 Jinshuo Lian1
Jinshuo Lian1 Yuan Yuan
Yuan Yuan Xinna Zhang
Xinna Zhang Chengyang Xu
Chengyang Xu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 03 February 2025
Sec. Terrestrial Microbiology
Volume 16 - 2025 | https://doi.org/10.3389/fmicb.2025.1506558
This article is part of the Research Topic Anthropogenic Effects on the Microbial Communities of Terrestrial Ecosystems View all 34 articles
Understanding the mechanisms by which seedlings respond to light and water regulation, as well as studying the response of rhizosphere microecology to drought stress, are crucial for forest ecosystem management and ecological restoration. To elucidate the response of the rhizosphere microecology of Quercus dentata and Quercus variabilis seedlings to water and light conditions, and to clarify how plants modulate the structure and function of rhizosphere microbial communities under drought stress, we conducted 12 water-light gradient control experiments. These experiments aimed to offer scientific theoretical support for the dynamic changes in rhizosphere soil enzyme activities and microbial community compositions of these two oak species under varying light and moisture conditions, and subsequently assist in the future breeding and cultivation efforts. The results are summarized as follows: (1) The activities of cellulase, urease, and chitinase in the rhizosphere soil of Q. dentata and Q. variabilis were significantly influenced by water and light treatments (p < 0.05). Urease was particularly sensitive to light, while sucrase exhibited sensitivity to light in Q. dentata and no significant difference in Q. variabilis. (2) Compared to Q. dentata, the rhizosphere bacteria of Q. variabilis demonstrated greater adaptability to drought conditions. Significant differences were observed in the composition of microorganisms and types of fungi in the rhizosphere soil of the two Quercus seedlings. The fungal community is significantly influenced by light and moisture, and appropriate shading treatment can increase the species diversity of fungi; (3) Under different water and light treatments, the rhizosphere soil microbial composition and dominant species differed significantly between the two Quercus seedlings. For instance, Streptomyces, Mesorhizobium, and Paecilomyces exhibited significant variations under different treatment conditions. Specifically, under L3W0 (25% light, 75–85% moisture) conditions, Hyphomonadaceae and SWB02 dominated in the Q. dentata rhizosphere, whereas Burkholderiales and Nitrosomonadaceae were prevalent in the Q. variabilis rhizosphere. Overall, the rhizosphere microecology of Q. dentata and Q. variabilis exhibited markedly distinct responses to varying light and water regimen conditions. Under identical conditions, however, the enzyme activity and microbial community composition in the rhizosphere soil of these two oak seedlings were found to be similar.
Seedlings constitute the fundamental cornerstone of natural regeneration within forest ecosystems, actively participating in and propelling the intricate succession process from establishment to maturity (Zhao et al., 2025). They serve as vital replenishers, counterbalancing tree mortality stemming from both natural and anthropogenic causes, thereby preserving the continuity and resilience of forest ecosystems. In the context of forest restoration endeavors, the complementary dynamic and functional attributes of seedlings hold paramount importance, not only facilitating vegetation recovery but also safeguarding biodiversity (Chen et al., 2018). Furthermore, as a pivotal microbial habitat, the soil environment exerts profound influences on the structure and diversity of microbial communities (Rasche et al., 2011; Richter et al., 2018). This intricate ecosystem encompasses a symphony of nutrients, enzymes, and microorganisms, all intricately intertwined in their influence on plant growth and development (Jang et al., 2020; Xiao D. et al., 2015; Xiao Y. et al., 2015). Notably, soil properties such as pH, texture, moisture content, temperature, organic carbon levels, and nutrient availability play pivotal roles in shaping the composition and diversity of soil microbial communities (Brockett et al., 2012; Cookson et al., 2007; Rousk et al., 2010). These factors, in turn, contribute to the overall health and functioning of forest ecosystems, underscoring the intricate interplay between seedlings, soil, and microbial communities in the grand tapestry of forest restoration and conservation. The rhizosphere environment is an active region for information and material exchange between plants, microorganisms and soil (Lin et al., 2020). Rhizosphere microorganisms have a significant influence on the biochemical cycling process in soil, and they play a crucial role in the health and development of roots (Birt et al., 2022). In terms of rhizosphere microecology, previous studies focused on the effects of crop patterns such as crop rotation, continuous cropping and co-cropping on rhizosphere microecology (Liu et al., 2024; Huang and Li, 2024; Yin et al., 2024), effects of different diseases on plant rhizosphere microecology (Liu, 2023; Zhang et al., 2023), effects of elements required for growth and biological reagents (hormones) on rhizosphere microecology (Wang et al., 2024; Li et al., 2024; Han et al., 2024), lack of external light and moisture conditions on the influence of research.
Soil enzymes represent a crucial indicator of soil microecology, facilitating an array of biochemical reactions that underpin essential processes such as organic matter decomposition and the promotion of carbon, nitrogen, phosphorus, sulfur, and other nutrient cycles. By assessing soil enzyme activity, we can gain indirect insights into soil quality, thereby serving as a vital metric for evaluating soil health and fertility (Lu and Li, 2020). Soil microorganisms, another indispensable element of soil ecosystems, contribute significantly to element cycling, organic matter decomposition, rhizosphere immunity enhancement, soil fertility improvement, and environmental health maintenance (Zhu Y. G. et al., 2021; Huang et al., 2024). Their diversity is intricately influenced by factors such as vegetation composition, soil type, temperature, moisture content, and management practices (Zhou and Lei, 2007). Soil fungi, a vital component of the microbial system, occupy a pivotal role in material circulation and energy flow within soil ecosystems (Zhao L. N. et al., 2020; Tiwari et al., 2021). Similarly, bacterial communities drive soil ecosystem restoration, significantly contributing to ecological restoration, soil fertility enhancement, and water conservation efforts (Yuan and Zhang, 2024; Shi et al., 2024).
Extensive research underscores the profound impact of rhizosphere microbial quantity, diversity, metabolic activity, and interactions on plant health. These microorganisms can either bolster plant resistance and growth or, conversely, act as pathogens, thereby modulating plant health outcomes (Berendsen et al., 2012; Sun et al., 2015; Terhonen et al., 2019). Investigations into rhizosphere microecology have delved into various influencing factors, with Chen Lanlan et al. comprehensively reviewing plant responses to drought stress, encompassing plant morphology, physiological molecular mechanisms, and rhizosphere microbial community dynamics (Chen et al., 2024). This work provides a solid theoretical foundation for future rhizosphere microecology research.
Furthermore, Shen Xintao has illuminated the mechanisms underlying rhizosphere microbial regulation of plant root architecture under acid stress, revealing that these microorganisms can modulate root architecture through hormone production, volatile organic compound release, and mineral nutrient regulation (Shen et al., 2025). This underscores the multifaceted role of rhizosphere microorganisms in shaping the rhizosphere environment beyond mere organic matter decomposition. Investigations into rhizosphere microecology under heavy metal and drought stress have also yielded significant insights. Li Na et al. discovered that cadmium stress alters the composition of plant rhizosphere microbial communities (Li et al., 2025). Meanwhile, Gao Yanting et al. explored the microbial carbon source metabolism response to varying drip irrigation levels in arid and semi-arid regions, revealing that moderate and mild water stress treatments can enhance soil microbial carbon source utilization diversity and metabolic capacity (Gao et al., 2022). These studies collectively highlight the intricate interplay between rhizosphere microorganisms, environmental stressors, and plant health.
Light and water constitute indispensable external stimuli for plant growth and development. Recent groundbreaking findings by Professor Xu Jianming have illuminated, for the first time, the phototropic modulation of the circadian rhythm within rice rhizosphere microbial communities (Zhao et al., 2021), demonstrating a pronounced decrease in the α-diversity of these microbial assemblages under light exposure. Furthermore, Xie et al. (2023) have observed a marked enhancement in the abundance of rhizosphere microorganisms, particularly those beneficial to plant growth (e.g., lysobacteria and Ramlibacter), along with heightened soil enzyme activity under LED illumination, underscoring the profound influence of light on soil biochemical processes. Concurrently, soil moisture status emerges as a pivotal environmental factor governing soil microbial dynamics and enzymatic activities, as evidenced by numerous studies (Ghezzehei et al., 2019; Siebielec et al., 2020). Notably, Cao Yajing has revealed that while chronic water scarcity prompts microbial adaptation strategies, acute drought stress can lead to microbial cell demise due to loss of resilience (Cao et al., 2023). Under drought conditions, the rhizosphere microbial landscape undergoes profound shifts in diversity, composition, and metabolic activity. Despite these insights, a notable gap persists in rhizosphere microecology research that specifically examines the combined effects of water and light as external factors. The relative significance of these two variables in modulating soil enzyme activities remains elusive. Given the paramount importance of fine-tuning soil enzyme activity and rhizosphere microbial proliferation during plant ontogeny for optimal seedling establishment, this study endeavors to dissect the differential responses of rhizosphere microecosystems in two Quercus species seedlings to light and water manipulations. The intricate interplay between these environmental cues, soil enzymes, and rhizosphere microbial communities remains an enigma, necessitating further exploration to unravel their complex relationships.
Q. dentata and Q. variabilis Blume, two prominent deciduous members of the Fagaceae family, exhibit widespread distribution and multifaceted applications (Liu, 2020; Sun et al., 2023). Their seeds are replete with starch, while their wood and bark are rich sources of tannin, amenable to extraction. In China, Quercus species form the backbone of numerous forest communities and occupy a preeminent position among dominant tree species, embodying the virtues of “native adaptability, longevity, stress resilience, food value, and aesthetic appeal.” Their unparalleled contribution to ecological, economic, cultural, and tourism sectors underscores their significance. Within the urban greenscape of Beijing, Q. variabilis, as a native species and a recommended tree for urban forestry, holds a pivotal role in the city’s “Underforest Supplementation Oak” initiative, exemplifying exceptional ecological potential (Huang et al., 2022; Zhang et al., 2024). While extensive research has delved into the functional traits, physiological and biochemical characteristics, and photosynthesis of Q. dentata and Q. variabilis (Qi et al., 2024; Sun et al., 2021; Hao et al., 2020), as well as their seedling cultivation, growth, and development (Cao et al., 2018; Wu et al., 2022; Wang et al., 2022), the intricate interplay between their habitat conditions, soil enzymes, and rhizosphere microbial communities remains an enigma. To unravel this complexity, our study devised a series of 12 experiments, each incorporating distinct water-light gradients, aiming to decipher the dual influence of light and water on the rhizosphere microecology of these two oak species. Leveraging bioinformatics and high-throughput sequencing technologies, we analyzed the dynamic fluctuations in rhizosphere soil enzyme activities and microbial community structures under varying light and water regimes. Our objectives are twofold: (Berendsen et al., 2012) to determine the extent and mechanisms by which light and water conditions impact rhizosphere soil enzyme activities in both oak species; (Birt et al., 2022) to elucidate how light and water conditions reshape the rhizosphere microbial community composition of the seedlings of these two oak species.
The research was conducted within the confines of the Nanda Wilderness Nursery, situated at 116.19°E, 39.89°N in the Shijingshan District of Beijing. To mimic the diverse understory light regimes in natural settings (Quero et al., 2006), a custom-built shade/canopy structure, spanning 36 square meters, was erected. The primary framework was fabricated from galvanized steel tubes, with transparent rigid plastic sheets serving as the covering for all four light intensity levels. For the three shading treatments, 25, 50, and 75% black shading screen fabrics were incorporated, respectively (Ncise et al., 2020).
This design allowed for flexibility in managing light exposure, with the plastic sheet being readily rolled up during sunny days to optimize light penetration and swiftly secured during rainy periods to shield against precipitation interference, ensuring a controlled environment conducive to experimentation. To maintain optimal ventilation and ensure the integrity of the experimental setup, special attention was paid to the canopy’s ventilation capabilities.
Furthermore, the study employed 1.5-year-old container seedlings of Q. dentata and Q. variabilis, both of which were bred under uniform provenance conditions and subjected to identical management and protection protocols.
In the experimental setup, a mixed substrate composed of vermiculite and peat in a 1:1 ratio was employed to establish an appropriate growth milieu. The potted seedlings were transferred to the nursery and subjected to a 45-day acclimatization period. During this stage, routine water and fertilizer management protocols were implemented. Watering was conducted at intervals of 3 to 5 days, contingent upon the prevailing weather conditions. Additionally, Osmocote 315 s (at a dosage of 3–5 grams per pot) was applied once to supply the essential nutrient elements, including nitrogen, phosphorus, potassium, etc., requisite for seedling growth. Simultaneously, the fungicide chlorothalonil was sprayed to safeguard against diseases, pests, and fungal and bacterial infections, with applications occurring once every 5 days for a total of 3 repetitions.
Samples seedlings were, respectively, drawn from a total of 120 seedlings under 12 treatment conditions. From this cohort, 36 pots of each species, exhibiting robust growth and free from pests and diseases, were selected to serve as the sample plants for this investigation.
The mean plant height of the potted Q. dentata seedlings was quantified at 31.03 cm, concomitant with a basal diameter of 4.31 mm; whereas the mean plant height of the potted Q. variabilis seedlings was ascertained to be 42.10 cm, along with a basal diameter of 6.37 mm. The initial growth status of the sample seedlings are detailed in Table 1.
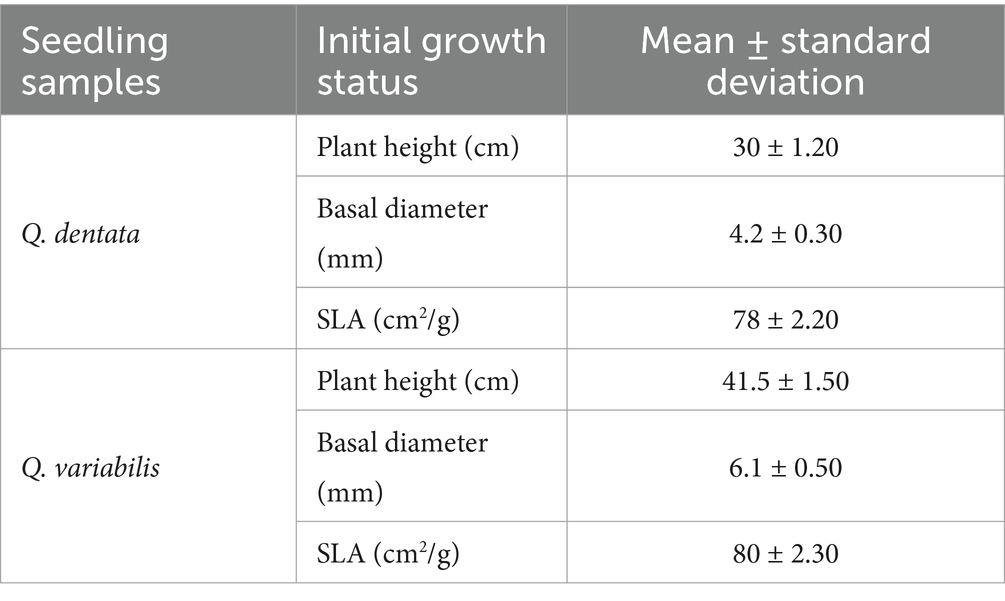
Table 1. The initial growth status of the sample seedlings.
Four distinct light intensity levels were established: natural light (L0, corresponding to 100% of natural illumination), light shading (L1, 75% of natural light), medium shading (L2, 50% of natural light), and deep shading (L3, 25% of natural light). During seedling growth, the light intensity under varying shade conditions was gauged via the “five-point method.” The sunshade’s configuration was promptly adjusted in accordance with weather changes, guaranteeing that all weather factors, except light intensity, remained relatively uniform across different light treatment zones.
To accurately assess and maintain soil water content across these gradients, a combined approach utilizing weighing methods alongside a soil temperature and humidity meter (YM-19-2) was employed. This methodology facilitated the calculation of soil water content and subsequent water supplementation, targeting specific treatment gradients: 75–85% of field water capacity (W0), 45–55% (W1), and 15–25% (W2). Based on prevailing weather conditions, soil water content measurements were conducted every 1–2 days in the evening, with three pots randomly selected from each treatment group. Irrigation was then administered as necessary to adhere to the prescribed water content ranges (Figure 1).
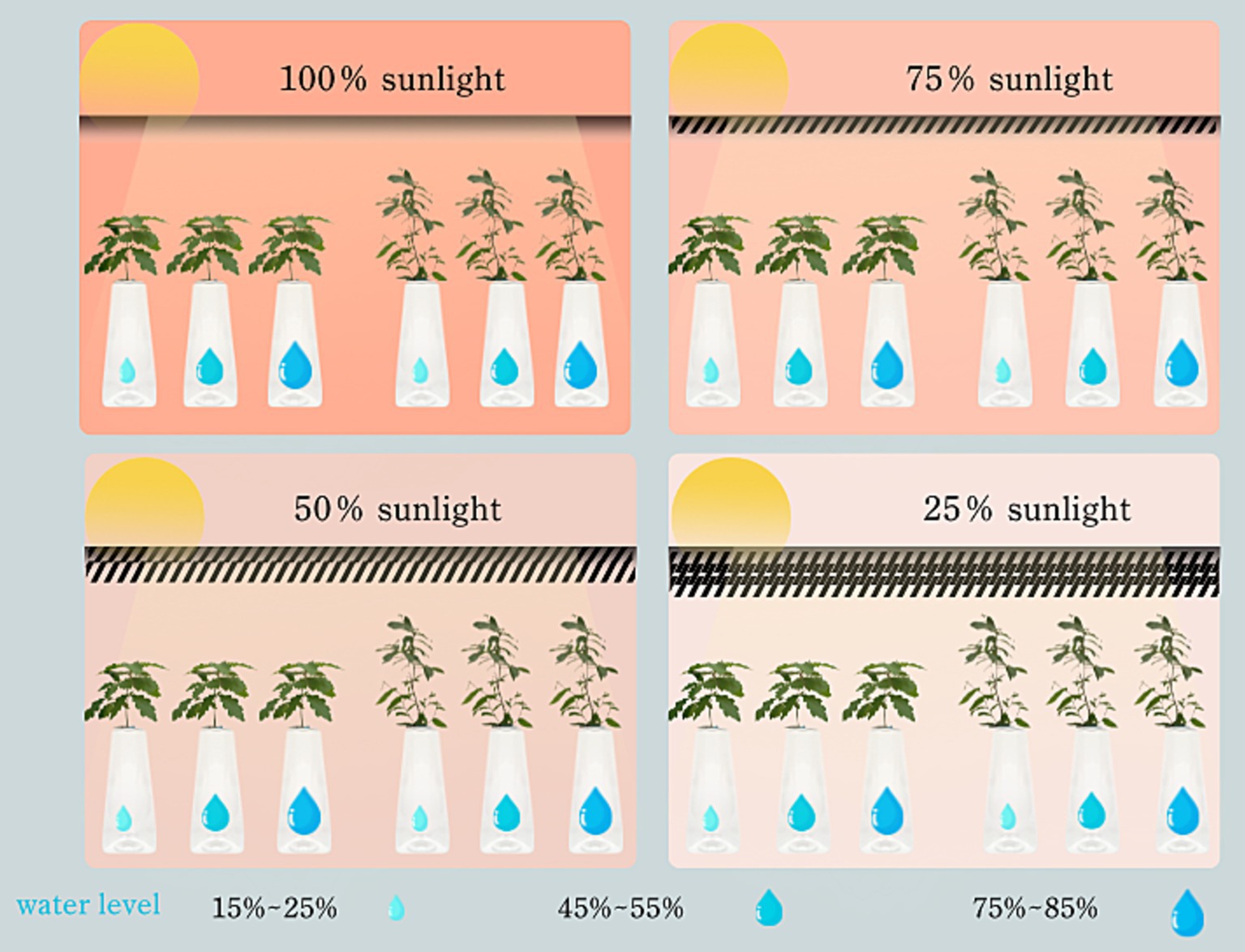
Figure 1. Schematic diagram of experimental design.
The experimental design adopted a completely randomized block design, ensuring robust statistical analysis and minimizing potential biases. The specific settings and configurations of this experimental setup are detailed in Tables 2, 3.
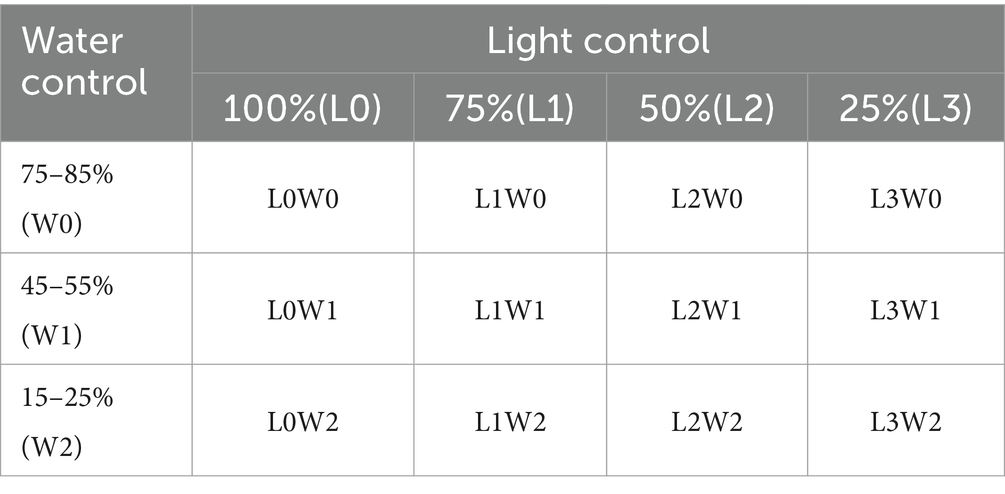
Table 2. Design of the experimental group.
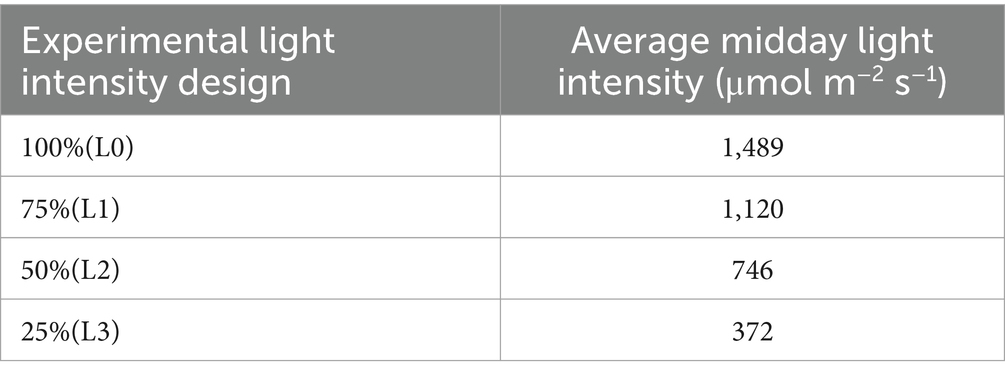
Table 3. The light intensity under different shading conditions.
In August 2023, we conducted a meticulous sampling of rhizosphere soil from a total of 72 seedlings belonging to two distinct species of Quercus plants. Upon reaching the sampling stage, the soil-root matrix within each container was transferred into sterile sampling bags to maintain the integrity and purity of the samples. Upon arrival at the laboratory, a delicate procedure was followed to separate the roots from the soil, which involved gently shaking the roots to dislodge loose soil particles. Subsequently, a sterile brush was employed to collect the residual soil adhering to the roots. The collected rhizosphere soil samples were then passed through a 20-mesh sterile sieve to eliminate larger particulate impurities, ensuring the purity and homogeneity of the samples. The resulting soil samples were promptly subjected to liquid nitrogen quick-freezing for preservation and stored at −80°C in a refrigerator to maintain their integrity until further analysis. From each soil lot, we prepared three replicate samples (yielding a total of 216 small samples), with each individual sample consisting of precisely 0.3 grams of rhizosphere soil. Once the sampling process was completed, all samples were stored in low-temperature conditions to preserve their quality and subsequently dispatched to Shanghai Personalbio Gene Technology Co., Ltd. for comprehensive sequencing analysis.
During the amplicon sequencing analysis, 0.3 g of soil was weighed for each sample. The soil was homogenized, the microorganisms were lysed, and the DNA was isolated and extracted according to the operating instructions in the DNA extraction kit. The purity of the extracted nucleic acid was detected using a NanoDrop micro-spectrophotometer. In addition, 2% agarose gel electrophoresis was employed to examine the integrity of the genomic DNA, to check for degradation and contamination such as proteins. The target fragments were excised and then recovered using the Axygen gel recovery kit. For PCR, the NEB Q5 DNA high-fidelity polymerase was used, and the hypervariable region V3-V4 of the bacterial ribosomal 16S rRNA was selected as the target region for polymerase chain reaction (PCR) amplification. The specific primers for the bacterial 16S rRNA V3-V4 region were 338F (5′-barcode+ACTCCTACGGGAGGCAGCA-3′) and 806R (5’-GGACTACHVGGGTWTCTAAT-3′). The barcode in the forward primer is an oligonucleotide sequence of 7–10 bases, which is used to distinguish different samples in the same library. Similarly, for fungal sequencing, the rDNA ITS sequence analysis technique ITS was utilized for PCR amplification. The ITS1 region primers ITS5 (GGAAGTAAAAGTCGTAACAAGG) and ITS2 (GCTGCGTTCTTCATCGATGC) were selected for amplification and sequencing. The barcode in the forward primer is an oligonucleotide sequence of 7–10 bases, which is used to distinguish different samples in the same library, and the amplification system was the same as that for bacteria.
The PCR products were quantified on a Microplate reader (BioTek, FLx800) using the Quant-iT PicoGreen dsDNA Assay Kit, and then the samples were pooled according to the required data amount for each sample. During the library construction process, the TruSeq Nano DNA LT Library Prep Kit of Ilumina was used for library construction. Firstly, in the end repair process, the End Repair Mix2 in the kit was used to remove the protruding bases at the 5′ end of the DNA, fill in the missing bases at the 3′ end, and add a phosphate group at the 5′ end. Finally, the library was subjected to quality inspection, data quality control, and analysis.
To assess the enzyme activity within the rhizosphere soil of two Quercus seedlings, we first weighed 0.1–0.5 grams of rhizosphere soil samples, passing them through a 60-mesh sieve to ensure homogeneity. These samples were then transferred into 2-mL centrifuge tubes for subsequent analysis. For the determination of urease activity, the samples were incubated at 37°C for 20 min using the sodium phenol-sodium hypochlorite colorimetric method, following which colorimetric analysis was performed at 578 nm using ultraviolet spectrophotometry, as described by Guan (1986). Similarly, soil catalase activity was determined using a standardized method, with colorimetric readings taken at 240 nm by ultraviolet spectrophotometry, adhering to the protocol outlined by Yang L. F. et al. (2011). For the quantification of soil sucrase and cellulase activities, we employed the 3,5-dinitrosalicylic acid colorimetric method, with colorimetric measurements conducted at 540 nm using ultraviolet spectrophotometry. Meanwhile, soil acid phosphatase activity was determined using the phenylene disodium phosphate colorimetric method, with colorimetric readings taken at 400 nm, as detailed by Li et al. (2008). Lastly, soil chitinase activity was measured by a specific method, with colorimetric analysis performed at 544 nm using ultraviolet spectrophotometry, according to the guidelines provided by Gu and Hu (1994). These rigorous methodologies ensured accurate and reliable assessments of the various enzyme activities present in the rhizosphere soil of the Quercus seedlings.
Experimental data integration and processing were facilitated using Microsoft Excel 2020 software, while statistical analyses were conducted with SPSS 27.0 software (SPSS, Chicago, IL). To investigate the responses of six soil enzymes in the rhizosphere of Q. dentata and Q. variabilis seedlings to varying water and light conditions, a two-way ANOVA was employed. For data visualization, Origin 2017, R version 4.1.2, and Python programming languages were utilized. Regarding microbiome bioinformatics, QIIME2 (version 2019.4) served as the primary tool for analysis. Primer fragments were excised using cutadapt, and subsequent steps including quality control, noise reduction, sequence assembly, and chimera removal were executed with DADA2. ASV (Amplicon Sequence Variants) feature sequences and tables were merged, and each ASV’s feature sequence was classified using QIIME2’s classify-sklearn algorithm, leveraging a Naive Bayes classifier against the Greengenes database for species annotation. To ensure consistent sample depth, the ASV table was rarefied to 95% of the minimum sample sequence size using QIIME2’s qiime feature-table rarefy function. All subsequent analyses were conducted on this rarefied ASV table, providing a robust foundation for our ecological insights.
The results presented in Figure 2 reveal several key findings regarding the enzymatic activities in the rhizosphere soil of Q. dentata and Q. variabilis. Specifically, the urease activity in the rhizosphere soil of Q. dentata was significantly influenced by light (p < 0.05) and the interaction between water and light (p < 0.01). In contrast, the urease activity in the rhizosphere soil of Q. variabilis exhibited significant differences under varying light (p < 0.001) and water-light interaction (p < 0.001) conditions, as well as under different water conditions (p < 0.01). The activity of acid phosphatase in the rhizosphere soil of Q. dentata was notably affected by the interaction between water and light (p < 0.05), whereas in Q. variabilis, this activity showed significant differences under different water conditions (p < 0.05). The activities of sucrase in the rhizosphere soil of both Quercus seedlings were not significantly impacted by water and light (p > 0.05). Within the L2 group, the sucrase activities in the rhizosphere soil of Q. variabilis seedlings ranged from 4.56 mg/d/g to 63.36 mg/d/g, while those in Q. dentata ranged from 2.94 mg/d/g to 87.80 mg/d/g, the range of sucrase activity in the rhizosphere of the same type of seedlings was relatively large.
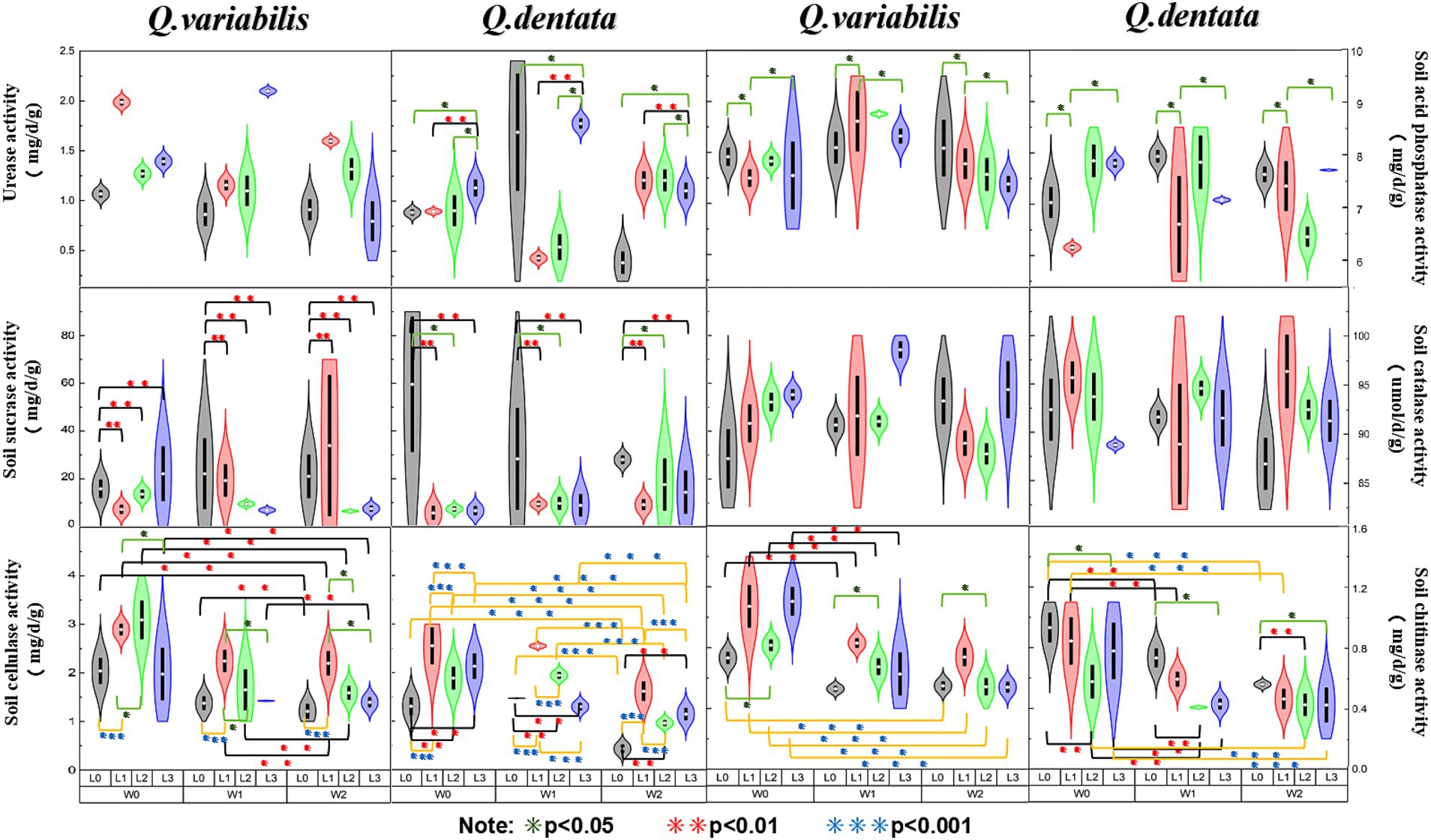
Figure 2. Soil enzyme activities among different seedling growth environments (*p < 0.05, **p < 0.01, ***p < 0.001).
The catalase activity in the rhizosphere soil of Q. variabilis showed significant differences under different light conditions (p < 0.05), whereas the influence of water on catalase activity in both Quercus species was not significant. Both water and light conditions had significant effects on the cellulase activity in the rhizosphere soil of Q. dentata and Q. variabilis seedlings (p < 0.01), with an extremely significant effect observed in Q. dentata seedlings (p < 0.001). Lastly, the chitinase activity in the rhizosphere soil of both Quercus plants was significantly influenced by light (p < 0.05) and water (p < 0.001).
The microbial diversity associated with Q. dentata exhibited the lowest levels under the L1W0 conditions (75–85% water, 75% light), encompassing 5 phyla, 11 classes, 21 orders, 27 families, 33 genera, and 37 species of fungi, as well as 21 phyla, 40 classes, 87 orders, 119 families, and 145 genera of bacteria. Notably, an identical claim of minimal diversity was inadvertently repeated for L2W0 conditions, which should be corrected to reflect unique findings if applicable; however, for the sake of this revision, we will focus on the intended comparison. Under the 50% light condition, the microbial diversity peaked, with a comprehensive tally of 6 phyla, 11 classes, 24 orders, 29 families, 48 genera, and 56 species of fungi, alongside 23 phyla, 48 classes, 97 orders, 136 families, and 159 genera of bacteria.
Among the 12 treatments examined, the highest fungal diversity was observed under L0W2 conditions (15–25% water, 100% light), featuring 4 phyla, 12 classes, 22 orders, 30 families, 38 genera, and 43 species of fungi. Conversely, bacterial diversity reached its zenith under L3W1 conditions (45–55% water, 25% light). Under L2W2 conditions (15–25% water, 50% light), a rich bacterial assemblage of 19 phyla, 40 classes, 91 orders, 128 families, 158 genera, and 27 species coexisted with a fungal community comprising 4 phyla, 9 classes, 15 orders, 22 families, 28 genera, and 32 species. Under full light exposure (100% light), bacterial diversity was notably depleted, consisting of 17 phyla, 35 classes, 81 orders, 109 families, 132 genera, and only 18 species. When comparing the rhizosphere bacteria of Q. variabilis and Q. dentata, the former demonstrated greater adaptability to drought conditions, whereas the latter exhibited a higher overall bacterial diversity (Figure 3).
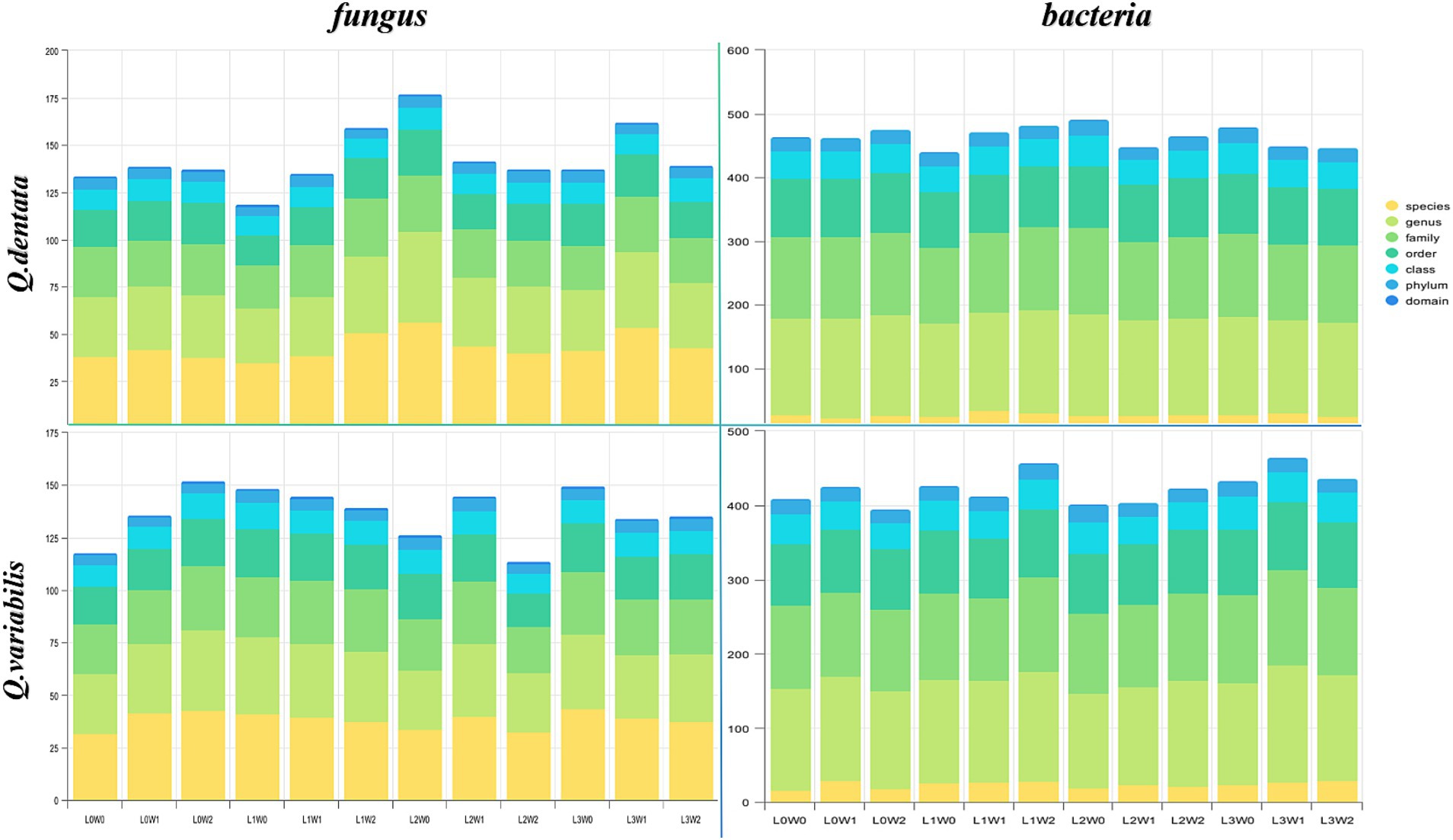
Figure 3. The number of taxa of rhizosphere microorganisms.
The results indicate significant variations in fungal and bacterial communities within the rhizosphere soil of both Q. dentata and Q. variabilis in response to different light and water treatments. Specifically, for Q. dentata, the Chao 1 (p < 0.01) and Simpson (p < 0.05) indices revealed that fungi exposed to the L1 light treatment exhibited significantly higher abundance compared to those under the L0 treatment, with the L1W2 treatment yielding the highest fungal richness. This suggests that fungi in the rhizosphere soil of Q. dentata thrive better under L1 light conditions. Furthermore, in the Pielou’s evenness index (p < 0.05), the highest distribution uniformity was observed under L2W0 conditions (75–85% water, 50% light), which distinctly differed from other treatments. Notably, for bacteria, the L2W0 conditions also demonstrated the highest diversity and most uniform community distribution in both the Simpson (p < 0.05) and Pielou (p < 0.05) indices, with these differences being more pronounced compared to other treatments. However, in the Chao 1 index (p > 0.05), no significant difference was observed in rhizosphere soil bacterial community richness (Figure 4).
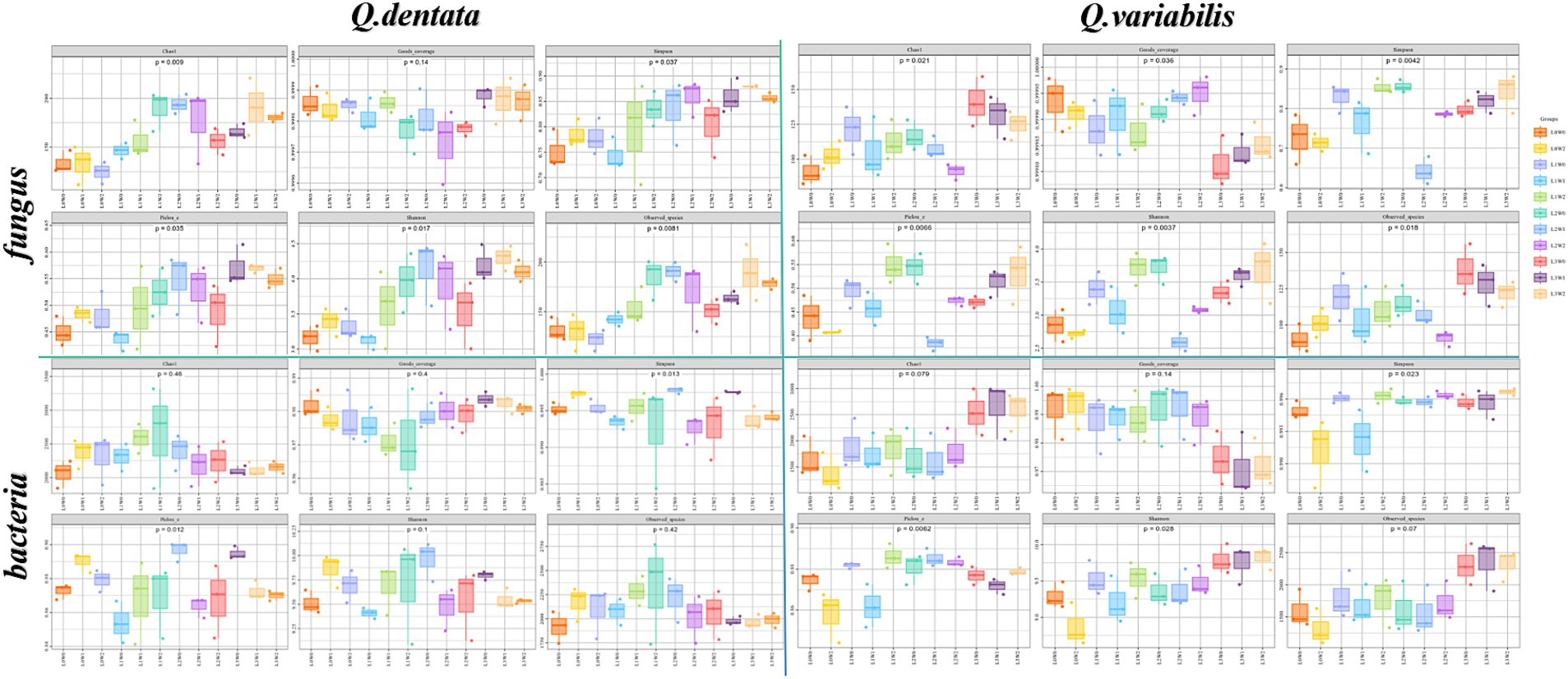
Figure 4. Rhizosphere microbial Alpha diversity index.
In the case of Q. variabilis, the Chao 1 index (p < 0.05) showed that fungal richness peaked under the L3 light condition, with the L3W0 treatment exhibiting the most significant difference compared to others. The Simpson index (p < 0.01) further confirmed that the L1 light treatment significantly outperformed the L0 treatment, while other treatments also showed significant differences. Additionally, the Pielou’s evenness index (p < 0.01) indicated that the distribution uniformity of fungi in rhizosphere soil was significantly influenced by both light and water, with notable differences observed between L2W0 (75–85% water, 50% light), L2W1 (45–55% water, 50% light), L3W2 (15–25% water, 25% light), L3W0, and other treatments. Analysis of bacterial groups revealed substantial bacterial diversity in the rhizosphere under L2 and L3 light conditions in the Simpson index (p < 0.05), with significant differences compared to L0W2 (15–25% water, 100% light) and L1W1 (45–55% water, 75% light). However, in the Shannon index (p < 0.05), L3 exhibited the greatest bacterial diversity under light conditions, with significant differences from other treatments (Figure 4).
The Principal Coordinates Analysis (PCoA) diagrams provide further insights into the bacterial community compositions. The first axis of the PCoA diagram for Q. dentata explains 14.4% of the inter-sample differences, while the second axis accounts for 9%. Visual inspection of the diagram reveals substantial differences among bacterial communities from treatments such as L1W1, L0W1, L2W1, L3W1, L0W0, L1W0, L2W0, and L3W0. Similarly, for Q. variabilis, the first axis of the PCoA diagram explains 16.4% of the inter-sample differences, and the second axis explains 8.5% (Figure 5).
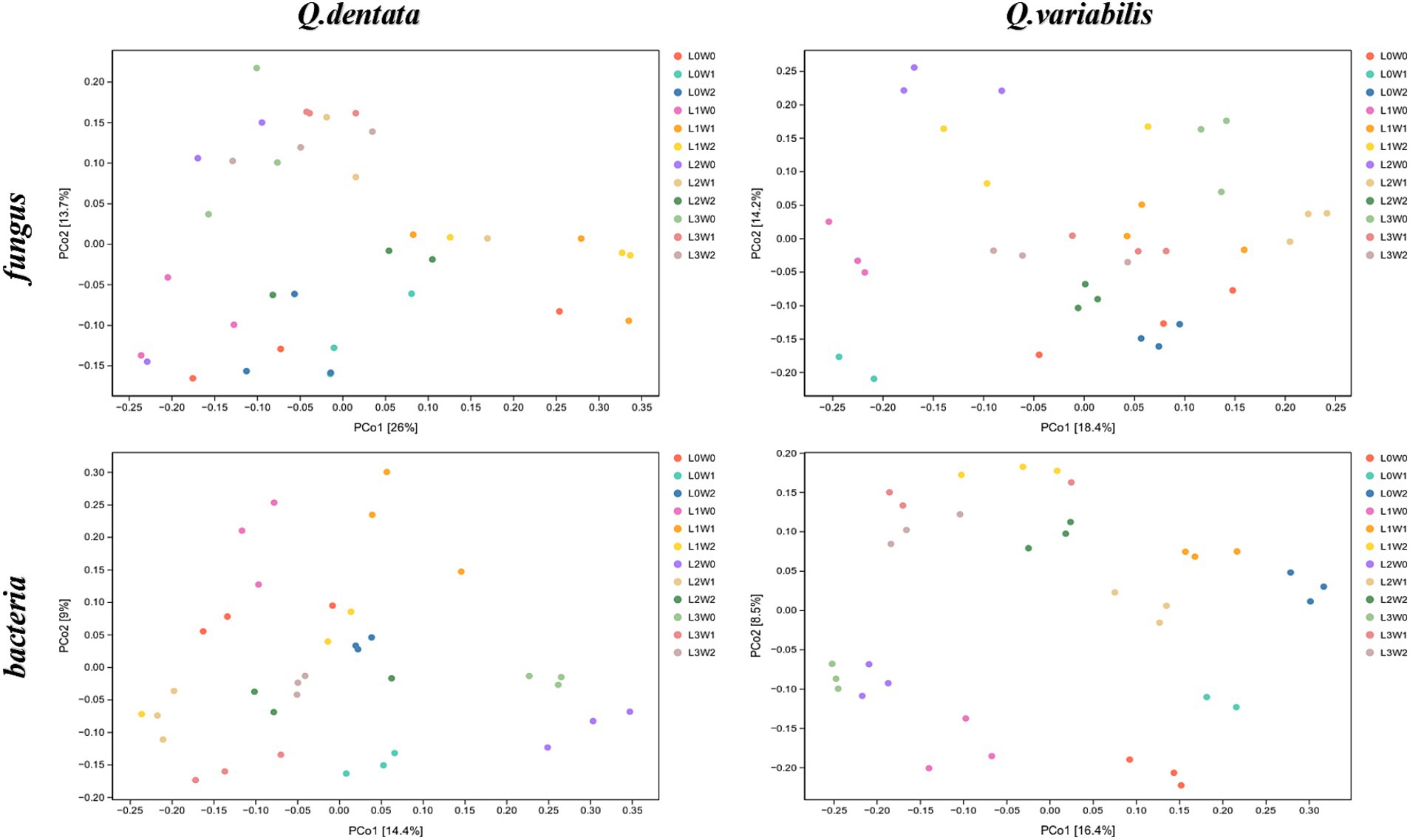
Figure 5. PCoA analysis of rhizosphere microorganisms.
The species difference analysis of rhizosphere fungal communities was conducted for two Quercus seedlings, with the Wayne diagram analysis revealing that 27 ASVs in the rhizosphere soil of Q. dentata were common across all 12 treatment groups, whereas 17 ASV species in the rhizosphere soil of Q. variabilis were similarly shared among these groups (Figure 6). Notably, the number of bacterial species in the rhizosphere significantly outnumbered the fungal species. Analysis of rhizosphere bacterial communities indicated that 324 ASV species were shared among the 12 treatment groups in the rhizosphere soil of both seedlings, with 218 ASVs commonly found in the rhizosphere soil of Q. variabilis (Figure 6).
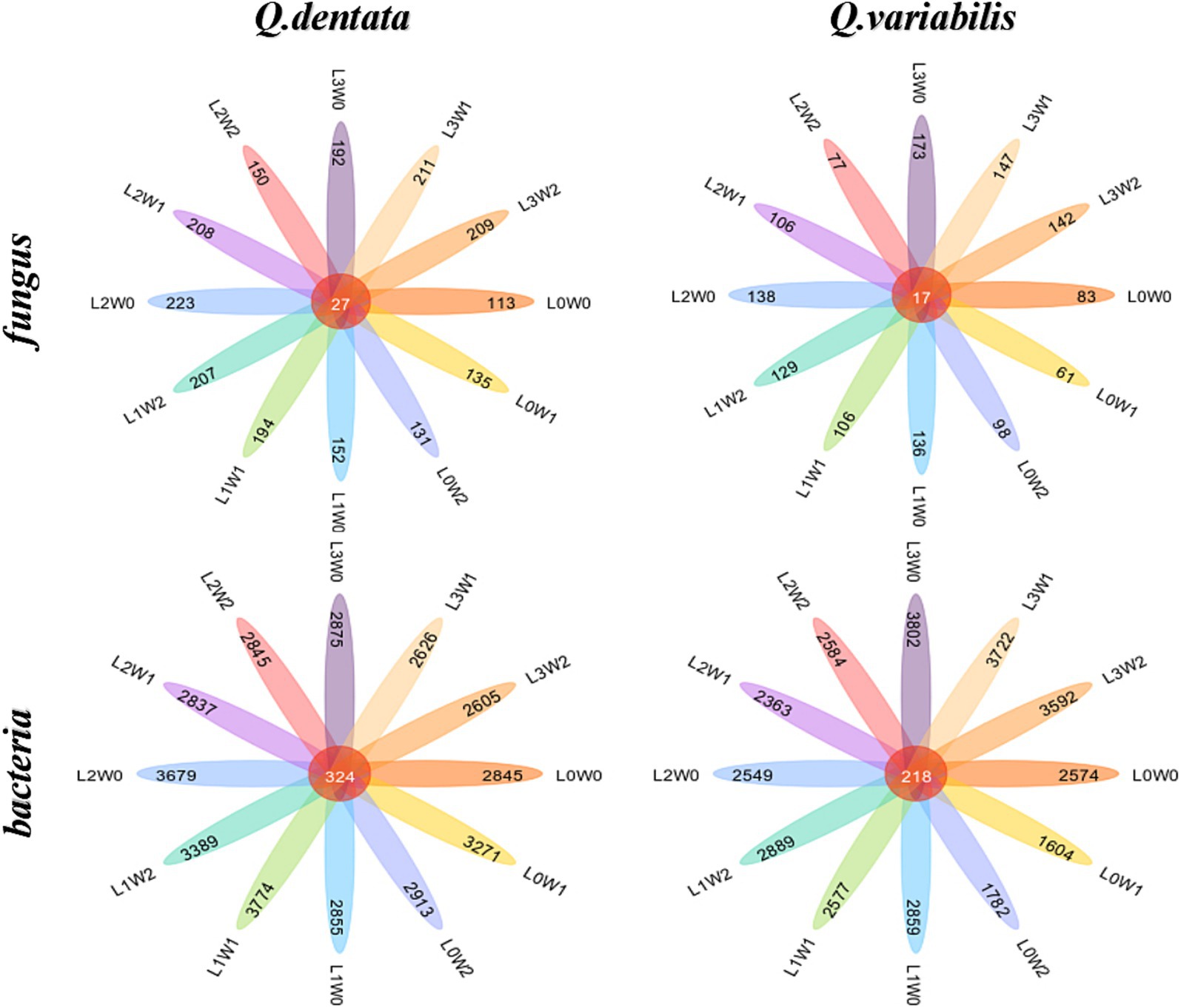
Figure 6. Wayne diagram of rhizosphere soil microbial ASV under different treatments.
To further compare species composition among the samples and visualize the trend of species abundance distribution, heat maps were generated using abundance data for the top 20 genera with the highest average abundance. Additionally, UPGMA clustering of the samples was performed based on the Euclidean distance of species composition data. The results demonstrated that species such as Cercospora, Botryotrichum, Frankia, and Sphingobium exhibited minimal variation under different treatments. Conversely, significant differences were observed among the microbiomes of Rozellomycota, Penicillium, Trichoderma, Streptomyces, Mesorhizobium, and Gemmatimonas. Specifically, the rhizospheres of Q. variabilis (with respect to Gemmatimonas), Fusarium (with respect to Fusarium), and Cephalotrichum (with respect to Cephalotrichum) showed limited differences under various treatments. However, significant variations were noted for Penicillium, Streptomyces, Mesorhizobium, Methylobacterium, and other microbiomes (Figure 7).
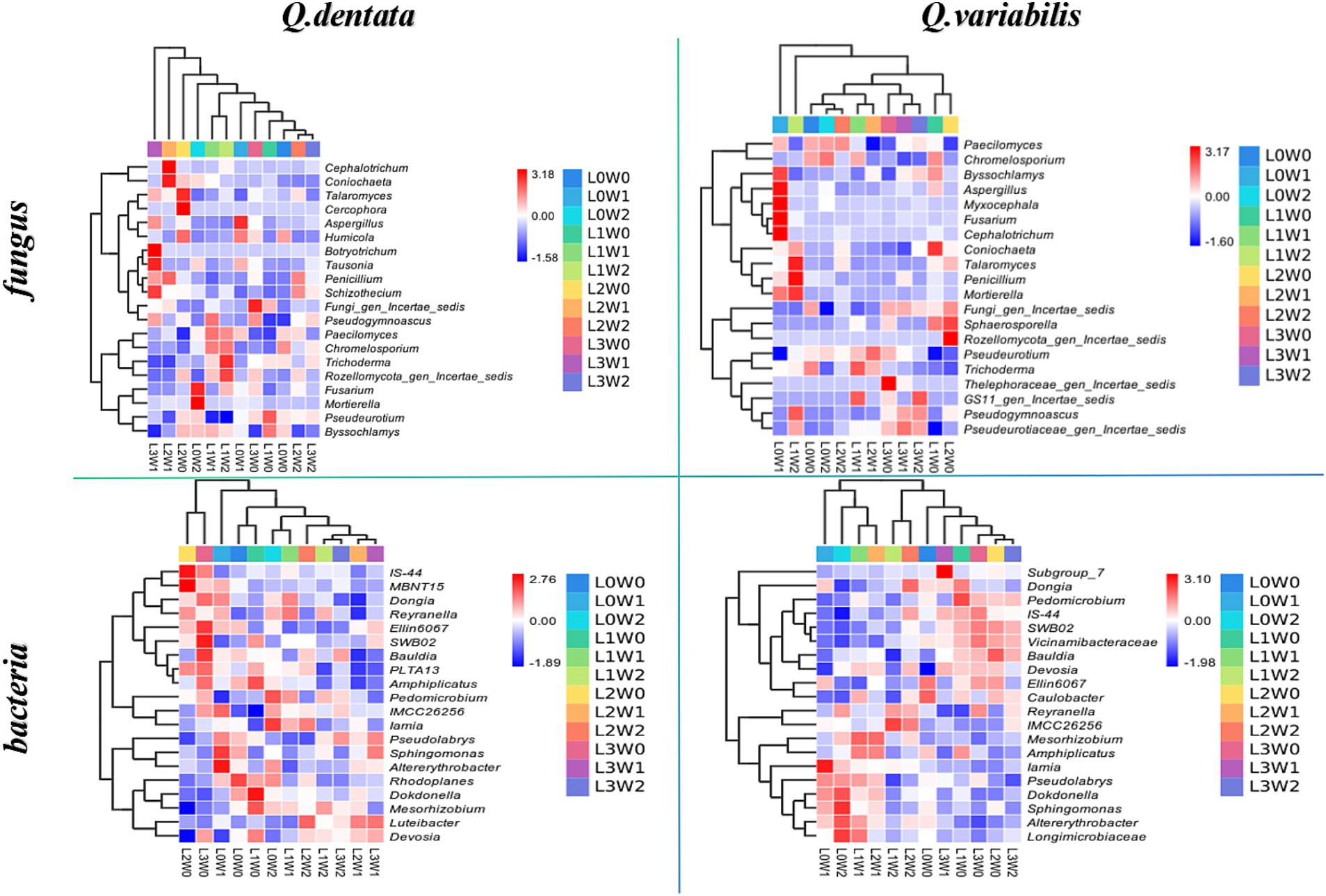
Figure 7. Heat map of rhizosphere soil microbial species composition under different treatments.
An LDA threshold of 2 was established, such that only species surpassing this threshold were deemed biomarker species with significantly higher abundance compared to other groups. The LDA histogram depicted the distinct rhizosphere microbial species in seedlings of Q. dentata and Q. variabilis under 12 water and light condition groups. The results highlighted significant differences in rhizosphere soil microbial composition between the two Quercus seedlings under different treatments. Notably, Ascomycota was found in the rhizosphere of Q. dentata under L0W0 (100% light, 75–85% water) treatment, while Nectriaceae fungi, Acidimicrobiia bacteria, and actinomycetes were detected in the rhizosphere of Q. variabilis under L0W1 (100% light, 45–55% water) treatment. Under L1W0 (75% light, 75–85% water) treatment, Thelebolales fungi and Leotiomycetes fungi were identified in the rhizosphere of Q. variabilis. Additionally, Cephalotrichum and Microascales were observed in the rhizosphere of Q. dentata under L2W1 (50% light, 45–55% water) treatment, while Gemmatimonadetes was found in the rhizosphere of Q. variabilis under L0W2 (100% light, 15–25% water) treatment. Furthermore, Bauldia was detected in the rhizosphere of Q. variabilis under L2W0 (50% light, 75–85% water) treatment, and Luteibacter was found in the rhizosphere of Q. dentata under L2W2 (50% light, 15–25% water) treatment. Under L3W0 (25% light, 75–85% moisture) treatment, Hyphomonadaceae and SWB02 were identified in the rhizosphere of Q. dentata, while the effect value of microorganisms such as Burkholderiales and Nitrosomonadaceae in the rhizosphere of Q. variabilis exceeded 4, indicating these microorganisms as significantly different species among the samples (Figure 8).
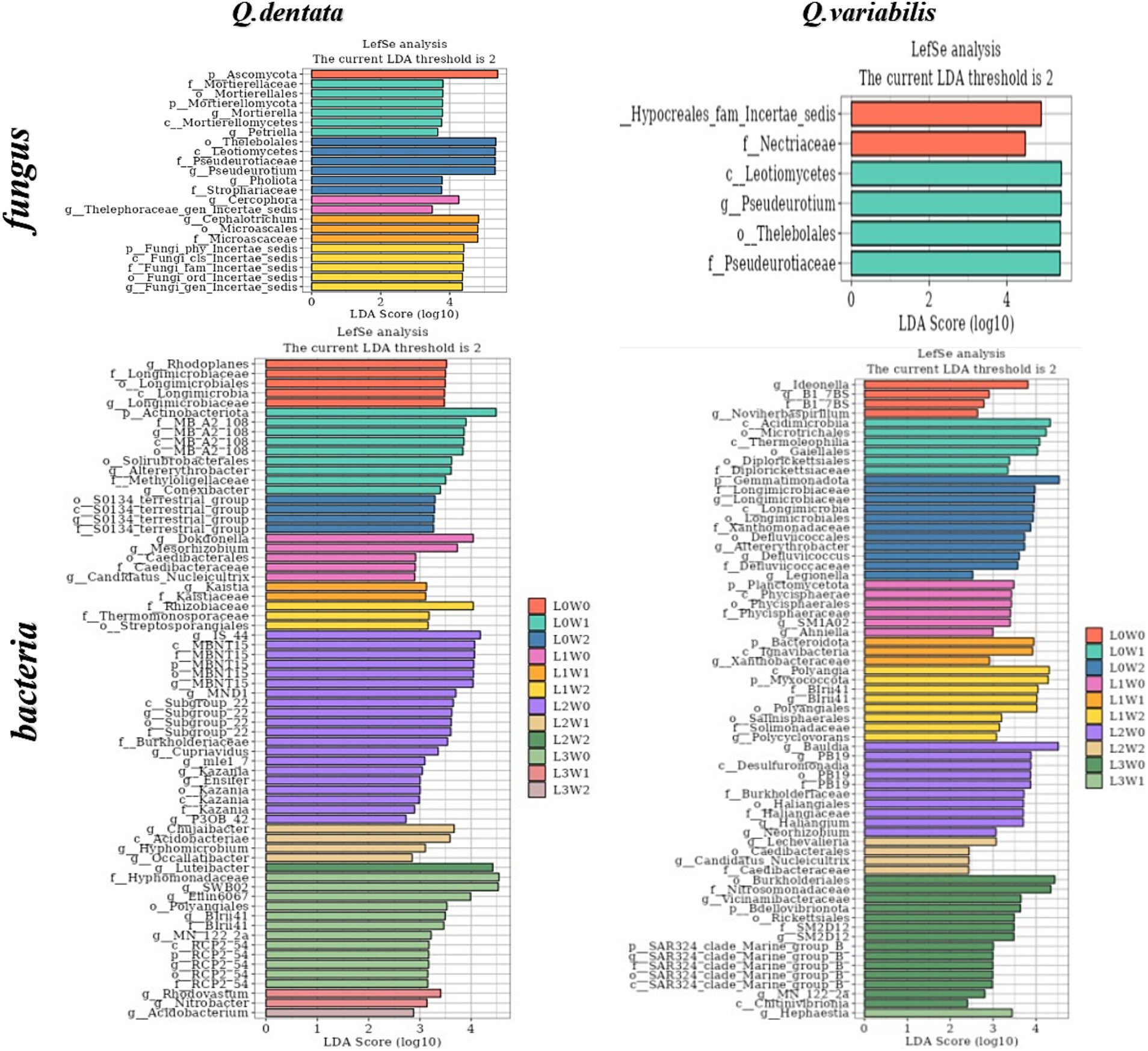
Figure 8. Histogram of rhizosphere microbial LDA.
Previous research on soil enzyme activities has predominantly emphasized the input of nutrients, particularly nitrogen and phosphorus. However, our focus shifts to the rhizosphere microenvironment, which serves as a dynamic and highly active zone for soil microorganisms. This microenvironment exhibits a marked responsiveness to alterations in the soil milieu, and this sensitivity is manifested through alterations in enzyme activities (Zhang et al., 2014). Notably, soil enzymes not only exert a direct influence on plant growth but can also indirectly impact plant health by disrupting the plant’s antioxidant system (Lian et al., 2024). Our findings align with these observations, indicating that the rhizosphere’s response to environmental cues is intricately linked to soil enzyme activities, which in turn regulate plant growth and resilience. However, it is important to note that not all microorganisms are susceptible to drought conditions. In fact, some microorganisms demonstrate remarkable drought resistance, which can alter the overall activity of extracellular enzymes in soil (McDaniel et al., 2013; Zuccarini et al., 2023). In specific instances, a compensatory mechanism arises between plants and microorganisms in response to drought stress. This mechanism helps to mitigate the detrimental effects of drought on extracellular enzymes, thereby bolstering the resilience of the entire ecosystem (Preece and Penuelas, 2016).
Several studies have highlighted that soil urease is not only a pivotal enzyme for urea hydrolysis but also possesses heat stability (Wang and Huang, 2003). Our study revealed that the activity of this enzyme is significantly influenced by light and water conditions. Specifically, excessive light may expedite the decomposition of urease, disrupting its structure, reducing its concentration, and consequently weakening its activity. These findings align with the research conducted by Cai et al. (2016), which observed significant differences in urease activity in the rhizosphere soil of Q. variabilis under varying light and water-light interaction conditions. Notably, compared to Q. dentata, the urease activity in the rhizosphere soil of Q. variabilis sinensis seedlings exhibited greater sensitivity to changes in water and light. Given the strong correlation between urease activity and soil nitrogen content, which facilitates the conversion of urea into ammonia (Craine et al., 2009), variations in urease activity serve as a reliable indicator of the intensity of soil nitrogen transformation processes (Tan, 2024).
Acid phosphatase (ACP) is a ubiquitous hydrolase in plants that plays a crucial role in phosphorus absorption and utilization (Zhao X. W. et al., 2020; Hu et al., 2024). Prior studies have shown that ACP activity is influenced by the plant’s phosphorus supply status (Song et al., 1999; Dracup et al., 1984). Our study revealed that the response of ACP in the rhizosphere soil of two Quercus species to changes in external environmental factors varied. Specifically, ACP activity in the rhizosphere soil of Q. dentata was affected by both light and water, whereas ACP activity in the rhizosphere soil of Q. variabilis was more significantly influenced by water alone. This difference may be attributed to the rhizosphere effect of Q. dentata seedlings, which is produced under the dual influence of light and water (Figure 2). The rhizosphere effect results in higher enzyme activity in rhizosphere soil compared to non-rhizosphere soil, and the presence of numerous microorganisms around plant roots facilitates the enhancement of phosphatase activity in rhizosphere soil (Spohn et al., 2015).
Soil sucrase is directly involved in the soil carbon cycle and is a key enzyme that catalyzes sucrose hydrolysis. It is commonly used as an indicator of soil carbon nutrition status (Chioti and Zervoudakis, 2017), and changes in its activity can reflect the activity of the soil carbon cycle (Li et al., 2005). Studies have shown that sucrase activity is significantly or extremely significantly correlated with most soil factors and the contents of nitrogen, phosphorus, and potassium in plants. In this study, we explored changes in sucrase activity in the rhizosphere soil of plants from the perspective of external environmental factors (light and water). The results indicated that there was no significant difference in sucrase activity in the rhizosphere soil of the two Quercus seedlings under different water and light conditions. However, the range of activity within each treatment group was large, which may be related to other external factors. This stability may be due to the regulation of the Q. dentata and Q. variabilis root systems, which maintain the pH and organic matter content of the rhizosphere soil within a relatively stable range, thereby ensuring the stability of sucrose content (Figure 2). Soil catalase is a key enzyme that can decompose soil hydrogen peroxide and is one of the key enzymes in the biological defense system, and its activity is closely related to soil respiration and microbial activities (Yang et al., 2019). In this study, it was found that light conditions had a significant effect on catalase activity, while water had no significant effect on it. The catalase activity of two kinds of Quercus seedlings was the highest under L3 (25% light) and L1 (75% light) treatment, respectively, which may be because light affects soil temperature and root secretion production (Figure 2). The change of temperature and secretion will affect the catalase activity in the soil, and the decrease of catalase activity means the decrease of the REDOX buffer capacity of the soil and the change of the REDOX state of the soil rhizosphere, which is not conducive to the absorption and transformation of plant nutrient elements. Some studies have shown that catalase activity increases under strong light and dark conditions, indicating that the antioxidant system is activated under high light stress (Gan et al., 2012), which is consistent with the results obtained in this study.
Soil cellulase (SCL) is a complex enzyme that acts as a biocatalyst in the decomposition of cellulose-containing litter (Xiao D. et al., 2015; Xiao Y. et al., 2015). Its activity is of significant scientific importance in characterizing the degree of soil maturation (Xu et al., 2022). Previous studies have indicated that a reduction in light intensity can influence soil enzyme activity by affecting soil moisture content (SMC), pH, and nutrient levels (Liu et al., 2022). By manipulating external environmental factors such as water and light, this study found that both factors had notable effects on the activity of cellulase in the rhizosphere soil of Q. dentata and Q. variabilis seedlings (Figure 2). Specifically, the differences in cellulase activity in the rhizosphere soil of Q. dentata seedlings were more pronounced under varying water and light conditions compared to Q. variabilis (Figure 2). Irrigation water can stimulate the growth and metabolism of soil microorganisms, thereby enhancing the activity of cellulase.
Chitin, a nitrogen-containing polysaccharide, is widely distributed in nature and is second only to cellulose in abundance (Shen, 2021). Chitinase decomposes chitin to release nutrients such as nitrogen, phosphorus, and potassium, which are beneficial for plant absorption and utilization. This process not only improves soil quality and fertility but also promotes plant growth and development, ultimately enhancing crop yields. Some studies have shown that soil water content is closely linked to enzyme activity, and optimal water content can enhance its activity (Li et al., 2018), which aligns with the findings of this study. Our study revealed that chitinase activity in the rhizosphere soil of both Quercus species was significantly influenced by light and water. As soil irrigation water decreased, soil moisture content diminished, inhibiting soil enzyme function and naturally reducing its activity (Zhu L. et al., 2021). This further suggests that chitinase thrives better in moist soil environments.
Our study demonstrated that soil enzymes exhibit optimal activity within a specific temperature range. Beyond this range, their activity may be weakened or even completely lost. Light and water, as crucial environmental factors, indirectly influence soil enzyme activity by modulating soil aeration, temperature, and the activity of soil microorganisms. Notably, rhizosphere soil enzyme activities of different plant species respond uniquely to external environmental factors. These findings not only underscore the sensitivity and adaptability of soil enzymes to environmental changes within the plant rhizosphere microenvironment but also offer valuable insights for further elucidating the role of soil enzymes in facilitating plant growth and development.
Research has indicated that numerous soil properties, including pH, salinity, temperature, and humidity, significantly influence the composition of microbial communities in intricate soil matrices (Frindte et al., 2019; Zheng et al., 2019). Furthermore, temporal variations such as weather patterns, root exudation, and seasonal inputs of organic matter can also affect the structure and activity of these microbial communities (Kuzyakov and Evgenia, 2015; Chernov and Zhelezova, 2020). Soil microbial communities play a crucial role in decomposing soil organic matter, enhancing nutrient storage conditions, and facilitating nutrient cycling through metabolic processes. These processes serve as direct indicators of soil microecological status and disease progression trends, and they represent sensitive early warning signals of soil ecosystem health (Dong et al., 2016; Zuppinger-Dingley et al., 2014). Notably, the diversity of soil microbial communities responds variably to environmental disturbances (Cai et al., 2020), aligning with the findings of the present study. Regarding the impact of water and light conditions on rhizosphere microbial community dynamics in seedlings, this study revealed that under varying water and light treatments, the microbial community richness, diversity, species abundance, and distribution uniformity of two Quercus species underwent significant alterations (Figures 3–5). Notably, there appears to be a substantial correlation between microbial diversity and soil enzyme activity, which is pivotal in maintaining ecosystem multifunctionality (Delgado-Baquerizo et al., 2017). In this study, fungi exhibited greater sensitivity to external light and water treatment conditions compared to bacteria. This may be attributed to the physiological adaptability and community structural differences between fungi and bacteria, resulting in their varied responsiveness to light and water. Fungi generally prefer dark and humid environments, whereas bacteria may exhibit broader adaptability (Figure 6). Consequently, changes in external light and water conditions may more profoundly affect the fungal community. Fungi are typically associated with the decomposition of C- and N-poor substrates, whereas bacteria are more sensitive to unstable substrates (Treseder et al., 2016; Xu et al., 2015). Intermediate decomposition products produced by fungi can serve as unstable materials for bacteria (Romaní et al., 2006). An increased abundance of microorganisms in the rhizosphere soil accelerates the decomposition rate of organic matter, providing more nutrients for seedling growth, thereby promoting their development.
Biodiversity is an intricate concept encompassing various components, such as species richness (the number of taxa) and composition (the identity and relative abundance of the organisms comprising the community) (Diaz and Cabido, 2001), among others. In this study, we observed that the fungal diversity in the rhizosphere soil of the two Quercus species was notably lower compared to bacterial diversity. Furthermore, the rhizosphere microbial communities of each sample exhibited high uniqueness and low similarity. At the phylum level, Ascomycota dominated the rhizosphere fungal communities of both Quercus seedlings, occupying a substantial proportion (Figure 7). Conversely, the rhizosphere bacterial communities were primarily composed of Proteobacteria, Actinobacteriota, and Acidobacteriota, with Proteobacteria being the most abundant. Whole genome sequencing (Trivedi et al., 2013) has provided insights into the distinct roles of Proteobacteria and Actinobacteriota in supporting crucial ecosystem processes like decomposition and nutrient cycling. Based on these findings, it can be deduced that Ascomycota and another fungal group play pivotal roles in the growth and development of plant seedlings. Notably, there were significant differences in the abundance of Paecilomyces, Streptomyces, and Mesorhizobium in the rhizosphere soil of Q. dentata and Q. variabilis, suggesting that these microorganisms are more sensitive to changes in light and water conditions compared to others.
Regarding the microbial community markers and species differences among samples, our findings revealed that the fungal composition in the rhizosphere soil of the two Quercus seedlings varied significantly under different treatments, albeit with fewer samples exhibiting differences compared to bacteria. Specifically, Ascomycetes, Trichospora, Microcomycetes, and Actinomycetes in the rhizosphere of Q. dentata, as well as Leotiomycetes, Acidomycetes, Burkella, Nitrosomonas, and other microflora in the rhizosphere of Q. variabilis, were notably distinct species. Notably, even seedlings of the same plant species exhibited significant differences in the dominant microbial species within their rhizosphere soil under varying light and water treatments. This could be attributed to microorganisms’ ability to adapt to different environments by modifying their metabolic pathways and growth strategies, thereby establishing dominance in the rhizosphere soil. For instance, as light conditions diminished, the abundance of rhizosphere bacteria and four physiological groups of bacterial grasses decreased, whereas the fungal population gradually increased, leading to a shift in the rhizosphere soil from a “bacterial-dominated” to a “fungal-dominated” type (Forey et al., 2015).
The results indicated that intense light hindered the growth of rhizosphere bacteria, whereas optimal light and humidity conditions fostered more vigorous rhizosphere bacterial community growth, suggesting that these environmental factors significantly influence rhizosphere bacterial flora (Hou et al., 2021; Zhang et al., 2022). Additionally, the rhizosphere bacteria of Q. variabilis exhibited greater drought tolerance, providing a survival advantage in arid environments. Although the rhizosphere bacteria of Q. dentata showed slightly less drought tolerance, their higher bacterial diversity implied more flexible adaptations to environmental stress. Furthermore, the increased water requirements of Q. variabilis under low light conditions likely helped mitigate the slower root growth resulting from insufficient light, emphasizing the crucial role of water in plant growth. On the other hand, fungal communities exhibited distinct responses to environmental conditions compared to bacteria. The fungal community was significantly impacted by light and water, and moderate shade treatment increased fungal species diversity (Yang D. et al., 2011), indicating fungi’s sensitivity to environmental changes. During the seedling stage, the types of bacteria in the rhizosphere microorganisms of Q. dentata and Q. variabilis showed some similarity under the dual influence of water and light, whereas the fungal types differed significantly (Figure 8). This might be attributed to the specific tendencies of plants at the seedling stage to demand and select microorganisms during their growth process. The variation in the abundance of rhizosphere microorganisms is influenced not only by microbial interactions but also by the external environment and the plant’s growth stage, highlighting the need to consider the role of multiple factors comprehensively when studying rhizosphere microorganisms.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
JL: Formal analysis, Writing – review & editing. KC: Data curation, Formal analysis, Methodology, Resources, Writing – review & editing. AY: Methodology, Project administration, Writing – review & editing. YY: Resources, Writing – review & editing. XZ: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Software, Writing – review & editing. CX: Resources, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This project is financially supported by the National Key R&D Program of China, reference number 2023YFF1304004-06. This work was supported by the grant from the National Natural Science Foundation of China, reference number 32271832.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Gen AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Berendsen, R., Pieterse, C., and Bakker, P. (2012). The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486. doi: 10.1016/j.tplants.2012.04.001
Birt, H. W. G., Courtney, L. T., Gordon, F. C., and Francisco, D. (2022). Root phenotypes as modulators of microbial microhabitats. Front. Plant Sci. 13:1003868. doi: 10.3389/fpls.2022.1003868
Brockett, B. F. T., Cindy, E. P., and Sue, J. G. (2012). Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven Biogeoclimatic zones in Western Canada. Soil Biol. Biochem. 44, 9–20. doi: 10.1016/j.soilbio.2011.09.003
Cai, Y., Liu, J. Y., Qin, J. L., Sun, S. G., Duan, S. S., and Xu, N. (2016). Effects of environmental factors on the growth and urease activity of the harmful dinoflagellate Prorocentrum donghaiense. Acta Ecol. Sin. 36, 1711–1718. doi: 10.5846/stxb201408251681
Cai, Y. J., Shen, J. P., Hong, J. D., Zhang, L. M., Zhang, C. J., and He, J. Z. (2020). Variation of soil nitrate and bacterial diversity along soil profiles in manure disposal maize field and adjacent woodland. J. Soils Sediments 20, 3557–3568. doi: 10.1007/s11368-020-02679-z
Cao, L., Chen, W. N., Hu, Z. H., and Leng, P. S. (2018). Effect of saline-alkali stress on the growth and photosynthesis characteristics of Quercus pedunculata seedlings. J. Beijing Agric. Univ. 33, 86–90. doi: 10.13473/j.cnki.issn.1002-3186
Cao, Y. J., Zhao, M. C., Zheng, C. Y., and Zhu, F. (2023). Rhizosphere microorganisms-mediated plant responses to drought stress. Chin. J. Eco-Agric. 31, 1330–1342. doi: 10.12357/cjea.20230127
Chen, C. F., Qin, L., Duan, Y. X., He, Y. T., Wang, P., Feng, Q. Y., et al. (2018). Effects of different management patterns on leaf functional traits and soil physicochemical properties of Mongolian oak secondary Forest. Acta Ecol. Sin. 38, 8371–8382. doi: 10.5846/stxb201801190146
Chen, L. L., Wang, L., Wu, Y. J., Chen, X. L., Kong, X., Gong, J. Y., et al. (2024). Molecular and microecological mechanisms of plant response to drought stress. Mol. Plant Breed., 1–15. Available at: http://kns.cnki.net/kcms/detail/46.1068.S.20230406.1634.006.html
Chernov, T. I., and Zhelezova, A. D. (2020). The dynamics of soil microbial communities on different timescales: A review. Eurasian Soil Sci. 53, 643–652. doi: 10.1134/S106422932005004X
Chioti, V., and Zervoudakis, G. (2017). Is root catalase a bifunctional catalase-peroxidase? Antioxidants. 6, 39–44. doi: 10.3390/antiox6020039
Cookson, W. R., Osman, M., Marschner, P., Abaye, D. A., Clark, I., Murphy, D. V., et al. (2007). Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol. Biochem. 39, 744–756. doi: 10.1016/j.soilbio.2006.09.022
Craine, J. M., Fiona, B., Michael, P., Nick, Z., Carl, M., and William, D. S. (2009). Grazing and landscape controls on nitrogen availability across 330 South African savanna sites. Austral Ecol. 34, 731–740. doi: 10.1111/j.1442-9993.2009.01978.x
Delgado-Baquerizo, M., Eldridge, D. J., Ochoa, V., Gozalo, B., Singh, B. K., and Maestre, F. T. (2017). Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecol. Lett. 20, 1295–1305. doi: 10.1111/ele.12826
Diaz, S., and Cabido, M. (2001). Vive la difference, plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 16, 646–655. doi: 10.1016/S0169-5347(01)02283-2
Dong, L. L., Jiang, X., Feng, G. Q., Li, X. W., and Chen, S. L. (2016). Soil bacterial and fungal community dynamics in relation to Panax Notoginseng death rate in a continuous cropping system. Sci. Rep. 6:31802. doi: 10.1038/srep31802
Dracup, M. N. H., Barrett-lennard, E. G., Greenway, H., and Robson, A. D. (1984). Effect of phosphorus deficiency on phosphatase activity of cell walls from roots of subterranean clove. J. Exp. Bot. 35, 466–480. doi: 10.1093/jxb/35.4.466
Forey, E., Sékou, F. M. C., and Matthieu, C. (2015). Flowering phenology of a herbaceous species (Poa annua) is regulated by soil Collembola. Soil Biol. Biochem. 90, 30–33. doi: 10.1016/j.soilbio.2015.07.024
Frindte, K., Pape, R., Werner, K., Löffler, J., and Knief, C. (2019). Temperature and soil moisture control microbial community composition in an Arctic–alpine ecosystem along Elevational and Micro-topographic gradients. ISME J. 13, 2031–2043. doi: 10.1038/s41396-019-0409-9
Gan, L. P., Ye, Y. X., Xiang, L. X., Zeng, X. L., and Chen, M. (2012). Effects of light intensity on pigment content and catalase activity of Pilea cadierei. J. South. Agric. 43, 1655–1659. doi: 10.3969/j:issn.2095-1191.2012.11.1655
Gao, Y. T., Yang, C. Y., Zhang, R., Lin, B. J., Dong, B., and Li, H. X. (2022). Effect of water stress on the carbon source metabolic activity of rhizosphere soil microorganisms in drip-irrigated vineyard. J. Irrig. Drain. 41, 9–18. doi: 10.13522/j.cnki.ggps.2021176
Ghezzehei, T. A., Benjamin, S., Chelsea, L. A., Nathaniel, A. B., and Asmeret, A. B. (2019). On the role of soil water retention characteristic on aerobic microbial respiration. Biogeosciences 16, 1187–1209. doi: 10.5194/bg-2018-265
Gu, X. Y., and Hu, Z. Y. (1994). A method for determining soil Chitinase activity. Soil Bull. doi: CNKI:SUN:TRTB.0.1994-06-014
Guan, S. Y. (1986). Soil enzymes and their research methods. Beijing: Agriculture Publishing House, 294–297.
Han, Y. L., Yin, J., Yang, Z., Fan, J. Y., and Li, S. (2024). Effects of plant nutrients (compound oligosaccharides) on the growth and soil enzyme activity of drip irrigation fruit tomatoes. Water Sav. Irrig. 6:121-1271-14. doi: 10.12396/jsgg.2023469
Hao, H., Cao, L., Chen, W. N., Hu, Z. H., and Leng, P. S. (2020). Effects of salt stress on the ion balance and physiological-biochemical characteristics of Quercus dentata seedlings. Acta Ecol. Sin. 40, 6897–6904. doi: 10.5846/stxb201906211314
Hou, S. J., Thorsten, T., Nathan, V., Fantin, M., Jörg, Z., Brigitte, P., et al. (2021). A microbiota–root–shoot circuit Favours Arabidopsis growth over defence under suboptimal light. Nat. Plants 7, 1078–1092. doi: 10.1038/s41477-021-00956-4
Hu, H. Y., Guo, W., Wei, N., and Xing, L. W. (2024). Effects of reduced chemical fertilizer combined with humic acid on soil properties, nutrient activity of corn plants, and dry matter accumulation. Jiangsu Agric. Sci. 52, 232–240. doi: 10.15889/j.issn.1002-1302.2024.01.0
Huang, W. J., and Li, M. (2024). Effects of Pogostemon cablin-Mentha haplocalyx rotation and continuous cropping of Pogostemon cablin on its quality and rhizosphere soil microecology. Southwest China J. Agric. Sci. 37, 276–285. doi: 10.16213/j.cnki.scjas.2024.2.006
Huang, Y. Y., Peng, X. H., Ou, G. N., Peng, X. X., Gan, L., Huang, Y. H., et al. (2024). Effects of continuous cropping of cassava on the succession of fungal community structure in rhizosphere and non-rhizosphere soils. Guihaia., 1–13. Available at: http://kns.cnki.net/kcms/detail/45.1134.Q.20240508.1057.004.html
Huang, J. H., Tang, J. C., Li, G. L., and Huang, Q. Z. (2022). Beijing will plant over 10 million native oak trees (holes) by 2025-1 million oak seedlings have taken root in Beijing. Greening Life 12, 4–8.
Jang, S., Nooruddin, B. S., Muhammad, H., Jehyeong, J., Taeksung, L., Jung-Seok, Y., et al. (2020). Silicon foliage spraying improves growth characteristics, morphological traits, and root quality of Panax Ginseng C.A. Mey. Ind. Crops Prod. 156:112848. doi: 10.1016/j.indcrop.2020.112848
Kuzyakov, Y., and Evgenia, B. (2015). Microbial hotspots and hot moments in soil: Concept & Review. Soil Biol. Biochem. 83, 184–199. doi: 10.1016/j.soilbio.2015.01.025
Li, Z. G., Luo, Y. M., and Teng, Y. (2008). Methods of soil and environmental microbiology research. Beijing: Science Press.
Li, N., Wang, J. F., Gong, J. Y., Chen, L. L., Chen, X. L., Wang, L., et al. (2025). Research Progress on physiological molecular and rhizosphere microecological mechanisms of plant response to cadmium stress. Jiangsu Agric. Sci., 1–11. Available at: https://ir.lzu.edu.cn/handle/262010/588289
Li, D. P., Wu, Z. J., Chen, L. J., Zhu, P., and Ren, J. (2005). Dynamics of invertase activity of black soil treated by a long-term located fertilization and its influence. Chin. J. Eco-Agric. 13, 102–105. Available at: http://www.ecoagri.ac.cn/en/article/id/2005234
Li, H. Y., Zhang, J. G., Yao, T., Yang, X. M., Gao, Y. M., Li, C. N., et al. (2018). Soil nutrients, enzyme activity, and ecological stoichiometric characteristics of degraded alpine grassland. J. Soil Water Conserv. 32, 287–295. doi: 10.13870/j.cnki.stbcxb.2018.05.045
Li, J. K., Zhu, L. Q., Huang, R. Z., Liu, P. Y., Wang, J. P., Fang, H. Y., et al. (2024). Effects of altered carbon input on soil chemical properties and microbial communities in subtropical coniferous and broad-leaved mixed forests. J. Agro-Environ. Sci., 1–11. Available at: http://www.aes.org.cn/nyhjkxxb/ch/reader/view_abstract.aspx?flag=2&file_no=202402180000002&journal_id=nyhjkxxb
Lian, Y. H., Shi, R. Y., Liu, J. Z., Aurang, Z., Wang, Q., Wang, J. L., et al. (2024). Effects of polystyrene, polyethylene, and polypropylene microplastics on the soil-rhizosphere-plant system: Phytotoxicity, enzyme activity, and microbial community. J. Hazard. Mater. 465:133417. doi: 10.1016/j.jhazmat.2023.133417
Lin, F., Wu, X., Dini-Andreote, A. F., and Wang, B. B. (2020). Changes in bulk soil affect the disease-suppressive rhizosphere microbiome against Fusarium wilt disease. Front. Agric. Sci. Eng. 7:307. doi: 10.15302/J-FASE-2020328
Liu, P. (2020). Study on hybridization and seed dispersal of three oak species in Beijing area. Beijing: Beijing Forestry University.
Liu, Y. (2023). The impact of sugarcane root rot on rhizosphere microecology and screening and identification of antagonistic Bacteria of pathogenic Bacteria. Guangxi University. doi: 10.27034/d.cnki.ggxiu.2022.001852
Liu, Q., Liu, R., Zhang, L. S., Chen, F. S., Guo, C. L., Wang, F. C., et al. (2022). Effects of fertilization and light on soil enzyme activity and its stoichiometric ratio in Liquidambar formosana cultivation. J. Jiangxi Agric. Univ. 44:12. doi: 10.13836/j.jjau.2022014
Liu, P. F., Wang, X. Q., Zhang, M. X., Zhang, W., Ma, J. N., Yu, J., et al. (2024). Effects of different rotation modes on the growth of Platycodon grandiflorum, rhizosphere soil microorganisms, and soil enzyme activities. Biol. Res. 46, 126–133. doi: 10.14188/j.ajsh.2024.02.003
Lu, Q., and Li, D. Q. (2020). Research progress on soil enzymes and their ecological indication function. J. Anhui Agric. Sci. 48, 14–17.
McDaniel, M. D., Kaye, J. P., and Kaye, M. W. (2013). Increased temperature and precipitation had limited effects on soil extracellular enzyme activities in a post-harvest Forest. Soil Biol. Biochem. 56, 90–98. doi: 10.1016/j.soilbio.2012.02.026
Ncise, W., Daniels, C. W., and Nchu, F. (2020). Effects of light intensities and varying watering intervals on growth, tissue nutrient content and antifungal activity of hydroponic cultivated Tulbaghia violacea L. under greenhouse conditions. Heliyon 6:e03906. doi: 10.1016/j.heliyon.2020.e03906
Preece, C., and Penuelas, J. (2016). Rhizodeposition under drought and consequences for soil communities and ecosystem resilience. Plant Soil 409, 1–17. doi: 10.1007/s11104-016-3090-z
Qi, Y. L., Meng, K., Wu, Z. Y., Si, H. Y., Liu, K. L., Han, Y., et al. (2024). Dynamics of female flower and seed growth and development in Quercus dentata. J. Beijing For. Univ. 46, 17–26. doi: 10.12171/j.1000-1522.20220505
Quero, J. L., Villar, R., Marañón, T., and Zamora, R. (2006). Interactions of drought and shade effects on seedlings of four Quercus species: physiological and structural leaf responses. New Phytol. 170, 819–834. doi: 10.1111/j.1469-8137.2006.01713.x
Rasche, F., Knapp, D., Kaiser, C., Koranda, M., Kitzler, B., Zechmeister-Boltenstern, S., et al. (2011). Seasonality and resource availability control bacterial and archaeal communities in soils of a temperate beech forest. ISME J. 5, 389–402. doi: 10.1038/ismej.2010.138
Richter, A. S., Schöning, I., Kahl, T., Bauhus, J., and Ruess, L. (2018). Regional environmental conditions shape microbial community structure stronger than local forest management intensity. For. Ecol. Manag. 409, 250–259. doi: 10.1016/j.foreco.2017.11.027
Romaní, A. M., Fischer, H., Mille-Lindblom, C., and Tranvik, L. J. (2006). Interactions of bacteria and fungi on decomposing litter: differential extracellular enzyme activities. Ecology 87, 2559–2569. doi: 10.1890/0012-9658(2006)87[2559:IOBAFO]2.0.CO;2
Rousk, J., Bååth, E., Brookes, P. C., Lauber, C. L., Lozupone, C., Caporaso, J. G., et al. (2010). Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 4, 1340–1351. doi: 10.1038/ismej.2010.58
Shen, Y. Y. (2021). Screening, identification and enzyme production characteristics of Chitinase-producing Bacteria in Cicada flower soil. Tianjin University of Commerce. doi: 10.27362/d.cnki.gtsxy.2021.000432
Shen, X. T., Tu, B. H., Liu, Z. Y., Zhao, Y., Ding, J. X., and Liang, Y. T. (2025). Mechanism of rhizosphere microorganisms regulating plant root architecture in acidic adverse soils. Environ. Sci. 46, 570–578. doi: 10.13227/j.hjkx.202312260
Shi, X. S., Reyimu, T., Yu, Z. W., Wumaieriang, S., and Liu, G. S. (2024). Effect of grazing on the bacterial characteristics of mountainous meadow soil. Agronomy 3, 8–16. doi: 10.11829/j.issn.1001-0629.2022-0673
Siebielec, S., Siebielec, G., Klimkowicz-Pawlas, K., Gałazka, A., Grzadziel, J., and Stuczynski, T. (2020). Impact of water stress on microbial community and activity in sandy and loamy soils. Agronomy 10:1429. doi: 10.3390/agronomy10091429
Song, K. M., Jiao, X. Z., Li, L., and Yan, J. Q. (1999). Relationship between the changes of acid phosphatase activities and pi uptake of tomato seedlings during phosphate starvation. Acta Bot. Yunnanica 21, 101–108. Available at: https://europepmc.org/article/CBA/325373
Spohn, M., Treichel, N. S., Cormann, M., Schloter, M., and Fischer, D. (2015). Distribution of phosphatase activity and various bacterial phyla in the rhizosphere of Hordeum vulgare L. depending on p availability. Soil Biol. Biochem. 89, 44–51. doi: 10.1016/j.soilbio.2015.06.018
Sun, J. Q., Cao, W. Q., Leng, P. S., and Hu, Z. H. (2021). Effects of inoculating four ectomycorrhizal Fungi on the growth, photosynthesis, and nutrient element activity of Quercus aliena var. Chelsea seedlings. J. Cent. South Univ. For. Technol. 41, 67–74. doi: 10.14067/j.cnki.1673-923x.2021.10.008
Sun, X. T., Li, L., Long, G. Q., Zhang, G. H., Meng, Z. G., Yang, S. C., et al. (2015). Research progress on obstacles to continuous cropping of Panax notoginseng. Chin. J. Ecol. 34, 885–893. doi: 10.13292/j.1000-4890.2015.0119
Sun, J. Y., Wang, Q., Liu, J. F., Chang, E. M., and Jia, Z. R. (2023). Geographic variation and influencing factors of cork thickness of Quercus variabilis. J. Beijing For. Univ. 45, 11–18. doi: 10.12171/j.1000-1522.20230029
Tan, Q. (2024). Effects of application of different types of sewage sludge on soil enzyme activities. Clean. World 40, 65–67. Available at: https://csti.cqvip.com/Qikan/Article/Detail?id=7111919845
Terhonen, E., Blumenstein, K., Kovalchuk, A., and Asiegbu, F. O. (2019). Forest tree microbiomes and associated fungal endophytes: functional roles and impact on forest health. Forests 10:42. doi: 10.3390/f10010042
Tiwari, P., Bose, S. K., and Bae, H. (2021). Plant growth-promoting soil microbiomes: beneficial attributes and potential applications. Sustainable Dev. Biodivers. 27, 1–30. doi: 10.1007/978-3-030-73507-4_1
Treseder, K. K., Marusenko, Y., Romero-Olivares, A. L., and Maltz, M. R. (2016). Experimental warming alters potential function of the fungal community in boreal forest. Glob. Chang. Biol. 22, 3395–3404. doi: 10.1111/gcb.13238
Trivedi, P., Anderson, I. C., and Singh, B. K. (2013). Microbial modulators of soil carbon storage, integrating genomic and metabolic knowledge for global prediction. Trends Microbiol. 21, 641–651. doi: 10.1016/j.tim.2013.09.005
Wang, J. H., and Huang, X. Q. (2003). Analysis of the ecological characteristics and causes of red tide of Protozoa with Denticles. Chin. J. Appl. Ecol. 14, 1065–1069. Available at: https://www.cjae.net/CN/abstract/abstract8718.shtml
Wang, Z., Ou, Q. H., Feng, Y. J., Wu, B., Zhao, Q. B., Deng, Y., et al. (2024). Effects of biostimulants on rhizosphere microecology and leaf quality of flue-cured tobacco. J. Northwest A&F Univ. (Nat. Sci. Ed). 52, 59–68. doi: 10.13207/j.cnki.jnwafu.2024.10.007
Wang, N., Wang, J. Q., Li, G. L., Li, J., Zhu, L., Li, T., et al. (2022). Germination and seedling characteristics of Quercus variabilis and physiological and biochemical changes. For. Sci. 58, 1–10. doi: 10.11707/j.1001-7488.20220401
Wu, Y. H., Xue, X. P., You, H. Z., Chen, B. C., Chen, F. C., and Yu, W. W. (2022). The response of Quercus variabilis seedlings to different light intensities. Tianjin Agric. Sci. 28, 1–5. Available at: https://www.fx361.cc/page/202211026/10704169.shtml
Xiao, D., Hao, Y., Yang, X., Sun, X. L., Wang, Y. B., and Liu, S. Y. (2015). Accumulation characteristics and correlation analysis of five Ginsenosides with different cultivation ages from different regions. J. Ginseng Res. 39, 338–344. doi: 10.1016/j.jgr.2015.03.004
Xiao, Y., Huang, Z. G., and Lu, X. G. (2015). Changes of soil labile organic carbon fractions and their relation to soil microbial characteristics in four typical wetlands of Sanjiang plain, Northeast China. Ecol. Eng. 82, 381–389. doi: 10.1016/j.ecoleng.2015.05.015
Xie, J. T., Lou, X. Q., Lu, Y. Z., Huang, H., Yang, Q., Zhang, Z. P., et al. (2023). Suitable light combinations enhance cadmium accumulation in Bidens Pilosa L. by regulating the soil microbial communities. Environ. Exp. Bot. 205:105128. doi: 10.1016/j.envexpbot.2022.105128
Xu, G., Chen, J., Berninger, F., Pumpanen, J., Bai, J. W., Yu, L., et al. (2015). Labile, recalcitrant, microbial carbon and nitrogen and the microbial community composition at two Abies faxoniana forest elevations under elevated temperatures. Soil Biol. Biochem. 91, 1–13. doi: 10.1016/j.soilbio.2015.08.016
Xu, M. P., Liu, H. Y., Zhang, Q., Zhang, Z. J., Ren, C. J., Feng, Y. Z., et al. (2022). Effect of forest thinning on soil organic carbon stocks from the perspective of carbon-degrading enzymes. Catena 218:106560. doi: 10.1016/j.catena.2022.106560
Yang, D., Duan, L. S., Xie, H. A., Li, Z. H., and Huang, T. X. (2011). Effect of pre-flowering light deficiency on the ecological effects of root zone soil microorganisms in Rice. Sci. Technol. Rev. 19, 347–352. doi: 10.3724/SP.J.1011.2011.00347
Yang, S., Yao, F., Ye, J., Fang, F., Wang, Z. R., Wang, R. Z., et al. (2019). Latitudinal pattern of soil lignin/cellulose content and the activity of their degrading enzymes across a temperate forest ecosystem. Ecol. Indic. 102, 557–568. doi: 10.1016/j.ecolind.2019.03.009
Yang, L. F., Zeng, Q., and Li, H. B. (2011). Measurement of soil catalase activity by ultraviolet spectrophotometry. Soil Bull. 2, 207–210. doi: 10.19336/j.cnki.trtb.2011.01.043
Yin, H., Luo, Z. E., Jin, Z. F., Luo, Q., Tan, R. X., Luo, W., et al. (2024). Effects of LW-1, a dominant fungus in rice-fish coculture farmland, on soil alkaline nitrogen, available phosphorus, soil enzyme activity, and cadmium bioavailability. Biol. Res. 46, 154–163. doi: 10.14188/j.ajsh.2024.02.006
Yuan, H. G., and Zhang, S. B. (2024). Impact of different modifiers on the structure of soil bacterial Community in red soil restoration forest land. Subtrop. Soil Water Conserv. 36, 1–5. doi: 10.3969/j.issn.1002-2651.2024.02.001
Zhang, X. C., Bilyera, N., Fan, L. C., Duddek, P., Ahmed, M. A., Carminati, A., et al. (2022). The spatial distribution of rhizosphere microbial activities under drought: water availability is more important than root-hair-controlled exudation. New Phytol. 237, 780–792. doi: 10.1111/nph.18409
Zhang, J., Cao, Y., Liu, J. H., Yang, J. T., Que, Y. X., You, C. H., et al. (2023). The impact of smut on sugarcane rhizosphere microecology and screening of antagonistic Bacteria. Chinese Society for Plant Pathology. China Agricultural Science and Technology Press. doi: 10.26914/c.cnkihy.2023.038691
Zhang, L., Ding, X. D., Chen, S. F., He, X. H., Zhang, F. S., Feng, G., et al. (2014). Reducing carbon: phosphorus ratio can enhance microbial phytin mineralization and lessen competition with maize for phosphorus. J. Plant Interact. 9, 850–856. doi: 10.1080/17429145.2014.977831
Zhang, X., Zhang, W. H., Jiang, M., Leng, P. S., and Hu, Z. H. (2024). Effect of shading treatment on the growth and physiology of Quercus seedlings. J. Beijing Agric. College. 39, 90–96. doi: 10.13473/j.cnki.issn.1002-3186.2024.0117
Zhao, K., Ma, B., and Xu, Y. (2021). Light exposure mediates circadian rhythms of rhizosphere microbial communities. ISME J. 15, 2655–2664. doi: 10.1038/s41396-021-00957-3
Zhao, J. B., Feng Zhi, P., Zong, W. Y., Wang, T., and Yang, X. T. (2025). Progress in the regulation mechanism of seedling growth-defense trade-off in Forest regeneration. J. Henan Agric. Univ. 59, 13–20. doi: 10.16445/.cnki.1000-2340.20240523.001
Zhao, L. N., Liu, Y. B., Wang, Z. R., Yuan, S. W., Qi, J. H., Zhang, W. L., et al. (2020). Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems. Plant Soil 447, 379–392. doi: 10.1007/s11104-019-04391-5
Zhao, X. W., Wu, N. L., Qiao, J. H., Li, X. K., Han, Y. H., Xing, G. F., et al. (2020). Identification of acid phosphatase-encoding family genes in millet and analysis of low phosphorus tolerance haplotype of SiACp1. Acta Agric. Bor.-Sin. 35, 35–45. Available at: https://jglobal.jst.go.jp/en/detail?JGLOBAL_ID=202002212858218169
Zheng, Q., Hu, Y. T., Zhang, S. S., Noll, L., Böckle, T., Dietrich, M., et al. (2019). Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil Biol. Biochem. 136:107521. doi: 10.1016/j.soilbio.2019.107521
Zhou, J., and Lei, T. (2007). Influencing factors and research methods of soil microbial diversity: status quo and prospect. Biodivers. Sci. 15, 306–311. doi: 10.1360/biodiv.070069
Zhu, L., Li, Y., Yang, W. Q., and Gao, Y. H. (2021). Effects of desertification on soil carbon, nitrogen, enzyme activity, and bacterial diversity in alpine grassland. J. Soil Water Conserv. 35, 350–358. doi: 10.13870/j.cnki.stbcxb.2021.03.048
Zhu, Y. G., Peng, J. J., Wei, Z., Shen, Q. R., and Zhang, F. S. (2021). Soil microbiome and soil health. Sci. China Life Sci. 51, 1–11. Available at: https://cstj.cqvip.com/Qikan/Article/Detail?id=7104049188
Zuccarini, P., Sardans, J., Asensio, L., and Peñuelas, J. (2023). Altered activities of extracellular soil enzymes by the interacting global environmental changes. Glob. Chang. Biol. 29, 2067–2091. doi: 10.1111/gcb.16604
Keywords: light, soil moisture, Quercus variabilis, Quercus dentata, rhizosphere microecology
Citation: Lian J, Cai K, Yin A, Yuan Y, Zhang X and Xu C (2025) Both light and soil moisture affect the rhizosphere microecology in two oak species. Front. Microbiol. 16:1506558. doi: 10.3389/fmicb.2025.1506558
Edited by:
Xingjia Xiang, Anhui University, ChinaReviewed by:
Mika Tapio Tarkka, Helmholtz Association of German Research Centres (HZ), GermanyCopyright © 2025 Lian, Cai, Yin, Yuan, Zhang and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xinna Zhang, emhhbmd4aW5uYUBiamZ1LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.