
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 03 February 2025
Sec. Microbe and Virus Interactions with Plants
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1463326
This article is part of the Research TopicAdvancing Mycorrhizal Research for Sustainable Ecosystem and Agricultural PracticesView all 5 articles
 Patricia Battie-Laclau1*†
Patricia Battie-Laclau1*† Adrien Taudière2†
Adrien Taudière2† Mathilde Bernard1
Mathilde Bernard1 Lucas Bodénan1
Lucas Bodénan1 Myriam Duchemin1
Myriam Duchemin1 Yvan de Roman1
Yvan de Roman1 Azimê Yol1
Azimê Yol1 Dominique Barry-Etienne1
Dominique Barry-Etienne1Background: Nature-based management of vineyards is at the heart of a sustainable development for the next decades. Although much is known about grapevine benefits from Arbuscular Mycorrhizal Fungi (AMF), little is known about the influence of vineyard terroir and farming practices on AMF communities.
Methods: We examined the relative effect of wine terroir and agricultural practices (organic, conversion, and conventional) on AMF abundance and diversity across 75 vineyards distributed over 14 wine terroirs in 6 winegrowing regions in France. We estimate AMF abundance by measuring spore density and root mycorrhization rates, and characterize AMF communities composition using metabarcoding by sampling both root and spore compartments for each vineyard.
Results: Organic farming slightly increases AMF abundance (spore density and mycorrhization rate). Vineyards under conversion and using organic practices display a higher AMF diversity than conventional ones. Terroirs vary widely in terms of AMF abundance and diversity, with the median of OTUs count per sample ranging from 9 (Côte des Blancs) to 35 (Gigondas). The composition of AMF communities is structured mainly by terroir and in a lesser extent by practice. The effect of terroir on AMF communities is partially explained by distance decay and soil properties, but the majority of variation is still explained only by the terroir identity. Organic practices improve both abundance and diversity of AMF in vineyards, possibly leading to more productivity and resilience of grapevines.
Conclusion: This large-scale study highlights the importance of terroir in our understanding of vineyard microbiome and paves the way to incorporation of AMF in microbial terroir studies and applications.
Recently, the negative impact of intensive agriculture on the environment and human health has led to a reevaluation of agricultural systems. Agroecology aims to improve agricultural production by employing natural processes, thereby reducing the reliance on synthetic inputs. The objective of agroecology is to utilize ecological processes and ecosystem services to develop and implement agricultural practices (Wezel et al., 2020). Nature-based management of the agroecosystem is at the heart of sustainable development for the next decades (Foley et al., 2011). The functioning of these agroecosystems is contingent upon soil microorganisms, which play a pivotal role in the health of soils, plants, animals, and humans (Banerjee and van der Heijden, 2023).
Vineyards combine a high economic and cultural value. France is the first grape producer globally, with an output of approximately 6 million tons, representing 20% of the production in the world (IOV, 2023). However, wine growers are facing numerous challenges, caught between stronger environmental constraints (e.g., higher levels of heat and water stress), economic viability and pressure to use fewer chemical inputs. Thanks to the great biodiversity they harbor (Paiola et al., 2020), organic vineyards support a wide array of ecosystem services (Winkler et al., 2017). Therefore, a sustainable viticulture will require a good comprehension of the grapevine microbiome, in order to adapt management practices, including possible microbial inoculation (Ochoa-Hueso et al., 2024).
The endomycorrhizal interaction between grapevine and arbuscular mycorrhizal fungi (AMF) is essential for both partners and soil health (Trouvelot et al., 2015). The benefits of AMF for grapevines are manifold and include enhanced nutrient uptake, improved access to water resources and protection from pathogens (see Torres et al., 2018; Aguilera et al., 2022 for a comprehensive list of benefits). AMF are also a key ecological compartment to maintain berry quality in grapevines under changing environments (Torres et al., 2018). However, AMF effectiveness may vary with changes in environmental conditions. Indeed, the beneficial effects of AMF can be exacerbated by conditions of limited nutrient and water availability in different ways. Resource availability can directly influence AMF growth and diversity. Studies have shown that increased precipitation reduces the density of AMF extra-radical hyphae (Wang et al., 2021), and, inversely, drought conditions increase sporulation and AMF diversity (Jafarian et al., 2024). Additionally, the alteration of host plants physiology and diversity in response to constraint conditions can affect AMF by selecting species as a function of stress (Biasi et al., 2023).
The diversity and composition of AMF communities are shaped by numerous variables, including plant species and genotypes (Guzman et al., 2021; Martin and van der Heijden, 2024). Environmental factors also shape AMF communities through both biotic (e.g., competition among AMF, Maherali and Klironomos, 2007) and abiotic (e.g., soil copper concentration, Betancur-Agudelo et al., 2023) characteristics. Biotic and abiotic factors can also interact in shaping AMF communities (Frew and Aguilar-Trigueros, 2023).
Likewise, AMF communities are affected by agricultural practices such as monoculture, soil tillage and elevated levels of fertilizer or biocide inputs (Trouvelot et al., 2015). Monocultures apply a strong negative selection pressure on biodiversity, including AMF, resulting in communities dominated by few taxa, better adapted to intensive agricultural practices (Verbruggen et al., 2010). Given the vulnerability of AMF mycelial networks, tillage has been demonstrated to affect AMF spore density, species richness and diversity (Säle et al., 2015; Thomopoulos et al., 2023). Indeed, deep plowing can result in the disappearance of certain species or the dispersal of their propagules to a deeper layer of soil, thereby reducing the level of root colonization. It has also been shown that synthetic herbicides harm root mycorrhization in herbal plant species (Zaller et al., 2018) and grapevines (Baumgartner et al., 2005) compared to mechanical weeding. Chemical components exert direct effects on AMF development and physiological metabolism (Yu et al., 2023).
Conversely, the establishment of cover crops promotes the proliferation of natural mycorrhizal communities (Rivera-Becerril et al., 2017). As an alternative to herbicides and to soil tillage for weed control, cover crops provide benefits to AMF communities by preventing soil disturbance and consequently mycelial destruction. Similarly, as AMF have a positive effect on plant diversity (van der Heijden, 2002), cultural practices based on high crop diversity enrich AMF communities (Guzman et al., 2021).
In the field of viticulture, the distinctions between agricultural systems reside in specific practices employed, generally related to the use of biocide (pesticides, fungicides, and herbicides). However, some practices can be shared by various agricultural systems, such as tillage, use of cover or organic inputs such as compost. As conventional agriculture uses large quantities of chemical fertilizers, biocides, and tillage to maximize crop yields, AMF spore density (Lin et al., 2020), species richness and diversity (Sheng et al., 2013; Oehl et al., 2004) tend to decline in comparison to organic plots. On the contrary, organic farming promotes AMF proliferation (Hart and Reader, 2002; Radić et al., 2014), colonization activity (Jiang et al., 2020) and AMF diversity (Oehl et al., 2004; Lin et al., 2012). The extensive use of organic inputs may counteract the detrimental effect of intensive tillage in organic farming, which is often used to replace herbicides (Rivera-Becerril et al., 2017; Van Geel et al., 2017).
We use the official definition of terroir as: “a concept which refers to an area in which collective knowledge of the interactions between the identifiable physical and biological environment and applied vitivinicultural practices develops, providing distinctive characteristics for the products originating from this area. Terroir includes specific soil, topography, climate, landscape characteristics and biodiversity features” (Resolution OIV/Viti 333/2010). The terroir concept can be considered a “black box” we need to explore (Brillante et al., 2020).
Although literature about terroir abounds, the incorporation of the microbial components of the terroir is poorly documented (Bokulich et al., 2016; Gilbert et al., 2014). A number of these studies concentrate upon the impact of microbes involved in the sulfur cycle or yeasts on the characteristics of wine (Mocali et al., 2020). The specific contributions of individual terroir components in explaining AMF community composition variation remain poorly understood. Recently, in a study of 200 vineyards, Gobbi et al. (2022) found that spatial distance was the primary explanatory variable for beta diversity of fungal and prokaryotic communities, at both global and local scales. The diversity of AMF is mainly influenced by the direct effects of climate (Jafarian et al., 2024). Inversely, according to Betancur-Agudelo et al. (2021), the most impactful component of terroir is soil properties, as soil directly influences nutrient availability and microbial habitats.
There is a paucity of data regarding the significance of AMF as a terroir component. Torres et al. (2019) postulated that infection with AMF might augment the amino acid content of the grapes, which may in turn affect the aromatic characteristics of the wine. This raises the question of the influence of terroir on AMF communities in vineyards across the country and how this terroir effect will interact with new ecoagricultural practices such as application of AMF-based biostimulants (Jindo et al., 2022). In order to utilize AMF-based biostimulants in a manner that does not adversely affect the natural AMF communities of a terroir, it is essential to evaluate the influence of soil properties, geographical distance, and other terroir characteristics on the communities that are naturally present in vineyard soils.
We aimed to quantify the effect of agricultural practices and terroirs on AMF communities. This was achieved by examining four key facets of AMF ecology:
i. Spore numbers.
ii. Mycorrhization rate.
iii. Alpha-diversity (local AMF diversity).
iv. Beta-diversity (change in AMF communities’ composition across samples).
The objective of our study was to investigate the abundance and diversity of AMF within the French vineyard. To this end, we conducted a comprehensive analysis of 75 different vineyards, distributed across 14 distinct wine terroirs and encompassing six major wine-producing regions in France: Bordeaux, Bourgogne, Camargue, Champagne, Côtes-du-Rhône, and Languedoc. Our study considers a wide range of pedoclimatic and management conditions, as detailed in Supplementary Table S1. Each terroir refers to a specific protected designation of origin (AOP), controlled designation of origin (AOC) or protected geographical indication (PGI) according to European Union regulation N° 1308/2013 dated 17th December 2013. Vineyard plots were selected in each terroir based on their agricultural practices (conventional, conversion, organic) and weed management in row and inter-row (chemical control by application of herbicide, mechanical control by scratching or plowing, grassed). The conventional plots primarily utilized synthetic products, though sulfur and copper treatments were intermittently employed, along with mineral fertilization. The organic plots were managed in accordance with the European Union Regulation (EEC) No. 834/2007, which excludes synthetic pesticides and inorganic fertilizers. The plots undergoing conversion to organic were in the first, second, or third year of conversion.
This large-scale study was conducted out over several years, spanning from 2019 to 2023. All plots within the same terroir were sampled in the same year. All soils and roots in each of the 75 plots were sampled at the same time, from late June to early July, regardless of the year. Indeed, early summer in France corresponds to grapevine fruit set, when arbuscular colonization reaches its highest level (Schreiner, 2005). Some previous studies carried out on AMF diversity in vineyards have had to deal with plots that had received AMF-based biostimulants (e.g., Bouffaud et al., 2016a). To avoid any bias in AMF communities’ composition, we verified beforehand with winegrowers that no fields had been inoculated with AMF in the past. In each of the 75 vineyards, we counted spores, measured mycorrhization rates, characterized AMF communities using metabarcoding. Additionally, we analyzed physicochemical soil characteristics for 49 vineyards.
In each vineyard, a total of 12 plants per plot were sampled by selecting 4 consecutive plants in three homogenous rows across the vineyard, avoiding border rows. Soil (approximately 150 g) and grapevine roots (enough to fill a 2 mL tube) were sampled early after fruit set (between late June and early July) at a soil depth of 20 cm at the base of each selected plant. Only thin and young grapevine roots with approximately 1 mm in diameter were harvested. Grapevine black roots are easily visually distinguishable from the roots of other plant species. Soil and root samples were combined in 3 pools from 4 neighbors’ plants, each totalizing 3 soil and root composite samples per plot. For all Vergèze and Côte des Blancs plots, all 12 root samples were pooled into a single composite sample (Figure 1).
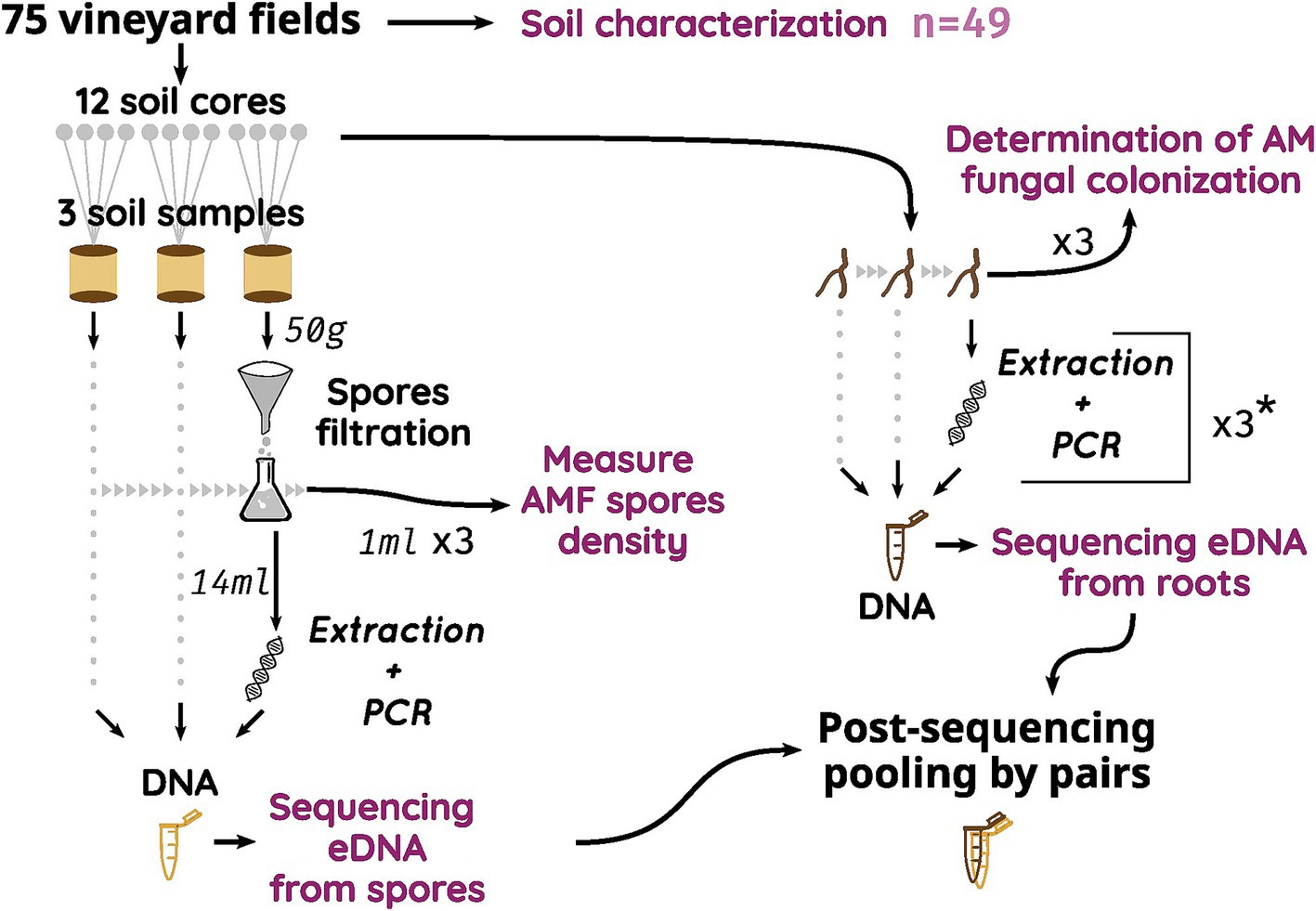
Figure 1. Schematic sampling strategy of the two AMF compartments. * For samples in Vergèze and Côte des Blancs terroirs, all three root samples were pooled before the unique extraction but 3 PCR were used to amplify DNA. eDNA, environmental DNA; AMF, Arbuscular Mycorrhizal Fungi.
All samples (soil and living roots) were stored in plastic bags at 4°C until processing, which was conducted within 1 week. Each root sample was washed free of soil and then divided into two portions. The first portion (a random subset of sixteen 1-cm-long root segments) was submerged in 70% ethanol and stored at 4°C before being colored for AM fungal colonization determination. The second portion was stored at −20°C until DNA extraction and subsequent molecular analyses of AM fungal communities. A 200 g soil aliquot was separated from each soil replicate sample, sieved at 2 mm and reserved for spore density and diversity analysis. To reach a good representation of AMF spore bank diversity, all spores found in soil samples were sorted out before DNA extraction. As we want to study the AMF communities in the Vitis vinifera rhizosphere as a whole, we combined environmental DNA (eDNA) samples from roots and from the spore bank (Figure 1) at the end of the bioinformatic pipeline.
For each plot (with the exception of some Vergèze plots and all plots of Côte des Blancs), soil physicochemical properties were analyzed from a 500 g soil aliquot. The resulting 49 soil samples were analyzed in the Laboratoire d’Analyses de Terres, de Végétaux et Environnementales of the Chambre d’Agriculture de l’Aude (France). In brief, air-dried soil was sieved through 2 mm sieves. In 1:5 soil-to-water (w/v) suspension, the pH and electrical conductivity (EC) of the soil were determined using a pH-EC meter. Particle-size distribution was determined by the hydrometer method (Bouyoucos, 1962). Organic carbon was determined using the wet-oxidation method by Walkley and Black and expressed as organic matter using Vant Hoff’s factor (1.72). Total nitrogen was quantified using the Semi-Micro Kjeldahl method (Bremner and Mulvaney, 1982). The Joret-Hébert method was used for phosphorus extraction. Exchangeable bases, calcium (Ca), magnesium (Mg), potassium (K), and sodium (Na) were assessed by soil saturation with neutral 1 M ammonium acetate and measured via atomic absorption spectrophotometry (Sumner and Miller, 1996). Cation exchange capacity (CEC) was determined by the Kjeldahl distillation method. The determination of available micronutrients, iron (Fe), copper (Cu), manganese (Mn) and zinc (Zn) involved diethylene triamine pentacetic acid (DTPA) (Lindsay and Norvell, 1978). Measurement of these micronutrients was performed using an atomic absorption spectrophotometer. EC1:2.5 was potentiometrically measured in a 1:2.5 soil-to-water ratio according to Okalebo et al. (2002).
AMF spores occurring in soil samples were extracted following the wet sieving method adapted from Gerdemann and Nicolson (1963) and Daniels and Skipper (1982). For each soil replicate sample, 50 g was sieved through three nested sieves with meshes of 1,000, 400, and 45 μm. Then, spores were purified by re-suspending the sieving in a 60% sucrose solution and centrifugation was carried out at 3,000 rpm for 2 × 3 min. The supernatant was removed and poured into the 45 μm sieve. Retrieved AMF spores were re-suspended in 15 mL of water. A 1 mL aliquot was placed in Petri dishes and spores were counted under a stereomicroscope (40 × magnification). Average numbers were calculated per 100 g of dry soil. The remaining 14 mL were stored at −20°C until DNA extraction and subsequent molecular analyses. As a result, each one of the 75 spore bank sample for eDNA diversity analysis correspond to spore bank of 140 g of soil (50 g × 14 mL / 15 mL × 3 replicates).
Sampled root fragments (3 replicates of 16 fragments of 1 cm long for each plot) were cleared in 10% KOH at ambient temperature for 10 h. Highly pigmented grapevine roots were additionally cleared in 3% w/v H2O2 for 40 min at 70°C and rinsed with distilled water. Root fragments were then colored with Schaeffer black ink, as described in Vierheilig et al. (1998). Thereafter, the samples were immersed in a mixture of 50% glycerol in water. Roots were mounted onto microscope slides and examined under 200–800 × magnification. The number of sections where mycorrhizal arbuscules, vesicles or hyphae were observed was noted separately for each structure type. For each replicate, the frequency of mycorrhiza (F %), root mycorrhization rate (M %), and arbuscular (A %) abundance in the root system were evaluated according to Trouvelot et al. (1986) using the MYCOCALC program.
AMF spores were extracted from each of the three soil replicate and ground in buffer solution (0.4 M NaCl, 10 mM Tris–HCl pH = 8, SDS 0.2%, 2 mM EDTA pH = 8). Spore DNA was extracted using the FastDNA Spin kit for Soil (MP Biomedicals, Europe) according to the manufacturer’s instructions. Elution of DNA was done using 200 μL of DNase-free water. The 3 DNA extracts replicates were mixed and diluted 1:50 in DNase-free water.
Root samples were ground in liquid nitrogen using a mortar and pestle. Genomic DNA was extracted from 250 mg of roots using the FastDNA Spin kit for Soil (MP Biomedicals, Europe) according to the manufacturer’s instructions. Elution of DNA was done using 100 μL of TE and stored at −20°C. The DNA extracts were diluted 1:10 in DNase-free water.
The DNA of arbuscular mycorrhizal fungi was amplified using 18S rRNA gene primers. The first PCR reactions were performed in triplicates. A fragment of 510–570 bp covering a variable region of the SSU was amplified using the universal eukaryotic primer NS31 (Simon et al., 1992; TTGGAGGGCAAGTCTGGTGCC) in combination with the AMF-specific primer AML2 (Lee et al., 2008; GAACCCAAACACTTTGGTTTCC) and included overhang adaptor sequences for the Nextera primer (Illumina Inc., CA, United States).
PCRs were performed in a total volume of 20 μL with 1 μL DNA, 0.2 μL of each specific primer (10 μM), 4 μL 5X Platinum II PCR buffer (Thermo Fisher, Massachusetts, Etats-Unis), 0.4 μL 10 mM dNTP mix, 0.32 μL Platinum II Taq Hot-Start DNA Polymerase, 13.88 μl of DNase-free water. The PCR cycle was as follows: 2 min at 94°C, (15 s at 94°C, 15 s at 55°C, 30 s at 72°C) for 35 cycles and a final elongation step at 72°C for 10 min. The PCR products were purified with magnetic beads (AMPure XP).
The second PCR was performed using a Nextera® XT Index Kit (Illumina, San Diego, United States) following the manufacturer’s instructions. After purification with magnetic beads (AMPure XP), these final PCR products were merged by triplicate (Figure 1), dosed with kit KAPA Library Quantification kit (Roche), multiplexed and sequenced on a MiSeq Illumina sequencer using MiSeq Reagent Kit v3 (600-cycle, Illumina).
DNA sequences were analyzed through the bioinformatics pipeline described in Supplementary Report S1. This pipeline draws 4.32 kWh (see Supplementary Note S1 for details) which results in a carbon footprint of 221 gCO2e (calculated using R package greenAlgoR, Taudière, 2024, algorithm based on Lannelongue et al., 2021). In short, primers were removed using cutadapt (v. 4.5, Martin, 2011). Sequences were quality filtered using filterAndTrim function from the dada2 package (v. 1.30.0; Callahan et al., 2016a) discarding sequences with default parameters. Then we followed dada2 classic pipeline (Callahan et al., 2016b) to obtain chimera-free amplicon sequence variants (ASV) using single forward (R1) sequences. Each ASV longer than 300 pb was then taxonomically assigned to two taxonomic databases with the assignTaxonomy function from dada2, which implements the RDP classifier of Wang et al. (2007). First, we used the PR2 database (v. 5.0.0; Guillou et al., 2012) to assign the taxonomy at the scale of Eukaryota. Second, we used the AMF specific database Maarjam (Öpik et al., 2010) to assign more specifically arbuscular mycorrhizal fungal OTUs.
Following recommendation by Tedersoo et al. (2022), we added a step of reclustering on ASV sequences to obtain a more classical version of OTU using the function asv2otu from the MiscMetabar package (v. 0.9.4; Taudière, 2023). The idea is to denoise using dada and then to cluster into taxonomic unit using vsearch software (v. 2.22.1; Rognes et al., 2016) at a 97% identity level. We also repeated the analysis on the ASV dataset in Supplementary Report S2 to identify potential differences in key results between the two approaches, as recommended by Joos et al. (2020).
After reclustering, we filtered out all non-AMF sequences using two filters. First, all sequences with less than 80% identity similarity with at least one sequence in Maarjam database were discarded (MiscMetabar::blast_pq function). Second, we also discarded OTUs assigned to other families than Mucoromycota by the PR2 database. Numbers of sequences across the major step are present in Table 1. Except for soil compartment analysis (spores vs. roots), we merged paired samples of spores and roots in one sample. We decided to pool root and spore samples because the two compartments bring different views on AMF communities, and we are interested in the whole rhizosphere of Vitis vinifera. Moreover, the different nature of the two compartments makes the result from metabarcoding difficult to compare and would bring a non-necessary level of complexity.
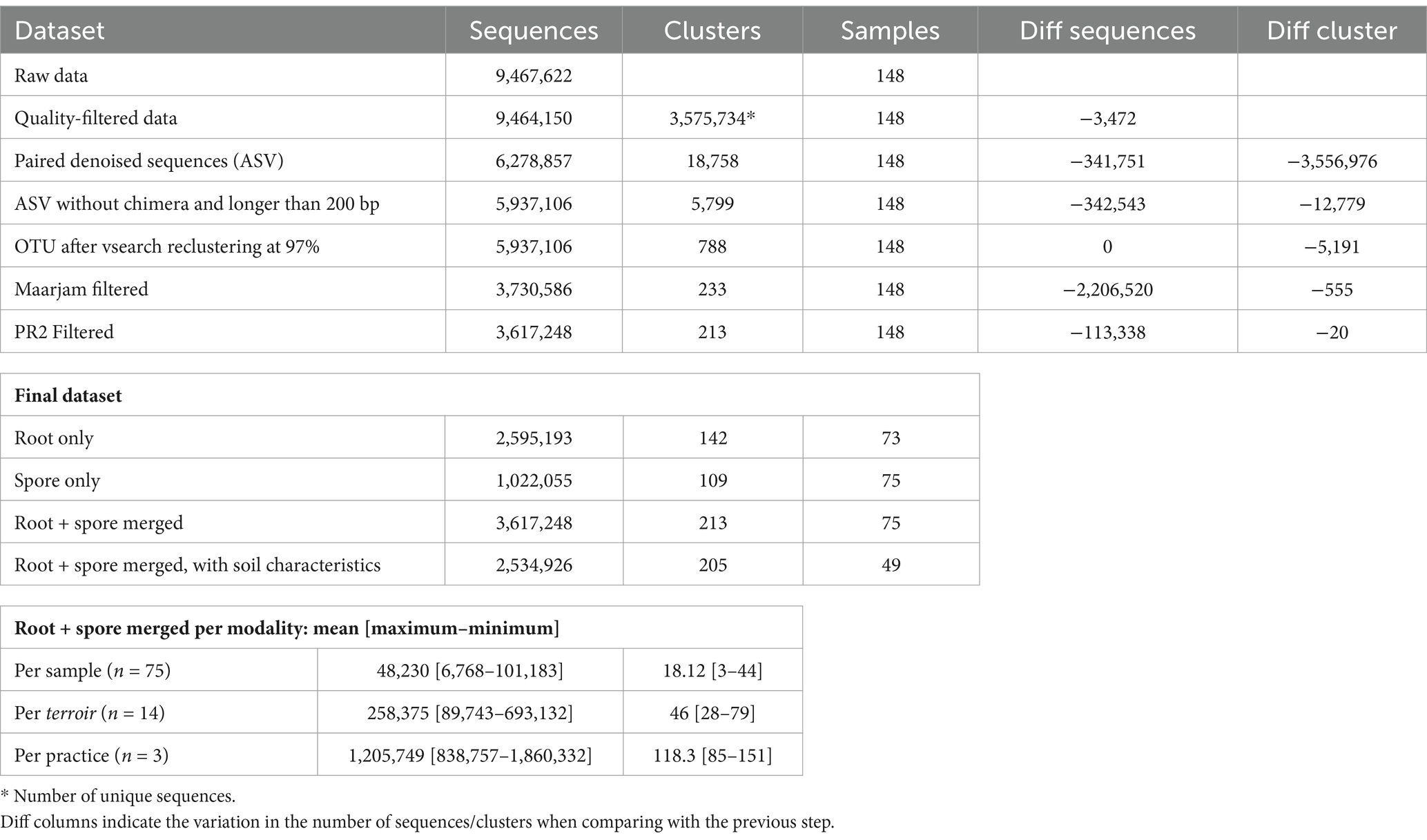
Table 1. Number of sequences, clusters (i.e., unique sequences, ASV or OTUs depending on the step) and samples across the main step of the bioinformatic pipeline.
All statistical analyses were carried out using R Studio software (Posit Team, 2024, version 2023.12.1) and R version 4.3.3 (R Core Team, 2023). Code for statistical analysis, tables, and figures is available in Supplementary Report S3. Most important packages are dada2 (v. 1.30; Callahan et al., 2016a), MiscMetabar (v. 0.9.4; Taudière, 2023), phyloseq (v. 1.46.0; McMurdie and Holmes, 2013), targets (v. 1.4.1 Landau, 2021), ggstatsplot (v. 0.12.3 Patil, 2021) and vegan (v. 2.6-4; Oksanen et al., 2022). Minimal graphical adjustments to improve the figures’ visibility were performed in Inkscape (Inkscape Team, 2023).
The local biodiversity of AMF (alpha-diversity) were assessed using the Hill number framework (Hill, 1973) recommended by Alberdi and Gilbert (2019) for DNA-based diversity analyses. The importance of the abundance distribution increases with increasing Hill order q. The Hill number for q = 0 (H0) is the richness, when q = 1 (H1), it is the exponential Shannon entropy and for q = 2 (H2), it is the inverse Simpson index.
To describe soil chemical properties, principal component analysis (PCA) was performed using ade4 R package (Dray and Dufour, 2007) and visualization with package FactoMiner (Le et al., 2008) and factoextra (Kassambara and Mundt, 2020). The PCAtest function from the PCAtest package (Camargo, 2024) was used to test for significance of PCA dimensions after correcting p-value for multiple-testing.
To study beta-diversity, we accounted for spatial autocorrelation in samples using distance-based Moran’s eigenvector maps (function dbmem from the adespatial package; Dray et al., 2023). The effect of terroir and practice on AMF beta-diversity was computed using a Permanova with the formula:
where nb_seq is the square roots of the number of reads per sample, MEM_1 and MEM_2 are the dbmem dimension; Dim.1, Dim.2, and Dim.3 correspond to the first three axes of soil PCA; practice corresponds to either organic, conventional, or conversion agricultural; rank and inter_rank correspond to weed management in the vineyard row and inter-row, terroir is the list of 14 terroirs.
Permanova were computed on bray-curtis distance. This allows us to consider differences in sample sequencing depth without discarding so many sequences. We show in supplementary tables the result of Permanova on bray-curtis distance after rarefaction, as well as the result of Permanova on robust-Aitchison distance. Test for multivariate homogeneity of groups dispersions (variances) were done using function vegan::betadisper (Supplementary Report S3).
Furthermore, we plotted beta-diversity results using Non-metric Multi-Dimensional Scaling (NMDS) ordination and upset plot (Lex et al., 2014). Indicator species were identified using the multipatt function from the indicspecies package (Cáceres and Legendre, 2009) with the IndVal.g metrics. Finally, soil, distance, terroir and practice influence on beta-diversity were measured using partition of variance of distance-based redundancy analysis function vegan::varpart (Oksanen et al., 2022). To circumvent the stochastic phenomenon of rarefaction, we ran 99 distinct rarefactions and report the mean value of adjust R2 only when at least 95% of rarefaction runs resulted in significant adjust R2 values (function MiscMetabar::var_par_rarperm_pq). As soil properties were only available for 49 samples, we computed Permanova and variance partioning on both the 75 samples without information about soil and the 49 samples with information about soil.
A PCA on 17 soil variables (Table 2) was computed for the 49 samples for which both soil properties and AM diversity are known. The first three dimensions (72.8% of variance explained) were selected using manual inspection of scree plot. The first axis contrasts samples with sand versus samples with clay, high nitrogen (N) and high carbon (C). The second axis mostly represents differences in phosphorus pentoxide (P2O5), copper (Cu) and silt. Lime, pH and coarse silt drive the third axis (Figure 2). The three first axes significantly vary across terroirs, but not practices (Supplementary Figure S1, Kruskal-Wallis test p-values in Supplementary Figure S1). There is also no effect of practice on copper concentration alone (Kruskal-Wallis χ2 = 0.61, p-value = 0.73, Supplementary Figure S2).
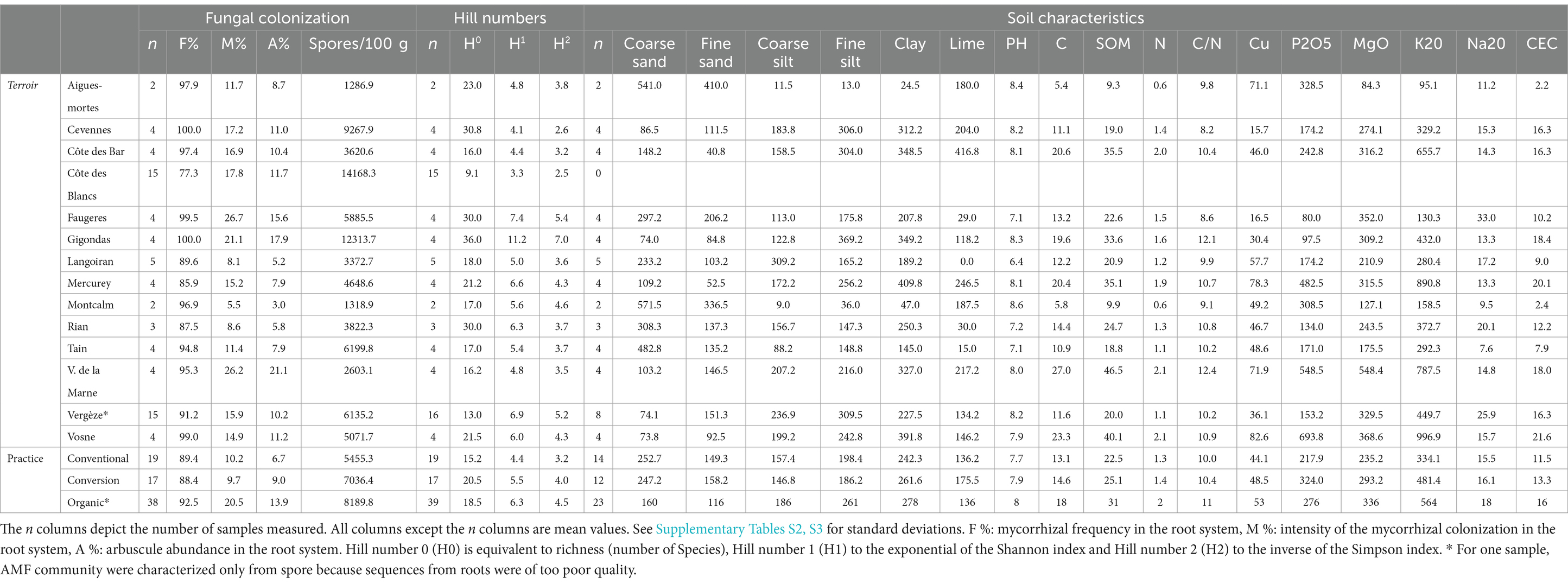
Table 2. Mycorrhization, diversity and soil characteristics for each terroir and practice.
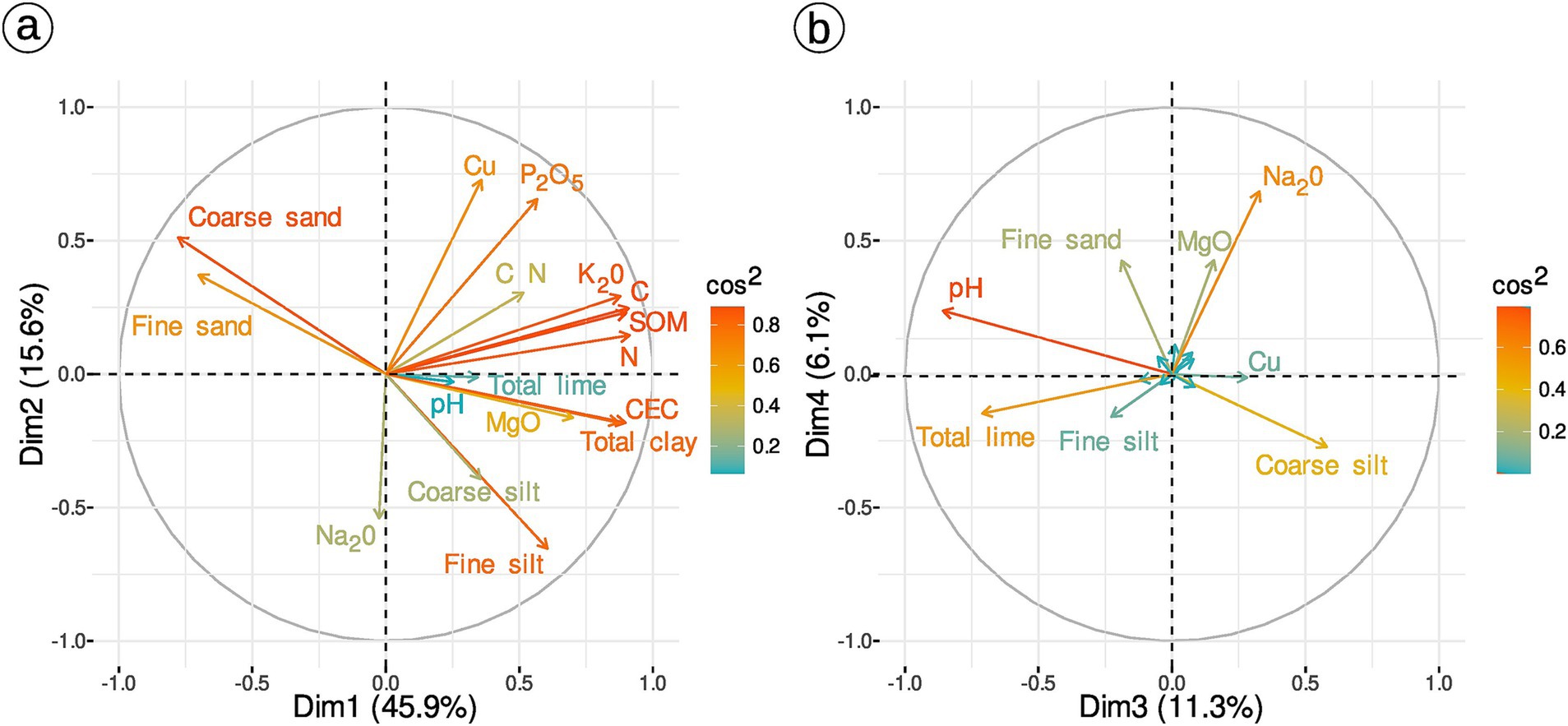
Figure 2. PCA of soil physicochemical variables (49 samples and 17 soil variables). Correlation graph of soil variables for the first two dimensions (A) and for dimension 3 and 4 (B).
There is no correlation between the number of spores and the number of AM fungal species (Spearman rank correlation test, p-value = 0.58; Supplementary Figure S3). We found no correlation between the three Hill numbers and the three mycorrhization measures A %, F %, and M % (Pearson test; Supplementary Figure S4). Spore counts are correlated with the three mycorrhization measures A % (Spearman test, p-value = 6e-3), F % (p-value = 0.001) and M % (p-value = 6e-4).
The number of spores significantly differs among terroirs (Kruskal-Wallis χ2 = 48.54, df = 14, p-value = 5e-6; Supplementary Figure S5). Using pairwise tests (Dunn with p-value Holm-adjustement), the terroir of Côte des Blancs shows a significant higher number of spores compared to Vergèze, Langoiran, Montcalm, Aigues-mortes and Vallée de la Marne. Mycorrhizal frequency (F %, p-value = 6e-10), intensity of the mycorrhizal colonization (M %, p-value = 1e-03), and arbuscule abundance in the root system (A %, p-value = 0.001) vary across terroirs.
The number of spores slightly varies with practices with organic vineyards presenting a significant higher number of spores than non-organic vineyards (Kruskal-Wallis, p-value = 0.044, Supplementary Figure S6). Organic agricultural practices (Kruskal-Wallis test and pairwise Dunn test) improve mycorrhizal frequency F % (mean = 92.5 vs. 88.4 for conversion and 89.4 for conventional; p-value = 0.046, Table 2), colonization intensity M % (mean = 20.5 vs. 9.7 for conversion and 10.2 for conventional; p-value = 5.3e-05), and arbuscules abundance A % (mean = 13.9 vs. 9 for conversion and 6.7 for conventional; p-value = 1e-03).
The bioinformatic pipeline identified 788 OTUs (5,937,106 sequences) from which 213 (3,617,248 sequences) were assigned to arbuscular mycorrhizal (AM) fungi using two criterions: 80% identity method to Maarjam database and PR2 assignation to Mucoromycota Family (Table 1). The first step filters out 37.1% of sequences and 70.4% of OTUs. Second, we also discarded OTUs assigned to other family than Mucoromycota in PR2, leading to an additional removal of 20 OTUs (113,338 sequences). The majority of non-Mucoromycota sequences are classified as Arthropoda (335 OTUs and 1,764,837 sequences), Nematoda (73 OTUs and 167,513 sequences), and Tardigrada (29 OTUs and 133,628 sequences, Supplementary Figures S7–S9). After merging paired spore and root samples, the sequencing depth varies from 6,768 to 101,183 (Table 1; mean = 48,230, sd = 19,620) AMF sequences per sample. Note that the 213 OTUs clustered a total of 3,695 ASV. All following results are robust to the decision to re-cluster ASV into OTUs (Supplementary Report S2).
Among the 9 families of the 213 OTUs, Glomeraceae is the most abundant (64.8% of OTUs, 81% of sequences) followed by Claroideoglomeraceae (8.5% of OTUs, 10.8% of sequences), Diversisporaceae (7% of OTUs, 3.9% of sequences), Paraglomeraceae (5.1% of OTUs, 2.5% of sequences) and Archaeosporaceae (5.1% of OTUs, 0.7% of sequences). The most abundant OTU is a Glomus species and accounts for 33% of the total number of sequences. Three Genus dominate the AMF communities using PR2 assignation: Glomus is the most abundant (42.3% of OTUs, 59.5% of sequences) followed by Rhizophagus (10.8% of OTUs, 18.8% of sequences) and Funneliformis (7.5% of OTUs, 12.7% of sequences).
Samples from spore and root compartments highly differ in terms of alpha and beta-diversity (Supplementary Figure S10). Root samples present a higher richness but similar hill numbers H1 and H2 (test de Mann–Whitney). For subsequent analysis, samples from roots and spores were merged by pairs leading to a total of 75 samples, 213 OTUs and 3,617,248 sequences (Table 1).
Terroir drives the AMF diversity (Table 2; Figure 3A; Supplementary Figure S11). The Gigondas terroir shows the highest richness (median = 33; 35.5 without rarefaction) whereas Côte des Blancs is the poorest (median = 9 with or without rarefaction). If we focus on the total diversity present in a given terroir, Cévennes (79 OTUs, Figure 4A) and Faugères (78 OTUs) present the highest diversity. Note that accumulation plots (Figure 3) are not here to assess the absolute soundness of our sampling because as our pipeline discards singletons, we care unable to draw a correct accumulation curve. However, comparison of curve shapes indicates that Rian, Faugères and Cévennes are not fully sampled with our sampling effort.
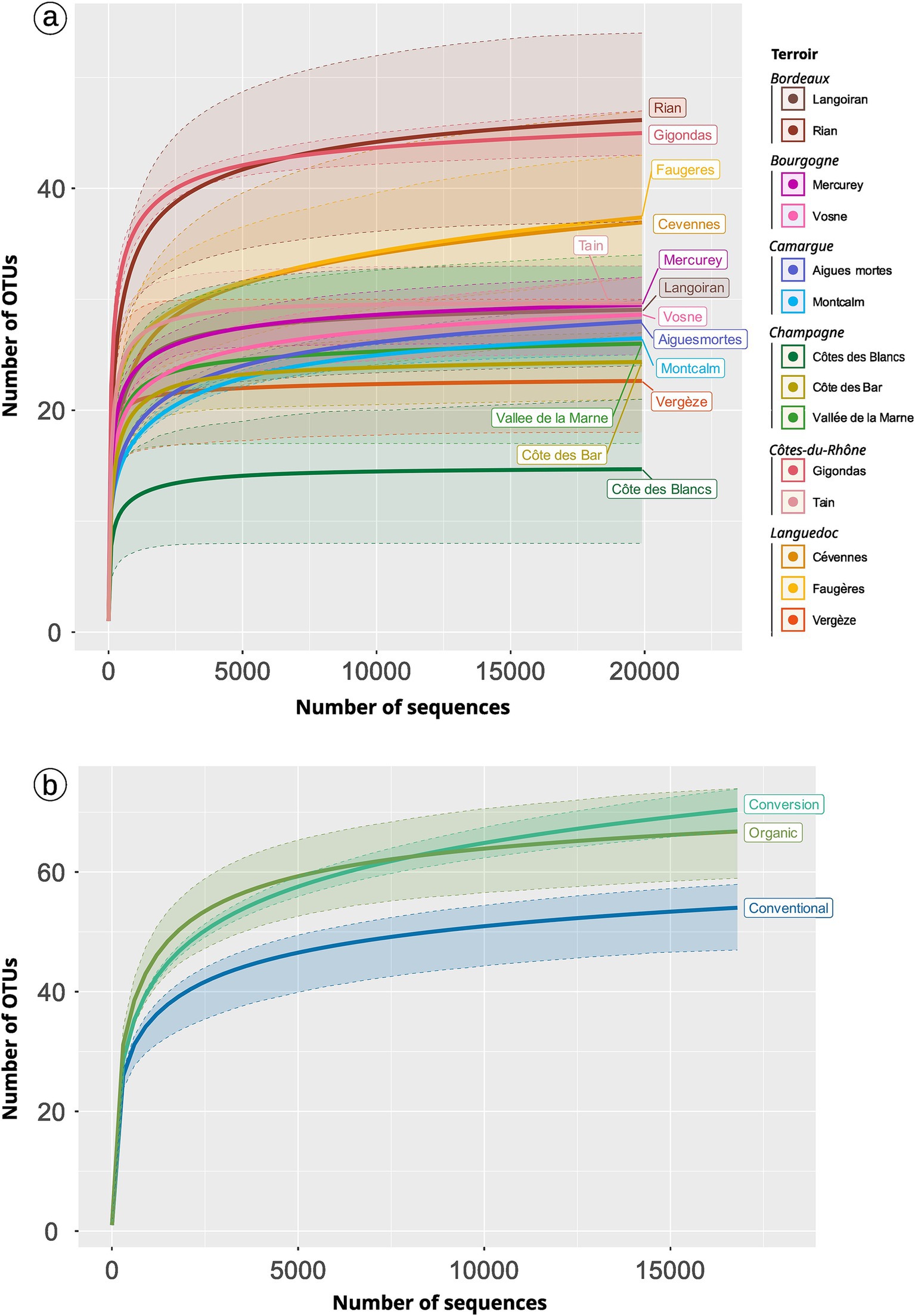
Figure 3. Pseudo-accumulation curves of AMF across terroirs (A) and practices (B). We rarefied the number of samples per modality as well as the total number of sequences per modality using 999 permutations (function MiscMetabar::accu_plot_balanced_modality). Filled areas show the 90% quantile distribution.
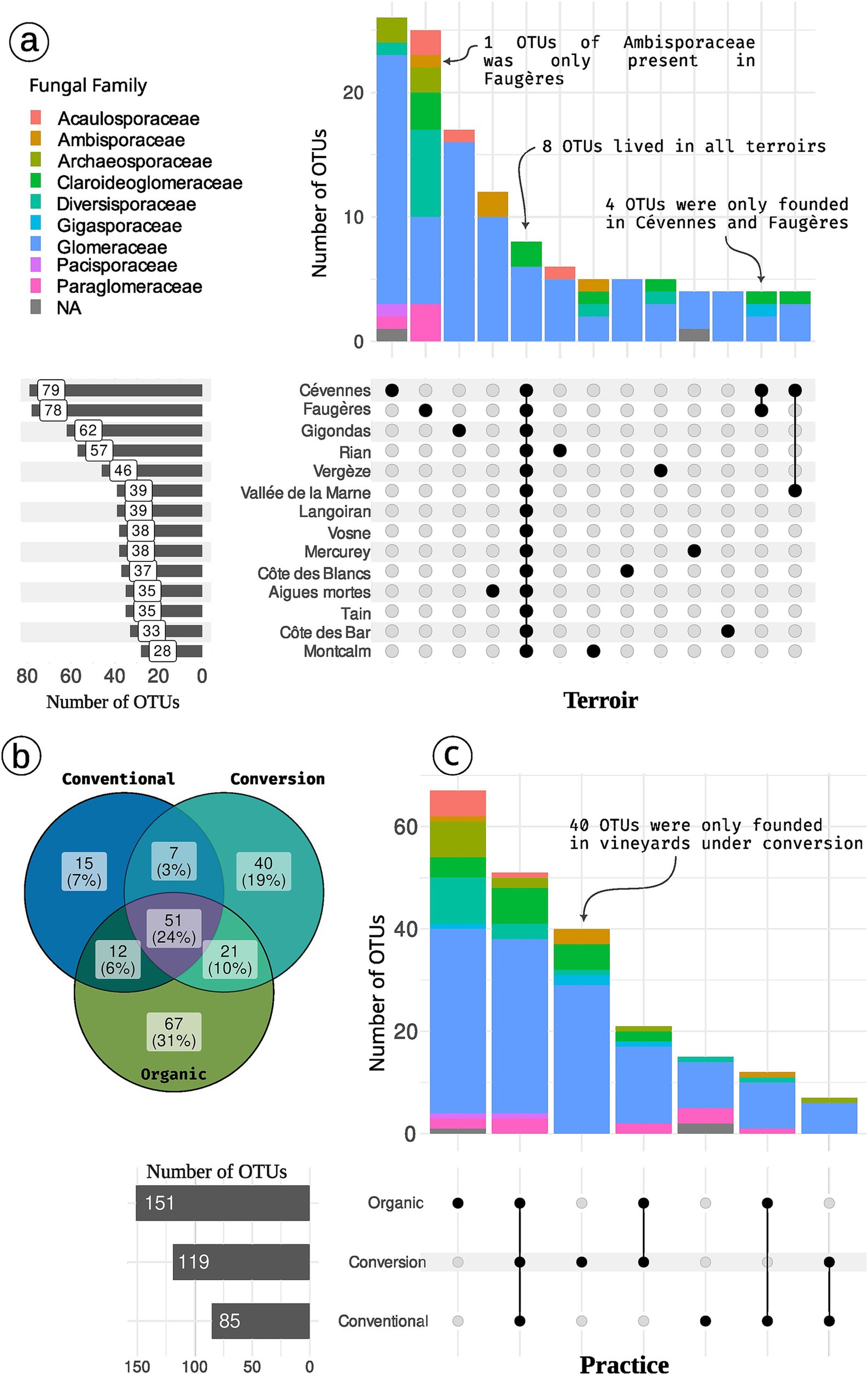
Figure 4. Distribution of OTUs across terroirs (A) and practices (B,C). Panel (B) is a venn diagramm depicting the number of shared OTUs. Panel (A,C) are upset plot. The matrix with point at the bottom of the figure represents the intersection between modalities (terroir or practice). In panel (A) only the case with at least 3 OTUs are plot. Colors depict OTUs Family. For example, 7 OTUs (6 Glomeraceae and 1 Archaeosporaceae) are found in vineyards under conversion and conventional practices but not in organic vineyards. Concerning terroir, 6 OTUs (5 Glomeraceae and 1 Acaulosporaceae) are specific to Rian. Cévennes terroir harbor 79 OTUs while 85 OTUs were found in all conventional farming samples.
Practices drive Hill number 1 and 2 (Figure 5 and Supplementary Figure S12 for analysis without rarefaction) with organic samples tending to harbor a higher AMF diversity than conventional ones. Focusing on the total diversity per farming practice gives a complementary vision for the richness facet. Organic vineyards and vineyards under conversion harbor a higher richness if we regroup samples per modality (Figures 4B, 3B). Organic samples display a total of 151 OTUs (39 samples), whereas conversion samples are represented by 119 OTUs (17 samples), and conventional ones by only 85 OTUs (19 samples). Thus, samples in organic and conversion vineyards harbor on average the same number of OTUs as conventional vineyards, but when considering all samples together, conventional vineyards harbor far less diversity.

Figure 5. Diversity of AMF across practices. Hill number 0 is equivalent to richness (number of Species), Hill number 1 to the exponential of the Shannon index and Hill number 2 to the inverse of the Simpson index.
Terroir is the major driver of AMF community composition, even if we control for soil properties and spatial autocorrelation (Table 3; Figures 6, 7). Spatial autocorrelation, and soil properties beyond terroir characteristics, also shape AMF communities. Eight OTUs were common to all 14 terroirs. Cévennes and Faugères terroirs, respectively, harbor 26 and 25 specific OTUs despite these terroirs being represented by only 4 samples. On the contrary, Côtes des blancs harbor only 5 unique OTUs despite 15 samples (Figure 4A). Moreover, among OTUs specific to Faugères, 7 out of 9 AMF families are present. Specific OTUs are OTUs found in only one terroir, but can be represented by only one sample. Thus, these specific OTUs may not be indicators of terroir if the number of samples is not sufficient to achieve statistical significance. Ten OTUs are indicators of terroir (Figure 8). Eight terroirs out of 14 present at least one indicator species. Gigondas (OTU_120, OTU_21, OTU_28 and OTU_65) and Aigues-mortes (OTU_198, OTU_43 and OTU_99) are the only terroirs characterized by specific indicator species.
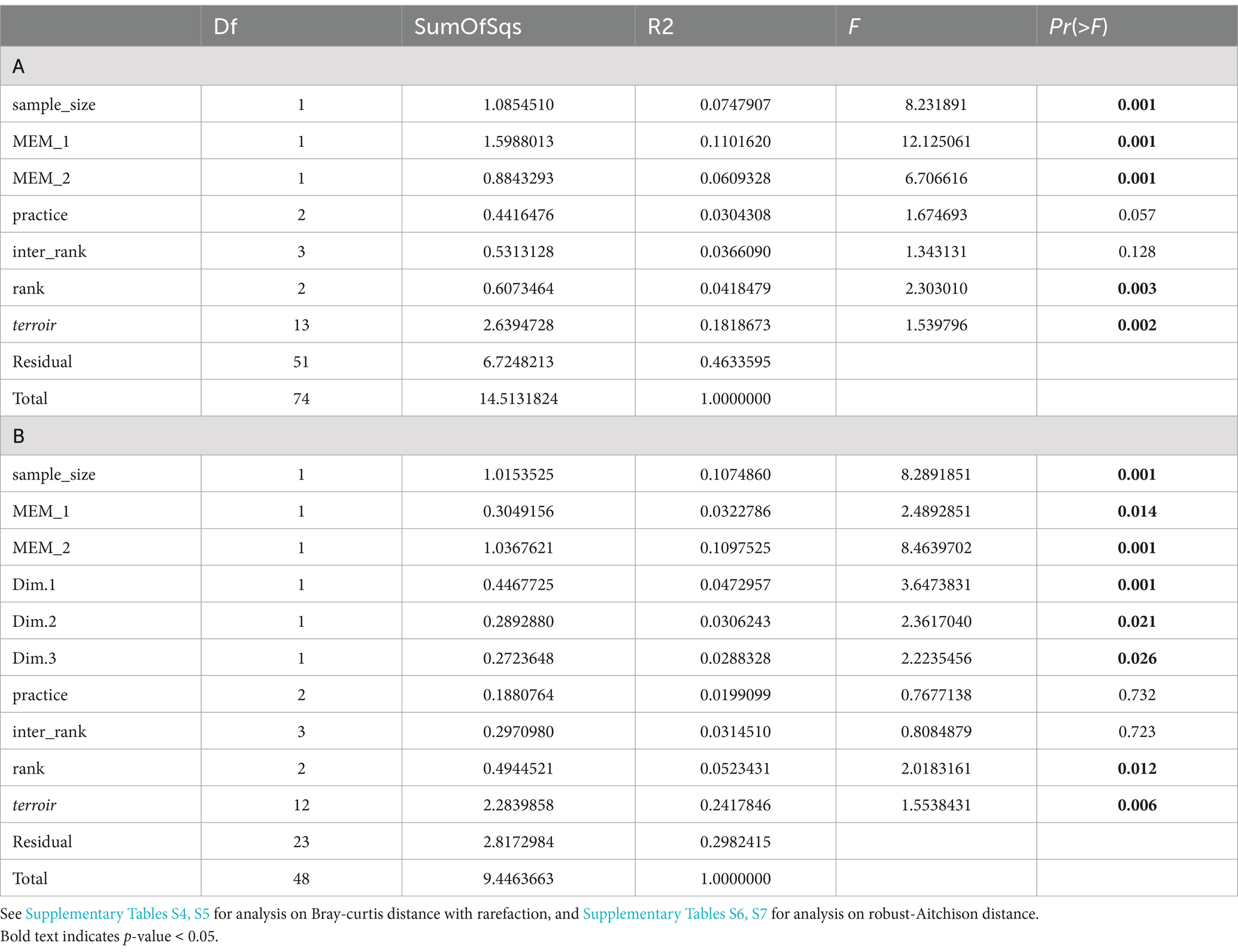
Table 3. Result of the Permanova (A with all 75 samples and B including soil physicochemical characteristics leading to only 49 samples) on Bray-curtis distance.
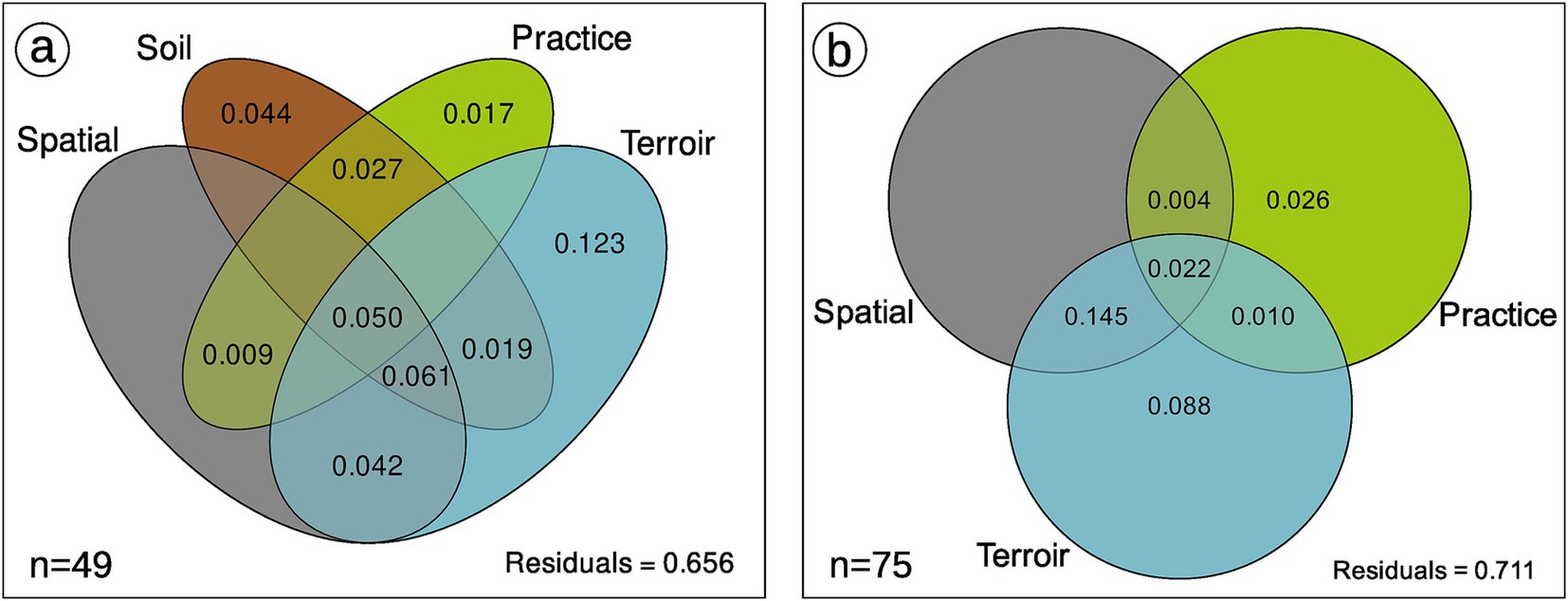
Figure 6. Variance partitioning of AMF community composition with (A) or without (B) soil component. This is the mean result of 99 rarefaction permutations (function MiscMetabar::var_par_rarperm_pq) on Bray Distance. The effect of soil, spatial and terroir components are statistically supported. When 5% of rarefaction permutations are below 0, values are not shown. See Supplementary Figure S13 for analysis with robust Aitchison distance.
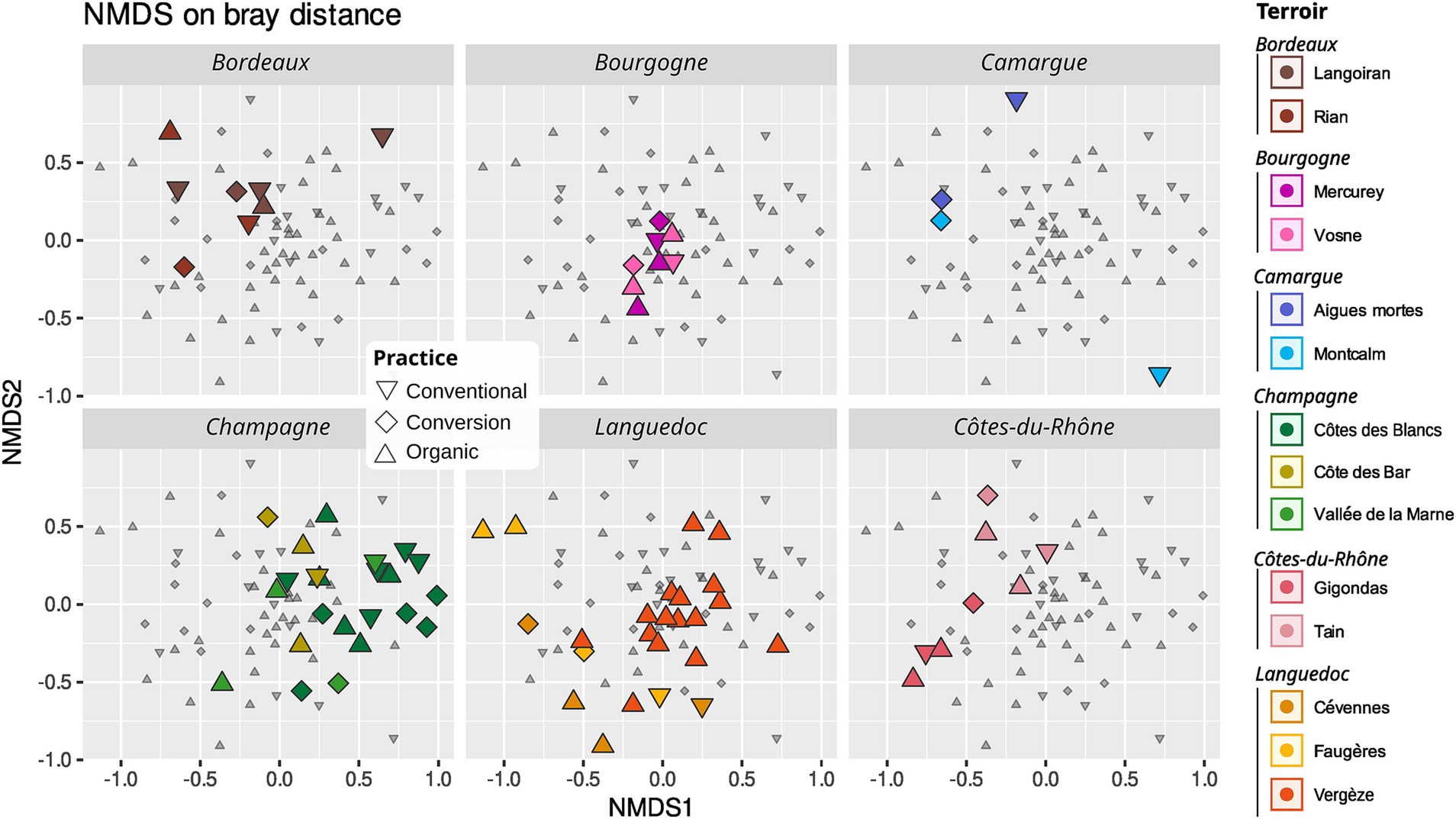
Figure 7. Non-metric dimensional scaling (NMDS) of AMF community using Bray-Curtis distance. Conventional farming: triangles pointing downwards, conversion farming: diamond, organic farming: triangles pointing upwards. Different colors represent different terroirs.

Figure 8. Indicator OTUs for terroir. Significant association between one, two or three terroirs with indicator OTUs are displayed using circles for IndVal.g metric. Terroir without indicator OTUs are not shown. None of indicator OTUs were found in association to agricultural practice.
Agricultural practices (organic, conventional, conversion) as well as rank and inter-rank work, slightly impact the AMF communities (Figure 6; Supplementary Table S4). These effects are even non-significant when we first control for soil effect and/or spatial autocorrelation in the Permanova (Table 3; Supplementary Table S5) even if we use robust Aitchison distance (Supplementary Tables S6, S7). Fifty-one OTUs (24%) are shared between all practices, less than the number of OTUs only found in organic vineyards (67 OTUs; Figures 4B,C). Fifteen OTUs (7%) are only present in vineyards under conventional practices, whereas 40 (19%) are specific to vineyards under conversion. Using indicators species analysis, we found no OTUs to be significantly linked to practice for both presence/absence and abundance-based analysis.
Whatever the practice and terroir, there is a high variability in the diversity of AMF (Table 2). Thirteen samples present a low diversity of AMF (H1 or H2 < 2; Table 4). Most low-diversity samples are conventional (8 vs. 3 conversion and 2 organic samples) and are located in Côte des Blancs terroir (7 out of 13, the 6 other samples represent 6 different terroirs). Interestingly, 5 out of 19 conventional samples were not treated with herbicide and none of these samples belong to the 8 conventional samples with low-diversity (χ2 = 4.94; p-value = 0.026).
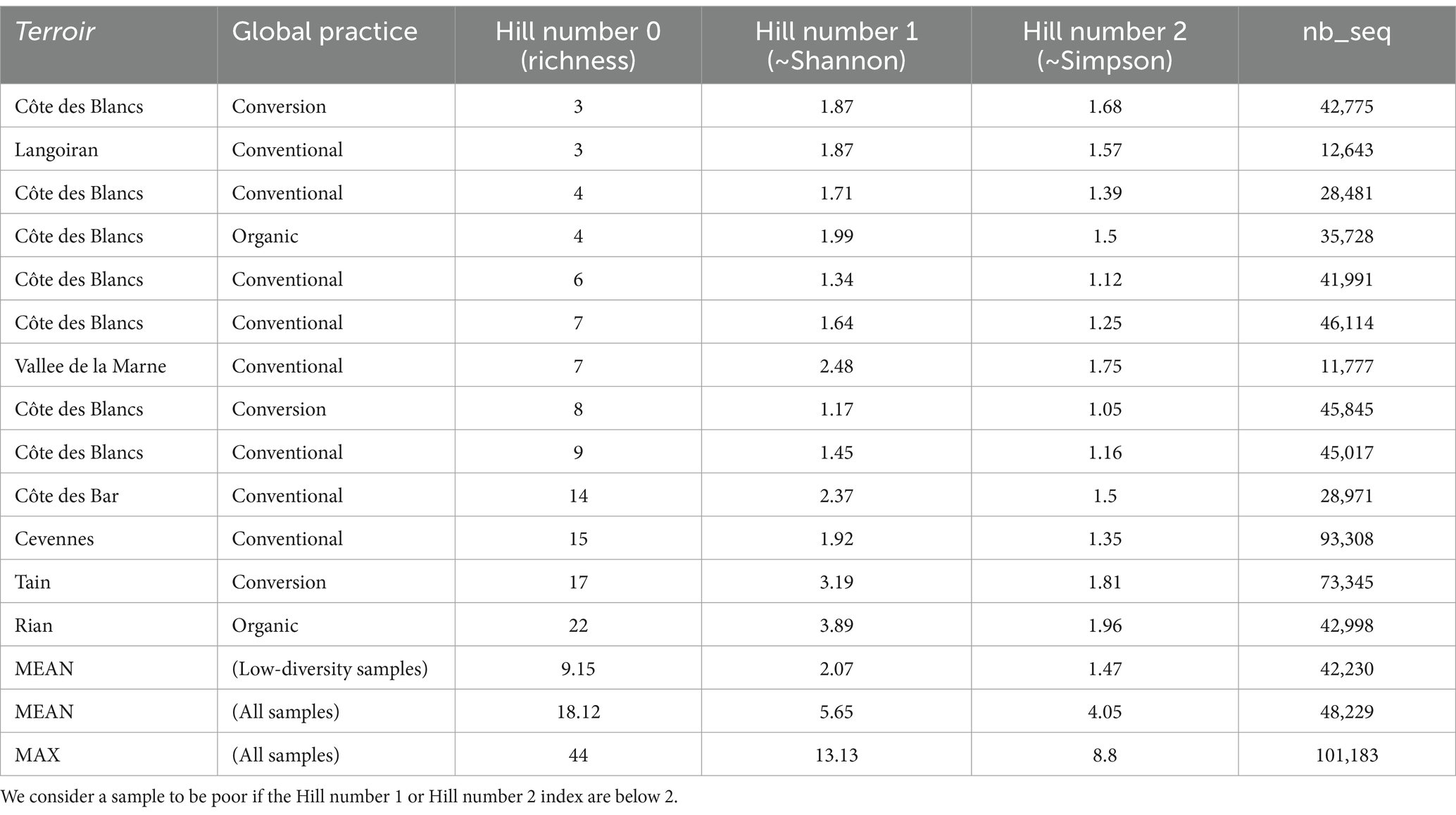
Table 4. Characteristics of low-diversity samples.
One of the most promising solutions for a sustainable viticulture is the incorporation of mutualist interactions between plants and microorganisms, such as arbuscular mycorrhizal fungi, into agricultural practice (Aguilera et al., 2022). Here, we characterize AMF communities in the wine rhizosphere across France, examining the influence of different agricultural practices in 14 terroirs (Figure 9). The taxonomy of AMF observed in French vineyards is consistent with that reported in previous studies conducted in France (Drain et al., 2019), Brazil (Bezerra et al., 2021), Canada (Holland et al., 2014), and Portugal (Fors et al., 2023). These studies have identified the genus Glomus, Rhizophagus, and Funneliformis as the dominant AMF genera in these communities.
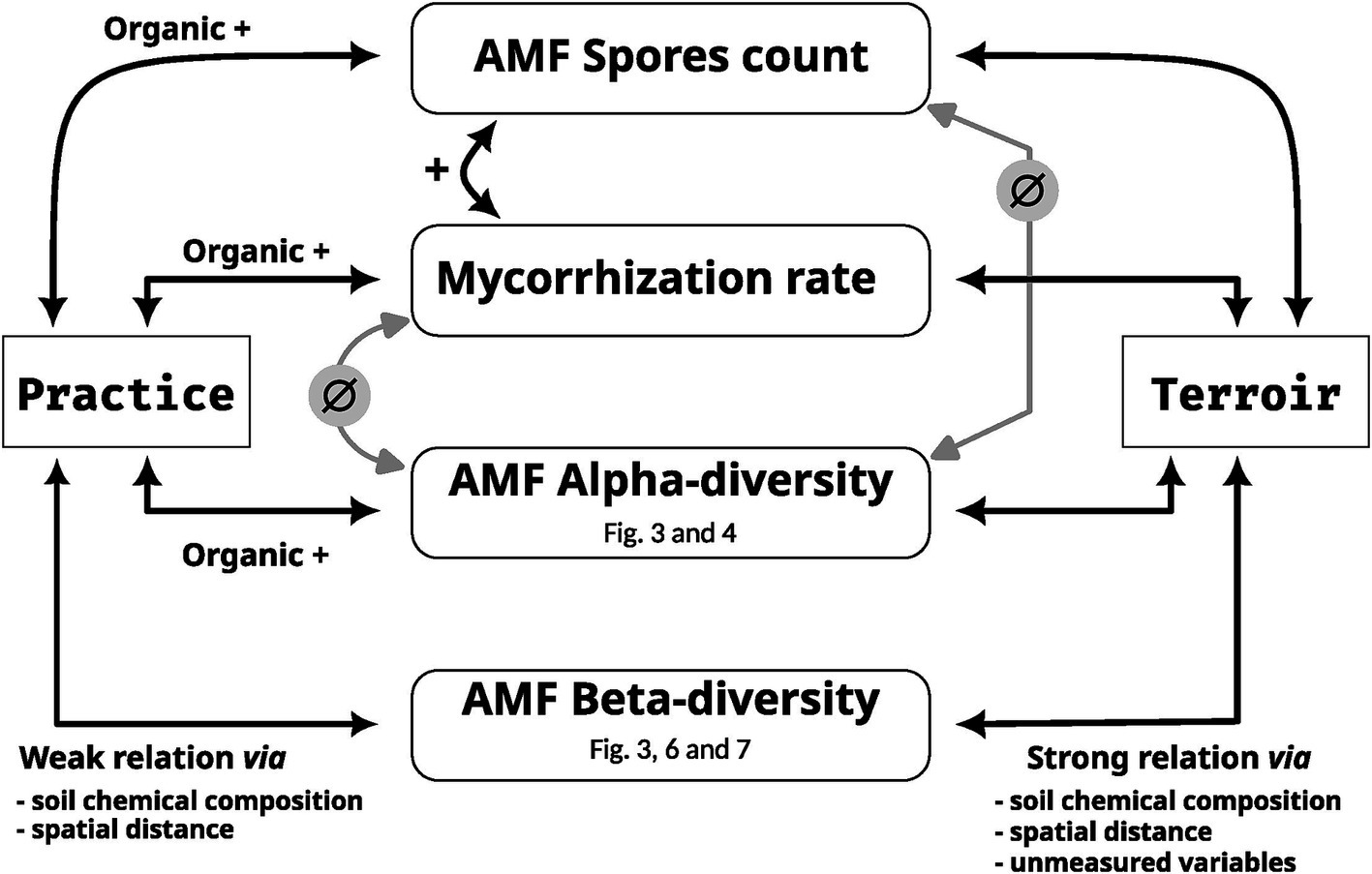
Figure 9. Schematic summary of the effect of practice and terroir on the four facets of AMF interactions. F %: mycorrhizal frequency in the root system, M %: intensity of the mycorrhizal colonization in the root system, A %: arbuscule abundance in the root system. All links are supported by correlations tests except for beta-diversity whose links are supported by Permanova and variance partitioning.
Spore-bank samples contain less AMF OTUs than root samples. This outcome may be attributed to variations in sampling, extraction, or amplification techniques between the two compartments. In particular, a notable discrepancy between spore and root samples is that we only sequenced a few thousand spores per sample, which is insufficient to encompass the total diversity present in the spore bank. Biological mechanisms can also lead to a lower diversity in spore-bank as some AMF might sporulate almost exclusively inside roots (Oehl et al., 2004). A better characterization of the AMF spores-bank requires simultaneous sampling of spores and bulk soil collected in different soil compartments, including rhizosphere without roots. As expected from previous work which separated soil and root samples (e.g., Massa et al., 2020), we found large differences in AMF communities’ composition from spore-bank and from roots. As we study the AMF from the entire rhizosphere of Vitis vinifera, the subsequent discussion is based on the samples resulting from the merging of spore and root samples in pairs. Future studies may benefit from a sampling scheme separating compartments including bulk soil samples, spore samples and root samples.
In our large-scale study, AMF spore number varied from 349 to 20,895 (mean = 4,370) spores/100 g of soil, depending on terroir and cultural practice. In the existing literature, the quantity of spores also varies considerably depending on numerous factors, including climate, soil type, grapevine variety and agricultural practices. Karagjannidis et al. (1997) found up to 280 spores/100 g of soil in healthy grapevine in Italy, while Nappi et al. (1985) counted up to 14,400 spores/100 g in the top layer of soil profile in vineyards covered with grass and bark mulch. In the same way as for spores, a large variability in arbuscules and vesicles abundance and intensity of mycorrhization in roots was found among vineyards, in the same order of values as other vineyard studies in France and Italy (Nappi et al., 1985; Bouffaud et al., 2016a).
Recent efforts have been made to consolidate the concept of terroir with data-driven analysis (e.g., Brillante et al., 2020; Bramley et al., 2023). Terroir is defined as a cultivated ecosystem in which the grapevine interacts with factors from the natural environment, principally soil and climate (Seguin, 1988). It is well established that major soil-related terroir parameters (e.g., soil temperature, soil mineral and water supply, Van Leeuwen et al., 2018) exert a profound influence on AMF establishment and development. Indeed, soil chemical attributes (Schreiner and Mihara, 2009) and climate (Shi et al., 2014) affect AMF abundance and diversity. Our focus is not to define terroir using soil or AMF communities, but to test for the possible impact of the well-defined terroir on AMF communities. The present study demonstrated a pronounced effect of wine terroir on AMF abundance and composition, indicating that certain environmental filters may be influencing the structure of AMF communities in French vineyards. Indeed, the terroir, through its soil characteristics, climate and other factors associated with geographical location and plant material, appears to be the primary determinant in shaping AMF communities. Bokulich et al. (2016), Gilbert et al. (2014) and Bouffaud et al. (2016a) also found significant differences in microbial community composition between vintages (years of production) and vineyard locations, showing that the soil microbiome is terroir specific. A significant dispersal limitations of AMF communities in vineyards were documented at the European scale (Bouffaud et al., 2016b).
In contrast with the finding of Betancur-Agudelo et al. (2023), our study did not provide evidence of mycorrhizal colonization and AMF richness decreases with increasing copper doses. Besides spatial and soil effect, terroir may influence AMF communities through plant material (Van Leeuwen et al., 2018) and unmeasured environmental or biotic variables. Thus, grapevine age, vigor, variety, and rootstocks also contribute to the community structure of arbuscular mycorrhizal fungi (Schreiner and Mihara, 2009; Noceto et al., 2023).
No clear subset of indicator species could be identified for terroir, with only 10 OTUs exhibiting relatively low indicator values. Further sampling is required to ascertain whether AMF composition can be employed as an indicator of terroir. This result is consistent with those of Gobbi et al. (2022) and Cruz-Silva et al. (2023), who identified bacterial indicator species for each terroir at the microbiome scale, but found no mycorrhizal indicator species. AMF are known to be relatively ubiquitous, with only a few hundred species forming associations with an estimated 200,000 plant species (van der Heijden et al., 2015). This may explain why the majority of AMF species do not demonstrate specificity (the presence is informative) or preference (the relative abundance is informative), yet are involved in a significant shift in AMF communities. Therefore, AMF community as a whole is clearly a part of terroir specificity and further work with more samples by terroir are necessary to describe the potential indicator communities (set of species) of terroir.
AMF abundance varies with cultural practice. We found higher root colonization and number of spores in the organic systems compared to the conversion and conventional systems. Numerous studies conducted on various crops, including grapevine, have also reported a positive effect of organic management on AMF communities (Oehl et al., 2004). Although organic farming is frequently associated with high copper use which can be detrimental to AMF populations (18% of total copper is used in organic vineyards which represent only 9% of French vineyards according to Anses, 2022), our data show that practice and soil copper concentration are not correlated. Thus, the strong decline in AMF observed in conventional systems must be linked to the use of synthetic inputs such as pesticides and chemical fertilizers (Fors et al., 2023). Synthetic fungicides (e.g., flutolanil, azoxystrobin, fenpropimorph, and fenhexamid; Zocco et al., 2008) and herbicides (e.g., glyphosate; Zaller et al., 2018) are known to inhibit, respectively, AMF spore germination, and extraradical mycelium growth and mycelial sporulation. Moreover, low-input systems such as organic farming are favorable to AMF persistence and development. Bezerra et al. (2021) observed the highest mycorrhizal incidence and index of arbuscules in vineyards with the lowest soil nutrient content. In those system, crops rely strongly on interaction with AMF to achieve a sufficient nutrient uptake (Verbruggen et al., 2010).
The data revealed no significant difference in root colonization or the number of spores between the grassed and weeded organic plots. Previous studies have yielded contradictory results. In contrast to the findings of Baumgartner et al. (2010), who observed no impact of weed flora and cover crops on AMF in vineyards, Radić et al. (2012) demonstrated that weed composition and density influence the development of fungal symbioses in the roots of grapevine. The formation of grapevine mycorrhizal networks with neighboring plants may depend on various factors including weed flora composition, plant density, and soil management practices employed to control weeds in vineyards (Baumgartner et al., 2010). Additionally, the plant host (here rootstock) may act as a natural filter selecting compatible AMF partners and thus reducing the potential influence of neighboring plants (van der Heijden et al., 2015).
The small positive effect of practice on the average number of OTUs per sample may be due to a confounding effect. On the one hand, the AMF in organic vineyards may benefit from fewer pesticides or from a higher relative investment of grapevine in the interaction. On the other hand, AMF in conventional vineyards may benefit from a possible better growth of grapevine plants and therefore a higher net investment in the interaction. In the same way, Jansa et al. (2002), Bouffaud et al. (2016a) and Fors et al. (2023) observed that different soil-tillage treatments and land use type only affected AMF community structure in vineyards, with no significant impact on α-diversity. Certain cultivation practices that can have an impact on AMF communities (e.g., soil tillage) are common to both conventional and organic plots and should therefore offset the effect of practices on the number of AMF species at terroir scale.
Even though cultural practices weakly affect α-diversity at sample scale, the number of AMF species is markedly higher in organic viticulture at global scale (142 OTUs in organic vs. 80 in conventional). This result corroborates the observation that organic farming exhibits a higher diversity of OTUs and a more complex structure of AMF communities compared to conventional farming (Chen et al., 2022). Moreover, conventional vineyards harbor 5 times less specific OTUs than organic vineyards and 3 times less than conversion vineyards. This suggests a higher diversity at the global scale in organic vineyards and, to a lesser but not significant extent, in conversion vineyards. These findings suggest that there are potential diverse OTUs resources currently present on conversion parcels.
AMF community structure is altered by conventional farming in French vineyards. However, most effects of practices are mediated by spatial proximity and soil chemical composition. The absence of indicator species for agricultural practice corroborates the relatively limited impact of practice on AMF communities in comparison to terroir. Although AMF are commonly found in a wide diversity of cultivated soils, conventional management practices such as soil tillage, fertilization, and the use of pesticides combined with low plant diversity negatively affect AMF abundance and diversity (Brito et al., 2012). These practices may favor a few fast-sporulating and generalist taxa over more specialist ones with higher symbiotic efficiency (Bouffaud et al., 2016b). The dominance of ubiquitous genera in disturbed sites such as Glomus, Claroideoglomus and Funneliformis can be related to their life strategy characterized by fast spore production, germination, and mycelium extension (Oehl et al., 2010). Consistently, 71% of the OTUs identified in conventional vineyards belong to the Glomeraceae family, against 68% in conversion and 62% in organic ones.
A higher biodiversity is often correlated with better ecosystem functioning and resilience (Tilman et al., 2014) and this relation is even stronger in stressful environments induced by global change drivers (Hong et al., 2022). Diversity-ecosystem function relationship in AMF was also found to be positive (Powell and Rillig, 2018). For a plant involved in mycorrhizal interactions, more fungal species may enhance the probability of finding a good partner (selection effect sensu Tilman et al., 2014) and interacting with functionally complementary fungi (complementarity effect). Consequently, by enhancing the abundance and the diversity of AMF, organic practices may improve productivity and resilience of vineyards to perturbations such as severe drought and fungal root pathogens. Additional research is necessary to quantify the effect of organic farming on vineyard ecosystem functioning through AMF.
The present study focuses on an essential functional part of the vineyard microbiome by describing the local AMF communities in the rhizosphere at a given date in French terroirs. To broaden our conclusions, future research should sequence more barcode markers (e.g., ITS, COI, 16S) to improve taxonomic resolution for AMF (Delavaux et al., 2020) and to expand the taxonomic scope to bacteria, non-mycorrhizal fungi and other Eukaryota. Further studies may also encompass other countries and incorporate the temporal dynamic of the AMF communities, particularly during the vineyard life cycle. Describing the seasonal and long-term variation in AMF diversity and composition will provide new insights crucial for future viticulture. Finally, multi-omics approaches could also elucidate new aspects of vineyard-AMF interactions.
Among the agroecological solutions that can assist in addressing the new constraints facing viticulture, the inoculation of vineyards with AMF is underexplored (Jindo et al., 2022). Our findings demonstrate that AMF communities in grapevines are highly dependent on terroir, underscoring the necessity for future studies on AMF inoculation in vineyards to consider the specific characteristics of each terroir.
The original contributions presented in the study are publicly available. This data can be found here: https://www.ebi.ac.uk/ena/browser/view/PRJEB83803.
PB-L: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AT: Data curation, Formal analysis, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. MB: Data curation, Formal analysis, Methodology, Writing – review & editing. LB: Conceptualization, Data curation, Formal analysis, Investigation, Writing – review & editing. MD: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing, Visualization. YR: Writing – review & editing, Visualization, Data curation, Formal analysis, Investigation, Methodology. AY: Formal analysis, Methodology, Software, Writing – review & editing, Visualization. DB-E: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was partially supported by LA REGION OCCITANIE under the Contrat Innovation number [19025370].
The authors are grateful to all the winegrowers for making their plots available for this study and for their support. We would like to thank Patrick Carroll for proofreading the article.
PB-L, AT, MB, LB, MD, YR, AY, and DB-E were employed by company Mycea.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1463326/full#supplementary-material
Aguilera, P., Becerra, N., Alvear, M., Ortiz, N., Turrini, A., Azcón-Aguilar, C., et al. (2022). Arbuscular mycorrhizal fungi from acidic soils favors production of tomatoes and lycopene concentration. J. Sci. Food Agric. 102, 2352–2358. doi: 10.1002/jsfa.11573
Alberdi, A., and Gilbert, M. T. P. (2019). A guide to the application of Hill numbers to DNA-based diversity analyses. Mol. Ecol. Resour. 19, 804–817. doi: 10.1111/1755-0998.13014
Anses, (2022). Cartographie des utilisations des produits phytopharmaceutiques à base de cuivre en France en considérant leur application en agriculture biologique et conventionnelle. (saisine 2021-AUTO-0060). Maisons-Alfort: Anses, 133.
Banerjee, S., and van der Heijden, M. G. A. (2023). Soil microbiomes and one health. Nat. Rev. Microbiol. 21, 6–20. doi: 10.1038/s41579-022-00779-w
Baumgartner, K., Fujiyoshi, P., Smith, R., and Bettiga, L. (2010). Weed for a and dormant-season cover crops have no effects on arbuscular mycorrhizae of grapevine. Weed Res. 50, 456–466. doi: 10.1111/j.1365-3180.2010.00793.x
Baumgartner, K., Smith, R. F., and Bettiga, L. (2005). Weed control and cover crop management affect mycorrhizal colonization of grapevine roots and arbuscular mycorrhizal fungal spore populations in a California vineyard. Mycorrhiza 15, 111–119. doi: 10.1007/s00572-004-0309-2
Betancur-Agudelo, M., Meyer, E., and Lovato, P. E. (2021). Arbuscular mycorrhizal fungus richness in the soil and root colonization in vineyards of different ages. Rhizosphere 17:100307. doi: 10.1016/j.rhisph.2021.100307
Betancur-Agudelo, M., Meyer, E., and Lovato, P. E. (2023). Increased copper concentrations in soil affect indigenous arbuscular mycorrhizal fungi and physiology of grapevine plantlets. Rhizosphere 27:100711. doi: 10.1016/j.rhisph.2023.100711
Bezerra, A. D. S., Betancur-Agudelo, M., Meyer, E., Kemmelmeier, K., Stürmer, S. L., Soares, C. R. F. S., et al. (2021). Occurrence and richness of arbuscular mycorrizal fungi in vineyards with grapevine decline and dieback symptoms. Ciência Rural 52:e20210011. doi: 10.1590/0103-8478cr20210011
Biasi, R., Brunori, E., Vanino, S., Bernardini, A., Catalani, A., Farina, R., et al. (2023). Soil–plant interaction mediated by indigenous AMF in grafted and own-rooted grapevines under field conditions. Agriculture 13:1051. doi: 10.3390/agriculture13051051
Bokulich, N. A., Collins, T. S., Masarweh, C., Allen, G., Heymann, H., Ebeler, S. E., et al. (2016). Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 7:e00631-16. doi: 10.1128/mBio.00631-16
Bouffaud, M.-L., Bernaud, E., Colombet, A., Van Tuinen, D., Wipf, D., and Redecker, D. (2016a). Regional-scale analysis of arbuscular mycorrhizal fungi: the case of Burgundy vineyards. OENO One 50:1. doi: 10.20870/oeno-one.2016.50.1.49
Bouffaud, M.-L., Creamer, R. E., Stone, D., Plassart, P., Van Tuinen, D., Lemanceau, P., et al. (2016b). Indicator species and co-occurrence in communities of arbuscular mycorrhizal fungi at the European scale. Soil Biol. Biochem. 103, 464–470. doi: 10.1016/j.soilbio.2016.09.022
Bouyoucos, G. J. (1962). Hydrometer method improved for making particle size analyses of soils 1. Agron. J. 54, 464–465. doi: 10.2134/agronj1962.00021962005400050028x
Bramley, R. G. V., Ouzman, J., Sturman, A. P., Grealish, G. J., Ratcliff, C. E. M., and Trought, M. C. T. (2023). Underpinning terroir with data: integrating vineyard performance metrics with soil and climate data to better understand within-region variation in Marlborough, New Zealand. Aust. J. Grape Wine Res. 2023, 1–23. doi: 10.1155/2023/8811402
Bremner, J. M., and Mulvaney, C. S. (1982) in “Total nitrogen” in methods of soil analysis part 2. eds. A. L. Page, R. H. Miller, and D. R. Keeney. 2nd ed (Wisconsin: ASA, SSSA Madison).
Brillante, L., Bonfante, A., Bramley, R. G. V., Tardaguila, J., and Priori, S. (2020). Unbiased scientific approaches to the study of terroir are needed! Front. Earth Sci. 8:539377. doi: 10.3389/feart.2020.539377
Brito, I., Goss, M. J., De Carvalho, M., Chatagnier, O., and Van Tuinen, D. (2012). Impact of tillage system on arbuscular mycorrhiza fungal communities in the soil under Mediterranean conditions. Soil Tillage Res. 121, 63–67. doi: 10.1016/j.still.2012.01.012
Cáceres, M. D., and Legendre, P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016a). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Callahan, B. J., Sankaran, K., Fukuyama, J. A., McMurdie, P. J., and Holmes, S. P. (2016b). Bioconductor workflow for microbiome data analysis: from raw reads to community analyses. F1000Res 5:1492. doi: 10.12688/f1000research.8986.2
Chen, J., Li, J., Yang, Y., Wang, Y., Zhang, Y., and Wang, P. (2022). Effects of conventional and organic agriculture on soil arbuscular mycorrhizal fungal Community in low-Quality Farmland. Front. Microbiol. 13:914627. doi: 10.3389/fmicb.2022.914627
Cruz-Silva, A., Laureano, G., Pereira, M., Dias, R., Silva, J. M. D., Oliveira, N., et al. (2023). A new perspective for vineyard terroir identity: looking for microbial Indicator species by long read Nanopore sequencing. Microorganisms 11:672. doi: 10.3390/microorganisms11030672
Daniels, B. A., and Skipper, H. D. (1982). “Methods for the recovery and quantitative estimation of propagules from soil” in Methods and principles of mycorrhizal research. ed. N. C. Schneck (St Paul, MN: American Phytopathological Society), 244.
Delavaux, C. S., Sturmer, S. L., Wagner, M. R., Schütte, U. M. E., Morton, J., and Bever, J. (2020). Utility of LSU for environmental sequencing of arbuscular mycorrhizal fungi: a new reference database and pipeline. New Phytol. 229, 3048–3052. doi: 10.1111/nph.17080
Drain, A., Bonneau, L., Recorbet, G., Van Tuinen, D., Wipf, D., and Courty, P.-E. (2019). “Characterization of arbuscular mycorrhizal communities in roots of vineyard plants” in Methods in rhizosphere biology research. eds. D. Reinhardt and A. K. Sharma (Singapore: Springer Singapore), 27–34.
Dray, S., Bauman, D., Blanchet, G., Borcard, D., Clappe, S., Guénard, G., et al. (2023). adespatial: Multivariate Multiscale Spatial Analysis. R package version 0.3-23. Available at: https://CRAN.R-project.org/package=adespatial.
Dray, S., and Dufour, A. (2007). The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22, 1–20. doi: 10.18637/jss.v022.i04
Foley, J. A., Ramankutty, N., Brauman, K. A., Cassidy, E. S., Gerber, J. S., Johnston, M., et al. (2011). Solutions for a cultivated planet. Nature 478, 337–342. doi: 10.1038/nature10452
Fors, R. O., Sorci-Uhmann, E., Santos, E. S., Silva-Flores, P., Abreu, M. M., Viegas, W., et al. (2023). Influence of soil type, land use, and rootstock genotype on root-associated arbuscular mycorrhizal Fungi communities and their impact on grapevine growth and nutrition. Agriculture 13:2163. doi: 10.3390/agriculture13112163
Frew, A., and Aguilar-Trigueros, C. A. (2023). Australia offers unique insight into the ecology of arbuscular mycorrhizal fungi: An opportunity not to be lost. Austral Ecol. 48, 1713–1720. doi: 10.1111/aec.13451
Gerdemann, J. W., and Nicolson, T. H. (1963). Spores of mycorrhizal Endogone species extracted by wet sieving and decanting. Trans. Br. Mycol. Soc. 46, 235–244. doi: 10.1016/S0007-1536(63)80079-0
Gilbert, J. A., van der Lelie, D., and Zarraonaindia, I. (2014). Microbial terroir for wine grapes. Proc. Natl. Acad. Sci. USA 111, 5–6. doi: 10.1073/pnas.1320471110
Gobbi, A., Acedo, A., Imam, N., Santini, R. G., Ortiz-Álvarez, R., Ellegaard-Jensen, L., et al. (2022). A global microbiome survey of vineyard soils highlights the microbial dimension of viticultural terroirs. Commun. Biol. 5:241. doi: 10.1038/s42003-022-03202-5
Guillou, L., Bachar, D., Audic, S., Bass, D., Berney, C., Bittner, L., et al. (2012). The Protist ribosomal reference database (PR2): a catalog of unicellular eukaryote small sub-unit rRNA sequences with curated taxonomy. Nucleic Acids Res. 41, D597–D604. doi: 10.1093/nar/gks1160
Guzman, A., Montes, M., Hutchins, L., DeLaCerda, G., Yang, P., Kakouridis, A., et al. (2021). Crop diversity enriches arbuscular mycorrhizal fungal communities in an intensive agricultural landscape. New Phytol. 231, 447–459. doi: 10.1111/nph.17306
Hart, M. M., and Reader, R. J. (2002). Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytol. 153, 335–344. doi: 10.1046/j.0028-646X.2001.00312.x
Hill, M. O. (1973). Diversity and evenness: A unifying notation and its consequences. Ecology 54, 427–432. doi: 10.2307/1934352
Holland, T., Bowen, P., Bogdanoff, C., and Hart, M. (2014). Arbuscular mycorrhizal fungal communities associated with Vitis vinifera vines under different frequencies of irrigation. Am. J. Enol. Vitic. 65, 222–229. doi: 10.5344/ajev.2014.13101
Hong, P., Schmid, B., De Laender, F., Eisenhauer, N., Zhang, X., Chen, H., et al. (2022). Biodiversity promotes ecosystem functioning despite environmental change. Ecol. Lett. 25, 555–569. doi: 10.1111/ele.13936
Inkscape Team. (2023). Inkscape. Available at: https://inkscape.org/.
IOV (2023). 2023 statistical report on world Vitiviniculture. Paris: International Organization of Vine and Wine.
Jafarian, N., Mirzaei, J., Omidipour, R., and Kooch, Y. (2024). Changes in climatic conditions drive variations in arbuscular mycorrhizal fungi diversity and composition in semi-arid oak forests. J. For. Res. 35:94. doi: 10.1007/s11676-024-01744-4
Jansa, J., Mozafar, A., Anken, T., Ruh, R., Sanders, I., and Frossard, E. (2002). Diversity and structure of AMF communities as affected by tillage in a temperate soil. Mycorrhiza 12, 225–234. doi: 10.1007/s00572-002-0163-z
Jiang, S., Hu, X., Kang, Y., Xie, C., An, X., Dong, C., et al. (2020). Arbuscular mycorrhizal fungal communities in the rhizospheric soil of litchi and mango orchards as affected by geographic distance, soil properties and manure input. Appl. Soil Ecol. 152:103593. doi: 10.1016/j.apsoil.2020.103593
Jindo, K., Goron, T. L., Pizarro-Tobías, P., Sánchez-Monedero, M. Á., Audette, Y., Deolu-Ajayi, A. O., et al. (2022). Application of biostimulant products and biological control agents in sustainable viticulture: A review. Front. Plant Sci. 13:932311. doi: 10.3389/fpls.2022.932311
Joos, L., Beirinckx, S., Haegeman, A., Debode, J., Vandecasteele, B., Baeyen, S., et al. (2020). Daring to be differential: metabarcoding analysis of soil and plant-related microbial communities using amplicon sequence variants and operational taxonomical units. BMC Genomics 21:733. doi: 10.1186/s12864-020-07126-4
Karagjannidis, N., Velemis, D., and Stavropoulos, N. (1997). Root colonization and spore population by VA-mycorrhizal fungi in four grapevine rootstocks. Vitis 36, 57–60.
Kassambara, A., and Mundt, F. (2020). Factoextra: extract and visualize the results of multivariate data analyses. R package version 1.0.7. Available at: https://CRAN.R-project.org/package=factoextra.
Landau, W. (2021). The targets R package: a dynamic make-like function-oriented pipeline toolkit for reproducibility and high-performance computing. JOSS 6:2959. doi: 10.21105/joss.02959
Lannelongue, L., Grealey, J., and Inouye, M. (2021). Green algorithms: quantifying the carbon footprint of computation. Adv. Sci. 8:2100707. doi: 10.1002/advs.202100707
Le, S., Josse, J., and Husson, F. (2008). Facto mine R: An R package for multivariate analysis. J. Stat. Softw. 25, 1–18. doi: 10.18637/jss.v025.i01
Lee, J., Lee, S., and Young, J. P. W. (2008). Improved PCR primers for the detection and identification of arbuscular mycorrhizal fungi: PCR primers for arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 65, 339–349. doi: 10.1111/j.1574-6941.2008.00531.x
Lex, A., Gehlenborg, N., Strobelt, H., Vuillemot, R., and Pfister, H. (2014). UpSet: visualization of intersecting sets. IEEE Trans. Vis. Comput. Graph. 20, 1983–1992. doi: 10.1109/TVCG.2014.2346248
Lin, X. G., Feng, Y. Z., Zhang, H. Y., Chen, R. R., Wang, J. H., Zhang, J. B., et al. (2012). Long-term balanced fertilization decreases arbuscular mycorrhizal fungal diversity in an arable soil in North China revealed by 454 pyrosequencing. Environ. Sci. Technol. 46, 5764–5771. doi: 10.1021/es3001695
Lin, C., Wang, Y., Liu, M., Li, Q., Xiao, W., and Song, X. (2020). Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 10:12260. doi: 10.1038/s41598-020-69213-6
Lindsay, W. L., and Norvell, W. A. (1978). Development of DTPA soil test for Zn, Fe, Mn and cu. Soil Sci. Soc. Am. J. 42, 421–428. doi: 10.2136/sssaj1978.03615995004200030009x
Maherali, H., and Klironomos, J. N. (2007). Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 316, 1746–1748. doi: 10.1126/science.1143082
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.j 17:10. doi: 10.14806/ej.17.1.200
Martin, F. M., and van der Heijden, M. G. A. (2024). The mycorrhizal symbiosis: research frontiers in genomics, ecology, and agricultural application. New Phytol. 242, 1486–1506. doi: 10.1111/nph.19541
Massa, N., Cesaro, P., Todeschini, V., Capraro, J., Scarafoni, A., Cantamessa, S., et al. (2020). Selected autochthonous rhizobia, applied in combination with AM fungi, improve seed quality of common bean cultivated in reduced fertilization condition. Appl. Soil Ecol. 148:103507. doi: 10.1016/j.apsoil.2020.103507
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Mocali, S., Kuramae, E. E., Kowalchuk, G. A., Fornasier, F., and Priori, S. (2020). Microbial functional diversity in vineyard soils: sulfur metabolism and links with grapevine plants and wine quality. Front. Environ. Sci. 8:75. doi: 10.3389/fenvs.2020.00075
Nappi, P., Jodice, R., Luzzati, A., and Corino, L. (1985). Grapevine root system and VA mycorrhizae in some soils of Piedmont (Italy). Plant Soil 85, 205–210. doi: 10.1007/BF02139624
Noceto, P.-A., Durney, C., Van Tuinen, D., De Sousa, J., Wipf, D., and Courty, P.-E. (2023). Arbuscular mycorrhizal fungal communities differ in neighboring vineyards of different ages. Mycorrhiza 33, 241–248. doi: 10.1007/s00572-023-01117-5
Ochoa-Hueso, R., Cantos-Villar, E., Puertas, B., Aguiar Del Rio, J. F., Belda, I., Delgado-Baquerizo, M., et al. (2024). Nature-based strategies to regenerate the functioning and biodiversity of vineyards. J. Sust. Agricult. Environ. 3:e12088. doi: 10.1002/sae2.12088
Oehl, F., Laczko, E., Bogenrieder, A., Stahr, K., Bösch, R., van der Heijden, M., et al. (2010). Soil type and land use intensity determine the composition of arbuscular mycorrhizal fungal communities. Soil Biol. Biochem. 42, 724–738. doi: 10.1016/j.soilbio.2010.01.006
Oehl, F., Sieverding, E., Mäder, P., Dubois, D., Ineichen, K., Boller, T., et al. (2004). Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 138, 574–583. doi: 10.1007/s00442-003-1458-2
Okalebo, J. R., Gathua, K. W., and Woomer, P. L. (2002). Laboratory methods of soil and plant analysis: A working manual 2nd. Nairobi: Sacred Africa, 21.
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., et al. (2022). Vegan: community ecology package. Available at: https://CRAN.R-project.org/package=vegan.
Öpik, M., Vanatoa, A., Vanatoa, E., Moora, M., Davison, J., Kalwij, J. M., et al. (2010). The online database Maarj AM reveals global and ecosystemic distribution patterns in arbuscular mycorrhizal fungi (Glomeromycota). New Phytol. 188, 223–241. doi: 10.1111/j.1469-8137.2010.03334.x
Paiola, A., Assandri, G., Brambilla, M., Zottini, M., Pedrini, P., and Nascimbene, J. (2020). Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 706:135839. doi: 10.1016/j.scitotenv.2019.135839
Patil, I. (2021). Visualizations with statistical details: the “ggstatsplot” approach. JOSS 6:3167. doi: 10.21105/joss.03167
Posit Team (2024). RStudio: Integrated development environment for R. Boston, MA: Posit Software, PBC Available at: http://www.posit.co/.
Powell, J. R., and Rillig, M. C. (2018). Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 220, 1059–1075. doi: 10.1111/nph.15119
R Core Team (2023). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing Available at: https://www.R-project.org/.
Radić, T., Hančević, K., Likar, M., Protega, I., Jug-Dujaković, M., and Bogdanović, I. (2012). Neighbouring weeds influence the formation of arbuscular mycorrhiza in grapevine. Symbiosis 56, 111–120. doi: 10.1007/s13199-012-0165-3
Radić, T., Likar, M., Hančević, K., Bogdanović, I., and Pasković, I. (2014). Occurrence of root endophytic fungi in organic versus conventional vineyards on the Croatian coast. Agric. Ecosyst. Environ. 192, 115–121. doi: 10.1016/j.agee.2014.04.008
Rivera-Becerril, F., Van Tuinen, D., Chatagnier, O., Rouard, N., Béguet, J., Kuszala, C., et al. (2017). Impact of a pesticide cocktail (fenhexamid, folpel, deltamethrin) on the abundance of Glomeromycota in two agricultural soils. Sci. Total Environ. 577, 84–93. doi: 10.1016/j.scitotenv.2016.10.098
Rognes, T., Flouri, T., Nichols, B., Quince, C., and Mahé, F. (2016). VSEARCH: a versatile open source tool for metagenomics. Peer J 4:e2584. doi: 10.7717/peerj.2584
Säle, V., Aguilera, P., Laczko, E., Mäder, P., Berner, A., Zihlmann, U., et al. (2015). Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 84, 38–52. doi: 10.1016/j.soilbio.2015.02.005
Schreiner, R. P. (2005). Spatial and temporal variation of roots, arbuscular mycorrhizal Fungi, and plant and soil nutrients in a mature pinot noir (Vitis vinifera L.) vineyard in Oregon, USA. Plant Soil 276, 219–234. doi: 10.1007/s11104-005-4895-0
Schreiner, R. P., and Mihara, K. L. (2009). The diversity of arbuscular mycorrhizal fungi amplified from grapevine roots (Vitis vinifera L.) in Oregon vineyards is seasonally stable and influenced by soil and vine age. Mycologia 101, 599–611. doi: 10.3852/08-169
Seguin, G. (1988). “Ecosystems of the great red wines produced in the maritime climate of Bordeaux” in Department of Horticultural Sciences. ed. Proceedings of the Symposium on Maritime Climate Winegrowing. Fuller-Perrine L. (Geneva, NY: Cornell University), 36–53.
Sheng, M., Lalande, R., Hamel, C., and Ziadi, N. (2013). Effect of long-term tillage and mineral phosphorus fertilization on arbuscular mycorrhizal fungi in a humid continental zone of eastern Canada. Plant Soil 369, 599–613. doi: 10.1007/s11104-013-1585-4
Shi, Z. Y., Wang, F. Y., Zhang, K., and Chen, Y. L. (2014). Diversity and distribution of arbuscular mycorrhizal fungi along altitudinal gradients in mount Taibai of the Qinling Mountains. Can. J. Microbiol. 60, 811–818. doi: 10.1139/cjm-2014-0416
Simon, L., Lalonde, M., and Bruns, T. D. (1992). Specific amplification of 18S fungal ribosomal genes from vesicular-arbuscular endomycorrhizal fungi colonizing roots. Appl. Environ. Microbiol. 58, 291–295. doi: 10.1128/aem.58.1.291-295.1992
Sumner, M. E., and Miller, W. P. (1996). Cation exchange capacity. In: methods of soil analysis part 3: chemical methods. SSSA Book Series 5, 1201–1229.
Taudière, A. (2023). Misc Metabar: an R package to facilitate visualization and reproducibility in metabarcoding analysis. J. Open Source Softw. 8:6038. doi: 10.21105/joss.06038
Tedersoo, L., Bahram, M., Zinger, L., Nilsson, R. H., Kennedy, P. G., Yang, T., et al. (2022). Best practices in metabarcoding of fungi: from experimental design to results. Mol. Ecol. 31, 2769–2795. doi: 10.1111/mec.16460
Thomopoulos, S., Elsgaard, L., Munkholm, L. J., and Ravnskov, S. (2023). Evaluation of the relation between soil biomass of arbuscular mycorrhizal fungi and glomalin-related soil protein in conservation agriculture. Soil Biol. Biochem. 187:109222. doi: 10.1016/j.soilbio.2023.109222
Tilman, D., Isbell, F., and Cowles, J. M. (2014). Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 45, 471–493. doi: 10.1146/annurev-ecolsys-120213-091917
Torres, N., Antolín, M. C., and Goicoechea, N. (2018). Arbuscular mycorrhizal Symbiosis as a promising resource for improving berry quality in grapevines under changing environments. Front. Plant Sci. 9:897. doi: 10.3389/fpls.2018.00897
Torres, N., Hilbert, G., Antolín, M. C., and Goicoechea, N. (2019). Aminoacids and flavonoids profiling in Tempranillo berries can be modulated by the arbuscular mycorrhizal Fungi. Plan. Theory 8:400. doi: 10.3390/plants8100400
Trouvelot, S., Bonneau, L., Redecker, D., Van Tuinen, D., Adrian, M., and Wipf, D. (2015). Arbuscular mycorrhiza symbiosis in viticulture: a review. Agron. Sustain. Dev. 35, 1449–1467. doi: 10.1007/s13593-015-0329-7
Trouvelot, A., Kough, J. L., and Gianinazzi-Pearson, V. (1986). “Mesure du taux de mycorrhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification functionelle” in Mycorrhizae: Physiology and genetics. eds. V. Gianinazzi-Pearson and S. Gianinazzi (Paris: INRA), 217–221.
van der Heijden, M. G. (2002). “Arbuscular mycorrhizal Fungi as a determinant of plant diversity: in search of underlying mechanisms and general principles” in Mycorrhizal Ecology, eds. M. G. A. HeijdenVan Der and I. R. Sanders (Springer Berlin Heidelberg: Berlin, Heidelberg), 243–265, 157.
van der Heijden, M. G., Martin, F. M., Selosse, M. A., and Sanders, I. R. (2015). Mycorrhizal ecology and evolution: the past, the present, and the future. New Phytol. 205, 1406–1423. doi: 10.1111/nph.13288
Van Geel, M., Verbruggen, E., De Beenhouwer, M., van Rennes, G., Lievens, B., and Honnay, O. (2017). High soil phosphorus levels overrule the potential benefits of organic farming on arbuscular mycorrhizal diversity in northern vineyards. Agric. Ecosyst. Environ. 248, 144–152. doi: 10.1016/j.agee.2017.07.017
Van Leeuwen, C., Roby, J.-P., and de Rességuier, L. (2018). Soil-related terroir factors: a review. OENO One 52, 173–188. doi: 10.20870/oeno-one.2018.52.2.2208
Verbruggen, E., Röling, W. F. M., Gamper, H. A., Kowalchuk, G. A., Verhoef, H. A., and van der Heijden, M. G. A. (2010). Positive effects of organic farming on below-ground mutualists: large-scale comparison of mycorrhizal fungal communities in agricultural soils. New Phytol. 186, 968–979. doi: 10.1111/j.1469-8137.2010.03230.x
Vierheilig, H., Coughlan, A. P., Wyss, U., and Piché, Y. (1998). Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal Fungi. Appl. Environ. Microbiol. 64, 5004–5007. doi: 10.1128/AEM.64.12.5004-5007.1998
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07
Wang, J., Zhang, J., Wang, C., Ren, G., Yang, Y., and Wang, D. (2021). Precipitation exerts a strong influence on arbuscular mycorrhizal fungi community and network complexity in a semiarid steppe ecosystem. Eur. J. Soil Biol. 102:103268. doi: 10.1016/j.ejsobi.2020.103268
Wezel, A., Herren, B. G., Kerr, R. B., Barrios, E., Gonçalves, A. L. R., and Sinclair, F. (2020). Agroecological principles and elements and their implications for transitioning to sustainable food systems. A review. Agron. Sustain. Dev. 40:40. doi: 10.1007/s13593-020-00646-z
Winkler, K. J., Viers, J. H., and Nicholas, K. A. (2017). Assessing ecosystem services and multifunctionality for vineyard systems. Front. Environ. Sci. 5:15. doi: 10.3389/fenvs.2017.00015
Yu, Z., Lu, T., and Qian, H. (2023). Pesticide interference and additional effects on plant microbiomes. Sci. Total Environ. 888:164149. doi: 10.1016/j.scitotenv.2023.164149
Zaller, J. G., Cantelmo, C., Santos, G. D., Muther, S., Gruber, E., Pallua, P., et al. (2018). Herbicides in vineyards reduce grapevine root mycorrhization and alter soil microorganisms and the nutrient composition in grapevine roots, leaves, xylem sap and grape juice. Environ. Sci. Pollut. Res. 25, 23215–23226. doi: 10.1007/s11356-018-2422-3
Keywords: symbiosis, microbiome, arbuscular mycorrhizal fungi, community ecology, wine, organic farming, terroir
Citation: Battie-Laclau P, Taudière A, Bernard M, Bodénan L, Duchemin M, de Roman Y, Yol A and Barry-Etienne D (2025) Terroir and farming practices drive arbuscular mycorrhizal fungal communities in French vineyards. Front. Microbiol. 15:1463326. doi: 10.3389/fmicb.2024.1463326
Edited by:
Monica Calvo-Polanco, University of Salamanca, SpainReviewed by:
Eva Nouri, Université de Fribourg, SwitzerlandCopyright © 2025 Battie-Laclau, Taudière, Bernard, Bodénan, Duchemin, de Roman, Yol and Barry-Etienne. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia Battie-Laclau, cGF0cmljaWEubGFjbGF1QG15Y2VhLmZy
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.