
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 05 January 2024
Sec. Phage Biology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1320345
This article is part of the Research TopicBacteriophage and Host InteractionsView all 15 articles
 Marta Gliźniewicz1*
Marta Gliźniewicz1* Dominika Miłek1Patrycja Olszewska1Artur Czajkowski1Natalia Serwin1Elżbieta Cecerska-Heryć1
Dominika Miłek1Patrycja Olszewska1Artur Czajkowski1Natalia Serwin1Elżbieta Cecerska-Heryć1 Barbara Dołęgowska1
Barbara Dołęgowska1 Bartłomiej Grygorcewicz1,2*
Bartłomiej Grygorcewicz1,2*Bacteria and fungi tend to coexist within biofilms instead of in planktonic states. Usually, such communities include cross-kingdom microorganisms, which make them harder to remove from abiotic surfaces or infection sites. Additionally, the produced biofilm matrix protects embedded microorganisms from antibiotics, disinfectants, or the host immune system. Therefore, classic therapies based on antibiotics might be ineffective, especially when multidrug-resistant bacteria are causative factors. The complexities surrounding the eradication of biofilms from diverse surfaces and the human body have spurred the exploration of alternative therapeutic modalities. Among these options, bacteriophages and their enzymatic counterparts have emerged as promising candidates, either employed independently or in synergy with antibiotics and other agents. Phages are natural bacteria killers because of mechanisms of action that differ from antibiotics, phages might answer worldwide problems with bacterial infections. In this review, we report the attempts to use bacteriophages in combating polymicrobial biofilms in in vitro studies, using different models, including the therapeutical use of phages. In addition, we sum up the advantages, disadvantages, and perspectives of phage therapy.
A biofilm is a structure composed of bacteria and other microorganisms (fungi, viruses) anchored in an extracellular matrix composed of organic substances produced by these microorganisms. Approximately 2–35% of the biofilm’s volume comprises microorganisms, while the matrix constitutes the remaining portion. Biofilm matrix primarily consists of proteins, lipids, polysaccharides, extracellular RNA and DNA, minerals, and ions suspended in water (Vu et al., 2009). Biofilm adheres to the biotic or abiotic surface. The biofilm’s structure may vary on homogeneous, composed of one species, or heterogeneous, consisting of many different strains of microorganisms. Biofilm is more challenging to eradicate than planktonic forms of microorganisms due to the protective properties of the matrix (Augustyniak et al., 2021). The properties of the biofilm enable pathogens to escape from the immune system, antibiotics, disinfectants, and other chemical substances (Jamal et al., 2018; Roy et al., 2018). Microorganisms regulate biofilm formation by expressing genes responsible for synthesizing and modifying extracellular components and communicating with each other by sending biochemical signals. This signaling network includes two-component systems (TCS), which regulate signal transduction via phosphorylation of cyclic di-GMP (c-di-GMP), diguanylate cyclase (DGC) systems which cooperate with TCS and coordinate the transition of bacteria from planktonic to biofilm growth mode, and quorum sensing (QS), mechanism that involves autoinducers which are small signal molecules and receptors (Guła et al., 2018). The signaling occurs interkingdom between microorganisms (bacteria, fungi) and host cells.
The main stages of biofilm formation are initial contact with a surface, irreversible contact with a surface, formation of microcolonies—expansion, maturation of the biofilm, and cell detachment of the individual cells from the matrix. Surfaces susceptible to bacterial adhesion encompass a variety of substrates such as sewage system pipes, soil particles, living tissues, and medical equipment (e.g., urological catheters, venous catheters, artificial heart valves, intrauterine coils, dental units, and contact lenses) (Vu et al., 2009; Stickler, 2014; Augustyniak et al., 2021). Additional cellular structures, such as fimbriae and flagella, bacterial proteins—adhesins, and physical forces, are responsible for the colonization. Environmental factors, such as the amount of available energy, surface structure, pressure, temperature, and orientation of bacterial cells, influence the possibility of adhesion to the substrate. The main physical forces involved in biofilm formation are van der Waals, steric, or electrostatic interactions associated with the cell membrane double layer (Delcaru et al., 2016).
Following the adhesion stage, there is a phase of microbial multiplication, leading to an expansion in the biofilm volume and the formation of a three-dimensional structure regulated by quorum sensing (QS). This mechanism relies on the secretion of proteins and autoinducers of the expression of genes coding for surface proteins, such as porins. This facilitates more effective nutrient absorption within the biofilm. The secretion of exopolysaccharides (EPSs), which stabilize the biofilm structure, also increases. Special channels are created in the entire biofilm volume to facilitate the removal of unnecessary metabolites and provide an appropriate gas environment and nutrients. Due to the static growth, the development of additional membrane structures responsible for the movement of bacteria is inhibited. In addition, a reduction in protease and phospholipase C synthesis, a decrease in the synthesis and release of toxins, and the production of rough and sometimes mucus-like polysaccharides to better adapt cells to specific conditions of the biofilm microenvironment are observed (Jamal et al., 2018; Narayanan et al., 2018; Amankwah et al., 2022).
The final phase of biofilm life occurs when the ratio of newly formed cells equals the number of dead ones. Environmental conditions such as oxygen depletion and nutrient unavailability result in the switching of bacterial metabolism. Enzymes (e.g., hydrolases and endonucleases) that break down the extracellular matrix, allowing individual bacteria to be released into the environment, are produced. In addition, the expression of genes leading to the formation of flagella returns, restoring the ability of bacteria to move and find a new location for biofilm expansion (Garrett et al., 2008).
Biofilm production by bacteria is related to their virulence and may imply the occurrence of chronic diseases in the host organism. This is related to many factors, e.g., the production of endotoxins or the protection of bacteria living in the biofilm against the mechanisms of the host immune system, such as phagocytosis or coating with antibodies (Roy et al., 2018). In addition, higher resistance to antibiotics is observed, which is associated with the problematic penetration of active drug ingredients through the biofilm, alternation in biofilm bacteria metabolic activity and presence of cells with a reduced metabolic activity called persister cells, multi-species biofilm, and facilitation of horizontal gene transfer (HGT) among bacteria (Ehrlich et al., 2010; Lehman and Donlan, 2015; Koo et al., 2017).
Biofilm eradication is an enlarging problem in medicine, agriculture, and the food industry. The Center for Disease Control and Prevention (CDC) estimates that even more than 65% of all chronic bacterial infections are caused by biofilm forms of pathogens (Amankwah et al., 2022). One of the novel antimicrobials is lactoferrin, mammalian transferrin with antimicrobial activity, which binds iron, preventing bacteria from using this metal. Another strategy is using molecules that inhibit the mechanisms of the QS system by suppressing signal generation, distribution or blocking signal receptors, and signal responses (Myszka and Czaczyk, 2010). The potential use of substances that influence the structure and work of efflux pumps, which are responsible for removing antibiotics from the bacterial cell, e.g., peptidomimetics, has also been investigated.
The possibility of using phages and phage-derived enzymes to combat bacteria in biofilm structures is also being explored. Furthermore, combination therapy using phages and/or phage-derived products with other antimicrobial agents, including antibiotics, nanoparticles, and antimicrobial peptides, is auspicious. Such a solution could be widely used in medicine to treat severe cases and the broadly understood industry (Herce-Ros et al., 2021; Srinivasan et al., 2021; Tanaka et al., 2021; Amankwah et al., 2022).
Mixed biofilms occur in many natural environments, e.g., the oral cavity, where many microorganisms form dental plaques, intestines, or vaginas. Certain multi-species biofilm-related diseases can arise when a single pathogen is introduced into an existing microbiome, leading to dysbiosis or when opportunistic pathogens become virulent due to environmental imbalances. Dysbiosis can develop gradually or rapidly and often leads to chronic destructive inflammation. Other situations occur when one pathogen first adheres to the infection site as first and prepares the environment for another. The initial pathogen that adheres to the surface may influence the subsequent bacterial cell selection and, consequently, the final composition of biofilm. It is called coaggregation and may occur when the secondary colonizer binds to specific molecules on the surface of a first one or several bacteria coordinate among themselves and favor some phenotypic changes that lead to the coaggregation on biofilms (Rickard et al., 2003; Peters et al., 2012; Szafrański et al., 2017). Moreover, due to the recruitment of a new species, the gene pool is broadened, and it helps control and regulate the survival mechanisms of individual members, such as adhesion, stimulation of host cellular senescence mechanisms to prevent the shedding of bacteria, and the production of plasma exudate for nutrition through local inflammation (Anju et al., 2022).
The interactions between microbes are complex and involve competition for space and nutrients. The biofilm community’s physiology and function often change and are regulated by various interspecies interactions. Bacterial species are organized into different spatial forms based on their type: interspecific segregation, coaggregation, and stratification (Liu et al., 2016; Anju et al., 2022). Microorganisms grouped in one community may act synergistically, antagonistically, or be indifferent to each other. Cooperation between bacteria facilitates their adhesion and growth of, resistance to antimicrobial agents, virulence, exopolysaccharide production, and protective properties of the whole biofilm.
Moreover, the exchange of nutrients and metabolic products may occur in some species’ relationships. For example, Fusobacterium nucleatum and Prevotella intermedia produce ammonia, which increases the pH and creates an environment suitable for the growth of Porphyromonas gingivalis. Another example is Pseudomonas aeruginosa, which produces substances that protect Staphylococcus aureus from aminoglycosides (Wolcott et al., 2013; Anju et al., 2022). The opposite behavior is observed when antagonistic interaction occurs. Then, one microorganism inhibits or kills the competing species, ensuring itself to avail available space, energy sources, and nutrients. Competition can be exploitative and involves the superiority of energy utilization or interference that produces compounds preventing other species’ growth (Mgomi et al., 2022).
Multi-species biofilms can also be characterized by the distribution of microorganisms within the matrix. Microorganisms may coexist in separate microcolonies, with limited interactions, in one style of organization. Another style is characterized by a thoroughly mixed arrangement where cells from different species randomly coexist throughout the biofilm. One species forms the bottom layer in the third organizational structure, while the second species places on top (Mgomi et al., 2022). Another scheme is frequently observed in bacteria–fungi biofilms where hyphae form a scaffold that carries bacteria cells (Bernard et al., 2020; Roszak et al., 2022). Different structures of biofilms generate different interactions between species and mechanisms of cellular responses for therapies.
Biofilm-related chronic infection is frequently polymicrobial. Coexisting in multi-species communities increases genetic material exchange between cells, metabolic cooperation, development of antibiotic resistance, niche optimization, host immune system modulation, and virulence induction (Kifelew et al., 2019; Mgomi et al., 2022). Creating a standard matrix on tissues or medical devices is a characteristic of population virulence, making the behavior of polymicrobial societies distinct from mono-species. These societies can alter their physical properties in response to the environment and evolve through mutation to better adapt to their surroundings (Ehrlich et al., 2010). Moreover, additional pathogens can be integrated into the biofilm, and the primary ones can mutate to improve the interaction with other resident species, producing a more stable and productive community. All these properties cause more severe disease symptoms than mono-species infections.
One of the biggest problems associated with polymicrobial infection is increased resistance to antimicrobial agents, which might be higher than in mono-species biofilm. It is caused by the extensive diversity of EPSs produced by heterogeneously distributed bacteria that disturb drug penetration (Topka-bielecka et al., 2021). Moreover, some bacteria and fungi can produce polysaccharides or other substances that protect themselves or cells of partner species from antibiotics and antifungal agents. Another threat is interspecies HGT, which results in gene exchange between evolutionarily distant species. This may create bacteria and fungi with different phenotypes with new features that may increase their virulence and drug resistance.
Biofilm-associated polymicrobial communities are responsible for many diseases, e.g., bone infections and osteomyelitis, gall bladder disease, various chronic middle-ear disease processes, and chronic rhinosinusitis, chronic infections of the urogenital systems, e.g., bacterial vaginosis, dental infections, tonsillitis, surgical site infections, chronic non-healing wounds such as venous and diabetic ulcers, pressure sores, and burn injuries, respiratory infections, e.g., cystic fibrosis and medical device-related infection (Ehrlich et al., 2010; Peters et al., 2012; Szafrański et al., 2017; Iszatt et al., 2021; Uyttebroek et al., 2021). Some of these diseases were subjected to phage therapy. In addition, many scientists investigated various possibilities for phage treatment in in vitro research.
Bacteriophages (phages) are viruses that infect bacteria and cannot multiply outside their host cells. Phages were discovered independently by Frederick Twort and Felix d’Hèrelle over 100 years ago and are the most diverse and numerous life forms on the earth. They show high host specificity, recognizing their host at the species and even strain level due to presence of characteristic receptors on the surface of the bacterial cell (Drulis-Kawa et al., 2015; Atshan et al., 2023). The use of bacteriophages is extensive. It covers many areas of life, e.g., medicine and veterinary (phage therapy), food industry (disinfectants of surfaces), agriculture (plant growth promoters), biotechnology and pharmacy (nanocarriers of drugs, biosensors, or diagnostic molecules), and diagnostic (phage typing) (Cowley et al., 2015; Drulis-Kawa et al., 2015). Currently, phages are classified by the European Union (EU) as medical products and by the Food and Drug Administration (FDA) as drugs. Good manufacturing practice (GMP) must be implemented during phage particle production. Nevertheless, all clinical trials are conducted as a therapy of a last chance according to Article 37 of the Declaration of Helsinki and need the permission of the ethical commission. The preparation of consistent legislation regarding the usage of phages in medicine is still ongoing (Patey et al., 2019).
The rate at which bacteria acquire antibiotic resistance is alarming, and the current epidemiological situation requires the search for alternative methods of combating bacterial infections. One of the options is to use bacteriophages (Iszatt et al., 2021). The usage of phages has many advantages, e.g., rapid clearance from organisms, self-propagation in the site of infection, host specificity, opportunity to make a genetic modification, easy isolation, stability, and relatively low-cost production (Łubowska et al., 2019; Mgomi et al., 2022). The right phage must be selected carefully because not all have good therapeutic results. When choosing phages, some rules should be followed: specificity to target bacteria, lytic activity, and the lack of genes encoding bacterial virulence factors, antibiotic resistance products, and toxins. Only fully sequenced bacteriophages can be used for treatment in medicine. Another concern about phage therapy is to optimize the dosage of virions and the method of administration to provide good delivery to the site of infection (Morrisette et al., 2019). The pharmacokinetics of phages are complicated due to their ability to self-replicate. After killing all pathogenic bacteria, the phages are removed from the body as they cannot multiply in eukaryotic cells. In addition, if selected carefully, phages are safe for the human microbiome (Sartini et al., 2021). On the other hand, they may be neutralized by the host’s immune system, removed from the body too quickly, and bring no profit. Another issue from the immune system might be an allergic reaction that may limit the scope of possible use of bacteriophages. Unfortunately, bacteria have developed multiple resistance mechanisms to phages (e.g., modification and blockage phage receptors on the bacterial surface), and even though phages have an equally impressive assortment of tools to overcome this resistance, it is better to use a cocktail of phages (Chegini et al., 2021). Notably, phage resistance observed in vitro may not necessarily translate to in vivo conditions. This discrepancy arises from the fact that the most prevalent resistance mechanisms often involve alterations in the cell surface that untenable host infection by the phage (Park et al., 2014; Iszatt et al., 2021). Another issue is that phage therapy could release bacterial endotoxins during bacterial cell lysis, which occurs as an effect of phage infection.
Next, to phages themselves, lytic enzymes that they produce are also considered as treatment factors. Bacteriophages synthesize enzymes such as peptidoglycan hydrolases, holins, and endolysins, which allow to release progeny virions by destroying bacterial cells walls (Sousa et al., 2023). Based on their mechanism of action, we can divide them into hydrolases and lyases. Both groups can degrade polysaccharides, including capsular polysaccharides (CPSs), lipopolysaccharides (LPSs), O-polysaccharides, or exopolysaccharides (EPSs), and sometimes polypeptides and lipids (Topka-bielecka et al., 2021). Endolysin can induce lysis from within as an antimicrobial agent active against pathogens. This refers mainly to Gram-negative bacteria, which need to treat with additional factors, e.g., holins that allow the endolysin to move through the cytoplasmic outer membrane and reach the peptidoglycan layer (Mgomi et al., 2022). Other phage enzymes—depolimerases, can be tail-spike proteins with the enzymatic domain or occurring as free molecules. Phage-delivered enzymes are usually unique and species-specific. However, sometimes enzymes can show activity across a broad spectrum between strains and species (Chegini et al., 2021). Treatment of polymicrobial communities usually requires several different enzymes or combined therapy. Bacteria rarely evolve resistance to lysins because they attack sites on the peptidoglycan cell wall critical to bacterial viability. Nevertheless, combining phage lysins and antibiotics, phages and other agents, or the production of genetically engineered enzymes may be more effective in infection elimination.
Lytic phage can be an effective weapon in the fight against biofilm, both in the context of preventing its formation and its eradication. The attempts to use phages as prevention factors include coating urinary catheters and disinfectants in hospital or industry environments (Curtin and Donlan, 2006; Lehman and Donlan, 2015; Melo et al., 2016; Santiago and Donlan, 2020). The activity of phages in mature biofilm depends on the bacteria growth phase, placement, coaggregation with other cells, nutrient availability, access to receptors, and diffusion capacity. Phages can be used not only in biofilms of multi-bacteria species but also in bacteria–yeast ones. P. aeruginosa infecting phage Pf4 can inhibit Candida albicans biofilm formation, possibly by sequestrating iron (Nazik et al., 2017; Pohl, 2022). Phages act differently from antibiotics. They produce enzymes, e.g., depolymerases (DP), that can destroy biofilm matrices made of polysaccharides, including EPS or alginase, breaking down the alginate matrix produced by P. aeruginosa (Peters et al., 2012; Santiago and Donlan, 2020). Moreover, phages can stimulate the host bacteria to produce EPS-degrading enzymes and proteases that degrade bacterial capsules. Phages oppositely to antibiotics can degrade bacteria that manifest low metabolic activity due to nutrient depletion. Another mechanism that helps overcome the matrix is to diffuse through water channels or to adsorb to motile bacteria and “have a ride” to the target site (Kifelew et al., 2019; Amankwah et al., 2022; Atshan et al., 2023). These actions collectively enhance the effectiveness of phages in combating biofilms. Initially, they facilitate the penetration of phages, allowing entry into the biofilm for subsequent replication within bacterial cells. The elevated bacterial density within biofilms significantly amplifies phage infection, leading to the release of new virions. Even when targeting cells with reduced metabolic activity, lytic phages prove effective by releasing intracellular materials. This release stimulates bacterial metabolism, ensuring sustained efficacy (Amankwah et al., 2022). Basting an expanded host range, polyvalent phages emerge as valuable assets in disrupting polymicrobial biofilms.
Phages also exhibit adaptability during isolation, potentially enriched by employing multiple bacterial hosts rather than a singular one. Additionally, an alternative strategy involves leveraging phages as quorum quenchers. Some phages eliminate bacteria conventionally and produce enzymes that disrupt bacterial signal molecules, providing a multifaceted approach to biofilm intervention (Kifelew et al., 2019; Santiago and Donlan, 2020).
Microbial communities have mechanisms of protection from phages that affect phage ability to adsorb, penetrate, diffuse, and proliferate in biofilm. The ability of biofilm to resist phage invasion depends on its age, shape, structure, and morphology. Bacteria can evolve to be insensitive to phage by changing their phenotypes in response to heterogeneous environments. P. aeruginosa may transform into a pili-defective variant to avoid infection of phages that use these structures as their receptors (Yamamoto et al., 2021). Biofilm matrix comprises many bacterial enzymes, e.g., amidases and peptidases, that may inactivate phages. Moreover, in deeper biofilm layers, more dead cells occur, and phages may adsorb to them without any benefits for therapy. Molecules can also catch virions in the matrix (Pires et al., n.d.). One of the ways of bacteria defense is the production of systems that interfere with phage nucleic acids, e.g., clustered regularly interspaced short palindromic repeats (CRISPR)–Cas9 (Yang et al., 2020). Bacteria may also prevent phage DNA integration by a superinfection exclusion system or use an abortive infection system to block the synthesis of phage particle compounds (Pires et al., n.d.). Another protection is related to hiding binding phage receptors by the production of curli polymer (CsgA) as extracellular fibers that curtain bacterial cells (Vidakovic et al., 2017; Santiago and Donlan, 2020).
To intensify the action of phages, they can also be used with various groups of antibiotics (phage-antibiotic synergy (PAS) phenomenon). However, not every drug acts synergistically with selected phages and every combination should be checked in in vitro studies. For example, the synergistic effect may arise from the stimulation of lytic phage development in the presence of beta-lactam antibiotics. Bacteria under beta-lactam stress change their morphology, facilitating phage assembly and increasing bacterial sensitivity to phage lysins (Comeau et al., 2007; Chegini et al., 2021). Another mode of action of phages is to interfere with bacterial efflux pumps, which increases the sensitivity to various drugs (Chan et al., 2016). Bacteriophages can also be combined with disinfectants such as chloride, hypochlorite, or quaternary ammonium compounds and enzymes, e.g., polysaccharides depolymerases. In addition, in the case of a biofilm with a heterogeneous structure, it is possible to use a phage cocktail composed of several bacteriophages showing bacteriolytic activity against various bacterial pathogens (Comeau et al., 2007). Another alternative to enhance phage penetration through the matrix is debriding biofilm mechanically before phage treatment (Pires et al., n.d.).
Another way to improve phage performance is to modify their genome or synthesize novel ones (Javed et al., 2019). Modified phages may have inserted gene coding for additional exopolysaccharide-degrading enzymes for better biofilm penetration. Bacteriostatic phages can be changed to suppress the DNA repair mechanism, or overexpression of sensitizing proteins, and to disrupt the cell–cell communication between the bacteria in the biofilm. Another target for phage engineering is to use phages as a modulators of antibiotic resistance or to make it possible to reach intracellularly bacteria present in eukaryotic cells (Hagens et al., 2004; Lu and Collins, 2007, 2009; Edgar et al., 2012; Pei and Lamas-Samanamud, 2014). Since sometimes bacterial lysis leads to release toxins and pro-inflammatory products, phages can be engineered to be toxic for bacteria but not lytic for their host (Szafrański et al., 2017). The possibilities of degrading polymicrobial biofilm using phage-mediated methods are presented in Figure 1.

Figure 1. Methods of disturbing polymicrobial biofilm by phage-mediated methods.
Bacteriophages should be recognized as a potent tool against pathogenic bacteria and integral components of healthy microbiomes, including those in the oral, intestinal, or vaginal environments. Phages interact with commensal bacteria, fungi, and chemical compounds and contribute to microbial communities assembly, stability, and function. They contribute to biofilm formation as extracellular DNA release through phage-mediated cell lysis may induce mobile genetic element transfer between microbes which, in turn, triggers a response of stabilizing the biofilm matrix (Amankwah et al., 2022). Some phages and their hosts developed reciprocal predator–prey relationships, e.g., in the intestine, phages may promote the evolution of bacterial resistance to phages in response to infection (Duerkop, 2018). Furthermore, phages may bind to mucin glycoproteins, providing phage-mediated antibacterial protection of animal mucosal surfaces (Barr et al., 2013).
Bacterial and phage composition in the intestine depends on diet and may drastically change during infection or other diseases. Increased or changed phage dsDNA levels were noticed during inflammatory bowel disease and type 1 diabetes in children (Zhao et al., 2017; Duerkop, 2018). Phages may stably multiplicate in their host for weeks but not lead to the elimination of pathogens. For example, enteroaggregative Escherichia coli (EAEC) and its phages may coexist without resolution, linked to persistent colonization and prolonged diarrhea (Maura et al., 2012). Beyond the intestine, in different niches, the phageome (bacteriophage community in the niche) of the bacterially infected site may be distinct from the healthy one, e.g., during cystic fibrosis (CF) (Reece et al., 2021). Phages, as a component of the polymicrobial community, may cooperate with the mammalian immune system and actively eliminate bacteria from the lungs during infection. Oppositely, P. aeruginosa prophages can stabilize biofilm in the lungs by promoting attachment to lung mucus and restricting the dispersal of cells from the biofilm. Moreover, phages may decrease the pro-inflammatory response of the immune system and lead to chronic infection (Duerkop, 2018). In another niche, lysogenic phages may modulate the number of vaginal lactobacilli during bacterial vaginosis (Jung et al., 2017). In addition, the taxonomic composition of phages may change during infection, e.g., chronic wound virome is more diverse than contralateral skin, which may influence microbial community and impact healing outcomes (Verbanic et al., 2022).
Numerous researchers explore phage therapy as a potential solution in the era of limited options for treating antibiotic-resistant bacterial infections. Various approaches are investigated, for example, using phages to eradicate mono-and multi-species biofilm, prevent biofilm formation, or change the composition of multi-species biofilm by removing only the harmful species. The in vitro studies implementing phage therapy and the research results employing various models are summed up in Table 1.
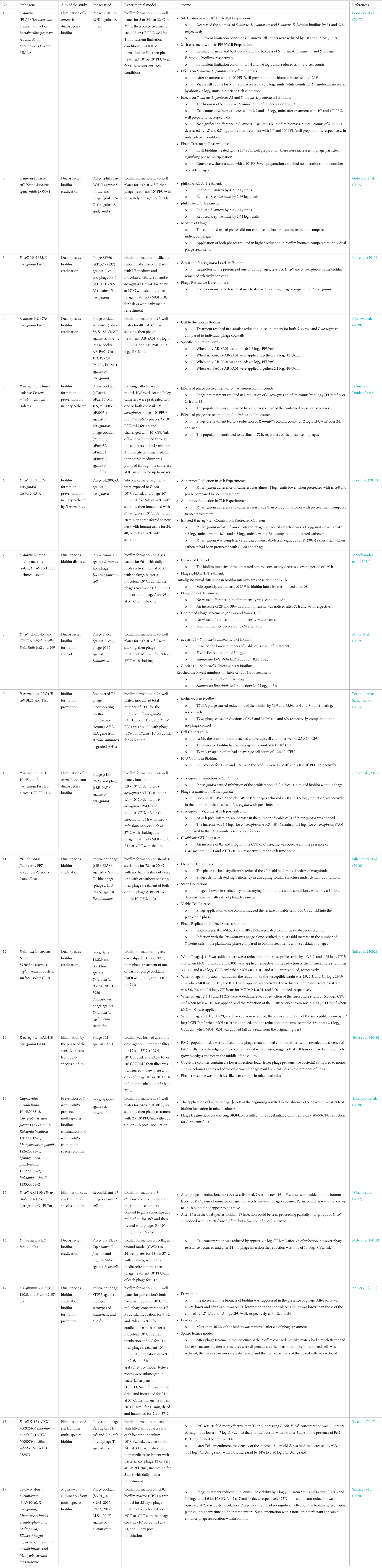
Table 1. Examples of phage treatment of polymicrobial biofilms in in vitro and in vivo studies with models.
Even though phages occurred to be effective to some point in biofilm-forming prevention and eradication of mature polymicrobial biofilm, some limitation of phage therapy must be overcome to achieve fully effectiveness. The main problems are acquiring phage resistance by bacteria in polymicrobial biofilm, reaching target bacteria in this complex structure, or selecting phages for all pathogens in biofilm. The solution might be combined therapy based on phages mixed with antibiotics, nanoparticles, other substances, or using phage-delivered enzymes with different properties than phages.
Different approaches to using phage-antibiotic synergy (PAS) therapy are listed in Table 2. The selection of good phage-antibiotic pair is strictly individual to the bacterial strain and case (Grygorcewicz et al., 2023). However, the effort is worth it and brings better results than monotherapy. Phage may prevent the development of antibiotic-resistant minority bacterial populations, and conversely, antibiotics may stimulate phage infection, changing the phenotype of the target host, and phages may interfere with drug-resistant mechanisms, making bacteria more vulnerable (Comeau et al., 2007; Chan et al., 2016; Dickey and Perrot, 2019; Chegini et al., 2021). In many cases, PAS is necessary for successful therapy due to its better ability to degrade biofilm complex. At the same time, more than one factor is applied and all of them present different mechanisms of action (Roszak et al., 2022). Phages are considered more effective against biofilm due to the production of polysaccharide depolymerases which loosen matrix structure and help antibiotics reach the bacteria cell surface. Another mode of action is the lysis of cell from exterior parts of biofilm which results in uncovering the deeper layers of cells, and giving them access to nutrients and oxygen. This makes bacteria more metabolically active and more susceptible to an antibiotic (Park et al., 2017). The effectiveness of such therapy also depends on the dosage of antibiotics and phages, time and order of administration, adsorption rate, burst size, latent period, and external physical factors such as pH and temperature (Morrisette et al., 2019).
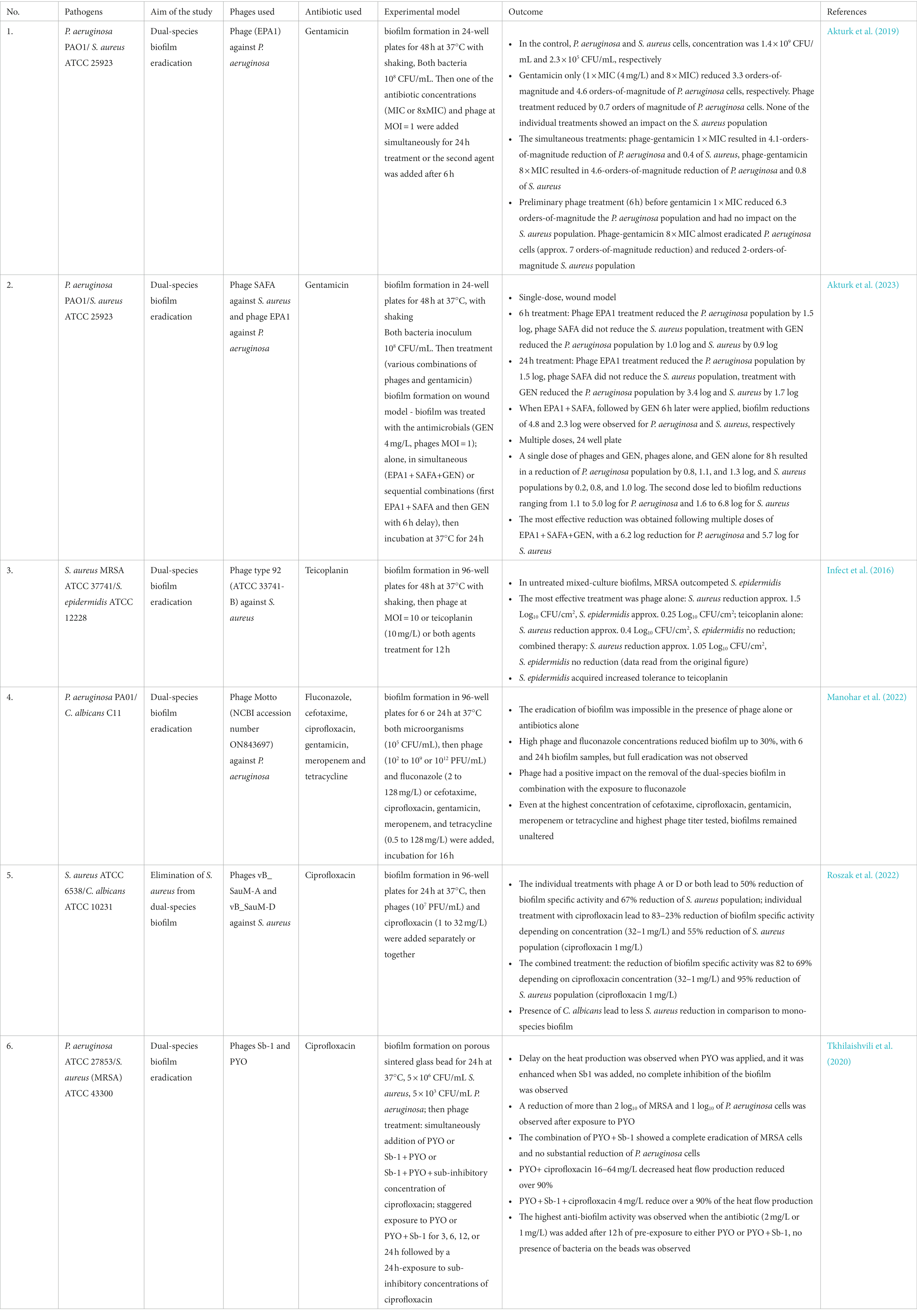
Table 2. Examples of phage-antibiotic synergy therapy in combating polymicrobial biofilm in in vitro studies.
Another approach is to combine phages with nanoparticles. That solution may enhance phage penetration through biofilm. Moreover, nanoparticle migration in matrix might be modulated by a magnetic field in ex vivo models. Li et al. (2017) investigated how polyvalent phages (PEL1) immobilized onto Fe3O4-based magnetic colloidal nanoparticle clusters (CNC) coated with chitosan (PEL1-CS-Fe3O4) penetrate P. aeruginosa/E. coli dual-species biofilm. The complex penetration was facilitated under a small magnetic field (660 gauss), leading to better plaque formation capability of PEL1 and removal of 88.7 ± 2.8% of the biofilm formed on a glass surface after 6 h of treatment. The usage of such a particle complex physically disrupts the biofilm and mitigates phage dilution, which, in turn, allow to keep a high concentration of phages and facilitate phage tail fibers exposition to the hosts (Li et al., 2017). Another study where phages were covalently conjugated with magnetic CNCs shows that this approach is noteworthy. Yu et al. (2019) used phages PEB1 or PEB2 conjugated with CNCs of different sizes to combat P. aeruginosa/ E. coli dual-species biofilm and P. aeruginosa/E. coli/B. subtilis and Shewanella oneidensis multi-species biofilm. Smaller complexes disrupted the biofilm bottom layer and detached the biofilm within 6 h with efficiency of 98.3 ± 1.4% for dual-species biofilm and 92.2 ± 3.1% for multi-species biofilm. Larger complexes were less effective, implying that the size of nano-phage complex matters (Yu et al., 2019). It was reported that magnetic field might influence bacteriophage development. Phages T4 for E. coli and vB_SauM_A for S. aureus exposed to a rotating magnetic field enhance their adsorption and propagation rate (Struk et al., 2017; Konopacki et al., 2020; Grygorcewicz et al., 2022). In addition, a magnetic field might modulate the metabolism of bacteria and other microorganisms (Jabłońska et al., 2022).
In addition to antibiotics and nanoparticles, other chemical compounds or groups of compounds might be combined with phages to minimize formation and eradicate polymicrobial biofilm. Chhibber et al. (2015) tested how bacteriophages combined with xylitol will eradicate K. pneumoniae/P. aeruginosa dual-species biofilm formed on polycarbonate disks. K. pneumoniae-specific depolymerase-producing phage KPO1K2 and P. aeruginosa specific non-depolymerase-producing phage Pa29 led to 2.13 and 1.27 log10 CFU/mL reduction of K. pneumoniae and P. aeruginosa cell counts, respectively in 1-day-old biofilm. They obtained slightly worse results for 2-day-old biofilm. The authors emphasize that depolymerase-producing phage was crucial for matrix disruption. The addition of xylitol to the system significantly enhanced the antibiofilm activity of phages and caused complete elimination of K. pneumoniae both in 1- and 2-day-old biofilms and also 3.5 and 3.02 log10 CFU/mL reduction of P. aeruginosa in 1- and 2-day-old biofilm, respectively. Xylitol may diffuse into the biofilm and accumulate as a toxic, non-metabolizable sugar alcohol phosphate, thus inhibiting bacterial growth, or it can hinder stress proteins that arise in the biofilm (Ichikawa et al., 2008; Chhibber et al., 2015). An interesting approach was presented by Oliveira et al. (2018), who used chestnut honey bacteriophages (vB_EcoS_CEB_EC3a and vB_PaeP_PAO1-D) against P. aeruginosa/E. coli dual-species biofilm formed on polystyrene and porcine skin. Honey has antimicrobial properties associated with high osmolarity, low availability of water, hydrogen peroxide production, acidic pH level, and the presence of methylglyoxal. The results of using different combinations of phage and honey showed that E. coli cell number reduction in biofilm depends on the applied treatment time and honey concentration. In the case of P. aeruginosa, combined treatment brought better results than phage or honey alone, however, without presenting a synergy effect on the polystyrene model. E. coli elimination from dual-species biofilm formed on porcine skin model was the most effective using phage and 50% honey and led to 1.4 log reduction at 24 h post-treatment. The combination of phage and honey acts synergistically in P. aeruginosa cell elimination at both concentrations (25 and 50%), leading to 2.2 log10 and 2.3 log10 higher cell reduction than the sum of phage and honey alone (Oliveira et al., 2018).
Phage-delivered enzymes are the next option to eliminate polymicrobial biofilm. Their main advantages are host specificity and easy matrix penetration and removal. Skillman and Sutherland (1999) proposed the usage of polysaccharide depolymerases isolated from a bacteriophage infecting E. agglomerans to degrade EPS in a dual-species biofilm formed with K. pneumoniae. Such treatment caused limited adhesion of E. agglomerans to K. pneumoniae, degradation of EPS, and effective removal of both species from the surface, even though the used enzyme was specific toward E. agglomerans only. This effect might have been caused by the proximity of both species or the larger contribution of E. agglomerans EPS in the mixed biofilm. Schuch et al. (2017) used bacteriophage lysin CF-301 and combined it with lysostaphin to target S. aureus and S. epidermidis in mixed biofilm formed on various surfaces (polystyrene, surgical mesh, and catheters). Dual-species biofilm was susceptible to disruption by CF-301 applied at concentrations down to 0.032 mg/L over 24 h. The reduction of both species on catheter and surgical mesh reached over 90% and over 80% on 24-well polystyrene plates. The good enzymatic activity against both species is reasonable because they belong to the same genus. However, more than one protein should be used when more phylogenetically distant species form a mixed biofilm. This approach was investigated by Manoharadas et al. (2023), who used two engineered enzybiotics (BP404 5 mg/L and P16-17/100 5 mg/L) against a dual-species biofilm formed by S. aureus and E. faecalis in an inert glass surface. The chimeric protein P16-17/100 was constructed, linking domains from endolysin P16 and minor tail protein P17 from phage φ44AHJD. Protein cocktail usage resulted in significant biofilm dispersal (absorbance OD575 reduction from 0.7 to less than 0.1) and more than 90% reduction of both species cells embedded in the matrix after 16 h of treatment.
Bacterial vaginosis (BV) is a common vaginal infection caused by anaerobic pathogens such as Gardnerella vaginalis, Fannyhessea vaginae, and Prevotella bivia, usually forming a polymicrobial biofilm. Therapy of BV usually relies on metronidazole and clindamycin treatment. However, sometimes, these antibiotics do not lead to the complete eradication of pathogens. The curation of biofilm-associated BV is challenging. Therefore, Landlinger et al. (2021) generated engineered endolysin Pm-477 encoded on Gardnerella prophages as an alternative treatment. The endolysin actively killed G. vaginalis in mono- and dual-species communities with Lactobacillus crispatus. Moreover, the efficacy of PM-477 was tested by fluorescence in situ hybridization on vaginal samples of 15 women with BV. Endolysin eliminated Gardnerella bacteria in 13 cases and physically dissolved the biofilm matrix. The remaining vaginal microbiome remained unaltered. Castro et al. (2022) also tested previously synthesized engineered phage endolysin PM-477 to disrupt dual-species biofilms composed of G. vaginalis/F. vaginae or G. vaginalis/P. bivia in in vitro study. In all dual-species biofilms, endolysin prevented biomass accumulation (from 24 to 48 h) but did not reduce existing ones. In G. vaginalis/F. vaginae, biofilm cell viability reduction was not obtained, but in G. vaginalis/P. bivia, biofilm reduction reached a 3 log10 CFU. The phage endolysin had high anti-G. vaginalis and slightly anti-P. bivia but no anti-F. vaginae activity.
Johnston et al. (2023) also investigated how endolysin therapy against G. vaginalis biofilm works in vitro. In their study, a four-species biofilm made of G. vaginalis, F. vaginae, P. bivia, and Mobiluncus curtisii was treated using an anti-Gardnerella endolysin (CCB7.1) as this species is the most abundant in polymicrobial community. The reduction of live cells of G. vaginalis reached 1–2 log10 after 24 h of endolysin treatment in all tested concentrations (128, 256, and 512 μg/mL) and a slight reduction of M. curtisii when the highest concentration of endolysin was applied. Worth mentioning is that CCB7.1 was ineffective against commensal lactobacilli. Novel endolysins against G. vaginalis are still being searched. Arroyo-Moreno et al. (2022) identified 84 diverse anti-Gardnerella endolysins and selected 5 (CCB2M94_8, CCB7.1, CCB8.1, CCB2.2, and CCB4.1) with the best properties. All of them could disturb G. vaginalis/Atopobium vaginae dual-species biofilm in the concentration of 200 μg/mL and had no activity against commensal lactobacilli.
In vitro studies provide valuable data about phage therapy efficiency against polymicrobial biofilms. Research shows that phages themselves or in combination with antibiotics or other substances can reduce biofilm formed on various surfaces, e.g., polystyrene, glass, stainless steel, or silicone (urine catheters) (Curtin and Donlan, 2006; Carson et al., 2010; Kaźmierczak et al., 2022). Promising results from in vitro studies allowed to start more comprehensive clinical trials using bacteriophages. Even though in some countries (Georgia, Russia, Poland) phage therapy has been used for many years, the Western world has only recently started the first attempts to treat patients with phages. Reported cases when phages or their enzymes were included in therapy refer to wound infections, bone infections, surgical site infections, etc.
Bone and joint infections are the hardest to cure and are usually related to post-traumatic or implant infections. Bacteria quickly form polymicrobial biofilm and can persist in osteoblasts or synovial cells, implicated in chronicity and recurrence, usually requiring heavy surgery with implant exchange. Bacteria mainly isolated from bone infections are S. aureus, coagulase-negative staphylococci, Cutibacterium acnes, Streptococcus spp., Enterobacteriaceae, and P. aeruginosa (Ferry et al., 2021). When antibiotics fail, phage therapy is proposed to patients.
Nir-Paz et al. (2019) successfully treated a 42-year-old male patient with a trauma-related left tibial infection caused by extensively drug-resistant Acinetobacter baumannii and multidrug-resistant K. pneumoniae. Patient with trauma was first treated with external fixation, irrigation, and debridement, plus left leg fasciotomies and a prolonged course of antibiotics: 6 weeks of piperacillin/tazobactam, initially followed by an 8-week course of meropenem and colistin. After 7 months of unsuccessful therapy, phages were included. The patient received a phage cocktail (ɸAbKT21phi3 and ɸKpKT21phi1 in concentration 5 × 107 PFU/ml each), colistin (4.5 × 106 units/bid), and meropenem intravenously. The first effect of curation was visible after a few days, and 8 months post-treatment, no bacteria were detected. Phage-antibiotic therapy saved the patient’s leg from amputation. Onsea et al. (2019) provide further instances of successful phage therapy. The group developed a protocol for intraoperative phage application and postoperative use of a draining system. They reported three successful curation of patients with polymicrobial bone infection: Patient 1 (infection: the trauma of pelvis; bacteria: P. aeruginosa, S. epidermidis; antibiotics used beside phages: for 3 months, vancomycin, rifampicin, moxifloxacin; phage therapy: for 7 days, BFC1 phage cocktail contains phages against S. aureus and P. aeruginosa 107 PFU/mL); patient 2 (infection: the trauma of femur; bacteria: P. aeruginosa, S. epidermidis; antibiotics used in addition to phages: for 6 weeks, vancomycin, colistin, fosfomycin; phage therapy: for 10 days, BFC1 phage cocktail); patient 3 (infection: trauma of femur; bacteria: S. agalactiae, S. aureus; antibiotics used in addition to phages: for 3 months, vancomycin, clindamycin, moxifloxacin; phage therapy: for 9 days, BFC1 phage cocktail). After 8 or 16 months, no signs of infection were observed (patients 1 and 3), and patient 2 needed further treatment. Van Nieuwenhuyse et al. (2021) report the case of a 13-year-old patient who developed chronic polymicrobial biofilm infection of a pelvic bone allograft. Clostridium hathewayi, P. mirabilis, Finegoldia magna, and methicillin-susceptible S. aureus were isolated from the infectious site. Conventional therapy (intravenous antibiotics and surgical debridement) with anti-S. aureus phage treatment (BFC1 phage cocktail) in situ was implemented. At first, therapy led to marked clinical and microbiological improvement, but it failed to prevent a recurrence of infection later.
Difficult to treat and chronic bacterial infections can occur at different sites of infection. They are born by various bacteria that cannot be eliminated with antibiotic therapy due to the formation of polymicrobial biofilm and the possibility of cell survival (Morozova et al., 2018). Phage therapy was proposed in many cases, referring to polymicrobial infections. Püschel et al. (2022) reported a case of successful treatment of drive line infection acquired after left ventricular assist device (LVAD) implantation with a combination of antibiotics, debridement, and local bacteriophage treatment. P. mirabilis and S. aureus were isolated due to unsuccessfully treated surgically for a driveline phage therapy was used. Phage cocktail containing phages against E. coli, S. aureus, P. aeruginosa, S. pyogenes, P. vulgaris, and P. mirabilis (107 PFU) was applied to the site of infection. The wound was healing well; the patient received cotrimoxazole for 20 days. Afterward, only S. aureus was detected in the infection site, and further flucloxacillin treatment was applied. In a follow-up examination 8 months later, the primary site of infection was free from bacteria.
Another example might be considered a success. However, the patient died long after phage therapy. Rubalskii et al. (2020). present a case of 52-year-old patients with a prosthetic infection after aortic arch replacement. Implant drainage and bronchial lavage were infected with S. aureus, E. faecium. P. aeruginosa, and E. faecium. Following the ineffectiveness of antibiotic therapy, a combination of phages (108 PFU/mL of Staphylococcus phage CH1, Enterococcus phage Enf1, Pseudomonas phage PA5, and Pseudomonas phage PA10), was applied in combination with two applications of gentamicin and daptomycin locally during the intraoperative phase, and a long-term intravenous application of cefepime, daptomycin, linezolid, and tobramycin was employed. After the intervention, S. aureus, E. faecium, and P. aeruginosa were undetected. However, the patient died after 2 months due to a new bacterial infection.
A research group from Eliava Phage Therapy Center, Tbilisi, Georgia (Nadareishvili et al., 2020) presents cases of successful phage treatment of polymicrobial infection related to biofilm (Nadareishvili et al., 2020). Patient 1, a 69-year-old male patient with a diabetic foot ulcer, was infected with following bacteria: Burkholderia cepacia, S. aureus, and E. faecalis. The staphylococcus phage and Intesti bacteriophage cocktail (consisting of Shigella spp., Salmonella spp., E. coli, Proteus spp., S. aureus, P. aeruginosa, and E. faecalis phages) were applied daily in the site of infection and orally for 40 days. The size of the wound was reduced after a few weeks; in addition, there was no recurrence after 1 year of treatment. Patient 2, a 68-year-old male patient with a postsurgical infection (after skin graft surgery), had two infections: the first infection was mono-species, and the second one was caused by S. aureus and Serratia marcescens. After the application of staphylococcus phage daily at the site of infection and orally for 3 months, the infection was resolved, and the tissue healed completely. Another example is reported in cooperation with Johri et al. (2021) group. Patient with chronic bacterial prostatitis (CBP) infected by methicillin-resistant S. aureus (MRSA), Staphylococcus haemolyticus, E. faecalis, and Streptococcus mitis was first unsuccessfully treated with antibiotics. Then, Pyo (a cocktail of phages against Streptococcus spp., Staphylococcus spp., E. coli, P. aeruginosa, and Proteus spp.) and Intesti bacteriophage cocktail, combined with additional Staphylococcal phage, was introduced in three forms: oral liquid, rectal suppositories, and urethral installations. After 5 days of therapy, the patient’s body temperature normalized. The therapy was prolonged, and isolated from patient bacteriophage against S. mitis was included. After almost a year, in a follow-up examination, semen and expressed prostatic secretion were free from bacteria, and the prostate was small and firm by rectal palpation.
In conclusion, bacteria and other microorganisms prefer to organize themselves in multi-species communities. Such biofilms are difficult to cure using antibiotic therapy and to remove from abiotic surfaces. Due to the persistence of multi-species biofilms, alternative methods of their eradication are being developed. Bacteriophages are one of the solutions. Studies have been conducted using both wild-type and genetically modified or polyvalent phages. In addition, they can be successfully used in combination with antibiotics or other chemical molecules. A separate group consists of enzymes and modified enzymes produced by phages. All these methods allow for better penetration of the biofilm matrix and reaching the surface of the target bacterial strains. The use of phage therapy is also increasingly used in medicine in the treatment of severe multi-species infections. However, the routine use of bacteriophages in medicine still requires a lot of research, including optimization and legislative work. However, despite further work required, bacteriophages and therapies using them to any extent are the future in treating bacterial infections. These viruses are and will be increasingly used to prevent bacteria in the hospital environment and other cases, e.g., in the food industry, veterinary medicine, or agriculture.
MG: Conceptualization, Data curation, Funding acquisition, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review & editing. DM: Writing – original draft. PO: Writing – original draft. AC: Writing – original draft. NS: Writing – original draft. EC-H: Writing – original draft. BD: Writing – original draft. BG: Conceptualization, Formal analysis, Funding acquisition, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the National Science Centre, Poland (PRELUDIUM 19, Project No. 2020/37/N/NZ9/02947).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Akturk, E., Melo, L. D. R., Oliveira, H., Crabbé, A., and Coenye, T. (2023). Biofilms 6. doi: 10.1016/j.bioflm.2023.100147
Akturk, E., Oliveira, H., Santos, S. B., Costa, S., Kuyumcu, S., Melo, L. D. R., et al. (2019). Synergistic action of phage and antibiotics: parameters to enhance the killing efficacy against mono and dual-species biofilms. Antibiotics 8:103. doi: 10.3390/antibiotics8030103
Amankwah, S., Abdella, K., Kassa, T., and Amankwah, S. (2022). Bacterial biofilm destruction: A focused review on the recent use of phage-based strategies with other Antibiofilm agents. Nanotechnol. Sci. Appl. 14, 161–177. doi: 10.2147/NSA.S325594
Anju, V. T., Busi, S., Imchen, M., Kumavath, R., Mohan, M. S., Salim, S. A., et al. (2022). Polymicrobial infections and biofilms: Clinical significance and eradication strategies. Antibiotics (Basel) 11:1731. doi: 10.3390/antibiotics11121731
Arroyo-Moreno, S., Cummings, M., Corcoran, D. B., Coffey, A., and McCarthy, R. R. (2022). Identification and characterization of novel endolysins targeting Gardnerella vaginalis biofilms to treat bacterial vaginosis. NPJ Biofilms Microbiom. 8, 29–12. doi: 10.1038/s41522-022-00285-0
Atshan, S. S., Hamat, R. A., Aljaberi, M. A., Chen, J., Huang, S., Lin, C., et al. (2023). Phage therapy as an alternative treatment modality for resistant Staphylococcus aureus infections. Antibiotics (Basel) 12:286. doi: 10.3390/antibiotics12020286
Augustyniak, A., Sikora, P., Grygorcewicz, B., Despot, D., Braun, B., Rakoczy, R., et al. (2021). Biofilms in the gravity sewer interfaces: making a friend from a foe. Rev. Environ. Sci. Biotechnol. 20, 795–813. doi: 10.1007/S11157-021-09582-0
Barr, J. J., Auro, R., Furlan, M., Whiteson, K. L., Erb, M. L., Pogliano, J., et al. (2013). Bacteriophage adhering to mucus provide a non-host-derived immunity. Proc. Natl. Acad. Sci. U. S. A. 110, 10771–10776. doi: 10.1073/pnas.1305923110
Bernard, C., Girardot, M., and Imbert, C. (2020). Candida albicans interaction with gram-positive bacteria within interkingdom biofilms. J Mycol Med 30:100909. doi: 10.1016/j.mycmed.2019.100909
Carson, L., Gorman, S. P., and Gilmore, B. F. (2010). The use of lytic bacteriophages in the prevention and eradication of biofilms of Proteus mirabilis and Escherichia coli. FEMS Immunol. Med. Microbiol. 59, 447–455. doi: 10.1111/j.1574-695X.2010.00696.x
Castro, J., Sousa, L. G. V., França, Â., Tisakova, L. P., Corsini, L., and Cerca, N. (2022). Exploiting the anti-biofilm effect of the engineered phage Endolysin PM-477 to disrupt in vitro single-and dual-species biofilms of vaginal pathogens associated with bacterial vaginosis. Antibiotics 11:558. doi: 10.3390/antibiotics11050558
Chan, B. K., Sistrom, M., Wertz, J. E., Kortright, K. E., Narayan, D., and Turner, P. E. (2016). Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 6, 1–8. doi: 10.1038/srep26717
Chegini, Z., Khoshbayan, A., Vesal, S., Moradabadi, A., Hashemi, A., and Shariati, A. (2021). Bacteriophage therapy for inhibition of multi drug-resistant uropathogenic bacteria: a narrative review. Ann. Clin. Microbiol. Antimicrob. 20, 1–13. doi: 10.1186/s12941-021-00433-y
Chhibber, S., Bansal, S., and Kaur, S. (2015). Disrupting the mixed-species biofilm of klebsiella pneumoniae B5055 and pseudomonas aeruginosa PAO using bacteriophages alone or in combination with xylitol. Microbiology (United Kingdom) 161, 1369–1377. doi: 10.1099/mic.0.000104
Comeau, A. M., Tétart, F., Trojet, S. N., Prère, M. F., and Krisch, H. M. (2007). Phage-antibiotic synergy (PAS): β-lactam and quinolone antibiotics stimulate virulent phage growth. PLoS One 2, 8–11. doi: 10.1371/journal.pone.0000799
Cowley, L. A., Beckett, S. J., Chase-Topping, M., Perry, N., Dallman, T. J., Gally, D. L., et al. (2015). Analysis of whole genome sequencing for the Escherichia coli O157: H7 typing phages. BMC Genomics 16, 1–13. doi: 10.1186/s12864-015-1470-z
Curtin, J. J., and Donlan, R. M. (2006). Using bacteriophages to reduce formation of catheter-associated biofilms by Staphylococcus epidermidis. Antimicrob. Agents Chemother. 50, 1268–1275. doi: 10.1128/AAC.50.4.1268
Delcaru, C., Alexandru, I., Podgoreanu, P., Grosu, M., Stavropoulos, E., Chifiriuc, M., et al. (2016). Microbial biofilms in urinary tract infections and prostatitis: etiology, pathogenicity, and combating strategies. Pathogens 5:65. doi: 10.3390/pathogens5040065
Dickey, J., and Perrot, V. (2019). Adjunct phage treatment enhances the effectiveness of low antibiotic concentration against Staphylococcus aureus biofilms in vitro. PLoS One 14, e0209390–e0209317. doi: 10.1371/journal.pone.0209390
Drulis-Kawa, Z., Majkowska-Skrobek, G., and Maciejewska, B. (2015). Bacteriophages and phage-derived proteins–application approaches. Curr. Med. Chem. 22, 1757–1773. doi: 10.2174/0929867322666150209152851
Duerkop, B. A. (2018). Bacteriophages shift the focus of the mammalian microbiota. PLoS Pathog. 14, e1007310–e1007316. doi: 10.1371/journal.ppat.1007310
Edgar, R., Friedman, N., Shahar, M. M., and Qimron, U. (2012). Reversing bacterial resistance to antibiotics by phage-mediated delivery of dominant sensitive genes. Appl. Environ. Microbiol. 78, 744–751. doi: 10.1128/AEM.05741-11
Ehrlich, G. D., Ahmed, A., Earl, J., Hiller, N. L., Costerton, J. W., Stoodley, P., et al. (2010). The distributed genome hypothesis as a rubric for understanding evolution in situ during chronic bacterial biofilm infectious processes. FEMS Immunol. Med. Microbiol. 59, 269–279. doi: 10.1111/j.1574-695X.2010.00704.x
Ferry, T., Kolenda, C., Briot, T., Souche, A., Lustig, S., Josse, J., et al. (2021). Past and future of phage therapy and phage-derived proteins in patients with bone and joint infection. Viruses 13, 1–20. doi: 10.3390/v13122414
Garrett, T. R., Bhakoo, M., and Zhang, Z. (2008). Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 18, 1049–1056. doi: 10.1016/j.pnsc.2008.04.001
González, S., Fernández, L., Campelo, A. B., Gutiérrez, D., Martínez, B., Rodríguez, A., et al. (2017). The behavior of Staphylococcus aureus dual-species biofilms treated with bacteriophage phiIPLA-RODI depends on the accompanying microorganism. Appl. Environ. Microbiol. 83:e02821-16. doi: 10.1128/AEM.02821-16
Grygorcewicz, B., Gliźniewicz, M., Olszewska, P., Miłek, D., Czajkowski, A., Serwin, N., et al. (2023). Response surface methodology application for bacteriophage–antibiotic Antibiofilm activity optimization. Microorganisms 11:2352. doi: 10.3390/MICROORGANISMS11092352
Grygorcewicz, B., Rakoczy, R., Roszak, M., Konopacki, M., Kordas, M., Piegat, A., et al. (2022). Rotating magnetic field-assisted reactor enhances mechanisms of phage adsorption on bacterial cell surface. Curr. Issues Mol. Biol 44, 1316–1325. doi: 10.3390/cimb44030088
Guła, G., Dorotkiewicz-Jach, A., Korzekwa, K., Valvano, M. A., and Drulis-Kawa, Z. (2018). Complex signaling networks controlling dynamic molecular changes in Pseudomonas aeruginosa biofilm. Curr. Med. Chem. 26, 1979–1993. doi: 10.2174/0929867325666180912110151
Gutiérrez, D., Vandenheuvel, D., Martínez, B., Rodríguez, A., Lavigne, R., and García, P. (2015). Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono-and dual-species staphylococcal biofilms. Appl. Environ. Microbiol. 81, 3336–3348. doi: 10.1128/AEM.03560-14
Hagens, S., Habel, A., Von Ahsen, U., Von Gabain, A., and Bläsi, U. (2004). Therapy of experimental Pseudomonas infections with a nonreplicating genetically modified phage. Antimicrob. Agents Chemother. 48, 3817–3822. doi: 10.1128/AAC.48.10.3817-3822.2004
Herce-Ros, N., Álvarez-Sagües, A., Álvarez-Losa, L., Nistal-Villan, E., Amador, U., Presa, J., et al. (2021). Antibacterial ability of sodium hypochlorite activated with PUI vs. XPF file against Bacteria growth on Enterococcus faecalis mature biofilm. Dent J (Basel) 9:9. doi: 10.3390/dj9060067
Ichikawa, T., Yano, Y., Fujita, Y., Kashiwabara, T., and Nagao, K. (2008). The enhancement effect of three sugar alcohols on the fungicidal effect of benzethonium chloride toward Candida albicans. J. Dent. 36, 965–968. doi: 10.1016/j.jdent.2008.07.013
Infect, J., Epidemiol, D., Riggs, J. M., Mclean, R. J. C., Rohde, R. E., and Aron, G. M. (2016). Development of teicoplanin tolerance by staphylococcus epidermidis and increased susceptibility to bacteriophage type 92 by methicillin-resistant Staphylococcus aureus in polymicrobial biofilms. J. Infect. Dis. Epidemiol. 2:009. doi: 10.23937/2474-3658/1510009
Iszatt, J. J., Larcombe, A. N., Chan, H., Stick, S. M., Garratt, L. W., and Kicic, A. (2021). Tract infections, 1–14.
Jabłońska, J., Dubrowska, K., Augustyniak, A., Kordas, M., and Rakoczy, R. (2022). Application of magnetically assisted reactors for modulation of growth and Pyocyanin production by Pseudomonas aeruginosa. Front. Bioeng. Biotechnol. 10, 1–7. doi: 10.3389/fbioe.2022.795871
Jamal, M., Ahmad, W., Andleeb, S., Jalil, F., Imran, M., Nawaz, M. A., et al. (2018). Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 81, 7–11. doi: 10.1016/j.jcma.2017.07.012
Javed, A., Parvaiz, F., and Manzoor, S. (2019). Bacterial vaginosis: an insight into the prevalence, alternative regimen treatments and it’s associated resistance patterns. Microb. Pathog. 127, 21–30. doi: 10.1016/j.micpath.2018.11.046
Johnston, W., Ware, A., Frederique, W., Delaney, C., Lee, J., Hagen, S., et al. (2023). Biofilm in vitro bacterial vaginosis biofilm community manipulation using endolysin therapy. Biofilms 5:100101. doi: 10.1016/j.bioflm.2022.100101
Johri, A. V., Johri, P., Hoyle, N., Pipia, L., Nadareishvili, L., and Nizharadze, D. (2021). Case report: chronic bacterial prostatitis treated with phage therapy after multiple failed antibiotic treatments. Front. Pharmacol. 12, 1–8. doi: 10.3389/fphar.2021.692614
Jung, H., Ehlers, M. M., Lombaard, H., Redelinghuys, M. J., and Kock, M. M. (2017). Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 43, 651–667. doi: 10.1080/1040841X.2017.1291579
Kay, M. K., Erwin, T. C., McLean, R. J. C., and Aron, G. M. (2011). Bacteriophage ecology in Escherichia coli and pseudomonas aeruginosa mixed-biofilm communities. Appl. Environ. Microbiol. 77, 821–829. doi: 10.1128/AEM.01797-10
Kaźmierczak, N., Grygorcewicz, B., Roszak, M., Bochentyn, B., and Piechowicz, L. (2022). Comparative assessment of bacteriophage and antibiotic activity against multidrug-resistant Staphylococcus aureus biofilms. Int. J. Mol. Sci. 23:1274. doi: 10.3390/IJMS23031274
Kifelew, L. G., Mitchell, J. G., and Speck, P. (2019). Mini-review: efficacy of lytic bacteriophages on multispecies biofilms Mini-review: efficacy of lytic bacteriophages on multispecies biofilms. Biofouling 35, 472–481. doi: 10.1080/08927014.2019.1613525
Kifelew, L. G., Warner, M. S., Morales, S., Thomas, N., Gordon, D. L., Mitchell, J. G., et al. (2020). Efficacy of lytic phage cocktails on staphylococcus aureus and pseudomonas aeruginosa in mixed-species planktonic cultures and biofilms. Viruses 12:559. doi: 10.3390/v12050559
Konopacki, M., Grygorcewicz, B., Kordas, M., Dołęgowska, B., and Rakoczy, R. (2020). Methods of bacteriophages production with application of alternate magnetic field. PAIC 2019: Practical Aspects of Chemical Engineering, 171–182.
Koo, H., Allan, R. N., Howlin, R. P., Stoodley, P., and Hall-Stoodley, L. (2017). Targeting microbial biofilms: current and prospective therapeutic strategies. Nat. Rev. Microbiol. 15, 740–755. doi: 10.1038/nrmicro.2017.99
Landlinger, C., Tisakova, L., Oberbauer, V., Schwebs, T., Muhammad, A., Latka, A., et al. (2021). Engineered phage endolysin eliminates gardnerella biofilm without damaging beneficial bacteria in bacterial vaginosis ex vivo. Pathogens 10, 1–19. doi: 10.3390/pathogens10010054
Lehman, S. M., and Donlan, R. M. (2015). Bacteriophage-mediated control of a two-species biofilm formed by microorganisms causing catheter-associated urinary tract infections in an in vitro urinary catheter model. Antimicrob. Agents Chemother. 59, 1127–1137. doi: 10.1128/AAC.03786-14
Li, L. L., Yu, P., Wang, X., Yu, S. S., Mathieu, J., Yu, H. Q., et al. (2017). Enhanced biofilm penetration for microbial control by polyvalent phages conjugated with magnetic colloidal nanoparticle clusters (CNCs). Environ. Sci. Nano 4, 1817–1826. doi: 10.1039/c7en00414a
Liao, K. S., Lehman, S. M., Tweardy, D. J., Donlan, R. M., and Trautner, B. W. (2012). Bacteriophages are synergistic with bacterial interference for the prevention of Pseudomonas aeruginosa biofilm formation on urinary catheters. J. Appl. Microbiol. 113, 1530–1539. doi: 10.1111/j.1365-2672.2012.05432.x
Liu, W., Røder, H. L., Madsen, J. S., Bjarnsholt, T., Sørensen, S. J., and Burmølle, M. (2016). Interspecific bacterial interactions are reflected in multispecies biofilm spatial organization. Front. Microbiol. 7, 1–8. doi: 10.3389/fmicb.2016.01366
Lu, T. K., and Collins, J. J. (2007). Dispersing biofilms with engineered enzymatic bacteriophage. Proc. Natl. Acad. Sci. U. S. A. 104, 11197–11202. doi: 10.1073/pnas.0704624104
Lu, T. K., and Collins, J. J. (2009). Engineered bacteriophage targeting gene networks as adjuvants for antibiotic therapy. Proc. Natl. Acad. Sci. U. S. A. 106, 4629–4634. doi: 10.1073/pnas.0800442106
Łubowska, N., Grygorcewicz, B., Kosznik-Kwaśnicka, K., Zauszkiewicz-Pawlak, A., Węgrzyn, A., Dołęgowska, B., et al. (2019). Characterization of the three new Kayviruses and their lytic activity against multidrug-resistant Staphylococcus aureus. Microorganisms 7:471. doi: 10.3390/microorganisms7100471
Manohar, P., Loh, B., Nachimuthu, R., Leptihn, S., and Nachimuthu Assistant Professor, R. (2022). Phage-antibiotic combinations to control Pseudomonas aeruginosa-Candida two-species biofilms. bio Rxiv 2022.08.18.504394 Available at: https://www.biorxiv.org/content/10.1101/2022.08.18.504394v1%0Ahttps://www.biorxiv.org/content/10.1101/2022.08.18.504394v1.abstract
Manoharadas, S., Ahmad, N., Altaf, M., Alrefaei, A. F., and Al-Rayes, B. F. (2023). An Enzybiotic cocktail effectively disrupts preformed dual biofilm of Staphylococcus aureus and Enterococcus faecalis. Pharmaceuticals 16:564. doi: 10.3390/ph16040564
Manoharadas, S., Altaf, M., Alrefaei, A. F., Hussain, S. A., Devasia, R. M., Badjah Hadj, A. Y. M., et al. (2021). Microscopic analysis of the inhibition of staphylococcal biofilm formation by Escherichia coli and the disruption of preformed staphylococcal biofilm by bacteriophage. Microsc. Res. Tech. 84, 1513–1521. doi: 10.1002/jemt.23707
Maura, D., Morello, E., du Merle, L., Bomme, P., Le Bouguénec, C., and Debarbieux, L. (2012). Intestinal colonization by enteroaggregative Escherichia coli supports long-term bacteriophage replication in mice. Environ. Microbiol. 14, 1844–1854. doi: 10.1111/j.1462-2920.2011.02644.x
Melo, L. D. R., Ferreira, R., Costa, A. R., Oliveira, H., and Azeredo, J. (2019). Efficacy and safety assessment of two enterococci phages in an in vitro biofilm wound model. Sci. Rep. 9, 6643–6612. doi: 10.1038/s41598-019-43115-8
Melo, L. D. R., Veiga, P., Cerca, N., Kropinski, A. M., Almeida, C., Azeredo, J., et al. (2016). Development of a phage cocktail to control Proteus mirabilis catheter-associated urinary tract infections. Front. Microbiol. 7:1024. doi: 10.3389/fmicb.2016.01024
Mgomi, F. C., Yuan, L., Chen, C., Zhang, Y. S., and Yang, Z. Q. (2022). Bacteriophages: a weapon against mixed-species biofilms in the food processing environment. J. Appl. Microbiol. 133, 2107–2121. doi: 10.1111/jam.15421
Milho, C., Silva, M. D., Alves, D., Oliveira, H., Sousa, C., Pastrana, L. M., et al. (2019). Escherichia coli and Salmonella Enteritidis dual-species biofilms: interspecies interactions and antibiofilm efficacy of phages. Sci. Rep. 9, 18183–18115. doi: 10.1038/s41598-019-54847-y
Morozova, V. V., Kozlova, Y. N., Ganichev, D. A., and Tikunova, N. V. (2018). Bacteriophage treatment of infected diabetic foot ulcers. Methods Mol. Biol. 1693, 151–158. doi: 10.1007/978-1-4939-7395-8_13
Morrisette, T., Kebriaei, R., Lev, K. L., Morales, S., and Rybak, M. J. (2019). Bacteriophage therapeutics: a primer for clinicians on phage-antibiotic combinations. Pharmacotherapy 40, 153–168. doi: 10.1002/phar.2358
Myszka, K., and Czaczyk, K. (2010). Quorum sensing mechanism as a factor regulating virulence of gram-negative bacteria. Postepy Hig. Med. Dosw. 64, 582–589.
Nadareishvili, L., Hoyle, N., Nakaidze, N., Nizharadze, D., Kutateladze, M., Balarjishvili, N., et al. (2020). Bacteriophage therapy as a potential management option for surgical wound infections. PHAGE: Ther. Appl. Res. 1, 158–165. doi: 10.1089/phage.2020.0010
Narayanan, A., Nair, M. S., Muyyarikkandy, M. S., and Amalaradjou, M. A. (2018). Inhibition and inactivation of uropathogenic escherichia coli biofilms on urinary catheters by sodium selenite. Int. J. Mol. Sci. 19:1703. doi: 10.3390/ijms19061703
Nazik, H., Joubert, L. M., Secor, P. R., Sweere, J. M., Bollyky, P. L., Sass, G., et al. (2017). Pseudomonas phage inhibition of Candida albicans. Microbiology (United Kingdom) 163, 1568–1577. doi: 10.1099/mic.0.000539
Nir-paz, R., Gelman, D., Khouri, A., Sisson, B. M., Yerushalmy, O., Bader, R., et al. (2019). Successful treatment of antibiotic-resistant poly-microbial bone infection with bacteriophages and antibiotics combination. Clin. Infect. Dis. 69, 2015–2018. doi: 10.1093/cid/ciz222
Oliveira, A., Sousa, J. C., Silva, A. C., Melo, L. D. R., and Sillankorva, S. (2018). Chestnut honey and bacteriophage application to control pseudomonas aeruginosa and escherichia coli biofilms: evaluation in an ex vivo wound model. Front. Microbiol. 9, 1–13. doi: 10.3389/fmicb.2018.01725
Onsea, J., Soentjens, P., Djebara, S., Merabishvili, M., Depypere, M., Spriet, I., et al. (2019). Bacteriophage application for difficult-to-treat musculoskeletal infections: development of a standardized multidisciplinary treatment protocol. Viruses 11:891. doi: 10.3390/v11100891
Park, K., Cha, K. E., and Myung, H. (2014). Observation of inflammatory responses in mice orally fed with bacteriophage T7. J. Appl. Microbiol. 117, 627–633. doi: 10.1111/jam.12565
Park, T., Chaudhry, W. N., Concepcio, J., Andleeb, S., Bull, J. J., and Levin, B. R. (2017). Synergy and order effects of antibiotics and phages in killing Pseudomonas aeruginosa biofilms. PLoS One 12:e0168615. doi: 10.1371/journal.pone.0168615
Patey, O., McCallin, S., Mazure, H., Liddle, M., Smithyman, A., and Dublanchet, A. (2019). Clinical indications and compassionate use of phage therapy: personal experience and literature review with a focus on osteoarticular infections. Viruses 11, 1–21. doi: 10.3390/v11010018
Pei, R., and Lamas-Samanamud, G. R. (2014). Inhibition of biofilm formation by T7 bacteriophages producing quorum-quenching enzymes. Appl. Environ. Microbiol. 80, 5340–5348. doi: 10.1128/AEM.01434-14
Peters, B. M., Jabra-Rizk, M. A., O’May, G. A., William Costerton, J., and Shirtliff, M. E. (2012). Polymicrobial interactions: impact on pathogenesis and human disease. Clin. Microbiol. Rev. 25, 193–213. doi: 10.1128/CMR.00013-11
Pires, D. P., Melo, L. D. R., Boas, D. V., and Sillankorva, S. (n.d.). Science direct phage therapy as an alternative or complementary strategy to prevent and control biofilm-related infections. Curr. Opin. Microbiol. 39, 48–56. doi: 10.1016/j.mib.2017.09.004
Pires, D. P., Silva, S., Almeida, C., Henriques, M., Anderson, E. M., Lam, J. S., et al. (2013). Evaluation of the ability of C. albicans to form biofilm in the presence of phage-resistant phenotypes of P. aeruginosa. Biofouling 29, 1169–1180. doi: 10.1080/08927014.2013.831842
Pohl, C. H. (2022). Recent advances and opportunities in the Study of Candida albicans Polymicrobial biofilms. Front. Cell. Infect. Microbiol. 12, 1–17. doi: 10.3389/fcimb.2022.836379
Püschel, A., Skusa, R., Bollensdorf, A., and Gross, J. (2022). Local treatment of driveline infection with bacteriophages. Antibiotics 11, 1–6. doi: 10.3390/antibiotics11101310
Reece, E., de Almeida Bettio, P. H., and Renwick, J. (2021). Polymicrobial interactions in the cystic fibrosis airway microbiome impact the antimicrobial susceptibility of pseudomonas aeruginosa. Antibiotics 10:827. doi: 10.3390/antibiotics10070827
Rickard, A. H., Gilbert, P., High, N. J., Kolenbrander, P. E., and Handley, P. S. (2003). Bacterial coaggregation: an integral process in the development of multi-species biofilms. Trends Microbiol. 11, 94–100. doi: 10.1016/S0966-842X(02)00034-3
Roszak, M., Dołȩgowska, B., Cecerska-Heryć, E., Serwin, N., Jabłońska, J., and Grygorcewicz, B. (2022). Bacteriophage-ciprofloxacin combination effectiveness depends on Staphylococcus aureus-Candida albicans dual-species communities’ growth model. Microb. Drug Resist. 28, 613–622. doi: 10.1089/mdr.2021.0324
Roy, R., Tiwari, M., Donelli, G., and Tiwari, V. (2018). Strategies for combating bacterial biofilms: a focus on anti-biofilm agents and their mechanisms of action. Virulence 9, 522–554. doi: 10.1080/21505594.2017.1313372
Rubalskii, E., Ruemke, S., Salmoukas, C., Boyle, E. C., Warnecke, G., Tudorache, I., et al. (2020). Bacteriophage therapy for critical infections related to cardiothoracic surgery. Antibiotics 9, 1–12. doi: 10.3390/antibiotics9050232
Santiago, A. J., Burgos-Garay, M. L., Kartforosh, L., Mazher, M., and Donlan, R. M. (2020). Bacteriophage treatment of carbapenemase-producing Klebsiella pneumoniae in a multispecies biofilm: a potential biocontrol strategy for healthcare facilities. AIMS Microbiol. 6, 43–63. doi: 10.3934/microbiol.2020003
Santiago, A. J., and Donlan, R. M. (2020). Bacteriophage infections of biofilms of health care-associated pathogens: Klebsiella pneumoniae. 9, 1–12. doi:doi: 10.1128/ecosalplus.ESP-0029-2019
Sartini, S., Permana, A. D., Mitra, S., Tareq, A. M., Salim, E., Ahmad, I., et al. (2021). Current state and promising opportunities on pharmaceutical approaches in the treatment of Polymicrobial diseases. Pathogens 10:245. doi: 10.3390/pathogens10020245
Schuch, R., Khan, B. K., Raz, A., Rotolo, J. A., and Wittekind, M. (2017). Bacteriophage lysin CF-301, a potent antistaphylococcal biofilm agent. Antimicrob Agents Chemother 61:e02666–16. doi: 10.1128/AAC.02666-16
Sillankorva, S., Neubauer, P., and Azeredo, J. (2010). Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 26, 567–575. doi: 10.1080/08927014.2010.494251
Sousa, L. G. V., Pereira, S. A., and Cerca, N. (2023). Fighting polymicrobial biofilms in bacterial vaginosis. Microb. Biotechnol. 16, 1423–1437. doi: 10.1111/1751-7915.14261
Srinivasan, R., Santhakumari, S., Poonguzhali, P., Geetha, M., Dyavaiah, M., and Xiangmin, L. (2021). Bacterial biofilm inhibition: a focused review on recent therapeutic strategies for combating the biofilm mediated infections. Front. Microbiol. 12, 1–19. doi: 10.3389/fmicb.2021.676458
Stickler, D. J. (2014). Clinical complications of urinary catheters caused by crystalline biofilms: something needs to be done. J. Intern. Med. 276, 120–129. doi: 10.1111/joim.12220
Struk, M., Grygorcewicz, B., Nawrotek, P., Augustyniak, A., Konopacki, M., Kordas, M., et al. (2017). Enhancing effect of 50 Hz rotating magnetic field on induction of Shiga toxin-converting lambdoid prophages. Microb. Pathog. 109, 4–7. doi: 10.1016/j.micpath.2017.05.018
Szafrański, S. P., Winkel, A., and Stiesch, M. (2017). The use of bacteriophages to biocontrol oral biofilms. J. Biotechnol. 250, 29–44. doi: 10.1016/j.jbiotec.2017.01.002
Tait, K., Skillman, L. C., and Sutherland, I. W. (2002). The efficacy of bacteriophage as a method of biofilm eradication. Biofouling 18, 305–311. doi: 10.1080/0892701021000034418
Tanaka, T., Yahata, Y., Handa, K., Venkataiah, S. V., Njuguna, M. M., Kanehira, M., et al. (2021). An experimental intraradicular biofilm model in the pig for evaluating irrigation techniques. BMC Oral Health 21, 177–112. doi: 10.1186/s12903-021-01536-w
Testa, S., Berger, S., Piccardi, P., Oechslin, F., Resch, G., and Mitri, S. (2019). Spatial structure affects phage efficacy in infecting dual-strain biofilms of Pseudomonas aeruginosa. Commun. Biol. 2, 405–412. doi: 10.1038/s42003-019-0633-x
Thompson, A. F., English, E. L., Nock, A. M., Willsey, G. G., Eckstrom, K., Cairns, B., et al. (2020). Characterizing species interactions that contribute to biofilm formation in a multispecies model of a potable water bacterial community. Microbiology (United Kingdom) 166, 34–43. doi: 10.1099/mic.0.000849
Tkhilaishvili, T., Wang, L., Perka, C., Trampuz, A., and Gonzalez Moreno, M. (2020). Using bacteriophages as a Trojan horse to the killing of dual-species biofilm formed by Pseudomonas aeruginosa and methicillin resistant Staphylococcus aureus. Front. Microbiol. 11:695. doi: 10.3389/fmicb.2020.00695
Topka-bielecka, G., Dydecka, A., Necel, A., and Bloch, S. (2021). Bacteriophage-derived Depolymerases against bacterial biofilm. Antibiotics (Basel) 10:175. doi: 10.3390/antibiotics10020175
Uyttebroek, S., Onsea, J., Metsemakers, W. J., Dupont, L., Devolder, D., Wagemans, J., et al. (2021). The potential role of bacteriophages in the treatment of recalcitrant chronic rhinosinusitis. Antibiotics 10, 1–13. doi: 10.3390/antibiotics10060675
Van Nieuwenhuyse, B., Galant, C., Brichard, B., Docquier, P. L., Djebara, S., Pirnay, J. P., et al. (2021). A case of in situ phage therapy against staphylococcus aureus in a bone allograft polymicrobial biofilm infection: outcomes and phage-antibiotic interactions. Viruses 13, 1–12. doi: 10.3390/v13101898
Verbanic, S., Deacon, J. M., and Chen, I. A. (2022). The chronic wound Phageome: Phage diversity and associations with wounds and healing outcomes. Microbiol Spectr. 10:e0277721. doi: 10.1128/spectrum.02777-21
Vidakovic, L., Singh, P. K., Hartmann, R., Nadell, C. D., and Drescher, K. (2017). Dynamic biofilm architecture confers individual and collective mechanisms of viral protection. Nat. Microbiol. 3, 26–31. doi: 10.1038/s41564-017-0050-1
Vu, B., Chen, M., Crawford, R. J., and Ivanova, E. P. (2009). Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 14, 2535–2554. doi: 10.3390/molecules14072535
Winans, J. B., Wucher, B. R., and Nadell, C. D. (2022). Multispecies biofilm architecture determines bacterial exposure to phages. PLoS Biol. 20, e3001913–e3001924. doi: 10.1371/journal.pbio.3001913
Wolcott, R., Costerton, J. W., Raoult, D., and Cutler, S. J. (2013). The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. 19, 107–112. doi: 10.1111/j.1469-0691.2012.04001.x
Yamamoto, K., Kusada, H., Kamagata, Y., and Tamaki, H. (2021). Parallel evolution of enhanced biofilm formation and phage-resistance in Pseudomonas aeruginosa during adaptation process in spatially heterogeneous environments. Microorganisms 9:569. doi: 10.3390/microorganisms9030569
Yang, D., Wang, Z., Ma, J., Fu, Q., Wu, L., Wang, H., et al. (2020). Glycine Cleavage System and cAMP Receptor Protein Co-Regulate CRISPR/cas3 Expression to Resist bacteriophage. Viruses 12:90. doi: 10.3390/v12010090
Yu, P., Mathieu, J., Yang, Y., and Alvarez, P. J. J. (2017). Suppression of enteric Bacteria by bacteriophages: importance of phage polyvalence in the presence of soil Bacteria. Environ. Sci. Technol. 51, 5270–5278. doi: 10.1021/acs.est.7b00529
Yu, P., Wang, Z., Marcos-Hernandez, M., Zuo, P., Zhang, D., Powell, C., et al. (2019). Bottom-up biofilm eradication using bacteriophage-loaded magnetic nanocomposites: a computational and experimental study. Environ. Sci. Nano 6, 3539–3550. doi: 10.1039/c9en00827f
Zhao, G., Vatanen, T., Droit, L., Park, A., Kostic, A. D., Poon, T. W., et al. (2017). Intestinal virome changes precede autoimmunity in type I diabetes-susceptible children. Proc. Natl. Acad. Sci. U. S. A. 114, E6166–E6175. doi: 10.1073/pnas.1706359114
Zhu, W., Ding, Y., Huang, C., Wang, J., Wang, J., and Wang, X. (2022). Genomic characterization of a novel bacteriophage STP55 revealed its prominent capacity in disrupting the dual-species biofilm formed by Salmonella typhimurium and Escherichia coli O157: H7 strains. Arch. Microbiol. 204, 1–17. doi: 10.1007/s00203-022-03208-x
Keywords: phage therapy, depolymerases, multi-species biofilm, phage-antibiotic synergy, polymicrobial infections
Citation: Gliźniewicz M, Miłek D, Olszewska P, Czajkowski A, Serwin N, Cecerska-Heryć E, Dołęgowska B and Grygorcewicz B (2024) Advances in bacteriophage-mediated strategies for combating polymicrobial biofilms. Front. Microbiol. 14:1320345. doi: 10.3389/fmicb.2023.1320345
Edited by:
Sylwia Bloch, University of Gdansk, PolandReviewed by:
Bozena Nejman-Falenczyk, University of Gdansk, PolandCopyright © 2024 Gliźniewicz, Miłek, Olszewska, Czajkowski, Serwin, Cecerska-Heryć, Dołęgowska and Grygorcewicz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Gliźniewicz, bWFydGEuZ2xpem5pZXdpY3pAcHVtLmVkdS5wbA==; Bartłomiej Grygorcewicz, YmFydGxvbWllai5ncnlnb3JjZXdpY3pAcHVtLmVkdS5wbA==; Yi5ncnlnb3JjZXdpY3pAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.