine protease-like proteins are pacemakers of allergic airway reactions to Staphylococcus aureus. J. Allergy Clin. Immunol. 139, 492.e–500.e. doi: 10.1016/j.jaci.2016.03.045
Sun, S., Ke, R., Hughes, D., Nilsson, M., and Andersson, D. I. (2012). Genome-wide detection of spontaneous chromosomal rearrangements in bacteria. PLoS ONE 7:e42639. doi: 10.1371/journal.pone.0042639
Thammavongsa, V., Missiakas, D. M., and Schneewind, O. (2013). Staphylococcus aureus degrades neutrophil extracellular traps to promote immune cell death. Science 342, 863–866. doi: 10.1126/science.1242255
Titus, B. C., and Irber, L. (2016). Sourmash: a library for MinHash sketching of DNA. J. Open Source Softw. 1:27. doi: 10.21105/joss.00027
Torell, E., Molin, D., Tano, E., Ehrenborg, C., and Ryden, C. (2005). Community-acquired pneumonia and bacteraemia in a healthy young woman caused by methicillin-resistant Staphylococcus aureus (MRSA) carrying the genes encoding Panton-Valentine leukocidin (PVL). Scand. J. Infect. Dis. 37, 902–904. doi: 10.1080/00365540500348945
Toussaint, J.-P., Farrell-Sherman, A., Feldman, T. P., Smalley, N. E., Schaefer, A. L., Greenberg, E. P., et al. (2017). Gene duplication in Pseudomonas aeruginosa improves growth on adenosine. J. Bacteriol. 199:e261-17.
Treangen, T. J., and Rocha, E. P. C. (2011). Horizontal transfer, not duplication, drives the expansion of protein families in prokaryotes. PLoS Genet. 7:e1001284. doi: 10.1371/journal.pgen.1001284
van der Woude, M. W., and Henderson, I. R. (2008). Regulation and function of Ag43 (flu). Annu. Rev. Microbiol. 62, 153–169. doi: 10.1146/annurev.micro.62.081307.162938
Werner, G., Coque, T. M., Hammerum, A. M., Hope, R., Hryniewicz, W., Johnson, A., et al. (2008). Emergence and spread of vancomycin resistance among enterococci in Europe. Euro. Surveill. 13:19046.
Xia, G., and Wolz, C. (2014). Phages of Staphylococcus aureus and their impact on host evolution. Infect. Genet. Evol. 21, 593–601. doi: 10.1016/j.meegid.2013.04.022
Zarb, P., Coignard, B., Griskeviciene, J., Muller, A., Vankerckhoven, V., Weist, K., et al. (2012). The european centre for disease prevention and control (ECDC) pilot point prevalence survey of healthcare-associated infections and antimicrobial use. Eur. Surveill. 17:20316. doi: 10.2807/ese.17.46.20316-en
Zhang, J. (2003). Evolution by gene duplication: an update. Trends Ecol. Evol. 18, 292–298. doi: 10.1016/s0169-5347(03)00033-8
Zhang, J., Liu, H., Zhu, K., Gong, S., Dramsi, S., Wang, Y.-T., et al. (2014). Antiinfective therapy with a small molecule inhibitor of Staphylococcus aureus sortase. Proc. Natl. Acad. Sci. U.S.A. 111, 13517–13522. doi: 10.1073/pnas.1408601111
Zhang, X., Bierschenk, D., Top, J., Anastasiou, I., Bonten, M. J. M., Willems, R. J. L., et al. (2013a). Functional genomic analysis of bile salt resistance in Enterococcus faecium. BMC Genomics 14:299. doi: 10.1186/1471-2164-14-299
Zhang, X., Top, J., de Been, M., Bierschenk, D., Rogers, M., Leendertse, M., et al. (2013b). Identification of a genetic determinant in clinical Enterococcus faecium strains that contributes to intestinal colonization during antibiotic treatment. J. Infect. Dis. 207, 1780–1786. doi: 10.1093/infdis/jit076
Keywords: gene duplication, Staphylococcus aureus, Enterococcus faecium, Enterococcus faecalis, bacterial genomics
Citation: Sanchez-Herrero JF, Bernabeu M, Prieto A, Hüttener M and Juárez A (2020) Gene Duplications in the Genomes of Staphylococci and Enterococci. Front. Mol. Biosci. 7:160. doi: 10.3389/fmolb.2020.00160
Received: 19 May 2020; Accepted: 24 June 2020;
Published: 23 July 2020.
Edited by:
Tatiana Venkova, Fox Chase Cancer Center, United StatesReviewed by:
Juan Carlos Alonso, National Center for Biotechnology (CNB), SpainCopyright © 2020 Sanchez-Herrero, Bernabeu, Prieto, Hüttener and Juárez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonio Juárez, YWp1YXJlekB1Yi5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.