
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 12 March 2025
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1539629
This article is part of the Research Topic Advancing Treatments for Protozoan Diseases: From Resistance Mechanisms to Novel Therapies View all 10 articles
 Belén Jesús Maciel1,2
Belén Jesús Maciel1,2 Chantal Reigada1,2
Chantal Reigada1,2 Fabio Augusto Digirolamo1,2
Fabio Augusto Digirolamo1,2 Marcos Rengifo1,2
Marcos Rengifo1,2 Claudio Alejandro Pereira1,2
Claudio Alejandro Pereira1,2 Mariana Reneé Miranda1,2*†
Mariana Reneé Miranda1,2*† Melisa Sayé1,2*†
Melisa Sayé1,2*†Chagas disease, caused by the parasite Trypanosoma cruzi, affects 6 million people worldwide. Although the drugs benznidazole (BZN) and nifurtimox are available to treat Chagas, they are not effective in the chronic phase when most patients are diagnosed. Moreover, long-term regimen and severe side effects often lead to poor adherence and treatment abandonment. These problems highlight the urgent need to develop new therapies to treat this neglected disease. Given that the antifungal drug nystatin (NYS) affects arginine uptake in yeasts, and fluctuations on arginine availability through transport processes in T. cruzi can negatively affect its viability, in this work we evaluated the potential of NYS for drug repurposing against T. cruzi. NYS inhibited arginine uptake and presented trypanocidal effect in both epimastigotes (IC50 0.17 μM) and trypomastigotes (IC50 4.90 μM). In addition, treatment of infected cells with NYS decreased the release of trypomastigotes with better efficacy than BZN (IC50s 4.83 μM and 8.60 μM, respectively) suggesting that NYS affects the progression of the intracellular life cycle. Furthermore, we observed a synergistic effect both in isolated trypomastigotes and infected cells when NYS was combined with BZN, which could enhance efficacy while improving treatment safety and adherence. As in yeasts, the mechanism of action of NYS in T. cruzi involved the plasma membrane disruption, and membrane transport processes, like amino acids and thymidine uptake, were affected prior to the disruption probably due to NYS interaction with the membrane. Drug repurposing is a recommended strategy by the World Health Organization to develop new therapeutic alternatives for neglected diseases. Our results indicate that NYS presents great potential to be repurposed as a trypanocidal drug to fight T. cruzi.
Chagas disease, caused by Trypanosoma cruzi, affects 6 million people worldwide and other 70 million are at risk of getting infected (World Health Organization, 2015). Only two medications are available to treat this neglected disease, nifurtimox and benznidazole (BZN). Both were discovered more than half century ago and the long-term therapies are associated with severe side effects that often lead to treatment abandonment. Moreover, these drugs are not effective in the chronic stage when most patients are diagnosed (Rassi et al., 2010; Zuma and de Souza, 2021) and the randomized trial BENEFIT (BENznidazole Evaluation For Interrupting Trypanosomiasis) further demonstrated that BZN does not halt disease progression (Morillo et al., 2015). These problems highlight the urgent need to develop new alternative therapies to treat Chagas disease.
Nystatin (NYS) is a polyene drug with antifungal and fungistatic activity that is used to treat several yeasts and fungi infections in humans. In addition to sterol binding and pore formation (Kristanc et al., 2019), NYS inhibits arginine uptake by targeting the arginine specific transporter CAN1 in Saccharomyces cerevisiae (Stachiewicz and Quastel, 1963; Opekarová and Tannerb, 1994). The structure, chemistry, action mechanism, and resistance of NYS resemble those of amphotericin B, a drug used to treat leishmaniasis, another neglected disease caused by the trypanosomatid Leishmania spp. (Kumari et al., 2022). NYS is included in the 23rd Model List of Essential Medicines (2023) as a safe and cost-effective medicine for the healthcare system (World Health Organization, 2023). Drug repurposing, which refers to the identification of new clinical applications for approved or investigational drugs, is a strategy recommended by the World Health Organization to accelerate the development of treatments for neglected diseases. This approach reduces time and investment costs because safety profile and pharmacological properties, for example, are already known (Ashburn and Thor, 2004).
Amino acid metabolism plays a crucial role in the parasite T. cruzi, since amino acids are involved in a wide variety of essential processes like protein synthesis, osmoregulation, stress resistance and parasite stage differentiation, among others (Silber et al., 2005). Particularly, arginine plays a unique role in T. cruzi as it participates in the cell energy management through arginine kinase and phosphoarginine (Pereira et al., 2000). Arginine can be obtained through protein degradation or active uptake and T. cruzi’s viability can be affected through fluctuations in arginine availability (Miranda et al., 2012). Furthermore, RNAi assays in T. brucei have demonstrate the essentiality of its arginine permease TbAAT5-3 (Mathieu et al., 2017).
Given the relevance of arginine in T. cruzi, its metabolism and availability can be considered promising therapeutic targets. A bibliographical search for compounds that inhibit arginine uptake or affect its intracellular concentration in different organisms allowed the identification of NYS. In this work, NYS was evaluated as an arginine transport inhibitor and as trypanocidal compound in T. cruzi in order to investigate its potential for drug repurposing against this parasite.
Epimastigotes of CL Brener strain were cultured at 28°C in plastic flasks containing brain-heart infusion-tryptose (BHT) medium supplemented with 10% fetal calf serum (FCS), 100 U/mL penicillin, 100 μg/mL streptomycin and 20 μg/mL hemin (Camargo, 1964).
HEK293 (human embryo kidney, ATCC CRL-1573), 3T3-L1 (mouse embryo fibroblast, ATCC CL-173) and Vero cells (african green monkey kidney, ATCC CCL-81) were cultured in Minimal Essential Media (MEM) supplemented with 10% FCS, 0.15% (w/v) NaHCO3, 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C, 5% CO2. Vero cells (5 × 104 per well) were infected with trypomastigotes (2.5 × 105 per well) for 24 h. After this period, the infected cells were washed twice with phosphate buffered saline (PBS), the MEM medium was replaced for MEM 4% FCS, and the cells were kept in culture at 37°C, 5% CO2. Trypomastigotes were obtained from infected Vero cells as previously described (Tonelli et al., 2004).
NYS was purchased from Magel (Magel SA Import Medicinal Drugs, Argentina) and benznidazole (BZN) was purified from ABARAX® (Elea Laboratorios, Argentina). Stock solutions were made in DMSO (NYS, 50 mM; BZN, 100 mM) and dilutions were prepared in the same solvent. DMSO did not exceed 1% final concentration, and was also used as no drug control in experiments.
1 × 107 epimastigotes were centrifuged at 8,000 × g for 30 s, and washed once with PBS. Cells were resuspended in 0.1 mL PBS and the assay started by the addition of 0.1 mL of the transport mix containing L-[3H] arginine, [3H] thymidine or [3H] amino acid mix (aspartate, glutamate, glutamine, glycine, leucine, lysine, phenylalanine, serine, tryptophan and valine) (PerkinElmer’s NEN Radiochemicals; 0.4 μCi) (Sayé et al., 2017). Following 15 min incubation at 28°C, reaction was stopped by adding 1 mL of ice-cold PBS. Cells were centrifuged as indicated above, and washed twice. Cell pellets were resuspended in 0.2 mL of water and counted for radioactivity in UltimaGold XR liquid scintillation cocktail (Packard Instrument Co., Meridien CT, United States). Non-specific uptake and carry over were assayed without incubation (T0), or incubated at 4°C. Normal parasite motility and morphology were confirmed by direct microscopic examination after incubation with the drug.
1 × 107 epimastigotes/mL in exponential growth phase were cultured in BHT medium with increasing concentrations of NYS and BZN in a 96-well plate to a final volume of 200 μL at 28°C. Parasite growth was determined after 24 h by counting in Neubauer chamber (Galceran et al., 2023).
1 × 106 trypomastigotes/mL were incubated with increasing concentrations of the drugs in a 96-well plate to a final volume of 200 μL for 24 h at 37°C. Survival was determined by counting in a Neubauer chamber. Only swimming parasites were counted (Galceran et al., 2023).
Membrane permeabilization assay was performed as previously described (Reigada et al., 2017). Briefly, 108 epimastigotes were incubated in PBS with increasing concentrations of NYS (0, 0.75, 1.0, 1.25, 1.5 and 2.0 μM). After 30 min of incubation with the drug at room temperature, the tubes were centrifuged, supernatants were kept on ice and pellets were resuspended in the same buffer. Permeabilization with digitonin was used as a positive control; cells were washed twice and resuspended in 50 mM Tris-HCl buffer, pH 7.5, 0.25 M sucrose and 10 μM E64. 108 parasites were mixed with 100 μL of the same buffer containing 0.3 mg/mL of digitonin. After incubation at room temperature, the tubes were centrifuged, supernatants were transferred to new tubes and pellets were resuspended in the same buffer. Samples of all supernatant and pellet fractions corresponding to 0.5 × 107 parasites were run on 15% SDS-PAGE gels and transferred onto a PVDF membrane. Western blot was performed with primary rabbit antibodies anti-glutamate dehydrogenase (1:5,000 dilution) followed by incubation with peroxidase-conjugated anti-rabbit (1:5,000 dilution). The peroxidase-labeled proteins were revealed using Super Signal West Pico Chemiluminescent substrate following the manufacturer instructions (Pierce, Waltham, MA, United States).
After treatment with NYS, membrane integrity was determined by ethidium bromide (EtBr) exclusion as described by Schoijet et al. (2008). Briefly, 107 epimastigotes resuspended in PBS were incubated with 50 μM EtBr for 5 min. Then, increasing concentrations of NYS (0, 0.5, 0.75, 1.0, 1.25, 1.5, 2.0, 2.5 and 3.0 μM) were added and fluorescence was monitored for 10 min. Finally, Triton X-100 0.1% was used to achieve maximum fluorescence (total loss of membrane integrity) and once it was added to the samples, fluorescence was monitored for another 10 min. EtBr fluorescence was recorded (λexcitation 365 nm, λemission 580 nm) with a Sinergy HTX (Biotek) spectrofluorimeter. Linear regression was applied for each concentration and the slopes were compared to the control condition (NYS 0 μM).
The toxicity of NYS was determined in HEK293, 3T3-L1 and Vero cells by the gentian violet staining assay as previously described (Reigada et al., 2019). Briefly, 1 × 104 cells/well were loaded onto a 96-well plate with MEM FCS 10% and maintained for 24 h at 37°C. Medium was replaced with 150 μL MEM containing increasing concentration of the drug and incubated for other 24 h. Cells were fixed and stained with 0.5% gentian violet. Cell viability was measured by absorbance at 570 nm. The selectivity index (SI) was calculated as the ratio between the IC50 obtained in mammalian cells and the IC50 for trypomastigotes.
3 × 104 Vero cells/well were settled on a 24-well plate with MEM 10% FCS 500 μL, and cells were infected with 3 × 105 trypomastigotes/well for 24 h. Then, medium was replaced with 500 μL of MEM 4% FCS containing increasing concentrations of NYS and BZN or their combinations and after 24 h of incubation the drugs were removed. Trypomastigotes were counted at days 8 and 9 post infection in a Neubauer chamber (Galceran et al., 2023). From day 6 post infection medium containing trypomastigotes was totally removed from the wells and then replaced with fresh MEM. Day 9 post infection was selected to report the IC50 values.
Trypomastigotes were treated with NYS and BZN following the same procedure described in 2.5. The concentrations were chosen below the IC50 values of each drug alone and combined in a 3 × 3 matrix. BZN: 0.1 μM, 0.5 μM and 1.0 μM; NYS: 0.1 μM, 0.5 μΜ and 1.0 μM. The two best combinations were also evaluated on infected Vero cells to study the effect on the trypomastigote release according to the procedure described in 2.8. Calculations were performed using CompuSyn software1 which is based on the median-effect equation (Chou, 1976) and its extension, the combination index (CI) recognized as the standard measure of combination effect (Chou and Talalay, 1984). Results are reported as CI, where CI < 1, CI = 1 and CI > 1 indicate synergism, additive effect and antagonism, respectively. The fraction affected (Fa) represents the fraction of parasites affected by the drugs (0–1), where 0 indicates no effect and 1 total effect (100% death). Dose-reduction index (DRI) represents the folds a dose of each drug in a synergistic combination may be reduced when compared with the dose of each drug alone to achieve the same effect. DRI >1 indicates favorable dose-reduction.
IC50 values were obtained by non-linear regression of dose-response logistic functions using GraphPad Prism 6.01 for Windows. Results are presented as IC50 value with their corresponding 95% confidence interval (95% CI). Slopes for EtBr exclusion were obtained by linear regression also using GraphPad Prism 6.01 for Windows. Combined treatment was analyzed using the free software CompuSyn as mentioned in section 2.9. The parameters CI and DRI are expressed as ± standard deviation (SD). All experiments were performed at least in triplicate.
Since NYS was identified through a bibliographical search of arginine uptake inhibitors, we first studied its effect on arginine transport in T. cruzi. By using increasing amounts of the drug, the concentration of NYS that inhibits 50% (IC50) of the transport activity in epimastigotes was calculated to be 1.02 μM (95% CI: 0.87–1.21) (Figure 1A).
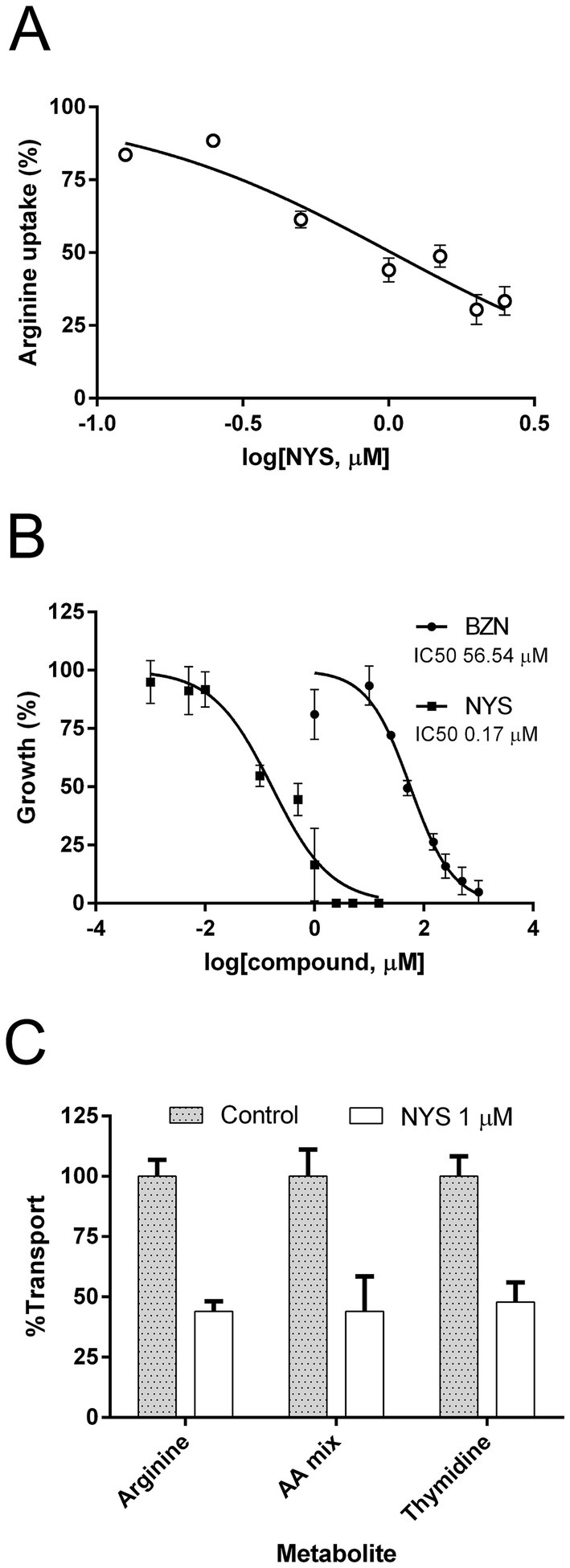
Figure 1. Effect of NYS on T. cruzi epimastigotes. (A) Inhibition of arginine uptake was evaluated using increasing concentration of NYS (from 0 to 2.5 μM). (B) Trypanocidal activity was evaluated incubating epimastigotes with concentrations of NYS (■, 0–15 μM) or BZN (●, 0–1,000 μM) for 24 h. Symbols represent mean values and error bars correspond to standard deviations (n = 3). Concentrations of drug inhibiting the 50% of arginine uptake and parasite growth (IC50) were calculated using the non-linear regression of dose-response logistic functions with GraphPad Prism software. (C) Uptake of different metabolites was evaluated in presence of 1 μM NYS. The radiolabeled amino acid mixture contained Asp, Glu, Gln, Gly, Leu, Lys, Phe, Ser, Trp, and Val. Graph shows mean values ± standard deviations (n = 3). NYS, nystatin; BZN, benznidazole; AA, amino acid.
The potential trypanocidal action of NYS was also assayed, yielding an IC50 of 0.17 μM (95% CI: 0.11–0.25) for epimastigote growth at 24 h (Figure 1B). The drug BZN was used as reference for trypanocidal effect as it is the current treatment for Chagas disease. Under the same experimental conditions, the IC50 for BZN was higher than the obtained for NYS with a value of 56.54 μM (95% CI: 45.05–70.94) (p-value <0.0001).
The large difference observed between the IC50 values for transport and growth suggests that trypanocidal effect of NYS is unlikely to be caused by arginine uptake inhibition alone.
To explore other possible targets in the parasite, we investigated the effect of NYS on additional membrane uptake processes. We measured the incorporation of a mixture of 10 amino acids (Asp, Glu, Gln, Gly, Leu, Lys, Phe, Ser, Trp, and Val) in the presence or absence of NYS. Since amino acids uptake in T. cruzi primarily occurs through permeases of the TcAAAP family (amino acid/auxin permeases), whose members share 75% of global sequence identity (Bouvier et al., 2004), it is plausible that an arginine transport inhibitor might also affect the activity of other TcAAAP proteins. Additionally, we determined the uptake of thymidine, an unrelated metabolite transported by TcrNT2, a member of the Equilibrative Nucleoside Transporters family (Campagnaro et al., 2018). In both cases, NYS at 1 μM produced a similar inhibitory effect to that observed for arginine transport. Inhibition levels were 56.0% ± 4.1 for arginine, 56.0% ± 14.5 for the amino acids mixture and 53.1% ± 8.12 for thymidine (Figure 1C). These results indicate that NYS does not act exclusively on arginine transport but also inhibits other membrane uptake processes.
The antifungal mechanism of action of NYS has been linked to membrane sterols binding, mainly ergosterol, and pore formation (Kristanc et al., 2019). In order to investigate whether similar mechanisms could be involved in T. cruzi, we studied the membrane stability of the parasites through a permeabilization assay with increasing concentrations of NYS (Figure 2A). The non-ionic detergent digitonin was used as positive control and the cytosolic protein glutamate dehydrogenase (GDH) (Cazzulo, 1994) served as a marker for membrane stability. Membrane permeabilization occurred and depended on NYS concentration, since GDH was only detected on supernatant fractions over 1.75 μM NYS.
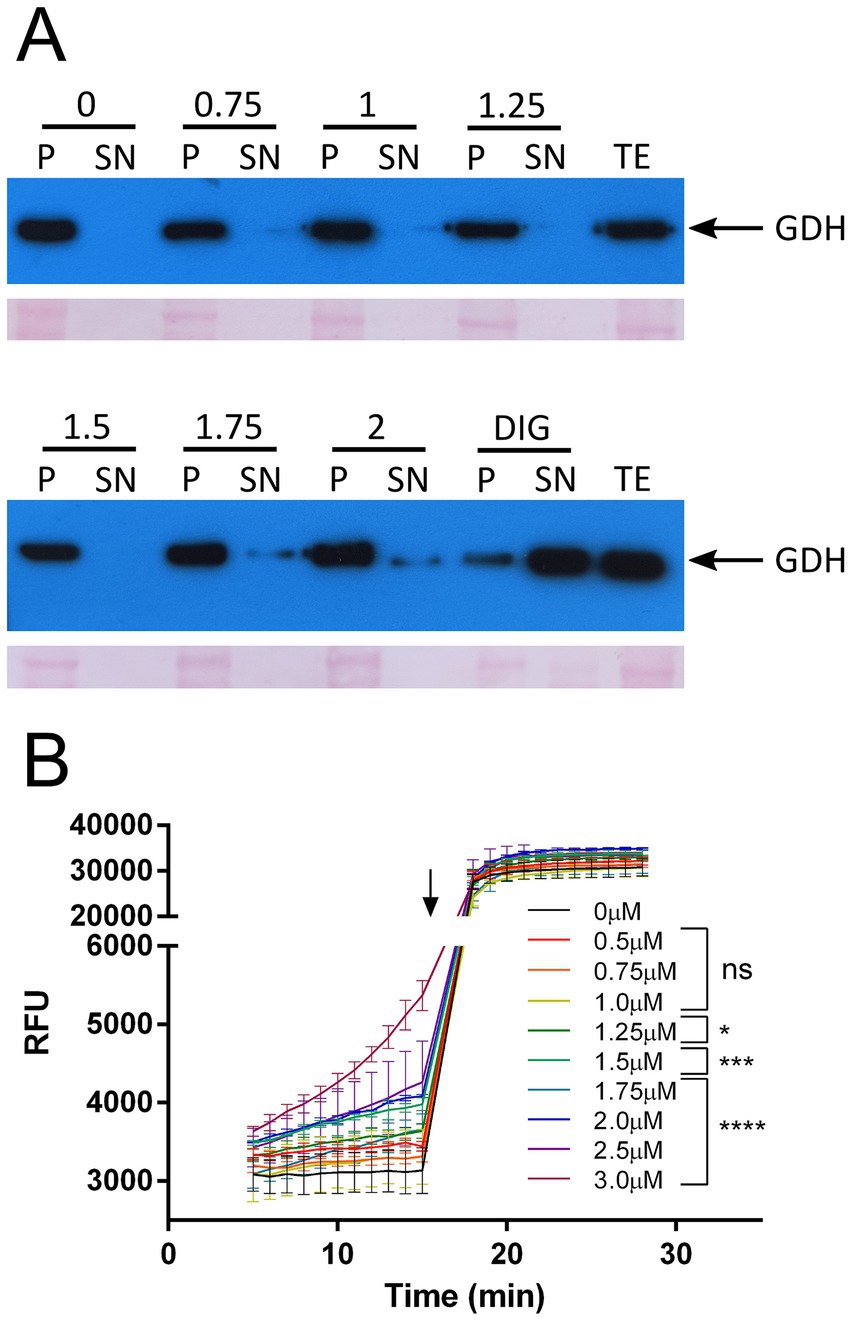
Figure 2. Membrane disruption by NYS. (A) Membrane stability was studied incubating 108 parasites with increasing concentrations of NYS (from 0 to 2 μM) or digitonin as positive control (0.3 mg/mL). Supernatant and pellet fractions were analyzed by Western blot using antibodies anti-T. cruzi glutamate dehydrogenase (GDH) which localizes in the cytosol (Cazzulo, 1994). Ponceau red stained membranes are also shown in the figure. (B) Membrane integrity was quantified using EtBr in presence of NYS (between 0.5 and 3.0 μM). Arrow indicates the time at which Triton X-100 was added. Slopes were calculated for each concentration between 5 and 15 min and compared to the control condition (NYS 0 μM). NYS, nystatin; DIG, digitonin; P, pellet fraction; SN, supernatant fraction; TE, total extract; EtBr, ethidium bromide; RFU, relative fluorescence units; ns, not significant. *p-value >0.05, ***p-value >0.001, and ****p-value >0.0001.
In order to quantify the membrane disruption caused by NYS, we performed an assay of ethidium bromide (EtBr) exclusion (Figure 2B). EtBr presents a fluorescence enhancement upon binding DNA, thus a compromise in membrane integrity would result in augmented fluorescence. The results confirmed that the plasma membrane is affected over 1.25 μM NYS since no significant increase in EtBr fluorescence could be observed with NYS 0.5, 0.75 and 1.0 μM (Figure 2B).
Altogether the results indicate that the mechanism of action of NYS in T. cruzi is through disruption of plasma membrane and at concentrations below 1.25 μM the interaction of NYS with the membrane might be enough to affect membrane transport processes.
Next, the effect of NYS was evaluated on trypomastigotes, the infective and non-replicative stage of T. cruzi in mammalian cells (Figure 3A). The IC50s for survival of trypomastigotes derived from infected Vero cells were similar for NYS and BZN, with values of 4.90 μM (95% CI: 4.46–5.38) and 3.72 μM (95% CI: 3.16–4.38), respectively (p-value = 0.08).
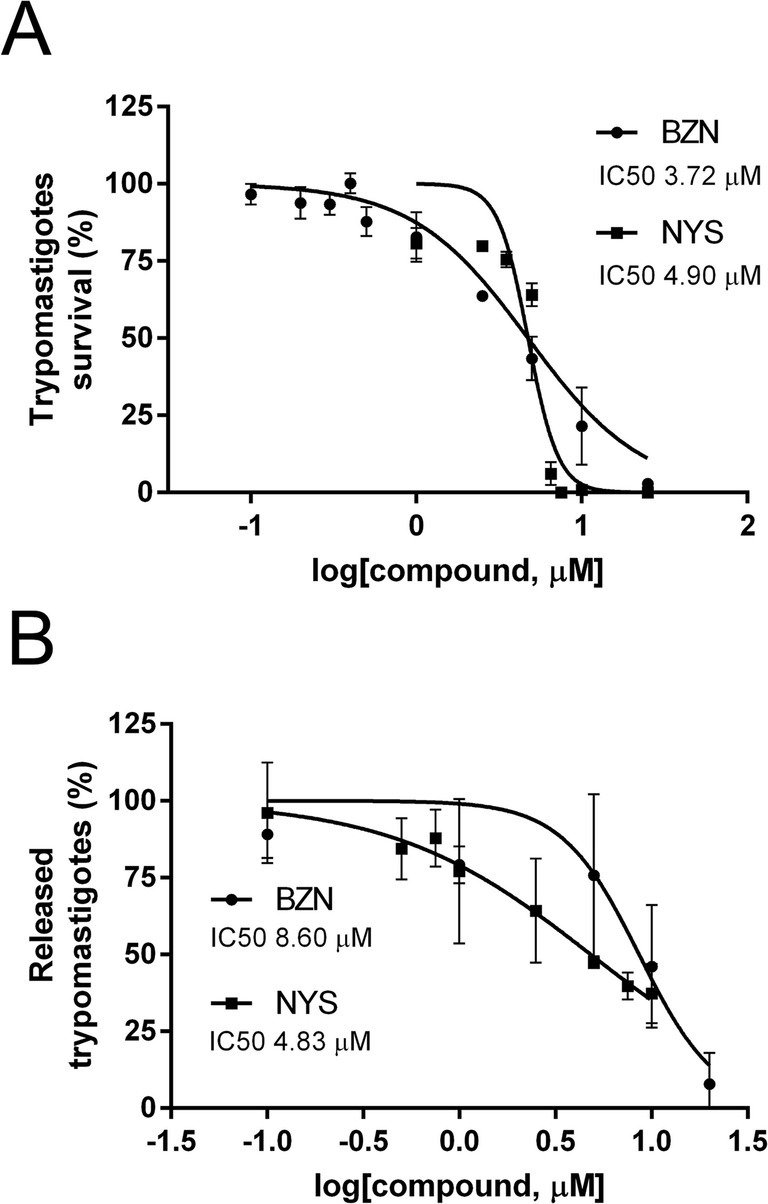
Figure 3. Effect of NYS on trypomastigotes and the intracellular life cycle. (A) Trypomastigotes were incubated for 24 h with increasing concentrations of NYS (■), or BZN (●), both from 0 to 25 μM. (B) Progression of intracellular life cycle was studied using increasing concentrations of the drugs in Vero cells already infected with trypomastigotes for 24 h. NYS (■) was evaluated from 0 to 10 μM and BZN (●) from 0 to 20 μM. Released trypomastigotes were counted at day 9 post-infection. Survival rate of Vero cells at NYS 5 μM was 90.32% ± 12.65. Symbols represent mean values and error bars correspond to standard deviations (n = 3). Concentrations of drug inhibiting the 50% of trypomastigote survival or release (IC50) were calculated using the non-linear regression of dose-response logistic functions with GraphPad Prism software. NYS, nystatin; BZN, benznidazole.
In order to calculate the selectivity index (SI), cytotoxicity was assessed in three different mammalian cell lines: HEK293 (human), 3T3-L1 (mouse) and Vero (monkey). The SI, defined as the ratio between cytotoxicity and trypanocidal activity, reflects the drug’s selectivity for parasites over host cells. NYS presented higher selectivity towards T. cruzi than for the three cell lines (Table 1).
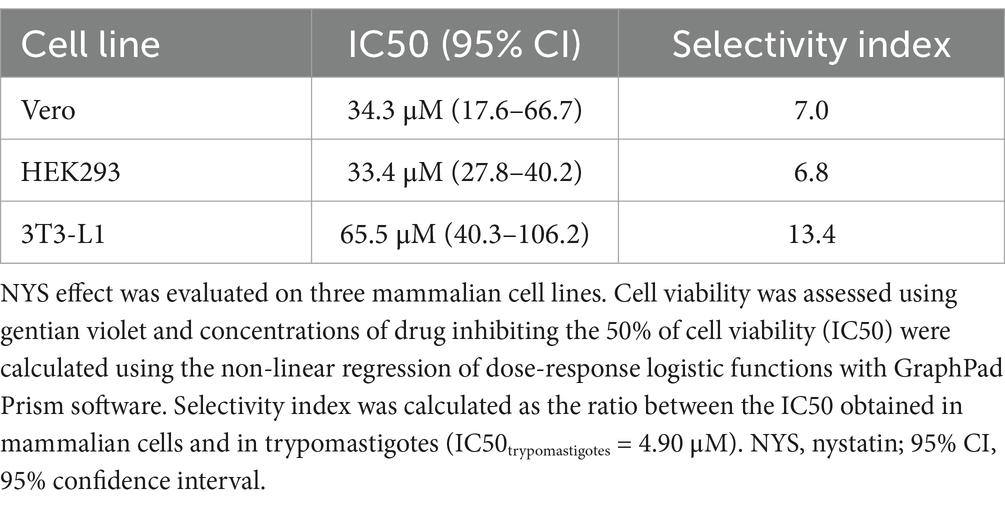
Table 1. NYS cytotoxicity in different mammalian cell lines.
Once the trypomastigotes have entered the host cell, the progression of the intracellular life cycle includes differentiation to amastigotes, replication of these forms, differentiation back to trypomastigotes and finally the cell burst with the release of the accumulated trypomastigotes to the medium. Vero cells were infected and then the cells were exposed to increasing concentrations of the drug. After 24 h treatment, the drug was washed out and, at day 9 post-infection, released trypomastigotes were quantified as a measure of the compound’s impact on the intracellular cycle. The effect of BZN was also assessed, and NYS proved to be more effective in reducing the trypomastigote burst, with IC50 values of 4.83 μM (95% CI 3.50–6.65) for NYS and 8.60 μM (95% CI 6.23–11.87) for BZN (p-value = 0.03) (Figure 3B). These results indicate that NYS affects the progression of the intracellular life cycle as the drug was added after the initial host cell infection.
The combined effect of NYS and BZN on trypomastigotes was explored, with drug interactions classified according to the combination index (CI). The CI is recognized as the standard measure of combination effect where CI < 1 means synergism, CI = 1 indicates additive effect, and CI > 1 is interpreted as antagonism. To preliminary analyze the interaction on isolated trypomastigotes, we selected sub-IC50 concentrations of NYS and BZN and constructed a 3 × 3 combination matrix using 0.1, 0.5 and 1.0 μM of each drug (Table 2). All treatments presented a synergistic effect, with CI values ranging from 0.07 to 0.62, suggesting that the two drugs were more effective in combination than individually. In addition, the dose-reduction index (DRI), which refers to the fold reduction in individual drug dosage achievable through combination for a given effect, was greater than 1 for all tested combinations of NYS and BZN on trypomastigotes. The combinations of 0.1 μM BZN with 0.1 μM and 0.5 μM NYS presented the best CI and DRI values and therefore were selected for evaluation during the progression of the intracellular life cycle on infected cells (Table 3). Both combinations presented synergistic effect with CI values of 0.26 and 0.51 and DRI values for BZN of 17.76 and 41.43. The survival rate of Vero cells with these combinations was 94.26% ± 11.61 and 89.50 ± 6.55, for the combination of 0.1 μM BZN with 0.1 μM and 0.5 μM NYS, respectively. Altogether, these results suggest that the combination have potential to allow dosage reduction and thus reduce adverse effects while keeping drug efficacy.

Table 2. Combined treatment of NYS and BZN on isolated tryposmatigotes.

Table 3. Combined treatment of NYS and BZN on infected Vero cells.
Although Chagas disease was discovered more than a century ago, there is still no effective cure for the chronic stage, where most patients are diagnosed. While posaconazole, fexinidazole and the compound E1224 showed promising results in culture and animal models, unfortunately they failed to cure Chagas in patients (Molina et al., 2014; Morillo et al., 2017; Torrico et al., 2023; Torrico et al., 2018; Pinazo et al., 2024). The development of new therapies to replace or complement conventional treatments remains an urgent need. In the search for these new therapeutic alternatives, drug repurposing and multidrug therapy have emerged as attractive strategies.
Drug repurposing, also known as drug repositioning, involves using an approved drug to treat a different pathology. This approach allows money and time savings because toxicity and pharmacokinetic profiles, among other features, are well-known and available (Ashburn and Thor, 2004). For example, miltefosine and eflornithine, initially discovered as cancer therapies are currently used to treat the trypanosomatid-caused neglected diseases leishmaniasis and human african trypanosomiasis (HAT), respectively (Andrews et al., 2014). Similarly, amphotericin B and paromomycin, which were developed originally to treat invasive mycosis and intestinal infections, are now used also for leishmaniasis (Andrews et al., 2014). Regarding Chagas disease, numerous medications have been studied to be repurposed as trypanocidal agents, including anticancer and antifungal compounds, and are reviewed in Porta et al. (2023). Targeting the TcAAAP family of amino acid and polyamines transporters in T. cruzi, some medicines have also been identified as trypanocidal compounds with potential to be repurposed. Isotretinoin, a drug used for severe acne, inhibits amino acids and polyamines uptake mediated by TcAAAP permeases and exhibits trypanocidal activity (Reigada et al., 2017). In a murine model of chronic Chagas disease, isotretinoin performed better than BZN at reducing parasitemia and anti-T. cruzi antibodies levels as well as preventing tissue damage (Rial et al., 2023). Other drugs evaluated as candidates for drug repurposing include the antihistamines loratadine and cyproheptadine, as well as the antibiotic clofazimine, all of which have been identified as proline uptake inhibitors with trypanocidal activity (Sayé et al., 2020). Recently, liposomal formulations of loratadine and isotretinoin have also demonstrated efficacy against the parasite (Reigada et al., 2024).
Through a bibliographical search, NYS stood out for inhibiting arginine uptake in S. cerevisiae and also for being a medication already approved by health control agencies. Since arginine availability affects the viability of T. cruzi (Miranda et al., 2012), compounds that alter the intracellular pool of this amino acid could exhibit trypanocidal activity. In this work, we investigated the potential of NYS to be repurposed as trypanocidal agent against T. cruzi. Previous reports have explored NYS as a possible repurposed drug for other pathologies such as migraine and viral infections with HIV-1 and SARS-CoV-2 (Selvam et al., 1993; Girotra et al., 2017; Virág, 2021). Furthermore, NYS presents leishmanicidal activity against promastigotes and amastigotes from several Leishmania species and also inhibits parasite entry into macrophages (Ali et al., 1997; Tewary et al., 2006; Yamamoto et al., 2018).
In this work, NYS presented trypanocidal effect and inhibited membrane uptake processes prior disruption of the plasma membrane. NYS affected membrane integrity in T. cruzi epimastigotes in a dose-dependent manner, as evidenced through the permeabilization and EtBr exclusion assays. These results are consistent with the primary mechanism of action of NYS in yeasts that involves sterol binding and pore formation (Kristanc et al., 2019). In T. cruzi, pores would begin to form after incubation with 1.25 μM NYS, allowing EtBr entry and enhanced fluorescence. At 1.75 μM NYS larger pores would be occurring that facilitate the release of proteins like GDH. In addition, NYS altered arginine transport in the parasite as well as the uptake of other amino acids and an unrelated metabolite such as thymidine prior to membrane destabilization, suggesting the non-specificity of NYS over these permeases and that NYS interaction with the membrane might be enough to alter membrane processes.
While the effect of NYS in epimastigotes was stronger than the effect of BZN, in trypomastigotes both drugs presented similar trypanocidal activity. This difference could be related to the membrane composition in these parasite stages and to the different affinity of NYS to cholesterol and ergosterol. Indeed, the plasma membrane of trypomastigotes is thinner than the membrane of epimastigotes suggesting different lipid and protein content (De Souza et al., 1978). For example, ergosterol is the major neutral lipid in the epimastigote forms (Franco Da Silveira and Colli, 1981), while in amastigotes, endogenous sterols are scarce and cholesterol is predominant and it is obtained from the host cell (Liendo et al., 1999). NYS has more affinity for ergosterol than cholesterol, and accordingly, we observed the higher effect of NYS in epimastigotes.
Since drug response can vary between different cell lines, it is important to assess cytotoxicity in diverse models. In this work we evaluated NYS cell lines from various mammalian origins, including human, mouse and monkey cells. Cytotoxicity was cell line-dependent, with Vero cells being more sensitive to NYS than HEK293 and 3T3-L1 cells. Nevertheless, we studied the progression of the intracellular life cycle in Vero cells because it is our routine in vitro infection model.
Combined therapy can offer advantages, such as delayed drug resistance emergence, reduced therapeutic dose and shorter treatment duration, ultimately improving efficacy and minimizing side effects. Although multidrug treatment may seem complex to implement, this approach has been successfully applied to treat neglected diseases, like leprosy (World Health Organization, 2004) and other trypanosomatid-caused diseases. For example, treatment regimens for leishmaniasis often include pentavalent antimonials as first-line drugs alone or in combination with other compounds such as miltefosine or amphotericin B (Berbert et al., 2018). Similarly, nifurtimox-eflornithine combined therapy, referred as NECT, is used to treat the second stage of T. b. gambiense HAT (Yun et al., 2010). These examples demonstrate the therapeutic potential of drug combination for neglected diseases. In this work, we explored the combined action of NYS and BZN on isolated trypomastigotes, resulting in a synergistic effect for all the assayed combinations. For example, the addition of 0.1 and 0.5 μM NYS to 0.1 μM BZN yielded the best CI and DRI values for BZN with a death effect of 0.36–0.49, where 1.0 equals maximum effect, and therefore were selected to be evaluated in the progression of the intracellular cycle. Both combinations presented synergistic effect on infected Vero cells, with CI values of 0.26 and 0.51. The combined treatment with 0.1 μM BZN and 0.5 μM NYS produced 47% death, allowing to reduce 41-folds the dose of BZN needed to individually achieve the same effect. The synergistic combinations have potential to allow reducing BZN dose which could lead to better tolerance and safer treatment while keeping drug efficacy, although further in vivo studies should be carried out to better understand the combined effect of these drugs.
NYS is prescribed primarily for candidal infections of the mucosa, skin, intestinal tract, and vagina. Although NYS is mainly administered topically, alternative drug delivery systems containing NYS have been explored and are reviewed in Sousa et al. (2023). Here we demonstrate that free NYS effectively kills the parasite T. cruzi in its epimastigote and trypomastigote forms with superior or equal efficacy to BZN. NYS alters membrane uptake processes prior to plasma membrane disruption and also affects the progression of the intracellular life cycle, diminishing the burst of trypomastigotes. Moreover, NYS exhibits synergistic effects in combination with BZN both in isolated trypomastigotes and on infected cells. We propose NYS as a promising candidate for drug repurposing to combat T. cruzi. However, further studies in animal models are needed to confirm NYS efficacy as an alternative therapy for Chagas disease.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical approval was not required for the studies on humans in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used. Ethical approval was not required for the studies on animals in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
BM: Investigation, Visualization, Writing – review & editing. CR: Funding acquisition, Investigation, Writing – review & editing. FD: Investigation, Writing – review & editing. MR: Investigation, Writing – review & editing. CP: Conceptualization, Funding acquisition, Writing – review & editing. MM: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – review & editing. MS: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, PIBAA 28720210100409, PIP 0792) and Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT, FONCYT PICT: 2018-1801, 2018-0585, 2018-01871, and 2019-02622).
The authors would like to thank Dr. Fernanda Rubio, Dr. Alejandra Palma, and Francisco Rosa from Laboratorio de Biología Molecular y Apoptosis (IDIM UBA-CONICET) for kindly provided HEK293 and 3T3-L1 cells, Dr. Guillermo Alonso, Dr. Alejandra Schoijet, and Dr. Salomé Vilchez Larrea from Instituto de Investigaciones en Ingeniería Genética y Biología Molecular (INGEBI CONICET), and Agustina Chidichimo and Liliana Sferco from Universidad de San Martín (IIB UNSAM CONICET).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ali, S. A., Iqbal, J., Nabeel, K. Y., Manzoor, A., Bukhari, I., et al. (1997). Leishmanicidal activity of nystatin (mycostatin): a potent polyene compound. J. Pak. Med. Assoc. 47, 246–248.
Andrews, K. T., Fisher, G., and Skinner-Adams, T. S. (2014). Drug repurposing and human parasitic protozoan diseases. Int. J. Parasitol. Drugs Drug Resist. 4, 95–111. doi: 10.1016/j.ijpddr.2014.02.002
Ashburn, T. T., and Thor, K. B. (2004). Drug repositioning: identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 3, 673–683. doi: 10.1038/nrd1468
Berbert, T. R. N., de Mello, T. F. P., Nassif, P. W., Mota, C. A., Silveira, A. V., Duarte, G. C., et al. (2018). Pentavalent antimonials combined with other therapeutic alternatives for the treatment of cutaneous and mucocutaneous leishmaniasis: a systematic review. Dermatol. Res. Pract. :9014726. doi: 10.1155/2018/9014726
Bouvier, A., Silber, A., Galva, C., Canepa, G., Miranda, M., Tonelli, R., et al. (2004). Post genomic analysis of permeases from the amino acid/auxin family in protozoan parasites. Biochem. Biophys. Res. Commun. 321, 547–556. doi: 10.1016/j.bbrc.2004.07.002
Camargo, E. (1964). Growth and differentiation in Trypanosoma cruzi. I. Origin of metacyclic trypanosomes in liquid media. Rev. Inst. Med. São Paulo 6, 93–100.
Campagnaro, G. D., de Freitas, N. J., Girard, R. B. M., Silber, A. M., and de Koning, H. P. (2018). Cloning and characterisation of the equilibrative nucleoside transporter family of Trypanosoma cruzi: ultra-high affinity and selectivity to survive in the intracellular niche. Biochim. Biophys. Acta Gen. Subj. 1862, 2750–2763. doi: 10.1016/j.bbagen.2018.08.015
Cazzulo, J. J. (1994). Intermediate metabolism in Trypanosoma cruzi. J. Bioenerg. Biomembr. 26, 157–165. doi: 10.1007/BF00763064
Chou, T. C. (1976). Derivation and properties of Michaelis–Menten type and hill type equations for reference ligands. J. Theor. Biol. 59, 253–276. doi: 10.1016/0022-5193(76)90169-7
Chou, T. C., and Talalay, P. (1984). Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 22, 27–55. doi: 10.1016/0065-2571(84)90007-4
De Souza, W., Martinez-Palomo, A., and Gonzalez-Robles, A. (1978). The cell surface of Trypanosoma cruzi: cytochemistry and freeze-fracture. J. Cell Sci. 33, 285–299. doi: 10.1242/jcs.33.1.285
Franco da Silveira, J., and Colli, W. (1981). Chemical composition of the plasma membrane from epimastigote forms of Trypanosoma cruzi. Biochim. Biophys. Acta Biomembr. 644, 341–350. doi: 10.1016/0005-2736(81)90392-8
Galceran, F., Digirolamo, F. A., Rengifo, M., Reigada, C., Saye, M., Maciel, B. J., et al. (2023). Identifying inhibitors of Trypanosoma cruzi nucleoside diphosphate kinase 1 as potential repurposed drugs for Chagas’ disease. Biochem. Pharmacol. 216:115766. doi: 10.1016/j.bcp.2023.115766
Girotra, P., Thakur, A., Kumar, A., and Singh, S. K. (2017). Identification of multi-targeted anti-migraine potential of nystatin and development of its brain targeted chitosan nanoformulation. Int. J. Biol. Macromol. 96, 687–696. doi: 10.1016/j.ijbiomac.2016.12.065
Kristanc, L., Božič, B., Jokhadar, Š. Z., Dolenc, M. S., and Gomišček, G. (2019). The pore-forming action of polyenes: from model membranes to living organisms. Biochim. Biophys. Acta Biomembr. 1861, 418–430. doi: 10.1016/j.bbamem.2018.11.006
Kumari, S., Kumar, V., Tiwari, R. K., Ravidas, V., Pandey, K., and Kumar, A. (2022). Amphotericin B: a drug of choice for visceral leishmaniasis. Acta Trop. 235:106661. doi: 10.1016/j.actatropica.2022.106661
Liendo, A., Visbal, G., Piras, M. M., Piras, R., and Urbina, J. A. (1999). Sterol composition and biosynthesis in Trypanosoma cruzi amastigotes. Mol. Biochem. Parasitol. 104, 81–91. doi: 10.1016/S0166-6851(99)00129-2
Mathieu, C., Peireia de Macedo, J., Hürlimann, D., Wirdnam, C., Suter Grotemeyer, M., Gonzales-Salgado, A., et al. (2017). Arginine and lysine transporters are essential for Trypanosoma brucei. PLoS One 12:e0168775. doi: 10.1371/journal.pone.0168775
Miranda, M. R., Sayé, M., Bouvier, L. A., de los Milagros Cámara, M., Montserrat, J., and Pereira, C. A. (2012). Cationic amino acid uptake constitutes a metabolic regulation mechanism and occurs in the flagellar pocket of Trypanosoma cruzi. PLoS One 7:e32760. doi: 10.1371/journal.pone.0032760
Molina, I., Gómezi Prat, J., Salvador, F., Treviño, B., Sulleiro, E., Serre, N., et al. (2014). Randomized trial of posaconazole and benznidazole for chronic Chagas’ disease. N. Engl. J. Med. 370, 1899–1908. doi: 10.1056/NEJMoa1313122
Morillo, C. A., Marin-Neto, J. A., Avezum, A., Sosa-Estani, S., Rassi, A., Rosas, F., et al. (2015). Randomized trial of benznidazole for chronic Chagas’ cardiomyopathy. N. Engl. J. Med. 373, 1295–1306. doi: 10.1056/NEJMoa1507574
Morillo, C., Waskin, H., Sosa-Estani, S., del Carmen Bangher, M., Cuneo, C., Milesi, R., et al. (2017). Benznidazole and Posaconazole in eliminating parasites in asymptomatic T. cruzi carriers. J. Am. Coll. Cardiol. 69, 939–947. doi: 10.1016/j.jacc.2016.12.023
Opekarová, M., and Tannerb, W. (1994). Nystatin changes the properties of transporters for arginine and sugars. An in vitro study. FEBS Lett. 350, 46–50. doi: 10.1016/0014-5793(94)00730-6
Pereira, C. A., Alonso, G. D., Paveto, M. C., Iribarren, A., Cabanas, M. L., Torres, N., et al. (2000). Trypanosoma cruzi arginine kinase characterization and cloning. A novel energetic pathway in protozoan parasites. J. Biol. Chem. 275, 1495–1501. doi: 10.1074/jbc.275.2.1495
Pinazo, M. J., Forsyth, C., Losada, I., Esteban, E. T., García-Rodríguez, M., Villegas, M. L., et al. (2024). Efficacy and safety of fexinidazole for treatment of chronic indeterminate Chagas disease (FEXI-12): a multicentre, randomised, double-blind, phase 2 trial. Lancet Infect. Dis. 24, 395–403. doi: 10.1016/S1473-3099(23)00651-5
Porta, E. O. J., Kalesh, K., and Steel, P. G. (2023). Navigating drug repurposing for Chagas disease: advances, challenges, and opportunities. Front. Pharmacol. 14:1233253. doi: 10.3389/fphar.2023.1233253
Rassi, A. J., Rassi, A., and Marin-Neto, J. A. (2010). Chagas disease. Lancet 375, 1388–1402. doi: 10.1016/S0140-6736(10)60061-X
Reigada, C., Digirolamo, F., Galceran, F., Sayé, M., Carrillo, C., Torres, P., et al. (2024). Trypanocidal activity of liposomal isotretinoin and loratadine formulations. J. Drug Deliv. Sci. Technol. 91:105241. doi: 10.1016/j.jddst.2023.105241
Reigada, C., Saye, M., Valera-Vera, E., Miranda, M. R., and Pereira, C. A. (2019). Repurposing terconazole as an anti Trypanosoma cruzi agent. Heliyon 5:e01947. doi: 10.1016/j.heliyon.2019.e01947
Reigada, C., Valera-Vera, E. A., Sayé, M., Errasti, A. E., Avila, C. C., Miranda, M. R., et al. (2017). Trypanocidal effect of isotretinoin through the inhibition of polyamine and amino acid transporters in Trypanosoma cruzi. PLoS Negl. Trop. Dis. 11:e0005472. doi: 10.1371/journal.pntd.0005472
Rial, M. S., Reigada, C., Prado, N., Bua, J., Esteva, M., Pereira, C. A., et al. (2023). Effectiveness of the repurposed drug isotretinoin in an experimental murine model of Chagas disease. Acta Trop. 2023:106920. doi: 10.1016/j.actatropica.2023.106920
Sayé, M., Fargnoli, L., Reigada, C., Labadie, G. R., and Pereira, C. A. (2017). Evaluation of proline analogs as trypanocidal agents through the inhibition of a Trypanosoma cruzi proline transporter. Biochim. Biophys. Acta Gen. Subj. 1861, 2913–2921. doi: 10.1016/j.bbagen.2017.08.015
Sayé, M., Gauna, L., Valera-Vera, E., Reigada, C., Miranda, M. R., and Pereira, C. A. (2020). Crystal violet structural analogues identified by in silico drug repositioning present anti-Trypanosoma cruzi activity through inhibition of proline transporter TcAAAP069. PLoS Negl. Trop. Dis. 14:e0007481. doi: 10.1371/journal.pntd.0007481
Schoijet, A. C., Miranda, K., Girard-Dias, W., De Souza, W., Flawia, M. M., Torres, H. N., et al. (2008). A Trypanosoma cruzi phosphatidylinositol 3-kinase (TcVps34) is involved in osmoregulation and receptor-mediated endocytosis. J. Biol. Chem. 283, 31541–31550. doi: 10.1074/jbc.M801367200
Selvam, M. P., Blay, R. A., Geyer, S., Buck, S. M., Pollock, L., Mayner, R. E., et al. (1993). Inhibition of HIV-1 replication in H9 cells by nystatin-a compared with other antiviral agents. AIDS Res. Hum. Retrovir. 9, 475–481. doi: 10.1089/aid.1993.9.475
Silber, A. M., Colli, W., Ulrich, H., Alves, M. J. M., and Pereira, C. A. (2005). Amino acid metabolic routes in Trypanosoma cruzi: possible therapeutic targets against Chagas’ disease. Curr. Drug Targets Infect. Disord. 5, 53–64. doi: 10.2174/1568005053174636
Sousa, F., Nascimento, C., Ferreira, D., Reis, S., and Costa, P. (2023). Reviving the interest in the versatile drug nystatin: a multitude of strategies to increase its potential as an effective and safe antifungal agent. Adv. Drug Deliv. Rev. 199:114969. doi: 10.1016/j.addr.2023.114969
Stachiewicz, E., and Quastel, J. H. (1963). Amino acid transport in yeast and effects of nystatin. Can. J. Biochem. Physiol. 41, 397–407. doi: 10.1139/y63-048
Tewary, P., Veena, K., Pucadyil, T. J., Chattopadhyay, A., and Madhubala, R. (2006). The sterol-binding antibiotic nystatin inhibits entry of non-opsonized Leishmania donovani into macrophages. Biochem. Biophys. Res. Commun. 339, 661–666. doi: 10.1016/j.bbrc.2005.11.062
Tonelli, R., Silber, A., Almeida-de-Faria, M., Hirata, I., Colli, W., and Alves, M. (2004). L-Proline is essential for the intracellular differentiation of Trypanosoma cruzi. Cell. Microbiol. 6, 733–741. doi: 10.1111/j.1462-5822.2004.00397.x
Torrico, F., Gascon, J., Ortiz, L., Alonso-Vega, C., Pinazo, M. J., Schijman, A., et al. (2018). Treatment of adult chronic indeterminate Chagas disease: proof-of-concept randomized placebo-controlled study of benznidazole and three E1224 dosing regimens. Lancet Infect. Dis. 18, 419–430. doi: 10.1016/S1473-3099(17)30538-8
Torrico, F., Gascón, J., Ortiz, L., Pinto, J., Rojas, G., Palacios, A., et al. (2023). A phase 2, randomized, multicenter, placebo-controlled, proof-of-concept trial of oral fexinidazole in adults with chronic indeterminate Chagas disease. Clin. Infect. Dis. 76, e1186–e1194. doi: 10.1093/cid/ciac579
Virág, E. (2021). Repurposed nystatin to inhibit SARS-CoV-2 and mutants in the GI tract. Biomed. J. Sci. Tech. Res. 40, 31854–31865. doi: 10.26717/BJSTR.2021.40.006392
World Health Organization (2004). Multidrug therapy against leprosy: development and implementation over the last 25 years. Geneva: World Health Organization, 1–179.
World Health Organization (2015). Chagas disease in Latin America: an epidemiological update based on 2010 estimates. Wkly. Epidemiol. Rec. 90, 33–44.
World Health Organization (2023). The selection and use of essential medicines 2023. Executive Summary of the report of the 24th WHO Expert Committee on Selection and Use of Essential Medicines. Geneva: World Health Organization.
Yamamoto, E. S., Jesus, J. A., Bezerra-Souza, A., Laurenti, M. D., Ribeiro, S. P., and Passero, L. F. D. (2018). Activity of fenticonazole, tioconazole and nystatin on new world Leishmania species. Curr. Top. Med. Chem. 18, 2338–2346. doi: 10.2174/1568026619666181220114627
Yun, O., Priotto, G., Tong, J., Flevaud, L., and Chappuis, F. (2010). NECT is next: implementing the new drug combination therapy for Trypanosoma brucei gambiense sleeping sickness. PLoS Negl. Trop. Dis. 4:e720. doi: 10.1371/journal.pntd.0000720
Keywords: nystatin, drug repurposing, Trypanosoma cruzi , trypanocidal compounds, Chagas disease
Citation: Maciel BJ, Reigada C, Digirolamo FA, Rengifo M, Pereira CA, Miranda MR and Sayé M (2025) The potential of the antifungal nystatin to be repurposed to fight the protozoan Trypanosoma cruzi. Front. Microbiol. 16:1539629. doi: 10.3389/fmicb.2025.1539629
Edited by:
Maria Carolina Touz, Medical Research Institute Mercedes and Martín Ferreyra (INIMEC), ArgentinaReviewed by:
Shahbaz Manzoor Khan, Johns Hopkins University, United StatesCopyright © 2025 Maciel, Reigada, Digirolamo, Rengifo, Pereira, Miranda and Sayé. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Reneé Miranda, bW1pcmFuZGFAY29uaWNldC5nb3YuYXI=; Melisa Sayé, bS5zYXllQGNvbmljZXQuZ292LmFy; bWVsaXNhLm1zYXllQGhvdG1haWwuY29t
†ORCID: Mariana Reneé Miranda, orcid.org/0000-0002-0692-5675
Melisa Sayé, orcid.org/0000-0003-0246-1356
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.