
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 12 April 2024
Sec. Terrestrial Microbiology
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1379879
This article is part of the Research TopicInsights in Terrestrial Microbiology: 2023/2024View all 10 articles
 Motunrayo Y. Jibola-Shittu1
Motunrayo Y. Jibola-Shittu1 Zhiang Heng1Nemat O. Keyhani2Yuxiao Dang1Ruiya Chen1
Zhiang Heng1Nemat O. Keyhani2Yuxiao Dang1Ruiya Chen1 Sen Liu1Yongsheng Lin1Pengyu Lai1Jinhui Chen1Chenjie Yang1Weibin Zhang1Huajun Lv1Ziyi Wu1
Sen Liu1Yongsheng Lin1Pengyu Lai1Jinhui Chen1Chenjie Yang1Weibin Zhang1Huajun Lv1Ziyi Wu1 Shuaishuai Huang3
Shuaishuai Huang3 Pengxi Cao3Lin Tian4Zhenxing Qiu5
Pengxi Cao3Lin Tian4Zhenxing Qiu5 Xiaoyan Zhang1*Xiayu Guan6*
Xiaoyan Zhang1*Xiayu Guan6* Junzhi Qiu1*
Junzhi Qiu1*Leaves of Camellia sinensis plants are used to produce tea, one of the most consumed beverages worldwide, containing a wide variety of bioactive compounds that help to promote human health. Tea cultivation is economically important, and its sustainable production can have significant consequences in providing agricultural opportunities and lowering extreme poverty. Soil parameters are well known to affect the quality of the resultant leaves and consequently, the understanding of the diversity and functions of soil microorganisms in tea gardens will provide insight to harnessing soil microbial communities to improve tea yield and quality. Current analyses indicate that tea garden soils possess a rich composition of diverse microorganisms (bacteria and fungi) of which the bacterial Proteobacteria, Actinobacteria, Acidobacteria, Firmicutes and Chloroflexi and fungal Ascomycota, Basidiomycota, Glomeromycota are the prominent groups. When optimized, these microbes’ function in keeping garden soil ecosystems balanced by acting on nutrient cycling processes, biofertilizers, biocontrol of pests and pathogens, and bioremediation of persistent organic chemicals. Here, we summarize research on the activities of (tea garden) soil microorganisms as biofertilizers, biological control agents and as bioremediators to improve soil health and consequently, tea yield and quality, focusing mainly on bacterial and fungal members. Recent advances in molecular techniques that characterize the diverse microorganisms in tea gardens are examined. In terms of viruses there is a paucity of information regarding any beneficial functions of soil viruses in tea gardens, although in some instances insect pathogenic viruses have been used to control tea pests. The potential of soil microorganisms is reported here, as well as recent techniques used to study microbial diversity and their genetic manipulation, aimed at improving the yield and quality of tea plants for sustainable production.
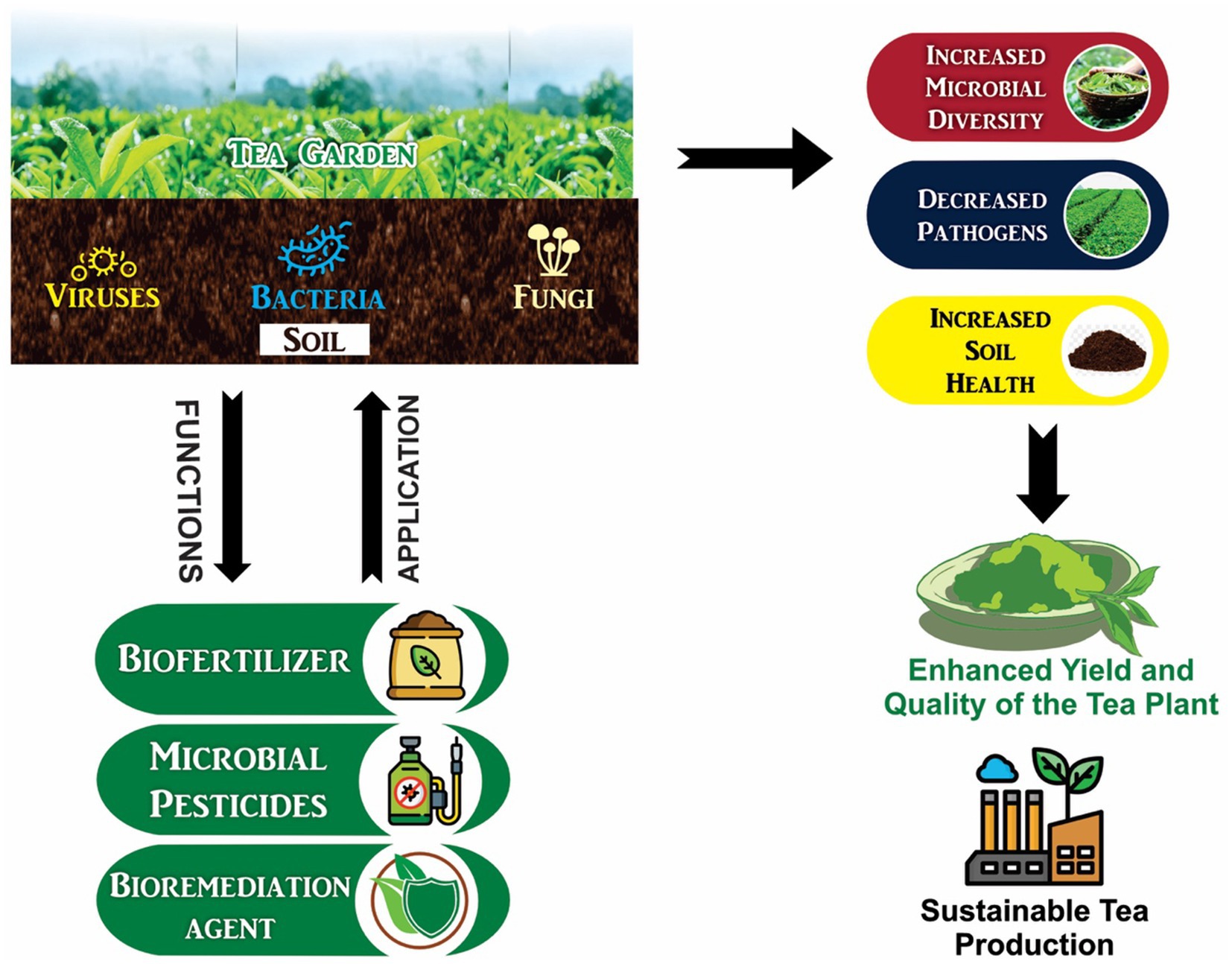
Graphical Abstract.
Camellia sinensis commonly referred to as the “tea plant” is an economically important crop, belonging to the family Thaeceae (Pandey et al., 2021). As a brewed beverage, it has been consumed for at least several millennia, with initial indications that leaves were first eaten raw or added to soups followed by fermentation and chewing of the leaves. Subsequently, it was turned into a beverage by mixing fresh or cured leaves with hot or boiling water, with early written description of tea drinking dating to at least the 3rd century A.D. in China. Modern tea consumption is second only to water, and tea production is dominated by India/Sri Lanka and China, with the latter accounting for about half of the tea produced in our world today (Xu et al., 2018, 2022). Tea is cultivated predominantly in tropical and subtropical regions of the world (Hazra et al., 2019), and global tea consumption is estimated to have increased ~43% from 2005 to 2020 (FAOSTAT, 2022). This is likely due in part to the many putative beneficial health and medicinal values of tea due to its wide range of bioactive constituents. One well known component of tea is caffeine, which can act as a stimulant increasing alertness, with levels of caffeine affected by leaf harvest time and various forms of post-harvest processing. In addition, tea contains a range of secondary metabolites, some of which may possess antioxidant, digestive, (putative) antimicrobial, and/or other health promoting benefits to the consumers (Pokharel et al., 2023; Ramphinwa et al., 2023). Aside from caffeine, potential bioactive compounds include polyphenols (e.g., flavonoids and catechins) as well as xanthines such as theobromine and theophylline. Tea consumption has been linked cancer-prevention, treatment of various cardiovascular problems, and improved circulation potentially due to the variety of polyphenols, antioxidants, and other compounds they possess, although definitive clinical data is lacking (Perez-Burillo et al., 2019; Abe and Inoue, 2021; Bag et al., 2022). Tea consumption also provides important comfort and social interactions in many societies worldwide. Nevertheless, the demand for tea is projected to see a continuous increase (Xi et al., 2023). Therefore, to effectively cater for the rising demand for tea, research should be aimed at the development of ecologically friendly and sustainable approaches to improving the quality and yield of tea.
Camellia sinensis is native to East Asia, with a purported origin along the Irrawady River, spreading into present day southeast China, India, and later to Sri Lanka. Two main cultivated varieties of traditional tea have thus far been described, C. sinensis var. sinensis and C. sinensis var. assamica, that are separated into distinct clades and likely have different parentages. However, C. sinensis var. assamica can be separated into two subtypes, namely, Southern Yunnan Assam (China) and Indian Assam (India), which although may have originated from the same parent, appears to represent two independent domestication events. Furthermore, some subvarieties appear to have undergone hybridization with closely related species such as C. taliensis and C. pubicosta (Mukhopadhyay and Mondal, 2017; Auria et al., 2022). In addition to the occurrence of different regional varieties, tea is further classified post-harvest via the different means and methods for processing of the leaves that result in significant differences in the final consumed product (Auria et al., 2022). These different post-harvest processing methods result in the commonly referred to black, green, white, oolong, dark, and yellow teas (among others), and involve a series of steps that can include, depending upon the final outcome, the following: (picking), (i) withering—drying of the leaves; under sun for darker teas, in a cool ventilated room for lighter teas, (ii) bruising—crushing, shaking, rolling, and/or other forms of manipulating the leaves; mainly for darker teas, (iii) oxidation—exposure leaves to air for different period of time; darkens teas depending upon time, (iv) heating—after oxidation, leaves are heated to stop oxidation process, also referred to as “fixation,” (v) yellowing—light heating in closed chamber, (vi) fermentation—leaves are allowed to ferment for a period of time; results in increase in sweetness, (vii) drying—remove moisture via baking, sun and/or air-drying, (viii) sorting and shaping—stems, seeds, and impurities are removed and the tea “shaped” into various forms, e.g., bricks, circles, etc., for aging and/or storage. Each “type” of tea has its own sequence of specific steps as outlined above, but not all. Thus, “black” tea involves withering, bruising, oxidation, shaping, and drying, “white” tea (freshly picked leaf buds): only withering, heating, shaping, and drying (with white tea often considered the least “processed” of the final tea forms). In addition, many of these steps can have important production differences with respect to leaf treatment for any given step in terms of time, temperature, and other conditions that can result in significant differences in the final products even if all are considered “black” teas (Auria et al., 2022; Aaqil et al., 2023).
Although post-harvest processing is relatively well-described, the soil support used to produce tea has been less studied despite anecdotal and regional recognition that variations in soil “quality” affects plant growth and subsequent leaf quality and production. It is well known that soil microorganisms (bacteria, fungi, and viruses, e.g., the soil microbiome) functions in mediating soil health, and subsequent plant growth and crop yield (Wang et al., 2017; Gu et al., 2019). These effects can be positive or negative with respect to plant health, with beneficial microbes helping to: (i) mobilize otherwise (plant) recalcitrant nutrients, particularly nitrogen and phosphorus, to the plant (often in exchange for carbon), (ii) facilitate plant resistance to abiotic stress including temperature and drought, and (iii) protect plants from infection and disease. In contrast, harmful microbes, e.g., biotrophic and necrotrophic plant pathogens can cause disease, and competition with some microbes may decrease overall plant access to nutrients. Overall, however, the diversity of soil microbial communities can serve as an indicator of soil fertility and soil health (Gui et al., 2022), and poor soil (in terms of mediating plat health) typically showing a reduction in soil microbial community diversity, which can then result in adverse effects on the sustainable utilization of soil resources (Chen et al., 2015). Therefore, maintaining the diversity of soil microbial communities, with an emphasis on identifying and enriching for beneficial microbes, can exert a significant impact on managing soil organic carbon and nutrient availability to plants thus increasing the sustainability of agricultural ecosystems (Bertola et al., 2021; Chauhan et al., 2023), particularly given that several studies have shown that soil microorganisms can have important positive effects on plant growth, plant health, resistance to abiotic stress, and overall agricultural productivity (Trivedi et al., 2020; Qiao et al., 2024). Due to the economic importance of tea plants, it is valuable to build models integrating the nature of soil microorganisms and the vital functions they perform with respect to tea cultivation. Here, we review current information concerning the diversity and potential functions of soil microbial communities in tea gardens, to provide insights into less reported factors that could be explored to improve tea cultivation by examining and potentially manipulating the diversity of soil microorganisms. We also highlight the various techniques used for studying soil microbial diversity within tea gardens. The identification of the diverse groups of soil microorganisms as well as their potential functions will help in meeting the growing demand for the sustainable production of tea plants with high quality and yield.
Soil microbial communities are an essential part of the soil ecosystem, consisting of diverse fungi, bacteria, and viruses (Naylor et al., 2022). Generally, soil microorganisms are involved in key processes in the soil ecosystem; they mediate organic matter decomposition, nutrient cycling, and gaseous fluxes, and impact soil geochemistry including pH, trace metal and other element content, and phosphorus availability, all of which have resultant effects on plant nutrient availability and resistance to stress (Bastida et al., 2021; Hartmann and Six, 2022). Although carbon is gained via photosynthesis, other primary nutrients such as nitrogen, phosphorus, sulphur and potassium, required for plant growth and development, are made available for plant uptake through cycling and transformation processes in the soil. These processes are actively mediated by soil microorganisms, and the availability of these nutrients for plant uptake is a determinant of soil fertility (Basu et al., 2021; Nabi, 2023). Highly fertile soils often exhibit increased bacterial diversity, predominantly those belonging to the Proteobacteria, Nitrospira, Chloroflexi, and Bacteroidetes, in addition to demonstrating enhanced functions such as nitrate reduction, ammonia oxidation and aromatic compound degradation in contrast with low fertile soils (Da Costa et al., 2024). For many crops, bacteria of prominent groups involved in nitrogen-cycling processes and maintaining soil nitrogen balance are indicative of soil fertility, and correlate with crop yield (Bayer et al., 2020; Hayatsu et al., 2021).
In addition, the soil microbiome can participate in the bioremediation of pollutants, heavy metals, and other compounds that can adversely affect plant health or could otherwise affect the quality (and human health safety) of the tea leaves (Bastida et al., 2021; Phillippot et al., 2023). Nitrogen which is a key element involved in the growth and quality tea plants leaves (Ma et al., 2021), is usually recycled by nitrogen-metabolizing microorganisms present in the rhizosphere, providing for enhanced nitrogen absorption of plant usable forms (NO−3 and NH+4) (Liu et al., 2017, 2022) that can then impact the growth and yield of tea. In tea root systems, ammonia is converted into theanine, a non-protein amino acid that adds a distinct rich flavor to tea as the root absorbs such nitrogen sources from the soil (Cheng et al., 2017; Dong et al., 2019). Theanine is then transported to the leaves and young shoots which are harvested during tea production (Zhang et al., 2023). Within this context, a consortium of nitrogen-metabolizing soil microorganisms predominantly belonging to the Proteobacteria and other phyla such as the Actinobacteria, Firmicutes, Chloroflexi, and Armatimonadetes have been reported in tea roots to help enhance ammonia uptake and subsequent theanine synthesis, thus contributing to the taste and quality of tea leaves (Xin et al., 2024).
Tea cultivation is unique in that an important number of tea “gardens” or areas of tea cultivation, have existed, i.e., been continuously cultivated with C. sinensis, for significant periods of times (generations or even more in some instances). With such relatively continuous cultivation in specific areas, it is likely strong co-interactions between the tea plants and resident microorganisms in the soil have developed, including potentially unique co-adaptations. However, many areas of tea cultivation have also had significant inputs (fertilizer, pesticides, even soil) from other areas that can impact the diversity of both beneficial and harmful (to the plant) microbes (Fu et al., 2021). In addition, tea gardens are faced with a variety of challenges from insect pests and microbial diseases, many of which are vectored by insects (Zhang X. et al., 2022). As suggested, to achieve high yield, significant amounts of fertilizers and pesticides are used in some tea gardens which can, after long term use, cause a decline in soil microbial diversity and hence results in a negative environmental impact (Wang et al., 2020). As tea gardens are usually found on elevated plains, application of chemical fertilizers and pesticides can easily run-off into downstream water bodies, causing eutrophication and pollution of the water (Xie et al., 2021).
Several studies on tea garden soils have revealed the presence of a vast array of microorganisms which are linked to the quality of tea produced from the soil (Fu et al., 2021; Kui et al., 2021; Bag et al., 2022), particularly as soil microbial communities participate in promoting soil health and suppressing plant pathogens (Wu et al., 2023). Based on culture-dependent approach and molecular identification of bacterial isolates through 16S rRNA gene sequencing, keystone bacteria genera such as Bacillus, Burkholderia, Serratia and Arthrobacter (Table 1) have been reported to display a wide range of growth promoting activities. For example, the phosphorus solubilizing abilities in Burkholderia and Bacillus isolated from rhizospheric soil samples of tea gardens in West Bengal, India have been characterized (Panda et al., 2017), and isolation of a Burkholderia pyrrocinia strain from the tea rhizosphere contributing to abiotic stress tolerance and possessing plant growth promoting activities through phosphorus solubilization and production of phytohormones has also been reported (Han et al., 2021). In addition, a biocontrol strain of Serratia marcescens was reportedly isolated from tea rhizosphere displayed significant biocontrol efficacy against fungal root pathogens of tea through the production of hydrolytic enzymes such as chitinase, protease, lipase and cellulase, as well as the production of antibiotics (Dhar Purkayastha et al., 2018). These studies indicate that knowledge concerning the microbial ecosystem of tea soils and the processes by which they help improve tea quality, should be considered an important avenue for further exploration aimed toward enhancing nutrient availability in tea gardens and the resultant yield and quality of tea plants.
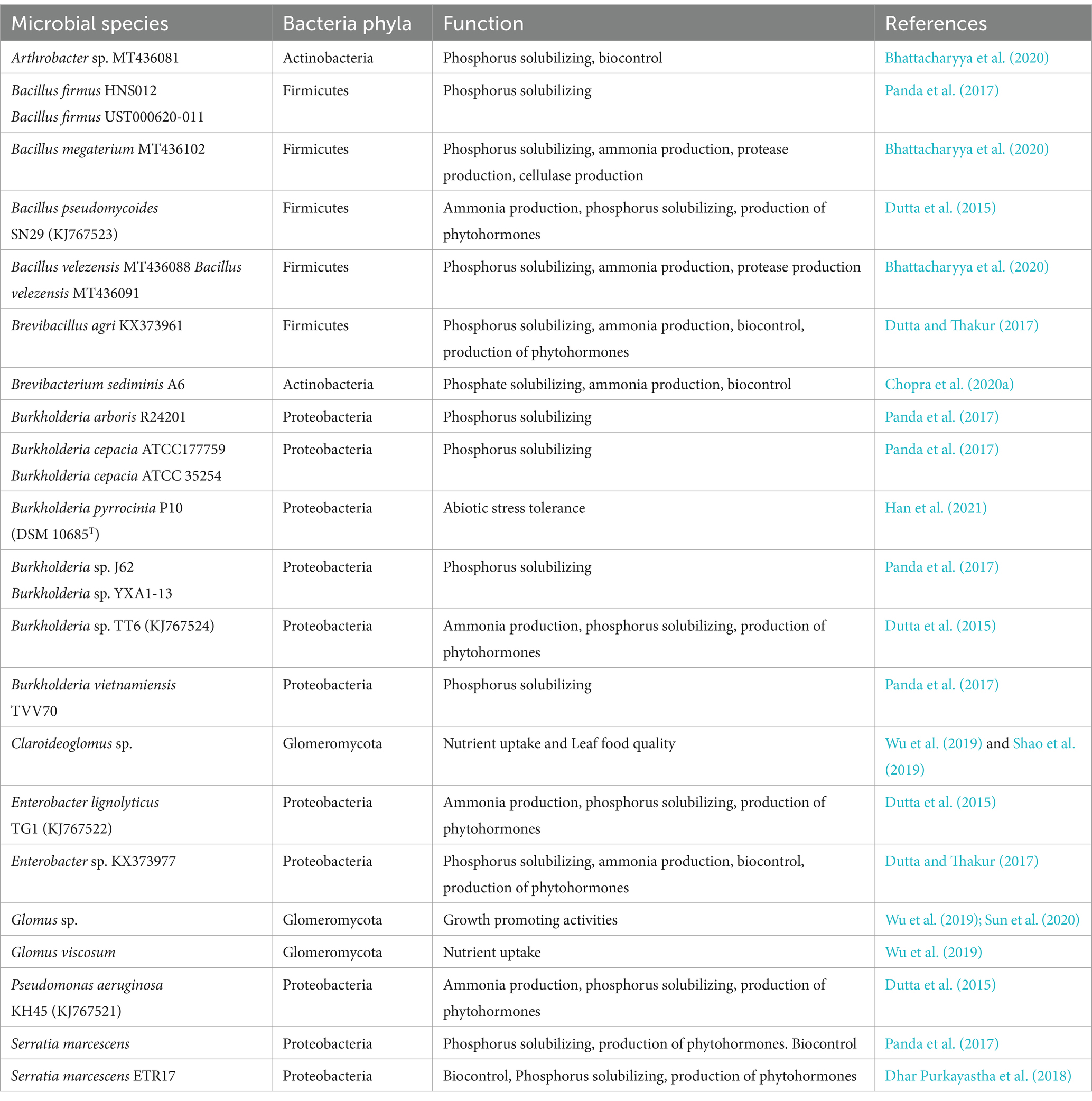
Table 1. Some key microbial species associated with soil ecosystem of tea gardens.
Exploring soil microbial diversity is beneficial for the development of agricultural ecosystems as well as testing the effectiveness of restoration measures (Deltedesco et al., 2020). For tea gardens such studies can help in understanding the functions of microorganisms and in harnessing them for better productivity of tea. Recently, the development of culture-independent metagenomics techniques, has contributed greatly to mapping soil microbial phylogeny (Su et al., 2017). The development and refinement of molecular techniques such as the high-throughput sequencing have greatly promoted the study and understanding of diversity and interactions of soil microorganisms (Wei et al., 2018; Song et al., 2023) and has given rise to a significant improvement in terms of both rDNA homology and descriptions of biosynthetic pathways (Wei et al., 2018). Consequently, metagenomic analyses display strong reliability and convenience for characterizing root-associated microorganisms (Bhattacharyya et al., 2016; Busby et al., 2017), and is being applied to characterize soil microbial diversity by directly capturing total soil microbial DNA (Sharma and Kaur, 2021; Parihar et al., 2022) providing a window into functional aspects of soil microorganisms (Wei et al., 2018).
Next,-generation sequencing technologies have also led to insights into various environmental factors contributing to soil microbial diversity (Egidi et al., 2019). As part of this, the majority of microbial species assigned to “RNA similarity groups” can help provide a deeper understanding of the changes in diversity and composition of soil microbial communities (Chen et al., 2017). Similarly, high through-put sequencing technologies have enabled microbiologists to sequence amplified gene markers (e.g., 16S ribosomal RNA), to determine phylogenetic and functional diversity profiles of soil microbial communities (Wu et al., 2015). High-throughput sequencing and molecular ecology network (MEN) analyses have been used to investigate soil microbial diversity, community structure, composition, and interaction networks of tea plantations, revealing the diversity, and dominance of Proteobacteria, Acidobacteria, and Chloroflexi in all tea plantation samples under different management practices (Tan et al., 2019). Although still limited, targeted gene manipulation, e.g., use of CRISPR/Cas technologies has been applied to alter the expression of genes, study genetic diversity, and or produce modified microorganisms, and/or transfer of genes have been applied to tea cultivation research (Bag et al., 2022).
Fungi are important drivers in soil ecosystems (Francioli et al., 2020); a rich fungal diversity in tea garden soils may help maintain healthy ecological functioning, including by facilitating nutrient cycling, organic matter decomposition, and plant productivity (Ma et al., 2022). In addition, (beneficial) fungi can play important roles in suppressing the activities of (microbial) plant pathogens present in the soil (Bollmann-Giolai et al., 2022). This latter function can be due to a variety of factors including excluding plant pathogen competitors to the production of certain metabolites targeting pathogenic microbes to stimulating plant antimicrobial defenses, thus suppressing tea pathogens, and enhancing tea yield and quality. Xu et al. (2022) have reported that fungal taxa that colonized tea shoots significantly inhibited fungal pathogens. These included fungal taxa corresponding to Myriangium and Mortierella which have been demonstrated to have plant growth-promoting abilities (Ozimek and Hanaka, 2021). Thus, these abilities may be linked to their ubiquity and potentials to protect plants against pathogens. Zheng et al. (2023) investigated the response of soil microbial communities and functions to long-term tea (C. sinensis) planting in a subtropical region and reported the relative abundance of fungal community in tea gardens to be largely dominated by Ascomycota (38.63–55.27%), Basidiomycota (19.45–39.13%), Mortierellomycota (1.8–10.1%), and Rozellomycota (0.12–7.41%). Similarly, Ma et al. (2022) in a study conducted on soils of tea plantations revealed that fungal community predominantly consisted of Ascomycota (44.7%), Mortierellomycota (17.7%) and Basidiomycota (11.4%) and accounted for 73.8% of total composition of fungal communities.
Earlier reports indicated that fungal communities in several tea gardens at the genus level are dominated primarily by Saitozyma (Ma et al., 2022; Wang et al., 2023). Members of the Saitozyma, have been shown to account for ~30% of the sequences in tea garden soils in the Southeast Asia region, followed by Mortierella (20%) and Pseudogymnoascus (10%) (Yan P. et al., 2022). Basidiomycota, such as Saitozyma, Russula and Hygrocybe commonly found in tea gardens, are well known to colonize lignin-rich surfaces and likely play significant roles in the degradation of lignin-rich plant litters (Guo et al., 2018; Kui et al., 2021), transforming these substrates to provide carbon and nitrogen as well as other nutrients for plant growth (Li et al., 2020). In addition, Penicillium, Trichoderma and Pseudogymnoascus are prominent members of the Ascomycota predominantly found in tea gardens (Kui et al., 2021; Wang et al., 2021; Yan L. et al., 2022). The abundance of the Ascomycota among soil fungal communities in tea gardens is perhaps because Ascomycota have been able to successfully evolve mechanisms to dominate soils globally (Egidi et al., 2019). These abilities of the Ascomycota include stress tolerance and production of secondary metabolites which can inhibit other microorganisms (Chen et al., 2017). Moreover, Ascomycota have been known to produce a wide range of antimicrobial agents, which can be advantageous to the protection of plants against pathogens.
The rhizosphere of tea gardens in Southeast Asia have been shown to contain a rich community of Glomeromycota, that include arbuscular mycorrhizal fungi (AMF), such as Claroideoglomus, Acaulospora, Rhizophagus and Glomus species. These fungi often colonize and form symbiotic relationships with the roots of tea plants (Bag et al., 2022; Zhang X. et al., 2022). This relationship with tea plant roots likely contributes to the ability of tea plants to thrive successfully for many years even under adverse environmental conditions that can include drought, salinity, and temperature. In particular, various AMF are known to provide host plants with essential mineral elements, confer resistance to pests, diseases and abiotic stress and promote plant health (Almario et al., 2022; Zhang X. et al., 2022). AMF members of the Glomus, Acaulospora and Gigaspora genera have all been reported in cultivated tea lands, e.g., in India and various locations of China (Sharma et al., 2013; Ji et al., 2022; Zhang Z. et al., 2022).
Bacterial communities are diverse and perform numerous functions in soils. Kui et al. (2021), through the direct extraction of total soil DNA from soil samples and sequencing using high through-put 16S rRNA and internal transcribed spacer amplicon sequencing techniques, characterized the soil microbiome in ancient tea plantations in Southwest region, China, identifiying Acidobacteria, Actinobacteria, and Proteobacteria phyla as the dominant bacterial community. As tea farming often occurs in one place over many generations and sometimes hundreds of years the dominance of Acidobacteria may indicate their importance in key ecological processes such as regulation of biogeochemical cycles and growth promoting activities (Kalam et al., 2020) in tea gardens. The relative abundance of Acidobacteria in tea garden soils is linked to increased age of tea plants (Wang et al., 2019). The long-term use of pesticides and fertilizers which contributes to the acidification of tea soil enables the Acidobacteria to thrive through many mechanisms they have developed. These mechanisms are genetically controlled and include acid tolerance, secondary metabolites, nitrogen metabolism, exopolysaccharide synthesis, hopanoids synthesis, siderophore synthesis (Kalam et al., 2020; Yadav et al., 2021).
Furthermore, using Illumina Miseq sequencing of the 16S rRNA targeting rhizospheric soil bacteria, Zi et al. (2020) found the bacterial community to be dominated by Proteobacteria, Acidobacteria and Actinobacteria with the relative abundance of 43.12, 21.61, and 14.84%, respectively, in Southwest tea cultivation region of China. The dominance of Proteobacteria in soils of tea gardens may be linked to their functioning in carbon and nitrogen cycling because these bacteria are known to be involved in ammonia oxidation and nitrification (Zhang Z. et al., 2022; Wang et al., 2023). Besides, the use of nitrogen-based fertilizers to increase yield in tea gardens could be responsible for the abundance of Proteobacteria in soil because they are actively involved in nitrogen conversions in soil. Likewise, a recent study conducted by Zheng et al. (2023) through the direct extraction of soil DNA and high-throughput sequencing to investigate soil microbial communities structure in tea plantations in the Southeast region of China, reported the relative abundance of bacterial phyla corresponding to Proteobacteria (20.96–41.40%), Acidobacteria (9.41–28.42%), Firmicutes (6.39–16.03%), Bacteriodetes (6.05–13.80%), Chloroflexi (3.35–13.27%) and Actinobacteria (2.37–11.52%) being dominant phyla. Lynn et al. (2017), through the direct extraction of soil microbial DNA and 16S rRNA sequencing also demonstrated that Actinobacteria, Chloroflexi, Acidobacteria, Proteobacteria, Firmicutes dominated the diverse bacterial communities in tea plantations found in Southern region of China. Actinobacteria have evolved mechanisms such as production of secondary metabolites, production of phytohormones, production of antimicrobials and stress tolerance, enabling them to thrive successfully in various soil ecosystem including adverse conditions, this may explain their high level of occurrence. The presence of Actinobacteria in tea gardens may also be helpful for the growth and successful yield of tea plants over the years. Actinobacteria have been reported to produce enzymes and secondary metabolites including a range of antibiotics some of which have been successfully exploited commercially and industrially (Barka et al., 2016; Jose et al., 2021). They are also known to act as biopesticides for agricultural benefit and play important roles in bioremediation of chemical pesticides, heavy metals, and other toxins (Alvarez et al., 2017; Banik et al., 2019). The rich abundance of Actinobacteria in soils of tea gardens likely helps account for the ability of tea plants to resist a wide range of phytopathogens, i.e, through their production of antimicrobial metabolites (Shan et al., 2018).
In addition, bacteria belonging to diverse genera known as the “mycorrhizal helper bacteria” (MHB) exist in soils. Frequently, these bacteria have a tripartite association with arbuscular mycorrhizal fungi and tea roots (Bidondo et al., 2016; Gupta and Chakraborty, 2020). Although their activities in tea gardens are poorly understood, these bacterial AMF enhancers generally have been reported to promote the functions of arbuscular mycorrhizal fungi leading to a better uptake of nutrients by plants and potentially increasing their ability to survive biotic and abiotic stresses (Sangwan and Prasanna, 2022). These bacteria may also benefit the AMF which would then benefit the plant. Bacteria that function actively as mycorrhizal fungal enhancers are found in (1) the Proteobacteria, specifically within the bacteria genera: Pseudomonas, Agrobacterium, Azospirillum, Azotobacter, Burkholderia, Bradyrhizobium, Enterobacter, Klebsiella and Rhizobium species, (2) the Actinobacteria including Rhodococcus, Streptomyces and Arthrobacter sp., and (3) the Firmicutes that include Bacillus, Brevibacillus and Paenibacillus sp. (Martin, 2016; Nasslahsen et al., 2022). These mycorrhizal helper bacteria may also perform several other functions not limited to enhancing AMF, such as plant growth promoting activities through the production of phytohormones (Sangwan and Prasanna, 2022), and they are widespread in tea garden soils.
Viruses are likely the most abundant and diverse organisms on earth, many of whom affect soil microbial communities and their functions (Berliner et al., 2018; Jansson and Wu, 2022). Examination of soil viruses remains understudied; however, it is known that viruses can regulate soil microbial communities (Chevallereau et al., 2021; Liao et al., 2022) and contribute significantly to soil ecological processes such as nutrient cycling (Bi et al., 2022). In particular, there is paucity of reports on specific activities of soil viruses in tea gardens, viruses can be very important as they infect other microbial communities such as the bacteria and fungi, hence shaping microbial composition, metabolism and probably influence major soil activities (Roux and Emerson, 2022). Since viruses are host specific, viruses that infect pathogenic bacteria, fungi, and insects, have been isolated and used as biocontrol agents targeting their respective hosts (Kizheva et al., 2021). In terms of insect pests, two viruses: Ectropis obliqua single-nucleocapsid nucleopolyhedrovirus (EcobSNPV) and Ectropis obliqua picorna-like viruses (EoPV) have been commercially used with high efficacy against E. obiqua which is a common pest of tea plants (Idris et al., 2020). However, effective viruses for other tea pests, e.g., Helopeltis theivora and Gyropsylla spegazziniana, have not yet been commercialized, and overall, the specific contributions of soil viruses to (tea) soil fertility and plant health remains unknown.
Depending upon the member community, soil microorganisms in tea garden could have beneficial or detrimental effects on tea plants (Figure 1). Beneficial microorganisms, such as members of the Bacillus, Rhizobium, Actinomycetes, Trichoderma, and Glomus promote soil health and enhance plant productivity through improving soil structure and promoting organic matter recycling (Hicks et al., 2021; Wei et al., 2024), e.g., functioning as decomposers of leaf litters and dead plant materials in tea gardens (Schroeter et al., 2022; Wang et al., 2023).

Figure 1. Composition and functions of soil microbial communities in tea garden.
Microbial communities can also help plant disease resistance, decrease soil load of (plant) pathogens, and increase (plant) environmental stress tolerances (Zhang X. et al., 2022). For example, beneficial microbes such as Bacillus spp. and Actinomycetes can help tea plants resist a range of fungal diseases, i.e., leaf blight and scab disease (Wang et al., 2021). Arbuscular AMF colonization in the tea rhizosphere, e.g., by Glomus, Rhizophagus, and Acaulospora, likely contributes to enhanced disease resistance in host tea plants. Tea roots colonization by AMF can also help tea plants to survive under adverse conditions, enhance photosynthesis, and increase nutrient (e.g., phosphorus) absorption (Bag et al., 2021; Almario et al., 2022). Conversely, some soil microorganisms, e.g., Fusarium and Pseudopestalotiopsis, can cause disease to tea plants (Arafat et al., 2020; Pandey et al., 2023), and cultivation methods such as long-term applications of chemical fertilizers (e.g., Urea- N and NPK) may result in enrichment of pathogenic fungi (e.g., Fusarium and Pseudopestalotiopsis) in tea gardens (Wang et al., 2023; Zheng et al., 2023).
Nitrosphaeraceae also play important roles in nitrogen cycling (Amoo and Babalola, 2017; Zhang Z. et al., 2022), and the rhizosphere of tea plants are frequently colonized by nitrogen fixing and ammonia oxidizing bacteria (e.g., Azotobacter and Nitrosomonas, respectively) which affect nutrient cycling in the soil and can regulate nutrient utilization in tea plants (Wright and Lehvirta-Morley, 2023). Beneficial bacteria such as Bacillus, Azotobacter and Pseudomonas have also been proven to be of great potential in soil remediation (Xiang et al., 2022). However, there is no information of their specific application in remediation of tea soils.
Viral lysis of microbial cells can release materials which are transformed into dissolved organic matters (Chen X. et al., 2022), thus impacting nutrient cycling processes. In addition, viruses can reprogram host metabolism by expressing virus-contained auxiliary metabolic genes during infection. These auxiliary metabolic genes are sometimes involved in numerous metabolic pathways and could boost host metabolism and supply energy, thereby enhancing viral propagation, consequently impacting biogeochemical cycles (Hurwitz and U’Ren, 2016; Bi et al., 2022). Several studies have demonstrated the presence of auxiliary metabolic genes in soils, particularly agricultural soils (Wang et al., 2016; Han et al., 2017). For example, viral-encoded carbon metabolism was identified in high organic matter peatsoils, demonstrating potential viral roles in carbon cycling processes (Emerson et al., 2018; Trubl et al., 2018). Viruses can also influence microbial/plant/animal evolution as agents of horizontal gene transfer by encoding other functional genes and mediating the transfer of genes between hosts (Trubl et al., 2018). Although assumed for many years that the tea plant was virus-free, Hao et al. (2018) reported two novel viruses belonging to the Blunervirus and Ilarvirus genera from tea plants using metagenomic analysis. These viruses infected the tea plant causing necrotic ring and discoloration of tea leaves, reducing the quality and yield of tea leaves.
Some soil microorganisms’ exhibit positive plant growth promoting (PGP) traits that impact the productivity of the plants that grows on such soils (Bag et al., 2022). Growth promoting traits in soil microorganisms (e.g., Bacillus subtilis, Trichoderma viridae, and Streptomyces griseus) in tea garden soils (Tables 2, 3) have been shown to impact phosphate solubilization, nitrogen fixation, siderophore production, antagonism to the pathogen, and act in the production of plant auxin hormone production, e.g., indole-3-acetic acid (Bhattacharyya and Sarmah, 2018; Kolandasamy et al., 2023), with tea rhizosphere bacteria also found to promote the growth of rice and maize seedlings (Bhattacharyya et al., 2020). A significant positive relationship between Nitrososphaeraceae in tea garden soils with ammonia oxidation and nitrification processes has been reported, suggesting the importance of these bacteria in sustaining nitrogen fixation (Zhang Z. et al., 2022). These results suggest that exploitation of identified beneficial tea rhizosphere microorganisms has the potential to be used as microbial-based fertilizers. A two-year field experiment comparing the effects of bio-organic fertilizers (Bacillus megaterium-based bio-organic fertilizer, Bacillus colloid-based bio-organic fertilizer and Bacillus subtilis-based bio-organic fertilizer) and conventional chemical fertilizers, reported that the microbial-based fertilizers increased significantly the contents of tea polyphenols, amino acids and caffeine compared with the conventional chemical fertilizer (Liu et al., 2023), perhaps, through nitrogen metabolism and nutrient solubilization processes which enhanced nutrient availability and uptake by tea roots. Similarly, a study conducted by Xin et al. (2024) reported that the inoculation of soil with a synthetic community (SynCom21) of 21 bacterial strains belonging to the phyla Proteobacteria and Actinobacteria isolated from the rhizosphere of highly productive tea plants, was able to enhance ammonia uptake and transport in tea plants, facilitate the synthesis of theanine and increase theanine content of the tea leaves in comparison with the controls. Thus, application of tea rhizosphere bacteria as microbial-based fertilizer can promote nutrient availability and absorption resulting in enhanced tea polyphenols and theanine content, increasing the quality of tea leaves.

Table 2. Regulation of the functions of major soil bacterial communities in tea gardens.
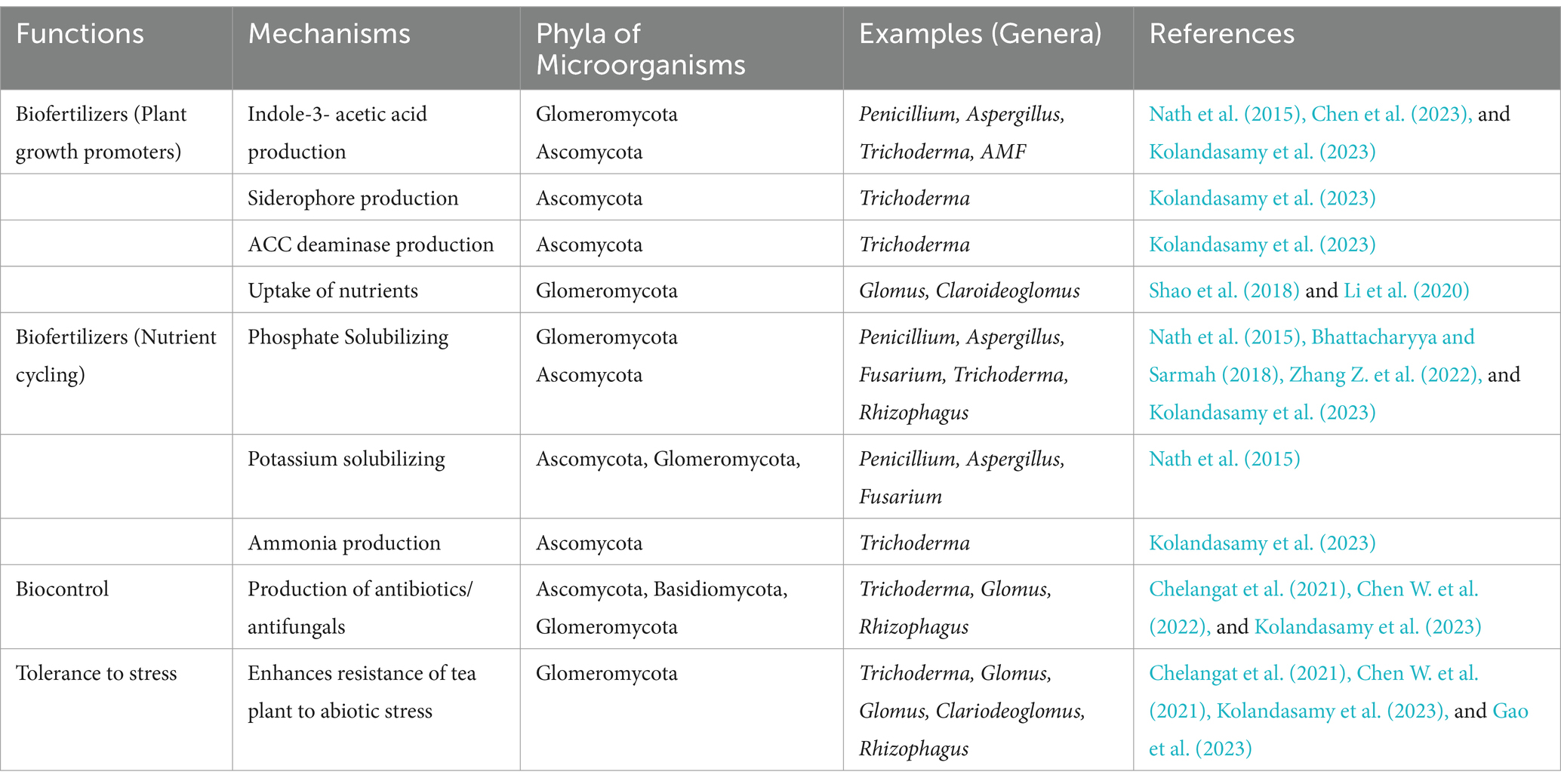
Table 3. Mechanisms of the functions of major soil fungal communities in tea gardens.
Soil microbe degradation of soil pollutants is another key ecological function that entails regulated gene expression and the activities of multiple enzymes (Wang et al., 2023). A variety of chemical pesticides, especially organochlorine pesticides (OCPs) such as Dichloro-diphenyl-trichloroethanes (DDT), Endosulfan and Dicofol to target tea scale insect, mites and tea mosquito bug are routinely used in tea plantations, resulting in residues on the tea plants themselves as well as in the soil (Lu et al., 2015; Fernandes et al., 2023). These chemicals can decrease the quality of both the tea and the soil, with such persistent organic pollutants (POPs), accumulating due to their low natural degradation rates (Negrete-Bolagay et al., 2021), potentially carcinogenic (Fernandes et al., 2023). However, rhizosphere microorganisms, through the action of degradative enzymes, have been reported to be able to degrade such persistent organic pollutants (Bishnu et al., 2012; Shi et al., 2015) in the soil of tea plants. The specific microorganisms involved in the degradation of organic pollutants in tea gardens have not been reported, but tea plant root secretions such as catechin, glucose, arginine and oxalic acid, have been reported to significantly influence the degradative abilities of soil microorganisms against persistent organic pollutants (POPs) by tea plant rhizosphere microorganisms. This was explained by the reduction in the binding energy of the complex protein to POP molecules in the presence of these root secretions (Du et al., 2022). These root secretions likely also attract and stimulate select microorganisms to produce degradative enzymes such as polyphenol oxidase, hydrolases, catalase and laccase (Wei et al., 2024), which can catalyze the degradation of the POPs. Moreover, root secretions can influence microorganisms present in the rhizosphere by acting as stimulants, signaling molecules or repellants (Olanrewaju et al., 2019; Xin et al., 2024). Various plant growth promoting microorganisms including Bacillus, Pseudomonas, and Trichoderma have also been shown to be involved in the remediation of pollutants and heavy metals in soil (Gond et al., 2021; Ren et al., 2023). The ability of bacteria such Acidobacteria and Chloroflexi to utilize complex organic compounds has been shown to increase with the age of tea planting (Wang et al., 2020). These data indicate that metabolic activities of microorganisms in the soil of tea gardens could potentially remove and degrade harmful substances such as chemical pesticides, heavy metals and organic pollutants in the soil.
Microorganisms also help maintain soil aggregates that are important to soil structure and fertility, root penetration and crop yield, through secretions of extracellular polymeric substances and other compounds including polysaccharides, polyuronic, and amino acids with adhesive properties which can bind soil particles together (Hartmann and Six, 2022). The soil fungal community can promote aggregate stability because of their filamentous growth and their hyphal networks in soil (Morris et al., 2019). Particularly, AMF produce hyphal networks and gromalin, a putative abundantly produced glycoprotein, which aids in soil resistance to erosion, and helps increase carbon storage and water-holding capacity (Rashid et al., 2016). AMF also increase the stability of soil macroaggregates in the soil ecosystem of tea gardens (Morris et al., 2019). A report on the dynamics of soil bacterial community diversity and composition at aggregate scales in tea gardens, revealed that soil aggregates exhibited complex bacterial communities which could provide biological buffering which could prevent individual bacterial species from gaining superiority via competition or predation (Wang et al., 2021). Because stable soils can provide a valuable ecosystem for tea plants to thrive, future studies should explore the potentials of microorganisms, particularly AMF stabilization of soil ecosystem in tea gardens.
Furthermore, some soil microorganisms colonizing the root of tea plants exhibit strong biocontrol activity against plant pathogens and pests (Bag et al., 2022). Plant growth promoting fungi such as Aspergillus, Fusarium, Trichoderma and bacteria such as Azotobacter, Azospirillum, Pseudomonas sp., have been shown to help increase tea plant growth and can help control soil-borne plant pathogens (Thabah and Joshi, 2022), as well as potentially improving tea plant resistance to diseases (Zhang X. et al., 2022). For instance, bacterial Bacillus and fungal Trichoderma strains isolated from the tea rhizosphere have been shown to display high biocontrol efficacy against Phomopsis theae, a fungi pathogen causing stem canker in tea plants (Kolandasamy et al., 2023). Similarly, isolates of B. subtilis has been reported to significantly improve the resistance of tea against several diseases including black rot, branch canker, blister blight and root diseases (Bhattacharyya et al., 2020; Bora and Bora, 2021). Actimomycetes such as Streptomyces, Microbacterium, and Norcardia sp. have been reported to produce secondary metabolites with antimicrobial potentials (Shan et al., 2018) and have been proven to be successful in managing tea diseases (Bhattacharyya et al., 2016). These data indicate that healthy and/or manipulation of tea plantations soils can be useful and effective approach toward helping tea plants resist attack by microbial pathogens.
Tea is a perennial plant that is usually propagated through seedlings developed from seeds by hardening in a nursery through stepwise exposure to full daylight. Tea plant is frequently pruned to enable the development of new shoots and maintain the shape and height and can take up to two years to maturity. The leaves are harvested by plucking new leaves and terminal buds from the tip of the branches at regular intervals from the second year onwards (Mukhopadhyay and Mondal, 2017; Auria et al., 2022). The plucking of the new leaves also enables the emergence of new buds and leaves. Tea soil ecosystem functions are often affected by multiple biotic and abiotic factors. The intensity and duration of tea planting have a significant impact on the microbial community structure, biomass, and its function (Kui et al., 2021), also impacting the soil physicochemical properties.
Soil physicochemical properties like temperature, humidity and pH values influence microbial community diversity in tea garden soils (Muneer et al., 2022). Bacterial and fungal communities during tea planting are strongly affected by changes in soil pH (Zheng et al., 2023) that can occur due to the long-term use of chemical fertilizers. Soil pH in tea gardens can be altered by agricultural practices such as the addition of fertilizers and pesticides. The heavy use of chemical fertilizers can decrease soil pH while the use of organic fertilizers can regulate soil pH. Ye et al. (2022) reported that the long-term use of chemical fertilizer led to a continuous decrease in soil pH from 3.07–2.82 in tea plantations in Southeast China while the long-term use of organic fertilizer led to a stable pH of 5.13–5.33, which is suitable for growth, improved yield, and quality of tea. Persistent decreases in soil pH in addition to decreasing microbial diversity, may result in the denaturing of soil enzymes, and lowered nutrient solubility and availability to plants, as well as increased aluminium and/or heavy metals toxicity as lowered soil pH can increase solubility of certain toxic chemicals, resulting both direct plant toxicity, decreased soil microbial diversity, and equally important in terms of relevance to human consumption, accumulation of toxic metals by the plant (Yan et al., 2021; Naz et al., 2022).
Like many metals, low levels of aluminium, copper, manganese, and others are needed by the plant and promote tea plant growth, however at high concentrations coupled to lowered pH, they can induce toxicity (in the plant and/or to the consumer) as insoluble forms, e.g., for aluminium, dissociate at pH < 5 and releasing (Al3+) ions into the soil, which can form complexes with the other compounds (e.g., phosphates) found in the rhizosphere of tea plants (Ray et al., 2022), which would not only increase metal contents in the plants, but could also reduce the availability of phosphorus to plants. As mentioned, decreases soil pH can lead to the accumulation of metals in the tea plant leaves (de Silva et al., 2016; Peng et al., 2018; Yan et al., 2018), that when cycled, leads to a further decrease in soil pH, successively affecting soil microbial community structure and function, and ultimately impairing healthy plant growth and reducing the quality of the tea leaves. Decreased pH would favor acidophilic bacteria, i.e., those that encode genes regulating acid tolerance and/or prefer acidic conditions for their growth such as Acidobacteria (Kalam et al., 2020), and the abundance of microorganisms such as Acidobacteria and Ascomycota in tea gardens was increased with lower soil pH (Zhang Z. et al., 2022).
Similarly, agricultural management practices (e.g., the use of pesticides, mulch, fertilizers, Figure 2) can have beneficial or detrimental effect on the health of (beneficial) soil microbial communities, leading to increased or decreased tea plants yields, respectively. Fertilizer and pesticide applications can affect the microbial communities of the rhizosphere (Liu et al., 2021), with nitrogen fertilizers improving tea yields but leading to rapid and continuous acidification of tea garden soils (Yan et al., 2020). This acidification can result in loss of important soil microorganisms which is exacerbated by continuous tea cultivation in the same soil, which can lead to erosion of tea quality and yield (Li et al., 2016; Yang et al., 2018; Ye et al., 2022). To combat this, there has been increasing use organic fertilizers/compost as these have been to improve (i.e., help alkalinize) acidified soils, improving soil microbial community health including enzyme activities that improve soil quality (Li et al., 2018; Lin et al., 2019; Xie et al., 2021). Thus, addition of organic fertilizers to tea garden soil can be one method for the remediation of acidified soil (Ye et al., 2022). Within this context, the application of a combination of compost and nitrogen fertilizer has been shown to increase soil microbial diversity, demonstrating the compatibility of this combined approach for promoting soil and subsequent plant health (Taha et al., 2016).
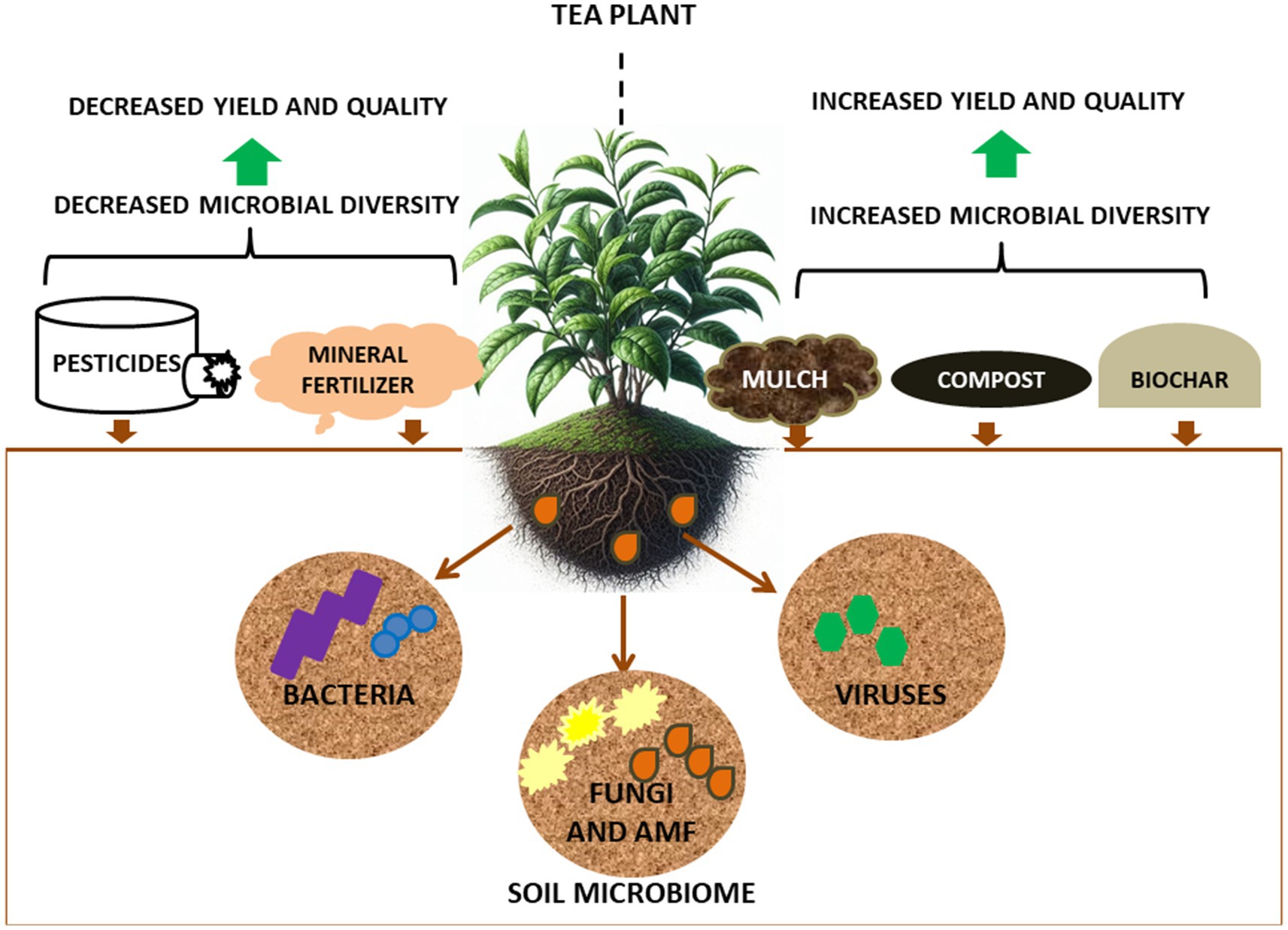
Figure 2. Agricultural management factors influencing microbial diversity in soils of tea gardens.
A study on the effect of organic mulching in tea plantations, reported that bacteria of the phylum Nitrospirae were more numerous in peanut hull mulched soils (3.24%) as compared to polyethylene mulched soils (1.21%) (Zhang et al., 2020). The abundance of Nitrospirae indicates the presence of ammonia-and nitrate-oxidizing bacteria which are important for nitrogen cycling processes (Chen Y. P. et al., 2021). Fungal Mortierellomycota and Basidiomycota were also higher in peanut mulched soils (33.72, 21.93%) as compared to polyethylene mulched soils (14.88, 6.53%) (Zhang et al., 2020), indicating that organic mulching of tea garden soils could have a positive effect on soil microbial communities, helping to improve soil fertility for higher tea plant yields.
More recently, the biochar, which consists of carbon, volatile matter, mineral matter (ash) and moisture, created by thermal burning of biomass has been applied to soils with the aim of improving soils (Armah et al., 2023). This innovation has gained prominence as an effective soil amendment for decreasing plant disease incidence and helping promote beneficial microbial populations in continuous cropping soils (Ge et al., 2023). Biochar application to soil has been reported to increase tea plant productivity and soil nutrient contents (Zou et al., 2023). Bamboo and rice straw biochar has also been shown to significantly improve tea growth, increase tea nutrients and reduced heavy metals in tea (Yan et al., 2021). Although the mechanism of biochar mediated effects on soil microorganisms in tea gardens remains unclear, biochar application has been shown to shape the tea soil fungal community (Zheng et al., 2019), which may be because fungi play important roles in organic matter turnover (Chen et al., 2014). The mechanism of how biochar influences specific soil microbial communities for improved tea yield is an important emerging field for sustainable tea production.
Tea cultivation has considerable economic and medicinal value, and to ensure sustainable tea production, it is necessary to study the role that soil microorganism play, including promoting an increase in the diversity of beneficial soil microorganisms to improve soil health and tea productivity. Targets of future research include:
i. Exploiting molecular techniques, including targeted gene manipulation (e.g., CRISPR/Cas) to enhance the beneficial characteristics of soil microorganisms, including their biofertilizing capabilities. For example, the potential of AMFs in soils (which can contribute to increased nutrient acquisition, stress tolerance and/or disease resistance) of tea plantations could be enhanced through the isolation and application of suitable strains for inoculation. In this context, molecular techniques can be used to directly manipulate tea varieties to achieve these desirable characteristics.
ii. Enhancing the biocontrol activity, especially toward fungal plant pathogens and insect pests, of soil bacteria and fungi in tea gardens to provide an ecologically friendly approaches disease and pest management.
iii. To explore and commercialize the use of plant growth-promoting microorganisms from other crops for tea cultivation and, conversely, the use of beneficial microbes derived from tea garden soils on other economically important crops for sustainable agriculture.
MJ-S: Conceptualization, Methodology, Writing – original draft. ZH: Resources, Writing – original draft. NK: Writing – review & editing. YD: Data curation, Writing – original draft. RC: Data curation, Writing – original draft. SL: Data curation, Writing – original draft. YL: Data curation, Writing – original draft. PL: Data curation, Writing – original draft. JC: Data curation, Writing – original draft. CY: Data curation, Writing – original draft. WZ: Data curation, Writing – original draft. HL: Resources, Writing – original draft. ZW: Resources, Writing – original draft. SH: Resources, Writing – original draft. PC: Resources, Writing – original draft. LT: Writing – original draft. ZQ: Writing – original draft. XZ: Writing – original draft. XG: Writing – original draft. JQ: Conceptualization, Funding acquisition, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was financed by the National Natural Science Foundation of China (no. 32270029, U1803232, 31670026), the National Key R&D Program of China (no. 2017YFE0122000), Social Service Team Support Program Project (no. 11899170165) and Science and Technology Innovation Special Fund (no. KFB23084) of Fujian Agriculture and Forestry University, Fujian Provincial Major Science and Technology Project (no. 2022NZ029017), Key Project from Fujian Provincial Department of Science and Technology (no. 2020N5005), and the Young and Middle-aged Teacher Education Research Project of Fujian Province (no. JAT210075).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aaqil, M., Peng, C., Kamal, A., Nawaz, T., Zhang, F., and Gong, J. (2023). Tea harvesting and processing techniques and its effects on phytochemical profile and final quality of black tea: a review. Food Secur. 12:4467. doi: 10.3390/foods12244467
Abe, S. K., and Inoue, M. (2021). Green tea and cancer and cardiometabolic diseases: a review of the current epidemiological evidence. Eur J. Clinical Nutr. 75, 865–876. doi: 10.1038/s41430-020-00710-7
Almario, J., Fabianska, I., Saridis, G., and Bucher, M. (2022). Unearthing the plant-microbe quid pro quo in root associations with beneficial fungi. New Phytol. 234, 1967–1976. doi: 10.1111/nph.18061
Alvarez, A., Saez, J. M., Costa, J. S. D., Colin, V. L., Fuentes, M. S., Cuozzo, S. A., et al. (2017). Actinobacteria: current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 166, 41–62. doi: 10.1016/j.chemosphere.2016.09.070
Amoo, A. E., and Babalola, O. O. (2017). Ammonia-oxidizing microorganisms: key players in the promotion of plant growth. J. Soil Sci. Plant Nutr. 17, 935–947. doi: 10.4067/S0718-95162017000400008
Arafat, Y., Din, I. U., Tayyab, M., Jiang, Y., Chen, T., Cai, Z., et al. (2020). Soil sickness in aged tea plantation is associated with a shift in microbial communities as a result of plant polyphenol accumulation in the tea gardens. Front. Plant Sci. 11:601. doi: 10.3389/fpls.2020.00601
Armah, E. K., Chetty, M., Adedeji, J. A., Estrice, D. E., Mutsvene, B., Singh, N., et al. (2023). Biochar: production, application and the future. In: Biochar-productive technologies, properties and applications. M. Bartoli, M. Giorcelli, and A. Tagliaferro (Eds.). Intech Open. pp. 1–26. doi: 10.5772/intechopen.105070
Auria, J. C. D., Cohen, S. P., Leung, J., Glockzin, K., Glockzin, K., Gervay-Hague, J., et al. (2022). United States tea: a synopsis of ongoing tea research and solutions to United States tea production issues. Front. Plant Sci. 13:934651. doi: 10.3389/fpls.2022.934651
Bag, S., Mondal, A., and Banik, A. (2022). Exploring tea (Camellia sinensis) microbiome: insights into the functional characteristics and their impact on tea growth promotion. Microbiol. Res. 254:126890. doi: 10.1016/j.micres.2021.126890
Bag, S., Mondal, A., Majumder, A., and Banik, A. (2021). Tea and its phytochemicals: hidden health benefits and modulation of signaling cascade by phytochemicals. Food Chem. 371:131098. doi: 10.1016/j.foodchem.2021.131098
Bagyalakshmi, B., Ponmurugan, P., and Balamurugan, A. (2017). Potassium solubilization, plant growth promoting substances by potassium solubilizing bacteria (KSB) from southern India tea plantation soil. Biocatal. and Agric. Biotechnol. 12, 116–124. doi: 10.1016/jbcab.2017.09.011
Banik, A., Chattopadhyay, A., Ganguly, S., and Mukhopadhyay, S. K. (2019). Characterisation of a tea pest specific Bacillus thuringiensis and identification of its toxin by MALDI-TOF mass spectrophotometry. Indust. Crops and Prod. 137, 549–556. doi: 10.1016/J.INDCROP.2019.05.051
Barka, E. A., Vatsa, P., Sanchez, L., Gaveau-Vaillant, N., Jacquard, C., Meier-Kolthoff, J. P., et al. (2016). Taxonomy, physiology and natural products of Actinobacteria. Microbiol. Biol. Rev. 80, 1–43. doi: 10.1128/mmbr.00019-15
Bastida, F., Eldridge, D. J., Garcia, C., Png, G. K., Bardgett, R. D., and Baquerizo, M. D. (2021). Soil microbial diversity-biomass relationships are driven by soil carbon content across global biomes. ISME J. 15, 2081–2091. doi: 10.1038/s41396-021-00906-0
Basu, S., Kumar, G., Chhabra, S., and Prasad, R. (2021). Chapter 13-role of microbes in biogeochemical cycle for enhancing soil fertility. In: New and future developments in microbial biotechnology and bioengineering. J. P. Verma, C. A. Macdonald, V. K. Gupta, and A. R. Podile (Eds.) Phytomicrobiome for sustainable agriculture. Elsevier. pp. 149–157.
Bayer, B., Saito, M. A., Mcllvin, M. R., Luker, S., Moran, D. M., Lankiewicz, T. S., et al. (2020). Metabolic versatility of the nitrite-oxidizing bacterium Nitrospira marina and its proteomic response to oxygen-limited conditions. ISME J. 15, 1025–1039. doi: 10.1038/s41396-020-00828-3
Berliner, A. J., Mochizuki, T., and Stedman, K. M. (2018). Astrovirology: viruses at large in the universe. Astrobiology 18, 207–223. doi: 10.1089/ast.2017.1649
Bertola, M., Ferrarini, A., and Visioli, G. (2021). Improvement of soil microbial diversity through sustainable agricultural practices and its evaluation by-omics approaches: a perspective for the environment, food quality and human safety. Microorganisms 9:1400. doi: 10.3390/microorganisms9071400
Bhaduri, J., Kundu, P., and Roy, S. K. (2018). Identification and molecular phylogeny analysis using random amplification of polymorphic DNA (RAPD) and 16SrRNA sequencing of N2 fixing tea field soil bacteria from North Bengal tea gardens. Afr. J. Microbiol. Res. 12, 655–663. doi: 10.5897/AJMR2018.8872
Bhattacharyya, C., Baberjee, S., Acharya, U., Mitra, A., Mallick, I., Haldar, A., et al. (2020). Evaluation of plant growth promotion properties and induction of oxidative defense mechanism by tea rhizobacteria of Darjeeling. India. Sci. Rep. 10:15536. doi: 10.1038/s41598-020-72439-z
Bhattacharyya, P. K. S., Roy, M., Dasa, S., Raya, D., Balachandar, S., Karthikeyan, A. K., et al. (2016). Elucidation of rice rhizosphere metagenome in relation to methane and nitrogen metabolism under elevated carbondioxide and temperature using whole genome metagenomic approach. Sci. Total Environ. 542, 886–898. doi: 10.1016/j.scitotenv.2015.10.154
Bhattacharyya, P., and Sarmah, S. R. (2018). “The role of microbes in tea cultivation” in Global tea science: Current status and future needs. eds. V. S. Sharma and M. T. Kumudini (Cambridge, UK: Burleigh Dodds Science Publishing), 1–35.
Bi, L., Yu, D., Han, L., Du, S., Yuan, C., He, J., et al. (2022). Unravelling the ecological complexity of soil viromes: challenges and opportunities. Sci. Total Environ. 812:152217. doi: 10.1016/j.scitotenv.2021.152217
Bidondo, L. F., Colombo, R., Bompadre, J., Benavides, M., Scorza, V., Silvani, V., et al. (2016). Cultivable bacteria associated with infective propagules of arbuscular mycorrhizal fungi: implications for mycorrhizal activity. Appl. Soil Ecol. 105, 86–90. doi: 10.1016/j.apsoil.2016.04.013
Bishnu, A., Chakraborty, A., Chakrabarti, K., and Saha, T. (2012). Ethion degradation and its correlation with microbial and biochemical parameters of tea soils. Biol. Fert. Soils. 48, 19–29. doi: 10.1007/s00374-011-0606-9
Bollmann-Giolai, A., Malone, J., and Arora, S. (2022). Diversity, detection and exploitation: linking soil fungi and plant diseases. Curr. Opin. Microbiol. 70:102199. doi: 10.1016/j.mib.2022.102199
Bora, P., and Bora, L. C. (2021). Microbial antagonists and botanicals mediated disease management in tea, Camellia sinensis (L.) O. Kuntze: An overview. Crop Protect. 148:105711. doi: 10.1016/j.cropro.2021.105711
Busby, P. E., Soman, C., Wagner, M. R., Friesen, M. L., Kremer, J., Bennett, A., et al. (2017). Research priorities for harnessing plant microbiomes in sustainable agriculture. PLoS Biol. 15:e2001793. doi: 10.1371/journal.pbio.2001793
Cernava, T., Chen, X., Krug, L., Li, H., Yang, M., and Berg, G. (2019). The tea leaf microbiome shows specific responses to chemical pesticides and biocontrol applications. Sci. Total Environ. 667, 33–40. doi: 10.1016/j.scitotenv.2019.02.319
Chauhan, P., Sharma, N., Tapwal, A., Kumar, A., Verma, G. S., Meena, M., et al. (2023). Soil microbiome: diversity, benefits and interactions with plants. Sustain. For. 15:14643. doi: 10.3390/su151914643
Chelangat, A., Gweyi-Onyango, J. P., Korir, N. K., and Mwangi, M. (2021). Influence of arbuscular mycorrhizae on callusing and root colonization of tea (Camellia sinensis) clones in Kenya. Asian Soil Res. J. 5, 21–26. doi: 10.9734/ASRJ/2021/v5i130098
Chen, Y., Deng, Y., Ding, J., Hu, H., Xu, T., Li, F., et al. (2017). District microbial communities in the active and permafrost layers on the Tibetan plateau. Mol. Ecol. 26, 6608–6620. doi: 10.1111/mec.14396
Chen, L., Hu, X., Yang, W., Xu, Z., Zhang, D., and Gao, S. (2015). The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in Populus cathayana. Ecotoxicol. Environ. Saf. 113, 460–468. doi: 10.1016/j.ecoenv.2014.12.033
Chen, H., Mothapo, N. V., and Shi, W. (2014). The significant contribution of fungi to soil N2O production across diverse ecosystems. Appl. Soil Ecol. 73, 70–77. doi: 10.1016/j.apsoil.2013.08.011
Chen, W., Shan, W., Niu, T., Ye, T., Sun, Q., and Zhang, J. (2023). Insight into regulation of adventitious root formation by arbuscular mycorrhizal fungus and exogeneous auxin in tea plant (Camellia sinensis L.) cuttings. Front. Plant Sci. 14:1258410. doi: 10.3389/fpls.2023.1258410
Chen, Y. P., Tsai, C. F., Rekha, P. D., Ghate, S. D., Huang, H. Y., Hsu, Y. H., et al. (2021). Agricultural management practices influence the soil enzyme activity and bacterial community structure in tea plantations. Bot. Stu. 62:8. doi: 10.1186/s40529-021-00314-9
Chen, X., Wei, W., Xiao, X., Wallace, D., Hu, C., Zhang, L., et al. (2022). Heterogeneous viral contribution to dissolved organic matter processing in a long-term macrocosm experiment. Environ. Internat. 158:106950. doi: 10.1016/j.envint.2021.106950
Chen, W., Ye, T., Sun, Q., Niu, T., and Zhang, J. (2021). Arbuscular mycorrhizal fungus alters root system architecture in Camellia sinensis L. as revealed by RNA-Seq analysis. Front. Plant Sci. 12:777357. doi: 10.3389/fpls.2021.777357
Chen, W., Ye, T., Sun, Q., Niu, T., and Zhang, J. (2022). Arbuscular mycorrhizal fungus alleviates anthracnose disease in tea seedlings. Front. Plant Sci. 13:1058092. doi: 10.3389/fpls.2022.1058092
Cheng, S., Fu, X., Wang, X., Liao, Y., Zeng, L., Dong, F., et al. (2017). Studies on the biochemical formation pathway of the amino acid L-theanine in tea (Camellia sinensis) and other plants. J. Agric. Food Chem. 65, 7210–7216. doi: 10.1021/acs.jafc.7b02437
Chevallereau, A., Pons, B. J., van Houte, S., and Westra, E. R. (2021). Interactions between bacterial and phage communities in natural environments. Nat. Rev. Microbiol. 20, 49–62. doi: 10.1038/s41579-021-00602-y
Chopra, A., Bobate, S., Rahi, P., Banpurkar, A., Mazumder, P. B., and Satpute, S. (2020b). Pseudomonas aeruginosa RTE4: a tea rhizobacterium with potential for plant growth promotion and biosurfactant production. Front. Bioeng. Biotechnol. 8:861. doi: 10.3389/fbioe.2020.00861
Chopra, A., Vandana, U. K., Rahi, P., Satpute, S., and Mazumder, P. B. (2020a). Plant promoting Brevibacterium sediminis A6 isolated from tea rhizosphere of Assam. India. Biocatal. Agric. Biotechnol. 27:101610. doi: 10.1016/j.bcab.2020.101610
Da Costa, D. P., da Silva, T. G. E., Araujo, A. S. F., Pereira, A. P. A., Mendes, L. W., Borges, W. S., et al. (2024). Soil fertility impact on recruitment and diversity of the soil microbiome in sub-humid tropical pastures in northeastern Brazil. Sci. Rep. 14:3919. doi: 10.1038/s41598-024-54221-7
De Silva, J., Tuwei, G., and Zhao, F. J. (2016). Environmental factors influencing aluminium accumulation in tea Camellia sinensis L. Plant Soil 400, 223–230. doi: 10.1007/s11104-015-2729-5
Deltedesco, E., Keiblinger, K. M., Piepho, H. P., Antonielli, L., Potsch, E. M., Zechmeis-Boltenstern, S., et al. (2020). Soil microbial community structure and function mainly respond to indirect effects in multifactorial climate manipulation experiment. Soil Biol. Biochem. 142:107704. doi: 10.1016/j.soilbio.2020.107704
Dhar Purkayastha, G., Mangar, P., Saha, A., and Saha, D. (2018). Evaluation of the biocontrol efficacy of Serratia marcescens strain indigenous to tea rhizosphere for the management of root rot disease in tea. PLoS One 13:e0191761. doi: 10.1371/journal.pone.0191761
Dong, C., Li, F., Yang, T., Feng, L., Zhang, S., Li, F., et al. (2019). Theanine transporters identified in tea plants (Camellia sinensis L.). Plant J. 101, 57–70. doi: 10.1111/tpj.14517
Du, M., Li, X., Cai, D., Zhao, Y., Li, Q., Wang, J., et al. (2022). In-silico study of reducing human health risk of POP residues’ direct (from tea) or indirect exposure (from tea garden soil): improved rhizosphere microbial degradation, toxicity control, and mechanism analysis. Ecotoxicol. Environ. Saf. 242:113910. doi: 10.1016/j.ecoenv.2022.113910
Dutta, J., Handique, P. J., and Thakur, D. (2015). Assessment of culturable tea rhizobacteria isolated from tea estates of Assam, India for growth promotion in commercial tea cultivars. Front. Microbiol. 6:1252. doi: 10.3389/fmicb.2015.01252
Dutta, J., and Thakur, D. (2017). Evaluation of multifarious plant growth promoting traits, antagonistic potential and phylogenetic affiliation of rhizobacteria associated with commercial tea plants grown in Darjeeling, India. PLoS One 12:e0182302. doi: 10.1371/journal.pone.0182302
Egidi, E., Delgado-Baquerizo, M., Plett, J. M., Wang, J., Eldridge, D. J., Bargett, R. D., et al. (2019). A few Ascomycota taxa dominate communities worldwide. Nat. Commun. 10:2369. doi: 10.1038/s41467-019-10373-z
Emerson, J. B., Roux, S., Brum, J. R., Bolduc, B., Woodcroft, B. J., Jang, H. B., et al. (2018). Host-linked soil viral ecology along a perma frost thaw gradient. Nat. Microbiol. 3, 870–880. doi: 10.1038/s41564-018-0190-y
FAOSTAT . (2022). Food and agriculture organization of the United Nations. Available at: https://faostat.fao.org
Fernandes, I. A. A., Maciel, G. M., Bortolini, D. G., Pedro, A. C., Rubio, F. T. B., de Carvalho, K. Q., et al. (2023). The bitter side of tea: pesticide residues and their impact on human health. Food Chem. Toxicol. 179:113955. doi: 10.1016/j.fct.2023.113955
Francioli, D., van Ruijven, J., Bakker, L., and Mommer, L. (2020). Drivers of total and pathogenic soil-borne fungal communities in grassland plant species. Fungal Ecol. 48:100987. doi: 10.1016/j.funeco.2020.100987
Fu, H., Li, H., Yin, P., Mei, H., Li, J., Zhou, P., et al. (2021). Integrated application of rapeseed cake and green manure enhances soil nutrients and microbial communities in tea garden soil. Sustain. For. 13:2967. doi: 10.3390/su13052967
Gao, X., Liu, Y., Liu, C., Guo, C., Zhang, Y., Ma, C., et al. (2023). Individual and combined effects of arbuscular mycorrhizal fungi and phytohormones on the growth and physicochemical characteristics of tea cutting seedlings. Front. Plant Sci. 14:1140267. doi: 10.3389/fpls.2023.1140267
Ge, S., Gao, J., Chang, D., He, T., Wang, M., Li, C., et al. (2023). Biochar contributes to resistance against root rot disease by stimulating soil polyphenol oxidase. Biochar 5, 1–17. doi: 10.1007/s42773-023-00257-3
Gond, D. P., Jha, S. S., Kumar, A., and Singh, S. K. (2021). Plant growth promoting bacteria and its role in green remediation. Sustain. Environ. Clean-up, 149–163. doi: 10.1016/B978-0-12-823828-8.00007-4
Gu, S., Hu, Q., Cheng, Y., Bai, L., Liu, Z., Xiao, W., et al. (2019). Application of organic fertilizer improves microbial community diversity and alters microbial network structure in tea (Camellia sinensis) plantation soils. Soil Tillage Res. 195:104356. doi: 10.1016/j.still.2019.104356
Gui, H., Fan, L. C., Wang, D. H., Yan, P., Li, X., Pang, Y. H., et al. (2022). Variations in soil nutrient dynamics and bacterial communities after the conversion of forests to long-term tea monoculture systems. Front. Microbiol. 13:896530. doi: 10.3389/fmicb.2022.896530
Guo, J. J., Liu, W. B., Zhu, C., Luo, G. W., Kong, Y. L., Ling, N., et al. (2018). Bacterial rather fungal community composition is associated with microbial activities and nutrient-use efficiencies in a paddy soil with short-term organic amendments. Plant Soil 424, 335–349. doi: 10.1007/s11104-017-3547-8
Gupta, S. K., and Chakraborty, A. P. (2020). Mycorrhiza helper bacteria: future prospects. Int. J. Res Rev. 7, 387–391.
Han, L. L., Yu, D. T., Zhang, L. M., Shen, J. P., and He, J. Z. (2017). Genetic and functional diversity of ubiquitous DNA viruses in selected Chinese agricultural soils. Sci. Rep. 7:45142. doi: 10.1038/srep45142
Han, L., Zhang, H., Xu, Y., Li, Y., and Zhou, J. (2021). Biological characteristics and salt-tolerant plant-growth promoting effects of an ACC deaminase-producing Burkholderia pyrrocinia strain isolated from the tea rhizosphere. Arch. Microbiol. 203, 2279–2290. doi: 10.1007/s00203-021-02204-x
Hao, X., Zhang, W., Zhao, F., Liu, Y., Qian, W., Wang, Y., et al. (2018). Discovery of plant viruses from tea plant (Camellia sinensis (L.) O. Kuntze) by metagenomic sequencing. Front. Microbiol. 9:2175. doi: 10.3389/fmicb.2018.02175
Hartmann, M., and Six, J. (2022). Soil structure and microbiome functions in agrosystems. Nat. Rev. Earth Environ. 4, 4–18. doi: 10.1038/s43017-022-00366-w
Hayatsu, M., Katsuyama, C., and Tago, K. (2021). Overview of recent researches on nitrifying microorganisms in soil. Soil Sci. Plant Nutri. 67, 619–632. doi: 10.1080/00380768.2021.1981119
Hazra, A., Dasgupta, N., Sengupta, C., Bera, B., and Das, S. (2019). “Tea: a worthwhile, popular beverage crop since time immemorial” in Agronomic crops. ed. M. Hasanuzzaman (Singapore: Springer Publishing), 507–531.
Hicks, L. C., Beat, F., Kjoller, R., Lukac, M., Moora, M., Weedon, J. T., et al. (2021). Toward a function-first framework to make soil microbial ecology perspective. Ecology 103:e03594. doi: 10.1002/ecy.3594
Hurwitz, B. L., and U’Ren, J. M. (2016). Viral metabolic reprogramming in marine ecosystems. Curr. Opin. Microbiol. 31, 161–168. doi: 10.1016/j.mib.2016.04.002
Idris, A. L., Fan, X., Muhammad, M. H., Guo, Y., Guan, X., and Huang, T. (2020). Ecologically controlling insect and mite pests of tea plants with microbial pesticides: a review. Arch. Microbiol. 202, 1275–1284. doi: 10.1007/s00203-020-01862-7
Jansson, J. K., and Wu, R. (2022). Soil viral diversity, ecology and climate change. Nat. Rev. Microbiol. 21, 296–311. doi: 10.1038/s41579-022-00811-z
Ji, L., Yang, X., Zhu, C., Ma, L., Chen, Y., Ling, N., et al. (2022). Land-use changes alter the arbuscular mycorrhizal fungal community composition and assembly in the ancient tea forest reserve. Agric. Ecosyst. Environ. 339:108142. doi: 10.1016/j.agee.2022.108142
Jose, P. A., Maharshi, A., and Jha, B. (2021). Actinobacteria in natural products research: progress and prospects. Microbiol. Res. 246:126708. doi: 10.1016/j.micres.2021.126708
Kalam, S., Basu, A., Ahmad, I., Sayyed, R. Z., El-Enshasy, H. A., Dailin, D. J., et al. (2020). Recent understanding of soil Acidobacteria and their ecological significance: a critical review. Front. Microbiol. 11:580024. doi: 10.3389/fmicb.2020.580024
Kizheva, Y., Eftimova, M., Rangelov, R., Micheva, N., Urshev, Z., Rasheva, I., et al. (2021). Broad host range bacteriophages found in rhizosphere soil of a healthy tomato plant in Bulgaria. Heliyon 7:e07084. doi: 10.1016/j.heliyon.2021.e07084
Kolandasamy, M., Mandal, A. K. A., Balasubramanian, M. G., and Ponnusamy, P. (2023). Multifaceted plant growth-promoting traits of indigenous rhizospheric microbes against Phomopsis theae, a causal agent of stem canker in tea plants. World J. Microbiol. Biotechnol. 39:237. doi: 10.1007/s11274-023-03688-z
Kui, L., Xiang, G., Wang, Y., Wang, Z., Li, G., Li, D., et al. (2021). Large-scale characterization of the soil microbiome in ancient tea plantations using high-throughput 16s rRNA and internal transcribed spacer amplicon sequencing. Front. Microbiol. 12:745225. doi: 10.3389/fmicb.2021.745225
Li, Y., Li, Z., Arafat, Y., and Lin, W. (2020). Studies on fungal communities and functional guilds shift in tea continuous cropping soils by high-through put sequencing. Ann. Microbiol. 70:7. doi: 10.1186/s13213-020-01555-y
Li, Y. C., Li, Z., Li, Z. W., Jiang, Y. H., Weng, B. Q., and Lin, W. X. (2016). Variations of rhizosphere bacterial communities in tea (Camellia sinensis L.) continuous cropping soil by high-throughput pyrosequencing approach. J. Appl. Microbiol. 121, 787–799. doi: 10.1111/jam.13225
Li, Y. C., Li, Z. W., Lin, W. W., Jiang, Y. H., Weng, B. Q., and Lin, W. X. (2018). Effects of biochar and sheep manure on rhizospheric soil microbial community in continuous rationing tea orchards. J. Appl. Ecol. 29, 1273–1282. doi: 10.13287/j.1001-9332.201804.036
Liao, H., Li, H., Duan, C. S., Zhou, X. Y., Luo, Q. P., An, X. L., et al. (2022). Response of soil viral communities to land use changes. Nat. Commun. 13:6027. doi: 10.1038/s41467-022-33771-2
Lin, W., Lin, M., Zhou, H., Wu, H., Li, Z., and Lin, W. (2019). The effects of chemical and organic fertilizer usage on rhizosphere soil in tea orchards. PLoS One 14:e0217018. doi: 10.1371/journal.pone.0217018
Liu, M. Y., Burgos, A., Zhang, Q., Tang, D., Shi, Y., Ma, L., et al. (2017). Analyses of transcriptome profiles and selected metabolites unravel the metabolic response of NH+4 and NO−3 as signaling molecules in tea plant (Camellia sinensis L.). Sci. Hort. 218, 293–303. doi: 10.1016/j.scienta.2017.02.036
Liu, W., Cui, S., Wu, L., Qi, W., Chen, J., Ye, Z., et al. (2023). Effects of bio-organic fertilizer on soil fertility, yield and quality of tea. J. Soil Sci. Plant Nutr. 23, 5109–5121. doi: 10.1007/s42729-023-01195-6
Liu, X., Hu, B., and Chu, C. (2022). Nitrogen assimilation in plants: current status and future prospects. J. Genetics Genomics. 49, 394–404. doi: 10.1016/j.jgg.2021.12.006
Liu, J., Li, X., and Yao, M. (2021). Research progress on assembly of plant rhizosphere microbialcommunity. Acta Microbiol Sin. 61, 231–248. doi: 10.13343/j.cnkiwsxb.20200154
Lu, Y., Song, S., Wang, R., Liu, Z., Meng, J., Sweetman, A. J., et al. (2015). Impacts of soil and water pollution on food safety and health risks in China. Environ. Int. 77, 5–15. doi: 10.1016/j.envint.2014.12.010
Lynn, T. M., Liu, Q., Hu, Y., Yuan, H., Wu, X., Khai, A. A., et al. (2017). Influence of land use on bacteria and archael diversity and community structures in three natural ecosystems and one agricultural soil. Arch. Microbiol. 199, 711–721. doi: 10.1007/s00203-017-1347-4
Ma, Z., Tanalgo, K. C., Xu, Q., Li, W., Wu, S., Ji, Q., et al. (2022). Influence of tea-Pleurotus ostreatus intercropping on soil fungal diversity and community structure. Can. J. Soil Sci. 102, 359–369. doi: 10.1139/CJSS-2021-0123
Ma, L., Yang, X., Shi, Y., Yi, X., Ji, L., Cheng, Y., et al. (2021). Response of tea yield, quality and soil bacteria characteristics to long-term nitrogen fertilization in an eleven-year field experiment. Appl. Soil Ecol. 166:103976. doi: 10.1016/j.apsoil.2021.103976
Morris, E. K., Morris, D. J. P., Vogt, S., Gleber, S. C., Bigalke, M., Wilcke, W., et al. (2019). Visualizing the dynamics of soil aggregation as affected by arbuscular mycorrhizal fungi. ISME J. 13, 1639–1646. doi: 10.1038/s41396-019-0369-0
Mukhopadhyay, M., and Mondal, T. K. (2017). Cultivation, improvement and environmental impacts of tea. Oxford Research Encyclopedia of Environmental Science. Oxford University Press. pp. 1–22. doi: 10.1093/acresfore/9780199389414.013.373
Muneer, M. A., Hou, W., Li, J., Huang, X. M., Kayani, M. U. R., Cai, Y. Y., et al. (2022). Soil pH: a key edaphic factor regulating distribution and functions of bacterial community along vertical soil profiles in redsoil of pomelo orchard. BMC Microbiol. 23:38. doi: 10.1186/s12866-02202452-x
Nabi, M. (2023). “Chapter eleven-role of microorganisms in plant nutrition and soil health” in Molecular inventions and advancements for crop improvement. Sustainable Plant Nutrition, 263–282.
Nasslahsen, B., Prin, Y., Ferhout, H., Smouni, A., and Duponnois, R. (2022). Mycorrhizae helper bacteria for managing the mycorrhizal soil infectivity. Front. Soil Sci. 2:979246. doi: 10.3389/fsoil.2022.979246
Nath, R., Sharma, G. D., and Barooah, M. (2015). Plant growth promoting endophytic fungi isolated from tea (Camellia sinensis) shrubs of Assam. India. Appl. Ecol. Environ. Res. 13, 877–891. doi: 10.15666/aeer/1303_877891
Naylor, D., McClure, R., and Jansson, J. (2022). Trends in microbial community composition and function by soil depth. Microorganisms 10:540. doi: 10.3390/microorganisms10030540
Naz, M., Dai, Z., Hussain, S., Tariq, M., Danish, S., Khan, I. U., et al. (2022). The soil pH and heavy metals revealed their impact on soil microbial community. J. Environ. Manag. 321:115770. doi: 10.1016/j.envman.2022.115770
Negrete-Bolagay, D., Zamora-Ledezma, C., Chuya-Sumba, C., DeSousa, F. B., Whitehead, D., Alexis, F., et al. (2021). Persistent organic pollutants: the trade-off between potential risks and sustainable remediation methods. J. Environ. Manag. 300:113737:113737. doi: 10.1016/j.jenvman.2021.113737
Olanrewaju, O. S., Ayangbenro, A. S., Glick, B. R., and Babalola, O. O. (2019). Plant health: feedback effect of root exudate-rhizobiome interactions. Appl. Microbial. Biotechnol. 103, 1155–1166. doi: 10.1007/s00253-018-9556-6
Ozimek, E., and Hanaka, A. (2021). Mortierella species as the plant growth-promoting fungi present in agricultural soils. Agriculture 11:7. doi: 10.3390/agriculture11010007
Panda, P., Choudhury, A., Chakraborty, S., Ray, D. P., Deb, S., Patra, P. S., et al. (2017). Phosphorus solubilizing bacteria from tea soils and their phosphate solubilizing abilities. Int. J. Bioresource Sci. 4, 113–125. doi: 10.5958/2454-9541.2017.00018.4
Pandey, A. K., Deka, B., Varshney, R., Cheramgoi, E. C., and Babu, A. (2021). Do the beneficial fungi manage phytosalinity problems in tea agro-ecosystem? Biol. Control 66, 445–462. doi: 10.1007/s10526-021-10084-9
Pandey, A. K., Hubbali, M., Dutta, V. P., and Babu, A. (2023). Characterisation and identification of fungicide insensitive Pestalotiopsis-like species pathogenic to tea crop in India. World J. Microbiol. Biotechnol. 39:34. doi: 10.1007/s11274-022-03474-3
Parihar, J., Parihar, S., Suravajhala, P., and Bagaria, A. (2022). Spatial metagenomic análisis in understanding the microbial diversity of Thar desert. Biol. 11:461. doi: 10.3390/biology11030461
Peng, C., Zhu, X. H., Hou, R. Y., Ge, G. F., Hua, R. M., Wan, X. C., et al. (2018). Aluminium and heavy metal accumulation in tea leaves: an interplay of environmental and plant factors and an assessment of exposure risks to consumers. J. Food Sci. 83, 1165–1172. doi: 10.1111/1750-3841.14093
Perez-Burillo, S., Jimenez-Zamora, A., Parraga, J., Rufian-Henares, J. A., and Pastoriza, S. (2019). Furosine and 5-hydroxymethylfurfural as chemical markers of tea processing and storage. Food Control 99, 73–78. doi: 10.1016/j.foodcont.2018.12.029
Phillippot, L., Chenu, C., Kappler, A., Rillig, M. C., and Fierer, N. (2023). The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 22, 226–239. doi: 10.1038/s41579-023-00980-5
Pokharel, S. S., Yu, H., Fang, W., Parajulee, M. N., and Chen, F. (2023). Intercropping cover crops for a vital ecosystem service: a review of the biocontrol of insect pests in tea agroecosystems. Plan. Theory 12:2361. doi: 10.3390/plants12122361
Qiao, Y., Wang, T., Huang, Q., Guo, H., Zhang, H., Xu, Q., et al. (2024). Core species impact plant health by enhancing soil microbial cooperation and network complexity during community coalescence. Soil Biol. Biochem. 188:109231. doi: 10.1016/j.soilbio.2023.109231
Ramphinwa, M. L., Mchau, G. R. A., Mashau, M. E., Madala, N. E., Chimonyo, V. G. P., Modi, T. A., et al. (2023). Eco-physiological response of secondary metabolites of teas: review of quality attributes of herbal tea. Front. Sustain. Food Syst. 7:990334. doi: 10.3389/fsufs.2023.990334
Rashid, M. I., Mujawar, L. H., Shahzad, T., Almeelbi, T., Ismail, I. M., and Oves, M. (2016). Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 183, 26–41. doi: 10.1016/j.mcres.2015.11.007
Ray, D., Baruah, P. M., and Agarwala, N. (2022). “Chapter 15-Aluminium in tea plants: phytotoxicity, tolerance and mitigation” in Harzardous and trace materials in soil and plants. eds. M. Naeem, T. Aftab, A. A. Ansari, S. S. Gill, and A. Macovei (Cambridge, MA: Academic Press), 217–229.
Ren, Z., Cheng, R., Chen, P., Xue, Y., Xu, H., Yin, Y., et al. (2023). Plant-associated microbe system in treatment of heavy metals-contaminated soil: mechanisms and applications. Water Air Soil Pollut. 234:39. doi: 10.1007/s11270-023-06061-w
Roux, S., and Emerson, J. B. (2022). Diversity in the soil virosphere: to infinity and beyond? Trends Microbiol. 30, 1025–1035. doi: 10.1016/j.tim.2022.05.003
Sangwan, S., and Prasanna, R. (2022). Mycorrhizae helper bacteria: unlocking their potential as bioenhancers of plant-arbuscular mycorrhizal fungal associations. Fungal Microbiol. 84, 1–10. doi: 10.1007/s00248-021-01831-7
Schroeter, S. A., Eveillard, D., Chaffron, S., Zoppi, J., Kampe, B., Lohmann, P., et al. (2022). Microbial community functioning during plant litter decomposition. Sci. Rep. 12:7451. doi: 10.1038/s41598-022-11485-1
Shan, W., Zhou, Y., Liu, H., and Yu, X. (2018). Endophytic Actinomycetes from tea plants (Camellia sinensis): isolation, abundance antimicrobial, and plant-growth-promoting activities. Biomed. Res. Int. 2018:1470305. doi: 10.1155/2018/1470305
Shao, Y. D., Zhang, D. J., Hu, X. C., Wu, Q. S., Jiang, C. J., Gao, X. B., et al. (2019). Arbuscular mycorrhiza improves leaf food quality of tea plants. Not. Bot. Horti. Agrobo. 47, 1842–4309. doi: 10.15835/nbha47311434
Shao, Y. D., Zhang, D. J., Hu, X. C., Wu, Q. S., Xia, T. J., Gao, X. B., et al. (2018). Mycorrhiza-induced changes in root growth and nutrient absorption of tea plants. Plant Soil Environ. 64, 283–289. doi: 10.17221/126/2018-PSE
Sharma, C., Gupta, R. K., Pathak, R. K., and Choudhary, K. K. (2013). Seasonal colonization of arbuscular mycorrhiza fungi in roots of Camellia sinensis (tea) in different tea gardens of India. ISRN Biodiversity 2013:593087, 1–6. doi: 10.1155/2013/593087
Sharma, S., and Kaur, S. (2021). “Soil microbial diversity and metagenomics” in Soil nitrogen ecology. eds. C. Cruz, K. Vishwakarma, D. K. Choudhary, and A. Varma, vol. 62 (Cham: Springer), 283–301.
Shi, J., Qu, R., Feng, M., Wang, X., Wang, L., Yang, S., et al. (2015). Oxidative degradation of decabromodiphenyl ether (BDE209) by potassium permanganate: reaction pathways, kinetics, and mechanisms assisted by density functional theory calculations. Environ. Sci. Technol. 49, 4209–4217. doi: 10.1021/es505111r
Song, Q., Deng, X., Song, R., and Song, X. (2023). Three plant growth-promoting rhizobacteria regulate the soil microbial community and promote the growth of maize seedlings. J. Plant Growth Regul. 42, 7418–7434. doi: 10.1007/s00344-023-11019-7
Su, J., Xia, Y., Yao, H. Y., Li, Y. Y., An, X. L., Singh, B. K., et al. (2017). Metagenomic assembly unravel microbial response to redox fluctuations in acid sulfate soil. Soil Biol. Biochem. 105, 244–252. doi: 10.1016/j.soilbio.2016.11.027
Sun, M., Yuan, D., Hu, X., Zhang, D., and Li, Y. (2020). Effects of mycorrhizal fungi on plant growth, nutrient absorption and phytohormones levels in tea under shading condition. Not. Bot. Horti. Agrobo. 48, 2006–2020. doi: 10.15835/48412082
Taha, M., Salama, A., El-Seedy, M., Elakhdar, I., Islam, M. S., Barutcular, C., et al. (2016). Potential impact of compost tea on soil microbial properties and performance of radish plant under sandy soil conditions-greenhouse experiments. Aust. J. Basic Appl. Sci. 10, 158–165.
Tan, L., Gu, S. S., Li, S., Ren, Z. H., Deng, Y., Liu, Z. H., et al. (2019). Responses of microbial communities and interaction networks to different management practices in tea plantation soils. Sustain. For. 11:4428. doi: 10.3390/su11164428
Thabah, S., and Joshi, S. R. (2022). Chapter 17-Plant growth promoting rhizobacteria from the perspectives of tea plantations and diseases. In: New and future developments in microbial biotechnology and bioengineering and engineering. Singh, H., Vaishnav, A. (Eds.). Sustainable Agriculture: Microorganisms as Biostimulants. Elsevier. pp. 315–332.
Trivedi, P., Leach, J. E., Tringe, S. G., Sa, T., and Singh, B. K. (2020). Plant–microbiome interactions: from community assembly to plant health. Nat. Rev. Microbiol. 18, 607–621. doi: 10.1038/s41579-020-0412-1
Trubl, G., Jang, H. B., Roux, S., Emerson, J. B., Solonenko, N., Vik, D. R., et al. (2018). Soil viruses are underexplored players in ecosystem carbon processing. mSystems 3, e00018–e00076. doi: 10.1128/mSystems.00076-18
Wang, Z., Geng, Y., and Liang, T. (2020). Optimization of reduced chemical fertilizer use in tea gardens based on the assessment of related environmental and economic benefits. Sci. Total Environ. 713:136439. doi: 10.1016/j.scitotenv.2019.136439
Wang, J., Ji, Y., Yuan, Z., Guan, J., Liu, C., and Lv, L. (2017). Analysis of bacterial community structure and diversity in different restoration methods in Qixing river wetland. Adv. J. Toxicol. Curr. Res. 1, 49–55.
Wang, S., Li, T., Zheng, Z., and Chen, H. Y. H. (2019). Soil aggregate associated bacterial metabolic activity and community structure in different aged plantations. Sci. Total Environ. 654, 1023–1032. doi: 10.1016/j.scitotenv.2018.11.032
Wang, X., Liu, J., Yu, Z., Jin, J., Liu, X., and Wang, G. (2016). Novel groups and unique distribution of phage phoH genes in paddy waters in Northeast China. Sci. Rep. 6:38428. doi: 10.1038/srep38428
Wang, S., Tang, L., and Ye, S. (2021). Dynamics of soil bacterial community diversity and composition at aggregate scales in chronosequence of tea gardens. Catena 206:105486. doi: 10.1016/j.catena.2021.105486
Wang, Z., Xu, M., Li, F., Bai, Y., Hou, J., Li, X., et al. (2023). Changes in soil bacterial communities and functional groups beneath coarse woody debris across a subalpine forest successional series. Global Ecol. Conserv. 43:e02436. doi: 10.1016/j.geco.2023.e02436
Wei, Z., Niu, S., Wei, Y., Liu, Y., Xu, Y., Yang, Y., et al. (2024). The role of extracellular polymetric substances (EPS) in chemical-degradation of persistent organic pollutants in soil: a review. Sci. Tot. Environ. 912:168877. doi: 10.1016/j.scitotenv.2023.168877
Wei, Y., Wu, Y., Yan, Y., Zou, W., Xue, J., Ma, W., et al. (2018). High-throughput sequencing of microbial community diversity in soil, grapes, leaves, grape juice and wine of grapevine from China. PLoS One 13:e0193097. doi: 10.1371/journal.pone0193097
Wright, C. L., and Lehvirta-Morley, L. E. (2023). Nitrification and beyond: metabolic versatility of ammonia oxidizing archaea. ISME J. 17, 1358–1368. doi: 10.1038/s41396-023-01467-0
Wu, Q. S., Shao, Y. D., Gao, X. B., Xia, T. J., and Kuca, K. (2019). Characterization of AMF-diversity of endosphere versus rhizosphere of tea (Camellia sinensis) crops. Indian J. Agric. Sci. 89, 348–352. doi: 10.56093/ijas.v89i2.87097
Wu, D., Wang, W., Yao, Y., Li, H., Wang, Q., and Niu, B. (2023). Microbial interactions within beneficial consortia promote soil health. Sci. Total Environ. 900:165801. doi: 10.1016/j.scitotenv.2023.165801
Wu, L., Wen, C., Qin, Y., Yin, H., Tu, Q., van Nostrand, J. D., et al. (2015). Phasing amplicon sequencing on illumine Miseq for robust environmental microbial community analysis. BMC Microbiol. 15, 1–12. doi: 10.1186/s12866-015-0450-4
Xi, S., Chu, H., Zhou, Z., Li, T., Zhang, S., Xu, X., et al. (2023). Effect of potassium fertilizer on tea yield and quality: a meta analysis. Eur. J. Agron. 144:126767. doi: 10.1016/j.eja.2023.126767
Xiang, L., Harindintwali, J. D., Redmile-Gordon, M., Chang, S. X., Fu, Y., He, C., et al. (2022). Integrating biochar, bacteria and plants for sustainable remediation of soils contaminated with organic pollutants. Environ. Sci. Technol. 56, 16546–16566. doi: 10.1021/acs.est.2c02976
Xie, S., Yang, F., Feng, H., Yu, Z., Liu, C., Wei, C., et al. (2021). Organic fertilizer reduced carbon and nitrogen in run-off and buffered soil acidification in tea plantations: evidence in nutrient content and isotope fractionations. Sci. Total Environ. 762:143059. doi: 10.1016/j.scitotenv.2020.143059
Xin, W., Zhang, J., Yu, Y., Tian, Y., Li, H., Chen, X., et al. (2024). Root microbiota of tea plants regulate nitrogen homeostasis and theanine synthesis to influence tea quality. Curr. Biol. 34, 868–880.e6. doi: 10.1016/j.cub.2024.01.044
Xu, P., Fan, X. Y., Mao, Y. X., Cheng, H. Y., Xu, A. A., Lai, W. Y., et al. (2022). Temporal metabolite responsiveness of microbiota in the tea plant phyllosphere promotes continuous suppression of fungal pathogens. J. Adv. Res. 39, 49–60. doi: 10.1016/j.jare.2021.10.003
Xu, G., Liu, Y., Long, Z., Hu, S., Zhang, Y., and Jiang, H. (2018). Effects of exotic plantation forests on soil edaphon and organic matter fractions. Sci. Total Environ. 626, 59–68. doi: 10.1016/j.scitotenv.2018.01.088
Yadav, A., Borrelli, J. C., Elshahed, M. S., and Youssef, N. H. (2021). Genomic analysis of family UBA6911 (group 18 Acidobacteria) expands the metabolic capacities of the phylum and highlights adaptations to terrestrial habitats. Appl. Environ. Microbiol. 87:e0094721. doi: 10.1128/AEM.00947-21
Yan, K., Abbas, M., Meng, L., Cai, H., and Zhao, X. (2021). Analysis of fungal diversity and community structure in Sichuan dark tea pile-fermentation. Front. Microbiol. 12:706714. doi: 10.3389/fmicb.2021.706714
Yan, L., Riaz, M., Liu, J., Yu, M., and Cuncang, J. (2022). The aluminium tolerance and detoxification in plants; recent advances and prospects. Crit. Rev. Environ. Sci. Technol. 52, 1491–1527. doi: 10.1080/10643389.2020.1859306
Yan, P., Shen, C., Fan, L. C., Li, X., Zhang, L. P., Zhang, L., et al. (2018). Tea planting affects acidification and nitrogen and phosphorus distribution in soil. Agric. Ecosys. Environ. 254, 20–25. doi: 10.1016/j.agee.2017.11.015
Yan, P., Shen, C., Zou, Z., Fan, L., Li, X., Zhang, L., et al. (2022). Increased soil fertility in tea gardens lead to declines in fungal diversity and complex subsoils. Agronomy 12:1751. doi: 10.3390/agronomy12081751
Yan, P., Wu, L. Q., Wang, D. H., Fu, J., Shen, C., Li, X., et al. (2020). Soil acidification in Chinese tea plantations. Sci. Total Environ. 715:136963. doi: 10.1016/j.scitotenv.2020.136963
Yang, X. D., Ni, K., Shi, Y. Z., Yi, X. Y., Zhang, Q. F., Fang, L., et al. (2018). Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 252, 74–82. doi: 10.1016/j.agee.2017.10.004
Ye, J., Wang, Y., Wang, Y., Hong, L., Jia, X., Kang, J., et al. (2022). Improvement of soil acidification in tea plantations by long-term use of organic fertilizers and its effect on tea yield and quality. Front. Plant Sci. 13:1055900. doi: 10.3389/fpls.2022.1055900
Zhang, Z., Ge, S. B., Fan, L. C., Guo, S., Hu, Q., Ahammed, G. J., et al. (2022). Diversity in rhizospheric microbial communities in tea varieties at different locations and tapping potential beneficial microorganisms. Front. Microbiol. 13:1027444. doi: 10.3389/fmicb.2022.1027444
Zhang, X., Long, H., Huo, D., Awan, M. I., Shao, J., Mahmood, A., et al. (2022). Insights into the functional role of tea microbes on tea growth, quality and resistance against pests and diseases. Not. Bot. Horti. Agrobo. 50:12915. doi: 10.15835/nbha50412915
Zhang, W., Ni, K., Long, L., and Ruan, J. (2023). Nitrogen transport and assimilation in tea plant (Camellia sinensis): a review. Front. Plant Sci. 14:1249202. doi: 10.3389/fpls.2023.1249202
Zhang, X. C., Wang, N. N., Hou, M. M., Wu, H. H., Jiang, H., Zhou, Z. W., et al. (2022). Contribution of K solubilizing bacteria (Burkholderia sp.) promotes tea plant growth (Camellia sinensis) and leaf polyphenols content by improving soil available K level. Funct. Plant Biol. 49, 283–294. doi: 10.1071/FP21193
Zhang, S., Wang, Y., Sun, L., Qiu, C., Ding, Y., Gu, H., et al. (2020). Organic mulching positively regulates the soil microbial communities and ecosystem functions in tea plantation. BMC Microbiol. 20:103. doi: 10.1186/s12866-020-01794-8
Zheng, X., Wu, Y., Xu, A., Lin, C., Wang, H., Yu, J., et al. (2023). Response of soil microbial communities and functions to long-term tea (Camellia sinensis L.) planting in a subtropical region. Forests 14:1288. doi: 10.3390/f14071288
Zheng, N., Yu, Y., Shi, W., and Yao, H. (2019). Biochar suppresses N2O emmissions and alters microbial communities in acidic tea soil. Environ. Sci. Pollut. Res. 26, 35978–35987. doi: 10.1007/s11356-019-06704-8
Zi, H., Jiang, Y., Cheng, X., Li, W., and Huang, X. (2020). Change of rhizospheric bacterial community of the ancient wild tea along elevational gradients in Ailao Mountain, China. Sci. Rep. 10:9203. doi: 10.1038/s41598-020-66173-9
Keywords: microbial communities, soil health, soil microorganisms, tea gardens, tea plant
Citation: Jibola-Shittu MY, Heng Z, Keyhani NO, Dang Y, Chen R, Liu S, Lin Y, Lai P, Chen J, Yang C, Zhang W, Lv H, Wu Z, Huang S, Cao P, Tian L, Qiu Z, Zhang X, Guan X and Qiu J (2024) Understanding and exploring the diversity of soil microorganisms in tea (Camellia sinensis) gardens: toward sustainable tea production. Front. Microbiol. 15:1379879. doi: 10.3389/fmicb.2024.1379879
Edited by:
Paola Grenni, National Research Council, ItalyReviewed by:
Kalpana Bhatt, Purdue University, United StatesCopyright © 2024 Jibola-Shittu, Heng, Keyhani, Dang, Chen, Liu, Lin, Lai, Chen, Yang, Zhang, Lv, Wu, Huang, Cao, Tian, Qiu, Zhang, Guan and Qiu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoyan Zhang, emhhbmd4aWFveWFuQHh0YmcuYWMuY24=; Xiayu Guan, Z3h5MzAyQDEyNi5jb20=; Junzhi Qiu, anVuemhpcWl1QDEyNi5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.