
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 24 February 2025
Sec. Microbiotechnology
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1542468
This article is part of the Research TopicExpert Opinions: Save the Microbes to Save the PlanetView all 12 articles
 Ani Paloyan1
Ani Paloyan1 Mane Tadevosyan2
Mane Tadevosyan2 Diana Ghevondyan2,3
Diana Ghevondyan2,3 Lev Khoyetsyan1
Lev Khoyetsyan1 Mariam Karapetyan1
Mariam Karapetyan1 Armine Margaryan2,3
Armine Margaryan2,3 Garabed Antranikian4
Garabed Antranikian4 Hovik Panosyan2,3*
Hovik Panosyan2,3*Polyhydroxyalkanoates (PHAs) are biobased and biodegradable polymers that offer a sustainable alternative to conventional plastics, addressing the escalating concerns over plastic pollution. While their environmental advantages are well-documented, the efficient degradation of PHAs in natural and engineered environments remains a critical component of their lifecycle. This review provides a comprehensive overview of PHA-degrading bacteria isolated from diverse ecosystems and highlights the pivotal role of PHA depolymerases in achieving PHA circularity. Microbial adaptation to diverse environmental conditions, such as extreme temperatures, salinity, and pH, significantly influences enzymes properties, including the stability, activity, and substrate specificity of PHA-degrading enzymes. These adaptations often enhance enzyme, performance, enabling functionality under challenging conditions. Consequently, extremophilic microorganisms are invaluable resources for discovering and engineering robust PHA depolymerases for industrial and environmental applications. This review underscores the urgent need for further research to improve the ecological and economic sustainability of PHA waste management.
The growing global reliance on plastics has created significant environmental challenges, primarily due to their persistence in natural ecosystems and the difficulties associated with their disposal (Covello et al., 2024). Conventional petrochemical-based plastics degrade extremely slowly, accumulating in terrestrial and aquatic environments and contributing to widespread pollution. To address these issues, bioplastics have emerged as sustainable alternatives, offering the potential to reduce environmental harm while retaining the versatility of traditional plastics (Yadav and Nikalje, 2024).
Among the various types of bioplastics, polyhydroxyalkanoates (PHAs) have garnered significant attention as promising biodegradable polymers. Synthesized by a wide range of microorganisms as intracellular carbon and energy reserves, PHAs are renewable, biodegradable, and environmentally friendly. These properties make PHAs particularly appealing for combating global plastic pollution (Getino et al., 2024). Furthermore, PHAs are biocompatible, enabling their application in medical fields such as drug delivery systems and surgical implants, thus broadening their potential uses.
However, the successful adoption of PHAs depends on a thorough understanding of their complete lifecycle, particularly their degradation in natural and engineered environments. PHA breakdown is essential for reintegrating these materials into natural biogeochemical cycles. This process heavily relies on microbial activity, where specialized bacteria and fungi produce extracellular depolymerases to hydrolyze PHA polymers into monomers and oligomers, which can then serve as carbon sources for microbial growth (Ray and Kalia, 2017). These microorganisms are found in diverse environments, including terrestrial soils, freshwater, marine ecosystems, and extreme conditions like geothermal springs and cryogenic soils, demonstrating their ecological adaptability.
The enzymatic degradation of PHAs is a critical factor in their biodegradability. Enzymes such as PHA depolymerases play a pivotal role in this process by hydrolyzing the ester bonds within PHA polymer chains, generating products that microorganisms can further metabolize (Millan and Hanik, 2023). PHA depolymerases exhibit remarkable specificity and efficiency, targeting short-chain-length (SCL-PHA) or medium-chain-length (MCL-PHA) polymers (Knoll et al., 2009). The efficiency of these enzymes is influenced by factors such as polymer composition, crystallinity, environmental conditions, and the presence of cofactors (Bhatt et al., 2011).
Despite significant progress in understanding PHA-degrading microorganisms and their enzymes, several knowledge gaps persist. For example, further research is needed to elucidate the interactions between environmental factors (e.g., pH, temperature, salinity) and enzymatic activity. Additionally, investigating the mechanisms underlying microbial adaptation to extreme environments, such as hot springs or cryogenic soils, could uncover novel biocatalysts. Addressing these gaps will enhance the development of efficient waste management systems and industrial processes for bioplastic recycling.
This review provides a comprehensive overview of the diversity and ecological distribution of PHA-degrading bacteria and their enzymes. It highlights the enzymatic mechanisms involved in PHA degradation, the structural and functional characteristics of depolymerases, and the factors influencing their activity. Furthermore, it explores the potential applications of PHA-degrading microorganisms and their enzymes in bioplastic recycling, waste management, and sustainable biotechnology. By synthesizing recent advances, this review aims to guide future research and innovation in leveraging microbial systems to address the global plastic crisis.
PHAs are biodegradable polymers that undergo degradation through various mechanisms, including photooxidative, catalytic, thermal, and mechanical degradation. Among these, microbial degradation plays a crucial role in the complete breakdown of PHAs in natural environments.
Photooxidative degradations occurs when PHA is exposed to light, particularly UV radiation. The energy from light causes the formation of free radicals within the polymer chains, leading to chain scission and degradation. This process is relatively slow and highly dependent on the intensity and duration of UV exposure (Lalonde et al., 2025).
Catalytic degradation involves the use of catalytic entities, including enzymes, transition metal ions (for the Fenton reaction), nanozymes are used to accelerate the breakdown of PHA. Catalysts specifically target the ester bonds in PHA, leading to faster degradation. However, the requirement for specific catalysts can make the process more complex and costly (Zandieh et al., 2024).
Thermal degradation occurs at high temperatures causing random chain scission in PHA polymer chains. This process results in the formation of smaller molecules, such as oligomers and monomers. While thermal degradation can be effective, it often produces unwanted byproducts and requires substantial energy input (Kunioka and Doi, 1990; Hablot et al., 2008).
Mechanical degradation involves physical forces such as grinding, shear stress, or mechanical wear that break down PHA polymers. This process primarily leads to fragmentation rather than chemical degradation. However it does not result in complete molecular breakdown but rather physical disintegration (Lin et al., 2020).
Microbial degradation of PHA is widely distributed among bacteria. PHA-degrading bacteria have been identified in diverse environmental niches, including soil (geothermal and cryogenic), hot springs, freshwater, and marine ecosystems, indicating the widespread distribution of PHA-degrading microorganisms (Hori et al., 2020; Volova et al., 2010; Boyandin et al., 2012; Takeda et al., 2002). These microorganisms play a vital role in the natural biodegradation of PHAs.
Soil environments harbor a diverse array of PHA-degrading bacteria across various genera, including Acidovorax, Acinetobacter, Arthrobacter, Bacillus, Burkholderia, Cytophaga, Rhodococcus, Cupriavidus, Mycobacterium, Nocardiopsis, Pseudomonas, Stenotrophomonas, Paraburkholderia, Streptomyces, Rhizobium, Variovorax, Xanthomonas, Alcaligenes, Lihuaxuella, Thermus, Schlegelella, Paenibacillus and Zoogloea (Hori et al., 2020; Boyandin et al., 2012; Volova et al., 2006; Prudnikova et al., 2021; Thomas et al., 2022; Pham et al., 2024). These bacteria exhibit significant functional diversity, with some strains demonstrating rapid degradation rates under laboratory conditions. For instance, Pseudomonas sp. DSDY0501, isolated from activated sludge, completely degraded poly (3-hydroxybutyrate) (PHB) films within 21 h in liquid culture, utilizing PHB as its sole carbon source (Di et al., 2019). Similarly, strains such as Bacillus subtilis PLA 2.3.1 and PET 2.2.1, Streptomyces griseorubens ACTY2-2, Rhizobium pusense PLA1-1, Priestia aryabhattai (A13, A34, L5, N7), isolated from plastic-contaminated soils in Armenia, have demonstrated the ability to degrade PHBV and PHBH. Additionally, Priestia aryabhattai A34, Priestia megaterium L7 and N7 were found to be capable of degrading PHB polymers (Khoyetsyan et al., 2024; Tadevosyan et al., 2024).
PHAs, biodegradable bioplastics, produced by various microorganisms as intracellular carbon and energy reserves (Arnosti, 2011), have also been targeted by Pseudomonas strains isolated from soil and compost. These strains exhibit promising potential for bioremediation and waste management, particularly in composting systems where they facilitate PHA degradation (Jaeger and Rosenau, 2004; Spasic et al., 2018).
The ability of Pseudomonas species to degrade PHAs, coupled with their production of specific depolymerases, highlights their importance in the environmental breakdown of bioplastics and the recycling of biodegradable waste (Arnosti, 2011; Jaeger and Rosenau, 2004).
Interestingly, the homopolymer poly-3-hydroxybutyrate (poly-3-HB) degraded faster than its copolymer counterpart, poly-3-HB/3-HV, demonstrating the impact of polymer composition on biodegradability. Additionally, microbial communities associated with the polymers are habitat-specific and differ significantly from those in the surrounding soil. Key bacterial genera such as Burkholderia, Bacillus, and Cupriavidus, alongside fungal species like G. butleri and Penicillium sp., have been identified as active degraders.
The interplay between environmental conditions and microbial ecosystems significantly influences PHA degradation, contributing to the variability of biodegradation rates across different geographies (Boyandin et al., 2013).
While soil environments host a diverse array of PHA-degrading bacteria with significant functional diversity, marine ecosystems also provide a unique habitat for these microorganisms, requiring adaptations to salinity and host interactions. Genera such as Enterobacter, Bacillus, Comamonas and Gracilibacillus have been identified as key PHA degraders in seawater and marine habitats (Volova et al., 2010; Boyandin et al., 2012; Volova et al., 2006; Schirmer et al., 1993).
Additional PHA degraders include Alcaligenes faecalis (from activated sludge), Ilyobacter delafieldii (from lake water and estuarine sediment), Caldimonas manganoxidans (from hot springs), and other species isolated from anaerobic sewage sludge (Takeda et al., 2002; Schirmer et al., 1993). Interestingly, some strains have been isolated from marine organisms, such as Acidovorax spp. from Siberian sturgeon, Acinetobacter spp. from European sea bass, and Ochrobactrum spp. from giant river prawns (Liu et al., 2010).
Studies have also identified microbial degradation of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBH) polymers by strains such as Glaciecola lipolytica E11, Aestuariibacter halophilus S23, and Pseudoalteromonas lipolytica S35. These microbes form biofilms on PHBH polymer surfaces within 3 days of incubation at 23°C in seawater. Notably, these strains do not degrade other types of bioplastics, demonstrating specificity toward PHBH. The formation of clear zones observed around colonies on PHBH-containing agar plates further confirm their PHA-degrading activity (Morohoshi et al., 2018).
Several bacterial species isolated from various marine environments have been identified as capable of degrading PHA, with Pseudomonas species playing a particularly prominent role in this process. Pseudomonas aeruginosa, Pseudomonas fluorescens, and Pseudomonas putida have been shown to produce extracellular PHA depolymerases —enzymes that break down PHA into smaller monomers, which can then be utilized by bacteria as a carbon source (Arnosti, 2011).
A novel tropical marine bacterium, Pseudomonas sp. NRRL B-300083, has demonstrated the ability to degrade PHB/PHV copolymers in seawater. Similarly, denitrifying bacteria such as Comamonas testosteroni and certain strains of the Variovorax genus have been found to degrade PHB and its copolymers, with faster degradation observed in copolymers containing higher proportions of hydroxyvalerate (Volova et al., 2006).
Extremophilic habitats, such as geothermal springs and cryogenic soils, host bacteria that exhibit remarkable adaptations to challenging conditions, including extreme temperatures, acidic or basic pH, and high salinity. These environments exert unique selection pressures that drive the development of specialized microbial adaptations. For example, Microbulbifer sp. SOL03 and Bacillus vietnamensis SOL04 demonstrate PHA-degrading activity across a wide temperature range (10–42°C), while Halobacillus trueperi SOL01 tolerates salinity levels as high as 15% NaCl (Park et al., 2021). Additionally, halophilic actinomycetes such as Nocardiopsis aegyptia efficiently degrade PHB and PHBV copolymers, achieving up to 89.94% weight loss within 30 days (Ghanem et al., 2005).
Thermophilic bacteria capable of degrading PHBH polymers have also been isolated from geothermal springs. For instance, Anoxybacillus sp. K99 and Anoxybacillus karvacharensis K1, isolated from the Karvachar geothermal spring in Nagorno-Karabakh, Parageobacillus toebii H-70 isolated from Hankavan hot spring, have demonstrated the ability to degrade PHBH at 55°C (Khoyetsyan et al., 2024; Tadevosyan et al., 2024).
Cryogenic soils in Siberia’s subarctic regions have yielded PHB-degrading bacteria such as Bacillus pumilus, Paraburkholderia sp., Pseudomonas sp., Rhodococcus sp., Stenotrophomonas rhizophila, Streptomyces prunicolor, and Variovorax paradoxus (Prudnikova et al., 2021). Among these, Ralstonia sp. exhibited the highest specific activity, maintaining functionality even at low pH levels (3.3–3.7).
The ability of PHA-degrading bacteria to thrive in extreme conditions is closely tied to the specialized characteristics of their enzymes, such as enhanced temperature and pH stability. These unique adaptations, evident across diverse extremophilic habitats, highlight the untapped potential of these strains for industrial applications under harsh conditions. By bridging the gap between microbial ecology and biotechnology, extremophilic PHA-degrading bacteria offer innovative solutions for industrial processes and waste management in challenging environments.
Several analytical methods are employed to study the microbial degradation of plastics, particularly bioplastics like PHAs. These methods can be categorized based on their purpose, such as visual assessment, structural analysis, chemical analysis, and molecular approaches.
Microbial degradation of bioplastics is often evidenced by the formation of clear zones around microbial colonies on plates where bioplastics serve as the sole carbon source. The diameter of this zone is commonly used to quantitative measure of biodegradation efficiency (Tezuka et al., 2004; Lee et al., 2005; Kumaravel et al., 2010). Advanced analytical techniques, such as scanning electron microscopy (SEM) and Fourier Transform Infrared (FTIR) spectroscopy, play crucial roles in understanding the structural and molecular changes occurring during microbial degradation. SEM is widely used to visualize the breakdown of PHAs, revealing surface irregularities and structural instability caused by microbial activity (Wu, 2009; Tachibana et al., 2013). FTIR spectroscopy complements SEM by detecting changes in bond intensities, providing insights into the enzymatic action on polymer chains.
A summary of the methods used for assessing plastic degradation is presented in Table 1.
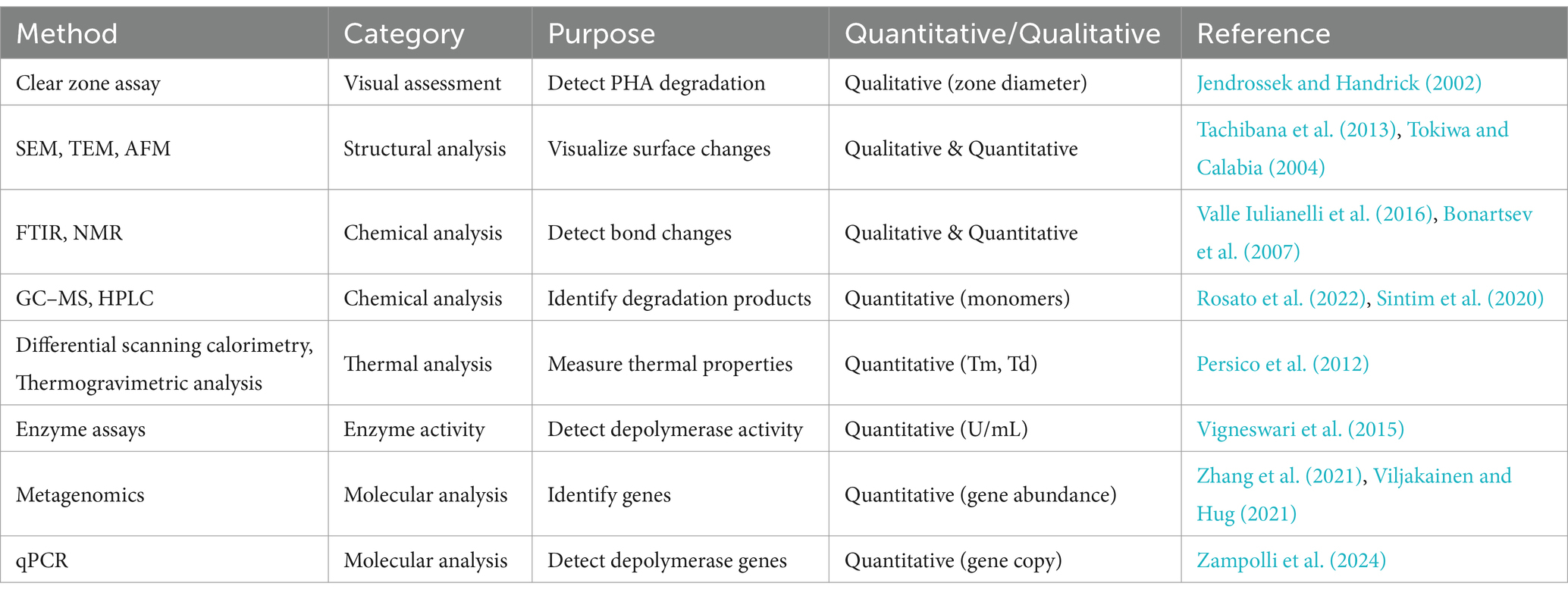
Table 1. Comprehensive list of commonly used methods to study the microbial degradation of bioplastics.
The efficiency of PHA biodegradation is influenced by several factors, including the polymer’s physicochemical properties and environmental conditions. Key polymer properties affecting degradation include shape, size, molecular weight, functional groups, crystallinity, hydrophobicity or hydrophilicity, and the presence of additives. Environmental factors such as temperature, light, oxygen availability, salinity, pH, and nitrate concentrations also play significant roles. Abiotic factors, such as temperature and light, enhance polymer accessibility to microbial degradation, while biotic factors, particularly enzyme activity, directly drive the breakdown process (Volova et al., 2010; Boyandin et al., 2013; Bonartseva et al., 2002). For instance, increased salinity typically reduces degradability, whereas higher temperatures and light levels promote it.
Seasonal studies conducted in the Sea of Japan and the Baltic Sea have demonstrated that PHA degradation rates are higher during the summer, driven by increased microbial activity under warmer temperatures (Zandieh et al., 2024; Tokiwa and Calabia, 2004). PHB degradation was most effective under aerobic conditions, with minimal degradation observed in anaerobic environments. The addition of nitrate further enhanced degradation across all aeration levels by stimulating denitrifying microbes. Notably, PHB degradation without nitrate occurred at 20°C but was inhibited at 5°C, whereas nitrate supplementation enabled degradation even at 5°C (Bonartseva et al., 2002).
Tropical soils also exhibit higher PHA degradation rates due to favorable climatic conditions, including elevated humidity and diverse microbial communities (Li et al., 2016).
Studies conducted in Lake Shira, a saline lake in Russia, revealed that PHA degradation occurs under both oxygen-rich (aerobic) and oxygen-deprived (anaerobic) conditions,. Under aerobic conditions, microorganisms convert PHAs into carbon dioxide and water, whereas anaerobic conditions yield methane and carbon dioxide (Volova et al., 2010).
The PHB degradation performance of Cutibacterium sp. SOL05 was notable, achieving 66% degradation within 7 days and 74% within 10 days. The addition of carbon sources significantly enhanced this efficiency. Among tested carbon sources-glucose, galactose, sucrose, xylose, lactose, and fructose-lactose had the most pronounced effect. A 1% lactose concentration increased degradation by 1.5 times compared to the control (Park et al., 2021).
The efficiency of PHA biodegradation is closely linked to the polymer’s physical and chemical properties. The degradation rate primarily depends on (i) crystallinity, (ii) the type of polymer (homopolymer vs. copolymer), and (iii) the copolymeric structure. Due to its high crystallinity, PHB is generally more resistant to biodegradation. Copolymers such as PHBV, which incorporate hydroxyvalerate into PHB, degrade more readily due to their increased amorphous regions. A higher hydroxyvalerate content reduces crystallinity, thereby enhancing the polymer’s biodegradability by facilitating enzyme adhesion and accelerating degradation (Meereboer et al., 2020). Films with larger surface areas degrade more rapidly than pellets, which have limited polymer-water interfaces and less space for microbial colonization (Volova et al., 2010).
The degradation of 3-PHB/3-PHV copolymers was faster than that of 3-PHB, with degradation rates increasing as the percentage of hydroxyvalerate in the copolymer rose. For example, in seawater, 58% of 3-PHB and 54% of 3-PHB/3-PHV films degraded after 160 days (Volova et al., 2010). It is worth noting that most P(3HB)- or P(3 HV)-degrading bacteria cannot degrade PHAs with six or more carbon atoms, suggesting that larger PHA monomers require specialized enzymes or microbes for degradation (Schirmer et al., 1993).
Additives, such as plasticizers used in polymer production to enhance physical properties like strength and flexibility, also affect degradation rates. While plasticizers improve PHB’s durability, they may alter its susceptibility to microbial activity. For instance, PHB films containing 10 and 20% tributyl citrate exhibited enhanced biodegradability. Tributyl citrate accelerated PHB degradation by 88% within the first day, although the final degradation levels after 3 days were comparable to non-plasticized films. By day three, all films achieved over 90% degradation. Despite slightly inhibiting the growth of Microbulbifer sp. SOL66, the degradation efficiency of plasticized PHB remained high (Cho et al., 2022). Adding plasticizers to PHB enhances its melt processability and optimizes its properties, which can potentially improve its biodegradability (Panaitescu et al., 2017). However, while some plasticizers are biodegradable, others are not. Thus, the choice of plasticizer significantly impacts the overall biodegradability of the polymer. Biodegradable plasticizers, such as certain citrate esters, can enhance the biodegradability of PHAs, whereas non-biodegradable plasticizers, like some phthalate-based compounds, may persist in the environment and potentially inhibit the biodegradation process (Meereboer et al., 2020). Therefore, while plasticizers can improve the biodegradability of PHAs by modifying their physical properties, the biodegradability of the plasticizers themselves is a crucial factor to consider in the design of environmentally friendly biopolymer systems. These findings highlight the critical interplay between environmental conditions, structural properties of PHAs, and microbial activity in determining biodegradation efficiency (Bonartseva et al., 2002).
Asummary of the factors influencing microbial PHA degradation is presented in Table 2.
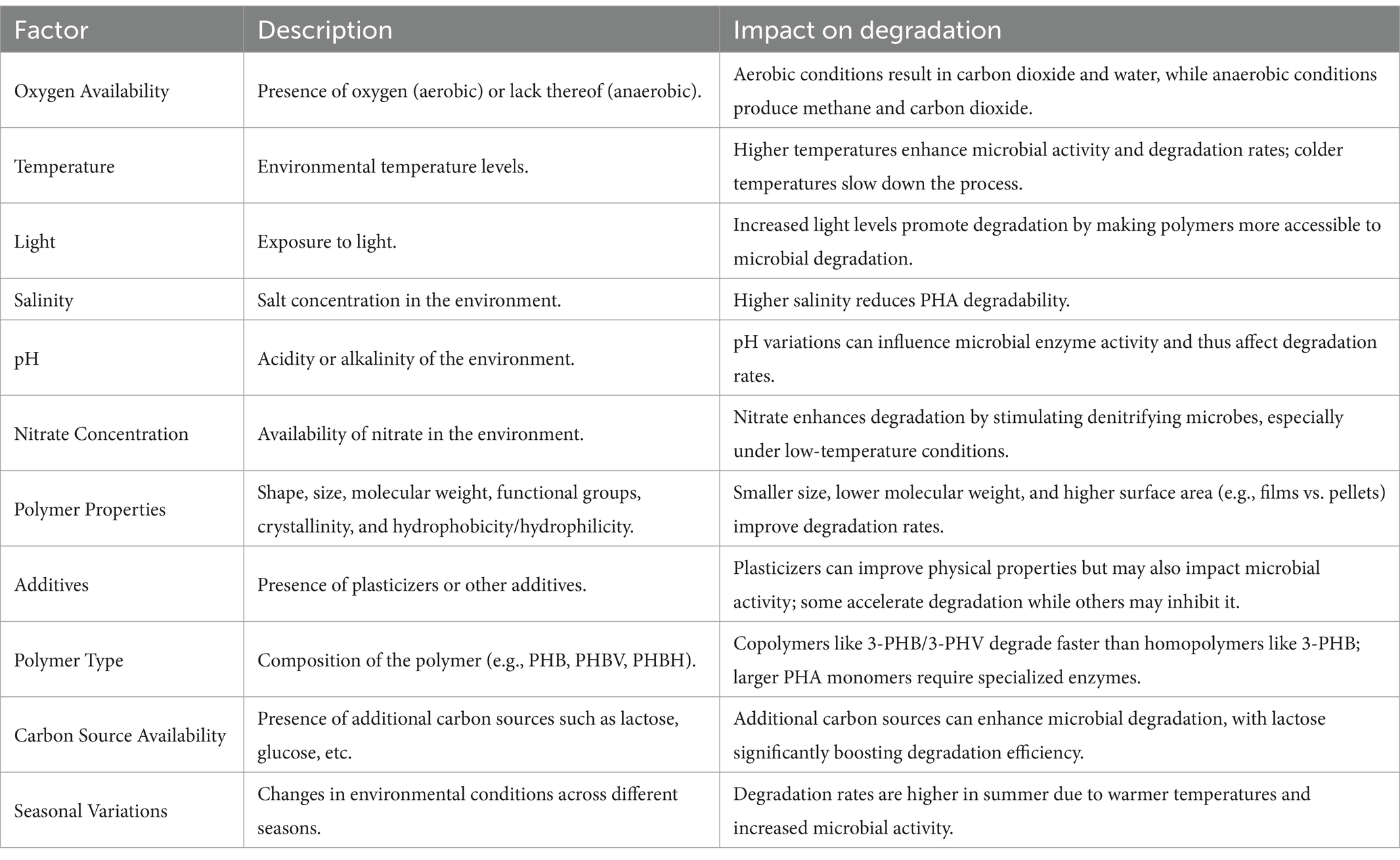
Table 2. Factors influencing microbial PHA degradation.
PHA depolymerases (EC 3.1.1.75, EC 3.1.1.76) are enzymes that catalyze the degradation of PHAs, breaking them into smaller molecules such as oligomers, dimers, and monomers. These hydroxyalkanoic acid monomers serve as carbon and energy sources for microorganisms, playing an essential role in microbial life cycles and PHA recycling. PHA depolymerases also hold great promise for biotechnological applications, particularly in the sustainable management of bioplastics.
PHA depolymerases are categorized based on their localization and substrate specificity.
• Extracellular (e-PHA depolymerases): Act on extracellular paracrystalline PHA granules.
• Intracellular (i-PHA depolymerases): Degrade “native” PHAs stored as intracellular granules.
• Periplasmic depolymerases: Found in the periplasmic space of certain bacteria, playing roles in microbial metabolism and predation.
• SCL-PHB Depolymerases: Target short-chain-length PHAs such as poly (3-hydroxybutyrate) (PHB), producing 3-hydroxybutyrate monomers. Examples include depolymerases from Rhodospirillum rubrum, Alcaligenes faecalis, Pseudomonas spp., and Bacillus spp.
• MCL-PHA Depolymerases: Degrade medium-chain-length PHAs such as poly (3-hydroxyhexanoate) and poly (3-hydroxyoctanoate), with microbial producers like Pseudomonas spp.
Intracellular PHA (i-PHA) depolymerases degrade native PHAs stored as amorphous granules. Once the protective protein layer surrounding these granules is, PHAs denature into a paracrystalline form, becoming accessible to extracellular PHA depolymerases. Notably, Pseudomonas lemoignei produces an extracellular PHB depolymerase (PhaZ7), that uniquely degrades native PHB granules despite lacking significant similarity to conventional hydrolases (Braaz et al., 2003).
Periplasmic PHA depolymerases, though less studied, have been identified in Rhodospirillum rubrum and Bdellovibrio bacteriovorus. In B. bacteriovorus, these enzymes contribute to its predatory lifestyle by degrading host PHAs during its developmental cycle (Zampolli et al., 2024; Bonartseva et al., 2002; Handrick et al., 2004). In R. rubrum, the physiological role of this enzyme remains unclear, as its PHB granules are stored intracellularly rather than in the periplasm.
e-PHA depolymerases are further classified into two major superfamilies based on substrate specificity:
1. e-PHASCL depolymerases (EC3.1.1.75): Degrade short-chain-length PHAs (3 to 5 carbon atoms) including PHB, poly (3-hydroxyvalerate) (PHV), poly (3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV), poly (4-hydroxybutyrate) (P4HB), poly (3-hydroxybutyrate-co-4-hydroxybutyrate) (P(3HB-co-4HB)), poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate) (PHBVH), poly (3-hydroxybutyrate-co-3-hydroxy-methylpropionate) (P(3HB-co-3MP)), etc.
2. e-PHAMCL depolymerases (EC3.1.1.76): Degrade medium-chain-length PHAs (6 to 14 carbon atoms) such as poly(3-hydroxyoctanoate) (P(3HO)), polyhydroxyoctanoate-co-hexanoate (P(HO-co-HH)) poly(hydroxyoctanoate-co-hydroxyhexanoate) (P(HO-co-HX)), poly(3-hydroxy-5-phenylvalerate) (PHPV), etc.
According to the PHA Depolymerase Engineering Database (Knoll et al., 2009), e-PHA depolymerases can be further categorized based on their catalytic domain:
• e-PHASCL depolymerases (Type 1 catalytic domain): Characterized by an N-terminal oxyanion hole.
• e-PHASCL depolymerases (Type 2 catalytic domain): Characterized by a C-terminal oxyanion hole.
• e-PHAMCL depolymerases: Specific for medium-chain-length PHAs.
• e-PHASCL depolymerase: Only active against native PHA granules, with Paucimonas lemoignei as a notable representative (Handrick et al., 2001).
Aschematic representation of this classification is shown in Figure 1, providing insights into the diversity and functionality of PHA depolymerases within microbial systems.
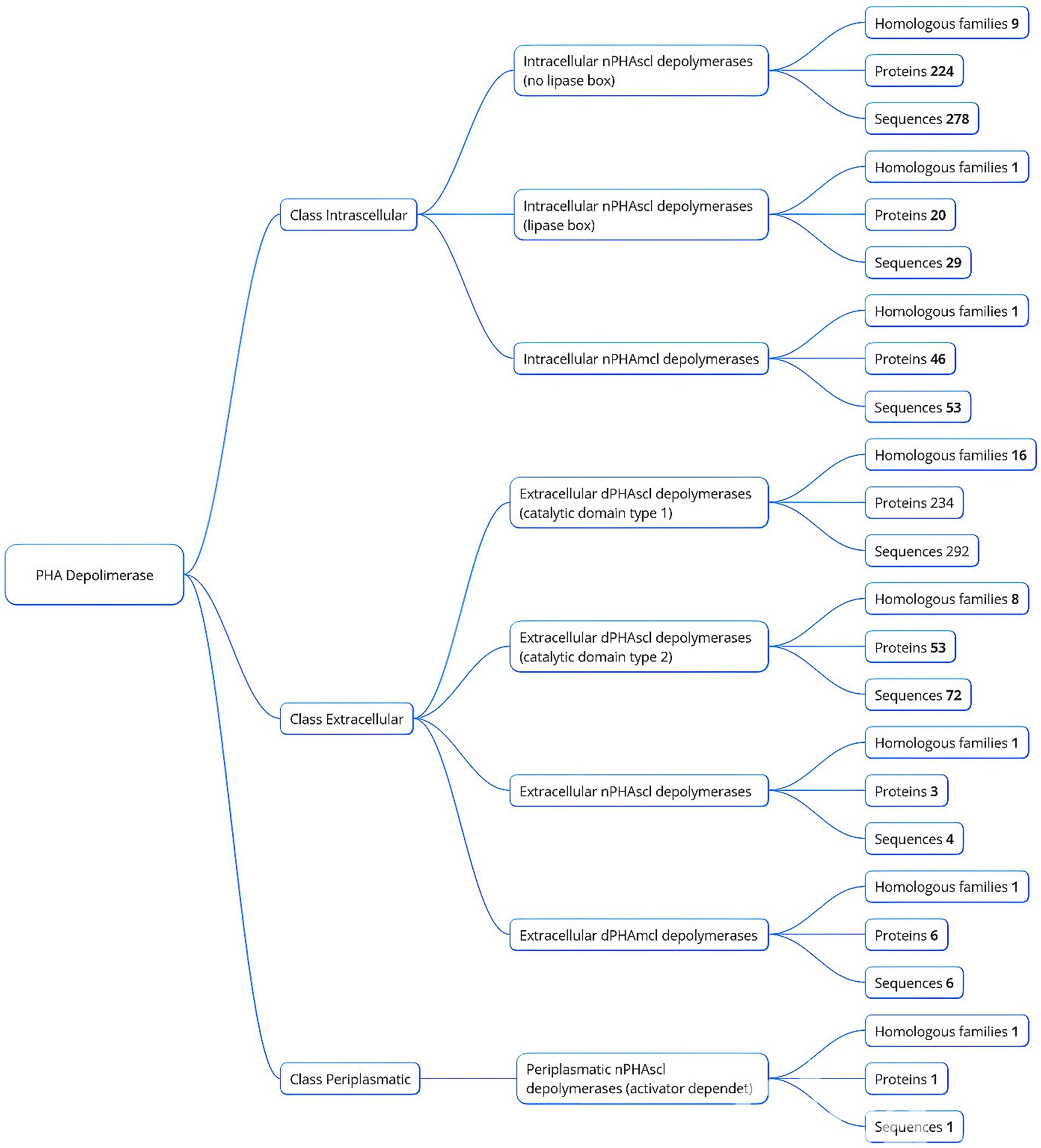
Figure 1. Classification of PHA depolymerases based on localization and homologous families.
While sequence based identification of PHA depolymerases is widespread, only a fraction of these enzymes have been purified and functionally validated. Compared to extracellular PHB (e-PHB) depolymerases, the biochemistry of intracellular PHB (i-PHB) depolymerases remains largely unexplored. Further research is needed to elucidate the structural determinants of substrate specificity and the catalytic mechanisms of these enzymes.
Efforts to optimize the use of PHA depolymerases for applications such as bioplastic recycling, waste management, and the production of PHA monomers are still in the early stages. From a biotechnological standpoint, extracellular PHA depolymerases hold significant promise due to their ability to hydrolyze paracrystalline PHA granules into monomers and oligomers. These degradation products can be metabolized by other microorganisms, making e-PHA depolymerases especially valuable for:
• Bioplastic recycling: enzymatic breakdown of PHA-based polymers for reuse.
• Waste management: accelerating the degradation of biodegradable plastics.
• PHA monomer recovery: facilitating sustainable bioplastic production.
The potential of e-PHA depolymerases to advance the circular bioeconomy is substantial. Future research should focus on comparative studies of e-PHASCL and e-PHAMCL depolymerases to optimize their industrial applications and improve our understanding of their ecological roles.
To date, various e-PHA depolymerases exhibiting activity toward SCL-PHA have been isolated and characterized from diverse bacterial origins. These include enzymes from Acidovorax (Vigneswari et al., 2015; Kobayashi et al., 1999; Wang et al., 2012), Aeromonas (Amir et al., 2024), Agrobacterium (Guo et al., 2016), Alcaligenes (Nojiri and Saito, 1997; Saito et al., 1989; Kita et al., 1997), Anoxybacillus (Colak et al., 2005), Arthrobacter (Asano and Watanabe, 2001), Aureobacterium (Sadocco et al., 1997), Bacillus (Ma et al., 2011; Takaku et al., 2006), Burkholderia (Azura Azami et al., 2019), Caldimonas (Lee et al., 2018), Comamonas (Kasuya et al., 1997; Shinomiya et al., 2006; Jendrossek et al., 1995), Leptothrix (Takeda et al., 2000; Takeda et al., 1998), Lihuaxuella (Thomas et al., 2022), Marinobacter (Kasuya et al., 2003), Microbacterium (Sadocco et al., 1997; Sayyed et al., 2019), Paucimonas (Braaz et al., 2003; Jendrossek et al., 1993; Briese et al., 1994; Jendrossek et al., 1995; Schöber et al., 2000), Pseudomonas (Di et al., 2019; Li et al., 2016; Ohura et al., 1999; Uefuji et al., 1997; Mao et al., 2013), Schlegelella (Elbanna et al., 2004), Streptomyces (Klingbeil et al., 1996; García-Hidalgo et al., 2012; Calabia and Tokiwa, 2006; Allen et al., 2011; Shah et al., 2007; Blevins et al., 2018), Thermus (Papaneophytou et al., 2009) (Table 2).
Characterized e-PHASCL depolymerases exhibit molecular weights ranging from 35 to 63 kDa (Amir et al., 2024; Ma et al., 2011), and display optimal activity across broad pH (Knoll et al., 2009; Bhatt et al., 2011; Lalonde et al., 2025; Zandieh et al., 2024; Kunioka and Doi, 1990; Hablot et al., 2008; Lin et al., 2020; Amir et al., 2024; Allen et al., 2011) and temperature ranges (30–80°C) (Sayyed et al., 2019; Elbanna et al., 2004; Allen et al., 2011). Among these, the most thermostable enzyme is derived from Schlegelella thermodepolymerans, exhibiting activity up to 90°C, with an optimal range of 75–90°C and stability after 24 h at 70°C (Elbanna et al., 2004). Another notable example is the enzyme from Streptomyces sp. IN1, which remains stable at 80°C for over 15 min and maintains activity at pH 12.0, highlighting its dual extremophilicity and industrial relevance (Allen et al., 2011). While certain enzymes, such as those from Schlegelella thermodepolymerans and Streptomyces sp. IN1, demonstrate exceptional stability, the structural basis for these properties remains unclear. Understanding these features could enhance the engineering of robust enzymes for industrial applications.
e-PHASCL depolymerases are commonly inhibited by reducing agents like dithiothreitol (DTT) and β-mercaptoethanol (ME), the importance of disulfide bonds for enzymatic activity. They are also sensitive to serine hydrolase inhibitors (e.g., PMSF, DFP) and chelating agents (e.g., EDTA). Most mature enzymes are inhibited by non-ionic (Tween 20, Tween 80, Triton X-100) and anionic (SDS) detergents. Metal ions also modulate activity; inhibitory effects are observed with Mn2+, Fe2+, and Ni2+, while activating effects are seen with Ca2+, Mg2+, Na+, and K+, likely due to structural stabilization or cofactor roles (Table 3).
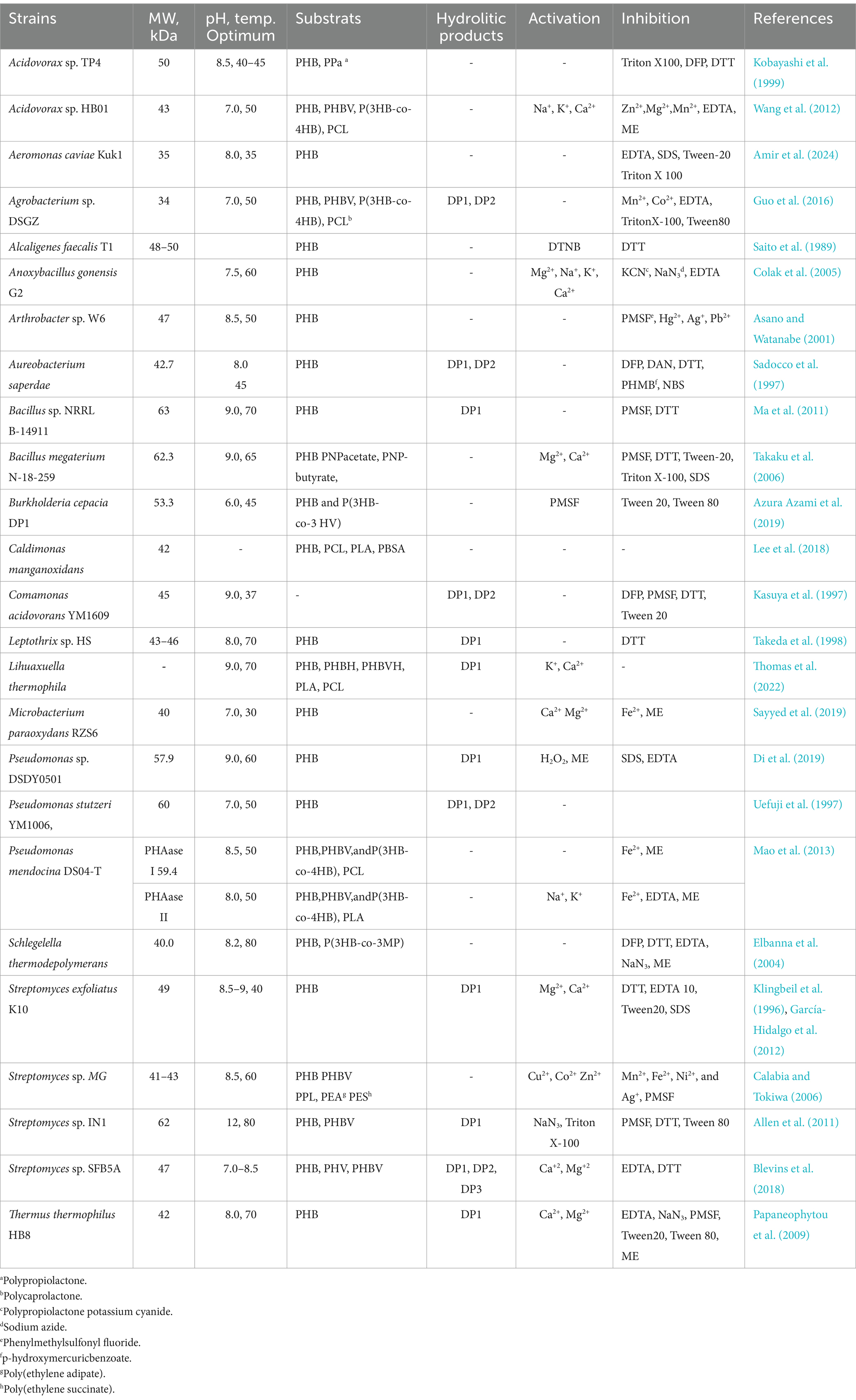
Table 3. Characteristics of e-PHASCL depolymerases from various microorganisms.
In the Bacillus genus, e-PHASCL depolymerases have been comprehensively studied only in Bacillus sp. strain B-14911 and Bacillus megaterium N-18-25-9 (Ma et al., 2011; Takaku et al., 2006). Our studies identified most strains as Priestia megaterium or Priestia aryabhattai (formerly B. megaterium). Whole-genome sequencing revealed that e-PHASCL depolymerase genes from P. megaterium N7 and P. aryabhattai L7 share 96% identity with B. megaterium N-18-25-9, with 22 amino acid differences suggesting variations in biochemical properties (Figure 2).
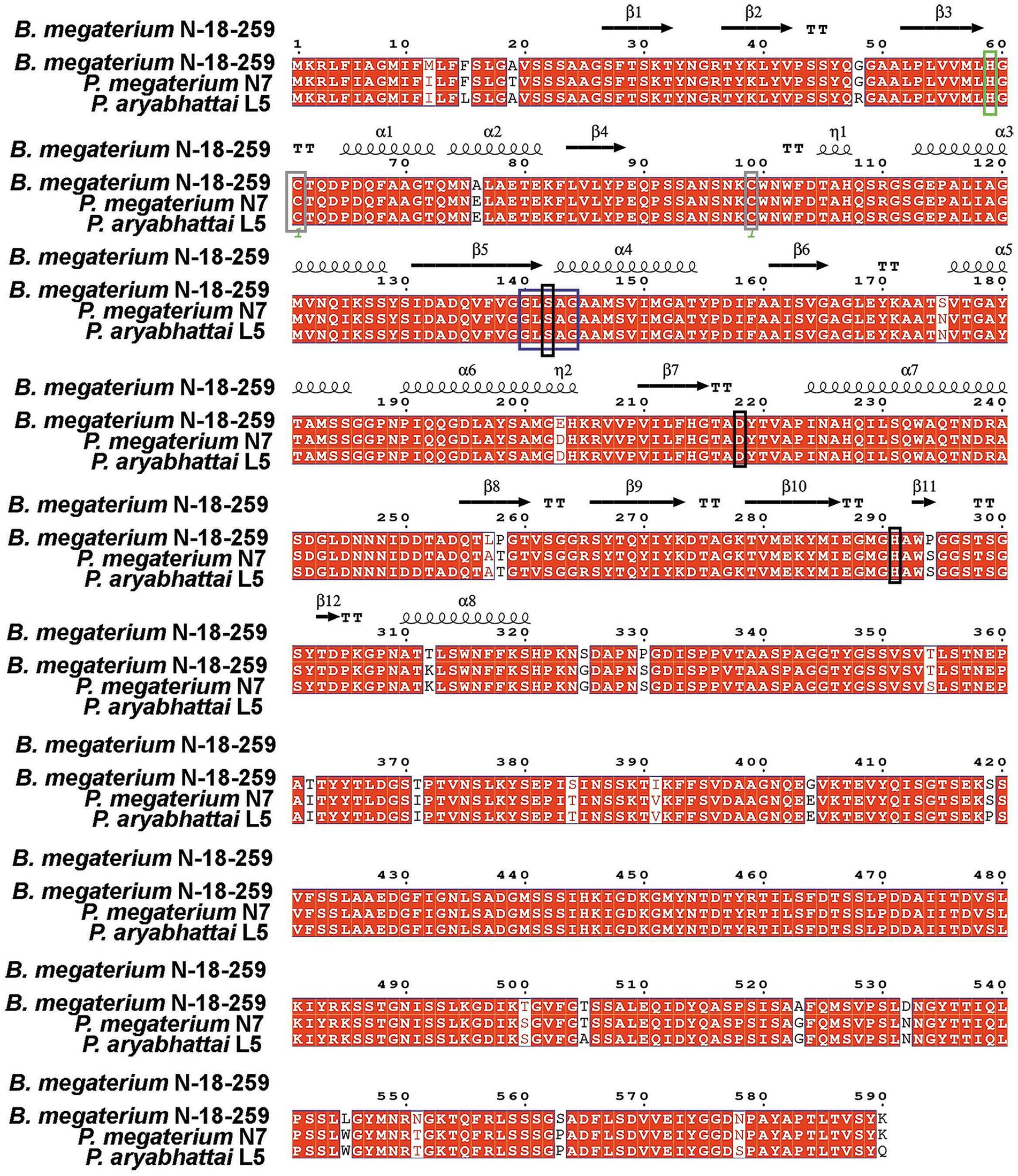
Figure 2. Comparative alignment of e-PHASCL depolymerase genes from B. megaterium N-18-25-9, P. megaterium N7, and P. aryabhattai L5, highlighting conserved catalytic triad, oxyanion hole, and structural motifs. The catalytic triad Ser146, Asp222, and His295 are marked in black, while the putative oxyanion hole residue His63 is highlighted in green. Two conserved cysteine residues are shown in brown, and the lipase box is marked in blue.
Our findings show that these enzymes degrade not only PHB but also copolymers like PHBV and PHBH. Interestingly, other strains exhibit different substrate specificities, underscoring the biochemical diversity of e-PHASCL depolymerases in Bacillus.
PHA depolymerization occurs in two steps: (Covello et al., 2024) adsorption of the binding domain onto the PHA surface, and (Yadav and Nikalje, 2024) hydrolysis of polyester chains by the catalytic domain (Numata et al., 2009). Enzyme-substrate interaction does not necessarily guarantee hydrolysis. For instance, PHA depolymerase from Alcaligenes faecalis adsorbs to five different substrates—PHB, PHP, P(4HB), P(2HP), and P(6Hx), but hydrolysis occurs only with PHB, PHP, and P(4HB). This suggests that the substrate-binding domain functions independently of the catalytic domain and exhibits broader interactions (Kasuya et al., 1996).
Polymer chain scission begins with endo-scission (random cleavage along the chain) followed by exo-scission (cleavage from the chain ends).
While all characterized e-PHASCL depolymerases degrade PHB some also degrade PHV and other copolymers, though they generally prefer PHB. Notably, PhaZ6 e-PHASCL depolymerase from Pseudomonas lemoignei displays higher activity toward PHV than PHB (Schöber et al., 2000). Understanding such substrate preferences could inform enzyme engineering for expanded specificity. Most of the characterized enzymes hydrolyze SCL-PHA down into monomers (Thomas et al., 2022; Di et al., 2019; Ma et al., 2011; Takeda et al., 1998; Klingbeil et al., 1996; García-Hidalgo et al., 2012; Allen et al., 2011), while others generate monomers, dimers and trimers (Guo et al., 2016; Sadocco et al., 1997; Kasuya et al., 1997; Uefuji et al., 1997; Blevins et al., 2018).
e-PHASCL depolymerases belong to the serine hydrolase family, characterized by the lipase-box motif Gly-X1-Ser-X2-Gly, where X1 is leucine/isoleucine and X2 is alanine/serine (Lee et al., 2018). The catalytic triad (Ser, Asp., His) and oxyanion hole (often cysteine-based) are critical for activity. Variations in these motifs suggest potential functional diversity. Many e-PHASCL depolymerases share similarities with lipases and esterases. However, Thomas et al. recently reported an e-PHASCL depolymerase from Lihuaxuella thermophila that resembles proteases and esterases but not lipases. This enzyme’s catalytic triad consists of Ser121, His270, and Asp197, while its oxyanion hole is formed by Cys40 (Thomas et al., 2022). The functional significance of motif variations remains poorly understood.
Most e-PHASCL depolymerases have four domains: signal peptide, catalytic domain, linker domain, and C-terminal substrate-binding domain (Ma et al., 2011; Takaku et al., 2006). The domain architecture of the e-PHASCL depolymerase from Bacillus megaterium N-18-25-9, identified through SMART protein analysis (Letunic et al., 2009) and modeled by AlphaFold3 (Abramson et al., 2024), is presented in Figure 3. Interestingly, the SMART database did not predict a distinct linking domain, a long linking region is observed, suggesting linker function.
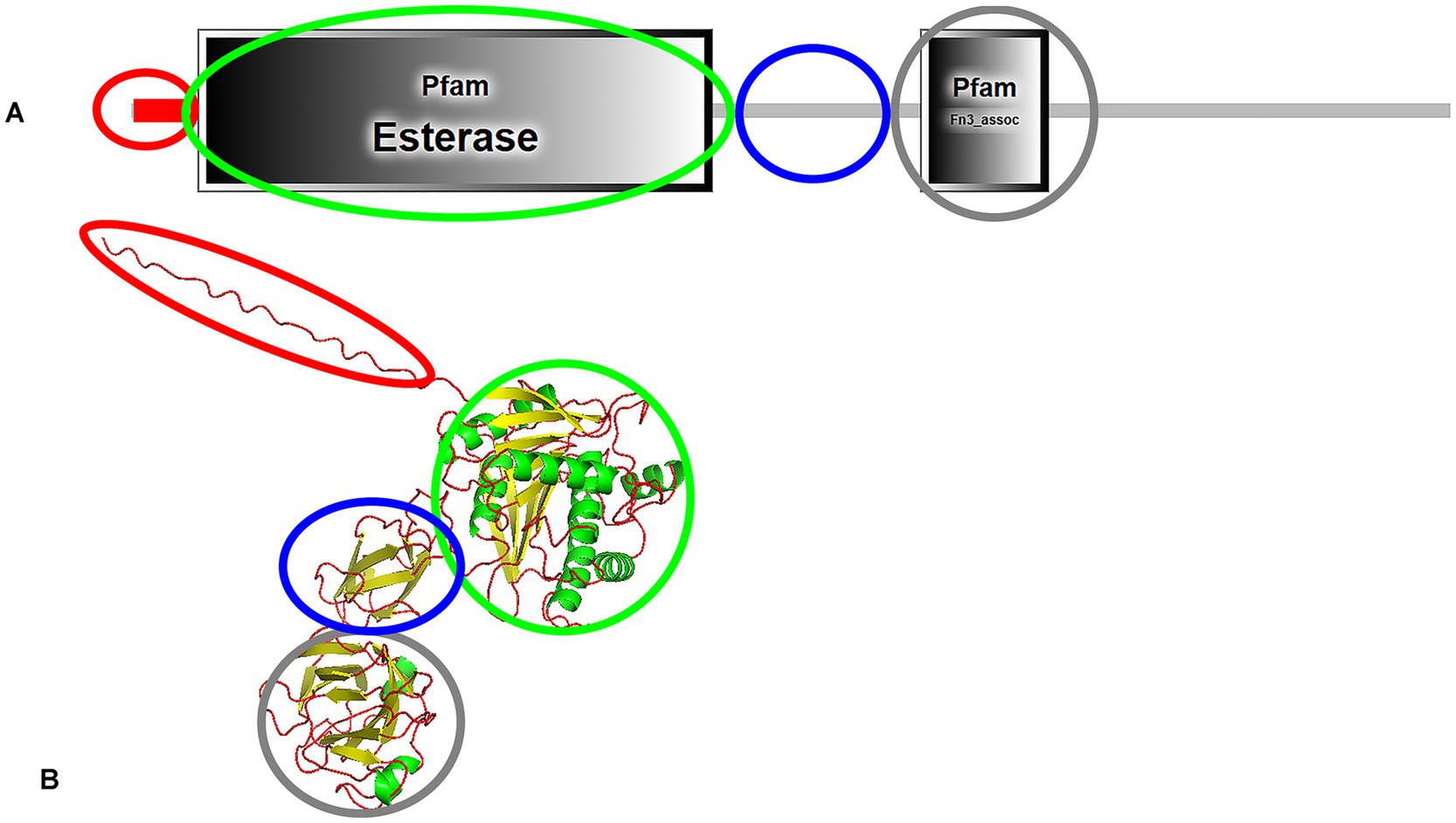
Figure 3. Domain architecture (A) and predicted structure (B) of e-PHASCL depolymerase from Bacillus megaterium N-18-25-9, highlighting functional domains. The signal peptide is marked in red, the catalytic domain in green, the linking domain in blue, and the C-terminal substrate-binding domain in grey. The structure was analyzed using SMART protein analysis and predicted by AlphaFold3.
Two types of catalytic domains (CD) exist: Type I, where the lipase-box is located in the middle of the sequence, and Type II, where it is near the N-terminus. Three types of linker domains—Fibronectin Type III (Fn3), Threonine-Rich (Thr-rich), and Cadherin-Like (Cad)—and two types of substrate-binding domains (SBD1 and SBD2) have been characterized (Jendrossek and Handrick, 2002). Bacillus derived e-PHASCL depolymerases possess a Type I catalytic domain (Ma et al., 2011; Takaku et al., 2006), while Comamonas acidovorans YM1609 and Leptothrix sp. strain HS feature Type II domains (Kasuya et al., 1997; Takeda et al., 1998). Despite structural variations in the catalytic domain—specifically, the position of the oxyanion hole—no significant differences in substrate specificity have been observed. This statement highlights an intriguing aspect of PHA depolymerase functionality. Regarding linker domains: Streptomyces sp. SFB5A, Comamonas acidovorans YM1609, and Leptothrix sp. strain HS contain an Fn3-type linking domain (Kasuya et al., 1997; Takeda et al., 1998; Blevins et al., 2018), Marinobacter sp. NK-1 has a Cad-type linking domain (Kasuya et al., 2003) and P. lemoignei features a Thr-rich linking domain (Briese et al., 1994). Notably, in Pseudomonas stutzeri, the linking domain has not been identified (Ohura et al., 1999). While different types of linking (e.g., Fn3, Cad, and Thr-rich) and substrate-binding domains (SBD1 and SBD2) have been characterized, their roles in substrate recognition, binding efficiency, and catalytic enhancement remain unclear. In some e-PHASCL depolymerases, such as in Pseudomonas stutzeri, the linking domain is absent, raising questions about alterantive structural mechanisms compensating for its absence. Limited availability of three-dimensional structures in the Protein Data Bank constrains our understanding of these enzymes.
The 3D structure of the PHB depolymerase PhaZ7 from Paucimonas lemoignei, which is active against amorphous PHB, has been extensively analyzed. This study revealed significant differences in substrate-binding strategies, active site accessibility, and conformational flexibility (Papageorgiou et al., 2008). Critical amino acid residues essential for substrate binding and enzymatic activity were identified, emphasizing a unique substrate-binding domain distinct from the catalytic core. Through mutagenesis, binding assays, and high-resolution structural analysis, researchers demonstrated how specific mutations influence PHB degradation (Jendrossek et al., 2013). More recently, the structure of a PHB depolymerase from the thermophilic Lihuaxuella thermophila provided further insights into broad substrate specificity, including activity against PLA and PCL. This enzyme’s shallow active site cleftfacilitates diverse substrates access, contrasting with P. lemoignei PhaZ7, where a mobile loop occludes the active site, reducing versatility (Thomas et al., 2022).
Microorganisms capable to degrade MCL-PHAs are less commonly found in the environment, and only a few e-PHAMCL depolymerases have been characterized to date. Most of the known e-PHAMCL depolymerases have been identified in species of Pseudomonas (Schirmer et al., 1993; Elbanna et al., 2004; Schirmer and Jendrossek, 1994; Schirmer et al., 1995; Kim et al., 2002) and Streptomyces (Martínez et al., 2015; Kim, 2003; Gangoiti et al., 2012; Santos et al., 2013). However, e-PHAMCL depolymerases have also been characterized in other genera, such as Bdellovibrio (Martínez et al., 2012), Thermus (Papaneophytou et al., 2011), Xanthomonas (Kim et al., 2000). The first e-PHAMCL depolymerase studied in detail at the molecular level was from Pseudomonas fluorescens GK13 (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995). Detailed results of charachterised e-PHAMCL are presented in Table 4.
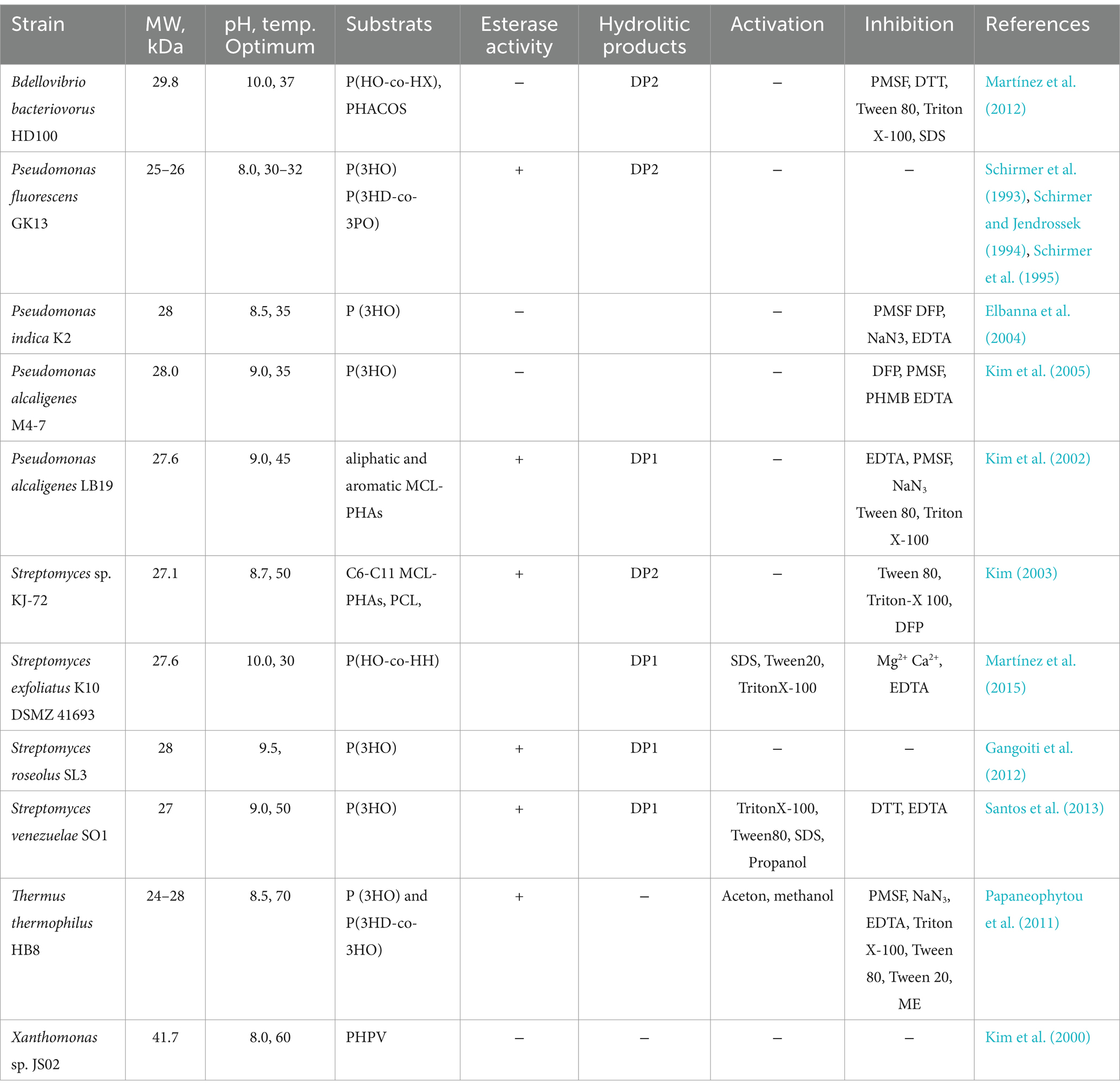
Table 4. Characteristics of e-PHAMCL depolymerases from various microorganisms.
The molecular weight of e-PHAMCL depolymerases generally ranges between 25 and 30 kDa (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995; Gangoiti et al., 2012; Martínez et al., 2012). However, to the best of our knowledge, the largest reported e-PHAMCL depolymerase derived from Xanthomonas sp. JS02 has a molecular weight of 41.7 kDa (Kim et al., 2000).
e-PHAMCL depolymerases exhibit pH optima between 8.0 and 10.0 and temperature optima ranging from 30°C to 70°C. The most thermostable e-PHAMCL depolymerase identified to date is from Thermus thermophilus HB8, with an optimum temperature of 70°C. In contrast, the e-PHAMCL depolymerases of P. fluorescens GK13 (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995) and Pseudomonas indica K2 (Elbanna et al., 2004) are most active at 30–35°C. Interestingly, dithiothreitol (DTT), which reduces disulfide bonds, does not affect the activity of e-PHAMCL depolymerases derived from Pseudomonas (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995; Kim et al., 2002). However, the e-PHAMCL depolymerases of Streptomyces venezuelae SO1 (Santos et al., 2013) and Bdellovibrio bacteriovorus HD100 (Martínez et al., 2012) are inhibited by DTT. This suggests that essential disulfide bonds are required for the enzymatic activity of these depolymerases, unlike those from Pseudomonas, which do not rely on such bonds for functionality. In some cases, EDTA exhibits an inactivating effect (Martínez et al., 2015), while in other cases, it has no impact (Kim, 2003). This suggests that the e-PHAMCL depolymerases are either not metalloenzymes or that the metal ions essential for activity are enclosed in a manner that prevents EDTA from chelating them. It is well established that e-PHAMCL depolymerases are serine hydrolases, which in most cases are inactivated by PMSF. Interestingly, e-PHAMCL depolymerases Streptomyces genera show minimal or no inhibition by PMSF compared with Pseudomonas (Martínez et al., 2015; Elbanna et al., 2004; Santos et al., 2013; Papaneophytou et al., 2011). This suggests that the structural architecture of these Streptomyces enzymes may differ significantly from that of Pseudomonas derived e-PHAMCL depolymerases, explaining their resistance to this inhibitor. Typically, e-PHAMCL depolymerases are inhibited by detergents. However, the activity of the enzymes from Streptomyces exfoliatus K10 and S. venezuelae SO1 demonstrated a notable enhancement in the presence of low concentrations of nonionic and anionic detergents, particularly with 0.01% SDS (Martínez et al., 2015; Santos et al., 2013). Most studied e-PHAMCL depolymerases are not activated by metals such as Ca2+ and Mg2+, suggesting that these enzymes do not require metal ions as cofactors for their activity (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995; Kim et al., 2002; Martínez et al., 2012). Furthermore, the depolymerase activity of S. exfoliatus was notably inhibited by CaCl₂ and MgCl₂ (Martínez et al., 2015).
Regarding the substrate polymers, most of the characterized enzymes exhibit activity toward MCL-PHA, degrading them into monomers (DP1) (Martínez et al., 2015; Kim et al., 2002; Gangoiti et al., 2012; Santos et al., 2013), However, enzymes from P. fluorescens GK13 (Schirmer et al., 1993; Schirmer and Jendrossek, 1994; Schirmer et al., 1995), Streptomyces sp. KJ-72 (Kim, 2003) and Bdellovibrio bacteriovorus HD100 (Martínez et al., 2012) primarily hydrolyze P(3HO) into dimeric forms (DP2). The enzyme from S. exfoliatus K10 DSMZ 41693 has been reported to function as an endo-exo-hydrolase, capable of cleaving both large and small PHA molecules. Remarkably, it requires only 30 min to produce monomers, whereas other enzymes demand higher enzyme loading and longer reaction times to achieve similar results (Gangoiti et al., 2012; Santos et al., 2013).
Some e-PHAMCL depolymerases also exhibit versatile, with applications extending to the hydrolysis of copolymers. For instance, the recombinant e-PHAMCL depolymerase from P. fluorescens GK13 can successfully degrade copolymer consisting of PHB and polyhydroxyoctanoate (PHO) (Millan and Hanik, 2023). Additionally, the e-PHAMCL depolymerase from S. exfoliatus K10 has been applied to hydrolyze functionalized polymers as PHACOS, yielding functional thioester-based monomers (Martínez et al., 2015).
The amino acid sequence of characterized e-PHAMCL depolymerases varies between 277 and 282 residues. These enzymes contain high proportion of amino acids with aromatic and uncharged aliphatic side chains, contributing to their hydrophobic nature. The amino acid sequence similarity among characterized e-PHAMCL depolymerases ranges from 69 to 98% (Kim et al., 2005). Sequences analysis revealed that they generally consist of a signal peptide, an N-terminal substrate-binding domain, and a C-terminal catalytic domain (Kim et al., 2002; Gangoiti et al., 2012; Papaneophytou et al., 2011). However, in the case of P. fluorescens GK13, only the signal peptide was identified, with substrate-binding domain likely located in N-terminal region (Schirmer and Jendrossek, 1994; Jendrossek and Handrick, 2002; Kim et al., 2005). Unlike e-PHASCL depolymerases, no linking domain has been identified between the substrate-binding region and the catalytic domain in e-PHAMCL depolymerases. e-PHAMCL depolymerases contain a conserved catalytic triad (Ser, Asp., His) in the active center (Martínez et al., 2015; Gangoiti et al., 2012). The catalytic domain of all e-PHAMCL depolymerases contains a lipase box (Gly-X1-Ser-X2-Gly), where X1 is an Ile and X2 is a Ser (Martínez et al., 2012). However, variations have been observed: inT. thermophilus, X1 is Gly and X2 is Tyr (Papaneophytou et al., 2011), in S. exfoliatus and S. roseolus, X1 is His and X2 is Gln (Martínez et al., 2015). Additionally, all PHA depolymerases contain an oxyanion hole residue essential for stabilizing the transition state during hydrolysis. This residue varies across species, being His in P. fluorescens GK13 (Schirmer and Jendrossek, 1994), Ser in Pseudomonas alcaligenes M4-7 (Kim et al., 2005), Asn111 in P. alcaligenes LB19 (Kim et al., 2005), Asn10 in T. thermophilus (Papaneophytou et al., 2011), and Gln147 in Streptomyces roseolus SL3 (Gangoiti et al., 2012).
Further ecological studies on MCL-PHA degraders, along with advancements in the biochemistry and molecular biology of MCL-PHA depolymerases, are essential to expanding our understanding of MCL-PHA degradation and its potential applications.
The vast majority of PHA-degrading microorganisms are known to produce only one type of PHA depolymerase acting upon either SCL-PHAs or MCL-PHAs. Very few bacterial strains have been identified with both e-PHASCL depolymerase and e-PHAMCL depolymerase activities. One such strain is Pseudomonas indica K2. Researchers demonstrated that this strain produces two distinct depolymerases: one specific for SCL-PHA and another for MCL-PHA. Additionally, they successfully purified and characterized the extracellular poly (3HO) depolymerase, which exhibited activity for MCL-PHA substrates but not for SCL-PHB, further confirming its role as an e-PHAMCL depolymerase (Elbanna et al., 2004). Similar results have been found for Xanthomonas sp. JS02 (Kim et al., 2000), S. exfoliatus K10 (Klingbeil et al., 1996; García-Hidalgo et al., 2012). Another example is Streptomyces sp. KJ-72 strain which also produces e-PHAMCL depolymerase. Interestingly it is expressed only if PHO was used as a carbon source in the cultivation media. It has also been reported that if PHB is used as a source of carbon PHB depolymerase is being produced (Kim, 2003). Additionally T. thermophilus produces an extra cellular e-PHAMCL depolymerase. In addition, it has been previously demonstrated that T. thermophilus is able to produce e-PHASCL depolymerase suggesting that this microorganism secretes different PHA depolymerases depending on the growth conditions. The extracellular e-PHASCL depolymerase of T. thermophilus was found to be secreted constitutively in the presence of MCL-alkanoates but not in the presence of glucose or SCL-alkanoates, similarly to the extracellular PHO depolymerase of P. fluorescens GK13 (Schirmer et al., 1993). According to Schirmer et al. high expression of the e-PHAMCL depolymerase depends on (i) the presence of 3HAs or similar compounds (as inducers) or (ii) carbon starvation. Thus, satisfactory amounts of PHA depolymerase can be produced using sodium octanoate as sole carbon source instead of PHO (Schirmer et al., 1993).
When PHA is disposed in the environment, it comes into contact with natural microbial communities. Specific microorganisms secrete PHA depolymerases, which catalyze the hydrolysis of ester bonds in the polymer, breaking it down into smaller units, primarily D-3-hydroxybutyrate and acetoacetate. These monomers or oligomers are taken up by the microorganisms and enter the microbial metabolic pathways, such as the β-oxidation pathway in anaerobic bacteria, or tricarboxylic acid cycle (TCA) cycle in aerobic bacteria. As a result, the monomers are further broken down into fundamental metabolites like acetyl-CoA. The final metabolic products include carbon dioxide (CO2), water (H2O), and microbial biomass under aerobic conditions. In anaerobic environments methane (CH4) may also be produced alongside CO2 (Figure 4) (Numata et al., 2009; Zhou et al., 2023).
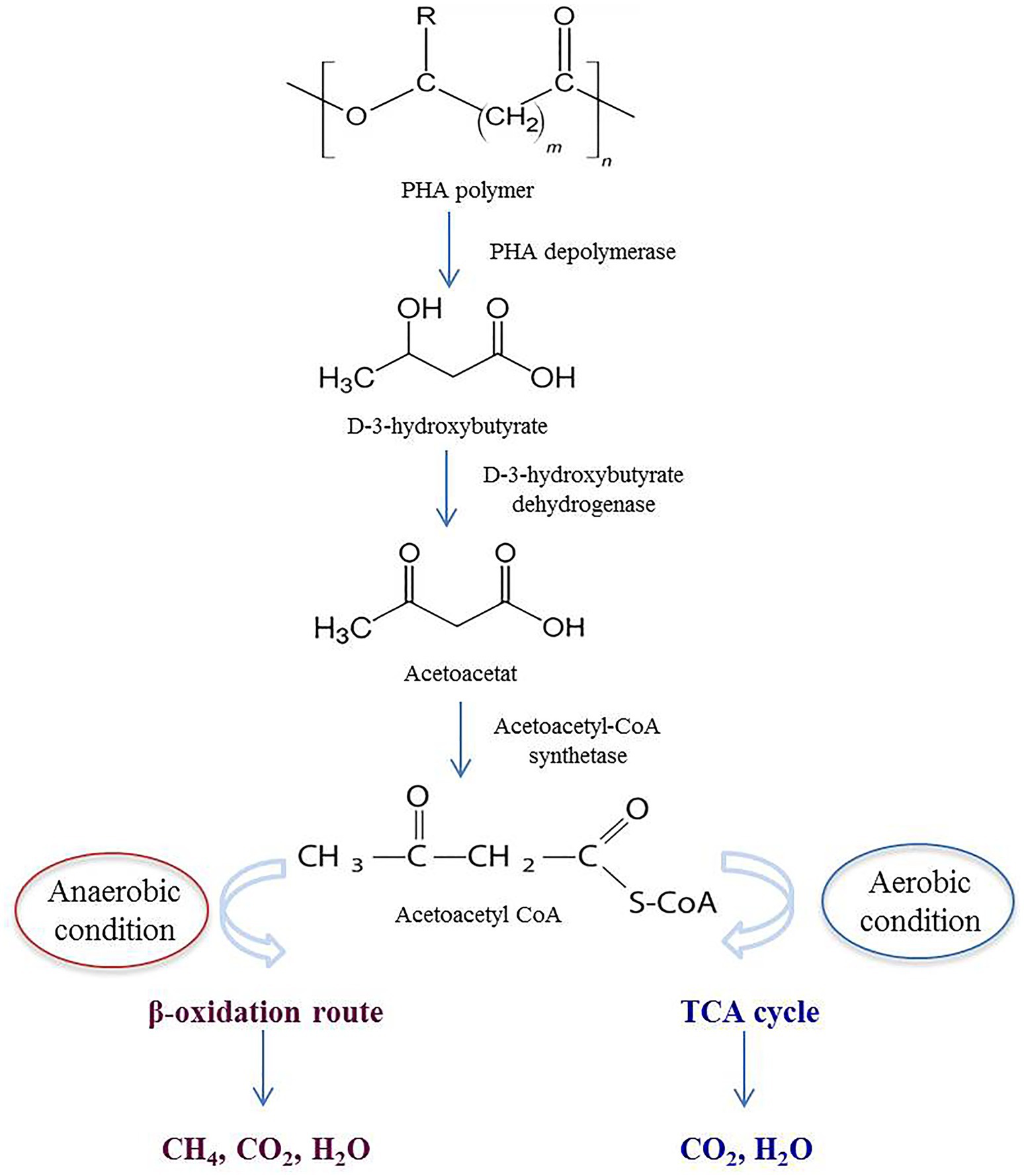
Figure 4. Schematic depiction of extracellular PHA degradation in both anaerobic and aerobic environments.
Unlike conventional plastics, PHA degrades into environmentally benign by-products without leaving toxic residues. The by-products contribute to the natural carbon cycle, potentially enhancing soil fertility or water quality (Shah et al., 2008). While the effectiveness of depolymerases has been well demonstrated in controlled laboratory conditions, their performance in real-world environments, such as landfills and soil, is significantly influenced by various environmental factors often not considered in laboratory studies, though degradation rates are generally lower compared to controlled settings. Studies have shown that microbial activity is the primery driver of PHA degradation with specific bacteria such as Pseudomonas and Acidovorax playing key roles. Additionally, degradation rates vary depending on the monomer composition of the PHA. For instance, P(3HB-co-4HB) degrades more rapidly than P(3HB) and P(3HB-co-3 HV). It has been reported that certain copolymers exhibit degradation rates of 98.9 ± 1.8% in soil and 81.5 ± 2.9% in lake environments after five weeks (Sevakumaran et al., 2019).
Furthermore, PHBV has shown significant biodegradation under compost and soil conditions. In controlled compost, PHBV degraded by 90% within 200 days, whereas in soil, the degradation rate was only 32% over the same period (Muniyasamy et al., 2016). Additionally marine microorganisms have been reported to degrade PHAs by secreting depolymerases that break down the polymer (Hachisuka et al., 2023).
However, the lack of standardized testing methods for deep-sea conditions presents challenges in accurately assessing PHA biodegradability in such environments (Meereboer et al., 2020).
For a successful bioindustrial process, enzymes must exhibit fast kinetics, long-term stability, and high activity within the specific environmental context. Special attention should be given to enzymes that are stable at high temperatures and resistant to various salts and detergents, as these properties are crucial for their functionality under harsh production conditions. Extremophilic microorganisms are valuable resources for identifying and engineering robust PHA depolymerases for industrial and environmental applications. Despite notable advancements in the purification and biochemical characterization of PHA depolymerases, structural studies remain limited. The lack of detailed 3D structural information hinders our understanding of critical mechanisms, such as enzymes-substrate recognition, binding and degradation, as well as how they maintain activity under extreme environmental conditions. By further advancing our understanding of microbial and enzymatic degradation processes, we can pave the way for a circular plastic bioeconomy, establishing PHAs as a viable and eco-friendly alternative to petroleum-based plastics.
AP: Funding acquisition, Supervision, Visualization, Project administration, Writing – original draft. MT: Writing – original draft, Data curation. DG: Data curation, Writing – original draft. LK: Data curation, Writing – original draft. MK: Data curation, Writing – original draft. AM: Funding acquisition, Project administration, Supervision, Visualization, Writing – review & editing. GA: Funding acquisition, Supervision, Visualization, Writing – review & editing, Conceptualization. HP: Conceptualization, Funding acquisition, Supervision, Visualization, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The research was conducted within the Science Committee of Armenia (24FP-2J041), ADVANCE Research Grant provided by the Foundation for Armenian Science and Technology (FAST) and Yerevan State University and ANSEF NS-microbio 2390.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abramson, J., Adler, J., Dunger, J., Evans, R., Green, T., Pritzel, A., et al. (2024). Accurate structure prediction of biomolecular interactions with alpha fold 3. Nature 630, 493–500. doi: 10.1038/s41586-024-07487-w
Allen, A. D., Anderson, W. A., Ayorinde, F., and Eribo, B. E. (2011). Isolation and characterization of an extracellular thermoalkanophilic P(3HB-co-3HV) depolymerase from Streptomyces sp. IN1. Int. Biodeterior. Biodegrad. 65, 777–785. doi: 10.1016/j.ibiod.2011.02.010
Amir, M., Bano, N., Gupta, A., and Zaheer Mohd, R. (2024). Purification and characterization of extracellular PHB depolymerase enzyme from Aeromonas caviae Kuk 1-(34) and their biodegradation studies with polymer films. Biodegradation 35, 137–153. doi: 10.1007/s10532-023-10051-4
Arnosti, C. (2011). Microbial extracellular enzymes and the marine carbon cycle. Annu. Rev. Mar. Sci. 3, 401–425. doi: 10.1146/annurev-marine-120709-142731
Asano, Y., and Watanabe, S. (2001). Isolation of poly (3-Hydroxybutyrate) (PHB)-degrading microorganisms and characterization of PHB-depolymerase from Arthrobacter sp. strain W6. Biosci. Biotechnol. Biochem. 65, 1191–1194. doi: 10.1271/bbb.65.1191
Azura Azami, N., Ira Aryani, W., Aik-Hong, T., and Amirul, A. A. (2019). Purification and characterization of new bio-plastic degrading enzyme from Burkholderia cepacia DP1. Protein Expr. Purif. 155, 35–42. doi: 10.1016/j.pep.2018.10.008
Bhatt, R., Patel, K., and Trivedi, U. (2011). “Biodegradation of poly (3-hydroxyalkanoates)” in A handbook of applied biopolymer technology: Synthesis, degradation and applications [internet]. eds. S. K. Sharma and A. Mudhoo (Cambridge, United Kingdom: The Royal Society of Chemistry).
Blevins, H. M., MkD, B., Cobbs, B. D., Ricotilli, T. A., Kyler, S. L., Shuey, C. T., et al. (2018). Characterization of an extracellular Polyhydroxyalkanoate Depolymerase from Streptomyces sp. SFB5A. J. Bioremediat. Biodegrad. 9:452. doi: 10.4172/2155-6199.1000452
Bonartsev, A., Myshkina, V., Nikolaeva, D., Furina, E., Makhina, T., Iordanskii, A., et al. (2007). Biosynthesis, biodegradation, and application of poly (3-hydroxybutyrate) and its copolymers - natural polyesters produced by diazotrophic bacteria. Commun Curr res Educ top trends. Appl. Microbiol. 1, 295–307.
Bonartseva, G. A., Myshkina, V. L., Nikolaeva, D. A., Rebrov, A. V., Gerasin, V. A., and Makhina, T. K. (2002). The biodegradation of poly-b-Hydroxybutyrate (PHB) by a model soil community: The effect of cultivation conditions on the degradation rate and the physicochemical characteristics of PHB. Cham: Springer.
Boyandin, A. N., Prudnikova, S. V., Karpov, V. A., Ivonin, V. N., Đỗ, N. L., Nguyễn, T. H., et al. (2013). Microbial degradation of polyhydroxyalkanoates in tropical soils. Int. Biodeterior. Biodegrad. 83, 77–84. doi: 10.1016/j.ibiod.2013.04.014
Boyandin, A. N., Rudnev, V. P., Ivonin, V. N., Prudnikova, S. V., Korobikhina, K. I., Filipenko, M. L., et al. (2012). Biodegradation of Polyhydroxyalkanoate films in natural environments. Macromol. Symp. 320, 38–42. doi: 10.1002/masy.201251004
Braaz, R., Handrick, R., and Jendrossek, D. (2003). Identification and characterisation of the catalytic triad of the alkaliphilic thermotolerant PHA depolymerase PHA Z7 of Paucimonas lemoignei. FEMS Microbiol. Lett. 224, 107–112. doi: 10.1016/S0378-1097(03)00425-7
Briese, B. H., Schmidt, B., and Jendrossek, D. (1994). Pseudomonas lemoignei has five poly (hydroxyalkanoic acid) (PHA) depolymerase genes: a comparative study of bacterial and eukaryotic PHA depolymerases. J. Environ. Polym. Degrad. 2, 75–87. doi: 10.1007/BF02074776
Calabia, B. P., and Tokiwa, Y. (2006). A novel PHB Depolymerase from a thermophilic Streptomyces sp. Biotechnol. Lett. 28, 383–388. doi: 10.1007/s10529-005-6063-5
Cho, J. Y., Kim, S. H., Jung, H. J., Cho, D. H., Kim, B. C., Bhatia, S. K., et al. (2022). Finding a benign plasticizer to enhance the microbial degradation of Polyhydroxybutyrate (PHB) evaluated by PHB degrader Microbulbifer sp. SOL66. Polymers 14:3625. doi: 10.3390/polym14173625
Colak, A., Sisik, D., Saglam, N., Guner, S., Canakci, S., and Belduz, A. (2005). Characterization of a thermoalkalophilic esterase from a novel thermophilic bacterium, G2. Bioresour. Technol. 96, 625–631. doi: 10.1016/j.biortech.2004.06.003
Covello, C., Di Vincenzo, F., Cammarota, G., and Pizzoferrato, M. (2024). Micro (nano) plastics and their potential impact on human gut health: a narrative review. Curr. Issues Mol. Biol. 46, 2658–2677. doi: 10.3390/cimb46030168
Di, Y., Xia, H., Jiao, Y., Zhang, X., Fang, Q., Li, F., et al. (2019). Biodegradation of polyhydroxybutyrate by Pseudomonas sp. DSDY0501 and purification and characterization of polyhydroxybutyrate depolymerase. 3 Biotech 9:359. doi: 10.1007/s13205-019-1871-9
Elbanna, K., Lütke-Eversloh, T., Jendrossek, D., Luftmann, H., and Steinbüchel, A. (2004). Studies on the biodegradability of polythioester copolymers and homopolymers by polyhydroxyalkanoate (PHA)-degrading bacteria and PHA depolymerases. Arch Microbiol [Internet] 182, 212–225. doi: 10.1007/s00203-004-0715-z
Gangoiti, J., Santos, M., Prieto, M. A., De La Mata, I., Serra, J. L., and Llama, M. J. (2012). Characterization of a novel subgroup of extracellular medium-chain-length Polyhydroxyalkanoate Depolymerases from Actinobacteria. Appl. Environ. Microbiol. 78, 7229–7237. doi: 10.1128/AEM.01707-12
García-Hidalgo, J., Hormigo, D., Prieto, M. A., Arroyo, M., and De La Mata, I. (2012). Extracellular production of Streptomyces exfoliatus poly (3-hydroxybutyrate) depolymerase in Rhodococcus sp. T104: determination of optimal biocatalyst conditions. Appl. Microbiol. Biotechnol. 93, 1975–1988. doi: 10.1007/s00253-011-3527-5
Getino, L., Martín, J. L., and Chamizo-Ampudia, A. (2024). A review of Polyhydroxyalkanoates: characterization, production, and application from waste. Microorganisms 12:2028. doi: 10.3390/microorganisms12102028
Ghanem, N. B., Mabrouk, M. E. S., Sabry, S. A., and El-Badan, D. E. S. (2005). Degradation of polyesters by a novel marine Nocardiopsis aegyptia sp. nov.: application of Plackett-Burman experimental design for the improvement of PHB depolymerase activity. J. Gen. Appl. Microbiol. 51, 151–158. doi: 10.2323/jgam.51.151
Guo, Z., Li, F., Liu, D., Xia, H., Yang, C., Chen, S., et al. (2016). Biodegradation of poly (3-hydroxybutyrate-co −4-hydroxybutyrate) by a novel P 3/4HB depolymerase purified from A grobacterium sp. J. Appl. Polym. Sci. 133:app. 42805. doi: 10.1002/app.42805
Hablot, E., Bordes, P., Pollet, E., and Avérous, L. (2008). Thermal and thermo-mechanical degradation of poly (3-hydroxybutyrate)-based multiphase systems. Polym. Degrad. Stab. 93, 413–421. doi: 10.1016/j.polymdegradstab.2007.11.018
Hachisuka, S. I., Sakurai, T., Mizuno, S., Kosuge, K., Endo, S., Ishii-Hyakutake, M., et al. (2023). Isolation and characterization of polyhydroxyalkanoate-degrading bacteria in seawater at two different depths from Suruga bay. Nikel PI, editor. Appl. Environ. Microbiol. 89, e01488–e01423. doi: 10.1128/aem.01488-23
Handrick, R., Reinhardt, S., Focarete, M. L., Scandola, M., Adamus, G., Kowalczuk, M., et al. (2001). A new type of Thermoalkalophilic hydrolase of Paucimonas lemoignei with high specificity for amorphous polyesters of short chain-length Hydroxyalkanoic acids. J. Biol. Chem. 276, 36215–36224. doi: 10.1074/jbc.M101106200
Handrick, R., Reinhardt, S., Kimmig, P., and Jendrossek, D. (2004). The “intracellular” poly(3-Hydroxybutyrate) (PHB) Depolymerase of Rhodospirillum rubrumis a periplasm-located protein with specificity for native PHB and with structural similarity to extracellular PHB Depolymerases. J. Bacteriol. 186, 7243–7253. doi: 10.1128/JB.186.21.7243-7253.2004
Hori, C., Sugiyama, T., Watanabe, K., Sun, J., Kamada, Y., Ooi, T., et al. (2020). Isolation of poly [d-lactate (LA)-co-3-hydroxybutyrate]-degrading bacteria from soil and characterization of d-LA homo-oligomer degradation by the isolated strains. Polym. Degrad. Stab. 179:109231. doi: 10.1016/j.polymdegradstab.2020.109231
Jaeger, K. E., and Rosenau, F. (2004). “Overexpression and secretion of Pseudomonas lipases” in Pseudomonas [internet]. ed. J. L. Ramos (Boston, MA: Springer US), 491–508.
Jendrossek, D., Backhaus, M., and Andermann, M. (1995). Characterization of the extracellular poly (3-hydroxybutyrate) depolymerase of Comamonas sp. and of its structural gene. Can. J. Microbiol. 41, 160–169. doi: 10.1139/m95-183
Jendrossek, D., Frisse, A., Behrends, A., Andermann, M., Kratzin, H. D., Stanislawski, T., et al. (1995). Biochemical and molecular characterization of the Pseudomonas lemoignei polyhydroxyalkanoate depolymerase system. J. Bacteriol. 177, 596–607. doi: 10.1128/jb.177.3.596-607.1995
Jendrossek, D., and Handrick, R. (2002). Microbial degradation of Polyhydroxyalkanoates. Ann. Rev. Microbiol. 56, 403–432. doi: 10.1146/annurev.micro.56.012302.160838
Jendrossek, D., Hermawan, S., Subedi, B., and Papageorgiou, A. C. (2013). Biochemical analysis and structure determination of Paucimonas lemoignei poly (3-hydroxybutyrate) (PHB) depolymerase PHA Z 7 muteins reveal the PHB binding site and details of substrate–enzyme interactions. Mol. Microbiol. 90, 649–664. doi: 10.1111/mmi.12391
Jendrossek, D., Müller, B., and Schlegel, H. G. (1993). Cloning and characterization of the poly (hydroxyalkanoic acid)-depolymerase gene locus, pha Z1, of Pseudomonas lemoignei and its gene product. Eur. J. Biochem. 218, 701–710. doi: 10.1111/j.1432-1033.1993.tb18424.x
Kasuya, K., Inoue, Y., and Doi, Y. (1996). Adsorption kinetics of bacterial PHB depolymerase on the surface of polyhydroxyalkanoate films. Int. J. Biol. Macromol. 19, 35–40. doi: 10.1016/0141-8130(96)01097-5
Kasuya, K., Inoue, Y., Tanaka, T., Akehata, T., Iwata, T., Fukui, T., et al. (1997). Biochemical and molecular characterization of the polyhydroxybutyrate depolymerase of Comamonas acidovorans YM 1609, isolated from freshwater. Appl. Environ. Microbiol. 63, 4844–4852. doi: 10.1128/aem.63.12.4844-4852.1997
Kasuya, K., Takano, T., Tezuka, Y., Hsieh, W. C., Mitomo, H., and Doi, Y. (2003). Cloning, expression and characterization of a poly (3-hydroxybutyrate) depolymerase from Marinobacter sp. NK-1. Int. J. Biol. Macromol. 33, 221–226. doi: 10.1016/j.ijbiomac.2003.08.006
Khoyetsyan, L., Karapetyan, M., Soghomonyan, T., Tadevosyan, M., Ghevondyan, D., Margaryan, A., et al. Isolation of bioplastic degrading bacterial strains and characterization of a corresponding enzyme. In: Proceedings of 14th international congress on extremophiles. (2024).
Kim, H. J. (2003). Characterization of an extracellular medium-chain-length poly(3-hydroxyalkanoate) depolymerase from Streptomyces sp. KJ-72. Antonie Van Leeuwenhoek 83, 183–189. doi: 10.1023/A:1023395527073
Kim, H., Ju, H. S., and Kim, J. (2000). Characterization of an extracellular poly(3-hydroxy-5-phenylvalerate) depolymerase from Xanthomonas sp. JS02. Appl. Microbiol. Biotechnol. 53, 323–327. doi: 10.1007/s002530050028
Kim, D. Y., Kim, H. C., Kim, S. Y., and Rhee, Y. H. (2005). Molecular characterization of extracellular medium-chain-length poly (3-hydroxyalkanoate) depolymerase genes from Pseudomonas alcaligenes strains. J. Microbiol. Seoul Korea 43, 285–294.
Kim, D. Y., Nam, J. S., and Rhee, Y. H. (2002). Characterization of an extracellular medium-chain-length poly (3-hydroxyalkanoate) Depolymerase from Pseudomonas alcaligenes LB19. Biomacromolecules 3, 291–296. doi: 10.1021/bm010113q
Kita, K., Mashiba, S., Nagita, M., Ishimaru, K., Okamoto, K., Yanase, H., et al. (1997). Cloning of poly (3-hydroxybutyrate) depolymerase from a marine bacterium, Alcaligenes faecalis AE122, and characterization of its gene product. Biochim. Biophys. Acta BBA Gene Struct. Expr. 1352, 113–122. doi: 10.1016/S0167-4781(97)00011-0
Klingbeil, B., Kroppenstedt, R. M., and Jendrossek, D. (1996). Taxonomic identification of Streptomyces exfoliatus K10 and characterization of its poly (3-hydroxybutyrate) depolymerase gene. FEMS Microbiol. Lett. 142, 215–221. doi: 10.1111/j.1574-6968.1996.tb08433.x
Knoll, M., Hamm, T. M., Wagner, F., Martinez, V., and Pleiss, J. (2009). The PHA Depolymerase engineering database: a systematic analysis tool for the diverse family of polyhydroxyalkanoate (PHA) depolymerases. BMC Bioinform. 10:89. doi: 10.1186/1471-2105-10-89
Kobayashi, T., Sugiyama, A., Kawase, Y., Saito, T., Mergaert, J., and Swings, J. (1999). No title found. J. Polym. Environ. 7, 9–18. doi: 10.1023/A:1021885901119
Kumaravel, S., Hema, R., and Lakshmi, R. (2010). Production of Polyhydroxybutyrate (bioplastic) and its biodegradation by Pseudomonas lemoignei and Aspergillus niger. J. Chem. 7:547. doi: 10.1155/2010/148547
Kunioka, M., and Doi, Y. (1990). Thermal degradation of microbial copolyesters: poly (3-hydroxybutyrate-co-3-hydroxyvalerate) and poly (3-hydroxybutyrate-co-4-hydroxybutyrate). Macromolecules 23, 1933–1936. doi: 10.1021/ma00209a009
Lalonde, J. N., Pilania, G., and Marrone, B. L. (2025). Materials designed to degrade: structure, properties, processing, and performance relationships in polyhydroxyalkanoate biopolymers. Polym. Chem. 16, 235–265. doi: 10.1039/D4PY00623B
Lee, K. M., Gimore, D. F., and Huss, M. J. (2005). Fungal degradation of the bioplastic PHB (Poly-3-hydroxy-butyric acid). J. Polym. Environ. 13, 213–219. doi: 10.1007/s10924-005-4756-4
Lee, M., Liu, E., Yang, C., Hsiao, L., Wu, T., and Li, S. (2018). Co-expression of ORF Cma with PHB Depolymerase (Pha Z Cma) in Escherichia coli induces efficient whole-cell biodegradation of polyesters. Biotechnol. J. 13:e1700560. doi: 10.1002/biot.201700560
Letunic, I., Doerks, T., and Bork, P. (2009). SMART 6: recent updates and new developments. Nucleic Acids Res. 37, D229–D232. doi: 10.1093/nar/gkn808
Li, W. C., Tse, H. F., and Fok, L. (2016). Plastic waste in the marine environment: a review of sources, occurrence and effects. Sci. Total Environ. 566-567, 333–349. doi: 10.1016/j.scitotenv.2016.05.084
Li, F., Zhang, C., Liu, Y., Liu, D., Xia, H., and Chen, S. (2016). Efficient production of (R)-3-hydroxybutyric acid by Pseudomonas sp. DS1001a and its extracellular poly (3-hydroxybutyrate) depolymerase. Process Biochem. 51, 369–373. doi: 10.1016/j.procbio.2015.12.016
Lin, Y., Kouznetsova, T. B., Chang, C. C., and Craig, S. L. (2020). Enhanced polymer mechanical degradation through mechanochemically unveiled lactonization. Nat. Commun. 11:4987. doi: 10.1038/s41467-020-18809-7
Liu, Y., De Schryver, P., Van Delsen, B., Maignien, L., Boon, N., Sorgeloos, P., et al. (2010). PHB-degrading bacteria isolated from the gastrointestinal tract of aquatic animals as protective actors against luminescent vibriosis: PHB-degrading isolates for Artemia protection. FEMS Microbiol. Ecol. 74, 196–204. doi: 10.1111/j.1574-6941.2010.00926.x
Ma, W. T., Lin, J. H., Chen, H. J., Chen, S. Y., and Shaw, G. C. (2011). Identification and characterization of a novel class of extracellular poly (3-Hydroxybutyrate) Depolymerase from Bacillus sp. strain NRRL B-14911. Appl. Environ. Microbiol. 77, 7924–7932. doi: 10.1128/AEM.06069-11
Mao, H., Jiang, H., Su, T., and Wang, Z. (2013). Purification and characterization of two extracellular polyhydroxyalkanoate depolymerases from Pseudomonas mendocina. Biotechnol. Lett. 35, 1919–1924. doi: 10.1007/s10529-013-1288-1
Martínez, V., De La Peña, F., García-Hidalgo, J., De La Mata, I., García, J. L., and Prieto, M. A. (2012). Identification and biochemical evidence of a medium-chain-length Polyhydroxyalkanoate Depolymerase in the Bdellovibrio bacteriovorus predatory hydrolytic arsenal. Appl. Environ. Microbiol. 78, 6017–6026. doi: 10.1128/AEM.01099-12
Martínez, V., De Santos, P. G., García-Hidalgo, J., Hormigo, D., Prieto, M. A., Arroyo, M., et al. (2015). Novel extracellular medium-chain-length polyhydroxyalkanoate depolymerase from Streptomyces exfoliatus K10 DSMZ 41693: a promising biocatalyst for the efficient degradation of natural and functionalized mcl-PHAs. Appl. Microbiol. Biotechnol. 99, 9605–9615. doi: 10.1007/s00253-015-6780-1
Meereboer, K. W., Misra, M., and Mohanty, A. K. (2020). Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites. Green Chem. 22, 5519–5558. doi: 10.1039/D0GC01647K
Millan, F., and Hanik, N. (2023). Degradation kinetics of medium chain length Polyhydroxyalkanoate degrading enzyme: a quartz crystal microbalance study. Front. Bioeng. Biotechnol. 11:1303267. doi: 10.3389/fbioe.2023.1303267
Morohoshi, T., Ogata, K., Okura, T., and Sato, S. (2018). Molecular characterization of the bacterial Community in Biofilms for degradation of poly (3-Hydroxybutyrate-co-3-Hydroxyhexanoate) films in seawater. Microbes Environ. 33, 19–25. doi: 10.1264/jsme2.ME17052
Muniyasamy, S., Ofosu, O., John, M. J., and Anandjiwala, R. D. (2016). Mineralization of poly (lactic acid) (PLA), poly (3-hydroxybutyrate-co-valerate) (PHBV) and PLA/PHBV blend in compost and soil environments. J. Renew Mater. 4, 133–145. doi: 10.7569/JRM.2016.634104
Nojiri, M., and Saito, T. (1997). Structure and function of poly (3-hydroxybutyrate) depolymerase from Alcaligenes faecalis T1. J. Bacteriol. 179, 6965–6970. doi: 10.1128/jb.179.22.6965-6970.1997
Numata, K., Abe, H., and Iwata, T. (2009). Biodegradability of poly (hydroxyalkanoate) materials. Materials 2, 1104–1126. doi: 10.3390/ma2031104
Ohura, T., Kasuya, K. I., and Doi, Y. (1999). Cloning and characterization of the Polyhydroxybutyrate Depolymerase Gene of Pseudomonas stutzeri and analysis of the function of substrate-binding domains. Appl. Environ. Microbiol. 65, 189–197. doi: 10.1128/AEM.65.1.189-197.1999
Panaitescu, D. M., Nicolae, C. A., Frone, A. N., Chiulan, I., Stanescu, P. O., Draghici, C., et al. (2017). Plasticized poly (3-hydroxybutyrate) with improved melt processing and balanced properties. J. Appl. Polym. Sci. 134:app. 44810. doi: 10.1002/app.44810
Papageorgiou, A. C., Hermawan, S., Singh, C. B., and Jendrossek, D. (2008). Structural basis of poly (3-Hydroxybutyrate) hydrolysis by Pha Z7 Depolymerase from Paucimonas lemoignei. J. Mol. Biol. 382, 1184–1194. doi: 10.1016/j.jmb.2008.07.078
Papaneophytou, C. P., Pantazaki, A. A., and Kyriakidis, D. A. (2009). An extracellular polyhydroxybutyrate depolymerase in Thermus thermophilus HB8. Appl. Microbiol. Biotechnol. 83, 659–668. doi: 10.1007/s00253-008-1842-2
Papaneophytou, C. P., Velali, E. E., and Pantazaki, A. A. (2011). Purification and characterization of an extracellular medium-chain length polyhydroxyalkanoate depolymerase from Thermus thermophilus HB8. Polym. Degrad. Stab. 96, 670–678. doi: 10.1016/j.polymdegradstab.2010.12.015
Park, S. L., Cho, J. Y., Choi, T. R., Song, H. S., Bhatia, S. K., Gurav, R., et al. (2021). Improvement of polyhydroxybutyrate (PHB) plate-based screening method for PHB degrading bacteria using cell-grown amorphous PHB and recovered by sodium dodecyl sulfate (SDS). Int. J. Biol. Macromol. 177, 413–421. doi: 10.1016/j.ijbiomac.2021.02.098
Persico, P., Ambrogi, V., Baroni, A., Santagata, G., Carfagna, C., Malinconico, M., et al. (2012). Enhancement of poly (3-hydroxybutyrate) thermal and processing stability using a bio-waste derived additive. Int. J. Biol. Macromol. 51, 1151–1158. doi: 10.1016/j.ijbiomac.2012.08.036
Pham, V. H. T., Kim, J., and Chang, S. (2024). A valuable source of promising extremophiles in microbial plastic degradation. Polymers 16:2109. doi: 10.3390/polym16152109
Prudnikova, S. V., Evgrafova, S. Y., and Volova, T. G. (2021). Metabolic activity of cryogenic soils in the subarctic zone of Siberia towards “green” bioplastics. Chemosphere 263:128180. doi: 10.1016/j.chemosphere.2020.128180
Ray, S., and Kalia, V. C. (2017). “Biological significance of degradation of Polyhydroxyalkanoates” in Microbial applications Vol 1 [internet]. eds. V. C. Kalia and P. Kumar (Cham: Springer international publishing), 125–139.
Rosato, A., Romano, A., Totaro, G., Celli, A., Fava, F., Zanaroli, G., et al. (2022). Enzymatic degradation of the Most common aliphatic bio-polyesters and evaluation of the mechanisms involved: an extended study. Polymers 14:1850. doi: 10.3390/polym14091850
Sadocco, P., Nocerino, S., Dubini-Paglia, E., Seves, A., and Elegir, G. (1997). Characterization of a poly (3-hydroxybutyrate) depolymerase fromaureobacterium saperdae: active site and kinetics of hydrolysis studies. J. Environ. Polym. Degrad. 5, 57–65. doi: 10.1007/BF02763569
Saito, T., Suzuki, K., Yamamoto, J., Fukui, T., Miwa, K., Tomita, K., et al. (1989). Cloning, nucleotide sequence, and expression in Escherichia coli of the gene for poly (3-hydroxybutyrate) depolymerase from Alcaligenes faecalis. J. Bacteriol. 171, 184–189. doi: 10.1128/jb.171.1.184-189.1989
Santos, M., Gangoiti, J., Keul, H., Möller, M., Serra, J. L., and Llama, M. J. (2013). Polyester hydrolytic and synthetic activity catalyzed by the medium-chain-length poly (3-hydroxyalkanoate) depolymerase from Streptomyces venezuelae SO1. Appl. Microbiol. Biotechnol. 97, 211–222. doi: 10.1007/s00253-012-4210-1
Sayyed, R. Z., Wani, S. J., Alyousef, A. A., Alqasim, A., Syed, A., and El-Enshasy, H. A. (2019). Purification and kinetics of the PHB depolymerase of Microbacterium paraoxydans RZS6 isolated from a dumping yard. PLoS One 14:e0212324. doi: 10.1371/journal.pone.0212324
Schirmer, A., and Jendrossek, D. (1994). Molecular characterization of the extracellular poly (3-hydroxyoctanoic acid) [P(3HO)] depolymerase gene of Pseudomonas fluorescens GK13 and of its gene product. J. Bacteriol. 176, 7065–7073. doi: 10.1128/jb.176.22.7065-7073.1994
Schirmer, A., Jendrossek, D., and Schlegel, H. G. (1993). Degradation of poly (3-hydroxyoctanoic acid) [P(3HO)] by bacteria: purification and properties of a P(3HO) depolymerase from Pseudomonas fluorescens GK13. Appl. Environ. Microbiol. 59, 1220–1227. doi: 10.1128/aem.59.4.1220-1227.1993
Schirmer, A., Matz, C., and Jendrossek, D. (1995). Substrate specificities of poly (hydroxyalkanoate)-degrading bacteria and active site studies on the extracellular poly (3-hydroxyoctanoic acid) depolymerase of Pseudomonas fluorescens GK13. Can. J. Microbiol. 41, 170–179. doi: 10.1139/m95-184
Schöber, U., Thiel, C., and Jendrossek, D. (2000). Poly (3-Hydroxyvalerate) Depolymerase of Pseudomonas lemoignei. Appl. Environ. Microbiol. 66, 1385–1392. doi: 10.1128/AEM.66.4.1385-1392.2000
Sevakumaran, V., Rashid, N. S. B. T., and Amirul, A. A. (2019). Bio-degradation of polyhydroxyalkanoates (PHA) films in soil and lake environment. Malays. Appl. Biol. 48, 193–198.
Shah, A. A., Hasan, F., Hameed, A., and Ahmed, S. (2007). Isolation and characterisation of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) degrading actinomycetes and purification of PHBV depolymerase from newly isolated Streptoverticillium kashmirense AF1. Ann. Microbiol. 57, 583–588. doi: 10.1007/BF03175359
Shah, A. A., Hasan, F., Hameed, A., and Ahmed, S. (2008). Biological degradation of plastics: a comprehensive review. Biotechnol. Adv. 26, 246–265. doi: 10.1016/j.biotechadv.2007.12.005
Shinomiya, M., Iwata, T., Kasuya, K., and Doi, Y. (2006). Cloning of the gene for poly (3-hydroxybutyric acid) depolymerase of Comamonas testosteroni and functional analysis of its substrate-binding domain. FEMS Microbiol. Lett. 154, 89–94. doi: 10.1111/j.1574-6968.1997.tb12628.x
Sintim, H. Y., Bary, A. I., Hayes, D. G., Wadsworth, L. C., Anunciado, M. B., English, M. E., et al. (2020). In situ degradation of biodegradable plastic mulch films in compost and agricultural soils. Sci. Total Environ. 727:138668. doi: 10.1016/j.scitotenv.2020.138668
Spasic, J., Mandic, M., Djokic, L., and Nikodinovic-Runic, J. (2018). Streptomyces spp. in the biocatalysis toolbox. Appl. Microbiol. Biotechnol. 102, 3513–3536. doi: 10.1007/s00253-018-8884-x
Tachibana, K., Urano, Y., and Numata, K. (2013). Biodegradability of nylon 4 film in a marine environment. Polym. Degrad. Stab. 98, 1847–1851. doi: 10.1016/j.polymdegradstab.2013.05.007
Tadevosyan, M., Ghevondyan, D., Khoyetsyan, L., Soghomonyan, T., Paloyan, A., Margaryan, A., et al. (2024). Bioplastic-degrading mesophiles from plastisphere and thermophiles from the teristerial geothermal springs. In: Proceedings of 14th international congress on extremophiles.
Takaku, H., Kimoto, A., Kodaira, S., Nashimoto, M., and Takagi, M. (2006). Isolation of a gram-positive poly (3-hydroxybutyrate) (PHB)-degrading bacterium from compost, and cloning and characterization of a gene encoding PHB depolymerase of Bacillus megaterium N-18-25-9. FEMS Microbiol. Lett. 264, 152–159. doi: 10.1111/j.1574-6968.2006.00448.x
Takeda, M., Kamagata, Y., Ghiorse, W. C., Hanada, S., and Koizumi, J. (2002). Caldimonas manganoxidans Gen. Nov., sp. nov., a poly (3-hydroxybutyrate)-degrading, manganese-oxidizing thermophile. Int. J. Syst. Evol. Microbiol. 52, 895–900. doi: 10.1099/00207713-52-3-895
Takeda, M., Kitashima, K., Adachi, K., Hanaoka, Y., Suzuki, I., Koizumi, J., et al. (2000). Cloning and expression of the gene encoding thermostable poly (3-hydroxybutyrate) depolymerase. J. Biosci. Bioeng. 90, 416–421. doi: 10.1016/S1389-1723(01)80011-6
Takeda, M., Koizumi, J. I., Yabe, K., and Adachi, K. (1998). Thermostable poly (3-hydroxybutyrate) depolymerase of a thermophilic strain of Leptothrix sp. isolated from a hot spring. J. Ferment. Bioeng. 85, 375–380. doi: 10.1016/S0922-338X(98)80080-9
Tezuka, Y., Ishii, N., Kasuya, K., and Mitomo, H. (2004). Degradation of poly (ethylene succinate) by mesophilic bacteria. Polym. Degrad. Stab. 84, 115–121. doi: 10.1016/j.polymdegradstab.2003.09.018
Thomas, G. M., Quirk, S., Huard, D. J. E., and Lieberman, R. L. (2022). Bioplastic degradation by a polyhydroxybutyrate depolymerase from a thermophilic soil bacterium. Protein Sci. 31:e4470. doi: 10.1002/pro.4470
Tokiwa, Y., and Calabia, B. P. (2004). Review degradation of microbial polyesters. Biotechnol. Lett. 26, 1181–1189. doi: 10.1023/B:BILE.0000036599.15302.e5
Uefuji, M., Kasuya, K., and Doi, Y. (1997). Enzymatic degradation of poly[(R)-3-hydroxybutyrate]: secretion and properties of PHB depolymerase from Pseudomonas stutzeri. Polym. Degrad. Stab. 58, 275–281. doi: 10.1016/S0141-3910(97)00058-X
Valle Iulianelli, G. C., Azevedo, R. D. S., Da Silva, P. S. R. C., and Tavares, M. I. B. (2016). PHB nanostructured: production and characterization by NMR relaxometry. Polym. Test. 49, 57–65. doi: 10.1016/j.polymertesting.2015.11.011
Vigneswari, S., Lee, T. S., Bhubalan, K., and Amirul, A. A. (2015). Extracellular Polyhydroxyalkanoate Depolymerase by Acidovorax sp. DP5. Enzyme Res. 2015, 1–8. doi: 10.1155/2015/212159
Viljakainen, V. R., and Hug, L. A. (2021). The phylogenetic and global distribution of bacterial polyhydroxyalkanoate bioplastic-degrading genes. Environ. Microbiol. 23, 1717–1731. doi: 10.1111/1462-2920.15409
Volova, T. G., Boyandin, A. N., Vasiliev, A. D., Karpov, V. A., Prudnikova, S. V., Mishukova, O. V., et al. (2010). Biodegradation of polyhydroxyalkanoates (PHAs) in tropical coastal waters and identification of PHA-degrading bacteria. Polym. Degrad. Stab. 95, 2350–2359. doi: 10.1016/j.polymdegradstab.2010.08.023
Volova, T. G., Gladyshev, M. I., Trusova, M. Y., and Zhila, N. O. (2006). Degradation of polyhydroxyalkanoates and the composition of microbial destructors under natural conditions. Microbiology 75, 593–598. doi: 10.1134/S0026261706050092
Wang, Z., Gao, J., Li, L., and Jiang, H. (2012). Purification and characterization of an extracellular poly (3-hydroxybutyrate-co-3-hydroxyvalerate) depolymerase from Acidovorax sp. HB01. World J. Microbiol. Biotechnol. 28, 2395–2402. doi: 10.1007/s11274-012-1048-8
Wu, C. S. (2009). Renewable resource-based composites of recycled natural fibers and maleated polylactide bioplastic: characterization and biodegradability. Polym. Degrad. Stab. 94, 1076–1084. doi: 10.1016/j.polymdegradstab.2009.04.002
Yadav, K., and Nikalje, G. C. (2024). Comprehensive analysis of bioplastics: life cycle assessment, waste management, biodiversity impact, and sustainable mitigation strategies. Peer J. 12:e18013. doi: 10.7717/peerj.18013
Zampolli, J., Vezzini, D., Brocca, S., and Di Gennaro, P. (2024). Insights into the biodegradation of polycaprolactone through genomic analysis of two plastic-degrading Rhodococcus bacteria. Front. Microbiol. 14:1284956. doi: 10.3389/fmicb.2023.1284956
Zandieh, M., Griffiths, E., Waldie, A., Li, S., Honek, J., Rezanezhad, F., et al. (2024). Catalytic and biocatalytic degradation of microplastics. Exploration 4:20230018. doi: 10.1002/EXP.20230018
Zhang, L., Chen, F., Zeng, Z., Xu, M., Sun, F., Yang, L., et al. (2021). Advances in metagenomics and its application in environmental microorganisms. Front. Microbiol. 12:766364. doi: 10.3389/fmicb.2021.766364
Keywords: polyhydroxyalkanoates (PHAs), biodegradation, PHA depolymerases, extremophiles, circular bioeconomy
Citation: Paloyan A, Tadevosyan M, Ghevondyan D, Khoyetsyan L, Karapetyan M, Margaryan A, Antranikian G and Panosyan H (2025) Biodegradation of polyhydroxyalkanoates: current state and future prospects. Front. Microbiol. 16:1542468. doi: 10.3389/fmicb.2025.1542468
Edited by:
Anna Maria Puglia, University of Palermo, ItalyReviewed by:
Jia Wang, The University of Tennessee, United StatesCopyright © 2025 Paloyan, Tadevosyan, Ghevondyan, Khoyetsyan, Karapetyan, Margaryan, Antranikian and Panosyan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hovik Panosyan, aHBhbm9zeWFuQHlzdS5hbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.