
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 18 February 2025
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1527755
This article is part of the Research TopicThe Interaction Between Food Ingredients and Gut Microbiome on Health and DiseaseView all 11 articles
 Suresh Kumar1*
Suresh Kumar1* Riya Mukherjee2,3
Riya Mukherjee2,3 Pratibha Gaur4,5
Pratibha Gaur4,5 Élcio Leal6
Élcio Leal6 Xiaoming Lyu7
Xiaoming Lyu7 Saheem Ahmad8
Saheem Ahmad8 Paridhi Puri9
Paridhi Puri9 Chung-Ming Chang2,3,10V. Samuel Raj5,6*
Chung-Ming Chang2,3,10V. Samuel Raj5,6* Ramendra Pati Pandey5,6*
Ramendra Pati Pandey5,6*The gut microbiome plays a pivotal role in human health, influencing digestion, immunity, and disease prevention. Beneficial gut bacteria such as Akkermansia muciniphila, Adlercreutzia equolifaciens, and Christensenella minuta contribute to metabolic regulation and immune support through bioactive metabolites like short-chain fatty acids (SCFAs). Dietary patterns rich in prebiotics, fermented foods, and plant-based bioactive compounds, including polyphenols and flavonoids, promote microbiome diversity and stability. However, challenges such as individual variability, bioavailability, dietary adherence, and the dynamic nature of the gut microbiota remain significant. This review synthesizes current insights into gut bacteria’s role in health, emphasizing the mechanisms by which dietary interventions modulate microbiota. Additionally, it highlights advancements in microbiome-targeted therapies and the transformative potential of personalized nutrition, leveraging microbiota profiling and artificial intelligence (AI) to develop tailored dietary strategies for optimizing gut health and mitigating chronic inflammatory disorders. Addressing these challenges requires a multidisciplinary approach that integrates scientific innovation, ethical frameworks, and practical implementation strategies.
The human gut is a fascinating ecosystem that harbors trillions of microorganisms collectively known as gut microbiota that play a crucial role in several physiological functions through their derived metabolites such as nutrient metabolism, immune system regulation, vitamin production, mental health, and brain function that contribute to overall maintaining of health. There is a wide distribution of microbes in the gut which is essential for better health (Figure 1). Some of the key gut bacteria, including Akkermansia muciniphila, Adlercreutzia equolifaciens, Barnesiella, Christensenella minuta, and Oxalobacter formigenes, Lactobacillus, Bifidobacterium, Faecalibacterium prausnitzii, Roseburia spp. contribute to unusual health benefits such as improved gut barrier function, better glucose metabolism, reduced inflammation, weight management, and prevention of kidney stone formation (Anhê et al., 2016; Maruo et al., 2008; Ubeda et al., 2013; Ang et al., 2023; Kaufman et al., 2008).
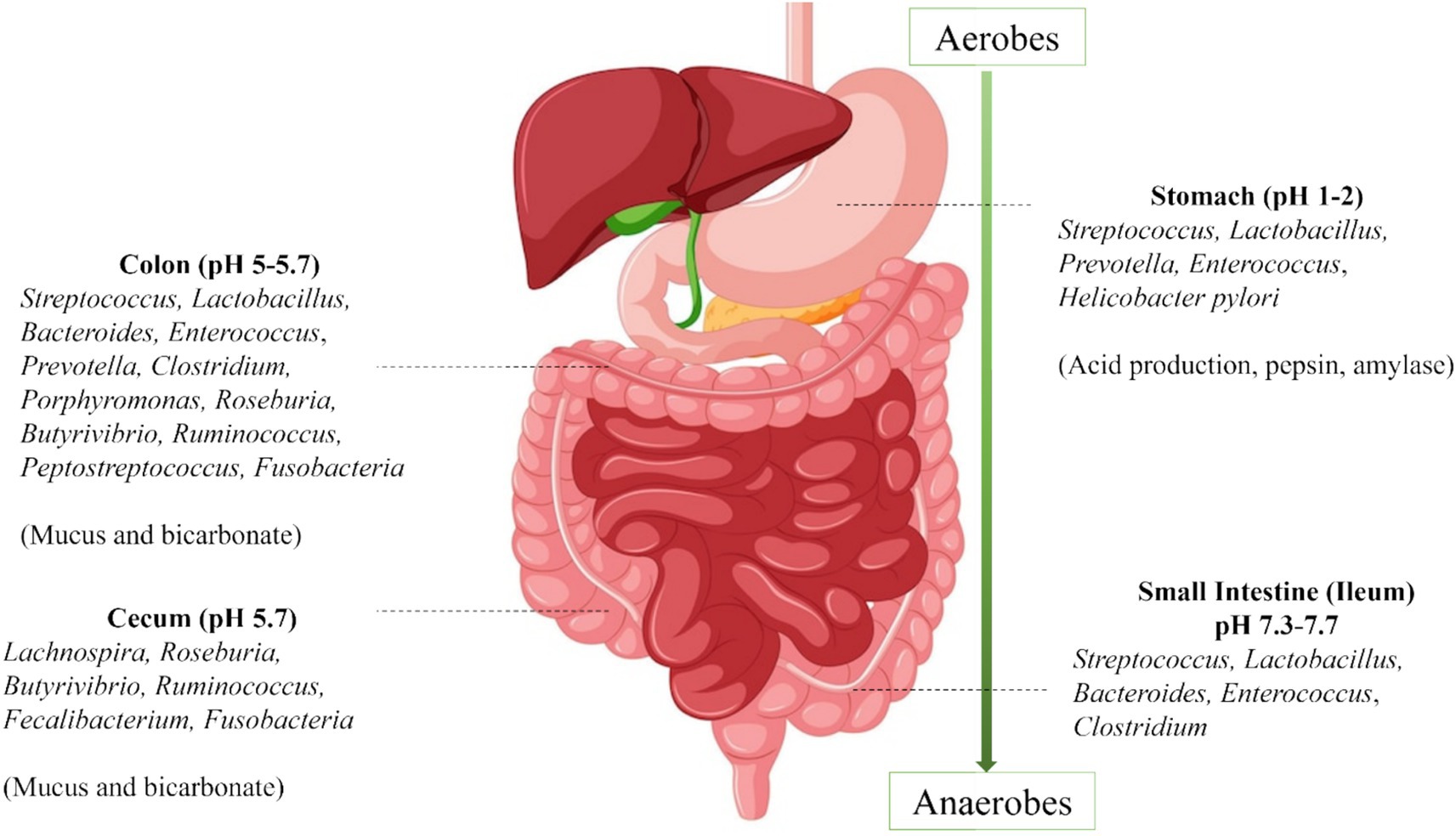
Figure 1. Distribution of gut microbiota.
The dietary sources such as polyphenols, alkaloids, capsaicin, and polysaccharides are essential constituents that support the growth of these beneficial bacteria, leading to improved digestive health and overall well-being (Chenbo et al., 2022; Othman et al., 2019). The metabolites are small molecules that produced by the gut microbiota as byproducts can target directly as well as indirectly both the bacteria themselves and the host, regulating the composition and function of the gut microbiota (Liu et al., 2022; Jandhyala et al., 2015). In recent years, the significance of gut microbiota and their metabolites in influencing human health has garnered significant attention.
The role of gut microbiota in disease initiation is gaining significant attention. Dysbiosis, an imbalance in microbial composition, is strongly associated with the onset of conditions such as inflammatory bowel disease (IBD), metabolic syndrome, and neurodegenerative disorders. Altered microbial populations lead to reduced production of beneficial metabolites like SCFAs and increased levels of pro-inflammatory compounds such as lipopolysaccharides (LPS), disrupting gut barrier integrity and systemic homeostasis (Tan et al., 2014). Additionally, microbial interactions with dietary components such as fibers, polyphenols, and omega-3 fatty acids further influence health outcomes by modulating inflammation, immune signaling, and metabolic pathways (Sultan et al., 2021).
Several studies have focused on elucidating the specific functions and mechanisms of these microorganisms in relation to various diseases (Lavelle and Sokol, 2020; Parada Venegas et al., 2019; Liu et al., 2022). There is currently a lack of a comprehensive summarization regarding the most beneficial gut bacteria, their derived metabolites, and their dietary sources that contribute to maintaining overall health. In this review, we provide the potential role of microbiota in human health. We also discuss the essential gut bacteria and emphasize the derived metabolites of these gut bacteria especially the functions of regulating the local and systemic immune system, energy metabolism, and neural activity. Finally, we discuss how dietary approaches impacts the gut microbial ecology, and foods that promote healthier and more resilient intestinal gut bacteria.
The microbial balance in the intestine is closely related to human diseases and wellness. Extensive research has uncovered the crucial link between gut bacteria and fundamental human biological functions. Because of the varied metabolic genes encoding distinct enzymes and biochemical pathways, the microbiota plays critical roles in energy and nutrition extraction from food (Hou et al., 2022). Furthermore, the creation of bioactive compounds, including vitamins, amino acids, and lipids, is heavily reliant on gut bacteria (Roberfroid et al., 2009). The human microbiota not only protects the host from external pathogens by creating antimicrobial compounds but also plays an essential role in the development of intestinal mucosa and the immune system.
One of the key mechanisms by which the gut microbiota influences health is through the production of short-chain fatty acids (SCFAs), primarily acetate, propionate, and butyrate, which are generated via the fermentation of dietary fibers. SCFAs play a multifaceted role in maintaining gut homeostasis. Butyrate serves as the primary energy source for colonocytes, promoting tight junction integrity and reducing gut permeability, thereby reinforcing the gut barrier. Propionate and acetate, on the other hand, influence systemic metabolism by modulating gluconeogenesis and lipid biosynthesis in the liver (Tan et al., 2014). Beyond metabolic regulation, SCFAs exert anti-inflammatory effects by interacting with G-protein-coupled receptors (GPCRs) and inhibiting histone deacetylases (HDACs), which regulate immune and inflammatory pathways (Yao et al., 2024; Liu et al., 2023).
The immune regulatory role of the microbiota is also mediated through the modulation of nuclear factor kappa B (NF-κB), a critical transcription factor that governs the expression of pro-inflammatory cytokines and antimicrobial peptides. Dysbiosis, or an imbalance in gut microbiota, can lead to overactivation of NF-κB, driving chronic inflammation and increasing susceptibility to inflammatory bowel disease and other gut-related disorders. SCFAs, particularly butyrate, have been shown to inhibit NF-κB activation by preventing the phosphorylation and degradation of inhibitor proteins (IκBs), thereby mitigating inflammatory responses and promoting gut homeostasis (Yao et al., 2024; Liu et al., 2023).
The gut microbiota demonstrates stability, resilience, and symbiotic relationships with the host in healthy environments. Its composition varies significantly across different anatomical sections of the gastrointestinal tract. For example, Akkermansia muciniphila resides in the mucus layer of the large intestine and is involved in maintaining intestinal integrity by stimulating mucin production and regulating gut permeability. Adlercreutzia equolifaciens is commonly found in the colon, where it metabolizes soy isoflavones into bioactive equol, a compound with antioxidant and estrogenic properties. Barnesiella has been discovered in multiple sections of the gastrointestinal tract, including the cecum, colon, and feces, where it contributes to the breakdown of complex carbohydrates and the modulation of immune responses (Afzaal et al., 2022).
Such discrepancies in microbial distribution are primarily attributable to differences in local conditions. The small intestine, for instance, has a rapid transit time and high bile content, while the colon exhibits slower flow rates, a gentler pH, and larger microbial communities dominated by anaerobic species (Afzaal et al., 2022). Aside from regional variation, gut microbiota composition also changes with age. Microbial diversity typically increases from childhood to adulthood and gradually declines after the age of 70, contributing to age-associated vulnerabilities in immune and metabolic health.
The composition and functionality of gut microbiota are profoundly influenced by specific dietary choices, with certain foods playing a pivotal role in promoting the growth of beneficial bacteria. Microbiota-accessible carbohydrates (MACs), including dietary fibers such as resistant starch, inulin, and pectin, serve as fermentable substrates for gut bacteria, particularly Bifidobacterium and Faecalibacterium prausnitzii. These fibers stimulate the production of short-chain fatty acids (SCFAs), which are essential for gut homeostasis. For example, resistant starch found in foods like legumes and whole grains has been shown to increase butyrate production, supporting colonic health and reducing gut inflammation. Similarly, inulin, a naturally occurring polysaccharide present in chicory root and Jerusalem artichoke, selectively enhances the abundance of Bifidobacterium, contributing to improved gut barrier function and immune modulation (Baxter et al., 2019) (Murga-Garrido et al., 2021).
Polyphenols, bioactive compounds abundant in plant-based foods such as berries, cocoa, and green tea, also play a critical role in supporting gut health. These compounds interact with gut microbiota to produce bioactive metabolites that exert anti-inflammatory and antioxidant effects. For instance, anthocyanins in blueberries have been shown to increase the abundance of Akkermansia muciniphila, a bacterium linked to enhanced metabolic health and reduced markers of systemic inflammation (Cano et al., 2024). However, the efficacy of these foods can be significantly influenced by processing methods. High-temperature cooking or refining can degrade fibers and polyphenols, reducing their availability to gut microbes. Minimally processed foods, such as raw fruits, vegetables, and whole grains, retain higher levels of beneficial compounds and are therefore more effective in nurturing gut bacteria (Sejbuk et al., 2024).
Prebiotics, defined as substrates selectively utilized by host microorganisms to confer health benefits, are fundamental to promoting gut health. Common prebiotics include inulin, fructooligosaccharides (FOS), galactooligosaccharides (GOS), and resistant starch, all of which foster the growth of beneficial bacterial taxa such as Bifidobacterium and Faecalibacterium (Krumbeck et al., 2018). Inulin, naturally present in foods like onions, garlic, and asparagus, is known for its ability to selectively enhance Bifidobacterium populations (Davani-Davari et al., 2019). Similarly, FOS, found in bananas, leeks, and artichokes, promotes microbial diversity and supports the production of SCFAs, which are crucial for maintaining gut barrier integrity and reducing inflammation. GOS, commonly derived from dairy products or synthesized for commercial use, has demonstrated particular benefits in infant gut microbiota, simulating the effects of human milk oligosaccharides (Davani-Davari et al., 2019).
Resistant starch, present in foods such as potatoes, green bananas, and legumes, has a unique role in promoting butyrate production. This SCFA is particularly beneficial for colonic health, as it serves as an energy source for colonocytes and reduces intestinal inflammation. However, the efficacy of prebiotics can be influenced by food processing. For example, cooking methods like boiling or steaming generally preserve prebiotic content, whereas high-temperature frying or prolonged storage may degrade these beneficial fibers, reducing their fermentability (Queen and Queen, 2020). To maximize the benefits of prebiotic-rich foods, dietary interventions should prioritize minimally processed options and explore novel methods for enhancing the stability of prebiotics during food preparation (Queen and Queen, 2020).
The intestinal mucosal barrier, which consists of physical, chemical, microbial, and immunological components, serves as one of the body’s major defensive barriers. It protects against bacterial invasion, prevents the entry of foreign antigens and toxins into the circulation, and minimizes water and nutrient loss. This barrier also regulates molecular exchange while supporting the coexistence and colonization of gut bacteria. Disruptions to the gut microbiota’s composition can have significant repercussions, influencing the development of major diseases (Figure 2).
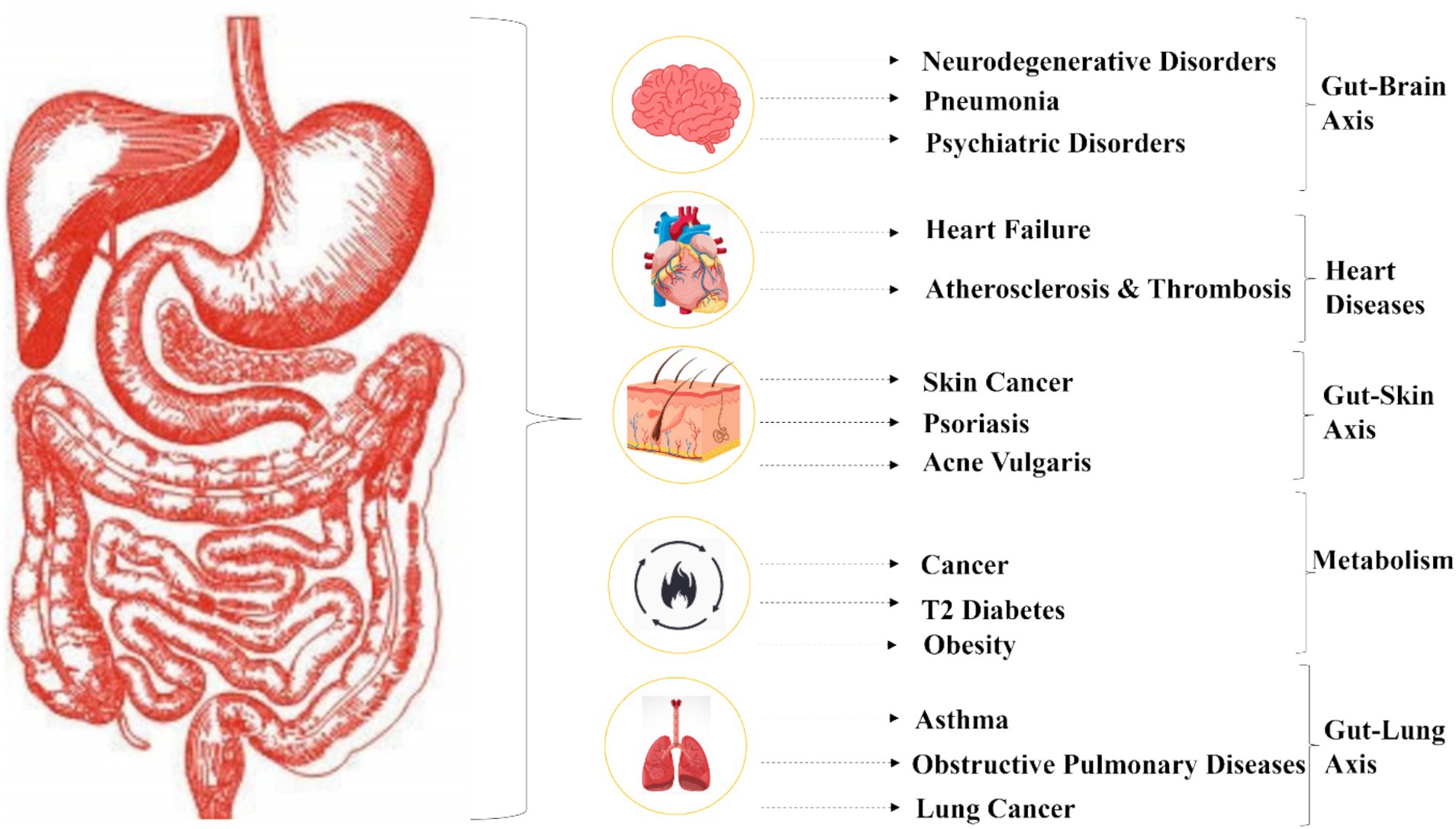
Figure 2. Dysbiosis of the human microbiota.
Dysbiosis, characterized by a loss of beneficial microbes and an overgrowth of pathogenic species, plays a central role in disease induction. The intestinal mucosal barrier becomes compromised, leading to increased gut permeability and systemic inflammation. For instance, in inflammatory bowel disease (IBD), the weakened mucus layer of the digestive tract allows luminal bacteria to penetrate epithelial cells (Bull and Plummer 2015), triggering proliferative and inflammatory processes (Parekh et al., 2015). Dysbiosis also affects the production of key microbial metabolites, such as short-chain fatty acids (SCFAs). Reduced levels of butyrate, a critical SCFA, impair gut barrier integrity and exacerbate inflammation, further contributing to the pathogenesis of IBD (Chae et al., 2024).
Emerging evidence has also highlighted the gut’s influence on systemic diseases such as obesity and type 2 diabetes (T2D). Obesity is associated with an increased Firmicutes to Bacteroides/Prevotella ratio, which elevates microbial genes involved in polysaccharide breakdown and raises SCFA production. This enhanced energy extraction from food contributes to weight gain. Additionally, microbial digestion of dietary choline and carnitine, common in Western diets, produces trimethylamine-N-oxide (TMAO), a compound linked to increased cardiovascular disease (CVD) risk. In T2D, dysbiosis results in a proliferation of Proteobacteria, Bacteroidetes, and Firmicutes, disrupting glucose metabolism and contributing to insulin resistance (Muscogiuri et al., 2017).
The gut microbiota’s role extends beyond metabolic and inflammatory disorders to include infectious diseases such as COVID-19. Studies suggest a strong gut-lung axis, wherein the gut microbiota upregulates the synthesis of ACE2, a receptor used by the SARS-CoV-2 virus to enter cells (Wang et al., 2023). Nutraceuticals, including polyphenols and probiotics, have been proposed to support immune responses and potentially mitigate disease severity.
Neurodegenerative diseases, such as Alzheimer’s and Parkinson’s, have also been linked to dysbiosis. Alterations in gut microbiota composition affect the gut-brain axis through microbial metabolites like SCFAs and tryptophan derivatives, influencing neuroinflammation and neurotransmitter synthesis (Junyi et al., 2024). For example, elevated levels of lipopolysaccharides (LPS) in dysbiotic states can disrupt the blood–brain barrier and activate microglia, exacerbating neurodegeneration.
The gut-brain axis represents a dynamic bidirectional communication network linking the gut microbiota and the central nervous system (CNS). This intricate system operates through multiple mechanisms, including microbial metabolites such as short-chain fatty acids (SCFAs), immune signaling, the vagus nerve, and neuroendocrine pathways via the hypothalamic–pituitary–adrenal (HPA) axis. Recent studies underscore the significant role of this axis in influencing brain function, mood regulation, and the pathophysiology of neurodegenerative diseases (Schneider et al., 2024).
Neurodegenerative diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) have been closely linked to gut dysbiosis, a condition characterized by imbalances in microbial composition (Schneider et al., 2024; Kulkarni et al., 2024; Caradonna et al., 2024). In Alzheimer’s disease, gut dysbiosis contributes to increased systemic inflammation through the release of microbial metabolites like lipopolysaccharides (LPS), which compromise the blood–brain barrier integrity. This allows amyloid-beta peptides to accumulate, exacerbating neuroinflammation and neuronal damage (Kulkarni et al., 2024; Caradonna et al., 2024). Similarly, Parkinson’s disease is associated with alterations in gut microbiota composition, particularly a reduction in anti-inflammatory taxa such as Akkermansia muciniphila. These changes are thought to facilitate the misfolding and aggregation of α-synuclein, a hallmark of PD pathology, and promote its transmission from the enteric nervous system to the CNS via the vagus nerve (Kulkarni et al., 2024; Caradonna et al., 2024).
Microbial metabolites play a pivotal role in modulating neuroinflammatory pathways. SCFAs, including butyrate and propionate, have been shown to regulate microglial activation, which is critical for maintaining CNS homeostasis. A reduction in SCFA levels, often observed in dysbiotic states, is correlated with increased neuroinflammation and cognitive decline (Fodor et al., 2023). Moreover, tryptophan metabolites derived from gut bacteria influence serotonin and kynurenine pathways, both of which are integral to mood regulation and cognitive processes. For instance, studies have highlighted the role of Bacteroides species in promoting the synthesis of indole derivatives, which have neuroprotective effects (Fodor et al., 2023).
Akkermansia muciniphila is an anaerobic gram-negative anti-obesity bacterium that grows singly or in pairs in mucin-rich medium (Liu et al., 2022). Akkermansia muciniphila has garnered significant attention due to its ability to degrade mucus that considers a protective layer that lines the gut and maintains a healthy balance that serves as a barrier against harmful microorganisms as excessive mucus production can lead to a range of health issues, including inflammatory bowel disease (IBD) and obesity (Geerlings et al., 2018; Kim et al., 2021). Mucin layer is primarily made up of gelatinous mucins secreted by goblet cells and also consist of a peptide backbone modified by O-linked glycans. The presence of proline and threonine in the peptide backbone contributes to the unique properties of mucin, including its gel-like consistency and protective function (Pelaseyed et al., 2014). Further, the production of butyrate, a short-chain fatty acid by Akkermansia muciniphila not only provides an energy source for the cells lining the colon but also contributes to reducing inflammation and improving gut barrier function (Portincasa et al., 2022). Another metabolite produced by Akkermansia muciniphila is propionate, which also has anti-inflammatory effects and regulate glucose metabolism. It also stimulates the production of molecules that promote the growth of beneficial bacteria in the gut, leading to a more diverse and balanced gut microbiota (Rodrigues et al., 2022).
Adlercreutzia equolifaciens is a Gram-positive bacterium that thrives in an anaerobic environment, slightly acidic pH range of 6.0 to 6.5 by using culture medium, such as Reinforced Clostridial Medium (RCM). This bacterium is of particular interest due to its ability to produce equol, a metabolite of the soy isoflavone daidzein, which is found in soy products and other legumes. Research has shown that equol has estrogenic and antioxidant properties, which may help alleviate symptoms associated with hormonal imbalances, such as menopause, hot flashes and night sweats (Mayo et al., 2019). Further equol have anti-inflammatory and anti-cancer effects. Individuals who consume a diet rich in soy products are more likely to have a higher abundance of Adlercreutzia equolifaciens in their gut (Tuli et al., 2022).
Barnesiella is an obligate anaerobic bacterium that can be cultivated under both anaerobic and microaerophilic conditions at a temperature range of 37–42°C in nutrient-rich media, such as brain-heart infusion (BHI) or tryptic soy broth (TSB), supplemented with appropriate carbon and nitrogen sources (Ubeda et al., 2013). Barnesiella is a commensal bacterium that found reasonably low level in the gastrointestinal tract of humans help in the breakdown of complex carbohydrates, production of SCFAs, modulation of immune responses, facilitate the clearance of intestinal VRE colonization and prevent the spread of highly antibiotic-resistant bacteria (Tao et al., 2021; Ubeda et al., 2013; Aindelis and Chlichlia, 2020).
Christensenella minuta is an obligate anaerobe that could be grow optimally under mesophilic conditions, between 30°C to 37°C by using a suitable culture medium, such as a nutrient-rich broth, to support its growth (Kropp et al., 2021; Ang et al., 2023). One of the unique metabolic features of Christensenella minuta is its ability to produce butyrate, a short-chain fatty acid linked with various health benefits, including anti-inflammatory and anti-cancer properties. Studies suggest that individuals with higher levels of Christensenella minuta may have a lower risk of type 2 diabetes, inflammatory bowel disease and obesity which sparked interest in the use in the development of probiotics and prebiotics (Mazier et al., 2021; Ang et al., 2023). In addition, this bacterium can influence the host’s response to certain medications, such as antidiabetic drugs and suggested that it may contribute to inter-individual differences in drug response and could potentially be targeted for personalized medicine approaches.
Oxalobacter formigenes is a unique bacterium that can be grow in anaerobic environment by using Oxalate-Minimal Medium, which consists of oxalate, mineral salts, vitamins and buffer system (Duncan et al., 2002; Daniel et al., 2021). This bacterium resides in the human gastrointestinal tract known for its unique ability to break down and metabolize oxalate, a compound that can form kidney stones in some individuals. Studies have shown that individuals with a higher abundance of Oxalobacter formigenes in their gut have a lower risk of developing kidney stones (Chmiel et al., 2022). This bacterium produces an enzyme called oxalyl-CoA decarboxylase, which converts oxalate into a less harmful compound called formate (Karamad et al., 2022).
Research has consistently shown that our dietary choices have a direct impact on the health and resilience of our gut microbiome (Singh et al., 2017; Su and Liu, 2021). The incorporation of prebiotic rich foods, fermented foods, and a wide variety of plants and fruits promote the thriving of these gut microbiome as detailed in Table 1 (Nambiar et al., 2023; Leeuwendaal et al., 2022). The colorful plant foods are not only visually appealing, but they also provide a wide range of health benefits for the microbiome and metabolism. One of the key benefits of colorful foods is their high content of phytochemicals bioactive compounds such as carotenoids, flavonoids, and anthocyanins that have been shown to have numerous health-promoting effects (Samtiya et al., 2021; Khoo et al., 2017).
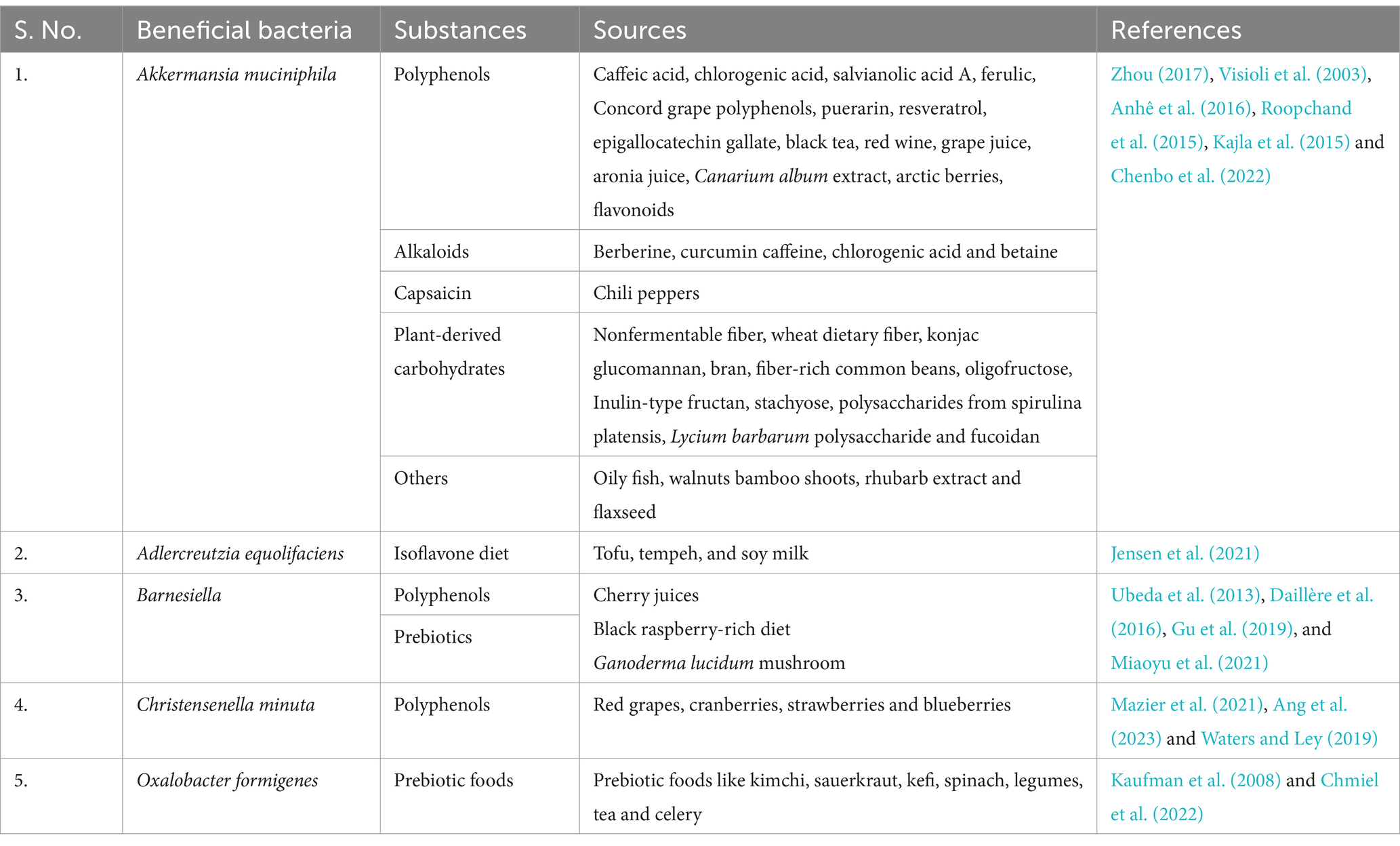
Table 1. Various dietary sources that produce and nourish good gut bacteria.
The microbiome is a living dynamic environment where the relative abundance of species may fluctuate daily, weekly, and monthly depending on diet, medication, exercise, and a host of other environmental exposures (Monda et al., 2017; York, 2019).
Gut microbiota breaks down the carotenoid’s rich foods, into various beneficial gut metabolites such as apocarotenoids and SCFAs having anti-inflammatory, antioxidant, and anticancer properties that contribute to decrease the risk of the development of chronic diseases, cardiovascular diseases, type 2 diabetes, obesity, brain-related diseases and certain types of cancer (Rowles and Erdman, 2020; Rocha et al., 2023; Eroglu et al., 2023; Min et al., 2023). They also act as antioxidants, protecting the body’s cells from damage caused by harmful free radicals. Their ability to protect against lipid peroxidation and damage caused by ROS makes them valuable in maintaining overall health and reducing the risk of chronic diseases (Lobo et al., 2010; Rocha et al., 2023).
Apo-carotenoids exhibit unique characteristics, including higher aqueous solubility and higher electrophilicity, which make them particularly suitable for targeting transcription factors such as NF-κB, PPARγ, and RAR/RXRs (Eroglu et al., 2023). These compounds hold potential for therapeutic applications in the fields of inflammation, metabolic disorders, and cell differentiation. Studies have found that carotenoids can directly influence the composition of the gut microbiota in a positive manner (Rocha et al., 2023). Beta-carotene such as carrots and sweet potatoes can increase the abundance of certain beneficial bacteria, such as Bifidobacteria and Lactobacillus (Rinninella et al., 2019; Zhiguo et al., 2023; Li et al., 2022a). Lycopene, found in tomatoes, has been shown to increase the levels of bacteria that produce short-chain fatty acids, which are beneficial for gut health (Eroglu et al., 2023). A. Lutein, found in leafy greens, has been associated with a more diverse gut microbiota (Dinsmoor et al., 2019).
Flavonoid-rich foods including fruits like berries, citrus fruits, vegetables, dark chocolate and tea, can lead to an increase in the diversity and abundance of gut metabolites that break down the flavonoids through fermentation processes into various metabolites compounds that have anti-inflammatory and antioxidant properties and reduce the risk of metabolic disorders such as cardiovascular disease by improving the insulin sensitivity (Joaquim et al., 2023; Wang et al., 2022; Pan et al., 2023). One important group of flavonoids known as flavan-3-ols has been shown to increase the levels of short-chain fatty acids (SCFAs) such as butyrate, which provide energy for the gut epithelial cells and have anti-inflammatory effects (Fotschki et al., 2015). Another bioactive compound flavanols associated with increase the production of urolithins metabolites in the gut that have been shown to have anti-inflammatory, antioxidant, and anticancer properties (Singh et al., 2019). Further flavonoids have also been found to increase the production phenolic acids, benzoic acid derivatives, and microbial-derived compounds that can influence various physiological processes in the body (Rahman et al., 2021). Previous research reported that flavonoids have the ability to modulate the relative abundance by increasing the relative abundance of Bifidobacterium, Lactobacillus, while decreasing the relative abundance of Lachnoclostridium and Bilophila, highlights their potential as dietary supplements or functional food ingredients to promote a healthy gut microbiota (Baky et al., 2022; Pan et al., 2023).
Anthocyanins that are responsible for bright red, purple, and blue colors to fruits like blueberries, blackberries, purple cabbage and cherries have been shown to have anti-inflammatory and antioxidant effects, as well as potential anti-cancer properties (Khoo et al., 2017; Bahare et al., 2020). They may also help regulate blood sugar levels and improve insulin sensitivity (Fernandes, 2019). One metabolite that has been extensively studied in relation to Anthocyanins increase nitric oxide (NO) metabolite known as signaling molecule that helps in the regulation of blood flow, neurotransmission, and immune responses Geum et al. (2020). Increased levels of nitric oxide relax and dilate blood vessels, leading to improved blood flow and reduced risk of hypertension, heart disease and diabetes (da Silva et al., 2021; Kumar et al., 2022). Studies have consistently shown that anthocyanin leads to an increase in the presence of beneficial bacteria such as Lactobacillus, Bifidobacterium, Blautia, Faecalibacterium, Prevotella, Akkermansia and stimulate the production SCFAs particularly butyrate (Verediano et al., 2021; Zhong et al., 2023; Liang et al., 2023).
Polyphenols are a group of compounds that are naturally found in many plant-based foods and beverage that undergo various transformations by the gut bacteria and converted into a wide range of metabolites having antioxidant and anti-inflammatory properties (Gizem et al., 2020; Bertelli et al., 2021). Studies have shown that certain polyphenols, such as those found in green tea, berries, and cocoa, can increase the production of short-chain fatty acids (SCFAs) and branched-chain amino acids (BCAAs) and could be useful in the treatment and prevention of various gastrointestinal disorders (Manach et al., 2004; Kumar Singh et al., 2019; Lippolis et al., 2023). Furthermore, polyphenols have been shown to modulate the composition of the gut microbiota, promoting the growth of beneficial bacteria like Lactobacillus, Lactiplantibacillus and Bifidobacterium while hindering the proliferation of pathogenic strains like Clostridium and Fusobacterium (Corrêa et al., 2019; Wang et al., 2022; Lippolis et al., 2023; Rahman et al., 2021).
Alkaloids have been known to possess various biological activities and have been used in traditional medicine for centuries (Heinrich et al., 2021). Studies have shown that certain alkaloids promote the growth of beneficial bacteria that leads to an increase in the production of short-chain fatty acids, such as butyrate, which have numerous health benefits (Feng et al., 2018; Dehau et al., 2023). Furthermore, alkaloids found in coffee, such as caffeine and chlorogenic acid, have been shown to increase the production of certain bile acids in the gut (Iriondo-DeHond et al., 2020; Chen et al., 2023). Bile acids are important for the digestion and absorption of dietary fats and also have regulatory roles in lipid and glucose metabolism (González-Regueiro et al., 2017). Research has shown that berberine has antimicrobial properties, specifically targeting harmful bacteria like Escherichia coli (E. coli) and Clostridium difficile (C. difficile), while promoting the growth of beneficial bacteria like Bifidobacterium and Lactobacillus (Cheng et al., 2022; Zhang et al., 2021; Peng et al., 2019).
Research has shown that capsaicin can stimulate the production of certain gut metabolites such as SCFAs known to have various beneficial effects, such as reducing inflammation and improving insulin sensitivity (Song et al., 2017; Kang et al., 2017). Furthermore, capsaicin has been shown to enhance the activity of certain enzymes such as enzyme lipase, which is responsible for breaking down dietary fat (Liu et al., 2021). This increased enzyme activity can lead to a more efficient digestion and utilization of nutrients, ultimately affecting the production of gut metabolites (Menden et al., 2022; Chandra et al., 2020). In addition to its direct effects on gut metabolites, capsaicin has also been found to influence the composition of the gut microbiota and increase the abundance of Akkermansia muciniphila (Li et al., 2022b).
The composition and functionality of the gut microbiota are profoundly influenced by dietary choices, making nutrition a cornerstone for maintaining microbial health. Specific dietary strategies, including the incorporation of fermented foods, synbiotic combinations, and anti-inflammatory diets, have been shown to promote microbial diversity, enhance the production of beneficial metabolites like short-chain fatty acids (SCFAs), and reduce inflammation. These approaches not only support digestive health but also play a critical role in systemic immunity and metabolic regulation. By tailoring dietary interventions to the unique needs of individuals, it is possible to foster a balanced gut microbiota that contributes to overall well-being and resilience against chronic diseases.
Prebiotics can be found in a variety of foods, including fruits, vegetables, whole grains, and legumes. Some common examples of prebiotic fibers include inulin, fructooligosaccharides (FOS), and galactooligosaccharides (GOS) (Lockyer and Stanner, 2019). The primary benefits of prebiotics are their ability to selectively stimulate the growth of beneficial bacteria, such as Bifidobacteria, Lactobacilli and enhance the production of beneficial metabolites like short-chain fatty acids (Markowiak and Śliżewska, 2017; Davani-Davari et al., 2019). Prebiotic rich foods such as Chicory root and Jerusalem Artichokes are excellent sources of inulin, a fiber that serves as a fuel for the beneficial gut bacteria (Carlson et al., 2018). Similarly, the content of pectin in apples and resistant starch in bananas acts as nourishment for good bacteria (Englyst and Cummings, 1986; Leonel and Alvarez-Leite, 2012).
Other prebiotic foods like oats, barley, and quinoa promote healthy gut bacteria by providing good source of prebiotic fibers (Slavin, 2013). Legumes foods, including lentils, chickpeas, and beans, are known for their high nutritional value and are an excellent source of prebiotics that promote the growth of beneficial bacteria in the gut (Kadyan et al., 2022). The content of fructooligosaccharides (FOS) in garlic and onions, and inulin and oligofructose in asparagus vegetables act as prebiotics by stimulating the growth of beneficial gut bacteria (Zhang et al., 2013; Guillamón et al., 2021).
Fermented foods are a rich source of probiotics that enhance gut health by introducing beneficial bacteria and improving microbial diversity. Popular examples include yogurt, kimchi, and sauerkraut. These foods support gut integrity by increasing levels of Lactobacillus and Bifidobacterium, which produce metabolites like short-chain fatty acids (SCFAs) that reduce inflammation and strengthen the gut barrier (Dimidi et al., 2019). In addition to these widely studied examples, global fermented foods such as miso and natto from Japanese cuisine, kefir from Eastern Europe, and dosa from India provide unique probiotic strains and bioactive compounds. Miso and natto, for instance, contain Bacillus subtilis, which has been shown to promote immune regulation and reduce markers of systemic inflammation (Oshiro et al., 2021). Kefir is particularly rich in lactic acid bacteria and yeast, offering a broad spectrum of probiotics with antimicrobial and gut-stabilizing properties (Tingirikari et al., 2024).
Fermented foods provide a natural and delicious way to support a healthy gut microbiome. With their probiotic properties, they offer numerous benefits, including improved digestive health, enhanced immune function, increased nutrient availability, and potential mental well-being (Leeuwendaal et al., 2022). Yogurt is a widely consumed fermented dairy product that augments beneficial bacteria, such as Lactobacillus and Bifidobacterium (Lisko et al., 2017). Kimchi is mainly produced by fermented vegetables, including cabbage, radishes, and garlic that contain Lactobacillus bacteria, which are known to promote gut health and improve digestion (Dimidi et al., 2019).
Another fermented food known as Kombucha made from a fermented tea beverage that is rich in bioactive compounds and a variety of beneficial acetic acid bacteria and yeasts, contributing to a healthy gut microbiome (Kitwetcharoen et al., 2023). A previous study conducted on rats revealed that tempeh, a fermented soybean product, has the potential to enhance the production of immunoglobulin A (IgA) and modulate the composition of gut microbiota. In addition to the study on rats, research involving supplementation of tempeh in humans for 16 days led to a significant increase in the abundance of beneficial gut bacteria like Akkermansia muciniphila (Stephanie et al., 2017).
Practical dietary recommendations include incorporating a variety of fermented foods into daily meals. For example, pairing kimchi with rice or adding yogurt to smoothies can provide both flavor and health benefits. Future research should explore the synergistic interactions between probiotics in fermented foods and the resident gut microbiota to optimize therapeutic applications.
Synbiotic foods provide a convenient and effective way to support a healthy gut microbiome (Jiang et al., 2022). By combining prebiotics and probiotics, these foods offer numerous health benefits, including improved digestion, enhanced immune function, increased nutrient absorption, and reduced inflammation (Yadav et al., 2022). Incorporating synbiotic foods into a balanced diet can be a valuable strategy for promoting overall health and well-being (Pandey et al., 2015). Synbiotic food having a combination of arabinose, lactulose and Lactobacillus plantarum, have gained attention for their potential to effectively regulate blood glucose, blood lipid, and body weight in patients with Type 2 Diabetes Mellitus (T2DM) (Jiang et al., 2022). Another preparation known as a banana smoothie made with kefir or yogurt is a delicious and nutritious beverage that maximizes the benefits for gut bacteria. Stir-fry made with tempeh, asparagus, garlic, and leeks prompts, you can actively healthy gut microbiome. The combination of yogurt and fruits such as blueberries creates a synbiotic effect, where the probiotics from yogurt and the prebiotics from blueberries work together to promote the growth and activity of healthy gut bacteria (Fernandez and Marette, 2017).
Anti-inflammatory foods such as fatty fish, fruits, vegetables, whole grains, and spices have long been recognized for their potential health benefits by supporting a diverse and balanced gut microbiome, reducing inflammation and promoting overall gut health (Bagheri et al., 2022). Fish like salmon, sardines, and anchovies in diet can have a positive impact on gut health. Their omega-3 fatty acids and ability to increase healthy gut bacteria make them a valuable addition to an anti-inflammatory diet (Costantini et al., 2017). Flax seeds are rich in dietary fiber, including both soluble and insoluble fiber that act as prebiotics and provide nourishment for the beneficial bacteria in the gut (Kajla et al., 2015; Mueed et al., 2022). When the gut bacteria ferment the fiber from flax seeds, they produce short-chain fatty acids (SCFAs), such as butyrate that help in maintaining a healthy gut environment (Arora et al., 2019).
Diet plays a pivotal role in shaping the composition, diversity, and metabolic activity of the gut microbiota. Long-term dietary patterns, including plant-based and high-protein diets, create distinct microbial environments that influence host health. Microbiota-accessible carbohydrates (MACs), a subset of dietary fibers, serve as the primary energy source for gut bacteria, fostering the growth of beneficial taxa and stimulating the production of short-chain fatty acids (SCFAs) (De Filippo et al., 2010). MACs are composed of complex polysaccharides, such as resistant starches, inulin, and pectin, that are indigestible by host enzymes but fermentable by gut microbes. These carbohydrates selectively enhance the abundance of health-promoting bacteria like Bifidobacterium and Faecalibacterium prausnitzii, both of which are associated with anti-inflammatory effects and improved gut barrier integrity (Hills et al., 2019). For example, resistant starch found in legumes and whole grains increases butyrate production, which supports colonic health. Similarly, inulin from chicory root and Jerusalem artichokes promotes the proliferation of Bifidobacterium, contributing to gut homeostasis and reduced systemic inflammation. These carbohydrates are made up of monosaccharides linked together by various forms of glycosidic connections. Any major differences in their chemical makeup, solubility, and size classify these carbohydrates into a wide range of biological niches. Dietary fibers are crucial sources of energy for the bacteria that live in the colon and cecum. In addition to reducing microbial diversity and short chain fatty acid (SCFA) production, a low intake of dietary fiber causes the gut microbial metabolism to shift toward the use of less advantageous substrates. The mucus barrier is harmed by a protracted absence of dietary fibers, which is also linked to an increase in the number of bacteria that break down mucins, like Akkermansia muciniphila (Makki et al., 2018).
As a result, a lack of dietary fibre and an increase in sugar and fat in human diets may lead to the extinction of particular bacterial species. These changes may lead to dysfunctions, which could exacerbate existing conditions like IBD, allergies, colorectal cancer, autoimmune illnesses, obesity, etc. A diet high in fiber helps to maintain a healthy gut microbiota that is more diverse and performs activities like producing short-chain fatty acids (SCFAs) (Figure 3). Low fiber intake, a diet high in protein and sugar, and an industrialized diet all contribute to altered gut bacterial function, including a significant decrease in their capacity to produce SCFAs, which is linked to the emergence of chronic inflammatory diseases (Makki et al., 2018).

Figure 3. Effect of high and low fiber diet on gut microbiota.
Advancements in microbiome research have led to the development of innovative therapies aimed at modulating gut microbiota to improve health outcomes, including CNS-related disorders. Microbiome-targeted therapies encompass probiotics, prebiotics, postbiotics, dietary interventions, and fecal microbiota transplantation (FMT), each offering unique mechanisms to restore microbial balance and enhance host health (Loh et al., 2024).
Probiotics, defined as live microorganisms that confer health benefits when consumed in adequate amounts, have garnered attention for their potential to influence the gut-brain axis. Specific strains, such as Lactobacillus rhamnosus and Bifidobacterium longum, have shown promise in alleviating symptoms of anxiety and depression by modulating the HPA axis and reducing systemic inflammation (Schächtle and Rosshart, 2021). Moreover, these probiotics enhance the production of neuroactive compounds like gamma-aminobutyric acid (GABA), which exerts calming effects on the CNS (Schächtle and Rosshart, 2021; Loh et al., 2024).
Prebiotics, including dietary fibers and oligosaccharides, serve as substrates for beneficial gut bacteria, stimulating the production of SCFAs and other bioactive metabolites. For example, inulin and fructooligosaccharides have been shown to increase butyrate levels, which play a critical role in reducing neuroinflammation and preserving neuronal integrity (Anand et al., 2025). Prebiotic interventions also hold potential for personalized nutrition, wherein diets are tailored to enhance specific microbial taxa associated with improved CNS health.
Fecal microbiota transplantation (FMT), a procedure involving the transfer of stool from a healthy donor to a recipient, has emerged as a promising therapeutic avenue for neuropsychiatric and neurodegenerative disorders. Preliminary clinical trials suggest that FMT can restore microbial diversity and ameliorate symptoms in conditions such as autism spectrum disorder and depression (Zhang et al., 2023; Peery et al., 2024). However, the standardization of FMT protocols and the identification of specific microbial communities responsible for therapeutic effects remain critical challenges (Zhang et al., 2023).
Future research should focus on the development of “designer probiotics” tailored to target specific diseases and the integration of microbiome data with genomic and metabolomic profiles to advance precision medicine. Additionally, leveraging computational tools to model host-microbiota interactions could facilitate the identification of novel therapeutic targets, paving the way for next-generation microbiome-based treatments.
The study of the human gut microbiota has advanced significantly in recent years, revealing its profound influence on the host’s metabolism, physiology, and immune system. Numerous factors, including nutrition, host genetics, age, medications, and lifestyle, shape the composition and functionality of the gut microbiota. These dynamic interactions play a pivotal role in human health, and alterations in gut microbial composition are now recognized as key contributors to the development of various diseases. A deeper understanding of the interplay between diet and microbiota holds the potential to develop tailored nutritional strategies aimed at reducing the prevalence of chronic inflammatory disorders.
Emerging evidence highlights the critical role of plant-based foods rich in phytochemicals and other specific dietary components in promoting the growth and maintenance of beneficial gut bacteria. However, challenges such as absorption, bioavailability, dietary interactions, and individual variations in metabolism can limit the efficacy of phytochemicals in fostering a healthy gut microbial community. The highly dynamic nature of gut microbiota, influenced by environmental factors, further complicates these interventions. Addressing issues such as standardization of dietary recommendations, variability in individual responses, and the long-term maintenance of dietary changes is essential for optimizing the benefits of specific foods on gut health.
In this review, we explored the intricate roles of beneficial gut bacteria and the significant impact of optimal diets on human health. Our findings underscore the transformative potential of personalized nutrition, customized to individual microbiota profiles, in revolutionizing healthcare. By leveraging the symbiotic relationship between gut bacteria and dietary patterns, targeted dietary interventions and preventive strategies can be developed to address chronic diseases. However, implementing such personalized recommendations on a large scale requires a paradigm shift in healthcare systems, moving towards individualized care. Ethical concerns, particularly related to privacy and the use of data in precision medicine, also present formidable challenges that must be carefully navigated.
Advancements in microbiome-targeted therapies, including probiotics, prebiotics, and synbiotics, along with the integration of artificial intelligence (AI), offer promising avenues for tailoring dietary interventions. AI-driven models can integrate microbiome, metabolome, and genomic data to generate precise nutritional recommendations, optimizing dietary strategies at an individual level. Future research should prioritize understanding the mechanisms underlying host-microbiota interactions, improving the bioavailability of key dietary components, and evaluating the long-term effects of dietary interventions on microbial ecology. Combining these scientific insights with robust ethical frameworks is essential to fully harness the potential of gut microbiota in promoting health and preventing disease. Through a multidisciplinary approach, we can unlock innovative solutions that bridge scientific discovery and practical implementation, advancing healthcare and improving lives.
SK: Conceptualization, Validation, Visualization, Writing – original draft, Writing – review & editing. RM: Validation, Visualization, Writing – original draft, Writing – review & editing. PG: Conceptualization, Validation, Writing – review & editing. ÉL: Validation, Visualization, Writing – review & editing. XL: Validation, Writing – review & editing, Investigation, Visualization. SA: Validation, Writing – review & editing. PP: Validation, Writing – review & editing, Formal analysis, Visualization. C-MC: Visualization, Writing – review & editing, Investigation, Supervision, Writing – original draft. VR: Investigation, Visualization, Writing – review & editing, Conceptualization, Validation. RP: Conceptualization, Investigation, Validation, Visualization, Writing – review & editing, Supervision, Writing – original draft.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by VtR Inc-CGU (SCRPD1L0221), DOXABIO-CGU (SCRPD1K0131), and a CGU grant (UZRPD1L0011, UZRPD1M0081).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Afzaal, M., Saeed, F., Shah, Y. A., Hussain, M., Rabail, R., Socol, C. T., et al. (2022). Human gut microbiota in health and disease: unveiling the relationship. Front. Microbiol. 13:99900. doi: 10.3389/fmicb.2022.99900
Aindelis, G., and Chlichlia, K. (2020). Modulation of anti-tumour immune responses by probiotic Bacteria. Vaccine 8:329. doi: 10.3390/vaccines8020329
Anand, A., Manjula, S. N., Fuloria, N. K., Sharma, H., and Mruthunjaya, K. (2025). Inulin as a prebiotic and its effect on gut microbiota. In inulin as a prebiotic and its effect on gut microbiota. pp. 113–135. doi: 10.1007/978-981-97-9056-2_6
Ang, W. S., Law, J. W., Letchumanan, V., Hong, K. W., Wong, S. H., Ab Mutalib, N. S., et al. (2023). A keystone gut bacterium Christensenella minuta-a potential biotherapeutic agent for obesity and associated metabolic diseases. Foods 12:2485. doi: 10.3390/foods12132485
Anhê, F. F., Pilon, G., Roy, D., Desjardins, Y., Levy, E., and Marette, A. (2016). Triggering Akkermansia with dietary polyphenols: a new weapon to combat the metabolic syndrome? Gut Microbes 7, 146–153. doi: 10.1080/19490976.2016.1142036
Arora, T., Rudenko, O., Egerod, K. L., Husted, A. S., Kovatcheva-Datchary, P., Akrami, R., et al. (2019). Microbial fermentation of flaxseed fibers modulates the transcriptome of GPR41-expressing enteroendocrine cells and protects mice against diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 316, E453–E463. doi: 10.1152/ajpendo.00391.2018
Bagheri, S., Zolghadri, S., and Stanek, A. (2022). Beneficial effects of anti-inflammatory diet in modulating gut microbiota and controlling obesity. Nutrients 14:3985. doi: 10.3390/nu14193985
Bahare, S., Javad, S., Francesca, C., Reiner, Ž., Zorzan, D., Imran, M., et al. (2020). The therapeutic potential of Anthocyanins: current approaches based on their molecular mechanism of action. Front. Pharmacol. 11:1300. doi: 10.3389/fphar.2020.01300
Baky, M. H., Elshahed, M., Wessjohann, L., and Farag, M. A. (2022). Interactions between dietary flavonoids and the gut microbiome: a comprehensive review. Br. J. Nutr. 128, 577–591. doi: 10.1017/S0007114521003627
Baxter, N. T., Schmidt, A. W., Venkataraman, A., Kim, K. S., Waldron, C., and Schmidt, T. M. (2019). Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. mBio 10:10.1128/mbio.02566-18. doi: 10.1128/mbio.02566-18
Bertelli, A., Biagi, M., Corsini, M., Baini, G., Cappellucci, G., and Miraldi, E. (2021). Polyphenols: from theory to practice. Foods 10:2595. doi: 10.3390/foods10112595
Bull, M. J., and Plummer, N. T. (2015). Part 2: Treatments for Chronic Gastrointestinal Disease and Gut Dysbiosis. Integr. Med. 14, 25–33.
Cano, R., Bermúdez, V., Galban, N., Garrido, B., Santeliz, R., Gotera, M. P., et al. (2024). Dietary polyphenols and gut microbiota Cross-talk: molecular and therapeutic perspectives for Cardiometabolic disease: a narrative review. Int. J. Mol. Sci. 25:9118. doi: 10.3390/ijms25169118
Caradonna, E., Nemni, R., Bifone, A., Gandolfo, P., Costantino, L., Giordano, L., et al. (2024). The brain–gut axis, an important player in Alzheimer and Parkinson disease: a narrative review. J. Clin. Med. 13:4130. doi: 10.3390/jcm13144130
Carlson, J. L., Erickson, J. M., Lloyd, B. B., and Slavin, J. L. (2018). Health effects and sources of prebiotic dietary Fiber. Curr. Dev. Nutr. 2:nzy005:nzy005. doi: 10.1093/cdn/nzy005
Chae, Y., Lee, Y. R., Kim, Y., and Park, H. (2024). Diet-induced gut dysbiosis and leaky gut syndrome. J. Microbiol. Biotechnol. 34, 747–756. doi: 10.4014/jmb.2312.12031
Chandra, P., Enespa, S. R., and Arora, P. K. (2020). Microbial lipases and their industrial applications: a comprehensive review. Microb. Cell Fact. 19:169. doi: 10.1186/s12934-020-01428-8
Chen, L., Wang, X. J., Chen, J. X., Yang, J. C., Lin, L., Cai, X. B., et al. (2023). Caffeine ameliorates the metabolic syndrome in diet-induced obese mice through regulating the gut microbiota and serum metabolism. Diabetol. Metab. Syndr. 15:37. doi: 10.1186/s13098-023-00993-3
Chenbo, Y., Chuanqi, C., Jianxin, Z., Hao, Z., Wei, C., and Qixiao, Z. (2022). Dietary strategies to promote the abundance of intestinal Akkermansia muciniphila, a focus on the effect of plant extracts. J. Funct. Foods 93:105093. doi: 10.1016/j.jff.2022.10509
Cheng, H., Liu, J., Tan, Y., Feng, W., and Peng, C. (2022). Interactions between gut microbiota and berberine, a necessary procedure to understand the mechanisms of berberine. J. Pharm. Anal. 12, 541–555. doi: 10.1016/j.jpha.2021.10.003
Chmiel, J. A., Carr, C., Stuivenberg, G. A., Venema, R., Chanyi, R. M., Al, K. F., et al. (2022). New perspectives on an old grouping: the genomic and phenotypic variability of Oxalobacter formigenes and the implications for calcium oxalate stone prevention. Front. Microbiol. 13:1011102. doi: 10.3389/fmicb.2022.1011102
Corrêa, T. A. F., Rogero, M. M., Hassimotto, N. M. A., and Lajolo, F. M. (2019). The two-way polyphenols-microbiota interactions and their effects on obesity and related metabolic diseases. Front. Nutr. 6:188. doi: 10.3389/fnut.2019.00188
Costantini, L., Molinari, R., Farinon, B., and Merendino, N. (2017). Impact of Omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 18:2645. doi: 10.3390/ijms18122645
Da Silva, G. M., da Silva, M. C., Nascimento, D. V. G., Lima Silva, E. M., Gouvêa, F. F. F., de França Lopes, L. G., et al. (2021). Nitric oxide as a central molecule in hypertension: focus on the Vasorelaxant activity of new nitric oxide donors. Biology 10:1041. doi: 10.3390/biology10101041
Daillère, R., Vétizou, M., Waldschmitt, N., Yamazaki, T., Isnard, C., Poirier-Colame, V., et al. (2016). Enterococcus hirae and Barnesiella intestinihominis facilitate cyclophosphamide-induced therapeutic immunomodulatory effects. Immunity 45, 931–943. doi: 10.1016/j.immuni.2016.09.009
Daniel, S. L., Moradi, L., Paiste, H., Wood, K. D., Assimos, D. G., Holmes, R. P., et al. (2021). Forty years of Oxalobacter formigenes, a gutsy oxalate-degrading specialist. Appl. Environ. Microbiol. 87:e0054421. doi: 10.1128/AEM.00544-21
Davani-Davari, D., Negahdaripour, M., Karimzadeh, I., Seifan, M., Mohkam, M., Masoumi, S., et al. (2019). Prebiotics: definition, types, sources, mechanisms, and clinical applications. Food Secur. 8:92. doi: 10.3390/foods8030092
De Filippo, C., Cavalieri, D., Di Paola, M., Ramazzotti, M., Poullet, J. B., Massart, S., et al. (2010). Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 107, 14691–14696. doi: 10.1073/pnas.1005963107
Dehau, T., Cherlet, M., Croubels, S., van Immerseel, F., and Goossens, E. (2023). A high dose of dietary Berberine improves Gut Wall morphology, despite an expansion of Enterobacteriaceae and a reduction in beneficial microbiota in broiler chickens. mSystems 8:e0123922. doi: 10.1128/msystems.01239-22
Dimidi, E., Cox, S., Rossi, M., and Whelan, K. (2019). Fermented foods: definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 11:1806. doi: 10.3390/nu11081806
Dinsmoor, A., Thompson, S., Edwards, C., Burd, N., Khan, N., Erdman, J., et al. (2019). Associations between serum lutein and human gut microbiota (P02-004-19). Current developments. Nutrition 3:nzz029. doi: 10.1093/cdn/nzz029.P02-004-19
Duncan, S. H., Richardson, A. J., Kaul, P., Holmes, R. P., Allison, M. J., and Stewart, C. S. (2002). Oxalobacter formigenes and its potential role in human health. Appl. Environ. Microbiol. 68, 3841–3847. doi: 10.1128/AEM.68.8.3841-3847.2002
Englyst, H. N., and Cummings, J. H. (1986). Digestion of the carbohydrates of banana (Musa paradisiaca sapientum) in the human small intestine. Am. J. Clin. Nutr. 44, 42–50. doi: 10.1093/ajcn/44.1.42
Eroglu, A., Al'Abri, I. S., Kopec, R. E., Crook, N., and Bohn, T. (2023). Carotenoids and their health benefits as derived via their interactions with gut microbiota. Adv. Nutr. 14, 238–255. doi: 10.1016/j.advnut.2022.10.007
Feng, W., Ao, H., and Peng, C. (2018). Gut microbiota, short-Chain fatty acids, and herbal medicines. Front. Pharmacol. 9:1354. doi: 10.3389/fphar.2018.01354
Fernandes, I. (2019). “Anthocyanins: Nutrition and health” in Bioactive Molecules in Food. Reference Series in Phytochemistry. eds. J. M. Mérillon and K. Ramawat (Cham: Springer).
Fernandez, M. A., and Marette, A. (2017). Potential health benefits of combining yogurt and fruits based on their probiotic and prebiotic properties. Adv. Nutr. 8, 155S–164S. doi: 10.3945/an.115.011114
Fodor, A., Cozma, A., Sitar-Taut, A., Vulturar, R., Suharoschi, R., Rusu, A., et al. (2023). The potential of epigenetic compounds in treating diabetes. Amsterdam: Elsevier.
Fotschki, B., Juśkiewicz, J., Sójka, M., Jurgoński, A., and Zduńczyk, Z. (2015). Ellagitannins and Flavan-3-ols from raspberry pomace modulate Caecal fermentation processes and plasma lipid parameters in rats. Molecules 20, 22848–22862. doi: 10.3390/molecules201219878
Geerlings, S. Y., Kostopoulos, I., de Vos, W. M., and Belzer, C. (2018). Akkermansia muciniphila in the human gastrointestinal tract: when, where, and how? Microorganisms 6:75. doi: 10.3390/microorganisms6030075
Geum, H. L., et al. (2020). Anthocyanins attenuate endothelial dysfunction through regulation of uncoupling of nitric oxide synthase in aged rats. Aging Cell 19:e13279. doi: 10.1111/acel.13279
Gizem, C., Koen, V., Luigi, L., Rocchetti, G., Delmas, D., Daglia, M., et al. (2020). Interaction of dietary polyphenols and gut microbiota: microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 1, 109–133. doi: 10.1002/fft2.25
González-Regueiro, J. A., Moreno-Castañeda, L., Uribe, M., and Chávez-Tapia, N. C. (2017). The role of bile acids in glucose metabolism and their relation with diabetes. Ann. Hepatol. 16, S21–S26. doi: 10.5604/01.3001.0010.5672
Gu, J., Thomas-Ahner, J. M., Riedl, K. M., Bailey, M. T., Vodovotz, Y., Schwartz, S. J., et al. (2019). Dietary black raspberries impact the colonic microbiome and phytochemical metabolites in mice. Mol. Nutr. Food Res. 63:e1800636. doi: 10.1002/mnfr.201800636
Guillamón, E., Andreo-Martínez, P., Mut-Salud, N., Fonollá, J., and Baños, A. (2021). Beneficial effects of organosulfur compounds from Allium cepa on gut health: a systematic review. Foods 10:1680. doi: 10.3390/foods10081680
Heinrich, M., Mah, J., and Amirkia, V. (2021). Alkaloids used as medicines: structural Phytochemistry meets biodiversity-an update and forward look. Molecules 26:1836. doi: 10.3390/molecules26071836
Hills, R. D., Pontefract, B. A., Mishcon, H. R., Black, C. A., Sutton, S. C., and Theberge, C. R. (2019). Gut microbiome: profound implications for diet and disease. Nutrients 11:1613. doi: 10.3390/nu11071613
Hou, K., Wu, Z., Chen, X., Wang, J., Zhang, D., and Xiao, C. (2022). Microbiota in health and diseases. Signal Transduction and Targeted Therapy, 7. doi: 10.1038/s41392-022-00974-4
Iriondo-DeHond, A., Uranga, J. A., Del Castillo, M. D., and Abalo, R. (2020). Effects of coffee and its components on the gastrointestinal tract and the brain-gut Axis. Nutrients 13:88. doi: 10.3390/nu13010088
Jandhyala, S. M., Talukdar, R., Subramanyam, C., Vuyyuru, H., Sasikala, M., and Nageshwar Reddy, D. (2015). Role of the normal gut microbiota. World J. Gastroenterol. 21, 8787–8803. doi: 10.3748/wjg.v21.i29.8787
Jensen, S. N., Cady, N. M., Shahi, S. K., Peterson, S. R., Gupta, A., Gibson-Corley, K. N., et al. (2021). Isoflavone diet ameliorates experimental autoimmune encephalomyelitis through modulation of gut bacteria depleted in patients with multiple sclerosis. Sci. Adv. 7:eabd4595. doi: 10.1126/sciadv.abd4595
Jiang, H., Cai, M., Shen, B., Wang, Q., Zhang, T., and Zhou, X. (2022). Synbiotics and gut microbiota: new perspectives in the treatment of type 2 diabetes mellitus. Foods 11:2438. doi: 10.3390/foods11162438
Joaquim, B., Bruno, S., Pedro, C., and Maria, H. (2023). Polyphenols in health and disease: gut microbiota, bioaccessibility, and bioavailability. Compounds 3, 40–72. doi: 10.3390/compounds3010005
Junyi, L., Yueyang, W., Bin, L., Xiaohong, D., Wenhui, C., Ning, Z., et al. (2024). Gut microbiota mediates Neuroinflammation in Alzheimer’s disease: unraveling key factors and mechanistic insights. Mol. Neurobiol. 62, 3746–3763. doi: 10.1007/s12035-024-04513-w
Kadyan, S., Sharma, A., Arjmandi, B. H., Singh, P., and Nagpal, R. (2022). Prebiotic potential of dietary beans and pulses and their resistant starch for aging-associated gut and metabolic health. Nutrients 14:1726. doi: 10.3390/nu14091726
Kajla, P., Sharma, A., and Sood, D. R. (2015). Flaxseed-a potential functional food source. J. Food Sci. Technol. 52, 1857–1871. doi: 10.1007/s13197-014-1293-y
Kang, C., Wang, B., Kaliannan, K., Wang, X., Lang, H., Hui, S., et al. (2017). Gut microbiota mediates the protective effects of dietary capsaicin against chronic low-grade inflammation and associated obesity induced by high-fat diet. MBio 8, e00470–e00417. doi: 10.1128/mBio.00470-17
Karamad, D., Khosravi-Darani, K., Khaneghah, A. M., and Miller, A. W. (2022). Probiotic oxalate-degrading Bacteria: new insight of environmental variables and expression of the oxc and frc genes on oxalate degradation activity. Foods 11:2876. doi: 10.3390/foods11182876
Kaufman, D. W., Kelly, J. P., Curhan, G. C., Anderson, T. E., Dretler, S. P., Preminger, G. M., et al. (2008). Oxalobacter formigenes may reduce the risk of calcium oxalate kidney stones. J. Am. Soc. Nephrol. 19, 1197–1203. doi: 10.1681/ASN.2007101058
Khoo, H. E., Azlan, A., Tang, S. T., and Lim, S. M. (2017). Anthocyanidins and anthocyanins: colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 61:1361779. doi: 10.1080/16546628.2017.1361779
Kim, S., Shin, Y. C., Kim, T. Y., Kim, Y., Lee, Y. S., Lee, S. H., et al. (2021). Mucin degrader Akkermansia muciniphila accelerates intestinal stem cell-mediated epithelial development. Gut Microbes 13, 1–20. doi: 10.1080/19490976.2021.1892441
Kitwetcharoen, H., Phung, L. T., Klanrit, P., Thanonkeo, S., Tippayawat, P., Yamada, M., et al. (2023). Kombucha healthy drink—recent advances in production chemical composition and health benefits. Fermentation 11:48. doi: 10.3390/fermentation9010048
Kropp, C., Le Corf, K., Relizani, K., Tambosco, K., Martinez, C., Chain, F., et al. (2021). The keystone commensal bacterium Christensenella minuta DSM 22607 displays anti-inflammatory properties both in vitro and in vivo. Sci. Rep. 11:11494. doi: 10.1038/s41598-021-90885-1
Krumbeck, J. A., Rasmussen, H. E., Hutkins, R. W., Clarke, J., Shawron, K., Keshavarzian, A., et al. (2018). Probiotic Bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 6:121. doi: 10.1186/s40168-018-0494-4
Kulkarni, R., Kumari, S., Dhapola, R., Sharma, P., Singh, S. K., Medhi, B., et al. (2024). Association between the gut microbiota and Alzheimer’s disease: an update on signaling pathways and translational therapeutics. Mol. Neurobiol. doi: 10.1007/s12035-024-04545-2
Kumar, S., Perumal, N., Yadav, P. K., Pandey, R. P., Chang, C. M., and Raj, V. S. (2022). Amoxicillin impact on pathophysiology induced by short term high salt diet in mice. Sci. Rep. 12:19351. doi: 10.1038/s41598-022-21270-9
Kumar Singh, A., Cabral, C., Kumar, R., Ganguly, R., Kumar Rana, H., Gupta, A., et al. (2019). Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 11:2216. doi: 10.3390/nu11092216
Lavelle, A., and Sokol, H. (2020). Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 17, 223–237. doi: 10.1038/s41575-019-0258-z
Leeuwendaal, N. K., Stanton, C., O'Toole, P. W., and Beresford, T. P. (2022). Fermented foods, health and the gut microbiome. Nutrients 14:1527. doi: 10.3390/nu14071527
Leonel, A. J., and Alvarez-Leite, J. I. (2012). Butyrate: implications for intestinal function. Curr. Opin. Clin. Nutr. Metab. Care 15, 474–479. doi: 10.1097/MCO.0b013e32835665fa
Li, Z., Dai, Z., Shi, E., Wan, P., Chen, G., and Zhang, Z. (2022a). Study on the interaction between β-carotene and gut microflora using an in vitro fermentation model. Food Science and Human Wellness, 12, 1369–1378. doi: 10.1016/j.fshw.2022.10.030
Li, J., Liao, X., Yin, X., Deng, Z., Hu, G., Zhang, W., et al. (2022b). Gut microbiome and serum metabolome profiles of capsaicin with cognitive benefits in APP/PS1 mice. Nutrients 15:118. doi: 10.3390/nu15010118
Liang, A., Leonard, W., Beasley, J. T., Fang, Z., Zhang, P., and Ranadheera, C. S. (2023). Anthocyanins-gut microbiota-health axis: a review. Critical reviews in food science and nutrition, 1–26. Adv. Online Pub. 64, 7563–7588. doi: 10.1080/10408398.2023.2187212
Lippolis, T., Cofano, M., Caponio, G. R., De Nunzio, V., and Notarnicola, M. (2023). Bioaccessibility and bioavailability of diet polyphenols and their modulation of gut microbiota. Int. J. Mol. Sci. 24:3813. doi: 10.3390/ijms24043813
Lisko, D. J., Johnston, G. P., and Johnston, C. G. (2017). Effects of dietary yogurt on the healthy human gastrointestinal (GI) microbiome. Microorganisms 5:6. doi: 10.3390/microorganisms5010006
Liu, X., Shao, J., Liao, Y., Wang, L., Jia, Y., Dong, P., et al. (2023). Regulation of short-chain fatty acids in the immune system. Front. Immunol. 14:1186892. doi: 10.3389/fimmu.2023.1186892
Liu, S. J., Wang, J., He, T. F., Liu, H. S., and Piao, X. S. (2021). Effects of natural capsicum extract on growth performance, nutrient utilization, antioxidant status, immune function, and meat quality in broilers. Poult. Sci. 100:101301. doi: 10.1016/j.psj.2021.101301
Liu, M. J., Yang, J. Y., Yan, Z. H., Hu, S., Li, J. Q., Xu, Z. X., et al. (2022). Recent findings in Akkermansia muciniphila-regulated metabolism and its role in intestinal diseases. Clin. Nutr. 41, 2333–2344. doi: 10.1016/j.clnu.2022.08.029
Lobo, V., Patil, A., Phatak, A., and Chandra, N. (2010). Free radicals, antioxidants and functional foods: impact on human health. Pharmacogn. Rev. 4, 118–126. doi: 10.4103/0973-7847.70902
Lockyer, S., and Stanner, S. (2019). Prebiotics – an added benefit of some fibre types. Nutr. Bull. 44, 74–91. doi: 10.1111/nbu.12366
Loh, J. S., Mak, W. Q., Tan, L. K. S., Ng, C. X., Chan, H. H., Yeow, S. H., et al. (2024). Microbiota–gut–brain axis and its therapeutic applications in neurodegenerative diseases. Signal transduction and targeted. Therapy 9:743. doi: 10.1038/s41392-024-01743-1
Makki, K., Deehan, E. C., Walter, J., and Bäckhed, F. (2018). The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 23, 705–715. doi: 10.1016/j.chom.2018.05.012
Manach, C., Scalbert, A., Morand, C., Rémésy, C., and Jiménez, L. (2004). Polyphenols: food sources and bioavailability. Am. J. Clin. Nutr. 79, 727–747. doi: 10.1093/ajcn/79.5.727
Markowiak, P., and Śliżewska, K. (2017). Effects of probiotics, prebiotics, and Synbiotics on human health. Nutrients 9:1021. doi: 10.3390/nu9091021
Maruo, T., Sakamoto, M., Ito, C., Toda, T., and Benno, Y. (2008). Adlercreutzia equolifaciens gen. Nov., sp. nov., an equol-producing bacterium isolated from human faeces, and emended description of the genus Eggerthella. Int. J. Syst. Evol. Microbiol. 58, 1221–1227. doi: 10.1099/ijs.0.65404-0
Mayo, B., Vázquez, L., and Flórez, A. B. (2019). Equol: a bacterial metabolite from the Daidzein Isoflavone and its presumed beneficial health effects. Nutrients 11:2231. doi: 10.3390/nu11092231
Mazier, W., Le Corf, K., Martinez, C., Tudela, H., Kissi, D., Kropp, C., et al. (2021). A new strain of Christensenella minuta as a potential biotherapy for obesity and associated metabolic diseases. Cells 10:823. doi: 10.3390/cells10040823
Menden, A., Hall, D., Hahn-Townsend, C., Broedlow, C. A., Joshi, U., Pearson, A., et al. (2022). Exogenous lipase administration alters gut microbiota composition and ameliorates Alzheimer's disease-like pathology in APP/PS1 mice. Sci. Rep. 12:4797. doi: 10.1038/s41598-022-08840-7
Miaoyu, L., Leilei, Y., Jianxin, Z., Hao, Z., Wei, C., Qixiao, Z., et al. (2021). Role of dietary edible mushrooms in the modulation of gut microbiota. J. Funct. Foods 83:104538. doi: 10.1016/j.jff.2021.104538
Min, W., Renzhe, T., Rui, Z., Yongxiang, Q., and Dongmei, D. (2023). The protective effect of serum carotenoids on cardiovascular disease: a cross-sectional study from the general US adult population. Front. Nutr. 10:10. doi: 10.3389/fnut.2023.1154239
Monda, V., Villano, I., Messina, A., Valenzano, A., Esposito, T., Moscatelli, F., et al. (2017). Exercise modifies the gut microbiota with positive health effects. Oxidative Med. Cell. Longev. 2017:972. doi: 10.1155/2017/3831972
Mueed, A., Shibli, S., Korma, S. A., Madjirebaye, P., Esatbeyoglu, T., and Deng, Z. (2022). Flaxseed bioactive compounds: chemical composition, functional properties, food applications and health benefits-related gut microbes. Foods 11:3307. doi: 10.3390/foods11203307
Murga-Garrido, S. M., Hong, Q., Cross, T. L., Hutchison, E. R., Han, J., Thomas, S. P., et al. (2021). Gut microbiome variation modulates the effects of dietary fiber on host metabolism. Microbiome 9:117. doi: 10.1186/s40168-021-01061-6
Muscogiuri, G., Balercia, G., Barrea, L., Cignarelli, A., Giorgino, F., Holst, J. J., et al. (2017). Gut: a key player in the pathogenesis of type 2 diabetes? Crit. Rev. Food Sci. Nutr. 58, 1294–1309. doi: 10.1080/10408398.2016.1252712
Nambiar, R. B., Perumal, A. B., Shittu, T., Sadiku, E. R., and Sellamuthu, P. S. (2023). Editorial: probiotics, prebiotics, synbiotics, postbiotics, & paraprobiotics - new perspective for functional foods and nutraceuticals. Front. Nutr. 10:1164676. doi: 10.3389/fnut.2023.1164676
Oshiro, N., Wong, S. K., Chan, H. T., Chan, E. W. C., and Kezuka, M. (2021). Natto and miso: an overview on their preparation, bioactive components and health-promoting effects. Food Research (Malaysia).
Othman, L., Sleiman, A., and Abdel-Massih, R. M. (2019). Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 10:911. doi: 10.3389/fmicb.2019.00911
Pan, L., Ye, H., Pi, X., Liu, W., Wang, Z., Zhang, Y., et al. (2023). Effects of several flavonoids on human gut microbiota and its metabolism by in vitro simulated fermentation. Front. Microbiol. 14:1092729. doi: 10.3389/fmicb.2023.1092729
Pandey, K. R., Naik, S. R., and Vakil, B. V. (2015). Probiotics, prebiotics and synbiotics- a review. J. Food Sci. Technol. 52, 7577–7587. doi: 10.1007/s13197-015-1921-1
Parada Venegas, D., De la Fuente, M. K., Landskron, G., González, M. J., Quera, R., Dijkstra, G., et al. (2019). Short Chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10:277. doi: 10.3389/fimmu.2019.00277
Parekh, P. J., Balart, L. A., and Johnson, D. A. (2015). The influence of the gut microbiome on obesity, metabolic syndrome and gastrointestinal disease. Clin. Transl. Gastroenterol. 6:e91. doi: 10.1038/ctg.2015.16
Peery, A. F., Kelly, C. R., Kao, D., Vaughn, B. P., Lebwohl, B., and Altayar, O. (2024). AGA Clinical Practice Guideline on Fecal Microbiota–Based Therapies for select Gastrointestinal Diseases. Gastroenterol. 166, 409–434. doi: 10.1053/j.gastro.2024.01.008
Pelaseyed, T., Bergström, J. H., Gustafsson, J. K., Ermund, A., Birchenough, G. M., Schütte, A., et al. (2014). The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 260, 8–20. doi: 10.1111/imr.12182
Peng, J., Zheng, T. T., Li, X., Liang, Y., Wang, L. J., Huang, Y. C., et al. (2019). Plant-derived alkaloids: the promising disease-modifying agents for inflammatory bowel disease. Front. Pharmacol. 10:351. doi: 10.3389/fphar.2019.00351
Portincasa, P., Bonfrate, L., Vacca, M., De Angelis, M., Farella, I., Lanza, E., et al. (2022). Gut microbiota and short Chain fatty acids: implications in glucose homeostasis. Int. J. Mol. Sci. 23:1105. doi: 10.3390/ijms23031105
Queen, R., and Queen, R. (2020). Resistant starch, butyrate and gut barrier integrity resistant starch. Resistant starch research resistant starch research and benefits explained in simple terms. Available at: https://resistantstarchresearch.com/posts/resistant-starch-butyrate-and-gut-barrier-integrity/#:~:text=Resistant%20starch's%20production%20of%20butyrate,intestinal%20inflammation%20is%20also%20beneficial
Rahman, M. M., Rahaman, M. S., Islam, M. R., Rahman, F., and Mithi, F. M.,and Alqahtani, T. (2021). Role of phenolic compounds in human disease: Current knowledge and future Prospects. Molecules, 27:233. doi: 10.3390/molecules27010233
Rinninella, E., Cintoni, M., Raoul, P., Lopetuso, L. R., Scaldaferri, F., Pulcini, G., et al. (2019). Food components and dietary habits: keys for a healthy gut microbiota composition. Nutrients 11:2393. doi: 10.3390/nu11102393
Roberfroid, M., Bornet, F., Bouley, C., and Cummings, J. (2009). Colonic microflora: nutrition and Health0. Summary and conclusions of an international Life Sciences Institute (ILSI) [Europe] workshop held in Barcelona, Spain. Nutr. Rev. 53, 127–130. doi: 10.1111/j.1753-4887.1995.tb01535.x
Rocha, H. R., Coelho, M. C., Gomes, A. M., and Pintado, M. E. (2023). Carotenoids diet: digestion, gut microbiota modulation, and inflammatory diseases. Nutrients 15:2265. doi: 10.3390/nu15102265
Rodrigues, V. F., Elias-Oliveira, J., Pereira, Í. S., Pereira, J. A., Barbosa, S. C., Machado, M. S. G., et al. (2022). Akkermansia muciniphila and gut immune system: a good friendship that attenuates inflammatory bowel disease, obesity, and diabetes. Front. Immunol. 13:934695. doi: 10.3389/fimmu.2022.934695
Roopchand, D., Carmody, R. N., Kuhn, P., Moskal, K., Rojas-Silva, P., Turnbaugh, P. J., et al. (2015). Dietary polyphenols promote growth of the gut BacteriumAkkermansia muciniphilaand attenuate high-fat diet–induced metabolic syndrome. Diabetes 64, 2847–2858. doi: 10.2337/db14-1916
Rowles, J. L., and Erdman, J. W. (2020). Carotenoids and their role in cancer prevention. Biochimica et biophysica acta. Mol. Cell Biol. Lipids 1865:158613. doi: 10.1016/j.bbalip.2020.158613
Samtiya, M., Aluko, R. E., Dhewa, T., and Moreno-Rojas, J. M. (2021). Potential health benefits of plant food-derived bioactive components: an overview. Foods 10:839. doi: 10.3390/foods10040839
Schächtle, M. A., and Rosshart, S. P. (2021). The microbiota-gut-brain axis in health and disease and its implications for translational research. Front. Cell. Neurosci. 15:698172. doi: 10.3389/fncel.2021.698172
Schneider, E., O’Riordan, K. J., Clarke, G., and Cryan, J. F. (2024). Feeding gut microbes to nourish the brain: unravelling the diet–microbiota–gut–brain axis. Nat. Metab. 6, 1454–1478. doi: 10.1038/s42255-024-01108-6
Sejbuk, M., Mirończuk-Chodakowska, I., Karav, S., and Witkowska, A. M. (2024). Dietary polyphenols, food processing and gut microbiome: recent findings on bioavailability, bioactivity, and gut microbiome interplay. Antioxidants 13:1220. doi: 10.3390/antiox13101220
Singh, R., Chandrashekharappa, S., Bodduluri, S. R., Baby, B. V., Hegde, B., Kotla, N. G., et al. (2019). Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 10:89. doi: 10.1038/s41467-018-07859-7
Singh, R. K., Chang, H. W., Yan, D., Lee, K. M., Ucmak, D., Wong, K., et al. (2017). Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 15:73. doi: 10.1186/s12967-017-1175-y
Slavin, J. (2013). Fiber and prebiotics: mechanisms and health benefits. Nutrients 5, 1417–1435. doi: 10.3390/nu5041417
Song, J. X., Ren, H., Gao, Y. F., Lee, C. Y., Li, S. F., Zhang, F., et al. (2017). Dietary capsaicin improves glucose homeostasis and alters the gut microbiota in obese diabetic Ob/Ob mice. Front. Physiol. 8:602. doi: 10.3389/fphys.2017.00602
Stephanie, R. N. K., Soka, S., and Suwanto, A. (2017). Effect of tempeh supplementation on the profiles of human intestinal immune system and gut microbiota. Microbiol. Indonesia 11, 11–17. doi: 10.5454/mi.11.1.2
Su, Q., and Liu, Q. (2021). Factors affecting gut microbiome in daily diet. Front. Nutr. 8:644138. doi: 10.3389/fnut.2021.644138
Sultan, S., El-Mowafy, M., Elgaml, A., Ahmed, T., Hassan, H., and Mottawea, W. (2021). Metabolic influences of gut microbiota dysbiosis on inflammatory bowel disease. Front. Physiol. 12:715506. doi: 10.3389/fphys.2021.715506
Tan, J., McKenzie, C., Potamitis, M., Thorburn, A. N., Mackay, C. R., and Macia, L. (2014). The role of short-chain fatty acids in health and disease. Adv. Immunol. 121, 91–119. doi: 10.1016/B978-0-12-800100-4.00003-9
Tao, F., Xing, X., Wu, J., and Jiang, R. (2021). Enteral nutrition modulation with n-3 PUFAs directs microbiome and lipid metabolism in mice. PLoS One 16:e0248482. doi: 10.1371/journal.pone.0248482
Tingirikari, J. M. R., Sharma, A., and Lee, H. (2024). Kefir: a fermented plethora of symbiotic microbiome and health. J. Ethnic Foods 11:252. doi: 10.1186/s42779-024-00252-4
Tuli, H. S., Kumar, A., Sak, K., Aggarwal, D., Gupta, D. S., Kaur, G., et al. (2022). Gut microbiota-assisted synthesis, cellular interactions and synergistic perspectives of Equol as a potent anticancer Isoflavone. Pharmaceuticals 15:1418. doi: 10.3390/ph15111418
Ubeda, C., Bucci, V., Caballero, S., Djukovic, A., Toussaint, N. C., Equinda, M., et al. (2013). Intestinal microbiota containing Barnesiella species cures vancomycin-resistant Enterococcus faecium colonization. Infect. Immun. 81, 965–973. doi: 10.1128/IAI.01197-12
Verediano, T. A., Stampini Duarte Martino, H., Dias Paes, M. C., and Tako, E. (2021). Effects of anthocyanin on intestinal health: a systematic review. Nutrients 13:1331. doi: 10.3390/nu13041331
Visioli, F., Risé, P., Barassi, M. C., Marangoni, F., and Galli, C. (2003). Dietary intake of fish vs. formulations leads to higher plasma concentrations of n-3 fatty acids. Lipids 38, 415–418. doi: 10.1007/s11745-003-1077-x
Wang, X., Qi, Y., and Zheng, H. (2022). Dietary polyphenol, gut microbiota, and health benefits. Antioxidants 11:1212. doi: 10.3390/antiox11061212
Wang, M., Yu, F., Zhang, Y., Chang, W., and Zhou, M. (2022). The effects and mechanisms of flavonoids on Cancer prevention and therapy: focus on gut microbiota. Int. J. Biol. Sci. 18, 1451–1475. doi: 10.7150/ijbs.68170
Wang, M., Zhang, Y., Li, C., Chang, W., and Zhang, L. (2023). The relationship between gut microbiota and COVID-19 progression: new insights into immunopathogenesis and treatment. Front. Immunol. 14:1180336. doi: 10.3389/fimmu.2023.1180336
Waters, J. L., and Ley, R. E. (2019). The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 17:83. doi: 10.1186/s12915-019-0699-4
Yadav, M. K., Kumari, I., Singh, B., Sharma, K. K., and Tiwari, S. K. (2022). Probiotics, prebiotics and synbiotics: safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 106, 505–521. doi: 10.1007/s00253-021-11646-8
Yao, Y., Liu, Y., Xu, Q., and Mao, L. (2024). Short chain fatty acids: essential weapons of traditional medicine in treating inflammatory bowel disease. Molecules 29:379. doi: 10.3390/molecules29020379
York, A. (2019). Your microbiome is what you eat. Nat. Rev. Microbiol. 17:721. doi: 10.1038/s41579-019-0287-1
Zhang, T., Gao, G., Kwok, L., and Sun, Z. (2023). Gut microbiome-targeted therapies for Alzheimer’s disease. Gut Microbes 15:613. doi: 10.1080/19490976.2023.2271613
Zhang, N., Huang, X., Zeng, Y., Wu, X., and Peng, X. (2013). Study on prebiotic effectiveness of neutral garlic fructan in vitro. Food Sci. Human Wellness 2, 119–123. doi: 10.1016/j.fshw.2013.07.001
Zhang, M., Li, A., Yang, Q., Li, J., Wang, L., Liu, X., et al. (2021). Beneficial effect of alkaloids from Sophora alopecuroides L. on CUMS-induced depression model mice via modulating gut microbiota. Front. Cell. Infect. Microbiol. 11:665159. doi: 10.3389/fcimb.2021.665159
Zhiguo, Z., Buyu, L., Xingquan, L., Weiwei, H., Zhang, C., Guo, Y., et al. (2023). Effects of steaming on sweet potato soluble dietary Fiber: content, structure, and Lactobacillus proliferation in vitro. Food Secur. 12:1620. doi: 10.3390/foods12081620
Zhong, H., Xu, J., Yang, M., Hussain, M., Liu, X., Feng, F., et al. (2023). Protective effect of Anthocyanins against neurodegenerative diseases through the microbial-intestinal-brain axis: a critical review. Nutrients 15:496. doi: 10.3390/nu15030496
Keywords: healthy gut bacteria, phytochemicals, prebiotic foods and probiotic foods, probiotics, fatty acid
Citation: Kumar S, Mukherjee R, Gaur P, Leal É, Lyu X, Ahmad S, Puri P, Chang C-M, Raj VS and Pandey RP (2025) Unveiling roles of beneficial gut bacteria and optimal diets for health. Front. Microbiol. 16:1527755. doi: 10.3389/fmicb.2025.1527755
Edited by:
Junling Shi, Northwestern Polytechnical University, ChinaReviewed by:
Malgorzata Ziarno, Warsaw University of Life Sciences, PolandCopyright © 2025 Kumar, Mukherjee, Gaur, Leal, Lyu, Ahmad, Puri, Chang, Raj and Pandey. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ramendra Pati Pandey, cmFtZW5kcmEucGFuZGV5QGdtYWlsLmNvbQ==; Suresh Kumar, ZHJzdXJlc2hrdW1hcm1vaGlsQGdtYWlsLmNvbQ==; V. Samuel Raj, ZGlyZWN0b3JjZDRAc3JtdW5pdmVyc2l0eS5hYy5pbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.