
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
CORRECTION article
Front. Microbiol. , 28 January 2025
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1561085
 Yuzhi Huang1†
Yuzhi Huang1† Lanmeng Xu1†Hang He2
Lanmeng Xu1†Hang He2 Lijuan Peng1Qinfeng Liao2Kun Wan1
Lijuan Peng1Qinfeng Liao2Kun Wan1 Simeng Qin1Lijing Cao3*Jie Zhang1*
Simeng Qin1Lijing Cao3*Jie Zhang1*A Corrigendum on
Effects of rosemary extract and its residue on production, immune performance, and gut microbiota in geese
by Huang, Y., Xu, L., He, H., Peng, L., Liao, Q., Wan, K., Qin, S., Cao, L., and Zhang, J. (2025). Front. Microbiol. 15:1483626. doi: 10.3389/fmicb.2024.1483626
In the published article, there was an error in Table 1 as published. The composition of basal diet was incomplete as it was missing the ingredient “Rice bran” with a content of 13.40%. The corrected Table 1 appear below.
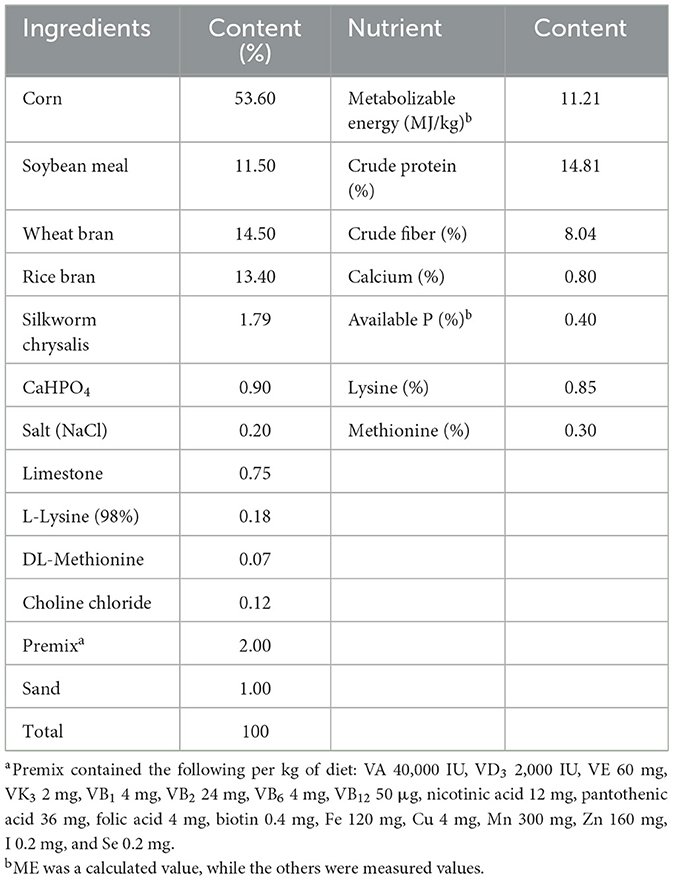
Table 1. Composition and nutrient level of basal diet (air-dry basis).
The authors apologize for this error and state that this does not change the scientific conclusions of the article in any way. The original article has been updated.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Keywords: rosemary extract, rosemary extract residue, production performance, immune performance, gut microbiota, geese
Citation: Huang Y, Xu L, He H, Peng L, Liao Q, Wan K, Qin S, Cao L and Zhang J (2025) Corrigendum: Effects of rosemary extract and its residue on production, immune performance, and gut microbiota in geese. Front. Microbiol. 16:1561085. doi: 10.3389/fmicb.2025.1561085
Received: 15 January 2025; Accepted: 16 January 2025;
Published: 28 January 2025.
Approved by:
Frontiers Editorial Office, Frontiers Media SA, SwitzerlandCopyright © 2025 Huang, Xu, He, Peng, Liao, Wan, Qin, Cao and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jie Zhang, emhhbmdqaWU4MTNAMTYzLmNvbQ==; Lijing Cao, NTE3Mjk2NTY2QHFxLmNvbQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.