
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 11 December 2024
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1467414
 Shilin Zhong1,2,3
Shilin Zhong1,2,3 Bingcai Yang1,2,3Yuzhen Liu1,2,3
Bingcai Yang1,2,3Yuzhen Liu1,2,3 Wenkui Dai1,2,3Guanglei Li4Juan Yang1,2,3Ao Yang1,2,3Ying Wang1,2,3Min Wang1,2,3Chang Xu5
Wenkui Dai1,2,3Guanglei Li4Juan Yang1,2,3Ao Yang1,2,3Ying Wang1,2,3Min Wang1,2,3Chang Xu5 Yuqing Deng1,2,3*
Yuqing Deng1,2,3*Introduction: Gut microbiota (GM) has been implicated in gestational diabetes mellitus (GDM), yet longitudinal changes across trimesters remain insufficiently explored.
Methods: This nested cohort study aimed to investigate GM alterations before 24 weeks of gestation and their association with GDM. Ninety-three Chinese participants provided fecal samples during the first and second trimesters. Based on oral glucose tolerance tests, 11 participants were classified as GDM, and 82 as non-diabetic (ND). Using 16S rRNA sequencing, we analyzed both cross-sectional and longitudinal differences in GM structure between those two groups.
Results: In the first trimester, GDM group exhibited lower levels of Bacteroides_H and Acetatifactor compared to ND group (p < 0.05). In the second trimester, GDM individuals showed increased abundance of Fusobacteriota and Firmicutes_D, and genera including Fusobacterium_A and Fournierella, while Anaerotruncus and others decreased (P<0.05). Inflammation-associated genera like Gemmiger_A_73129 and Enterocloster increased, while Megamonas decreased in overweight or obese GDM women, which was not identified in normal-weight women. The ratios of relative abundance of genera Streptococcus, Enterocloster, and Collinsella exceeded 1.5 in the GDM group, particularly in overweight or obese individuals. Inflammatory pathways related to African trypanosomiasis and Staphylococcus aureus infection were predicted to be up-regulated in overweight or obese GDM individuals but not in normal-weight GDM women.
Discussion: This study suggests that GM of women with GDM undergoes significant alterations between the first and second trimesters, potentially linked to inflammation, with more pronounced changes observed in overweight or obese individuals.
Gestational diabetes mellitus (GDM) is defined as diabetes diagnosed in the second or third trimester of pregnancy that was not clearly overt diabetes prior to gestation (American Diabetes Association, 2020). As a prevalent obstetric complication, GDM affects over one in six pregnant women globally (He et al., 2023). This not only leads to adverse maternal outcomes such as primary cesarean delivery and preeclampsia (Catalano et al., 2012), but also impacts fetal outcomes, including hypoglycemia, macrosomia, and preterm delivery (Domanski et al., 2018). Furthermore, GDM has long-term health implications for both mothers and their offspring (Bianco and Josefson, 2019). Women with GDM face a significantly increased risk of metabolic diseases, including type 2 diabetes mellitus (T2DM) (Sheiner, 2020) and obesity. Their offspring also have a heightened risk of cardiovascular alterations (Di Bernardo et al., 2023) and diabetes (Sweeting et al., 2022; Mitanchez et al., 2015). Prenatal exposure to untreated GDM is a significant and independent risk factor for impaired glucose tolerance (IGT) in childhood (Lowe et al., 2019). Notably, GDM treatment may not substantially reduce adverse metabolic outcomes in children (Bianco and Josefson, 2019). With China’s recent fertility policy removing fertility restrictions and potentially increasing the proportion of older and multiparous women, the incidence of GDM could rise (Wang D. et al., 2024). Therefore, it is crucial to explore GDM risk factors in early pregnancy and implement preventative measures to manage its occurrence.
Increasing evidence suggests that gut microbiota (GM) plays a critical role in regulating glucose metabolism homeostasis (Palmnäs-Bédard et al., 2022). Dysbiosis in GM contributed to glucose intolerance and insulin resistance, which were linked to T2DM (Gurung et al., 2020) and GDM (Medici Dualib et al., 2021; Balleza-Alejandri et al., 2024). Mechanisms underlying those links include low-grade endotoxemia due to increased gut permeability (Ye et al., 2023), an imbalance in the production of short-chain fatty acids (Bielka et al., 2022) and branched-chain amino acids (Pedersen et al., 2016), as well as disruptions in bile acid metabolism (Cai et al., 2022). GM alterations may serve as biomarkers for diagnosing glucose metabolism abnormalities and, more importantly, as potential targets for GDM intervention (Crudele et al., 2023; Li H.-Y. et al., 2021; Hu et al., 2023). GDM may also alter the structure of breastmilk microflora, which may affect the health of offspring (Gámez-Valdez et al., 2021). Thus, examining GM characteristics and markers in women with GDM between the first trimester (FT) and second trimester (ST) could aid in diagnosing, treating, or preventing GDM.
Recently, several studies have examined the structure and diversity of GM in pregnant women with GDM (Chen F. et al., 2021; Chen T. et al., 2021; Hu et al., 2021; Zheng et al., 2020). Several differentially abundant taxa have been identified between the GDM and control groups at both genus and species levels (Chen F. et al., 2021). Changes in GM compositions during the second trimester, prior to GDM diagnosis, are linked to fasting serum metabolite levels (Chen T. et al., 2021). GM dynamics in early pregnancy differed significantly between GDM and normoglycemic women (Hu et al., 2021), suggesting potential as early biomarkers for GDM (Zheng et al., 2020). Beneficial gut microorganisms were inversely associated with GDM, while opportunistic pathogens increased GDM risk, correlating with higher OGTT glucose levels (Hu et al., 2021).
However, significant inconsistencies existed among these studies (Wang S. et al., 2024). For instance, Streptococcus decreased in one study (Zheng et al., 2020) but increased in another (Zhang et al., 2021), and Lachnospiraceae exhibited opposite trends (Hu et al., 2021; Ma et al., 2020). Such heterogeneity may stem from variations in study population characteristics, gestational age at sampling, sequencing methods, and analysis techniques, which pose challenges in cross-sectional comparisons. Importantly, only a few studies have investigated GM changes from the first to the second trimester in women with GDM (Zheng et al., 2020; Sun et al., 2023). Additionally, obesity or overweight is also significantly associated with gut microbiota (Afzaal et al., 2022), and there is a complex interaction between obesity/overweight and GDM. Therefore, a longitudinal cohort study with matching and stratification based on initial BMI is warranted to reduce the effect of confounding factors.
This nested cohort study from China aims to examine the gut microbiota characteristics in women with GDM during the first trimester (FT) and second trimester (ST). It also seeks to explore the correlation between longitudinal changes in gut microbiota and GDM, utilizing the variance of the relative abundance (VRA) and ratio of the relative abundance (RRA) indices. Additionally, stratified analysis based on the initial BMI was conducted. The findings from this study are anticipated to contribute valuable insights for the prediction, prevention, and management of GDM.
This nested cohort study was conducted at Peking University Shenzhen Hospital from April 2023 to November 2023. Participants were eligible if they were aged 18–45 years, had a gestational age less than 14 weeks at enrollment, and provided informed consent. Exclusion criteria comprised pre-pregnancy diabetes, hypertension, intrahepatic cholestasis of pregnancy, chronic diarrhea, acute or chronic infectious diseases, and use of antibiotics/probiotics within the last 3 months. Participants diagnosed with GDM were assigned to the GDM group, while those without GDM were placed in the non-diabetic (ND) group (Figure 1). Based on the BMI in the first trimester, participants were further categorized into normal weight and overweight/obese subgroups within GDM and ND groups. According to a previous Chinese study, 14 cases of GDM were sufficient to demonstrate significant GM differences in the second trimester between GDM and control groups (Li et al., 2023). Given the 15% GDM incidence in China (Juan and Yang, 2020), a sample size of 93 participants were calculated. Accounting for a 15% dropout rate, a total of 110 women were recruited. This study received approval from the Ethics Committee of Peking University Shenzhen Hospital (No. 2022-007).
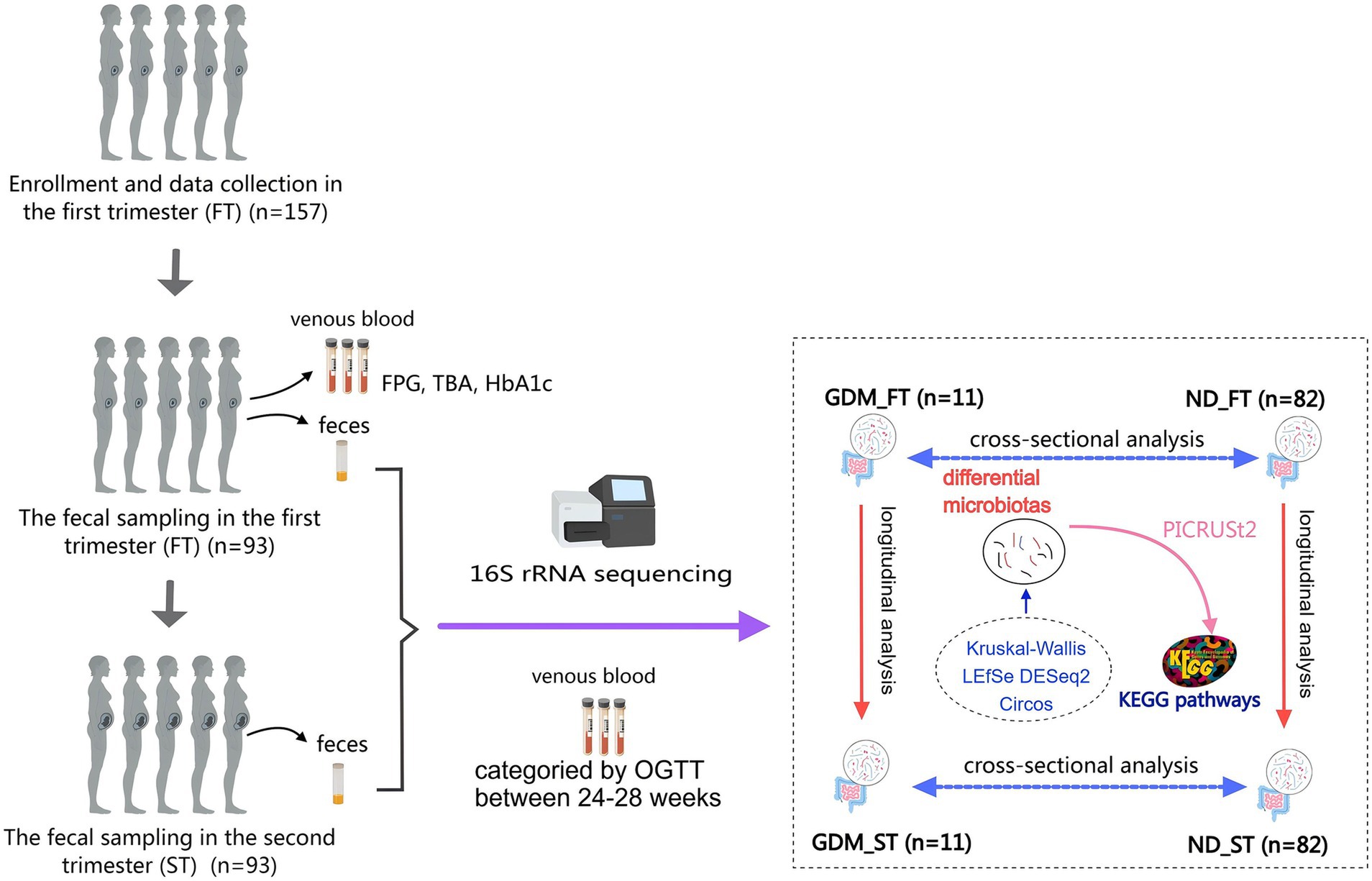
Figure 1. The flow chart of this study. GDM, gestational diabetes mellitus; ND, non-diabetics; HbA1c, glycated hemoglobin A1c; OGTT, oral glucose tolerance test; TBA, total bile acid; LEfSe, Linear discriminant analysis Effect Size; PICRUSt, Phylogenetic Investigation of Communities by Reconstruction of Unobserved States; KEGG, Kyoto Encyclopedia of Genes and Genomes.
A 5-mL venous blood sample was collected from all participants after a 10-h fast between 11–13 weeks and 6 days of gestation. This sample was used to measure fasting plasma glucose (FPG), total bile acids, and glycosylated hemoglobin levels. During 24–28 weeks of gestation, a 75 g oral glucose tolerance test (OGTT) was performed to determine the presence of GDM. Venous blood samples were taken at fasting (0-h), 1-h, and 2-h after consuming a 75 g glucose solution. FPG, 1-h blood glucose, and 2-h blood glucose levels during the OGTT were measured using the hexokinase method with a Glucose assay kit on a Beckman AU5800 analyzer (Agencourt, Beckman Coulter, Brea, CA, United States). GDM was diagnosed according to the criteria of the International Association of Diabetes and Pregnancy Study Group (IADPSG) (He et al., 2022), where OGTT 0-h blood glucose ≥5.1 mmol/L, 1-h blood glucose ≥10.0 mmol/L, or 2-h blood glucose ≥8.5 mmol/L indicated GDM. HbA1c was measured using the high-performance liquid chromatography (HPLC) method on a Glycosylated hemoglobin analyzer D-100 (Bio-Rad Laboratories, Berkeley, CA, United States) with an HbA1c Kit. Total bile acids (TBA) were examined using the Total Bile Acid Assay Kit (Leadman bio, Beijing, China) with the enzymatic cycling method on a Beckman AU5800 analyzer.
Body weight was recorded in the first trimester (11–14 weeks) and second trimester (21–24 weeks), with BMI calculated as weight (kg) divided by height (m2). BMI in the first trimester between 18.5 and 24 kg/m2 was considered normal weight, while BMI ≥ 24 kg/m2 indicated overweight or obesity. Data on maternal age, ethnicity, parity, occupation, and education were also collected and compared between the GDM and ND groups.
Fecal samples were collected using a disposable sampling kit (Miraclean Tech Co., Ltd., Shenzhen, China) during the first trimester (11 weeks to 13 weeks and 6 days) and the second trimester (21 week to 23 weeks and 6 days). Over 20 grams of feces were collected under aseptic conditions and immediately placed into cryotubes containing preservation solution. Samples were stored at −80°C within 24 h and transported to the laboratory (Wekemo Tech Group Co., Ltd., Shenzhen, China) for subsequent testing. During transport, the samples were kept on dry ice.
Total fecal genomic DNA was extracted from approximately 250 mg-500 mg of feces using Magnetic Stool DNA Kit (Cat# DP712, TIANGEN Biotech Co. Ltd., Beijing, China), following the manufacturer’s instructions. After concentrating and purifying the DNA, primers 515F (5’-GTGCCAGCMGCCGCGGTAA-3′) and 907R (5’-CCGTCAATTCCTTTGAGTTT-3′) were used to amplify the V4 to V5 regions of 16S rRNA genes via PCR. PCR products were purified using a Universal DNA Kit (Cat# DP214, TianGen, China). Sequencing libraries were generated using the NEBNext® Ultra DNA Library Prep Kit (Cat# E7370L, Illumina, San Diego, CA, United States), following the manufacturer’s guidelines. Library quality was assessed using the Agilent 5400 system (Agilent Technologies Co. Ltd., United States). Sequencing was performed on an Illumina platform, producing 250 bp paired-end reads.
The analysis followed the QIIME2 tutorial.1 Raw FASTQ files were imported into a QIIME2-compatible format using the QIIME tools import program. Sequences from each sample were demultiplexed and filtered for high quality, trimmed, and de-noised. Chimeric sequences were identified and removed using the QIIME2 dada2 plugin to obtain the amplicon sequence variant (ASV) feature table. ASV sequences were aligned to the pre-trained Greengenes2 database2 (McDonald et al., 2024) using the QIIME2 feature-classifier plugin to generate a taxonomy table. Contaminating mitochondrial and chloroplast sequences were filtered out. Relative abundance (RA) was used to reflect the proportion of a certain genus or phylum in the total microbiota, with bacterial flora having RA > 0.1% defined as dominant flora. Kruskal-Wallis, Linear discriminant analysis Effect Size (LEfSe), DESeq2, and Circos analysis were applied to identify bacteria with different RA among groups (Figure 1). Diversity metrics were calculated using the core-diversity plugin within QIIME2. Alpha diversity was assessed using the Chao1 and Shannon indices, while beta diversity was investigated using nonmetric multidimensional scaling (NMDS) and principal coordinate analysis (PCoA). Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt) (Langille et al., 2013) predicted potential functional Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways associated with changes in gut microbiota composition in the GDM group (Figure 1). Samples were divided into first-trimester GDM (GDM_FT), first-trimester ND (ND_FT), second-trimester GDM (GDM_ST), and second-trimester ND (ND_ST) groups for microbiota abundance comparison.
In this study, variance of the relative abundance (VRA) and ratio of the relative abundance (RRA) were used to analyze microbiota differences between the first and second trimesters in the GDM and ND groups. VRA was defined as the difference in RA between the second and first trimesters for a given bacterial flora, with the average VRA compared between the GDM and ND groups. RRA was calculated as the ratio of the mean RA in the second trimester to the mean RA in the first trimester in the GDM group, divided by the corresponding ratio in the ND group, using the formula: RRA = (RAGDM_ST / RAGDM_FT) / (RAND_ST / RAND_FT). RRA reflects the change in VR of a given flora between trimesters, adjusted for background changes. An RRA > 1.5 or < 0.8 was considered statistically significant.
Statistical analysis was performed using SPSS 26.0 software (SPSS Inc., Chicago, IL, United States). Categorical variables were expressed as frequencies and compared using the chi-square test. Quantitative data following a normal distribution were presented as mean ± standard deviation and compared using the t-test. For non-normally distributed data, the median (Q1, Q3) was provided, and comparisons were made using the Mann–Whitney U test or Kruskal-Wallis test. For relative abundance comparisons, p-values or Q-values (false discovery rate adjusted) < 0.05 were considered significant. In the LEfSe analysis, microbiotas with linear discriminant analysis (LDA) scores >2 were identified as potential biomarkers. Statistical significance was defined as p < 0.05.
Of the 157 pregnant women who met the inclusion criteria, 93 successfully provided fecal samples in both the first and second trimesters and were included in the study (Figure 1). The participants’ ages ranged from 19 to 42 years, with an average age of 31.7 ± 3.4 years. The gestational age at the time of the first sample collection ranged from 11 weeks and 3 days to 13 weeks and 6 days, while for the second sample, it ranged from 21 weeks to 23 weeks and 5 days. In the second trimester, 11 women (11.83%) were diagnosed with GDM. The GDM group exhibited significantly higher BMIs and OGTT blood glucose levels compared to ND group (p < 0.05). However, no significant differences were observed between the groups in terms of maternal age, ethnicity, parity, occupation, educational status, gestational weeks at either sampling time point, fasting plasma glucose, total bile acid, or glycosylated hemoglobin levels (Table 1).
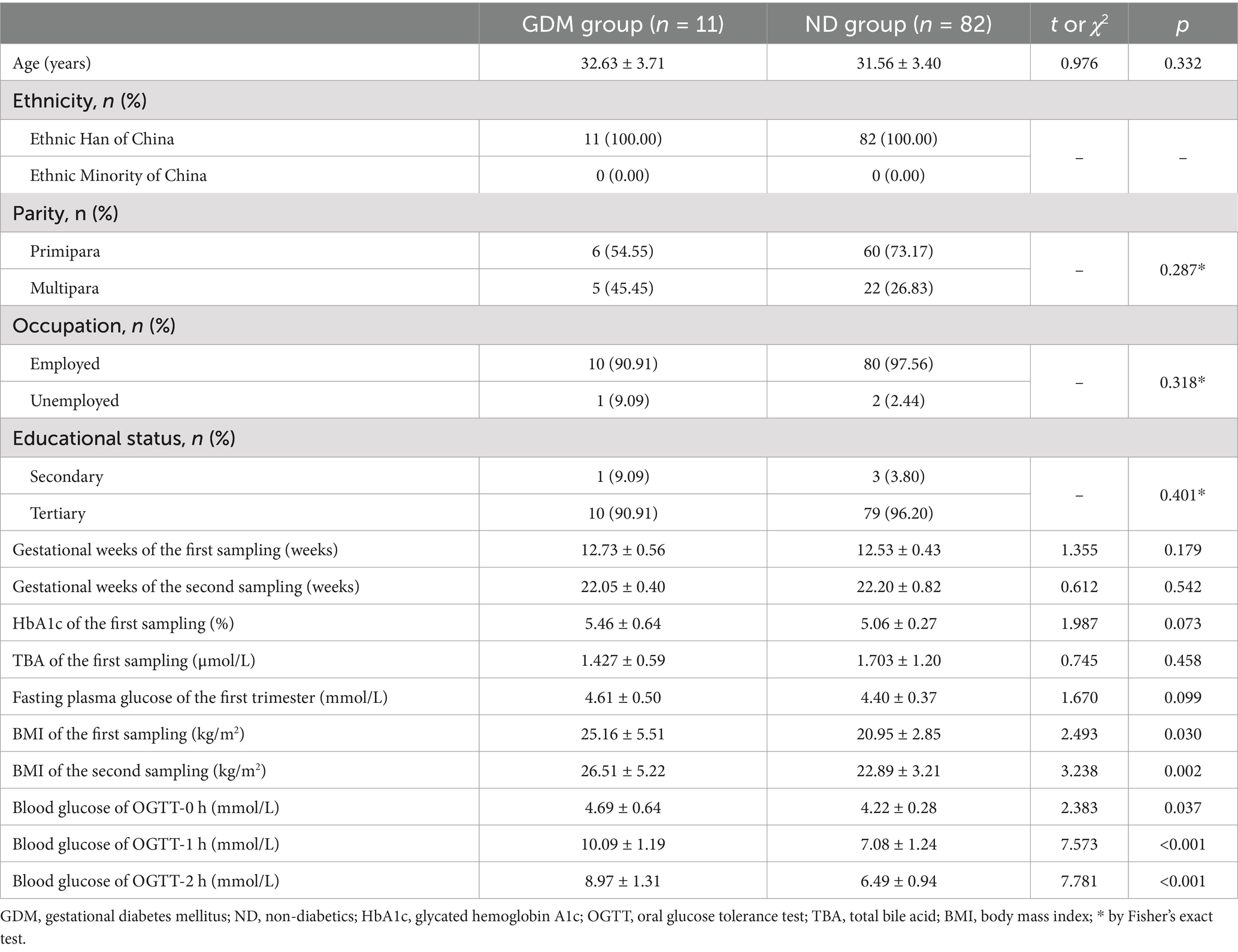
Table 1. Comparison of clinical characteristics of the participants between GDM group and ND group.
Sequencing the 16S rRNA genes from 186 fecal samples produced a total of 21,280,018 high-quality reads, averaging 114,408.7 clean reads per sample. Using a 100% similarity threshold, clean reads were clustered via sklearn species classification, resulting in 80,829 amplicon sequence variants (ASVs). These ASVs were subsequently matched to the Greengenes2 database for taxonomic annotations. At a confidence coefficient of 0.7, all the ASVs were identified, with 60,908 classified at the genus level.
At the phylum level, the dominant microbiotas (with abundance over 1%) were Bacteroidota (43.59%), Firmicutes_A (36.28%), Proteobacteria (7.19%), Firmicutes_C (5.37%), Actinobacteriota (2.97%), and Firmicutes_D (2.83%), collectively accounting for approximately 98.24% of the total microbiota. In the first trimester, Bacteroidota (47.75%) and Firmicutes_A (34.53%) were the most prevalent phyla. In the second trimester, Bacteroidota (39.43%) and Firmicutes_A (38.03%) remained dominant (Supplementary Table S1).
At the genus level, the dominant genera (with relative abundance over 1%) were Phocaeicola_A_858004, Prevotella, Faecalibacterium, Bacteroides_H, Gemmiger_A_73129, Escherichia_710834, Lachnospira, Bifidobacterium_388775, Blautia_A_141781, Acinetobacter, Megamonas, Collinsella, Agathobacter_164117, Phascolarctobacterium_A, Parabacteroides_B_862066, Dialister, Anaerostipes, Fusicatenibacter, Streptococcus, Enterocloster, Acetatifactor, Roseburia, and Klebsiella_724518, which together constituted 74% of the total abundance. In the first trimester, the most abundant genera (>5%) were Phocaeicola_A_858004, Prevotella, Faecalibacterium, and Bacteroides_H. In the second trimester, Gemmiger_A_73129 joined this group (>5%) (Supplementary Table S2).
In the first trimester, the GDM group showed slight, non-significant variations in the dominant phyla compared to the ND group (Figure 2A). While Prevotella increased in the GDM group, this increase was not statistically significant (Figure 2B). However, Bacteroides_H (p = 0.020) (Figure 2C), Acetatifactor (p = 0.049) (Figure 2D), and Megasphaera_A_38685 (p = 0.018) (Figure 2E) significantly decreased in the GDM group. Alpha diversity (Figures 2F,G) and beta diversity (Figures 2H,I) showed no significant differences between the groups. The volcano plot indicated a significant increase in CAG_417 and a decrease in Scybalousia, Megasphaera_A_38685, UBA1394, Burkholderia, Choladousia, and JC017 in the GDM group (Figure 2J). Anaeroglobus was identified as a GDM marker, while Bacteroides_H, Acetatifactor, Megasphaera_A_38685, Enterococcaceae, and Enterococcus_E were markers for the ND group (Figure 2K). The Circos plot suggested a weaker association of Acetatifactor, Scybalousia, Megasphaera_A_38685, and Burkholderia with GDM (Figure 2L).
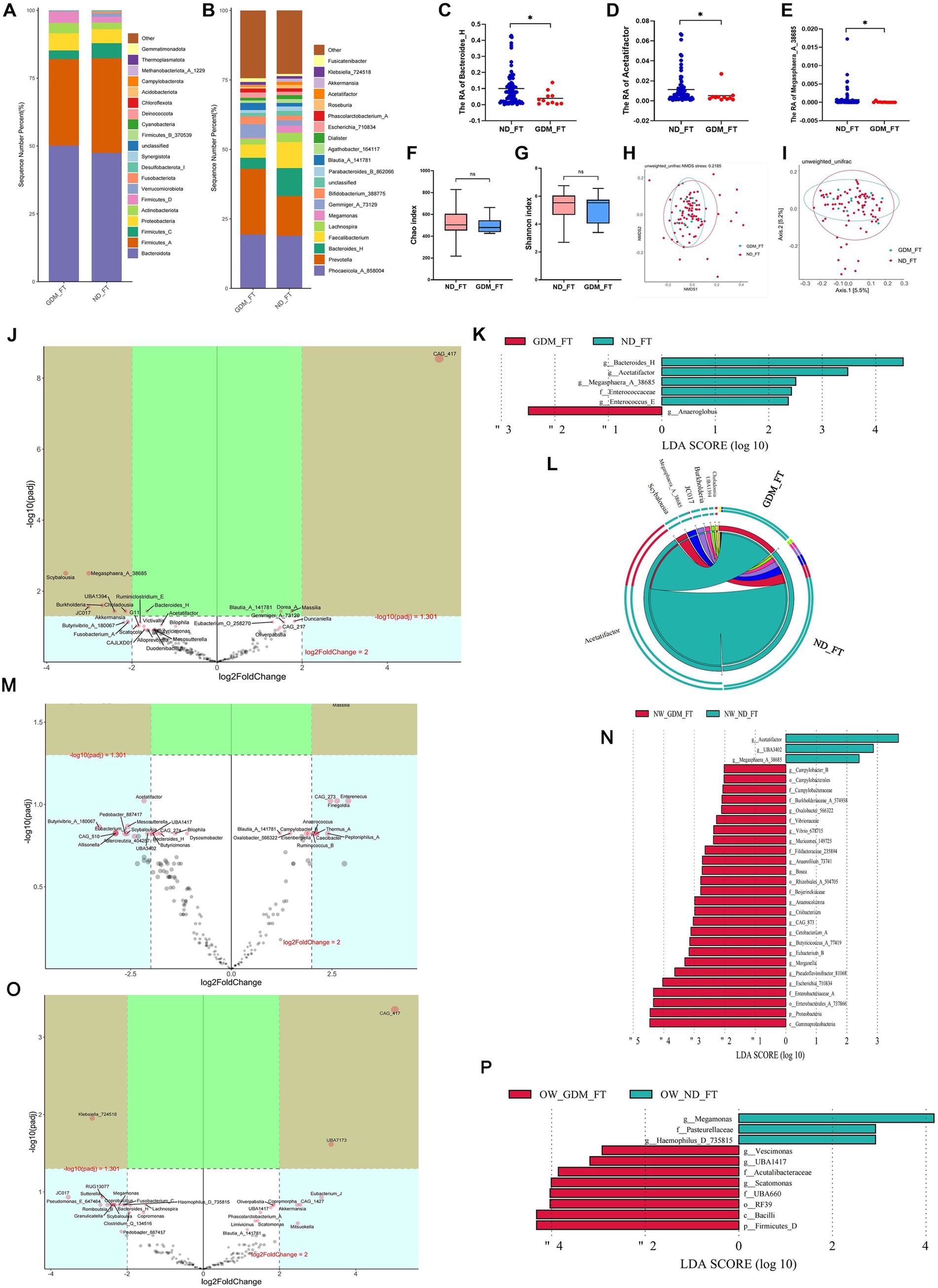
Figure 2. Comparison of the structure and diversity of gut microbiota between the GDM and ND groups in the first trimester. The dominant phyla (A) and genera (B) differed between ND and GDM groups in FT. Bacteroides_H (C), Acetatifactor (D), and Megasphaera_A_38685 (E) were significantly lower in the GDM group (p < 0.05). No significant differences were found in the Chao (F) or Shannon indices (G). NMDS (H) and PCoA (I) showed no distinct microbiota separation. The volcano plot (J) highlighted six genera that increased and one that decreased significantly in the GDM group. LEfSe analysis (K) identified one biomarker for GDM and five for ND in FT. The Circos plot (L) showed a closer relationship between Acetatifactor, Scybalousia, Megasphaera_A_38685, and Burkholderia with ND. Significant differential genus related to GDM were identified in participants with normal weight (M), and those overweight or obese (O). LEfSe analysis also pinpointed biomarkers for both groups among participants who were normal weight (N) and those overweight or obese (P). * p < 0.05.
In normal-weight women, the GDM group showed a significant increase in Escherichia_710834, Blautia_A_141780, Massilia, Thermus_A, Campylobacter_B, Oxalobacter_566322, and Slackia_A, while Megasphaera_A_38685, UBA3402, and Acetatifactor decreased (p < 0.05) (Supplementary Table S3). The volcano plot highlighted a significant increase in Massilia and a decrease in Megasphaera_A_38685 (Figure 2M). Linear discriminant analysis (LDA) identified 26 marker bacterial communities in the GDM group and 3 in the ND group (Figure 2N).
In overweight or obese GDM women, the abundance of Vescimonas, UBA1417, CAG_510, Gordonibacter, Brevundimonas, Eubacterium_O_258270, and Scatomonas significantly increased, while Haemophilus_D_735815 decreased (p < 0.05) (Supplementary Table S4). CAG_417 and UBA7173 were notably higher, and Klebsiella_724518 was lower in GDM patients, as shown by volcano plot (Figure 2O). LDA identified 8 marker bacterial communities in the GDM group and 3 in the ND group (Figure 2P). No significant difference in microbiota diversity was observed between the GDM and ND groups, regardless of BMI, in the first trimester (Supplementary Figures S1–S4).
In the second trimester, the composition of dominant phyla (Figure 3A) and genera (Figure 3B) differed between the GDM and ND groups. At the phylum level, Fusobacteriota (Figure 3C) and Firmicutes_D (Figure 3D) were significantly higher in the GDM group (p < 0.05). At the genus level, Fusobacterium_A (Figure 3E), Scatomonas (Figure 3F), and Fournierella (Figure 3G) significantly increased, while Anaerotruncus (Figure 3H), Coprobacter (Figure 3I), and Angelakisella (Figure 3J) significantly decreased in the GDM group (p < 0.05). However, alpha diversity (Figures 3K,L) and beta diversity (Figures 3M,N) did not significantly differ between the GDM and ND groups. The Volcano plot (Figure 3O) showed that in the second trimester, Butyribacter and Allisonella were significantly increased, while OM05_12, QALR01, Fenollaria, Duncaniella, Scatocola, Ezakiella, Limisoma, Sutterella, Coprobacter, Acidaminococcus, Cryptobacteroides, and Onthocola_B were significantly decreased in the GDM group. LEfSe analysis identified 13 microbiotas as biomarkers for the GDM group and 15 for the ND group (Figure 3P). The Circos plot suggested a close association between Butyribacter, Duncaniella, and Allisonella with the GDM group (Figure 3Q).
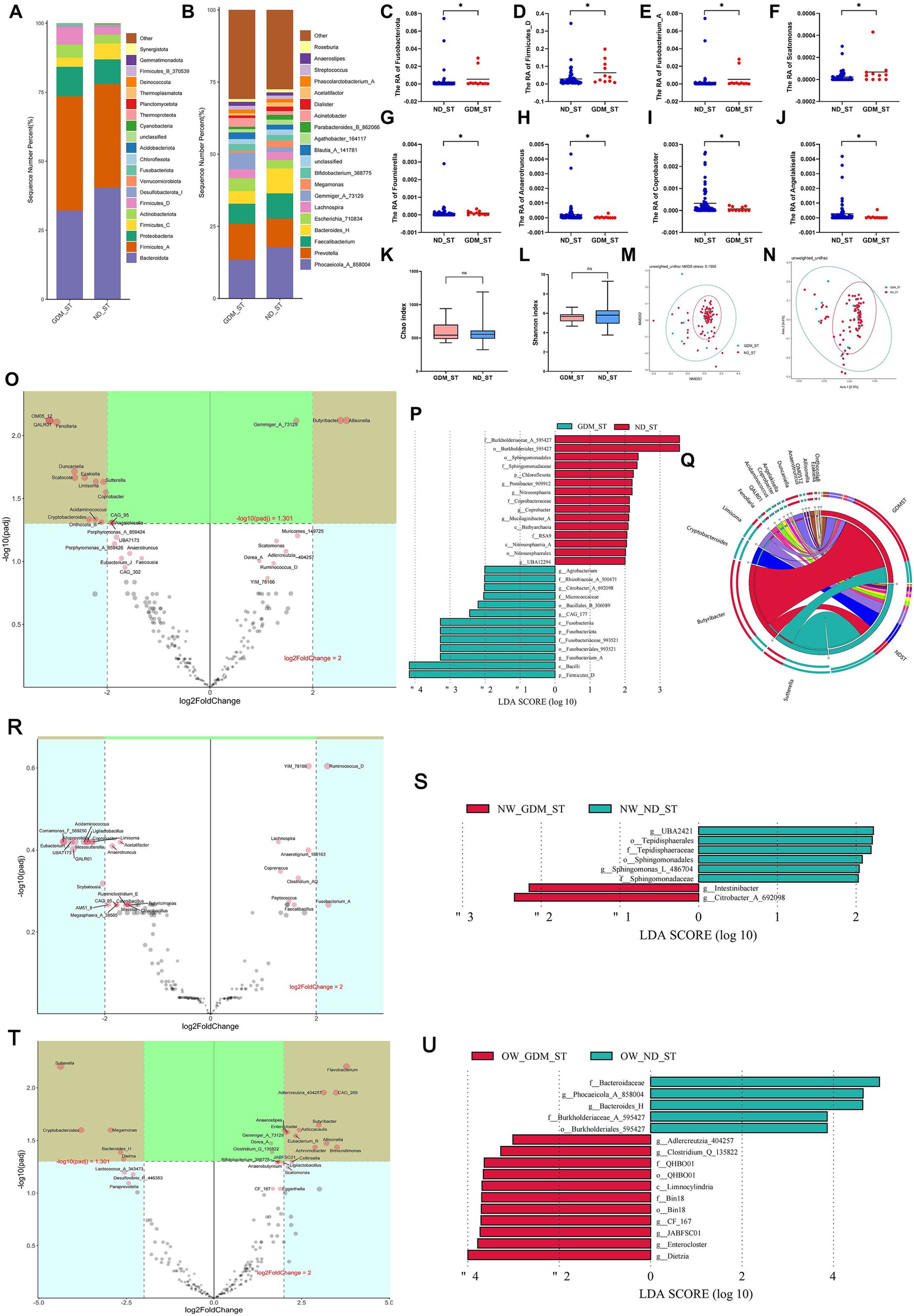
Figure 3. Comparison of the structure and diversity of gut microbiota between the GDM and ND groups in the second trimester. The proportion of dominant phyla (A) and genera (B) varied between the ND and GDM groups in the second trimester (ST). At the phylum level, Fusobacteriota (C) and Firmicutes_D (D) significantly decreased in GDM group. At the genus level, Fusobacterium_A (E), Scatomonas (F), and Fournierella (G) significantly increased, whereas Anaerotruncus (H), Coprobacter (I), and Angelakisella (J) significantly decreased in GDM group. No significant differences were noted in the Chao (K) or Shannon indices (L). NMDS (M) and PCoA (N) showed no microbiota separation between groups. The volcano plot (O) showed two genera increased and twelve decreased in GDM. LEfSe analysis (P) identified thirteen GDM biomarkers and fifteen ND biomarkers. The Circos plot (Q) highlighted close relationships between Butyribacter, Duncaniella, and Allisonella with GDM. The volcano plot showed no significant differential genera related to GDM in normal-weight participants (R), but overweight or obese women had increases in Enterocloster, Gemmiger_A_73129, and nine other genera, and decreases in Megamonas, Bacteroides_H, and three others (T). LEfSe analysis identified biomarker for the GDM group and ND group in the participants with normal weight (S), overweight or obese (U). *p < 0.05.
In normal-weight women, Christensenella, Coprenecus, and Intestinibacter were significantly more abundant in the GDM group during the second trimester (p < 0.05) (Supplementary Table S5). However, no significant difference was found between the GDM and ND groups in the volcano plot (Figure 3R). LDA identified 2 marker bacterial communities in the GDM group and 6 in the ND group (Figure 3S).
For overweight or obese participants, Enterocloster, Clostridium_Q_135822, and Adlercreutzia_404257 increased, whereas Dielma, Phocaeicola_A_858004, and Bacteroides_H decreased in the GDM group (Supplementary Table S6). The volcano plot indicated significant increases in Flavobacterium, Adlercreutzia_404257, CAG_269, Butyribacter, Asticcacaulis, Enterocloster, Gemmiger_A_73129, Eubacterium_R, Allisonella, Brevundimonas, and Achromobacter, while Sutterella, Cryptobacteroides, Megamonas, Bacteroides_H and Dielma significantly decreased (Figure 3T). LDA identified 11 marker communities in the GDM group and 5 in the ND group (Figure 3U). No significant difference in GM alpha or beta diversity was observed between the GDM and ND groups in the second trimester, regardless of initial BMI (Supplementary Figures S5–S8).
Matched comparative analyses were used to investigate longitudinal changes in the microbiota from the first trimester (FT) to the second trimester (ST). In the GDM group, the relative abundance of 14 genera during the second trimester was markedly higher than in the first trimester; however, only 8 of these genera showed a significant increase in the ND group. Specifically, the abundance of Streptococcus rose from 0.288 to 1.308% in the GDM group, compared to an increase from 0.521 to 1.166% in the ND group (Supplementary Table S7).
At the genus level, compared with the ND group, the variances of relative abundance (VRAs) of Eubacterium_R (Figure 4A), Allisonella (Figure 4B), Fournierella (Figure 4C), Intestinibacter (Figure 4D), and CAG_41 (Figure 4E) significantly increased in the GDM group (p < 0.05). Conversely, Angelakisella (Figure 4F) exhibited a significant decrease in VRA (p < 0.05) (Supplementary Table S8). In overweight or obese participants, the VRA of Megamonas in the GDM group was significantly lower than in the ND group (Figure 4G), whereas this difference was not observed in women of normal weight. Among normal-weight women, the VRA of Faecalibacterium was higher (Figure 4H), while the VRA of Escherichia_710834 was lower (Figure 4I), compared to the ND group.
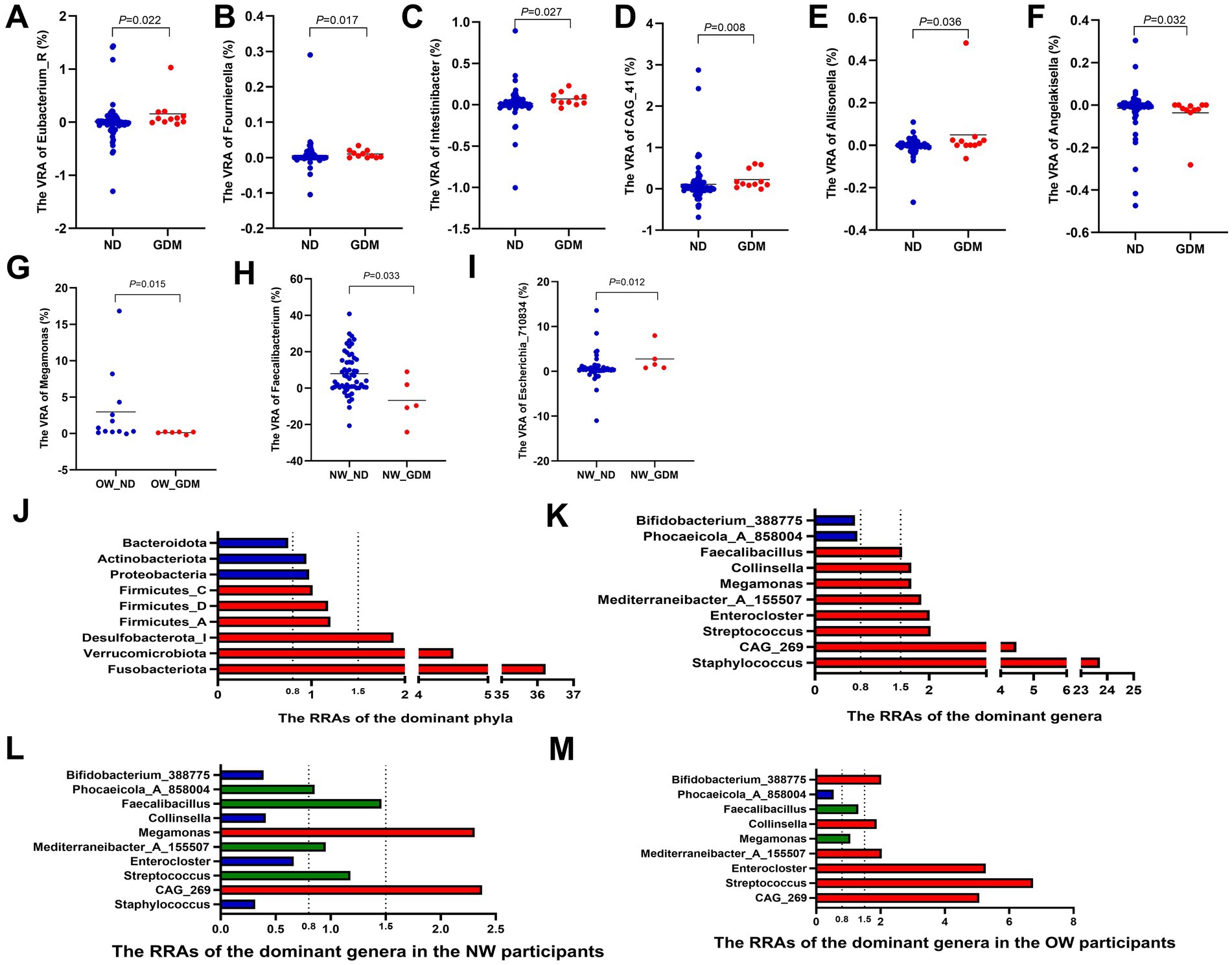
Figure 4. Variations in the relative abundance of dominant phyla and genera between the GDM and ND groups. The VRAs of Eubacterium_R (A), Allisonella (B), Fournierella (C), Intestinibacter (D) and CAG_41 (E) increased, while Angelakisella (F) decreased in GDM group. Overweight or obese GDM women showed increased VRA of Megamonas (G), whereas normal-weight GDM women had higher VRA of Faecalibacterium (H) and lower VRA of Escherichia_710834 (I). Phyla Fusobacteriota, Verrucomicrobiota, and Desulfobacterota_I had RRAs over 1.5, while Bacteroidota was below 0.8 (J). Genera Staphylococcus, Streptococcus, Enterocloster, Collinsella, and Faecalibacillus exceeded 1.5, with Phocaeicola_A_858004 and Bifidobacterium_388775 below 0.8 (K). In normal-weight women, Staphylococcus, Enterocloster, and Collinsella had RRAs less than 0.8, with Streptococcus, Mediterraneibacter_A_155507, and Faecalibacillus between 0.8 and 1.5, and only CAG_269 and Megamonas above 1.5 (L). In overweight or obese women, RRAs of CAG_269, Streptococcus, Enterocloster, Mediterraneibacter_A_155507, and Collinsella exceeded 1.5 (M). RA, relative abundance; VRA, variance of the relative abundance from the first to the second trimester; RRA, ratio of the relative abundance. * p < 0.05.
The relative abundance ratios (RRAs) of Fusobacteriota, Verrucomicrobiota, and Desulfobacterota_I in the GDM group all exceeded 1.5 (36.22, 4.50, and 1.88, respectively) (Figure 4I), indicating a higher magnitude of increase from the first to the second trimester in the GDM group compared to the ND group. In contrast, the RRA of Bacteroidota was only 0.75, indicating a lesser change in relative abundance in the GDM group (Figure 4J; Supplementary Table S9). At the genus level, the RRAs of Staphylococcus, CAG_269, Streptococcus, Enterocloster, Mediterraneibacter_A_155507, Megamonas, Collinsella, and Faecalibacillus were all greater than 1.5 (23.72, 4.48, 2.03, 2.01, 1.86, 1.69, 1.69, and 1.53, respectively), while the RRAs of Phocaeicola_A_858004 and Bifidobacterium_388775 were both less than 0.8 (0.74 and 0.70, respectively) (Figure 4K; Supplementary Table S10).
In normal-weight women, the RRAs of Staphylococcus, Enterocloster and Collinsella were all less than 0.8. The RRAs of Streptococcus, Mediterraneibacter_A_155507 and Faecalibacillus were between 0.8 and 1.5, while only CAG_269 and Megamonas maintained RRAs greater than 1.5 (Figure 4L). However, in overweight or obese women, the RRAs of CAG_269, Streptococcus, Enterocloster, Mediterraneibacter_A_155507, and Collinsella were all larger than 1.5 (Figure 4M).
Spearman analysis revealed positive correlations between the variances of relative abundance (VRAs) of certain microbiotas and blood glucose levels during OGTT. Specifically, Fusobacterium_A (r = 0.258, p = 0.015), Gemella (r = 0.285, p = 0.007), Schaedlerella (r = 0.225, p = 0.035) and Porphyromonas_A_859426 (r = 0.251, p = 0.018) showed positive relationships with OGTT-0 h blood glucose levels. Similarly, Fusobacterium_A (r = 0.223, p = 0.037) and Fusobacterium_C (r = 0.211, p = 0.048) were positively correlated with OGTT-1 h blood glucose levels, while Erysipelatoclostridium (r = 0.224, p = 0.036), Fusobacterium_C (r = 0.216, p = 0.043), and Peptococcus (r = 0.240, p = 0.025) were positively correlated with OGTT-2 h blood glucose levels (Figure 5).
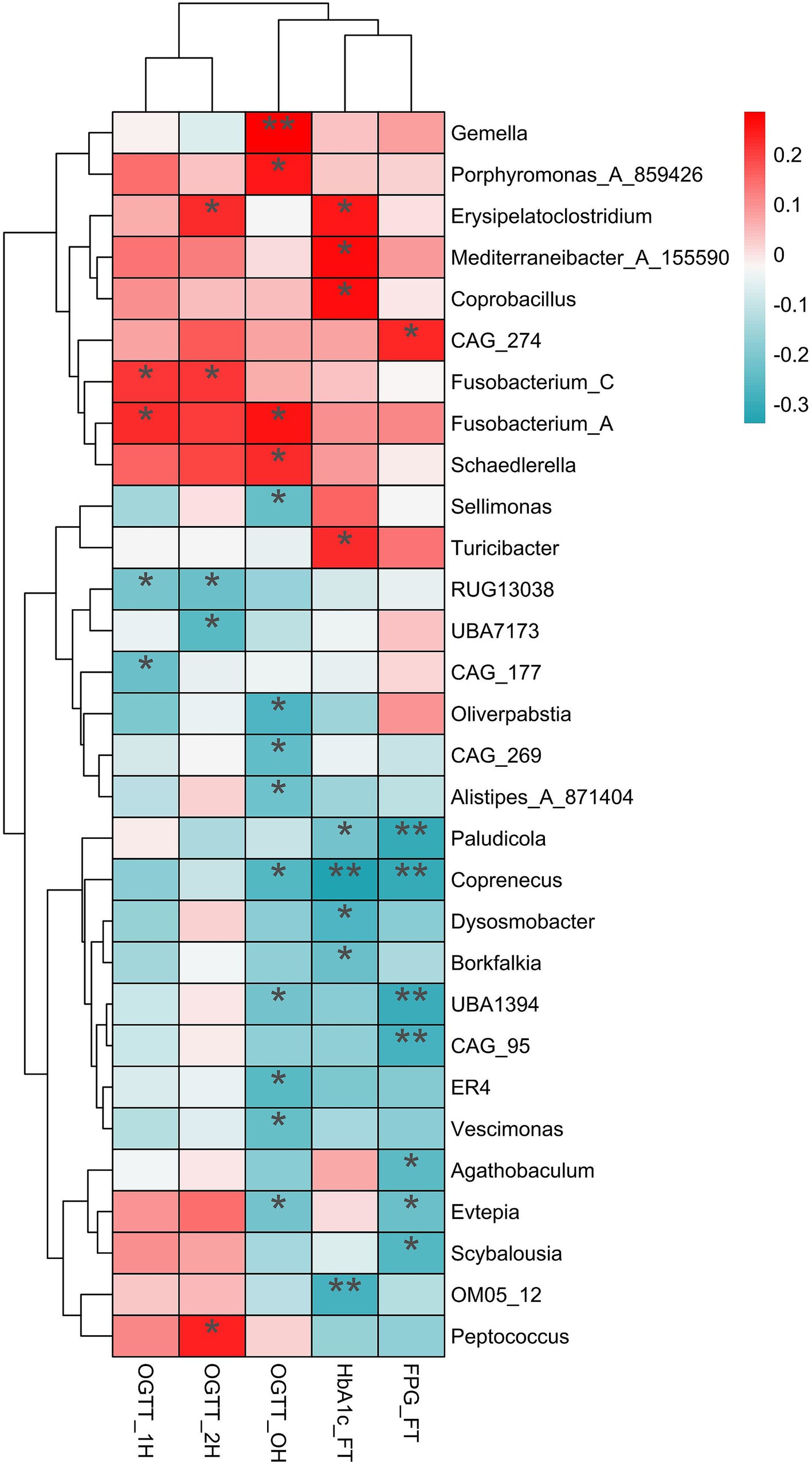
Figure 5. The relationship of variances of relative abundance with the blood glucose and HblAc. The significantly positive relationship (red) and negative relationship (dark turquoise) were label by asterisk. OGTT, oral glucose tolerance test; HblAc, glycated hemoglobin A1c; FPG, fasting plasma glucose; * p < 0.05, **p < 0.01.
Additionally, the VRAs of Erysipelatoclostridium (r = 0.251, p = 0.020), Mediterraneibacter_A_155590 (r = 0.271, p = 0.012), Coprobacillus (r = 0.261, p = 0.016), and Turicibacter (r = 0.227, p = 0.037) positively correlated with HbA1c levels in the first trimester (p < 0.05). Conversely, the VRAs of Sellimonas (r = −0.235, p = 0.028), Oliverpabstia (r = −0.265, p = 0.013), CAG_269 (r = −0.243, p = 0.023), Alistipes_A_871404 (r = −0.225, p = 0.035), Coprenecus (r = −0.259, p = 0.015), and Vescimonas (r = −0.232, p = 0.029) were negatively related to OGTT-0 h blood glucose (Figure 5).
PICRUSt2 software analysis predicted differences in KEGG pathways between the groups. In the first trimester, one pathway [Methane metabolism (p = 0.030)] was significantly up-regulated, while five KEGG pathways [ECM-receptor interaction (p = 0.022), Furfural degradation (p = 0.002), Lipoarabinomannan (LAM) biosynthesis (p = 0.003), Lipoic acid metabolism (p = 0.038), Neomycin, kanamycin and gentamicin biosynthesis (p = 0.026)] were significantly down-regulated in the GDM group compared to the ND group (Figure 6A).
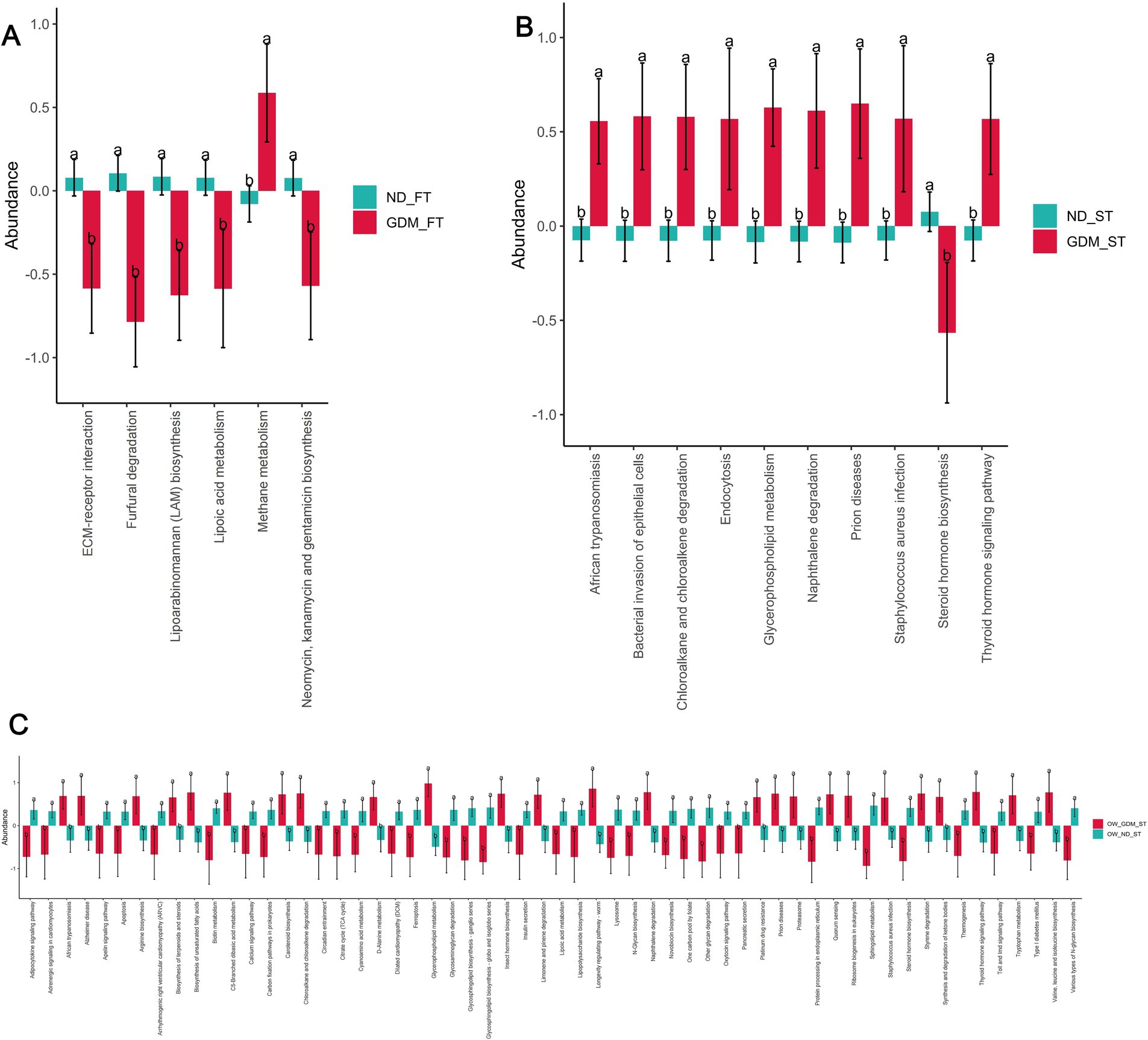
Figure 6. The KEGG pathways predicted by PICRUSt2 differed between the GDM and ND groups. The up-regulated KEGG pathways (red) and down-regulated pathways (dark turquoise) were predicted in the total participants of the first trimester (A), second trimester (B) and the overweight or obese participants in the second trimester (C). ND, non-diabetic; GDM, gestational diabetes mellitus; FT, first trimester; ST, second trimester; ECM, extracellular matrix. * p < 0.05, **p < 0.01.
In the second trimester, however, nine pathways, including African trypanosomiasis (p = 0.018), Bacterial invasion of epithelial cells (p = 0.016), Chloroalkane and chloroalkene degradation (p = 0.034), Endocytosis (p = 0.038), Glycerophospholipid metabolism (p = 0.006), Naphthalene degradation (p = 0.008), Prion diseases (p = 0.013), Staphylococcus aureus infection (p = 0.042), and Thyroid hormone signaling pathway (p = 0.016), were up-regulated in the GDM group. Concurrently, one pathway—Steroid hormone biosynthesis (p = 0.022)—was down-regulated in the GDM group. Notably, four of the nine up-regulated pathways were related to inflammation: African trypanosomiasis, bacterial invasion of epithelial cells, prion diseases, and Staphylococcus aureus infection (Figure 6B).
PICRUSt2 did not predict any pathways with significant between-group differences among individuals with normal weight during the first trimester (Supplementary Table S11). However, among overweight or obese individuals in the first trimester, PICRUSt2 predicted 25 significantly up-regulated KEGG pathways and 33 significantly down-regulated pathways in the GDM group (Figure 6C). Among the significantly up-regulated KEGG pathways, two were inflammation-related: African trypanosomiasis (p = 0.012) and Staphylococcus aureus infection (p = 0.046) (Figure 6C).
This nested cohort study delved into the cross-sectional characteristics of gut microbiota in pregnant women with gestational diabetes mellitus (GDM) and examined the longitudinal changes from the first to the second trimester. Certain gut genera identified early in pregnancy distinguished GDM from non-diabetic women, with more distinct microbiota profiles emerging in the second trimester. Critically, the longitudinal changes in gut microbiota, rather than merely cross-sectional characteristics, differed significantly based on GDM diagnosis. Compared to non-diabetic women, those with GDM showed significant microbiota shifts potentially linked to inflammation, particularly in overweight or obese individuals. These findings may provide insights into the connection between GDM and gut microbiota.
Previous studies have extensively examined the characteristics of intestinal flora in women with GDM across different trimesters. During the first trimester, the families Ruminococcaceae UCG (Ma et al., 2020) and genera Bifidobacterium (Chen F. et al., 2021; Hu et al., 2021; Zhang et al., 2021), and Prevotella (Pinto et al., 2023; Vavreckova et al., 2022) were all reduced in GDM women, whereas Ruminococcus gnavus (Li et al., 2023) increased in GDM. However, the relationship between some microbiotas and GDM varied across studies. In Zheng et al.’s (2020) study, Streptococcus was reduced in GDM, while in Zhang et al.’s (2021) study, it increased. Lachnospiraceae also exhibited opposite trends in different studies (Hu et al., 2021; Ma et al., 2020). In the first trimester, our study observed decreased levels of Bacteroides_H and Acetatifactor in the GDM group, identified as biomarkers for the non-diabetic group. These findings contrast with one previous study (Dualib et al., 2022) but align with another (Li et al., 2023), emphasizing the heterogeneity in gut microbiota profiles across different populations and methodologies. The decrease in Bacteroides_H and Acetatifactor, both known producers of lithocholic acid (Pathak et al., 2018), suggests abnormalities in bile acid-metabolizing bacteria that may influence glucose metabolism. Bacteroides is suggested to be a harmful bacterium associated with obesity (Ponzo et al., 2019) and Acetatifactor was increased in animal models with a high-fat diet (Zhu et al., 2023). Given that the Greengenes2 annotation database used in this study was recently updated (McDonald et al., 2024), Bacteroides_H annotated by this new database has not been previously reported as related to GDM. Further investigation is warranted to elucidate the correlation between these genera and GDM in early pregnancy.
Moving into the second trimester, previous studies have consistently showed that phyla Bifidobacterium (Chen F. et al., 2021) and Actinobacteria (Su et al., 2021; Chen T. et al., 2021), and genera Lachnospiraceae (Chen T. et al., 2021; Wang et al., 2020), Eubacterium (Chen F. et al., 2021; Su et al., 2021), Prevotella (Chen F. et al., 2021; Ye et al., 2023), Collinsella (Su et al., 2021; Tanaka et al., 2022), and Faecalibacterium (Ye et al., 2019, 2023) decreased in GDM patients, while Blautia (Chen F. et al., 2021; Ye et al., 2019), Parabacteroides (Su et al., 2021; Kuang et al., 2017), and Megamonas (Ye et al., 2023; Kuang et al., 2017) increased. However, phylum Bacteroidetes, Verrucomicrobia, and genera Bacteroides, Subdoligranulum, Eggerthella, and Clostridium manifested inconsistent results across studies (Wang S. et al., 2024). In our study, phyla Fusobacteriota and Firmicutes_D, as well as genera Fusobacterium_A, Scatomonas, and Fournierella, all increased in the GDM group during the second trimester. LEfSe analysis indicated that Firmicutes_D and Fusobacteria were markers of GDM. Spearman analysis revealed a positive correlation between Fusobacterium_A and OGTT-0 h and 1 h glucose levels. The changes of Firmicutes in our study align with some previous studies (Liu et al., 2023; Wei et al., 2021) and underscore the importance of its alterations during this trimester in the occurrence of GDM. Firmicutes levels also rose in the GDM group during the third trimester (Li G. et al., 2021). Additional, specific Firmicutes phyla such as Collinsella, Olsenella, and Clostridium were significantly elevated in postpartum women diagnosed with GDM (Crusell et al., 2018). However, the phylum Firmicutes and class Clostridia were notably decreased in men with type 2 diabetes (T2DM) (Larsen et al., 2010). The variations in subject demographics and types of diabetes might account for these discrepancies. Regarding Fusobacteria, Wang et al.’s study (Wang et al., 2018) indicated an increase in women with GDM and a negative correlation between the Faecalibacterium/Fusobacterium ratio and GDM, aligning with Cortez et al.’s findings during late pregnancy in GDM women (Cortez et al., 2019). Significantly, a higher Faecalibacterium/Fusobacterium ratio has been linked to obesity-induced low-grade inflammation (Roselli et al., 2017). Our findings suggest that changes in intestinal flora structure during the second trimester are closely related to the development of GDM.
To mitigate the impact of population background factors and gestational age on microbiota analyses, we utilized the indices of the variance of relative abundance (VRA) and the ratios of relative abundance (RRA). The RRA values revealed substantial increases or decreases in dominant microbiota from the first to the second trimester in the GDM group, after adjusting for background changes. Specifically, the RRAs of phyla and genera associated with inflammation, such as Staphylococcus, Streptococcus, Enterococcus, and Collinsella, exceeded the cut-off value of 1.5, indicating a positive correlation with GDM. These findings are consistent with previous studies reporting increased levels of inflammatory bacteria in GDM patients (Wei et al., 2021; Vavreckova et al., 2022; Rold et al., 2022) and suggest that inflammation may play a crucial role in the gut microbiota changes associated with GDM. Staphylococcus, Streptococcus, and Enterococcus are identified as opportunistic pathogens (Mao et al., 2024), which are linked to intestinal inflammation. Pathogenic microbiotas may trigger Toll-like receptor 4 via lipopolysaccharides (LPS) in the gut mucosal barrier, contributing to metabolic inflammation (Tang et al., 2021). Inflammation-associated bacteria such as Collinsella and Streptococcus were markedly elevated not only in individuals with T2DM (Zhong et al., 2019; Takagi et al., 2020) but also in those with GDM (Hasain et al., 2020). Collinsella is recognized for its pro-inflammatory influence during pregnancy (Gomez-Arango et al., 2018) and has been positively linked to elevated maternal insulin levels (Gomez-Arango et al., 2016, 2018). Furthermore, our study observed that the relative abundance of Bifidobacterium_388775 was below 0.8 in the GDM group, aligning with previous research findings (Chen F. et al., 2021). Bifidobacterium species are renowned for their anti-inflammatory properties in neonatal diabetes (Alsharairi, 2023) and acute pancreatitis (Li et al., 2022). Supplementation with Bifidobacterium probiotics helps mitigate inflammation and oxidative stress in GDM (Zhang et al., 2019). T2DM patients typically have reduced numbers of Bifidobacterium species (Lê et al., 2013). Consequently, the results of this study might indicate a future risk of developing T2DM in these individuals and could therefore be pivotal for preventing T2DM following GDM.
PICRUSt2 analysis predicted the upregulation of inflammatory pathways in the second trimester within the GDM group in our study, further supporting the link between gut microbiota dysbiosis, inflammation, and GDM. It has been reported that the low-grade inflammation caused by gut microbiota dysbiosis may contribute to insulin resistance and hyperglycemia in women with GDM (Hu et al., 2023). These results suggest that inflammation may play a role in the gut microbiota changes associated with GDM. Nevertheless, we did not detect any clinically apparent inflammation in the GDM group (Supplementary Table S12). We attribute this finding to two potential reasons. Firstly, the relatively small sample size of the GDM group might not have been sufficient to demonstrate a significant statistical difference. Secondly, based on findings from other studies (Hasain et al., 2020), the inflammation associated with GDM is often considered low-grade, which may explain the absence of a typical inflammatory response.
Stratified analysis by BMI revealed distinct microbiota changes in overweight or obese women with GDM compared to normal-weight women. Specifically, inflammation-associated genera like Streptococcus, Enterocloster, Collinsella, Gemmiger_A_73129 and Enterocloster increased, while the beneficial butyrate-producer Megamonas decreased in overweight or obese GDM women. These findings, along with the upregulation of inflammatory pathways predicted by PICRUSt2 analysis, suggest that inflammatory microbiota and pathways may play a significant role in the development of GDM in this subset of the population. Higher Gemmiger levels correlate with inflammation (Lang et al., 2021), Enterocloster is linked to frailty and inflammatory markers (Dong et al., 2024), and lower Megamonas levels are associated with systemic inflammation (Chen Yan et al., 2024). Similarly, the colostrum of women who are obese or have GDM showed an overabundance of Staphylococcus compared to the control group (Gámez-Valdez et al., 2021). A metabolomic study found elevated levels of branched-chain amino acids and glycoprotein acetylation, a marker of low-grade inflammation, in overweight or obese women with GDM (Mokkala et al., 2020). BMI critically mediates the link between systemic inflammation and diabetes (Chen Yongze et al., 2024). These findings suggest that inflammatory microbiota and pathways may play a significant role in GDM development in this group, warranting further research.
This study has several limitations. Firstly, the incidence of GDM in the enrolled population was lower than expected, resulting in a limited sample size for the GDM group. This may have reduced the statistical power of group comparisons and led to missed detection of significantly different bacterial flora. Secondly, information on diet, exercise, and other potential confounders was not collected, which may have influenced gut microbiota composition. However, the longitudinal comparison from the first to the second trimester helped mitigate these confounding effects. Thirdly, the 16S rRNA sequencing technology used in this study has inherent limitations and may not fully capture the genetic information of bacterial flora. Further investigation using metagenomic and metabolomic analyses is needed to reveal the specific functions and metabolic pathways of different bacterial communities.
In conclusion, this nested cohort study investigated the dynamics of gut microbiota from the first to the second trimester before GDM diagnosis and uncovered significant alterations in the GDM cohort compared to the control group. The key bacteria abundant in the GDM group, such as phyla Fusobacteriota, Firmicutes_D, and genus Fusobacterium_A, showed notable increases between the first and second trimesters. Additionally, there was an escalation in microbiota associated with inflammation, such as Staphylococcus, Streptococcus, and Enterocloster, in GDM patients, particularly among overweight or obese individuals. These findings highlight the intricate link between disruptions in gut microbiota and GDM, suggesting that elevated inflammatory responses related to microbiota dysbiosis in overweight or obese women during this critical period may influence GDM development. Further research into the role of inflammatory responses driven by gut microbiota in GDM among obese women is essential to elucidate the underlying mechanisms and develop targeted interventions.
The original contributions presented in the study are publicly available. This data can be found in the NCBI repository: https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA1193797 and the sequencing data: https://www.ncbi.nlm.nih.gov/sra/?term=SRP549371.
The studies involving humans were approved by the Research Ethics Committee of Peking University Shenzhen Hospital. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin. The study was conducted in accordance with the local legislation and institutional requirements.
SZ: Formal analysis, Funding acquisition, Methodology, Visualization, Writing – original draft, Writing – review & editing. BY: Data curation, Investigation, Writing – review & editing. YL: Formal analysis, Writing – original draft, Writing – review & editing. WD: Formal analysis, Methodology, Visualization, Writing – review & editing. GL: Methodology, Writing – review & editing. JY: Data curation, Writing – review & editing. AY: Data curation, Formal analysis, Writing – review & editing. YW: Writing – review & editing. MW: Writing – review & editing. CX: Formal analysis, Methodology, Writing – review & editing. YD: Investigation, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Shenzhen Science and Technology Program (JCYJ20210324110206017), Research Project of Peking University Shenzhen Hospital (LCYJ2021010), Shenzhen High-level Hospital Construction Fund (YBH2019-260), and Sanming Project of Medicine in Shenzhen (No.SZSM202011016).
We would like to thank all the participants of this study. Furthermore, we would like to thank the Wekemo Technology Group Company (Shenzhen, China) which provided the service of sequencing service and data analysis.
GL was employed by CheerLand Biological Technology Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1467414/full#supplementary-material
American Diabetes Association (2020). 2. Classification and diagnosis of diabetes: standards of medical Care in Diabetes—2020. Diabetes Care 43, S14–S31. doi: 10.2337/dc20-S002
Afzaal, M., Saeed, F., Shah, Y. A., Hussain, M., Rabail, R., Socol, C. T., et al. (2022). Human gut microbiota in health and disease: unveiling the relationship. Front. Microbiol. 13, 999001. doi: 10.3389/fmicb.2022.999001
Alsharairi, N. A. (2023). Exploring the diet-gut microbiota-epigenetics crosstalk relevant to neonatal diabetes. Genes 14, 1017. doi: 10.3390/genes14051017
Balleza-Alejandri, L. R., Peña-Durán, E., Beltrán-Ramírez, A., Reynoso-Roa, A. S., Sánchez-Abundis, L. D., García-Galindo, J. J., et al. (2024). Decoding the gut microbiota–gestational diabetes link: insights from the last seven years. Microorganisms 12:1070. doi: 10.3390/microorganisms12061070
Bianco, M. E., and Josefson, J. L. (2019). Hyperglycemia during pregnancy and Long-term offspring outcomes. Curr. Diab. Rep. 19:143. doi: 10.1007/s11892-019-1267-6
Bielka, W., Przezak, A., and Pawlik, A. (2022). The role of the gut microbiota in the pathogenesis of diabetes. Int. J. Mol. Sci. 23:480. doi: 10.3390/ijms23010480
Cai, J., Rimal, B., Jiang, C., Chiang, J. Y. L., and Patterson, A. D. (2022). Bile acid metabolism and signaling, the microbiota, and metabolic disease. Pharmacol. Ther. 237:108238. doi: 10.1016/j.pharmthera.2022.108238
Catalano, P. M., McIntyre, H. D., Cruickshank, J. K., McCance, D. R., Dyer, A. R., Metzger, B. E., et al. (2012). The hyperglycemia and adverse pregnancy outcome study. Diabetes Care 35, 780–786. doi: 10.2337/dc11-1790
Chen, F., Gan, Y., Li, Y., He, W., Wu, W., Wang, K., et al. (2021). Association of Gestational Diabetes Mellitus with changes in gut microbiota composition at the species level. BMC Microbiol. 21, 1–8. doi: 10.1186/s12866-021-02207-0
Chen, Y., Huang, R., Mai, Z., Chen, H., Zhang, J., Zhao, L., et al. (2024). Association between systemic immune-inflammatory index and diabetes mellitus: mediation analysis involving obesity indicators in the NHANES. Front. Public Health 11, 1331159. doi: 10.3389/fpubh.2023.1331159
Chen, Y., Le, D., Xu, J., Jin, P., Zhang, Y., and Liao, Z. (2024). Gut microbiota Dysbiosis and inflammation dysfunction in late-life depression: an observational cross-sectional analysis. Neuropsychiatr. Dis. Treat. 20, 399–414. doi: 10.2147/NDT.S449224
Chen, T., Zhang, Y., Zhang, Y., Shan, C., Zhang, Y., Fang, K., et al. (2021). Relationships between gut microbiota, plasma glucose and gestational diabetes mellitus. J. Diabetes Investig. 12, 641–650. doi: 10.1111/jdi.13373
Cortez, R. V., Taddei, C. R., Sparvoli, L. G., Ângelo, A. G. S., Padilha, M., Mattar, R., et al. (2019). Microbiome and its relation to gestational diabetes. Endocrine 64, 254–264. doi: 10.1007/s12020-018-1813-z
Crudele, L., Gadaleta, R. M., Cariello, M., and Moschetta, A. (2023). Gut microbiota in the pathogenesis and therapeutic approaches of diabetes. EBioMedicine 97:104821. doi: 10.1016/j.ebiom.2023.104821
Crusell, M. K., Hansen, T. H., Nielsen, T., Allin, K. H., Rühlemann, M. C., Damm, P., et al. (2018). Gestational diabetes is associated with change in the gut microbiota composition in third trimester of pregnancy and postpartum. Microbiome 6:89. doi: 10.1186/s40168-018-0472-x
di Bernardo, S. C., Lava, S. A. G., Epure, A. M., Younes, S. E., Chiolero, A., Sekarski, N., et al. (2023). Consequences of gestational diabetes mellitus on neonatal cardiovascular health: MySweetHeart cohort study. Pediatr. Res. 94, 231–238. doi: 10.1038/s41390-022-02390-4
Domanski, G., Lange, A. E., Ittermann, T., Allenberg, H., Spoo, R. A., Zygmunt, M., et al. (2018). Evaluation of neonatal and maternal morbidity in mothers with gestational diabetes: a population-based study. BMC Pregnancy Childbirth 18:367. doi: 10.1186/s12884-018-2005-9
Dong, S., Zeng, Q., He, W., Cheng, W., Zhang, L., Zhong, R., et al. (2024). Effect of Lactobacillus Plantarum BFS1243 on a female frailty model induced by fecal microbiota transplantation in germ-free mice. Food Funct. 15, 3993–4009. doi: 10.1039/D3FO05282F
Dualib, P. M., Taddei, C. R., Fernandes, G., Carvalho, C. R. S., Sparvoli, L. G., Silva, I. T., et al. (2022). Gut microbiota across Normal gestation and gestational diabetes mellitus: a cohort analysis. Meta 12, 1–13. doi: 10.3390/metabo12090796
Gámez-Valdez, J. S., García-Mazcorro, J. F., Montoya-Rincón, A. H., Rodríguez-Reyes, D. L., Jiménez-Blanco, G., Rodríguez, M. T. A., et al. (2021). Differential analysis of the bacterial Community in Colostrum Samples from women with gestational diabetes mellitus and obesity. Sci. Rep. 11:24373. doi: 10.1038/s41598-021-03779-7
Gomez-Arango, L. F., Barrett, H. L., McIntyre, H. D., Callaway, L. K., Morrison, M., Dekker Nitert, M., et al. (2016). Connections between the gut microbiome and metabolic hormones in early pregnancy in overweight and obese women. Diabetes 65, 2214–2223. doi: 10.2337/db16-0278
Gomez-Arango, L. F., Barrett, H. L., Wilkinson, S. A., Callaway, L. K., McIntyre, H. D., Morrison, M., et al. (2018). Low dietary Fiber intake increases Collinsella abundance in the gut microbiota of overweight and obese pregnant women. Gut Microbes 9, 189–201. doi: 10.1080/19490976.2017.1406584
Gurung, M., Li, Z., You, H., Rodrigues, R., Jump, D. B., Morgun, A., et al. (2020). Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 51:102590. doi: 10.1016/j.ebiom.2019.11.051
Hasain, Z., Mokhtar, N. M., Kamaruddin, N. A., Mohamed Ismail, N. A., Razalli, N. H., Gnanou, J. V., et al. (2020). Gut microbiota and gestational diabetes mellitus: a review of host-gut microbiota interactions and their therapeutic potential. Front. Cell. Infect. Microbiol. 10:188. doi: 10.3389/fcimb.2020.00188
He, Y., Ching Wan Ma, R., McIntyre, H. D., Sacks, D. A., Lowe, J., Catalano, P. M., et al. (2022). Comparing IADPSG and NICE diagnostic criteria for GDM in predicting adverse pregnancy outcomes. Diabetes Care 45, 2046–2054. doi: 10.2337/dc22-0579
He, L.-R., Li, Y., and Guo, Y. (2023). Birth weight and large for gestational age trends in offspring of pregnant women with gestational diabetes mellitus in southern China, 2012-2021. Front. Endocrinol. 14, 1166533. doi: 10.3389/fendo.2023.1166533/full
Hu, P., Chen, X., Chu, X., Fan, M., Ye, Y., Wang, Y., et al. (2021). Association of gut Microbiota during early pregnancy with risk of incident gestational diabetes mellitus. J. Clin. Endocrinol. Metabol. 106, e4128–e4141. doi: 10.1210/clinem/dgab346
Hu, R., Liu, Z., Geng, Y., Huang, Y., Li, F., Dong, H., et al. (2023). Gut microbiota and critical metabolites: potential target in preventing gestational diabetes mellitus? Microorganisms 11, 1–18. doi: 10.3390/microorganisms11071725
Juan, J., and Yang, H. (2020). Prevalence, prevention, and lifestyle intervention of gestational diabetes mellitus in China. Int. J. Environ. Res. Public Health 17, 9517. doi: 10.3390/ijerph17249517
Kuang, Y. S., Lu, J.-H., Li, S.-H., Li, J.-H., Yuan, M.-Y., He, J.-R., et al. (2017). Connections between the human gut microbiome and gestational diabetes mellitus. GigaScience 6, 1–12. doi: 10.1093/gigascience/gix058
Lang, S., Martin, A., Zhang, X., Farowski, F., Wisplinghoff, H., Vehreschild, M. J. G. T., et al. (2021). Combined analysis of gut microbiota, diet and PNPLA3 polymorphism in biopsy-proven non-alcoholic fatty liver disease. Liver Int. 41, 1576–1591. doi: 10.1111/liv.14899
Langille, M. G. I., Zaneveld, J., Caporaso, J. G., McDonald, D., Knights, D., Reyes, J. A., et al. (2013). Predictive functional profiling of microbial communities using 16S RRNA marker gene sequences. Nat. Biotechnol. 31, 814–821. doi: 10.1038/nbt.2676
Larsen, N., Vogensen, F. K., van den Berg, F. W. J., Nielsen, D. S., Andreasen, A. S., Pedersen, B. K., et al. (2010). Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One 5:e9085. doi: 10.1371/journal.pone.0009085
Lê, K.-A., Li, Y., Xu, X., Yang, W., Liu, T., Zhao, X., et al. (2013). Alterations in fecal Lactobacillus and Bifidobacterium species in type 2 diabetic patients in southern China population. Front. Physiol. 3, 496. doi: 10.3389/fphys.2012.00496
Li, H., Xie, J., Guo, X., Yang, G., Cai, B., Liu, J., et al. (2022). Bifidobacterium Spp. and their metabolite lactate protect against acute pancreatitis via inhibition of pancreatic and systemic inflammatory responses. Gut Microbes 14, 1–23. doi: 10.1080/19490976.2022.2127456
Li, G., Yin, P., Chu, S., Gao, W., Cui, S., Guo, S., et al. (2021). Correlation analysis between GDM and gut microbial composition in late pregnancy. J. Diabetes Res. 2021, 1–17. doi: 10.1155/2021/8892849
Li, M., Zhang, G., Cui, L., Zhang, L., Zhou, Q., Mu, C., et al. (2023). Dynamic changes in gut microbiota during pregnancy among Chinese women and influencing factors: a prospective cohort study. Front. Microbiol. 14, 1–10. doi: 10.3389/fmicb.2023.1114228
Li, H.-Y., Zhou, D.-D., Gan, R.-Y., Huang, S.-Y., Zhao, C.-N., Shang, A., et al. (2021). Effects and mechanisms of probiotics, prebiotics, Synbiotics, and Postbiotics on metabolic diseases targeting gut microbiota: a narrative review. Nutrients 13, 3211. doi: 10.3390/nu13093211
Liu, N., Sun, Y., Wang, Y., Ma, L., Zhang, S., and Lin, H. (2023). Composition of the intestinal microbiota and its variations between the second and third trimesters in women with gestational diabetes mellitus and without gestational diabetes mellitus. Front. Endocrinol. 14, 1–10. doi: 10.3389/fendo.2023.1126572
Lowe, W. L., Scholtens, D. M., Kuang, A., Linder, B., Lawrence, J. M., Lebenthal, Y., et al. (2019). Hyperglycemia and adverse pregnancy outcome follow-up study (HAPO FUS): maternal gestational diabetes mellitus and childhood glucose metabolism. Diabetes Care 42, 372–380. doi: 10.2337/dc18-1646
Ma, S., You, Y., Huang, L., Long, S., Zhang, J., Guo, C., et al. (2020). Alterations in gut microbiota of gestational diabetes patients during the first trimester of pregnancy. Front. Cell. Infect. Microbiol. 10, 58. doi: 10.3389/fcimb.2020.00058
Mao, L., Gao, B., Chang, H., and Shen, H. (2024). Interaction and metabolic pathways: elucidating the role of gut microbiota in gestational diabetes mellitus pathogenesis. Meta 14:43. doi: 10.3390/metabo14010043
McDonald, D., Jiang, Y., Balaban, M., Cantrell, K., Zhu, Q., Gonzalez, A., et al. (2024). Greengenes2 unifies microbial data in a single reference tree. Nat. Biotechnol. 42, 715–718. doi: 10.1038/s41587-023-01845-1
Medici Dualib, P., Ogassavara, J., Mattar, R., Mariko Koga da Silva, E., Atala Dib, S., and de Almeida Pititto, B. (2021). Gut microbiota and gestational diabetes mellitus: a systematic review. Diabetes Res. Clin. Pract. 180:109078. doi: 10.1016/j.diabres.2021.109078
Mitanchez, D., Yzydorczyk, C., Siddeek, B., Boubred, F., Benahmed, M., and Simeoni, U. (2015). The offspring of the diabetic mother – short- and long-term implications. Best Pract. Res. Clin. Obstet. Gynaecol. 29, 256–269. doi: 10.1016/j.bpobgyn.2014.08.004
Mokkala, K., Vahlberg, T., Houttu, N., Koivuniemi, E., and Laitinen, K. (2020). Distinct metabolomic profile because of gestational diabetes and its treatment mode in women with overweight and obesity. Obesity 28, 1637–1644. doi: 10.1002/oby.22882
Palmnäs-Bédard, M. S. A., Costabile, G., Vetrani, C., Åberg, S., Hjalmarsson, Y., Dicksved, J., et al. (2022). The human gut microbiota and glucose metabolism: a scoping review of key bacteria and the potential role of SCFAs. Am. J. Clin. Nutr. 116, 862–874. doi: 10.1093/ajcn/nqac217
Pathak, P., Xie, C., Nichols, R. G., Ferrell, J. M., Boehme, S., Krausz, K. W., et al. (2018). Intestine Farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid Receptor-1 signaling to improve metabolism. Hepatology 68, 1574–1588. doi: 10.1002/hep.29857
Pedersen, H. K., Gudmundsdottir, V., Nielsen, H. B., Hyotylainen, T., Nielsen, T., Jensen, B. A. H., et al. (2016). Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 535, 376–381. doi: 10.1038/nature18646
Pinto, Y., Frishman, S., Turjeman, S., Eshel, A., Nuriel-Ohayon, M., Shrossel, O., et al. (2023). Gestational diabetes is driven by microbiota-induced inflammation months before diagnosis. Gut 72, 918–928. doi: 10.1136/gutjnl-2022-328406
Ponzo, V., Fedele, D., Goitre, I., Leone, F., Lezo, A., Monzeglio, C., et al. (2019). Diet-gut microbiota interactions and gestational diabetes mellitus (GDM). Nutrients 11, 330. doi: 10.3390/nu11020330
Rold, L. S., Bundgaard-Nielsen, C., Niemann Holm-Jacobsen, J., Glud Ovesen, P., Leutscher, P., Hagstrøm, S., et al. (2022). “Characteristics of the gut microbiome in women with gestational diabetes mellitus: a systematic review”. PLoS One 17:e0262618. doi: 10.1371/journal.pone.0262618
Roselli, M., Devirgiliis, C., Zinno, P., Guantario, B., Finamore, A., Rami, R., et al. (2017). Impact of supplementation with a food-derived microbial community on obesity-associated inflammation and gut microbiota composition. Genes Nutr 12, 1–12. doi: 10.1186/s12263-017-0583-1
Sheiner, E. (2020). Gestational diabetes mellitus: Long-term consequences for the mother and child grand challenge: how to move on towards secondary prevention? Front. Clin. Diabetes Healthc. 1, 546256. doi: 10.3389/fcdhc.2020.546256
Su, Y., Wang, H. K., Gan, X. P., Chen, L., Cao, Y. N., Cheng, D. C., et al. (2021). Alterations of gut microbiota in gestational diabetes patients during the second trimester of pregnancy in the Shanghai Han population. J. Transl. Med. 19, 1–13. doi: 10.1186/s12967-021-03040-9
Sun, Z., Pan, X.-F., Li, X., Jiang, L., Hu, P., Wang, Y., et al. (2023). The gut microbiome dynamically associates with host glucose metabolism throughout pregnancy: longitudinal findings from a matched case-control study of gestational diabetes mellitus. Adv. Sci. 10, 1–12. doi: 10.1002/advs.202205289
Sweeting, A., Wong, J., Murphy, H. R., and Ross, G. P. (2022). A clinical update on gestational diabetes mellitus. Endocr. Rev. 43, 763–793. doi: 10.1210/endrev/bnac003
Takagi, T., Naito, Y., Kashiwagi, S., Uchiyama, K., Mizushima, K., Kamada, K., et al. (2020). Changes in the gut microbiota are associated with hypertension, hyperlipidemia, and type 2 diabetes mellitus in Japanese subjects. Nutrients 12:2996. doi: 10.3390/nu12102996
Tanaka, K., Harata, G., Miyazawa, K., He, F., Tanigaki, S., and Kobayashi, Y. (2022). The gut microbiota of non-obese Japanese pregnant women with gestational diabetes mellitus. Biosci. Microbiota Food Health 41, 4–11. doi: 10.12938/bmfh.2021-025
Tang, J., Lingqi, X., Zeng, Y., and Gong, F. (2021). Effect of gut microbiota on LPS-induced acute lung injury by regulating the TLR4/NF-KB signaling pathway. Int. Immunopharmacol. 91:107272. doi: 10.1016/j.intimp.2020.107272
Vavreckova, M., Galanova, N., Kostovcik, M., Krystynik, O., Ivanovova, E., Roubalova, R., et al. (2022). Specific gut bacterial and fungal microbiota pattern in the first half of pregnancy is linked to the development of gestational diabetes mellitus in the cohort including obese women. Front. Endocrinol. 13, 970825. doi: 10.3389/fendo.2022.970825
Wang, S., Cui, Z., and Yang, H. (2024). Interactions between host and gut microbiota in gestational diabetes mellitus and their impacts on offspring. BMC Microbiol. 24, 1–17. doi: 10.1186/s12866-024-03255-y
Wang, X., Liu, H., Li, Y., Huang, S., Zhang, L., Cao, C., et al. (2020). Altered gut bacterial and metabolic signatures and their interaction in gestational diabetes mellitus. Gut Microbes 12, 1–13. doi: 10.1080/19490976.2020.1840765
Wang, D., Wei, T., Zhao, F., and Huang, J. (2024). Fertility policy changes, maternal and foetal characteristics and birth timing patterns at a tertiary referral Centre in Beijing: a ten-year retrospective study. BMJ Open 14:e076987. doi: 10.1136/bmjopen-2023-076987
Wang, J., Zheng, J., Shi, W., du, N., Xu, X., Zhang, Y., et al. (2018). Dysbiosis of maternal and neonatal microbiota associated with gestational diabetes mellitus. Gut 67, 1614–1625. doi: 10.1136/gutjnl-2018-315988
Wei, J., Qing, Y., Zhou, H., Liu, J., Qi, C., and Gao, J. (2021). 16S RRNA gene amplicon sequencing of gut microbiota in gestational diabetes mellitus and their correlation with disease risk factors. J. Endocrinol. Investig. 45, 279–289. doi: 10.1007/s40618-021-01595-4
Ye, D., Huang, J., Wu, J., Xie, K., Gao, X., Yan, K., et al. (2023). Integrative metagenomic and Metabolomic analyses reveal gut microbiota-derived multiple hits connected to development of gestational diabetes mellitus in humans. Gut Microbes 15, 1–17. doi: 10.1080/19490976.2022.2154552
Ye, G., Zhang, L., Wang, M., Chen, Y., Gu, S., Wang, K., et al. (2019). The gut microbiota in women suffering from gestational diabetes mellitus with the failure of glycemic control by lifestyle modification. J. Diabetes Res. 2019, 1–12. doi: 10.1155/2019/6081248
Zhang, J., Ma, S., Wu, S., Guo, C., Long, S., and Tan, H. (2019). Effects of probiotic supplement in pregnant women with gestational diabetes mellitus: a systematic review and meta-analysis of randomized controlled trials. J. Diabetes Res. 2019, 1–12. doi: 10.1155/2019/5364730
Zhang, H., Qi, C., Zhao, Y., Lu, M., Li, X., Zhou, J., et al. (2021). Depletion of gut secretory immunoglobulin a coated Lactobacillus Reuteri is associated with gestational diabetes mellitus-related intestinal mucosal barrier damage. Food Funct. 12, 10783–10794. doi: 10.1039/D1FO02517A
Zheng, W., Xu, Q., Huang, W., Yan, Q., Chen, Y., Zhang, L., et al. (2020). Gestational diabetes mellitus is associated with reduced dynamics of gut microbiota during the first half of pregnancy. mSystems 5, e00109–20. doi: 10.1128/mSystems.00109-20
Zhong, H., Ren, H., Lu, Y., Fang, C., Hou, G., Yang, Z., et al. (2019). Distinct gut metagenomics and metaproteomics signatures in prediabetics and treatment-Naïve type 2 diabetics. EBioMedicine 47, 373–383. doi: 10.1016/j.ebiom.2019.08.048
Keywords: gestational diabetes mellitus, gut microbiota, body mass index, obesity, inflammation
Citation: Zhong S, Yang B, Liu Y, Dai W, Li G, Yang J, Yang A, Wang Y, Wang M, Xu C and Deng Y (2024) Dynamic changes of gut microbiota between the first and second trimester for women with gestational diabetes mellitus and their correlations with BMI: a nested cohort study in China. Front. Microbiol. 15:1467414. doi: 10.3389/fmicb.2024.1467414
Edited by:
Shenghui Li, Puensum Genetech Institute, ChinaReviewed by:
Cuauhtemoc Licona-Cassani, Monterrey Institute of Technology and Higher Education (ITESM), MexicoCopyright © 2024 Zhong, Yang, Liu, Dai, Li, Yang, Yang, Wang, Wang, Xu and Deng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuqing Deng, ZGVuZ3l1cWluZzE4NTJAMTYzLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.