 Lenka Mészárošová
Lenka Mészárošová Eliška Kuťáková3,4,5†
Eliška Kuťáková3,4,5† Petr Kohout
Petr Kohout Zuzana Münzbergová
Zuzana Münzbergová Petr Baldrian
Petr Baldrian
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 02 October 2024
Sec. Terrestrial Microbiology
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1416515
This article is part of the Research TopicResponse of Soil Microbial Structure and Function with Vegetation Succession in Temperate EcosystemsView all 8 articles
Introduction: Revegetation of barren substrates is often determined by the composition and distance of the nearest plant community, serving as a source of colonizing propagules. Whether such dispersal effect can be observed during the development of soil microbial communities, is not clear. In this study, we aimed to elucidate which factors structure plant and soil bacterial and fungal communities during primary succession on a limestone quarry spoil heap, focusing on the effect of distance to the adjoining xerophilous grassland.
Methods: We established a grid of 35 plots covering three successional stages – initial barren substrate, early successional community and late successional grassland ecosystem, the latter serving as the primary source of soil colonization. On these plots, we performed vegetation surveys of plant community composition and collected soil cores to analyze soil chemical properties and bacterial and fungal community composition.
Results: The composition of early successional plant community was significantly affected by the proximity of the source late successional community, however, the effect weakened when the distance exceeded 20 m. Early successional microbial communities were structured mainly by the local plant community composition and soil chemical properties, with minimal contribution of the source community proximity.
Discussion: These results show that on small spatial scales, species migration is an important determinant of plant community composition during primary succession while the establishment of soil microbial communities is not limited by dispersal and is primarily driven by local biotic and abiotic conditions.
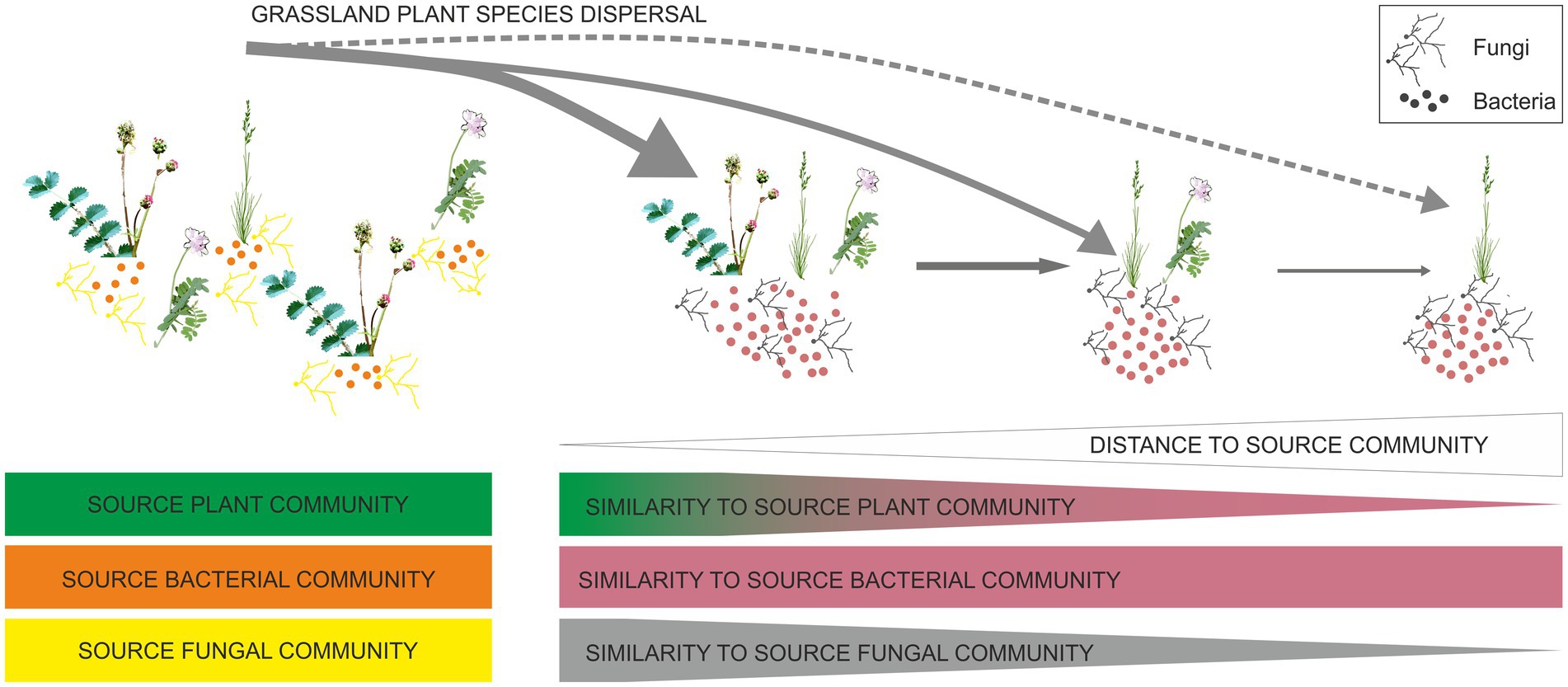
Graphical Abstract.
Surface mining activities, such as limestone mining, degrade large areas of land, leaving behind substrates that typically contain little to no organic matter, have low nutrient levels and are virtually devoid of residual biological legacy. As such, they present unique ecological niches to study primary succession, i.e., a process of de novo ecosystem development (Walker and Moral, 2011) in areas that are not constrained by extreme climate such as areas after glacier retreat. Primary succession has mostly been examined from a plant perspective, but it is crucial to recognize that soil microbes are important initial colonizers of newly exposed substrates (Nemergut et al., 2007; Harantová et al., 2017). Since microbes play pivotal roles in the accumulation and cycling of nutrients and soil organic matter before the onset of plant colonization (Kallenbach et al., 2016) and facilitate plant establishment (Schulz et al., 2013), they contribute, alongside plants, to ecosystem recovery. Primary succession should be thus seen and studied with a focus on both microbes and plants.
One of the key factors driving plant community assembly and development is propagule availability (Lichter, 2000; Lanta and Lepš, 2009). Propagule availability is closely linked to dispersal, which is especially important early in succession (Knappová and Münzbergová, 2015; Makoto and Wilson, 2019) when plants colonize empty niches and plant–plant interactions are minimal. Typically, most seeds disperse over a short distance from the parent plant with the number of seeds dispersing at longer distances dropping sharply (Cheplick, 2022). Accordingly, Novák and Konvička (2006) have shown that during the revegetation of abandoned quarries, the likelihood of a species establishing successfully depends on the size and proximity of its population in surrounding undisturbed vegetation patches.
Microbes are generally much less dispersal-limited than plants (Junker et al., 2021). However dispersal still influences the assembly of soil microbial communities (Wang et al., 2022a; Lemoine et al., 2023), with fungi being substantially more dispersal limited than bacteria (Schmidt et al., 2014; Powell et al., 2015; Zhang et al., 2021). Soil microbes disperse through a variety of vectors, including animals, rain and wind (Huffman et al., 2013; Golan and Pringle, 2017), with patches of vegetation serving as sources of microbial propagules (Redondo et al., 2020), analogous to their role in plant colonization. While information on the effect of the distance from source vegetation on microbial dispersal dynamics is sparse, especially for bacteria, it is conceivable that microbes exhibit dispersal patterns similar to plants (Barbour et al., 2023). Indeed, recent studies have shown that the abundance of fungal propagules decreases with increasing distance from potential spore sources (Peay et al., 2012; Grosdidier et al., 2018). Besides serving as potential reservoirs of microbial propagules, plants further contribute directly to microbial dispersal by introducing microbes into soil in and on seeds (Herrera Paredes and Lebeis, 2016), and indirectly, by attracting specific microbes via relatively long-distance (tens of cm) belowground chemical signalling (Bever et al., 2012; Schulz-Bohm et al., 2018).
This study aimed to investigate the effects of distance from the putative source habitat (approximately 10 to 50 meters) on the composition of plant and soil microbial communities and the drivers of their assembly in a limestone quarry spoil deposit undergoing primary succession. This deposit adjoins a late successional xerophilous grassland, which serves as a major source of both plant and microbial propagules. We hypotheize that (i) the composition of the early successional (ES) plant community on the spoil heap depends on distance from source plant community, meaning that ES plots closer to the late successional (LS) grassland will have a plant community more similar to the LS community; (ii) due to the effect on vegetation on soil microbiomes, similar distance-dependent spatial pattern in microbial community composition will be observed. This effect is anticipated to be stronger in fungi, which are known to be more responsive to vegetation than bacteria (Urbanová et al., 2015; Harantová et al., 2017).
The study was carried out on a limestone quarry spoil deposit and an adjacent species-rich dry calcareous grassland located in the Czech Karst Protected Landscape Area in the Central Bohemia, Czech Republic (49.96154 N°, 14.16454° E–49.96365° N, 14.16845° E). The surrounding hilly karstic landscape is characterized by relatively warm climate and mild winters (mean annual temperatures 8–9°C, mean annual precipitation 530 mm), the vegetation consists of thermophilic and xerothermic grasslands alongside deciduous forests and human settlements. Since 2009, the abandoned parts of the quarry have been gradually filled with clayey spoil from the deep layers of the quarry, and thus likely not containing any soil biota, and left to spontaneous revegetation. The oldest part of the spoil heap (ca. 150 × 100 m) represents an early primary successional habitat and is directly adjacent to a species-rich calcareous grassland with scattered trees, a late successional habitat, which serves as a source community for plant species colonizing the spoil heap (Kuťáková et al., 2020). Since 2005, the grassland has been grazed by sheep and goats.
The sampling took place on June 25, 2015. To determine how the proximity of the source community influences the composition of plant and soil microbial communities during early succession, we established five sampling transects, each perpendicular to the grassland edge (Supplementary Figure S1). At each transect, the first 1 m2 study plot was located within the late successional grassland, approx. 5 m from the grassland edge (LS) and another five points were located within the early successional (ES) spoil deposit area, 10 m (ES10), 20 m (ES20), 30 m (ES30), 40 m (ES40), and 50 m (ES50) from the first plot. Transects were parallel to each other and adjacent transects were separated by >10 meters (Supplementary Figure S1). To get an idea how the microbial communities looked at the beginning of succession, we established additional five plots in the area with the most recent spoil deposition from 2014, representing the initial succession (IS) phase. These plots were located approximately 200 meters away from the last early successional plots. At each plot, we collected 5 soil cores (diameter of 35 mm), one in the centre and the other four in each corner of the plot. The samples were brought to the laboratory and processed within 24 h. Soil up to the depth of 10 cm from all cores from each plot were pooled, litter, stones and roots were removed, and the soil was sieved through a 5 mm sieve. The samples were freeze-dried, weighed and stored at −40°C for further analyses.
On each plot, vegetation composition was assessed by recording all species of vascular plants and visually estimating their percentage ground cover (Supplementary Table S2). In order to capture all plant species present in the plots in their phenological optima, the vegetation sampling was conducted twice during the season of 2015, in the end of May and in the beginning of August, and the resulting dataset consisted of averaged values of the two observations. To assess which plant species are likely migrating from the adjacent grassland to the spoil deposit area, we used a detailed floristic survey performed in 2011 to a distance up to 100 m from the spoil heap (Kuťáková, unpublished data). The species found solely on the grassland but not in the other surrounding habitats were classified as grassland species. The nomenclature followed Kubát and Bělohlávková (2002).
To characterize soil chemical properties, soil samples were analyzed for pH (1,10 w/v soil-to-water ratio), available phosphorus (Pav), total nitrogen (Ntot) and total carbon (Ctot). Ntot, Ctot (using Carlo Erba Instrument NC 2500) and active pH (Zbíral, 2002) were measured by the Analytical laboratory of the Institute of Botany, Czech Academy of Sciences, Průhonice, Czech Republic. Pav was assessed in samples shaken with water (1,10, w,v) for 18 h, filtered through 0.22 μm membrane filter and measured with the malachite green method (Řezáčová et al., 2019). Soil moisture content was calculated based on the sample masses before and after freeze-drying. See Supplementary Table S1 for the overview and soil properties of the study plots.
Fungal and bacterial communities in soil samples were characterized by sequencing the internal transcribed spacer (ITS2) region and the V4 region of the 16S ribosomal RNA gene, respectively. DNA from each freeze-dried sample (350 mg of soil) was extracted in duplicates using the method of Miller modified by Sagova-Mareckova et al. (2008). DNA extracts were purified using Geneclean Turbo Kit (Biogenic) following the manufacturer’s instructions, duplicates were pooled and stored in −20°C before further use. For the microbial community analysis, PCR amplification of the fungal ITS2 region from DNA was performed using barcoded primers fITS7 and ITS4 (Ihrmark et al., 2012). The V4 region of bacterial 16S rRNA was amplified using the barcoded primers 515F and 806R (Caporaso et al., 2012). PCR was performed in triplicate for each sample as described previously (Tláskal et al., 2016; Žifčáková et al., 2016). The resulting amplicons were purified, pooled, and libraries prepared with the TruSeq DNA PCR-Free Kit (Illumina) were sequenced in house on the Illumina MiSeq (2 × 250-base reads).
The amplicon sequencing data processing was done using the pipeline SEED 2.0.3 (Větrovský et al., 2018). Briefly, paired-end reads were merged using fastq-join (Aronesty, 2013) and fungal ITS2 region was extracted using ITSx 1.0.9 (Bengtsson-Palme et al., 2013). ITS and 16S sequences were then clustered at 97% similarity into operational taxonomic units (OTUs) using UPARSE implemented in USEARCH7 (Edgar, 2013), detected chimeric sequences were deleted. The most abundant sequence was determined for each cluster, and its closest hit at a genus or species level was identified using blastn against the Ribosomal Database Project (Cole et al., 2014) and UNITE 8 (Nilsson et al., 2019). Sequences identified as nonbacterial or nonfungal were discarded. Fungal guilds were determined based on the primary and secondary lifestyle in the FungalTraits database (Põlme et al., 2020). Sequence data have been deposited in the SRA under accession number PRJNA603889.
Illumina Miseq Sequencing yielded 596,904 bacterial and 433,179 fungal sequences. Bacterial sequences clustered into 15,122 OTUs, including 6,533 singletons; fungal sequences clustered into 13,191 OTUs, of which 8,268 were singletons. For microbial alpha diversity calculations, bacterial and fungal datasets including singletons were subsampled to the minimum number of reads (6,506 for bacteria, 3,390 for fungi). For further analyses, singletons and doubletons were filtered out and bacterial and fungal datasets were subsampled to 10,000 and 6,000 reads, respectively. We ended up with 336,757 bacterial sequences, clustering into 6,404 OTUs, and 202,720 fungal sequences clustering into 3,079 OTUs. Statistical analyses were performed on datasets containing only OTUs occurring with an abundance of 0.1% in at least three samples (3,072 bacterial OTUs, 96% of sequences; 1775 fungal OTUs, 97% of sequences).
Statistical analyses were performed using R software v. 4.1.0 (R Core Team and R Development Core Team, 2021), package vegan (Oksanen et al., 2022). The R-Code used in this article is available from the authors upon request.
Prior to statistical analyses, soil physicochemical variables except for pH (total soil N and C, available soil P, soil moisture) were log-transformed to normalize distribution and plant species cover data and bacterial and fungal OTU abundances were Hellinger-transformed. One of the fungal LS samples was dropped due to poor sequencing performance.
The observed species richness and evenness of bacterial and fungal communities were calculated on datasets rarefied to the minimum number of reads. Evenness was calculated as Shannon diversity index/ln(observed species richness).
We used linear regression to investigate how individual soil chemical characteristics, plant species richness and cover, and soil microbial species richness change with distance from the source grassland community. The grassland plant species cover ratio for each ES plot was calculated the cover of grassland plant species divided by the total plant cover in that plot. To assess variation in beta diversities among microbial groups and plots, we measured the multivariate homogeneity of group dispersions using the function betadisper based on Bray–Curtis dissimilarities (Anderson et al., 2006); the results were visualized as mean distance-to-centroid values. As we wanted to detect the effect of source community proximity on microbial community assembly on the spatial scale, we compared the community composition dissimilarity (Bray–Curtis) of sample pairs within each transect considering their distances. Compositional dissimilarities within the ES distance classes and between ES and IS transect plots were calculated analogously. The overall correlation between bacterial and fungal communities was calculated on full Bray–Curtis dissimilarity matrices using Mantel test.
To test if soil chemical properties, plant community composition and soil microbial community composition differed across transects, we performed permutational multivariate analysis of variance (PERMANOVA) on respective datasets using Euclidean distance matrix for soil chemical properties and Bray–Curtis distance matrix for microbial and plant data. We used PCA to visualize the compositional differences in soil chemical properties among the samples. For microbial and plant communities, we used NMDS, using the envfit function to fit the environmental variables onto the ordinations. For plants, we depicted only those species that contributed more than 30% of the variation (p < 0.05), as determined by envfit.
To assess the relative importance of space, soil chemistry, and plant community composition for structuring soil microbial communities during early succession, we performed the variation partitioning analysis (Legendre et al., 2005) using the function varpart. Variation partitioning combines redundancy analysis (RDA) and partial RDA to partition the variation of a response dataset among multiple explanatory matrices. Each explanatory matrix uniquely explains a portion of the variation in the response dataset. Additionally, another portion of the variation might be explained by the combined effect of two or more matrices, indicating correlations between the explanatory matrices. The residual variation in the response dataset is not explained by any explanatory matrix or their combination.
Plant community data were represented by the scores of the NMDS axes. The spatial predictors were derived from the GPS coordinates of individual samples using the principal coordinates of neighbourhood matrix (PCNM) method (Borcard and Legendre, 2002). To select the most relevant predictors of microbial data variation, independent double-stop forward selections (adespatial) (Dray et al., 2023) of soil chemical variables and PCNM vectors were carried out prior to the variation partitioning. To disentangle the effects of distance and distance-unrelated spatial variability, we linearly detrended bacterial, fungal, and plant community data to remove the distance aspect, represented as linear and second-order polynomial terms. The residuals were then used to select relevant PCNM vectors via forward selection, capturing distance-unrelated spatial variability. Variation partitioning was subsequently performed using the non-detrended microbial and plant datasets, with distance included as an independent variable.
Soil chemistry differed by habitat, with soil nutrient content increasing and soil pH decreasing with habitat successional stage. The early successional plots (ES) were more similar to the initial (IS) than to the late successional (LS) soil and did not show significant differences among distance classes (Figure 1). However, soil pH of the ES plots significantly increased with distance from the late successional habitat (R2adj = 0.18, p < 0.05, Figure 1).
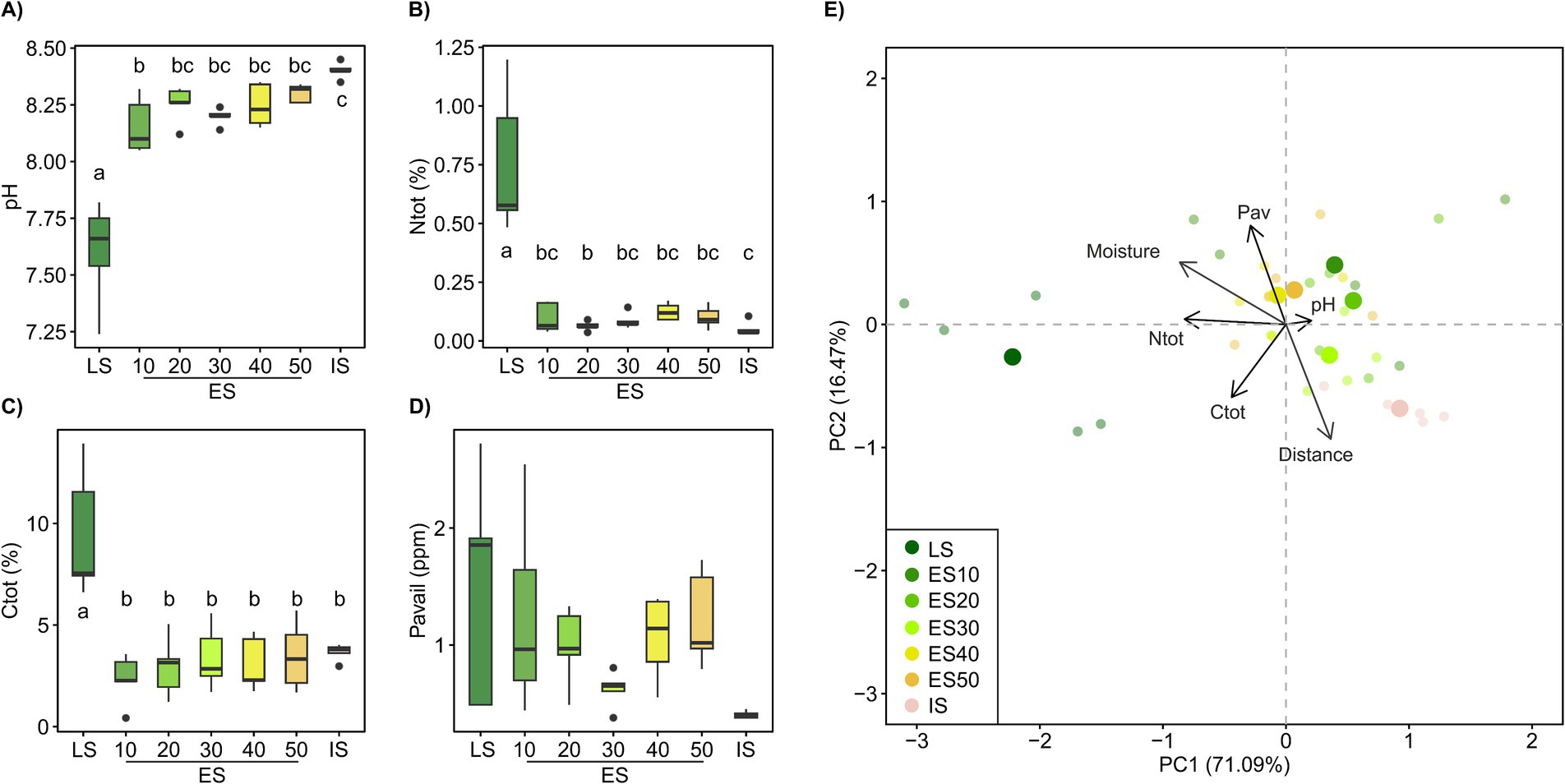
Figure 1. Soil properties, pH, total N (Ntot), total C (Ctot), and available P (Pav), across a successional grassland sequence. In (A–D), treatments with different letters are significantly different (p < 0.05, ANOVA, Tukey’s test). (E) The principal component analysis (PCA) biplot shows clustering of samples based on their soil chemical properties, the “distance” vector represents the distance to the LS and was added to the ordination as a supplementary variable using envfit. LS, late successional plots; ES, early successional plots, numbers denote distance (in m) to the LS; IS, initial soil.
Plant species richness and vegetation cover of the ES plots were not significantly different from those of the LS community (Figures 2A,B), but both tended to decrease with distance from the LS (Supplementary Figure S2). Plant species richness across the ES transects was significantly negatively correlated with soil Ntot and Pav (Supplementary Figure S2). The grassland plant species cover significantly decreased with distance to the LS (R2adj = 0.43, p < 0.001, Figures 2C,D). Plant community composition in ES significantly differed not only from LS plant community but also among ES distance classes, with ES10 being significantly different from ES30-50 and ES20 falling in between (Figure 2E). The more distant the ES locations, the more dissimilar to the LS plant communities they were (Figure 3C); plant cover showed marginally significant decrease (Supplementary Figure S2). A significant decrease of similarity with distance was also observed among ES communities (Figure 3F). ES plant community variation exhibited strong spatial structure. The most significant factor influencing plant community composition across the ES plots was the distance to the LS, accounting for over 11% of the community variation. Additionally, spatial heterogeneity unrelated to distance explained another 5% of the community variation (Supplementary Figure S3). These results are consistent with our first hypothesis, confirming the decisive role of source community proximity in the assembly of ES plant communities. In line with the findings of Novák and Konvička (2006), who demonstrated distance-dependent differences in successional trajectories and establishment success of source grassland species in abandoned basalt quarries, we observed a tipping point in ES plant community assembly at a distance of 30 meters from LS (Figure 2E).
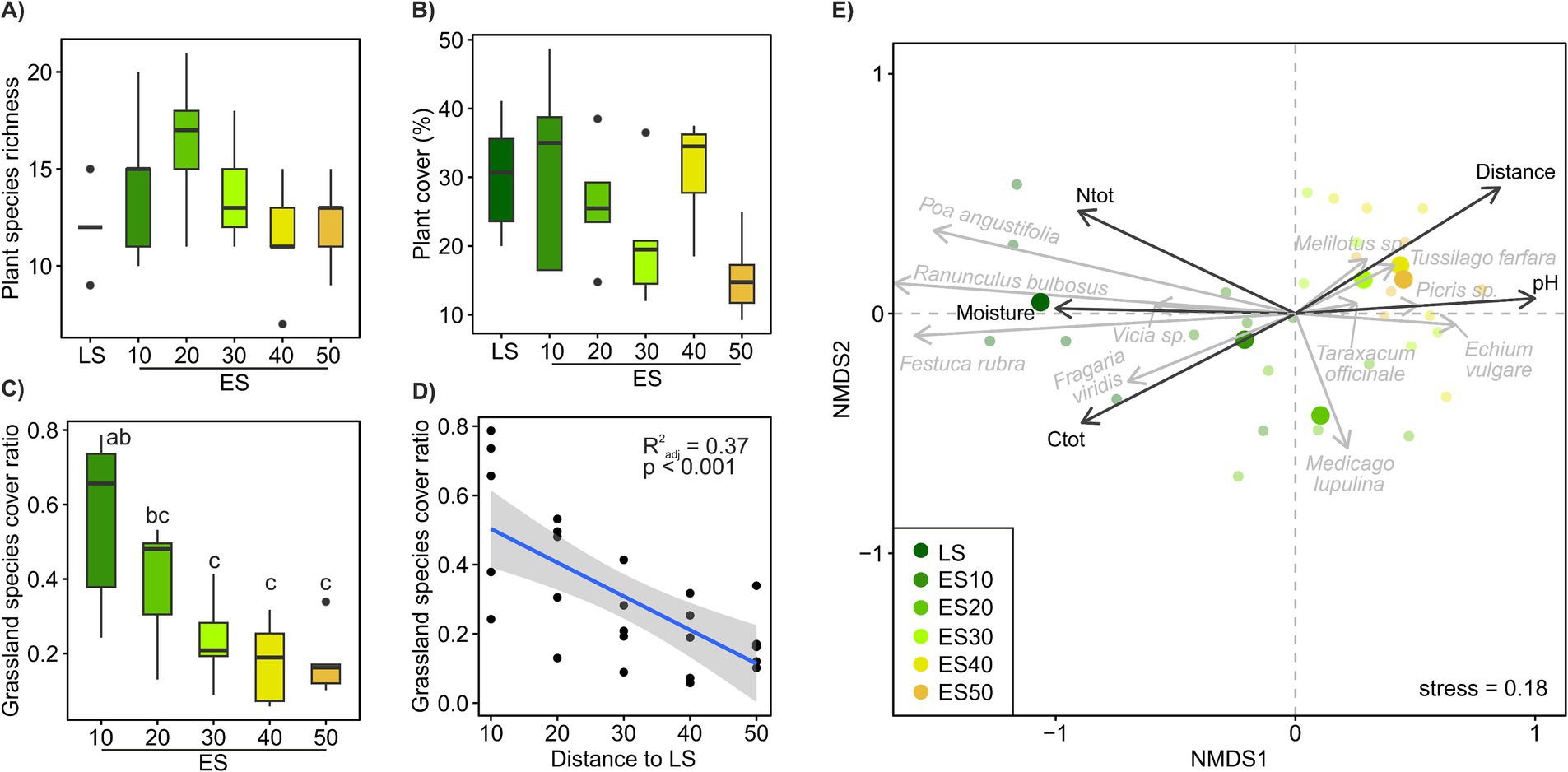
Figure 2. Plant community composition across a successional grassland sequence. (A–D) Plant community properties. Treatments with different letters are significantly different (p < 0.05, ANOVA, Tukey’s test). The scatter plot illustrates the relationship between the share of grassland plant species cover in ES plots and their distance from the LS plots. (E) Beta-diversity of the plant community. Non-metric multidimensional scaling (NMDS) ordination based on Bray–Curtis distances calculated on Hellinger-transformed plant species cover data (grey arrows), environmental variables significantly correlated (p < 0.05) with ordination axes are shown as vectors (black arrows). Ntot, total N; Ctot, total C; Pav, available P.
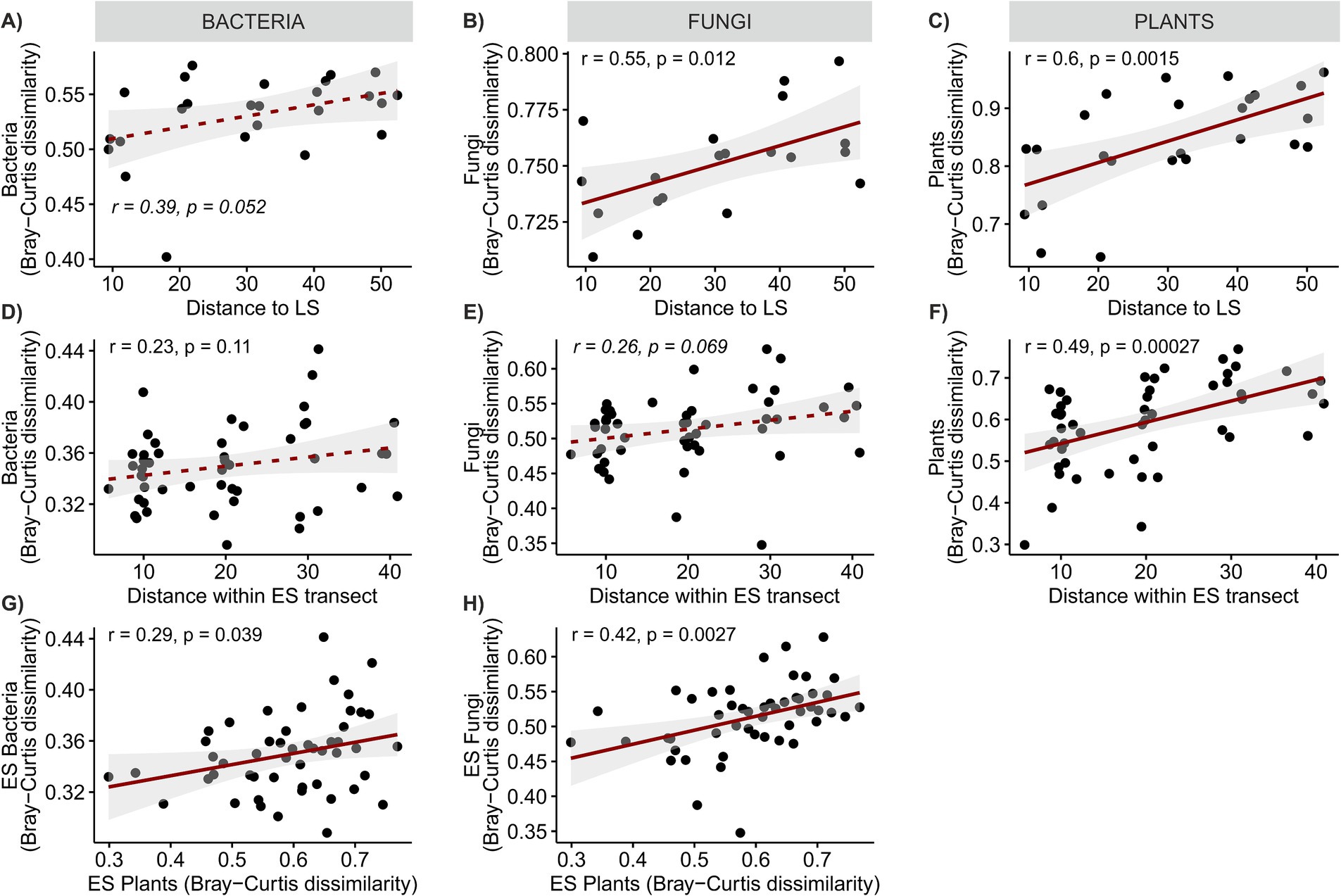
Figure 3. Linear regressions of (A–C) the dissimilarity of ES plot communities from the LS community and distance to LS, (D–F) dissimilarity of ES plant communities and distance between ES plots, and (G,H) similarity of microbial and plant community composition. Solid lines denote significant relationships at p < 0.05, dashed lines denote non-significant relationship.
The differences in taxonomic composition of both bacterial and fungal communities were especially pronounced at finer taxonomic levels (Supplementary Figure S4). Bacterial communities exhibited higher overall evenness compared to fungal communities, and both microbial groups exhibited significant differences in community evenness across the three successional habitats, with the highest evenness observed in the ES communities. Fungal IS community was particularly uneven with almost 70% of sequences belonging to top five OTUs, whereas the LS and ES communities were more balanced (Supplementary Figures S4C,D).
Both bacterial and fungal OTU richness tended to be higher in the ES soil than in the LS or IS soil (Figures 4A,B,D,E). Bacterial richness in ES was influenced neither by distance to the LS transect, nor by any of the soil properties or plant species richness and/or cover. Contrastingly, ES fungal richness was significantly positively correlated with plant species richness and soil moisture (Supplementary Figure S2). The relationship between plant and soil microbial species richness has been extensively studied, with varied results. Some studies in grasslands have reported a positive correlation between plant and soil fungal (Chen et al., 2017; Dassen et al., 2017; Yang et al., 2017) or bacterial (Yang et al., 2018) species richness, while others found no significant relationship (Prober et al., 2015; Navrátilová et al., 2018; Guasconi et al., 2023). Our results suggest a stronger link between plant and soil fungal richness compared to bacterial richness, consistent with the findings by Wang et al. (2022b) or Lepinay et al. (2024).
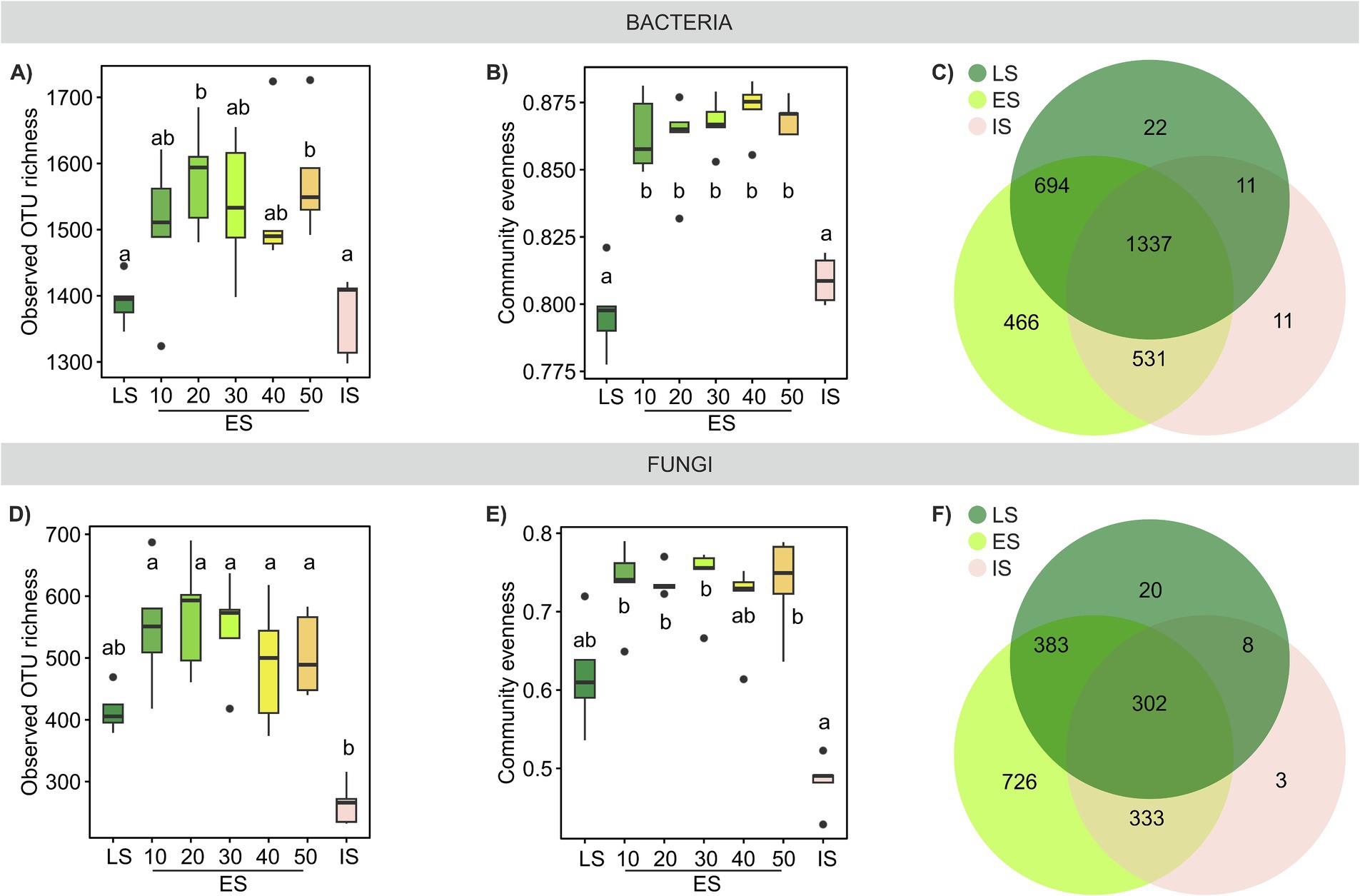
Figure 4. Microbial richness and evenness across a successional grassland sequence. (A,D) Observed OTU richness and (B,E) community evenness of bacterial and fungal communities; treatments with different letters are significantly different (p < 0.05, ANOVA, Tukey’s test). (C,F) Venn diagrams illustrate the number of OTUs specific to each habitat or shared among them.
Plant species richness influences fungal richness in several ways. Firstly, higher plant species diversity affects the quantity and chemical diversity of plant litter (Schnitzer et al., 2011; Santonja et al., 2017), modulates metabolite production of individual plant species (Scherling et al., 2010), and enhances the amount and diversity of plant exudates (Steinauer et al., 2016; Eisenhauer et al., 2017). With fungi being the main decomposers of recalcitrant plant-derived organic matter (López-Mondéjar et al., 2018) and receivers of a substantial portion of recent plant photosynthate carbon (Hawkins et al., 2023), the diversity of plant-derived nutrients may directly influence the diversity of soil fungal communities (Broeckling et al., 2008; Cline and Zak, 2015). Secondly, higher plant species richness has been associated with greater diversity in communities of fungal plant pathogens (Liu et al., 2021) and AMF (Hiiesalu et al., 2014), fungal guilds particularly abundant in the ES community (Supplementary Figure S4E). The positive correlation between fungal species richness and soil moisture suggests a potential role for microclimate, possibly through increased shading of more species-rich ES plots (Fischer et al., 2019). Alternatively, the higher plant cover in ES plots closer to the grassland may also play a role. Early plant establishment results in longer accumulation of organic matter, subsequently increasing water retention potential of the soil (Lal and Rattan Lal, 2020).
Bacterial and fungal community similarities across the three successional habitats were substantially correlated (Supplementary Figure S5A); however, the beta diversity of bacterial communities was significantly lower than that of fungal communities (Supplementary Figure S5B). While both bacterial and fungal communities significantly differed by habitat (PERMANOVA R2adj = 0.46, p < 0.001 and R2adj = 0.31, p < 0.001, respectively, Figures 5A,B), bacterial taxa were more ubiquitous than those of fungi, resulting in a larger community overlap among ES, LS, and IS compared to fungal OTUs. The proportion of OTUs unique to IS was notably low for both bacteria and fungi (Figures 4C,F).
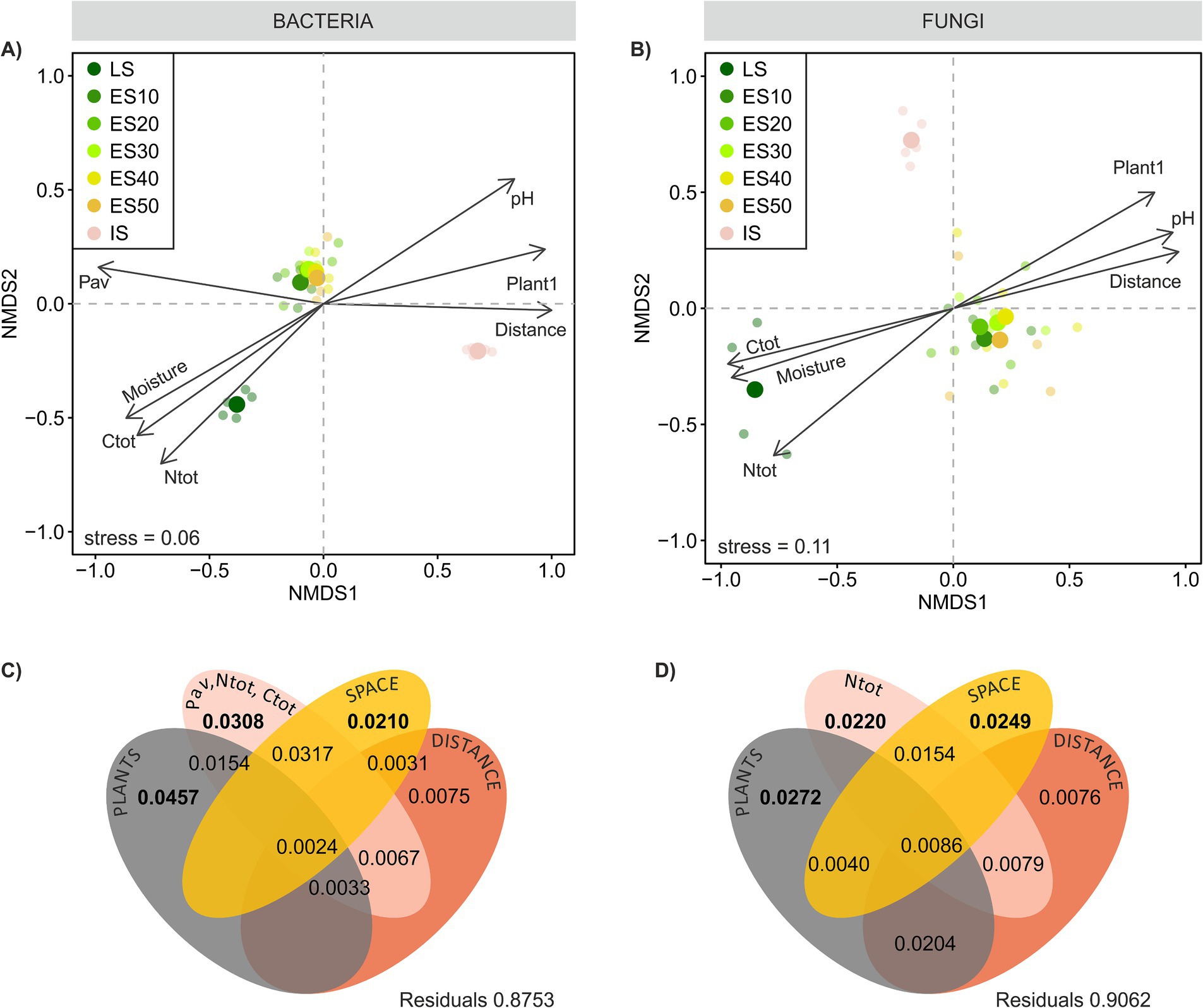
Figure 5. Bacterial and fungal community composition across a successional grassland sequence. (A,B) Beta-diversity of the microbial communities. Non-metric multidimensional scaling (NMDS) ordination based on Bray–Curtis distances calculated on Hellinger-transformed OTU abundance data, environmental variables significantly correlated (p < 0.05) with ordination axes were fitted onto the ordinations as vectors using envfit. Ntot, total N; Ctot, total C; Pav, available P; Plant1, scores of NMDS axis 1 (C,D) Partitioning of variance in ES microbial community composition among plant community composition, soil chemistry, spatial variables, and their joint effects as predictors. For each testable fraction, an adjusted R2 is given, and significance (p < 0.05) is indicated in bold.
We expected to observe a clear distance gradient in microbial ES community composition, however, our assumptions were only partially met. Microbial communities across ES transects were rather homogeneous, in bacteria more so than in fungi, and their composition did not exhibit any apparent distance-related patterns akin to those observed in plants (Figures 3D,E, 5A,B). These results concur with those of García de León et al. (2016) who found no clear effect of presumed propagule source distance on the small-scale dispersal (tens of meters) of AMF during early succession, Bacterial ES community composition was more similar to the LS than IS community (average pairwise community dissimilarity was 0.53 ± 0.007 and 0.60 ± 0.008, respectively, p < 0.001), while fungal ES community resembled more the IS than the LS community (average pairwise community dissimilarity was 0.70 ± 0.015 and 0.75 ± 0.005, respectively, p < 0.01). Both bacterial and fungal ES communities became progressively more dissimilar to the LS community with increasing distance (Figures 3A,B). Additionally, both bacterial and fungal ES community dissimilarity was significantly positively correlated with plant ES community dissimilarity (Figures 3G,H). In both cases, these phenomena were more pronounced for fungi. The prominent role of plant community composition in structuring the ES soil microbial communities was confirmed by the variation partitioning. The analysis explained 9.4 and 12.5% of the variation in ES fungal and bacterial community composition, respectively, and identified the ES plant community composition as the most important determinant of variation in both the bacterial and the fungal communities, which is in line with previous studies on microbial succession (Knelman et al., 2012; Harantová et al., 2017). The second strongest driver of microbial community composition was soil nutrient content, followed by distance-unrelated spatial structuring. Bacteria, in particular, appeared sensitive to spatial gradients of soil nutrients, as evidenced by the shared effect of space and soil chemistry, which was as strong as the effect of soil chemistry alone. This might reflect the fact that their small body size and limited motility make bacteria more sensitive than fungi to even very local and small-scale nutrient fluctuations in their immediate environment (Vos et al., 2013).
The distance to the LS per se did not directly affect bacterial or fungal community composition. Instead, its impact on fungal, but not bacterial, community composition manifested indirectly through shared effects with plants, reflecting spatial gradients within the ES plant community composition (Figures 5C,D). In any case, the spatial effects were rather weak, and our results suggest that during primary succession on small spatial scales, the development of soil microbial communities is likely more constrained by environmental filtering than by dispersal limitations. Alternatively, the absence of a distance gradient in the composition of ES microbial communities might be caused by the limited time of development of only 6 years. Our previous study on succession (Harantová et al., 2017) showed that fungal communities started to diverge later, alongside more profound changes in vegetation composition, and changes in bacterial community composition were even less evident. Since the ES plant community composition was the primary determinant of the ES microbial community composition, we can expect that soil microbes, particularly fungi, will follow future changes in plant community composition. Over time the microbiome composition may either differentiate with vegetation differentiation or, at a later stage, converge as plants overcome dispersal limitations. It should be noted that the plant effect is stronger in root microbiomes than in surrounding soil (Mészárošová et al., 2024). One can thus not rule out that the differences in plant communities in the ES, while not affecting soil microbes, might possibly be visible in the root microbiome.
Source habitat proximity was the main structuring force of early successional plant community composition, while having only minimal influence over the early successional microbial communities. Soil microbial community variation was determined primarily by early successional plant community composition and soil nutrient content, indicating local conditions play a more significant role than dispersal limitations.
The datasets analyzed for this study can be found in the Sequence Read Archive [https://www.ncbi.nlm.nih.gov/sra] under the accession number PRJNA603889.
LM: Data curation, Formal analysis, Methodology, Visualization, Writing – original draft, Writing – review & editing, Investigation, Conceptualization, Project administration. EK: Data curation, Methodology, Visualization, Writing – review & editing, Conceptualization, Investigation. PK: Writing – review & editing. ZM: Conceptualization, Funding acquisition, Writing – review & editing, Resources. PB: Conceptualization, Funding acquisition, Supervision, Writing – review & editing, Methodology, Project administration, Resources, Validation.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Czech Science Foundation (21-17749S) and by the Ministry of Education, Youth and Sports of the Czech Republic (CZ.02.01.01/00/22_08/0004635 - AdAgriF - Advanced methods of greenhouse gases emission reduction and sequestration in agriculture and forest landscape for climate change mitigation).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1416515/full#supplementary-material
Anderson, M. J., Ellingsen, K. E., and McArdle, B. H. (2006). Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 9, 683–693. doi: 10.1111/J.1461-0248.2006.00926.X
Aronesty, E. (2013). Comparison of sequencing utility programs. Open Bioinf. J. 7, 1–8. doi: 10.2174/1875036201307010001
Barbour, K. M., Barrón-Sandoval, A., Walters, K. E., and Martiny, J. B. H. (2023). Towards quantifying microbial dispersal in the environment. Environ. Microbiol. 25, 137–142. doi: 10.1111/1462-2920.16270
Bengtsson-Palme, J., Ryberg, M., Hartmann, M., Branco, S., Wang, Z., Godhe, A., et al. (2013). Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 4, 914–919. doi: 10.1111/2041-210X.12073
Bever, J. D., Platt, T. G., and Morton, E. R. (2012). Microbial population and community dynamics on plant roots and their feedbacks on plant communities. Ann. Rev. Microbiol. 66, 265–283. doi: 10.1146/annurev-micro-092611-150107
Borcard, D., and Legendre, P. (2002). All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Model. 153, 51–68. doi: 10.1016/S0304-3800(01)00501-4
Broeckling, C. D., Broz, A. K., Bergelson, J., Manter, D. K., and Vivanco, J. M. (2008). Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 74, 738–744. doi: 10.1128/AEM.02188-07
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 6, 1621–1624. doi: 10.1038/ismej.2012.8
Chen, Y. L., Xu, T.-L., Veresoglou, S. D., Hu, H. W., Hao, Z. P., Hu, Y. J., et al. (2017). Plant diversity represents the prevalent determinant of soil fungal community structure across temperate grasslands in northern China. Soil Biol. Biochem. 110, 12–21. doi: 10.1016/j.soilbio.2017.02.015
Cheplick, G. P. (2022). Philomatry in plants: why do so many species have limited seed dispersal? Am. J. Bot. 109, 29–45. doi: 10.1002/AJB2.1791
Cline, L. C., and Zak, D. R. (2015). Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 96, 3374–3385. doi: 10.1890/15-0184.1
Cole, J. R., Wang, Q., Fish, J. A., Chai, B., McGarrell, D. M., Sun, Y., et al. (2014). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, D633–D642. doi: 10.1093/nar/gkt1244
Dassen, S., Cortois, R., Martens, H., de Hollander, M., Kowalchuk, G. A., van der Putten, W. H., et al. (2017). Differential responses of soil bacteria, fungi, archaea and protists to plant species richness and plant functional group identity. Mol. Ecol. 26, 4085–4098. doi: 10.1111/mec.14175
Dray, S., Bauman, D., Blanchet, G., Borcard, D., Clappe, S., Guénard, G., et al., (2023), {adespatial}: Multivariate Multiscale Spatial Analysis.
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Eisenhauer, N., Lanoue, A., Strecker, T., Scheu, S., Steinauer, K., Thakur, M. P., et al. (2017). Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 7:44641. doi: 10.1038/srep44641
Fischer, C., Leimer, S., Roscher, C., Ravenek, J., de Kroon, H., Kreutziger, Y., et al. (2019). Plant species richness and functional groups have different effects on soil water content in a decade-long grassland experiment. J. Ecol. 107, 127–141. doi: 10.1111/1365-2745.13046
García de León, D., Moora, M., Öpik, M., Jairus, T., Neuenkamp, L., Vasar, M., et al. (2016). Dispersal of arbuscular mycorrhizal fungi and plants during succession. Acta Oecol. 77, 128–135. doi: 10.1016/j.actao.2016.10.006
Golan, J. J., and Pringle, A. (2017). Long-distance dispersal of fungi. Microbiol. Spectrum 5:FUNK-0047-2016. doi: 10.1128/microbiolspec.funk-0047-2016
Grosdidier, M., Ioos, R., Husson, C., Cael, O., Scordia, T., and Marçais, B. (2018). Tracking the invasion: dispersal of Hymenoscyphus fraxineus airborne inoculum at different scales. FEMS Microbiol. Ecol. 94:fiy049. doi: 10.1093/FEMSEC/FIY049
Guasconi, D., Juhanson, J., Clemmensen, K. E., Cousins, S. A. O., Hugelius, G., Manzoni, S., et al. (2023). Vegetation, topography, and soil depth drive microbial community structure in two Swedish grasslands. FEMS Microbiol. Ecol. 99:fiad080. doi: 10.1093/FEMSEC/FIAD080
Harantová, L., Mudrák, O., Kohout, P., Elhottová, D., Frouz, J., and Baldrian, P. (2017). Development of microbial community during primary succession in areas degraded by mining activities. Land Degrad. Dev. 28, 2574–2584. doi: 10.1002/ldr.2817
Hawkins, H. J., Cargill, R. I. M., Van Nuland, M. E., Hagen, S. C., Field, K. J., Sheldrake, M., et al. (2023). Mycorrhizal mycelium as a global carbon pool. Curr. Biol. 33, R560–R573. doi: 10.1016/J.CUB.2023.02.027
Herrera Paredes, S., and Lebeis, S. L. (2016). Giving back to the community: microbial mechanisms of plant–soil interactions. Funct. Ecol. 30, 1043–1052. doi: 10.1111/1365-2435.12684
Hiiesalu, I., Pärtel, M., Davison, J., Gerhold, P., Metsis, M., Moora, M., et al. (2014). Species richness of arbuscular mycorrhizal fungi: associations with grassland plant richness and biomass. New Phytol. 203, 233–244. doi: 10.1111/nph.12765
Huffman, J. A., Prenni, A. J., Demott, P. J., Pöhlker, C., Mason, R. H., Robinson, N. H., et al. (2013). High concentrations of biological aerosol particles and ice nuclei during and after rain. Atmos. Chem. Phys. 13, 6151–6164. doi: 10.5194/acp-13-6151-2013
Ihrmark, K., Bödeker, I. T. M., Cruz-Martinez, K., Friberg, H., Kubartova, A., Schenck, J., et al. (2012). New primers to amplify the fungal ITS2 region—evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 82, 666–677. doi: 10.1111/j.1574-6941.2012.01437.x
Junker, R. R., He, X., Otto, J. C., Ruiz-Hernández, V., and Hanusch, M. (2021). Divergent assembly processes? A comparison of the plant and soil microbiome with plant communities in a glacier forefield. FEMS Microbiol. Ecol. 97:fiab135. doi: 10.1093/femsec/fiab135
Kallenbach, C. M., Frey, S. D., and Grandy, A. S. (2016). Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 7:13630. doi: 10.1038/ncomms13630
Knappová, J., and Münzbergová, Z. (2015). Low seed pressure and competition from resident vegetation restricts dry grassland specialists to edges of abandoned fields. Agric. Ecosyst. Environ. 200, 200–207. doi: 10.1016/J.AGEE.2014.11.008
Knelman, J. E., Legg, T. M., O’Neill, S. P., Washenberger, C. L., González, A., Cleveland, C. C., et al. (2012). Bacterial community structure and function change in association with colonizer plants during early primary succession in a glacier forefield. Soil Biol. Biochem. 46, 172–180. doi: 10.1016/j.soilbio.2011.12.001
Kuťáková, E., Mészárošová, L., Baldrian, P., and Münzbergová, Z. (2020). Evaluating the role of biotic and chemical components of plant-soil feedback of primary successional plants. Biol. Fertil. Soils 56, 345–358. doi: 10.1007/s00374-019-01425-z
Lal, R., and Rattan Lal, C. (2020). Soil organic matter and water retention. Agron. J. 112, 3265–3277. doi: 10.1002/AGJ2.20282
Lanta, V., and Lepš, J. (2009). How does surrounding vegetation affect the course of succession: a five-year container experiment. J. Veg. Sci. 20, 686–694. doi: 10.1111/J.1654-1103.2009.01061.X
Legendre, P., Borcard, D., and Peres-Neto, P. R. (2005). Analyzing beta diversity: partitioning the spatial variation of community composition data. Ecol. Monogr. 75, 435–450. doi: 10.1890/05-0549
Lemoine, N. P., Adams, B. J., Diaz, M., Dragone, N. B., Franco, A. L. C., Fierer, N., et al. (2023). Strong dispersal limitation of microbial communities at Shackleton glacier. MSystems 8:e0125422. doi: 10.1128/msystems.01254-22
Lepinay, C., Větrovský, T., Chytrý, M., Dřevojan, P., Fajmon, K., Cajthaml, T., et al. (2024). Effect of plant communities on bacterial and fungal communities in a central European grassland. Environ. Microb. 19, 1–19. doi: 10.1186/s40793-024-00583-4
Lichter, J. (2000). Colonization constraints during primary succession on coastal Lake Michigan sand dunes. J. Ecol. 88, 825–839. doi: 10.1046/J.1365-2745.2000.00503.X
Liu, X., Zhang, L., Huang, M., and Zhou, S. (2021). Plant diversity promotes soil fungal pathogen richness under fertilization in an alpine meadow. J. Plant Ecol. 14, 323–336. doi: 10.1093/JPE/RTAA099
López-Mondéjar, R., Brabcová, V., Štursová, M., Davidová, A., Jansa, J., Cajthaml, T., et al. (2018). Decomposer food web in a deciduous forest shows high share of generalist microorganisms and importance of microbial biomass recycling. ISME J. 12, 1768–1778. doi: 10.1038/s41396-018-0084-2
Makoto, K., and Wilson, S. D. (2019). When and where does dispersal limitation matter in primary succession? J. Ecol. 107, 559–565. doi: 10.1111/1365-2745.12988
Mészárošová, L., Kuťáková, E., Kohout, P., Münzbergová, Z., and Baldrian, P. (2024). Plant effects on microbiome composition are constrained by environmental conditions in a successional grassland. Environ. Microb. 19:8. doi: 10.1186/s40793-024-00550-z
Navrátilová, D., Tláskalová, P., Kohout, P., Dřevojan, P., Fajmon, K., Chytrý, M., et al. (2018). Diversity of fungi and bacteria in species-rich grasslands increases with plant diversity in shoots but not in roots and soil. FEMS Microbiol. Ecol. 95:fiy208. doi: 10.1093/femsec/fiy208
Nemergut, D. R., Anderson, S. P., Cleveland, C. C., Martin, A. P., Miller, A. E., Seimon, A., et al. (2007). Microbial community succession in an unvegetated, recently deglaciated soil. Microb. Ecol. 53, 110–122. doi: 10.1007/s00248-006-9144-7
Nilsson, R. H., Larsson, K. H., Taylor, A. F. S., Bengtsson-Palme, J., Jeppesen, T. S., Schigel, D., et al. (2019). The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 47, D259–D264. doi: 10.1093/nar/gky1022
Novák, J., and Konvička, M. (2006). Proximity of valuable habitats affects succession patterns in abandoned quarries. Ecol. Eng. 26, 113–122. doi: 10.1016/j.ecoleng.2005.06.008
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., et al., (2022). Vegan: community ecology package
Peay, K. G., Schubert, M. G., Nguyen, N. H., and Bruns, T. D. (2012). Measuring ectomycorrhizal fungal dispersal: macroecological patterns driven by microscopic propagules. Mol. Ecol. 21, 4122–4136. doi: 10.1111/j.1365-294X.2012.05666.x
Põlme, S., Abarenkov, K., Henrik Nilsson, R., Lindahl, B. D., Clemmensen, K. E., Kauserud, H., et al. (2020). FungalTraits: a user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 105, 1–16. doi: 10.1007/s13225-020-00466-2
Powell, J. R., Karunaratne, S., Campbell, C. D., Yao, H., Robinson, L., and Singh, B. K. (2015). Deterministic processes vary during community assembly for ecologically dissimilar taxa. Nat. Commun. 6:8444. doi: 10.1038/ncomms9444
Prober, S. M., Leff, J. W., Bates, S. T., Borer, E. T., Firn, J., Harpole, W. S., et al. (2015). Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 18, 85–95. doi: 10.1111/ele.12381
R Core Team and R Development Core Team, (2021). R: a Language and Environment for Statistical Computing
Redondo, M. A., Berlin, A., Boberg, J., and Oliva, J. (2020). Vegetation type determines spore deposition within a forest-agricultural mosaic landscape. FEMS Microbiol. Ecol. 96:fiaa082. doi: 10.1093/femsec/fiaa082
Řezáčová, V., Slavíková, R., Konvalinková, T., Zemková, L., Řezáč, M., Gryndler, M., et al. (2019). Geography and habitat predominate over climate influences on arbuscular mycorrhizal fungal communities of mid-European meadows. Mycorrhiza 29, 567–579. doi: 10.1007/s00572-019-00921-2
Sagova-Mareckova, M., Cermak, L., Novotna, J., Plhackova, K., Forstova, J., and Kopecky, J. (2008). Innovative methods for soil DNA purification tested in soils with widely differing characteristics. Appl. Environ. Microbiol. 74, 2902–2907. doi: 10.1128/AEM.02161-07
Santonja, M., Rancon, A., Fromin, N., Baldy, V., Hättenschwiler, S., Fernandez, C., et al. (2017). Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biol. Biochem. 111, 124–134. doi: 10.1016/J.SOILBIO.2017.04.006
Scherling, C., Roscher, C., Giavalisco, P., Schulze, E. D., and Weckwerth, W. (2010). Metabolomics unravel contrasting effects of biodiversity on the performance of individual plant species. PLoS One 5:e12569. doi: 10.1371/journal.pone.0012569
Schmidt, S. K., Nemergut, D. R., Darcy, J. L., and Lynch, R. (2014). Do bacterial and fungal communities assemble differently during primary succession? Mol. Ecol. 23, 254–258. doi: 10.1111/mec.12589
Schnitzer, S. A., Klironomos, J. N., HilleRisLambers, J., Kinkel, L. L., Reich, P. B., Xiao, K., et al. (2011). Soil microbes drive the classic plant diversity-productivity pattern. Ecology 92, 296–303. doi: 10.1890/10-0773.1
Schulz, S., Brankatschk, R., Dümig, A., Kögel-Knabner, I., Schloter, M., and Zeyer, J. (2013). The role of microorganisms at different stages of ecosystem development for soil formation. Biogeosciences 10, 3983–3996. doi: 10.5194/bg-10-3983-2013
Schulz-Bohm, K., Gerards, S., Hundscheid, M., Melenhorst, J., De Boer, W., and Garbeva, P. (2018). Calling from distance: attraction of soil bacteria by plant root volatiles. ISME J. 12, 1252–1262. doi: 10.1038/S41396-017-0035-3
Steinauer, K., Chatzinotas, A., and Eisenhauer, N. (2016). Root exudate cocktails: the link between plant diversity and soil microorganisms? Ecol. Evol. 6, 7387–7396. doi: 10.1002/ece3.2454
Tláskal, V., Voříšková, J., and Baldrian, P. (2016). Bacterial succession on decomposing leaf litter exhibits a specific occurrence pattern of cellulolytic taxa and potential decomposers of fungal mycelia. FEMS Microbiol. Ecol. 92:fiw177. doi: 10.1093/femsec/fiw177
Urbanová, M., Šnajdr, J., and Baldrian, P. (2015). Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biology and Biochemistry 8453–64. doi: 10.1016/j.soilbio.2015.02.011
Větrovský, T., Baldrian, P., and Morais, D. (2018). SEED 2: a user-friendly platform for amplicon high-throughput sequencing data analyses. Bioinformatics 34, 2292–2294. doi: 10.1093/bioinformatics/bty071
Vos, M., Wolf, A. B., Jennings, S. J., and Kowalchuk, G. A. (2013). Micro-scale determinants of bacterial diversity in soil. FEMS Microbiol. Rev. 37, 936–954. doi: 10.1111/1574-6976.12023
Wang, C., Ma, L., Zuo, X., Ye, X., Wang, R., Huang, Z., et al. (2022a). Plant diversity has stronger linkage with soil fungal diversity than with bacterial diversity across grasslands of northern China. Glob. Ecol. Biogeogr. 31, 886–900. doi: 10.1111/GEB.13462
Wang, J., Wang, Y., Qu, M., and Li, J. (2022b). Dispersal limitation dominates the community assembly of abundant and rare fungi in dryland montane forests. Front. Microbiol. 13:929772. doi: 10.3389/FMICB.2022.929772/BIBTEX
Yang, T., Adams, J. M., Shi, Y., He, J. S., Jing, X., Chen, L., et al. (2017). Soil fungal diversity in natural grasslands of the Tibetan plateau: associations with plant diversity and productivity. New Phytol. 215, 756–765. doi: 10.1111/nph.14606
Yang, F., Wu, J., Zhang, D., Chen, Q., Zhang, Q., and Cheng, X. (2018). Soil bacterial community composition and diversity in relation to edaphic properties and plant traits in grasslands of southern China. Appl. Soil Ecol. 128, 43–53. doi: 10.1016/j.apsoil.2018.04.001
Zbíral, J. (2002). Analýza půd I. Laboratorní odbor: Ústřední kontrolní a zkušební Ústav zemědělský.
Zhang, G., Wei, G., Wei, F., Chen, Z., He, M., Jiao, S., et al. (2021). Dispersal limitation plays stronger role in the community assembly of fungi relative to bacteria in rhizosphere across the arable area of medicinal plant. Front. Microbiol. 12:713523. doi: 10.3389/FMICB.2021.713523/BIBTEX
Keywords: soil bacterial community, soil fungal community, primary succession, source habitat proximity, temperate grassland
Citation: Mészárošová L, Kuťáková E, Kohout P, Münzbergová Z and Baldrian P (2024) Closing the gap: examining the impact of source habitat proximity on plant and soil microbial communities in post-mining spoil heap succession. Front. Microbiol. 15:1416515. doi: 10.3389/fmicb.2024.1416515
Edited by:
Xin Sui, Heilongjiang University, ChinaReviewed by:
Mia Rose Maltz, University of Connecticut, United StatesCopyright © 2024 Mészárošová, Kuťáková, Kohout, Münzbergová and Baldrian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lenka Mészárošová, aGFyYW50b3ZhLmVsQGdtYWlsLmNvbQ==
†ORCID: Lenka Mészárošová, https://orcid.org/0000-0002-8672-8611
Eliška Kuťáková, https://orcid.org/0000-0003-1442-2848
Petr Kohout, https://orcid.org/0000-0002-3985-2310
Zuzana Münzbergová, https://orcid.org/0000-0002-4026-6220
Petr Baldrian, https://orcid.org/0000-0002-8983-2721
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.