
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 07 March 2024
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 15 - 2024 | https://doi.org/10.3389/fmicb.2024.1366181
This article is part of the Research Topic New and advanced mechanistic insights into the influences of the infant gut microbiota on human health and disease View all 23 articles
 Wei Liu1,2
Wei Liu1,2 Hanlei Yan1,2
Hanlei Yan1,2 Wanying Jia1,2Jingjing Huang1,2Zihao Fu1,2Wenyao Xu1,2Hui Yu1,2Weili Yang1Weikang Pan1Baijun Zheng1
Wanying Jia1,2Jingjing Huang1,2Zihao Fu1,2Wenyao Xu1,2Hui Yu1,2Weili Yang1Weikang Pan1Baijun Zheng1 Yong Liu2
Yong Liu2 Xinlin Chen2Ya Gao1*
Xinlin Chen2Ya Gao1* Donghao Tian1,2*
Donghao Tian1,2*Background: Several studies have pointed to the critical role of gut microbiota (GM) and their metabolites in Hirschsprung disease (HSCR) pathogenesis. However, the detailed causal relationship between GM and HSCR remains unknown.
Methods: In this study, we used two-sample Mendelian randomization (MR) analysis to investigate the causal relationship between GM and HSCR, based on the MiBioGen Consortium’s genome-wide association study (GWAS) and the GWAS Catalog’s HSCR data. Reverse MR analysis was performed subsequently, and the sensitivity analysis, Cochran’s Q-test, MR pleiotropy residual sum, outlier (MR-PRESSO), and the MR-Egger intercept were used to analyze heterogeneity or horizontal pleiotropy. 16S rDNA sequencing and targeted mass spectrometry were developed for initial validation.
Results: In the forward MR analysis, inverse-variance weighted (IVW) estimates suggested that Eggerthella (OR: 2.66, 95%CI: 1.23–5.74, p = 0.01) was a risk factor for HSCR, while Peptococcus (OR: 0.37, 95%CI: 0.18–0.73, p = 0.004), Ruminococcus2 (OR: 0.32, 95%CI: 0.11–0.91, p = 0.03), Clostridiaceae1 (OR: 0.22, 95%CI: 0.06–0.78, p = 0.02), Mollicutes RF9 (OR: 0.27, 95%CI: 0.09–0.8, p = 0.02), Ruminococcaceae (OR: 0.16, 95%CI: 0.04–0.66, p = 0.01), and Paraprevotella (OR: 0.45, 95%CI: 0.21–0.98, p = 0.04) were protective factors for HSCR, which had no heterogeneity or horizontal pleiotropy. However, reverse MR analysis showed that HSCR (OR: 1.02, 95%CI: 1–1.03, p = 0.049) is the risk factor for Eggerthella. Furthermore, some of the above microbiota and short-chain fatty acids (SCFAs) were altered in HSCR, showing a correlation.
Conclusion: Our analysis established the relationship between specific GM and HSCR, identifying specific bacteria as protective or risk factors. Significant microbiota and SCFAs were altered in HSCR, underlining the importance of further study and providing new insights into the pathogenesis and treatment.
Hirschsprung disease (HSCR) is one of the most common forms of neurocristopathy in children, an intestinal peristaltic disorder showing obstruction and proximal megacolon that affects 1 in 5,000 human births. The pathogenesis is generally characterized by aganglionosis in the distal intestine (Gui et al., 2017; Fu et al., 2020). Most patients with heritable variation are diagnosed by rectal biopsy in the neonatal period due to significant constipation; however, a subset of children (~20%) develop symptoms until early childhood or even adolescence (De Lorijn et al., 2005). Surgery is the primary treatment method, but commonly gives rise to medical complications, such as enterocolitis (~35% after surgery), soiling, anastomotic stricture, or leak with abscess and chronic constipation, threatening crucially the growth and development of children (Heuckeroth, 2018).
It is well known that HSCR primally arises from genetic factors, such as RET, EDNRB, RARB, GATA2, and SOX10 mutations. Recently, increasing reports indicate that abnormal intestinal microenvironment contributed to the HSCR pathogenesis, especially gut microbiota (GM; Obata and Pachnis, 2016; Kuil et al., 2021; Vincent et al., 2023). The GM begins to colonize after birth, and changes significantly from the neonatal period to early childhood, which coincides with the development of the enteric nervous system (ENS; Backhed et al., 2015; Gritz and Bhandari, 2015; Yassour et al., 2016; Stewart et al., 2018). Some researchers transplanted fecal bacteria into the nerve-free colon and found that the GM played a positive role in ENS development (Obata and Pachnis, 2016). Meanwhile, 16S rDNA sequencing and targeted mass spectrometry were used to develop a detailed profile of how the gut microbiome and its metabolites benefited the aganglionosis animal model (Soret et al., 2020; Tian et al., 2023). However, it remains to be investigated to determine whether and which GM has a causal relationship with HSCR.
Mendelian randomization (MR) is a new statistical method that can explore the causal relationship between exposed factors and disease. MR follows Mendelian’s second law and relies on independent random assignment of genetic variants during meiosis to achieve randomization effects similar to randomized controlled trials (RCTs), which can effectively overcome the confounding factors (Ma et al., 2023; Vincent et al., 2023; Yan et al., 2023; Zanoaga et al., 2023). MR has been widely used to analyze the causal relationship between GM and spinal arthropathy (Lai et al., 2023), ophthalmic disease (Li and Lu, 2023), tumors (Jiang et al., 2023), psychiatric disorders (Zeng et al., 2023), and circulatory system disease (Dai et al., 2023). However, the relationship between GM and HSCR has rarely been studied based on MR methods.
In this study, a two-sample MR analysis was performed using pooled statistics from the genome-wide association study (GWAS) of MiBioGen and the GWAS Catalog to assess the causal relationship between GM and HSCR, which could provide new insights into the potential pathogenesis of how GM contributes to HSCR and the assignment of effective treatment strategies.
This study utilized pooled-level genetic data to conduct a bidirectional, two-sample MR analysis, aiming to test for the association between GM and HSCR. In order to reduce bias and obtain reliable results, we tried to satisfy the following three hypotheses when using MR analysis: (1) Correlation hypothesis: instrumental variables (IVs) are closely related to exposure; (2) Exclusivity hypothesis: IVs do not affect outcomes by other means; and (3) Independence hypothesis: IVs are not affected by confounding factors (Davies et al., 2018; Figure 1).
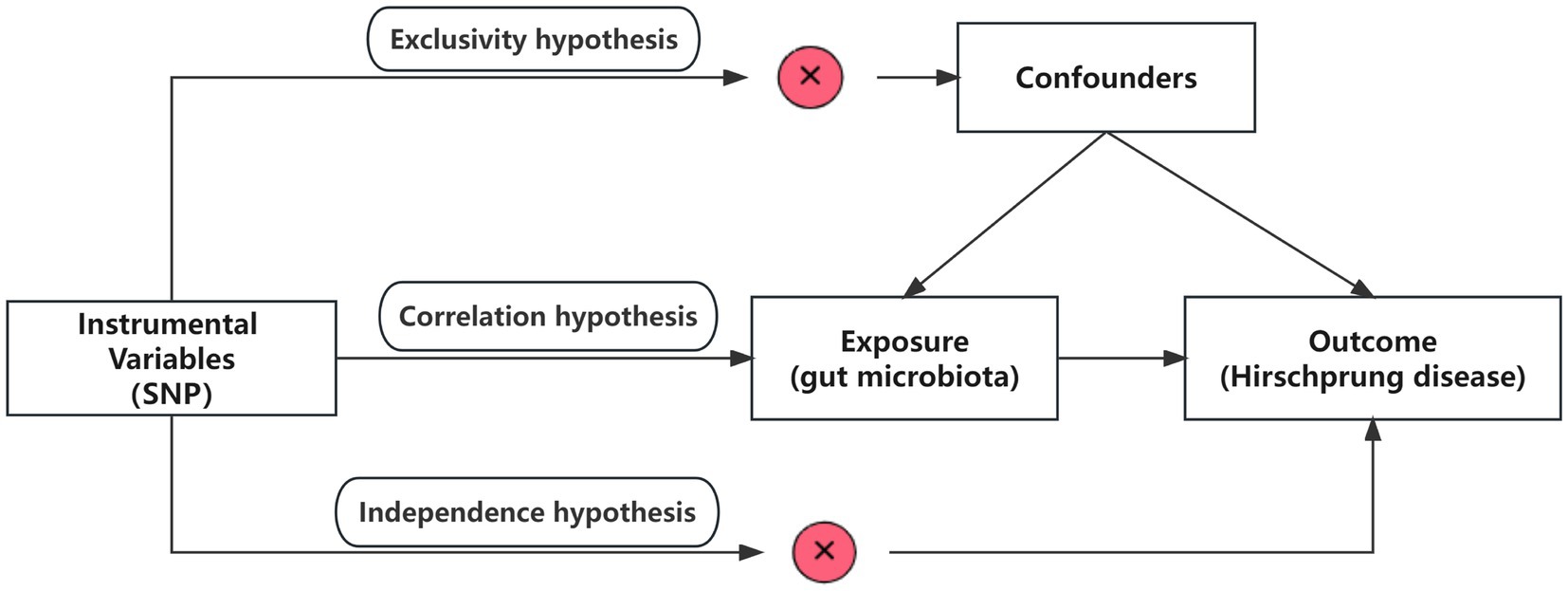
Figure 1. Overview of the study design. SNPs, single-nucleotide polymorphisms.
Genetic variation for GM comes from the largest genome-wide meta-analysis of GM composition published by Kurilshikov et al. (2021) and MiBioGen Consortium (n.d.). They collected genome-wide genotypes and 16S fecal microbiome data from 18,340 individuals across 24 cohorts based on the MiBioGen consortium. A total of 196 taxa have been included in the study, classified under 5 biological categories: phylum, class, order, family, and genus. It should be noted that participants of European origin were exclusively considered, and 15 GM taxa (unknown family or genus) without specific species names were excluded from the analysis. GWAS summary statistics for HSCR were downloaded from the NHGRI-EBI GWAS Catalog (Sollis et al., 2023) on 10 November 2023 for study GCST005289 (Fadista et al., 2018). This study used the phenotype of HSCR and combined the most extensive collection of sporadic Hirschsprung cases to date, including 586 cases and 5,620 controls of European ancestry, which combined data from Denmark, the U.S., Finland, and Sweden. In the process of data analysis, strict quality control was carried out on the samples (Fadista et al., 2018).
The following selection criteria were used to choose the IVs: (1) single-nucleotide polymorphisms (SNPs) associated with each genus at the locus-wide significance threshold (p < 1.0 × 10−5) were selected as potential IVs (Li et al., 2023); (2) 1000 Genomes project European samples data were used as the reference panel to calculate the linkage disequilibrium (LD) between the SNPs, and among those SNPs that had R2 < 0.001 (clumping window size = 10,000 kb), only the SNPs with the lowest p-values were retained (Fan et al., 2023); (3) SNPs with minor allele frequency (MAF) ≤ 0.01 were removed; and (4) when palindromic SNPs existed, the forward strand alleles were inferred using allele frequency information. (5) In order to control bias caused by IVs, the strength of each IV was evaluated using the formula F = β2/SE2 to calculate the F-statistic, where β represents the effect size of SNP on exposure, and SE represents the standard deviation of SNP on exposure (Burgess et al., 2011). If the corresponding F-statistic was >10, it was considered that there was no significant weak instrumental bias (Burgess et al., 2011). Power calculations were conducted with the online Power Calculator Tool (Burgess, 2014; Online sample size and power calculator for Mendelian randomization with a binary outcome, n.d.).
MR is a statistical analysis method used for causal inference. It uses genetic variation as an IV to deduce the causal relationship between exposure and outcome, which can effectively avoid the influence of confounding bias in traditional epidemiological studies (Emdin et al., 2017). In this study, we analyzed the potential causal relationship between GM and HSCR with two-sample MR and strictly reduced the effect of confounders. The MR analysis process is shown in Figure 2. In the process of analyzing causal correlation, we used a variety of methods to evaluate it, including inverse-variance weighted (IVW; Burgess et al., 2017), MR-Egger test (Bowden et al., 2015), weighted median (WM), weighted mode (WMO), simple mode, and MR-PRESSO. The IVW method uses a meta-analysis method to combine Wald estimates of each SNP to summarize GM’s overall estimate of HSCR. If there is no horizontal pleiotropy, the IVW results are highly effective. MR-Egger tests the existence of the intercept term and uses it to evaluate pleiotropy. If the intercept term is very close to zero, then the MR-Egger regression model is very close to IVW, but if the intercept term is very different from zero, it indicates that there may be horizontal pleiotropy between these IVs. The WM is a method of considering the weight of the data when calculating the median, which can be used when the weight of the data should be considered more. The WMO takes into account the weights of the data points and is able to give a suitable mode if there are duplicate values with different weights. The simple mode can be used when each data point in the data set is weighted equally. MR-PRESSO improves the accuracy of causal associations by detecting and removing outliers. In the present study, we used the IVW method as the primary analysis method, while other methods were used to evaluate the stability of our results.
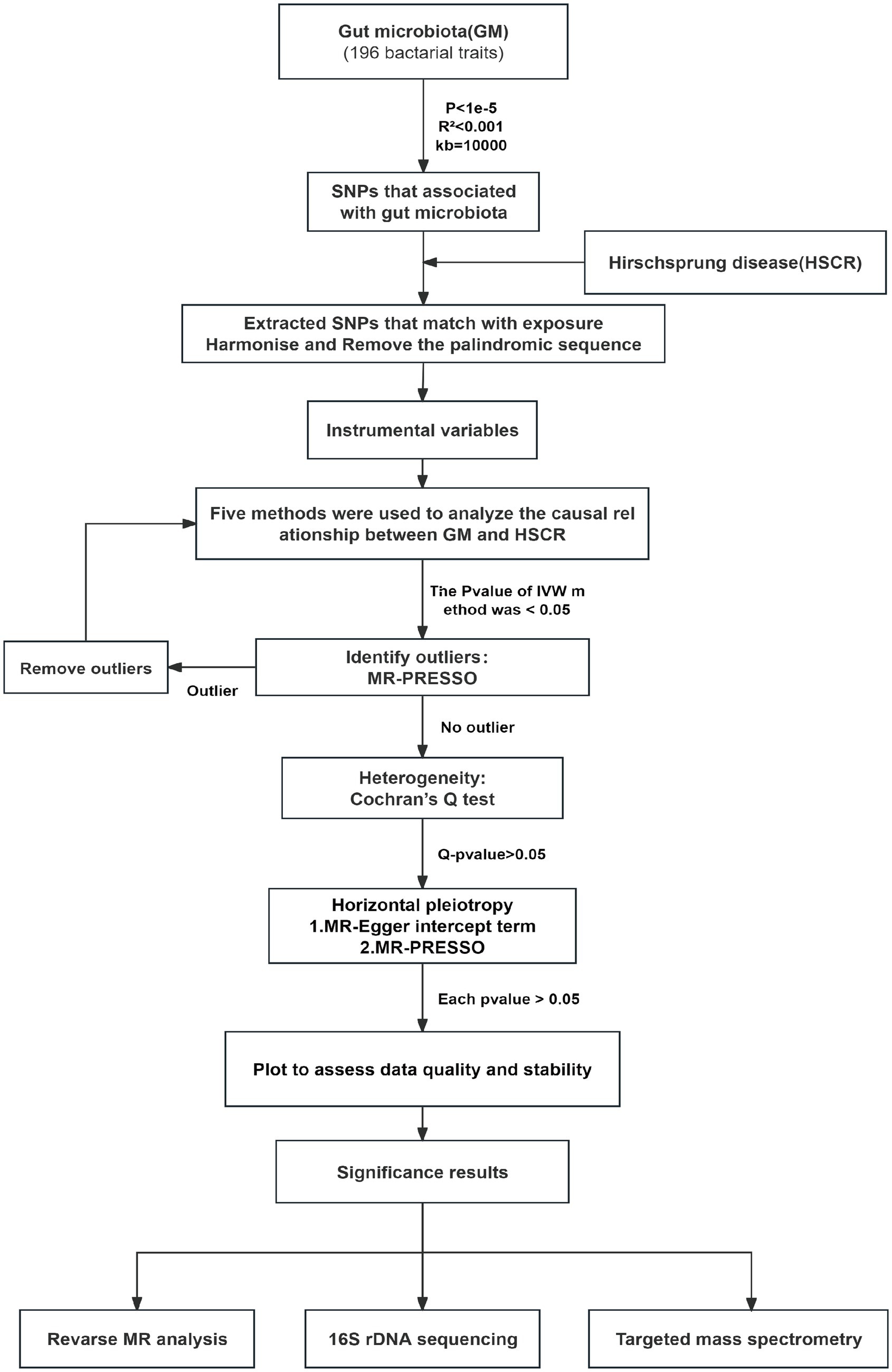
Figure 2. Flow chart of forward MR analysis. MR, Mendelian randomization; IVs, instrumental variable; IVW, inverse-variance weighted; SNPs, single-nucleotide polymorphisms.
Heterogeneity was examined using Cochran’s Q-test, and a p-value of <0.05 was considered heterogeneous. The horizontal pleiotropy of IVs was measured by the MR-Egger intercept term. Meanwhile, MR-PRESSO was used to analyze pleiotropy further and pick out outliers in IVs. In addition, to identify potential heterogeneous SNPs, the “leave-one-out” analysis was performed by omitting each instrumental SNP in turn, aiming to verify the reliability and stability of the causal association (Chen et al., 2022).
To further evaluate whether there is a causal relationship between intestinal flora and HSCR, we performed a reverse MR analysis. Bacteria with a significant causal association with positive MR were selected as outcome variables, and HSCR was used as exposure for reverse MR analysis. The analysis methods and SNP selection conditions were consistent with those of positive MR.
False discovery rate (FDR) correction was conducted by applying the q-value procedure, with a FDR q-value of <0.1 (Storey and Tibshirani, 2003). Genera of GM and HSCR were considered to have a suggestive association when p < 0.05 but q ≥ 0.1 and a significant association when p < q < 0.05.
To characterize the bacterial community taxonomically, we performed 16S rDNA amplicon sequencing. First, genomic DNA was extracted from feces using the QIAamp Fast DNA Stool Mini Kit. Illumina-compatible primers (338F, 5′-ACTCCTACGGGAGGCAGCAG-30; 806R, 5′-GGACTACHVGGGTWTCTAAT-30) were then used to amplify the 16S rDNA V3–V4 regions by conventional PCR (Tian et al., 2023). The amplification products were subjected to gel purification prior to quantification by QuantiFluor-ST. After reverse transcription, the cDNA library was processed by fragmentation, end repair, and A-tailing using the TruSeq DNA PCR-free sample preparation kit. Then, the pooled library was run and sequenced on the NovaSeq600. According to QIIM, the effective Tags were obtained after chimera picking and quality filtering. Clustering by Uparse v7.0.1001 was performed to generate a list of open reference operational taxonomic units (OTUs) with an identity of 97%. The taxonomic assignment was subsequently achieved with the SSU rRNA database (Tian et al., 2023).
A targeted mass spectrometry assay was developed to detect the short-chain fatty acid (SCFA) levels in feces. Briefly, acetic, propionic, butyric, isobutyric, valeric, isovaleric, and hexanoic acids were purchased from Sigma Aldrich, as standard solutions with 10 concentration gradients (0.02, 0.1, 0.5, 1, 2, 5, 10, 25, 50, and 100 μg/mL). The sample was mixed with 50 μL of 15% phosphoric acid, 10 μL of 75 μg/mL of isohexanoic acid (internal standard), and 140 μL of diethyl ether, homogenated for 1 min, then centrifuged at 15,000 g at 4°C for 10 min. Next, the supernatant was collected for the test. The Agilent HP-INNOWax column was used for split injection, and the sample volume was 1 μL, the split injection ratio was 10:1, the inlet temperature was 250°C, the ion source temperature was 230°C, the transmission line temperature was 250°C, and the quadrupole temperature was 150°C. The programmed temperature increased from 90°C to 120°C at 10°C/min, next to 150°C at 5°C/min, and finally, to 250°C at 25°C/min for 2 min. The helium carrier gas flow rate was 1.0 mL/min. Mass spectrometry conditions included an electron bombardment ionization (EI) source, SIM scanning mode, and electron energy of 70 eV. The samples and standard solutions of SCFAs were measured by MS. The area under the curve for standard SCFA solution normalized to the internal standard was used as a single point on that SCFA standard curve. A standard curve for each SCFA was established based on the concentration gradient and served to calculate the concentration of this SCFA in the samples. All the samples were measured repeatedly, at least six times (Tian et al., 2023).
R software (version 4.3.1) and GraphPad Prism (version 8.0.1) were used to conduct the statistical analysis. We performed MR by the TwoSampleMR (version 0.5.7), MR-PRESSO (version 1.0; Verbanck et al., 2018), q-value (version 2.34.0; Storey and Tibshirani, 2003), and ggplot2 (version 3.3.3) R packages. Normally distributed data were presented as means standard error of the mean (SEM), and non-normally distributed data were presented as medians (interquartile range). Depending on the distribution and variances, a two-tailed Student’s t-test was applied to compare differences between groups. Significance was set at a p-value of <0.05. All figure panels represent data independently repeated at least three times, yielding similar results.
According to the selection criteria for IVs, 2,699 SNPs (p < 1.0 × 10−5) were selected for 196 intestinal flora in our MR analysis process. Details of all SNPs are provided in Supplementary Table S1.
IVW estimates were employed as the primary MR assay, and seven intestinal flora significantly associated with HSCR were identified. As shown in Figure 3, Eggerthella (OR: 2.66, 95%CI: 1.23–5.74, p = 0.01) was associated with an increased risk of HSCR, while Peptococcus (OR: 0.37, 95%CI: 0.18–0.73, p = 0.004), Ruminococcus2 (OR: 0.32, 95%CI: 0.11–0.91, p = 0.03), Clostridiaceae1 (OR: 0.22, 95%CI: 0.06–0.78, p = 0.02), Mollicutes RF9 (OR: 0.27, 95%CI: 0.09–0.8, p = 0.02), Ruminococcaceae (OR: 0.16, 95%CI: 0.04–0.66, p = 0.01) and Paraprevotella (OR: 0.45, 95%CI: 0.21–0.98, p = 0.04) were associated with reduced risks of HSCR. We corrected all p-values with FDR, and the Q-values were all greater than 0.05 (Supplementary Table S2). The other four methods are consistent with the direction of the IVW beta value (Figure 4).

Figure 3. Mendelian randomization analysis of gut microbiota and Hirschsprung disease. OR, odds ratio; 95% CI, 95% confidence interval; MR, Mendelian randomization; SNPs, single-nucleotide polymorphisms.
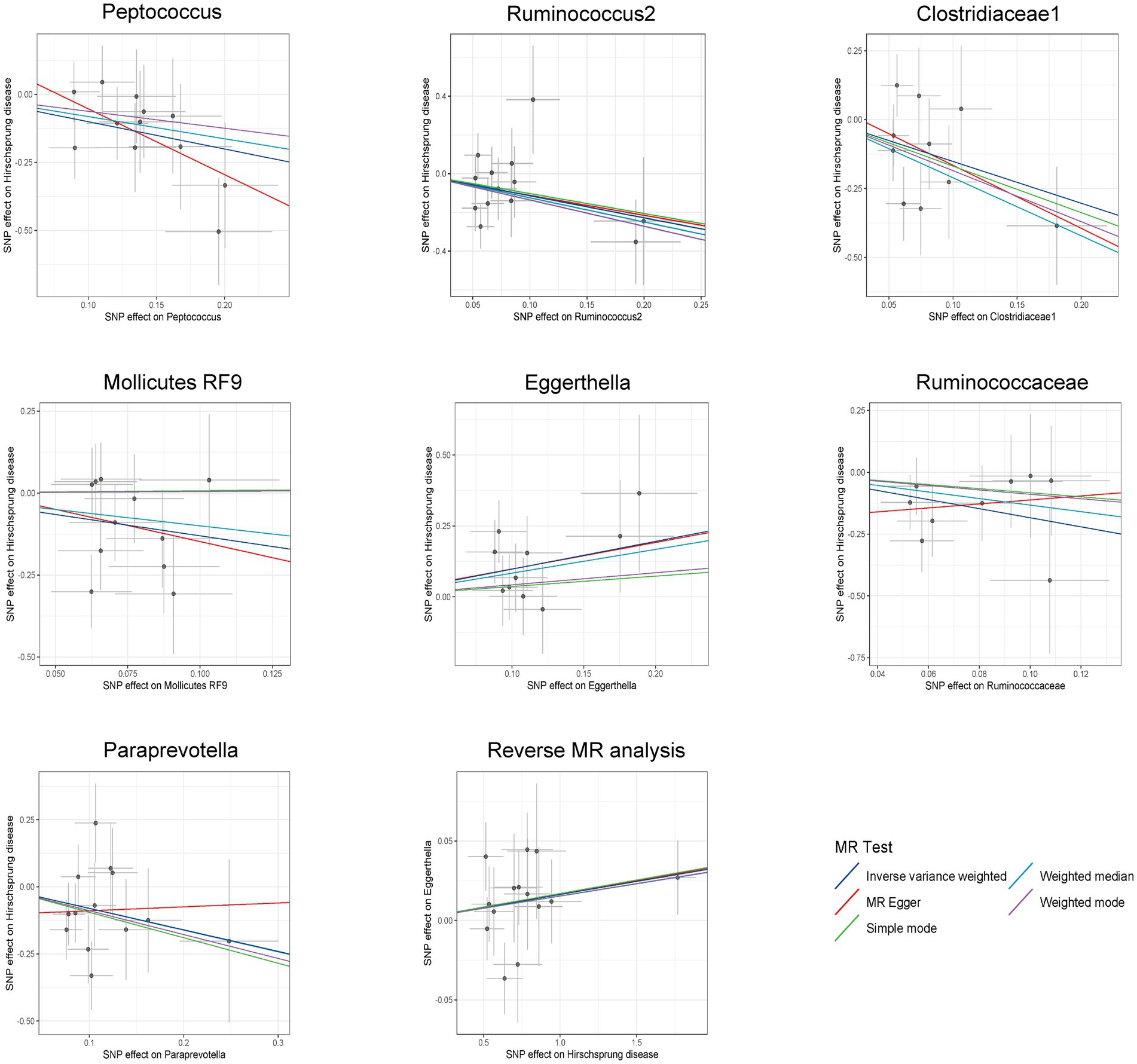
Figure 4. Scatter plots for bidirectional MR analyses of the causal effect of gut microbiota on Hirschsprung disease. SNPs, single-nucleotide polymorphisms; MR, Mendelian randomization.
Among these seven causal associations, the F-statistics of the IVs ranged from 17.42 to 32.26, eliminating the bias of weak IVs (Supplementary Table S3). As shown in Table 1, Cochran’s Q-test showed no significant heterogeneity for these IVs (p > 0.05; Supplementary Table S4), the MR-Egger regression intercept analysis found no horizontal pleiotropy (p > 0.05; Supplementary Table S5), and the MR-PRESSO test showed no outliers (global test p > 0.05; Supplementary Table S6). Moreover, after correcting the outliers, there is still no pleiotropy, which is supplemented by the MR-Egger regression intercept analysis. Subsequently, by drawing scatter plots (Figure 4) and leave-one-out plots (Figure 5), we did not find significant outliers for all IVs.
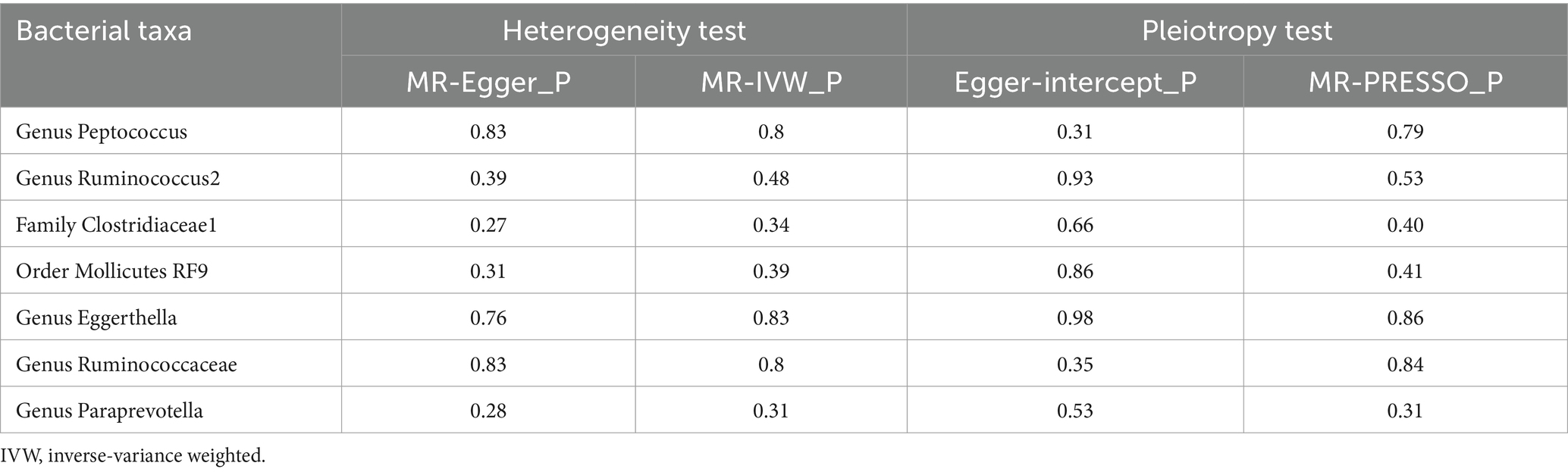
Table 1. Sensitivity analysis between gut microbiota and Hirschsprung disease.

Figure 5. Leave-one-out plots for the bidirectional MR analysis of the causal effect of gut microbiota on Hirschsprung disease. MR, Mendelian randomization; SNPs, single-nucleotide polymorphisms.
Reverse MR analysis showed that HSCR was associated with an increased abundance of Eggerthella (Supplementary Table S7; Figure 4), while no significant causal association was found with the other six bacteria (Supplementary Table S7). Cochran’s Q-test showed no significant heterogeneity in the IVs of HSCR (p > 0.05; Supplementary Table S8). Meanwhile, the analysis results of MR-Egger and MR-PRESSO (p > 0.05; Supplementary Table S8) show that horizontal pleiotropy is insignificant.
To investigate the alteration of the significant intestinal flora in HSCR, 16S rDNA sequencing was employed. Compared with the control, the relative abundance of Clostridiaceae (P: 0.002), Ruminococcus (P: 0.0003), and Paraprevotella (P: 0.00003) was significantly decreased (p < 0.05), while Eggerthellaceae was increased without statistical significance (p < 0.1) in HSCR. There were no significant changes in Ruminococcaceae (P: 0.671) or Peptococcus (P: 0.185; Figure 6A; Supplementary Tables S9, S10).
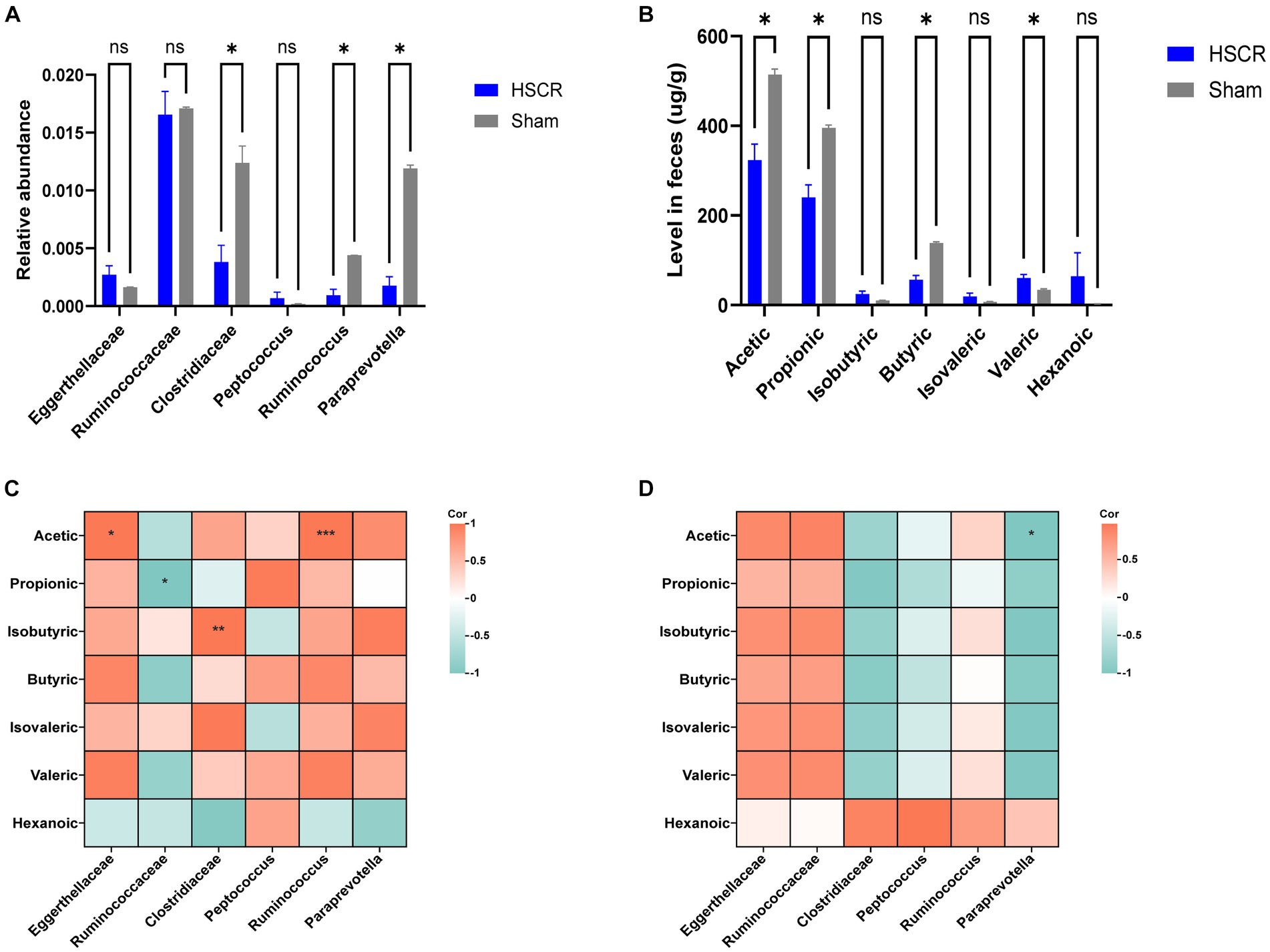
Figure 6. Detection of microbiota and short-chain fatty acids. (A) Relative abundance of GM. (B) Level of SCFAs in feces. (C) Correction analysis of GM and SCFAs in the HSCR group. (D) Correction analysis of GM and SCFAs in the control group. HSCR, Hirschsprung disease; GM, gut microbiota; SCFAs, short-chain fatty acids. Cor, correlation coefficient. Data were represented as means ± SEM (n = 3 biological replicates per group). *p < 0.05, **p < 0.01, and ***p < 0.001.
According to MR and the difference analysis, Clostridiaceae and Ruminococcus, as protective flora, showed a significant decrease in HSCR, and were reported as the producers of SCFAs (Vojinovic et al., 2019; Wu et al., 2023). Thus, we performed metabolically targeted mass spectrometry for SCFAs in fecal samples. Compared with the control, acetic, propionic, and butyric acid levels were lower in the HSCR group, while caproic acid levels were higher (Figure 6B; Supplementary Tables S11, S12). We further analyzed the correlation between significant flora and SCFAs. In the HSCR group, Ruminococcaceae was negatively correlated with acetic acid (R: −0.998, P: 0.041), while Clostridiaceae was positively correlated with isobutyric acid (R: 1.000, P: 0.002), and Ruminococcus was positively correlated with acetic acid (R: 1.000, P: 0.001; Figure 6C; Supplementary Table S13). However, in the control group, only Paraprevotella was negatively associated with acetic acid (R: −1.000, P: 0.012; Figure 6D; Supplementary Table S14).
These results suggested that the protective flora alternation was closely associated with SCFA levels in the intestinal microenvironment of HSCR patients.
HSCR is characterized by abnormal structure and function of the ENS, which is supposed to be dominated by genetic factors (Montalva et al., 2023). HSCR occurs unusually in infants who delay or have no meconium excretion; it is also diagnosed in preschool children (Parathan et al., 2020). There is further development and maturity of ENS after birth (Radenkovic et al., 2018), during which the GM colonizes and changes significantly (Stewart et al., 2018), indicating the microbiota is correlated with HSCR pathogenesis. Studies have shown that the abnormal intestinal microenvironment contributed crucially to the HSCR pathogenesis, especially gut GM, but without details (Obata and Pachnis, 2016; Vincent et al., 2023). In this study, we performed bidirectional two-sample MR to analyze the causal relationship between GM and HSCR, as well as 16S rDNA sequencing and targeted mass spectrometry to characterize the bacterial community taxonomically and metabolically. We found that Eggerthella was associated with an increased risk of HSCR, and HSCR was identified to promote Eggerthella flora colonizing the intestine. Meanwhile, Peptococcus, Ruminococcus, Clostridiaceae, Mollicutes RF9, Ruminococcaceae, and Paraprevotella were identified as protective factors for HSCR, of which the relative abundance of Clostridiaceae, Ruminococcus, and Paraprevotella in HSCR was significantly decreased and was correlated with SCFAs.
The development of the ENS is a gradual process, going through key developmental stages and gradually maturing in the few years after birth (Gershon and Ratcliffe, 2004; Nagy and Goldstein, 2017; Radenkovic et al., 2018; Parathan et al., 2020). Coincidentally, the GM, as a major factor in the intestinal microenvironment, begins to colonize immediately after birth, with significant changes from the neonatal period to early childhood (Backhed et al., 2015; Gritz and Bhandari, 2015; Yassour et al., 2016; Stewart et al., 2018). This may explain why many patients do not have obvious symptoms in early life but occur intestinal obstruction in childhood or even adolescence and are diagnosed with HSCR by internal diameter biopsy (Barnes et al., 1986; Wheatley et al., 1990; Pemmada et al., 2023). Some studies have shown that people with HSCR have different GM than healthy children, and the microbiota diversity is lower (Yan et al., 2014; Li et al., 2016; Neuvonen et al., 2018). For HSCR, some protective flora and its metabolites may promote enteric neural stem cells phenotype, thereby ameliorating the intestinal motivity due to innate genetic mutations (De Vadder et al., 2018; Tian et al., 2023). Conversely, risk factors related to GM may lead to chronic inflammation and affect local intestinal immunity, even intestinal glial cell maturation (Obata and Pachnis, 2016; Behzadi et al., 2024), which all contribute to the HSCR.
SCFAs are one of the main metabolites of GM. In our MR results, some SCFA-producing bacteria, such as Peptococcus (Wang H. et al., 2023), Ruminococcus2 (Wu et al., 2023), Clostridiaceae1 (Vojinovic et al., 2019), and Ruminococcaceae (D'Amato et al., 2020; Zhou et al., 2022) were identified to be protective to HSCR, some of which in HSCR was significantly decreased, and were correlated with SCFAs. Animal studies suggested that SCFAs could independently stimulate the enteric neural stem cell phenotype, thereby improving enteric neural disorders (Fung et al., 2021; Tian et al., 2023; Wang L. et al., 2023). Recent studies revealed that SCFA promoted enterochromaffin cells (ECs) to secrete 5-hydroxytryptamine (5-HT; Atarashi et al., 2013; Furusawa et al., 2013; Reigstad et al., 2015; Yano et al., 2015), which is regulated by the calcitonin selective reuptake transporter (SERT) of epithelial cells and plays crucial roles in enteric neuroregeneration and neuroprotection (Bian et al., 2007). Intestinal SERT is expressed lower in children compared to adults, and therefore 5-HT is more available in the early stages of ENS development and maturity (Bian et al., 2007). Additionally, SCFAs specifically increase the proportion of excitatory cholinergic neurons in the colon and regulate motor function (Soret et al., 2010). Moreover, Ruminococcus2 and Clostridiaceae1 participate in the metabolism of cholic acid (Tanaka et al., 2020; Wang et al., 2020; Molinero et al., 2022), which can activate the G-protein-coupled bile acid receptor (TGR5; Lin et al., 2023). Secondary bile acids have been shown to stimulate secretion of 5-HT, glucagon-like peptide 1 (GLP-1), and calcitonin gene-related peptide (CGRP), all of which significantly regulate the ENS and intestinal motility (Bampton et al., 2002; Kidd et al., 2008; Alemi et al., 2013). These results suggested that the flora (Peptococcus, Ruminococcus, Clostridiaceae, and Ruminococcaceae) with metabolite SCFAs and bile acids contribute to ENS development and functional maturation, which play a potential protective role and thereby reduce the risk of HSCR.
Eggerthella was the only significant factor both in the MR and the reverse MR analyses. In our study, Eggerthella colonization increased the risk of HSCR, and the HSCR patients were more conducive to Eggerthella survival, which was consistent with 16S rDNA sequencing. According to a recent study, Eggerthella could induce intestinal Th17 activation (Alexander et al., 2022), while the pro-inflammatory factor IL-17 significantly inhibited the proliferation and migration and induced apoptosis of enteric neural stem cells (Tian et al., 2023). This implies that Eggerthella may contribute to HSCR by promoting the release of inflammatory factors. In reverse, HSCR patients may promote Eggerthella colonization. This finding provides new clues about the etiological mechanism of HSCR and helps clinical antibiotic selection to prevent and cure Hirschsprung-associated enterocolitis (HAEC).
Moreover, whether and how the microbiome induces HSCR during pregnancy remains unclear. It is reported that the mother’s GM changes significantly during pregnancy (Goltsman et al., 2018) and plays a vital role in fetal health (Di Simone et al., 2020). One animal study confirmed that maternal GM metabolites circulated between the mother and fetus, such as SCFAs, regulate placental growth and blood vessel formation (Pronovost et al., 2023). Furthermore, the inflammatory factors produced by the mother’s gut flora impacted early fetal development (Apostol et al., 2020). Therefore, it is much more valuable to explore how the above six protective GMs reduce the risk of HSCR during pregnancy, just as the Eggerthella increases the risk of HSCR.
This study has some limitations because aggregated data were used rather than raw data, it was not possible to group by fetal, neonatal, and infantile analyses. Therefore, we cannot determine at which growth stage the gut flora most significantly contributes to HSCR, which also prevents us from further exploring the details of the causal association between GM and HSCR. For sensitivity analysis and pleiotropy testing, there seems to be a need for more genetic variants as IVs. Therefore, SNPs in the MR analysis failed to reach the traditional GWAS significance threshold (p < 5.0 × 10−8). With the gradual improvement of GWAS data, it is suggested that future MR studies can be stratified based on pre-birth and post-birth studies to obtain more precise and complete results. Furthermore, MR has the advantage of evaluating the direct effect of exposure factors on the outcome, while intermediate effects or effects via other pathways may be ignored (Sekula et al., 2016). All the above may limit the full understanding of the causal chain, which needs to be investigated further.
In conclusion, our bidirectional two-sample MR study demonstrates a causal association between GM and HSCR. Specifically, Peptococcus, Ruminococcus, Clostridiaceae, Mollicutes RF9, Ruminococcaceae, and Paraprevotella are protective factors against HSCR, while Eggerthella may increase the risk for HSCR. Moreover, reverse MR also confirmed that HSCR was a risk factor for Eggerthella. 16S rDNA sequencing and targeted mass spectrometry presented significant alteration of Clostridiaceae, Ruminococcus, and Paraprevotella, as well as correlation with SCFAs, which underline the importance of further study and provide new insights into the pathogenesis and treatment. However, the details need further study.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
WL: Data curation, Methodology, Validation, Visualization, Writing – original draft. HYa: Software, Supervision, Visualization, Writing – review & editing. WJ: Formal analysis, Methodology, Resources, Writing – review & editing. JH: Conceptualization, Investigation, Validation, Writing – review & editing. ZF: Methodology, Software, Writing – review & editing. WX: Data curation, Methodology, Validation, Writing – review & editing. HYu: Funding acquisition, Software, Writing – review & editing. WY: Data curation, Methodology, Writing – review & editing. WP: Supervision, Validation, Writing – review & editing. BZ: Investigation, Writing – review & editing. YL: Investigation, Writing – review & editing. XC: Investigation, Writing – review & editing. YG: Funding acquisition, Methodology, Supervision, Writing – review & editing. DT: Funding acquisition, Investigation, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The study was supported by grants from the National Natural Science Foundation of China (nos: 82300575, 82071692, and 82170531), Xi’an Jiaotong University (no: YXJLRH2022053), and the General Project of Shaanxi Science and Technology Department (no: 2022SF-133/033).
The authors sincerely thank the scientists who shared their data on the public database. We thank the creators of the packages TwoSampleMR, MR-PRESSO, q-value, and ggplot2.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2024.1366181/full#supplementary-material
Alemi, F., Poole, D. P., Chiu, J., Schoonjans, K., Cattaruzza, F., Grider, J. R., et al. (2013). The receptor TGR5 mediates the prokinetic actions of intestinal bile acids and is required for normal defecation in mice. Gastroenterology 144, 145–154. doi: 10.1053/j.gastro.2012.09.055
Alexander, M., Ang, Q. Y., Nayak, R. R., Bustion, A. E., Sandy, M., Zhang, B., et al. (2022). Human gut bacterial metabolism drives Th17 activation and colitis. Cell Host Microbe 30, 17–30.e9. doi: 10.1016/j.chom.2021.11.001
Apostol, A. C., Jensen, K. D. C., and Beaudin, A. E. (2020). Training the fetal immune system through maternal inflammation-a layered hygiene hypothesis. Front. Immunol. 11:123. doi: 10.3389/fimmu.2020.00123
Atarashi, K., Tanoue, T., Oshima, K., Suda, W., Nagano, Y., Nishikawa, H., et al. (2013). Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 500, 232–236. doi: 10.1038/nature12331
Backhed, F., Roswall, J., Peng, Y., Feng, Q., Jia, H., Kovatcheva-Datchary, P., et al. (2015). Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 17, 690–703. doi: 10.1016/j.chom.2015.04.004
Bampton, P. A., Dinning, P. G., Kennedy, M. L., Lubowski, D. Z., and Cook, I. J. (2002). The proximal colonic motor response to rectal mechanical and chemical stimulation. Am. J. Physiol. Gastrointest. Liver Physiol. 282, G443–G449. doi: 10.1152/ajpgi.00194.2001
Barnes, P. R., Lennard-Jones, J. E., Hawley, P. R., and Todd, I. P. (1986). Hirschsprung's disease and idiopathic megacolon in adults and adolescents. Gut 27, 534–541. doi: 10.1136/gut.27.5.534
Behzadi, P., Dodero, V., and Golubnitschaja, O. (2024). Systemic inflammation as the health-related communication tool between the human host and gut microbiota in the framework of predictive. Prevent Personal Med. 18, 203–241. doi: 10.1007/978-3-031-46891-9_15
Bian, X., Patel, B., Dai, X., Galligan, J. J., and Swain, G. (2007). High mucosal serotonin availability in neonatal guinea pig ileum is associated with low serotonin transporter expression. Gastroenterology 132, 2438–2447. doi: 10.1053/j.gastro.2007.03.103
Bowden, J., Davey Smith, G., and Burgess, S. (2015). Mendelian randomization with invalid instruments: effect estimation and bias detection through egger regression. Int. J. Epidemiol. 44, 512–525. doi: 10.1093/ije/dyv080
Burgess, S. (2014). Sample size and power calculations in Mendelian randomization with a single instrumental variable and a binary outcome. Int. J. Epidemiol. 43, 922–929. doi: 10.1093/ije/dyu005
Burgess, S., Bowden, J., Fall, T., Ingelsson, E., and Thompson, S. G. (2017). Sensitivity analyses for robust causal inference from Mendelian randomization analyses with multiple genetic variants. Epidemiology 28, 30–42. doi: 10.1097/EDE.0000000000000559
Burgess, S., and Thompson, S. G.Collaboration CCG (2011). Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 40, 755–764. doi: 10.1093/ije/dyr036
Chen, M., Xie, C. R., Shi, Y. Z., Tang, T. C., and Zheng, H. (2022). Gut microbiota and major depressive disorder: a bidirectional Mendelian randomization. J. Affect. Disord. 316, 187–193. doi: 10.1016/j.jad.2022.08.012
Dai, H., Hou, T., Wang, Q., Hou, Y., Zhu, Z., Zhu, Y., et al. (2023). Roles of gut microbiota in atrial fibrillation: insights from Mendelian randomization analysis and genetic data from over 430,000 cohort study participants. Cardiovasc. Diabetol. 22:306. doi: 10.1186/s12933-023-02045-6
D'Amato, A., Di Cesare, M. L., Lucarini, E., Man, A. L., Le Gall, G., Branca, J. J. V., et al. (2020). Faecal microbiota transplant from aged donor mice affects spatial learning and memory via modulating hippocampal synaptic plasticity-and neurotransmission-related proteins in young recipients. Microbiome. 8:140. doi: 10.1186/s40168-020-00914-w
Davies, N. M., Holmes, M. V., and Davey, S. G. (2018). Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ 362:k601. doi: 10.1136/bmj.k601
De Lorijn, F., Reitsma, J. B., Voskuijl, W. P., Aronson, D. C., Ten Kate, F. J., Smets, A. M., et al. (2005). Diagnosis of Hirschsprung's disease: a prospective, comparative accuracy study of common tests. J. Pediatr. 146, 787–792. doi: 10.1016/j.jpeds.2005.01.044
De Vadder, F., Grasset, E., Mannerås Holm, L., Karsenty, G., Macpherson, A. J., Olofsson, L. E., et al. (2018). Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. U. S. A. 115, 6458–6463. doi: 10.1073/pnas.1720017115
Di Simone, N., Santamaria Ortiz, A., Specchia, M., Tersigni, C., Villa, P., Gasbarrini, A., et al. (2020). Recent insights on the maternal microbiota: impact on pregnancy outcomes. Front. Immunol. 11:528202. doi: 10.3389/fimmu.2020.528202
Emdin, C. A., Khera, A. V., and Kathiresan, S. (2017). Mendelian randomization. JAMA 318, 1925–1926. doi: 10.1001/jama.2017.17219
Fadista, J., Lund, M., Skotte, L., Geller, F., Nandakumar, P., Chatterjee, S., et al. (2018). Genome-wide association study of Hirschsprung disease detects a novel low-frequency variant at the RET locus. Eur. J. Hum. Genet. 26, 561–569. doi: 10.1038/s41431-017-0053-7
Fan, J., Zhou, Y., Meng, R., Tang, J., Zhu, J., Aldrich, M. C., et al. (2023). Cross-talks between gut microbiota and tobacco smoking: a two-sample Mendelian randomization study. BMC Med. 21:163. doi: 10.1186/s12916-023-02863-1
Fu, A. X., Lui, K. N.-C., Tang, C. S.-M., Ng, R. K., Lai, F. P.-L., Lau, S.-T., et al. (2020). Whole-genome analysis of noncoding genetic variations identifies multiscale regulatory element perturbations associated with Hirschsprung disease. Genome Res. 30, 1618–1632. doi: 10.1101/gr.264473.120
Fung, C., Cools, B., Malagola, S., Martens, T., Tack, J., Kazwiny, Y., et al. (2021). Luminal short-chain fatty acids and 5-HT acutely activate myenteric neurons in the mouse proximal colon. Neurogastroenterol. Motil. 33:e14186. doi: 10.1111/nmo.14186
Furusawa, Y., Obata, Y., Fukuda, S., Endo, T. A., Nakato, G., Takahashi, D., et al. (2013). Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450. doi: 10.1038/nature12721
Gershon, M. D., and Ratcliffe, E. M. (2004). Developmental biology of the enteric nervous system: pathogenesis of Hirschsprung's disease and other congenital dysmotilities. Semin. Pediatr. Surg. 13, 224–235. doi: 10.1053/j.sempedsurg.2004.10.019
Goltsman, D. S. A., Sun, C. L., Proctor, D. M., DiGiulio, D. B., Robaczewska, A., Thomas, B. C., et al. (2018). Metagenomic analysis with strain-level resolution reveals fine-scale variation in the human pregnancy microbiome. Genome Res. 28, 1467–1480. doi: 10.1101/gr.236000.118
Gritz, E. C., and Bhandari, V. (2015). The human neonatal gut microbiome: a brief review. Front. Pediatr. 3:17. doi: 10.3389/fped.2015.00017
Gui, H., Schriemer, D., Cheng, W. W., Chauhan, R. K., Antinolo, G., Berrios, C., et al. (2017). Whole exome sequencing coupled with unbiased functional analysis reveals new Hirschsprung disease genes. Genome Biol. 18:48. doi: 10.1186/s13059-017-1174-6
Heuckeroth, R. O. (2018). Hirschsprung disease—integrating basic science and clinical medicine to improve outcomes. Nat. Rev. Gastroenterol. Hepatol. 15, 152–167. doi: 10.1038/nrgastro.2017.149
Jiang, Z., Mou, Y., Wang, H., Li, L., Jin, T., Wang, H., et al. (2023). Causal effect between gut microbiota and pancreatic cancer: a two-sample Mendelian randomization study. BMC Cancer 23:1091. doi: 10.1186/s12885-023-11493-y
Kidd, M., Modlin, I. M., Gustafsson, B. I., Drozdov, I., Hauso, O., and Pfragner, R. (2008). Luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants, and olfactants. Am. J. Physiol. Gastrointest. Liver Physiol. 295, G260–G272. doi: 10.1152/ajpgi.00056.2008
Kuil, L. E., MacKenzie, K. C., Tang, C. S., Windster, J. D., Le, T. L., Karim, A., et al. (2021). Size matters: large copy number losses in Hirschsprung disease patients reveal genes involved in enteric nervous system development. PLoS Genet. 17:e1009698. doi: 10.1371/journal.pgen.1009698
Kurilshikov, A., Medina-Gomez, C., Bacigalupe, R., Radjabzadeh, D., Wang, J., Demirkan, A., et al. (2021). Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 53, 156–165. doi: 10.1038/s41588-020-00763-1
Lai, B., Jiang, H., Gao, Y., and Zhou, X. (2023). Causal effects of gut microbiota on scoliosis: a bidirectional two-sample mendelian randomization study. Heliyon. 9:e21654. doi: 10.1016/j.heliyon.2023.e21654
Li, C., and Lu, P. (2023). Association of gut Microbiota with age-related macular degeneration and Glaucoma: a bidirectional Mendelian randomization study. Nutrients 15:646. doi: 10.3390/nu15214646
Li, Y., Poroyko, V., Yan, Z., Pan, L., Feng, Y., Zhao, P., et al. (2016). Characterization of intestinal microbiomes of Hirschsprung's disease patients with or without enterocolitis using Illumina-MiSeq high-throughput sequencing. PLoS One 11:e0162079. doi: 10.1371/journal.pone.0163239
Li, N., Wang, Y., Wei, P., Min, Y., Yu, M., Zhou, G., et al. (2023). Causal effects of specific gut microbiota on chronic kidney diseases and renal function-a two-sample Mendelian randomization study. Nutrients 15:360. doi: 10.3390/nu15020360
Lin, S., Wang, S., Wang, P., Tang, C., Wang, Z., Chen, L., et al. (2023). Bile acids and their receptors in regulation of gut health and diseases. Prog. Lipid Res. 89:101210. doi: 10.1016/j.plipres.2022.101210
Ma, T., Chen, M., Cheng, X., and Bai, Y. (2023). Assessment of bidirectional relationships between frailty and mental disorders: a bidirectional Mendelian randomization study. J. Am. Med. Dir. Assoc. doi: 10.1016/j.jamda.2023.10.009
MiBioGen Consortium. (n.d.). MiBioGen. Available at: https://mibiogen.gcc.rug.nl/ (Accessed 16 September 2022).
Molinero, N., Conti, E., Walker, A. W., Margolles, A., Duncan, S. H., and Delgado, S. (2022). Survival strategies and metabolic interactions between Ruminococcus gauvreauii and Ruminococcoides bili, isolated from human bile. Microbiol Spectr. 10:e0277621. doi: 10.1128/spectrum.02776-21
Montalva, L., Cheng, L. S., Kapur, R., Langer, J. C., Berrebi, D., Kyrklund, K., et al. (2023). Hirschsprung disease. Nat. Rev. Dis. Primers. 9:54. doi: 10.1038/s41572-023-00465-y
Nagy, N., and Goldstein, A. M. (2017). Enteric nervous system development: a crest cell's journey from neural tube to colon. Semin. Cell Dev. Biol. 66, 94–106. doi: 10.1016/j.semcdb.2017.01.006
Neuvonen, M. I., Korpela, K., Kyrklund, K., Salonen, A., de Vos, W., Rintala, R. J., et al. (2018). Intestinal microbiota in Hirschsprung disease. J. Pediatr. Gastroenterol. Nutr. 67, 594–600. doi: 10.1097/MPG.0000000000001999
Obata, Y., and Pachnis, V. (2016). The effect of microbiota and the immune system on the development and Organization of the Enteric Nervous System. Gastroenterology 151, 836–844. doi: 10.1053/j.gastro.2016.07.044
Online sample size and power calculator for Mendelian randomization with a binary outcome. (n.d.). Available at: https://sb452.shinyapps.io/power/
Parathan, P., Wang, Y., Leembruggen, A. J., Bornstein, J. C., and Foong, J. P. (2020). The enteric nervous system undergoes significant chemical and synaptic maturation during adolescence in mice. Dev. Biol. 458, 75–87. doi: 10.1016/j.ydbio.2019.10.011
Pemmada, V., Bhat, G., and Bhat, B. K. (2023). Adult Hirschsprung's Disease. J. Gastrointest. Surg. 27, 1029–1031. doi: 10.1007/s11605-022-05560-1
Pronovost, G. N., Yu, K. B., Coley-O'Rourke, E. J. L., Telang, S. S., Chen, A. S., Vuong, H. E., et al. (2023). The maternal microbiome promotes placental development in mice. Sci. Adv. 9:eadk1887. doi: 10.1101/2023.02.15.528712
Radenkovic, G., Radenkovic, D., and Velickov, A. (2018). Development of interstitial cells of Cajal in the human digestive tract as the result of reciprocal induction of mesenchymal and neural crest cells. J. Cell. Mol. Med. 22, 778–785. doi: 10.1111/jcmm.13375
Reigstad, C. S., Salmonson, C. E., Rainey, J. F. 3rd, Szurszewski, J. H., Linden, D. R., Sonnenburg, J. L., et al. (2015). Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. FASEB J. 29, 1395–1403. doi: 10.1096/fj.14-259598
Sekula, P., Del Greco, M. F., Pattaro, C., and Köttgen, A. (2016). Mendelian randomization as an approach to assess causality using observational data. J. Am. Soc. Nephrol. 27, 3253–3265. doi: 10.1681/ASN.2016010098
Sollis, E., Mosaku, A., Abid, A., Buniello, A., Cerezo, M., Gil, L., et al. (2023). The NHGRI-EBI GWAS catalog: knowledgebase and deposition resource. Nucleic Acids Res. 51, D977–D985. doi: 10.1093/nar/gkac1010
Soret, R., Chevalier, J., De Coppet, P., Poupeau, G., Derkinderen, P., Segain, J. P., et al. (2010). Short-chain fatty acids regulate the enteric neurons and control gastrointestinal motility in rats. Gastroenterology 138, 1772–1782. doi: 10.1053/j.gastro.2010.01.053
Soret, R., Schneider, S., Bernas, G., Christophers, B., Souchkova, O., Charrier, B., et al. (2020). Glial cell-derived neurotrophic factor induces enteric neurogenesis and improves Colon structure and function in mouse models of Hirschsprung disease. Gastroenterology 159, 1824–38 e17. doi: 10.1053/j.gastro.2020.07.018
Stewart, C. J., Ajami, N. J., O'Brien, J. L., Hutchinson, D. S., Smith, D. P., Wong, M. C., et al. (2018). Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 562, 583–588. doi: 10.1038/s41586-018-0617-x
Storey, J. D., and Tibshirani, R. (2003). Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. U. S. A. 100, 9440–9445. doi: 10.1073/pnas.1530509100
Tanaka, M., Sanefuji, M., Morokuma, S., Yoden, M., Momoda, R., Sonomoto, K., et al. (2020). The association between gut microbiota development and maturation of intestinal bile acid metabolism in the first 3 y of healthy Japanese infants. Gut Microbes 11, 205–216. doi: 10.1080/19490976.2019.1650997
Tian, D., Xu, W., Pan, W., Zheng, B., Yang, W., Jia, W., et al. (2023). Fecal microbiota transplantation enhances cell therapy in a rat model of hypoganglionosis by SCFA-induced MEK1/2 signaling pathway. EMBO J. 42:e111139. doi: 10.15252/embj.2022111139
Verbanck, M., Chen, C.-Y., Neale, B., and Do, R. (2018). Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 50, 693–698. doi: 10.1038/s41588-018-0099-7
Vincent, E., Chatterjee, S., Cannon, G. H., Auer, D., Ross, H., Chakravarti, A., et al. (2023). Ret deficiency decreases neural crest progenitor proliferation and restricts fate potential during enteric nervous system development. Proc. Natl. Acad. Sci. U. S. A. 120:e2211986120. doi: 10.1073/pnas.2211986120
Vojinovic, D., Radjabzadeh, D., Kurilshikov, A., Amin, N., Wijmenga, C., Franke, L., et al. (2019). Relationship between gut microbiota and circulating metabolites in population-based cohorts. Nat. Commun. 10:5813. doi: 10.1038/s41467-019-13721-1
Wang, Q., Hao, C., Yao, W., Zhu, D., Lu, H., Li, L., et al. (2020). Intestinal flora imbalance affects bile acid metabolism and is associated with gallstone formation. BMC Gastroenterol. 20:59. doi: 10.1186/s12876-020-01195-1
Wang, L., Lv, W. Q., Yang, J. T., Lin, X., Liu, H. M., Tan, H. J., et al. (2023). Enteric nervous system damage caused by abnormal intestinal butyrate metabolism may lead to functional constipation. Front. Microbiol. 14:1117905. doi: 10.3389/fmicb.2023.1117905
Wang, H., Shen, Q., Fu, Y., Liu, Z., Wu, T., Wang, C., et al. (2023). Effects on diabetic mice of consuming lipid extracted from foxtail millet (Setaria italica): gut microbiota analysis and serum metabolomics. J. Agric. Food Chem. 71, 10075–10086. doi: 10.1021/acs.jafc.3c02179
Wheatley, M. J., Wesley, J. R., Coran, A. G., and Polley, T. Z. Jr. (1990). Hirschsprung's disease in adolescents and adults. Dis. Colon Rectum 33, 622–629. doi: 10.1007/BF02052222
Wu, X., Li, Q., Lin, D., Cai, J., Huang, H., and Tan, H. (2023). Gut microbiota and hypertensive disorders in pregnancy: evidence from the Mendelian randomization study. Aging 15, 9105–9127. doi: 10.18632/aging.205019
Yan, W., Jiang, M., Hu, W., Zhan, X., Liu, Y., Zhou, J., et al. (2023). Causality investigation between gut microbiota, derived metabolites, and obstructive sleep apnea: a bidirectional Mendelian randomization study. Nutrients 15:544. doi: 10.3390/nu15214544
Yan, Z., Poroyko, V., Gu, S., Zhang, Z., Pan, L., Wang, J., et al. (2014). Characterization of the intestinal microbiome of Hirschsprung's disease with and without enterocolitis. Biochem. Biophys. Res. Commun. 445, 269–274. doi: 10.1016/j.bbrc.2014.01.104
Yano, J. M., Yu, K., Donaldson, G. P., Shastri, G. G., Ann, P., Ma, L., et al. (2015). Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 161, 264–276. doi: 10.1016/j.cell.2015.02.047
Yassour, M., Vatanen, T., Siljander, H., Hamalainen, A. M., Harkonen, T., Ryhanen, S. J., et al. (2016). Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci. Transl. Med. 8:343ra81. doi: 10.1126/scitranslmed.aad0917
Zanoaga, M. D., Friligkou, E., He, J., Pathak, G. A., Koller, D., Cabrera-Mendoza, B., et al. (2023). Brain-wide Mendelian randomization study of anxiety disorders and symptoms. Biol. Psychiatry. S0006-3223(23)01718-3. doi: 10.1016/j.biopsych.2023.11.006
Zeng, H., Zhou, K., Zhuang, Y., Li, A., Luo, B., and Zhang, Y. (2023). Unraveling the connection between gut microbiota and Alzheimer's disease: a two-sample Mendelian randomization analysis. Front. Aging Neurosci. 15:1273104. doi: 10.3389/fnagi.2023.1273104
Zhou, F., Jiang, H., Kong, N., Lin, J., Zhang, F., Mai, T., et al. (2022). Electroacupuncture attenuated anxiety and depression-like behavior via inhibition of hippocampal inflammatory response and metabolic disorders in TNBS-induced IBD rats. Oxid. Med. Cell. Longev. 2022:8295580. doi: 10.1155/2022/8295580
Keywords: Mendelian randomization analysis, Hirschsprung disease, gut microbiota, causality, bidirectional
Citation: Liu W, Yan H, Jia W, Huang J, Fu Z, Xu W, Yu H, Yang W, Pan W, Zheng B, Liu Y, Chen X, Gao Y and Tian D (2024) Association between gut microbiota and Hirschsprung disease: a bidirectional two-sample Mendelian randomization study. Front. Microbiol. 15:1366181. doi: 10.3389/fmicb.2024.1366181
Edited by:
Renqiang Yu, Women's Hospital of Jiangnan University, ChinaReviewed by:
Qiri Mu, Inner Mongolia International Hospital, ChinaCopyright © 2024 Liu, Yan, Jia, Huang, Fu, Xu, Yu, Yang, Pan, Zheng, Liu, Chen, Gao and Tian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ya Gao, eWdhb0B4anR1LmVkdS5jbg==; Donghao Tian, eWQxNzIwQHhqdHUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.