 Noora Barzkar
Noora Barzkar Stanislav Sukhikh
Stanislav Sukhikh Olga Babich
Olga Babich
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Microbiol. , 08 January 2024
Sec. Aquatic Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1285902
The marine environment has remained a source of novel biological molecules with diversified applications. The ecological and biological diversity, along with a unique physical environment, have provided the evolutionary advantage to the plant, animals and microbial species thriving in the marine ecosystem. In light of the fact that marine microorganisms frequently interact symbiotically or mutualistically with higher species including corals, fish, sponges, and algae, this paper intends to examine the potential of marine microorganisms as a niche for marine bacteria. This review aims to analyze and summarize modern literature data on the biotechnological potential of marine fungi and bacteria as producers of a wide range of practically valuable products (surfactants, glyco-and lipopeptides, exopolysaccharides, enzymes, and metabolites with different biological activities: antimicrobial, antitumor, and cytotoxic). Hence, the study on bioactive secondary metabolites from marine microorganisms is the need of the hour. The scientific novelty of the study lies in the fact that for the first time, the data on new resources for obtaining biologically active natural products — metabolites of marine bacteria and fungi — were generalized. The review investigates the various kinds of natural products derived from marine microorganisms, specifically focusing on marine bacteria and fungi as a valuable source for new natural products. It provides a summary of the data regarding the antibacterial, antimalarial, anticarcinogenic, antibiofilm, and anti-inflammatory effects demonstrated by marine microorganisms. There is currently a great need for scientific and applied research on bioactive secondary metabolites of marine microorganisms from the standpoint of human and animal health.
A large portion of this planet is covered by oceans that harbor a variety of living forms with enormous potential to produce natural compounds. Indeed, marine organisms are reportedly a better source for the production of bioactive compounds than their terrestrial counterparts (Karthikeyan et al., 2022). Life on this planet evolved in a marine environment and hence the marine organisms have witnessed all the evolutionary periods and have acquired the ability to produce diversified compounds with unique structural and functional properties (Jiménez, 2018; Karthikeyan et al., 2022). Approximately 28,500 marine natural products (including polysaccharides, peptides, polyketides, polyphenolic compounds, sterol-like products, and alkaloids, among others) had been identified by the end of 2016 (Blunt et al., 2018). Marine natural products present a broad range of biological activities, including anticancer (Carroll et al., 2023), antibacterial (Mohie et al., 2023), antifungal (El-Hossary et al., 2017), and antiviral effects (Ji et al., 2018). Such a range of activities makes the secondary metabolites promising for the development of novel drug prototypes. Despite the promise of marine microorganisms, the literature has not given them enough attention. As a result, our knowledge of their capacities and bioactive qualities is currently restricted. Unexplored marine worlds characterized by high biodiversity are a resource for discovering new structures with unique characteristics. The long evolution of marine organisms has resulted in organisms with rare genes. The environment in which marine microorganisms must survive includes high pressure (up to 1,100 atmospheres), anaerobic conditions, temperatures below 0°C, high acidity (pH 2.8), and ambient temperatures (above 100°C) in hot springs. It was also necessary to adapt to high salinity, radiation, light, and reduced nutrients (Poli et al., 2017). In such cases, extreme conditions promoted genetic and metabolic diversity in marine microorganisms, leading to specific adaptive mechanisms, especially the synthesis of rare protective compounds. It has now been shown that marine microorganisms can synthesize many unique metabolites with different biological properties, which are expected to be used in the pharmaceutical, cosmetic, and medical industries (Puttaswamygowda et al., 2019). For example, microbial hydrolases work in conditions that result in the deposition or denaturation of proteins produced by mesophilic (terrestrial) microorganisms (Ruginescu et al., 2022). Furthermore, seawater, which is similar to human blood plasma in terms of physiology and chemistry, contains biomolecules with lower toxicity and greater therapeutic effectiveness compared to traditional enzymes. As a result, it is not unexpected that there is a growing number of publications each year on the exploration of marine microorganisms as sources of biologically active substances (Schofield et al., 2015; Gui et al., 2017; Ye et al., 2017; Zhang et al., 2018; Hanif et al., 2019; Sun et al., 2019; Cho et al., 2020; Martyniuk et al., 2020; Pedrosa et al., 2020; Ameen et al., 2021; Chung et al., 2023; Nugraha et al., 2023). About 400 reviews have been published in the last five years with the keyword “marine microorganisms.” Such estimates, however, are “narrowly specific” because marine microorganisms produce antibiotics (Ye et al., 2017; Ameen et al., 2021), antitumor compounds (Gui et al., 2017), enzymes (Sun et al., 2019), polysaccharides (Martyniuk et al., 2020), or those that account for the ability of marine fungi (Schofield et al., 2015; Cho et al., 2020), algae (Nugraha et al., 2023) to synthesize some metabolites. Also, most of these reviews focus on marine microbial biodiversity, the determination of chemical composition, structure of metabolites (Gui et al., 2017; Martyniuk et al., 2020; Pedrosa et al., 2020; Chung et al., 2023; Nugraha et al., 2023) and their biological activity (Gui et al., 2017; Sun et al., 2019; Cho et al., 2020; Ameen et al., 2021; Nugraha et al., 2023). However, the prospects of marine microorganisms for use in biotechnology are not considered.
This review intends to examine and condense recent literature findings on the biotechnological capabilities of marine fungi and bacteria in producing a diverse array of valuable products. These products include surfactants, glyco-and lipopeptides, exopolysaccharides, enzymes, as well as metabolites with various biological activities such as antimicrobial, antitumor, and cytotoxic properties.
The presence of marine microorganisms in the marine habitat is very diverse, including in shallow and deep waters, deep-sea hydrothermal vents, polar regions and in varied coral reefs (Poli et al., 2017). The surface of marine organisms, such as corals, fish, sponges and algae serves as a niche for marine bacteria since these microorganisms frequently have symbiotic or mutualistic relationships with higher organisms (Puttaswamygowda et al., 2019). It is estimated that only 0.01% of marine bacteria have been so far characterized (Ruginescu et al., 2022). Analyses of marine metagenomes indicate a large number of bacteria and archaea phylogenetic groups, that are still unexploited (Martyniuk et al., 2020). True marine bacteria represent a treasure for the discovery of novel bioactive compounds (Pedrosa et al., 2020). However, only 179 novel bioactive compounds were characterized in marine bacteria in 2016 (Blunt et al., 2018). For instance, the marine-derived strain Micromonospora harpali produces a group of new spirotetronate analogs (e.g., Microsporanates A (1)–F and tetrocarcins p (2), A, B, and AC6H that display antibacterial activity against Gram-positive bacteria (MIC of 0.016 to 8 μg mL−1) (Gui et al., 2017). Chromomycins (e.g., chromomycin Ap, (3) isolated from cultures of the marine-derived actinomycete Streptomyces sp. MBTI36 showed potent antibacterial activity against Gram-positive bacteria including methicillin-resistant Staphylococcus aureus (MRSA) (Cho et al., 2020). The bacterium Micromonospora sp. produces thiocoraline (4), a depsipeptide which inhibits DNA polymerase-α, and has been applied for the treatment of cancer in preclinical research (Ameen et al., 2021). Moreover, salinosporamide A (5) isolated from cultures of the marine actinobacterium Salinispora tropica (Sun et al., 2019) is in clinical trials for the treatment of multiple myeloma, solid tumors, and lymphomas (Chung et al., 2023; Nugraha et al., 2023).
The bacterium Streptomyces sp. P11-238 creates two cyclodepsipeptides known as streptodepsipeptides P11A and P11B. These compounds have been found to slow the growth of various glioma cell lines, with IC50 values ranging from 0.1 to 1.4 μM. Streptodepsipeptide P11A specifically has been shown to halt the cell cycle in the G0/G1 phase, induce apoptosis, and reduce the activity of certain tumor metabolic enzymes (HK2, PFKFB3, PKM2, LLS, and FASN) (Ye et al., 2017). (−)-Ecteinascidin 743 (Yondelis), on the other hand, was first discovered in the Caribbean ascidian Ecteinascidia turbinate but was later found to be produced by a γ-proteobacterial endosymbiont called Candidatus endoecteinascidia frumentensis (Schofield et al., 2015). ET-743 is currently used in medical treatments for advanced soft tissue sarcoma and recurrent platinum-sensitive ovarian cancer when combined with liposomal doxorubicin (Hanif et al., 2019).
These examples show how marine bacteria can manufacture bioactive chemicals that could lead to the creation of brand-new medicinal medicines.
A literature survey between 2017 and 2021 of publications in the top leading scientific journals reporting novel bioactive metabolites from marine bacteria provided the following results. Streptoseomycin (8) has been isolated from cultures of the marine-derived Streptomyces seoulensis A01 and displayed very good antibacterial activity against Helicobacter pylori, Lactobacillus acidophilus, Bifidobacterium bifidum, Eubacterium brachy, Propionibacterium acnes, Staphylococcus aureus, Micrococcus luteus and Bacillus subtilis, with MICs between 2 and 64 μg/mL (Zhang et al., 2018). Pyrroloformamides C (9) and D were isolated from cultures of Streptomyces sp. CB02980 and displayed moderate activity against S. aureus ATCC 29213, MethicillinResistant Staphylococcus aureus, Escherichia coli, Klebsiella pneumoniae (Zhou et al., 2020). The anthranilate derivatives anthranosides A, B and C (10) have been isolated from cultures of the marine bacterium Streptomyces sp. CMN-62. Anthranoside C (10) displayed anti-viral activity with IC50 of 171 μM against the influenza A H1N1 virus (ribavirin as positive control, IC50 133 μM) (Che et al., 2018). The alkaloids bulbimidazoles A (11) B and C have been isolated from cultures of the marine bacterium Microbulbifer sp. DC3-6 and displayed antibacterial activity against Kocuria rhizophila ATCC9341 and against S. aureus FDA209P JC-1 (Karim et al., 2020). Only these three articles reporting novel bioactive metabolites from marine bacteria were published in leading chemistry journals between 2017 and 2021, illustrating that the knowledge of marine bacteria secondary metabolism is still very incipient.
Thus, based on the literature review presented in this section, it can be concluded that there are few studies reporting novel bioactive metabolites of marine bacteria. It was found that the properties of secondary metabolites in marine bacteria are poorly understood. However, it is clear that the potential use of marine bacterial metabolites is great, especially for the treatment of malignant tumors, bacterial and viral infections.
Obligate and facultative marine fungi are the two categories by which these organisms are conventionally classified. Regarding diversity and ecological significance, this divide is still debatable (Deshmukh et al., 2018). Oversimplified descriptions have left the definition of marine fungus uncertain. It is frequently forgotten that marine mushrooms may adapt to a variety of marine settings, including saltmarshes, hydrothermal zones, and the deep sea (Gladfelter et al., 2019). Nearly all maritime habitats have been shown to include fungi, including sediments (Barone, 2022), the saltwater column (Xu et al., 2018), and combinations with other marine organisms such as sponges, corals, and algae (Zhao et al., 2019; de Sá et al., 2022; Giddings and Newman, 2022; Hafez Ghoran et al., 2023). Both deep and surface seas include marine fungi, which are a reliable source of many healthy substances like antimicrobials, antioxidants, and anticancer chemicals (Deshmukh et al., 2018).
A marine Aspergillus sp. fungus produces dehydroxychlorofusarielin B (12), which exhibits antibacterial activity against methicillin-resistant and multidrug-resistant (MDR) Staphylococcus aureus (Gladfelter et al., 2019). Linear peptides, simplicilliumtides A (13), E, G, and H were isolated from cultures of Simplicillium obclavatum EIODSF 020. Simplicilliumtide A (13) and G showed weak cytotoxicity toward human leukemia HL-60 cell line with IC50 values of 64.7 and 100 μM, while simplicilliumtides E and H showed weak cytotoxicity toward the K562 cell line with IC50 values of 39.4 and 73.5 μM (Deshmukh et al., 2018). Chaetoxanthone B (14) was isolated from cultures of a marine Chaetomium sp. and showed very good antimalarial activity against Plasmodium falciparum (IC50 = 0.5 μg mL−1) and activity against Trypanosoma cruzi, the causative agent of Chagas disease (IC50 = 1.5 μg mL−1) (Barone, 2022). Five new benzophenone derivatives, including a new eremophilane derivative, have been isolated from cultures of marine-derived Phomopsis lithocarpus (Xu et al., 2018). These compounds possess a rare naturally occurring aldehyde functionality within this family. One of the new compounds, tenellone H (15), has exhibited cytotoxic activity against HepG-2 and A549 cell lines with IC50 values of 16.0 and 17.6 μM, respectively. Another marine-derived fungus, Aspergillus niger, produced malformin C (16), which showed strong anti-HIV-1 activity (Hafez Ghoran et al., 2023). Additionally, stachyflin (17), a terpenoid isolated from cultures of marine-derived Stachybotrys, demonstrated modest activity against the influenza A virus (H1N1) with an IC50 of 3,910 − 3 μM (Zhao et al., 2019). These examples highlight the potential for research on marine fungal secondary metabolism as a source for developing drug targets since nearly 38% of microbial bioactive metabolites are of fungal origin, but only 5% of the world’s fungal taxa have been described so far (Giddings and Newman, 2022).
The researchers de Sá et al. found that Neosartorya, a type of marine fungi, are closely related to the group A. fumigatus. This suggests that the secondary metabolites produced by Neosartorya fungi will be similar to those produced by Aspergillus species. In fact, they have discovered that there are common features in the secondary metabolites of both Neosartorya and Aspergillus species, particularly indoles, meroterpenoids, and polyketides (de Sá et al., 2022). They have also identified different types of metabolites within the same species, depending on the environment they are found in. These include indole alkaloids, prenylated indoles, 1,4-benzodiazepen-2,5-dione-containing prenylated indoles, and many others such as peptides, terpenoids, sterols and sterones, polyketides, benzoic acid derivatives, and nucleosides.
Compounds isolated from representatives of the genus Neosartorya were tested for biological and pharmacological activity, mainly in vitro. Like all other natural products, most of the natural compounds isolated from Neosartorya species exhibited anticancer,cytotoxic and antimicrobial activities (de Sá et al., 2022).
Biologically active substances from marine microorganisms are isolated or purified using preparative column chromatography and high-performance liquid chromatography (HPLC) on direct and reversed phases. The structures of the compounds are determined using state-of-the-art NMR spectroscopy, including 1H, 13C, COSY, NOESY, HSQC, HMBC, TOCSY, and mass spectrometry (EI, LSI, ESI, MALDI-TOF), including tandem mass spectrometry. Chemical transformations are used to obtain derivatives of biologically active substances from marine microorganisms (Makarieva et al., 2014). Absolute configurations of asymmetric centers are established using various methods. For example, acetylated derivatives of (−)-2-octyl-glycosides of the corresponding monosaccharides are analyzed using gas–liquid chromatography to determine the absolute configuration of the monosaccharides. Relative and absolute configurations of asymmetric side chain centers of new steroids and terpenoids were determined using the modified Mosher method (Makarieva et al., 2014).
In the environments where we live, bacteria are extensively dispersed. Infections brought on by pathogenic bacteria have a tremendous impact on human health despite the fact that only a limited number of bacteria may cause them (Packiavathy et al., 2021). Bacterial infections are generally less severe compared to fungal or viral infections (Shafiekhani et al., 2022). However, the issue of antimicrobial resistance is becoming increasingly urgent and surpasses the negative effects of fungal and viral infections (Dadgostar, 2019).
Microbes like bacteria and fungi produce secondary metabolites that are effective against other microbes or particular physiological states in sick organisms. Particularly, bacteria are known for producing a wide variety of structurally distinct bioactive chemicals with powerful biological activity. Consequently, they are regarded as an intriguing source for such compounds (Mulani et al., 2019).
The production of new therapeutic compounds is possible through the bacteria present in seawater, sediments, and marine organisms (Mulani et al., 2019). These marine bacteria utilize nutrients from their host, such as carbon, to protect against environmental pollutants by releasing bioactive chemicals (Schultz et al., 2020).
In a recent study, Contreras-Castro et al. (2020) conducted a recent study on bacteria found in marine sediments in Punta Arena de la Ventana, Mexico. They utilized 16S rRNA gene identification and multilocus sequence analysis to identify 71 isolates as members of the Salinispora genus. The researchers then examined the antibacterial properties of various Salinispora spp., with 10 demonstrating strong inhibitory activity against ESKAPE infection (Stonik et al., 2020). Out of the tested isolates, 23 were found to be positive for one or more bacterial pathogens. Streptomyces sp. was also found to possess antibacterial activity against drug-resistant bacteria like E. faecium, Staphylococcus aureus, and A. baumannii. A new acidic fluorophore called chlorocaterin, which is a chlorinated catecholic acid, was discovered (Yang et al., 2023). Streptomyces has also shown the ability to produce biologically active compounds like lactic acid and indole-3, which exhibit potent antibacterial activity against C. albicans and Mucor mihei (Fijan et al., 2022).
Bacteroidia, Cytophaga, Flavobacteria, and Sphingobacteria represent the four major classes of Bacteroidetes. The bacteria are classified as cephalosporin-negative bacteria, bacteriophage bacteria, and non-sporulating bacteria and can survive in both aerobic and anaerobic environments (Li et al., 2019). Bacteroidetes bacteria are found not only in marine habitats such as sediments, seawater, and soil, but also in biological habitats such as animal skin and intestines (Huang, 2023).
Cyanobacteria are unique in that they are the sole type of bacteria that generate energy through photosynthesis. Additionally, they are the only group of photosynthetic prokaryotes that are able to generate oxygen and take in CO2 (Kariyazono et al., 2022). For a while now, marine cyanobacteria’s secondary metabolites have been recognized for their pharmacological traits, such as their ability to fight off bacteria, viruses, fungi, and more (Perera et al., 2023).
According to studies (Brumley et al., 2018), Fischerella ambigua, a cyanobacterium, has an antibacterial effect on Mycobacterium tuberculosis, Ambigane, and Misonitrile, with values of the minimum limiting concentration ranging from 6.6 to 7.5 μM. New (alkylphenols, anaephenes AC) were isolated from the cyanobacterium Hormoscilla sp. They demonstrated moderate inhibition of S. aureus growth (Giddings and Newman, 2022). Anaephenes A, B, and C were able to completely inhibit the visible growth of S. aureus at concentrations of 22, 6.1, and 22 μg/mL, respectively. When it comes to antiviral drugs isolated from marine bacteria, there are only a few compared to antibacterial and antifungal drugs. Glycolipids are heterosteric compounds that can be found in cyanobacteria and form thylakoid membranes in all prokaryotic and eukaryotic photosynthetic organisms (Brumley et al., 2018). Numerous studies have demonstrated that glycolipids are antiviral agents against the influenza virus (Stincone and Brandelli, 2020). Glycolipids were also found to be effective against HIV. For example, cyanobacteria (Phormidium tenue and Yngbya lagerheimii) secrete glycolipids containing sulfonic acid, which inhibit HIV-1 reverse transcriptase (Saurav et al., 2017). The cholesterol glycolipids of the cyanobacteria Oscillatoria laoin and Cytonema sp. secrete HIV-1 reverse transcriptase at a concentration of 10 μM (Yasir, 2018). Cyanobacterial lipoprotein complexes containing sulfoquinosylparanosillipids inhibit the enzymatic cleavage of HIV-1 virus. At an IC50 concentration of 24 nM, esom sulfonide inhibited the HIV-1 DNA polymerase reverse transcriptase pathway in humans. Calcium cyproline is an antiviral that protects against HIV-1, HPV-1, influenza A, and herpes simplex.
Recently, a new antibacterial compound based on oxygenated cycloketones was isolated from rivers containing B. strocoris. The isolated compounds inhibited the growth of aquatic bacteria of the genera Aeromonas and Vibrio. Furthermore, a secondary vaxillin antagonist metabolite has been discovered in the seaweed B. amyloliquefaciens MTCC 10456. Wang et al. have published a report on the antibacterial activity of a new thiopeptide antibiotic micrococcin, which was obtained from B. stratosphericus in a marine environment. This antibiotic has shown potent activity against Gram-positive bacterial pathogens at concentrations up to 10 μM (Kuo et al., 2019). Marine sponges are known to play a crucial role in benthic communities worldwide, as they can impact both biomass and pelagic processes (Maslin et al., 2021). These sponges are part of a diverse microbial community that includes bacteria, fungi, archaea, and viruses – all of which are valuable sources of natural products. These natural products can be produced by sponges themselves, their microbial symbionts, or through interactions between sponges and symbionts. Consequently, many researchers have focused on studying related bacteria for the purpose of screening and isolating biologically active compounds.
Altuğ et al. demonstrated that methanolic extracts of seaweed-associated bacteria are effective against various bacterial pathogens such as Staphylococcus aureus, Vibrio vulnificus, and Escherichia coli. Bacterial diseases were studied (Altuğ et al., 2021). About 22% of the biologically active compounds were isolated from marine fungi associated with Athymycota. A recent study by Fahmy et al. included the antibacterial compound isolated from Streptomyces sp. (Fahmy and Abdel-Tawab, 2021). Cyclopeptides isolated from fungi were applied to B. cerevisiae at a concentration of 25 μg/mL. In another study, the new thiopeptide antibiotics YM-266183 and YM-266184 were associated with the marine bacteria B. cereus QN03323 and the salmon Halychondria japonica. Moreover, both of these thiopeptide antibiotics have demonstrated good inhibitory activity against Gram-positive bacterial infections (Wang et al., 2020).
Pseudomonas, a marine bacteria found in association with Diginea sp. algae, yielded a novel cyclic tetrapeptide complex, cyclo-[phenylalanyl-prolyl-leucyl-prolyl], which exhibits potent inhibitory effects against V. anguillarum and B. subtilis (Rungprom et al., 2008). Avila and Angulo-Preckler demonstrated that P. tunicata-produced marine bacteria have a competitive edge over their fungi counterparts in surface colonization. Additionally, P. tunicata-derived yellow chemical molecule, tamjamine, is believed to possess antibacterial properties (Avila and Angulo-Preckler, 2020) that effectively suppress the growth of staphylococci and bacilli (Tan, 2023).
Another bioactive compound, 2,4-diacetylfloroglucin, was isolated from a new strain of Pseudomonas. Pseudomonas isolated from Padina tetrastromatica algae inhibited the growth of Gram-negative bacterial pathogens such as Pseudomonas aeruginosa and Pseudomonas aeruginosa at a dose of 300 mg (Arslan et al., 2021). Pelagiobacter variabilis, a new marine bacterium isolated from macroalgae, produces a number of chemical molecules of peragomycin from A to C (Srinivasan et al., 2021).
The content of secondary metabolites in marine bacteria exhibiting antibacterial activity is presented in Table 1.
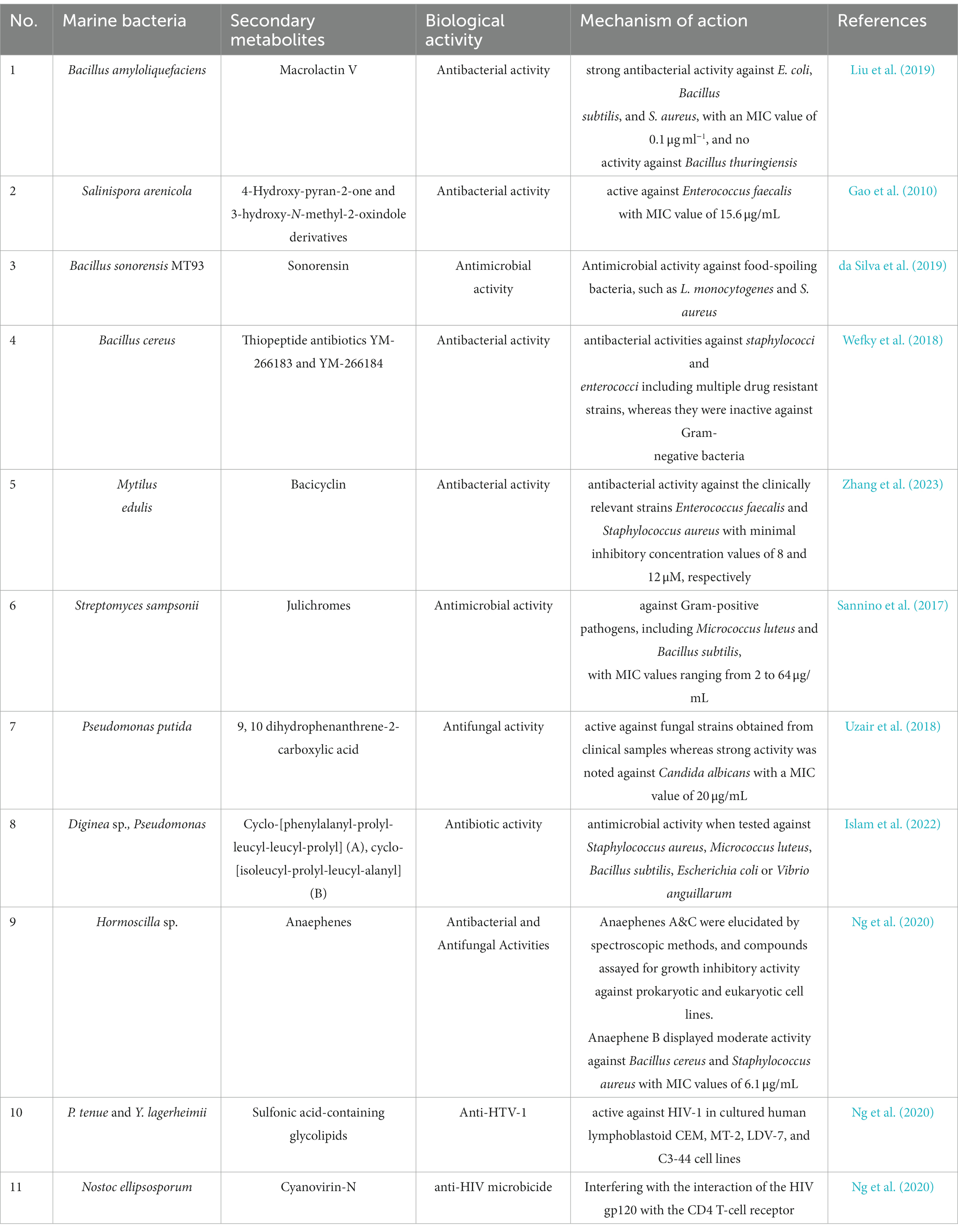
Table 1. Content of secondary metabolites of marine bacteria with antibacterial activity.
It is known that various mediators, such as prostaglandins, leukotrienes and kinins, platelet activation factors are involved in the development of inflammation. The study (Srilekha et al., 2017) presents the results of carrageenan injection from the marine bacterium Brevibacterium sp. in inflammation, indicating that the inflammatory mediator serotonin is released in phase 1, causing inflammation. This first phase occurs within the first hour after carrageenan injection. Prostaglandins, inflammatory mediators, are released in phase 2 and begin to exert their inflammatory effects around the third hour of the inflammatory process.
Oxidative stress plays an important role in endothelial dysfunction (Miksch et al., 2021), lung disease, gastrointestinal dysfunction, and atherosclerosis, all of which are associated with inflammatory responses (Ribeiro et al., 2023). The crude extract of pigment was used as an anti-inflammatory agent to induce paw edema in male Wistar rats. A significant reduction in edema and inflammation was noted. Marine bacteria are considered very valuable because they produce various antibiotics and other therapeutically useful compounds with diverse biological activities (Anguita-Maeso et al., 2020).
Chemical extracts of marine Actinobacteria isolates were screened for their antimicrobial and anti-inflammatory activities as described in (Ribeiro et al., 2023). Extracts of two Streptomyces strains showed activity against Candida albicans. In addition, 15 extracts (derived from marine microorganisms Brachybacterium, Brevibacterium, Microbacterium, Microbacterium, Rhodococcus, and Streptomyces) demonstrated anti-inflammatory potential in the RAW 264.4 cell model assay without concomitant cytotoxic response.
Elbandy discovered that neuroinflammation is closely linked to the development and progression of various neurodegenerative conditions such as Alzheimer’s disease, Huntington’s disease, and Parkinson’s disease (Elbandy, 2023). The activation of astrocytes and microglia serves as the brain’s defense mechanism against damaged tissue and harmful pathogens, but their chronic activation can lead to neuroinflammation, which in turn worsens or triggers neurodegeneration. Currently available therapeutic drugs only provide temporary relief for these disorders, and there are no treatments available that can slow down or stop the progression of neurodegeneration. Therefore, natural compounds that may have a protective effect against these disorders hold therapeutic potential. Marine sources have already yielded numerous chemical compounds such as bioactive peptides, fatty acids, pigments, alkaloids, and polysaccharides which possess anti-inflammatory properties and can be effective in treating and preventing neuroinflammatory diseases. The anti-inflammatory components found in marine compounds are utilized as functional food ingredients for preventing and treating neurological disorders (Elbandy, 2023).
A biofilm is an aggregation of microorganisms attached to a surface (O’Toole et al., 2000). It consists of microbial cells encapsulated in an extracellular polymeric matrix containing various biopolymers such as proteins, nucleic acids, lipids and other substances (Flemming and Wingender, 2010). Biofilms consist of one or more species of microorganisms colonizing biological surfaces (Hall-Stoodley et al., 2004). The complex biofilm structure protects microorganisms within the biofilm and provides the spatial proximity and internal communication necessary for growth and development (Dang and Lovell, 2016). This film can protect microbial cells from various external influences, such as antimicrobial therapy, toxins, antiprotozoal therapy, and host immune defense. For example, biological preparations reduce the sensitivity of microorganisms to antimicrobial drugs by 10–1,000 times. In addition, multispecies organisms are less susceptible to antimicrobial therapy than single-species organisms due to complex host-organism interactions (Hengzhuang et al., 2011).
In the marine environment, microorganisms rapidly colonize living and abiotic surfaces and subsequently form a biota consisting of bacteria, diatoms, fungi, unicellular algae and protozoa (Qian et al., 2007), which is a serious problem for humans. Marine biomass plays an important role in the biodegradation of the marine environment. Biomass plays an important role in the habitat selection and distribution of many surface-dwelling marine species. For example, invertebrate larvae can differentiate between biomes composed of different microbial community structures that may or may not form colonies (Dobretsov and Qian, 2006). Various substances are broken down by microbial metabolites such as hydrogen sulfide, various acids, and ammonia. Every year, biofouling and biodegradation cause significant economic losses worldwide in industries such as heat transfer, oil and gas processing, storage, and transportation, and drinking water and wastewater treatment (De Carvalho, 2018; Plaza and Achal, 2020). Furthermore, the persistence and spread of harmful or pathogenic microorganisms and their genetic material in marine biota poses a serious threat to humans (Hengzhuang et al., 2011).
Given the increasing economic losses and potential risks of marine biofilm formation, effective and efficient control mechanisms are needed. Antibiotic/antioxidant coatings are now widely used and provide a simple method of controlling the marine environment. However, chemical inhibitors such as amines, amides, organotin compounds and copper oxides used in traditional adhesives are toxic and harmful, have no favorable environmental impact and are prone to biological oxidation (Figure 1). Special attention is paid to the development of effective and safe adhesives using natural products. Natural compounds with antimicrobial activity produced by the metabolic mechanism of microorganisms can replace traditional chemical and biological agents and have environmentally friendly properties such as low toxicity and biodegradability. However, insufficient funding and complexity limit the development of naturally synthesizable composites (Ohno et al., 1978).
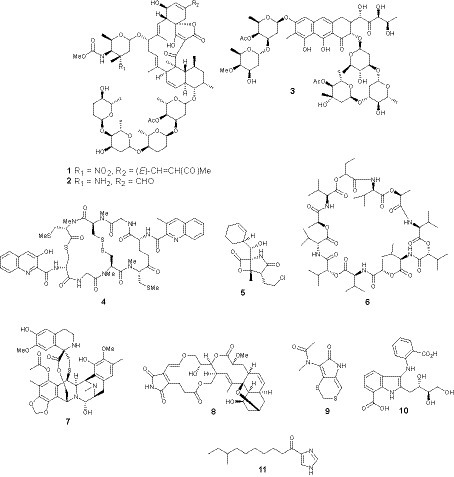
Figure 1. Bioactive metabolites isolated from marine bacteria.
Bacteria, as major colonizers in marine biota, are important determinants of the structure and function of mature biomass (De Carvalho, 2018). Thus, marine biofilm bacteria may be an excellent target for the discovery of compounds that inhibit biofilm formation. Compounds that inhibit biofilm formation are being sought using bacteria isolated from marine biofilms, leading to the discovery of potent activity of elasnin against biofilms. The structural formula of elasnin is shown in Figure 2.
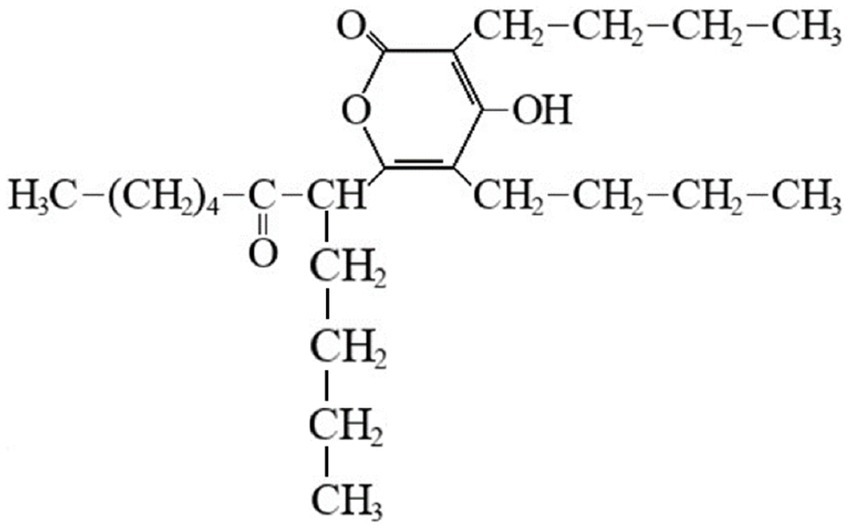
Figure 2. Structural formula of elasnin.
In a screening program of antibiofilm compounds against marine biofilms, a potent biofilm inhibitory activity of elasnin was found (Hengzhuang et al., 2011). Elasnin effectively inhibited biofilm formation of seven bacterial strains isolated from marine biofilms. With high performance, elastin-based coatings were fabricated in a simple and cost-effective manner that showed excellent results in suppressing the formation of multispecies biofilms and attachment of large biofouling organisms in marine environments. 16S-amplicon and anti-larvae assays showed that elasnin could prevent biofouling through indirect effects of altered microbial biofilm composition and direct inhibitory effects on larval settlement with low toxic effects. These results indicate the potential application of elasnin for biofilm and biofouling control in marine environments (Hengzhuang et al., 2011).
Ten strains isolated from marine biofilms (Vibrio alginolyticus B1, Erythrobacter sp.HKB8, Rugeria B32, Staphylococcus aureus B04, Staphylococcus hominis N32, Staphylococcus arlettae OM, Microbacterium asteromaticum N22, Idiomarina sediminum N28, Pseudoalteromonas L001, and Escherichia coli N57) were used as targets in the analysis of minimum biofilm inhibitory concentrations (MIC). MBIC is the lowest concentration of a compound that specifically reduces the number of attached cells, whereas MIC is the lowest concentration required to inhibit significant planktonic cell growth. Seven of the ten strains tested formed biofilms successfully during the experiment, while three strains (V. alginolyticus B1, Erythrobacter sp. HKB8, and Rugeria B32) failed to form biofilms under the experimental conditions. For the seven biofilm-forming strains, the biofilms of the four Gram-positive bands were sensitive to arginine levels: MBIC 90 was between 2.5 and 5 mg/mL, and MBIC 50 was between 1.25 and 5 mg/mL. Both MBIC 90 and MBIC 50 g were sensitive to enzyme action in a concentration-dependent manner. Enzyme concentrations ranged from 5 to 10 mg/mL and 1.25 to 10 mg/mL, respectively. The BICs of nine strains were determined, except for M. asteraromaticum N22, which failed to grow under the experimental conditions. Elasnin inhibited the growth of planktonic cells of S. aureus B04 and I. sediminum N28 with BICs between 5 and 10 mg/mL, whereas the BIC of elasnin exceeded 10 mg/mL for the other species. Overall, elasnin inhibits biofilm formation more effectively than it does antibacterial activity.
The dependence of the degree of biofilm eradication on elastin concentration is presented in Figure 3.
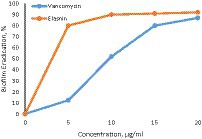
Figure 3. Dependence of biofilm eradication degree on elastin concentration (vancomycin – control).
As a result, a highly effective elasnin-based coating was developed and its activity against a wide range of natural biological agents was evaluated in clinical trials (Long et al., 2021).
Elasnin was first discovered in 1978 as a novel elastase inhibitor with low toxicity to rats and high selectivity for human granulomatous elastase (Nsanzabana, 2019), but its antibacterial and antibiotic activity was not detected at that time. According to the experimental results, elasnin not only inhibited the formation of single and multi-organism biofilms, but also inhibited macroorganism colonization. 16S-amplicon analysis suggests that the altered biome may not promote microbial colonization, so elasnin-based biofilms may reduce coat biodiversity, indirectly limiting biocolonization. In addition, antibody assays showed that elasnin inhibited larval colonization with low toxicity. Considering the effective concentration of elasnin and its mobility in the marine environment, the toxic effects of elasnin should be ignored. Elasnin shows great potential for biocide and biological control applications in the marine environment due to its low toxicity, high efficacy and high productivity (Hengzhuang et al., 2011).
More than 40% of the world’s population now lives in areas where malaria is common. There were 219 million clinical cases of malaria in 2017, and 435,000 people died from the disease (Lackner et al., 2017). Existing antimalarials quickly become ineffective due to the rapid emergence of resistant bacteria. Artemisinin therapy is recommended for the treatment of malaria even in malaria-endemic countries, but widespread resistance has been shown to be alarming in Southeast Asia (Newman and Cragg, 2020). Given the alarming situation in malaria treatment, there is an urgent need for new chemotherapeutic agents for next-generation antimicrobial therapy.
Throughout history, natural products have played a crucial role in uncovering new potential for pharmaceuticals. Between 1981 and 2014, a total of 1,562 fresh drugs were created, with 59.5% being derived from natural raw materials (WHO, 2020). Natural products are particularly valuable as sources of anti-cancer and antimicrobial agents, such as quinine-based antimalarials. Artemisinin, which was initially used in phytotherapy and is derived from pepper, has recently emerged as a leading treatment for drug-resistant malaria. Even though most approved natural products and naturally occurring chemicals come from terrestrial habitats, the marine environment remains largely uncharted. This presents an opportunity to discover new therapeutic compounds from a variety of marine organisms. Invertebrates and algae have traditionally been the primary targets for discovering new therapeutic medications, but marine biotechnology is now focusing on marine microbes (Domínguez-Oliva et al., 2023).
Many compounds originally isolated from marine invertebrates are thought to come from marine bacteria (McCarthy et al., 2019). Marine surfaces usually contain 10 5 microbial cells per mL (Nielsen et al., 2017). The so-called microbial decoctions of some algae contain a large number of related microorganisms with a density of 108 to 1010/g, which amounts to 20 to 30% of the mollusc biomass. There is growing evidence indicating that these communities of microorganisms play a crucial role in the metabolism of molluscs, including the production of natural substances (Nicoletti et al., 2018). Manzamin A, an antiviral substance produced by actinomycetes associated with fungi, is one of the earliest examples of mollusc metabolites that have been demonstrated to be produced by cultivated microorganisms (Ruiz-Gil et al., 2020). Marine microorganisms have been found to produce various antibacterial compounds. For instance, Pseudomonas (Masschelein et al., 2017) associated with sponges produces 2-undecyl-4-quinolone while Calothrix (Dyshlovoy and Honecker, 2018), a type of cyanobacterium, produces calotrixins A and B. Despite the potential of the marine microbiome for discovering new chemotypes, it has not been fully utilized for discovering new antimalarials.
A study (Bunbamrung et al., 2020) mentions that the phylogenetic relationships of 2,290 heterotrophic marine bacterial isolates were determined by analyzing restriction fragment length polymorphism and 16SU rRNA gene sequences (Meyer et al., 2020). The resulting similarity was <97%. Common fungi isolated from the sea, such as Penicillium and Aspergillus, have been found to produce new metabolites (Xu et al., 2020).
Marine bacteria that have antimalarial activity come from many taxa. There are five known isolates of Penicillium fungi (Tajuddeen and Van Heerden, 2019). There is a wealth of information available on terrestrial penicillins and other naturally occurring fungi-derived products (Trial.gov, C, 2023). Recently, however, marine isolates of these fungi have become a source of new natural antimalarial products (Álvarez-Bardón et al., 2020). The same applies to common mangrove endophytes, which have been found to produce a wide range of natural antimalarial products, including xantoquinidin-like tritilachium compounds isolated from marine fungi (Bank, D, 2019).
Six active marine actinomycetes (Gram-positive bacteria) usually produce biologically active antimalarial products (Miksch et al., 2021). In screening studies, six isolates of marine Gram-negative bacteria were found to have antimalarial activity (da Silva et al., 2019). Recently, marine gram-negative bacteria have become recognized as a valuable source for antimalarial products, including polyketides, non-ribosomal peptides, and polyketide-neribosomal hybrid peptides (Wefky et al., 2018). Of the three active isolates discovered, all belonged to the genus Marinobacter and demonstrated the ability to produce the siderophore petrobactin (Dyshlovoy and Honecker, 2018).
The oil-degrading bacterium Alcanivorax is known to produce the active ingredients and α-pyrone alkaniborone (ClinicalTrials.gov, 2023a), but the biological activity of these molecules has not been reported. Endozoiconado spp. can be associated with marine fungi, sponges, and octocorals and can dominate microbial populations. Although their secondary metabolites have not been studied, one article describes the antimalarial activity of endozoicomonad extracts (Martyniuk et al., 2020).
These results support the hypothesis that marine microbes can produce new antimalarial chemicals. Based on literature review (Bank, D, 2019; ClinicalTrials.gov, 2023a; DrugBank Online, 2023a), the following marine organisms were found to have significant antimalarial activity: Streptomyces sp., Nocardiopsis sp., Micromonospora sp., Unidentified actinomycete, Penicillium sp., Endozoicomonas numazuensis, Alcanivorax sp., Marinobacter sp., Talaromyces rotundus, and Tritirachium sp. Taxonomic diversity is achieved among individuals with high levels of activity, and it is expected to develop into chemical diversity of antimalarials as research progresses (ClinicalTrials.gov, 2023a). Table 2 presents the activity of various marine microorganisms against malaria.
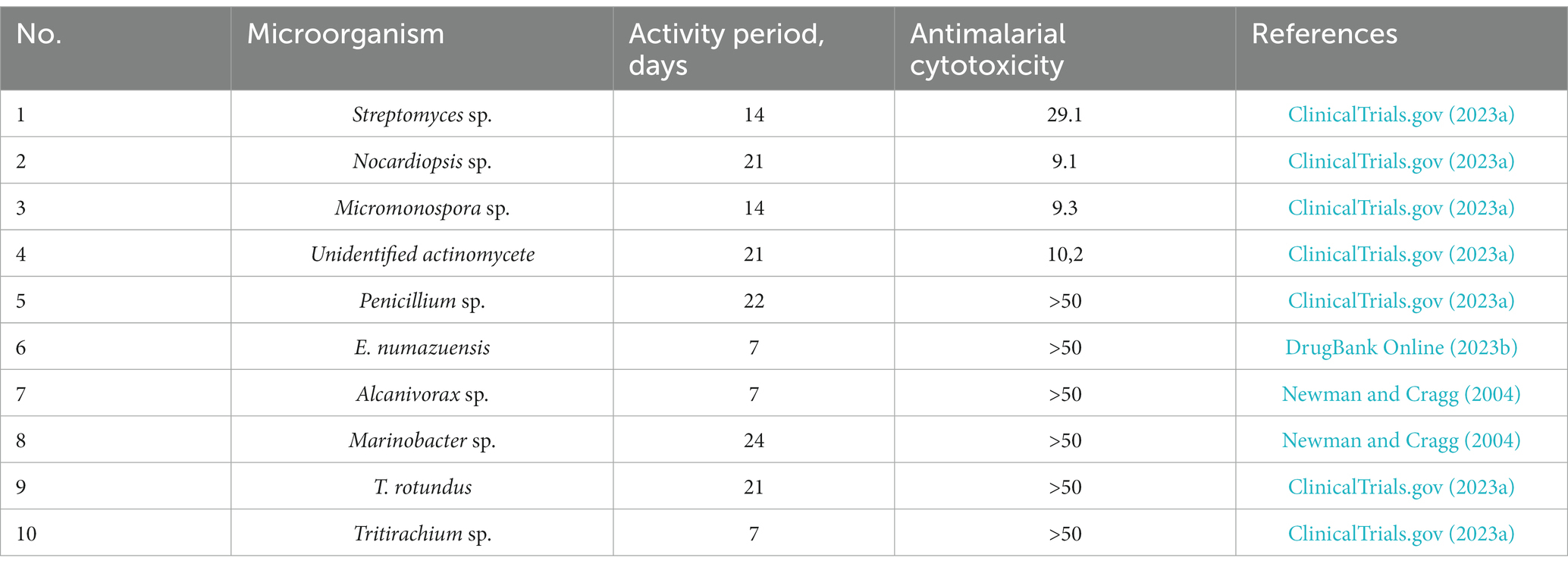
Table 2. Antimalarial activity of marine microorganisms.
It has been found that bioactive molecules derived from nature have the ability to eliminate cancer cells by targeting their macromolecules in oncogenic signaling pathways (Bank, D, 2019). Numerous metabolites obtained from the ocean have demonstrated the ability to impede the growth of human tumor cells in both laboratory and animal models (specifically rats), as well as in clinical trials (ClinicalTrials.gov, 2023a; DrugBank Online, 2023a). By utilizing advanced technology and natural marine products, researchers have come across a new generation of anticarcinogenic agents that are currently undergoing clinical testing (DrugBank Online, 2023b). Marine microbes possess immense potential for the discovery of new substances that can be advantageous for cancer prevention and treatment. The diversity of bacteria found in natural marine products is increasingly attracting attention for its ability to overcome challenges in drug discovery (Newman and Cragg, 2004). In general, natural products serve as a significant source of compounds that can effectively treat various forms of cancer and present opportunities for exploring novel mechanisms of action (Khotimchenko, 2010).
Live non-pathogenic marine bacterial species can selectively proliferate in tumors and inhibit their growth. More than 70 compounds belonging to different structural classes of marine microorganisms, such as polyketides, indolocarbozoles, isoprenoids, macrolides, non-ribosomal proteins, etc., have been proven to have antitumor effects on various cell lines (Zong et al., 2023). Due to their selectivity towards tumor tissues, these marine bacteria and their spores (Zong et al., 2023) also serve as ideal vectors for the delivery of therapeutic proteins to tumors. Marine bacterial toxins have also emerged as a promising strategy for cancer treatment (Rommasi, 2022). The most potential and promising strategy is marine bacteria-based gene-directed enzymatic prodrug therapy. This method has been shown to be effective in the treatment of cancer in vivo (Jha and Zi-rong, 2004). Marine microorganisms produce unusual macrolactins that inhibit proliferation of melanoma B16-F10 cells in rodents, suppress replication of mammalian herpes simplex viruses, and protect T lymphocytes from human immunodeficiency virus (HIV) (DrugBank Online, 2023c). A schematic overview of the role of marine bacteria in cancer therapy is presented in Figure 4.
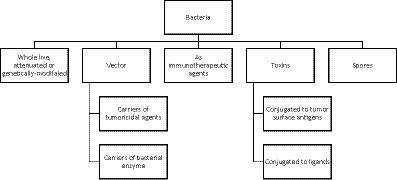
Figure 4. Schematic overview of the role of marine bacteria in cancer therapy.
Over the course of the last half-century, data has indicated that a variety of natural products derived from marine plants and microbes (as listed in Table 3) have promising potential for preventing and treating cancer. This includes medications such as cytarabine, eribulin mesylate, brentuximab vedotin, and trabectedin, which have been utilized to treat leukemia, breast cancer, soft tissue sarcoma, and cervical cancer (ClinicalTrials.gov, 2023b). To accelerate the development of new anti-tumor drugs that are highly effective with minimal side effects from marine resources, a comprehensive integrated approach has been proposed. The marine organisms examined in this review (Streptomyces sp., Aplidium albicans, Nostoc sp., Dolabella auricularia, Ecteinascidia turbinate, Halichondria okadai, Bryopsis sp., Elysia rufescens, Sponges, Mollusc) had high anticancer activity (Table 3).
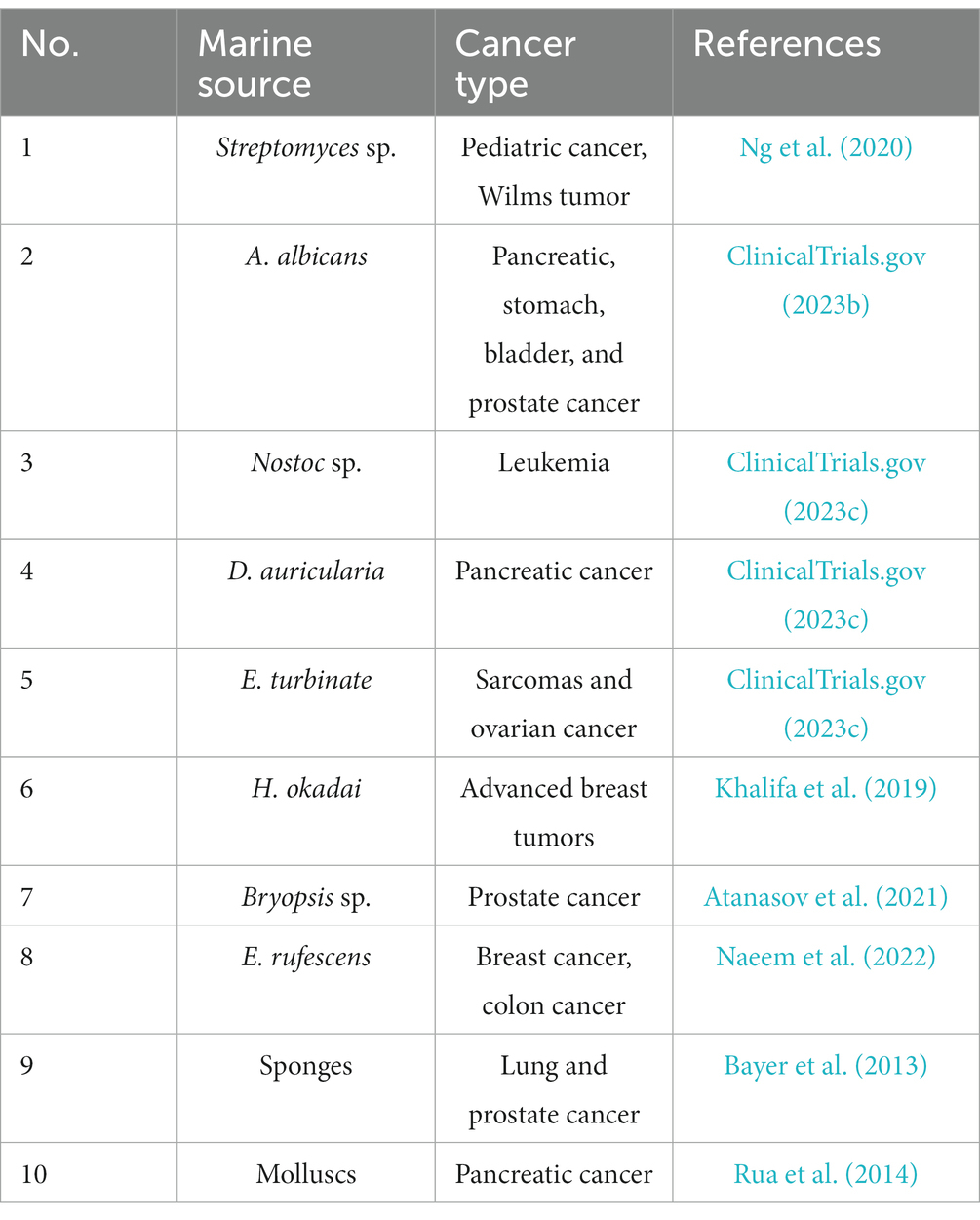
Table 3. Potential anti-cancer effects of marine sources.
A research study has reported the identification of a vast number of microorganisms, approximately 3.7 × 1030 derived from marine organisms (Pike et al., 2013). Although most of these bacteria cannot be grown in laboratory conditions, they have the ability to produce numerous natural compounds with potential medical applications (Nobili et al., 2009). The impressive chemical and pharmacological capabilities observed in marine organisms can be attributed to their adaptation to extreme conditions, which prompts them to synthesize secondary metabolites that are resistant to heat, salt, stress, and predators. In the past century, various marine organisms including bacteria, actinomycetes, cyanobacteria, fungi, microbes, macromycetes (Shahidi and Santhiravel, 2022), invertebrates, molluscs, soft corals, sea sponges, sea urchins, holojamberts, shellfish, and other sea creatures have been discovered and extensively investigated for their anticancer properties (Bhanot et al., 2011). Furthermore, advancements in marine chemistry have facilitated the utilization of metabolomics and other novel tools to gain new insights into aquatic products (Obaid et al., 2023).
Microbes are responsible for the production of more than 22,000 secondary metabolites, with actinomycetes accounting for 70%, fungi for 20%, Bacillus spp. for 7%, and other bacteria for 1–2% (Rocha-Martin et al., 2014). However, only a small portion (10%) of biologically active natural products can be attributed to microbial sources. Several anticancer drugs, including briostatin 1, ET-743, and dolastatin 10, have undergone successful clinical trials. Briostatin 1 recently completed phase II clinical trials and showed efficacy in treating melanoma, Hodgkin’s lymphoma, kidney cancer, and colorectal cancer (Boopathy and Kathiresan, 2010). Its mechanism of action involves promoting the proliferation of normal osteoblasts (White et al., 2014). ET-743 is a tetrahydroisoquinolone alkaloid derived from erythropoietin that has passed phase I clinical trials due to its selective targeting of guanine residues in the small DNA groove. Dolabella aureus, a mollusc peptide, has progressed to phase II clinical trials by inhibiting microbial aggregation and inducing cell cycle metaphase arrest (White et al., 2014).
Cyanobacteria, also known as blue-green algae, offer a diverse range of more than 400 newly discovered metabolites. Among these metabolites, distinct bioactive peptides and polyketides (Bayona et al., 2022) have demonstrated the ability to induce apoptosis in cancer cells and positively impact cellular signal transduction through the protein kinase C family (Ngamcharungchit et al., 2023). In experiments involving 41 cyanobacterial strains, approximately half exhibited the capability to induce cell death in cancer cells (Ngamcharungchit et al., 2023). Dolastin 10 and curazine A, derived from cyanobacteria and possessing antibacterial properties, are currently being clinically evaluated for their potential in cancer therapy and have served as foundational structures for the development of several synthetic analogues and derivatives (Gao et al., 2021). Calothrix A and B are five metabolites isolated from Cyanobacterium calothrix that exhibit potent anti-tumor activity against human HeLa cancer cells in virological studies, with IC50 values of 40 and 350 nM, respectively. Additionally, Uritiaciclamide synthesized by the cyanobacteria Prochromon spp. and pateramide synthesized by Lissoclinum patella have demonstrated significant cytotoxic activity against human nasopharyngeal carcinoma cell lines, with IC50 values of 17 and 3,000 ng/mL, respectively, (Premarathna et al., 2024).
Borofins, which are metabolites containing boron, have been obtained from the marine cyanobacteria Nostoc linckia and Nostoc spongiaeforme var. tenue (Encarnação et al., 2015); these compounds have shown strong cytotoxic effects against human epidermal carcinoma (LOVO) and human colorectal adenocarcinoma (KB) cell lines. The septic toxicity of Nostoc sp. has also been observed in human tumor cells (GSV 224) and tumor cells, with an IC50 value of 0.005 for KB and LOVO cell lines. The Leptolyngbya strain exhibits significant cytotoxicity (LC50 < 23 nM) against NCIH460 cells and rat lung neuronal cells. These cells have the ability to regulate enzymatic kinases and inhibit the growth of human fibroblasts and endothelial cells. Curacin A, extracted from organic materials found in the Lyngbya majuscule collection in Curaçao, is a notable antiviral agent with strong potency. It can prevent tubulin polymerization and selectively inhibit Burkitt leukemic lymphocytes (Robles-Bañuelos et al., 2022). Apratocin is another cyanobacterial compound that shows inhibition of various cancer cells at nanomolar doses.
Autophagic activity against chronic myelogenous leukocytes was observed in various strains of cyanobacteria, while benign cells such as hepatocytes and cardiomyocytes remained unaffected (Tan, 2007). Current research indicates that benthic cyanobacteria cultivated in a temperate marine environment hold great promise as a previously untapped source of novel anti-leukemia drugs (Khotimchenko, 2010). Nonetheless, certain compounds isolated from marine sources, including calcroxin A, B, euletiaclamide, betylamide, borofin, and largasol, have not yet undergone clinical trials and exhibit distinct modes of action. Consequently, further investigation is required to determine their potential biological activity and potential for clinical application.
Biologically active compounds derived from marine Pseudomonas are diverse and include pyrroles, pseudoanes, pyrrolidenediones, floroglucines, phenazines, benzaldehydes, quinines, quinolones, phenanthenes, phthalates, anhydrides, moiramides, chaffrin, and bouchrin. Some of the biologically active compounds in these antimicrobials are dibutyl phthalate and di (2-ethylhexyl) phthalate, which were reported to be cathepsin B inhibitors (Zong et al., 2023). Discodermold, briostatin, sarcoidin, and eletrobin are among the antibiotics produced mainly by marine bacteria (Naeem et al., 2022).
Based on the generalization of literature sources, it was found that marine microorganisms are a more valuable source of biologically active secondary metabolites than terrestrial ones. This is attributed to the fact that marine organisms have therefore survived all periods of evolution and have acquired the ability to produce a variety of compounds with unique structural and functional properties (Barzkar et al., 2017, 2018, 2019, 2021a,b,c, 2022a,b, 2023a,b; Jahromi and Barzkar, 2018a,b; Barzkar, 2020; Barzkar and Sohail, 2020).
However, only 179 new bioactive compounds have been characterized in marine bacteria. Therefore, the relevance of studying the types and properties of secondary metabolites in marine microorganisms is beyond doubt. The scientific novelty of the study was that for the first time, data on new resources for obtaining biologically active natural productsmetabolites of marine bacteria and fungi - were generalized. The review presents information on types of natural products derived from marine microorganisms (bacteria and fungi), summarizes data on the antibacterial, antimalarial, anticarcinogenic, and anti-inflammatory effects of marine microorganisms. Properties and mechanisms of antibacterial action of the following marine microorganisms have been studied: Pseudomonas sp., M. producens, B. amyloliquefaciens, S. arenicola, B. sonorensis MT93, B. cereus, M. edulis, S. sampsonii, P. putida, Diginea sp., Pseudomonas, Hormoscilla sp., P. tenue and Y. Lagerheimii, and N. ellipsosporum. Therefore, there is a great need for scientific and applied research on the bioactive secondary metabolites of marine microorganisms from the viewpoint of human and animal health.
NB: Conceptualization, Writing – original draft, Writing – review & editing. SS: Writing – review & editing. OB: Writing – original draft, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Altuğ, G., Çiftçi Türetken, P. S., Kalkan, S., and Topaloğlu, B. (2021). The distribution and antibacterial activity of marine sponge-associated bacteria in the Aegean Sea and the sea of Marmara Turkey. Curr. Microbiol. 78, 2275–2290. doi: 10.1007/s00284-021-02489-7
Álvarez-Bardón, M., Pérez-Pertejo, Y., Ordóñez, C., Sepúlveda-Crespo, D., Carballeira, N. M., Tekwani, B. L., et al. (2020). Screening marine natural products for new drug leads against Trypanosomatids and malaria. Mar. Drugs 18:187. doi: 10.3390/md18040187
Ameen, F., AlNadhari, S., and Al-Homaidan, A. A. (2021). Marine microorganisms as an untapped source of bioactive compounds. Saudi J. Biol. Sci. 28, 224–231. doi: 10.1016/j.sjbs.2020.09.052
Anguita-Maeso, M., Olivares-García, C., Haro, C., Imperial, J., Navas-Cortés, J. A., and Landa, B. B. (2020). Culture-dependent and culture-independent characterization of the olive xylem microbiota: effect of sap extraction methods. Front. Plant Sci. 10:1708. doi: 10.3389/fpls.2019.01708
Arslan, E., Çobanoğlu, Ş., and Yazici, A. (2021). Antimicrobial activity of pigments extracted from Auxenochlorella protothecoides SC3 against Pseudomonas aeruginosa Tr. J. Nature Sci. 10, 163–167. doi: 10.46810/tdfd.930388
Atanasov, A. G., Zotchev, S. B., and Dirsch, V. M. (2021). Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 20, 200–216. doi: 10.1038/s41573-020-00114-z
Avila, C., and Angulo-Preckler, C. (2020). Bioactive compounds from marine Heterobranchs. Mar. Drugs 18:657. doi: 10.3390/md18120657
Barone, G. (2022). Local environmental conditions promote high turnover diversity of benthic Deep-Sea Fungi in the Ross Sea (Antarctica). J. Fungi 8:65 8, 65. doi: 10.3390/jof8010065
Barzkar, N. (2020). Marine microbial alkaline protease: an efficient and essential tool for various industrial applications. Int. J. Biol. Macromol. 161, 1216–1229. doi: 10.1016/j.ijbiomac.2020.06.072
Barzkar, N., Attaran, F. G., and Taheri, A. (2017). Proximate composition and mineral contents in the body wall of two species of sea cucumber from Oman Sea. Environ. Sci. Pollut. Res. 24, 18907–18911. doi: 10.1007/s11356-017-9379-5
Barzkar, N., Homaei, A., Hemmati, R., and Patel, S. (2018). Thermostable marine microbial proteases for industrial applications: scopes and risks. Extremophiles 22, 335–346. doi: 10.1007/s00792-018-1009-8
Barzkar, N., Jahromi, S. T., Poorsaheli, H. B., and Vianello, F. (2019). Metabolites from marine microorganisms, micro, and macroalgae: immense scope for pharmacology. Mar. Drugs 17:464. doi: 10.3390/md17080464
Barzkar, N., Jahromi, S. T., and Vianello, F. (2022a). Marine microbial fibrinolytic enzymes: an overview of source, production, biochemical properties and thrombolytic activity. Mar. Drugs 20:46. doi: 10.3390/md20010046
Barzkar, N., Khan, Z., Jahromi, S. T., Pourmozaffar, S., Gozari, M., and Nahavandi, R. (2021a). A critical review on marine serine protease and its inhibitors: a new wave of drugs? Int. J. Biol. Macromol. 170, 674–687. doi: 10.1016/j.ijbiomac.2020.12.134
Barzkar, N., Rungsardthong, V., Tamadoni Jahromi, S., Laraib, Q., das, R., Babich, O., et al. (2023a). A recent update on fucoidonase: source, isolation methods and its enzymatic activity. Front. Mar. Sci. 10:1129982. doi: 10.3389/fmars.2023.1129982
Barzkar, N., Sukhikh, S., Babich, O, Venmathi Maran, B. A., and Tamadoni Jahromi, S. (2023b). Marine collagen: purification, properties and application Front. Mar. Sci 10, 1245077. doi: 10.3389/fmars.2023.1245077
Barzkar, N., Sheng, R., Sohail, M., Jahromi, S. T., Babich, O., Sukhikh, S., et al. (2022b). Alginate lyases from marine bacteria: an enzyme ocean for sustainable future. Molecules 27:3375. doi: 10.3390/molecules27113375
Barzkar, N., and Sohail, M. (2020). An overview on marine cellulolytic enzymes and their potential applications. Appl. Microbiol. Biotechnol. 104, 6873–6892. doi: 10.1007/s00253-020-10692-y
Barzkar, N., Sohail, M., Tamadoni, J. S., Gozari, M., Poormozaffar, S., Nahavandi, R., et al. (2021b). Marine bacterial esterases: emerging biocatalysts for industrial applications. Appl. Biochem. Biotechnol. 193, 1187–1214. doi: 10.1007/s12010-020-03483-8
Barzkar, N., Sohail, M., Tamadoni, J. S., Nahavandi, R., and Khodadadi, M. (2021c). Marine microbial l-glutaminase: from pharmaceutical to food industry. Appl. Microbiol. Biotechnol. 105, 4453–4466. doi: 10.1007/s00253-021-11356-1
Bayer, T., Neave, M. J., Alsheikh-Hussain, A., Aranda, M., Yum, L. K., Mincer, T., et al. (2013). The microbiome of the Red Sea coral Stylophora pistillata is dominated by tissue-associated Endozoicomonas bacteria. Appl. Environ. Microbiol. 79, 4759–4762. doi: 10.1128/AEM.00695-13
Bayona, L. M., de Voogd, N. J., and Choi, Y. H. (2022). Metabolomics on the study of marine organisms. Metabolom. Off. J. Metabol. Soc. 18:17.
Bhanot, A., Sharma, R., and Noolvi, M. N. (2011). Natural sources as potential anti-cancer agents: a review. Int. J. Phytomed. 3, 9–26.
Blunt, J. W., Carroll, A. R., Copp, B. R., Davis, R. A., Keyzers, R. A., and Prinsep, M. R. (2018). Marine natural products. Nat. Prod. Rep. 35, 8–53. doi: 10.1039/C7NP00052A
Boopathy, N. S., and Kathiresan, K. (2010). Anticancer drugs from marine flora: an overview. J. Oncol. 2010, 1–18. doi: 10.1155/2010/214186
Brumley, D., Spencer, K. A., Gunasekera, S. P., Sauvage, T., Biggs, J., Paul, V. J., et al. (2018). Isolation and characterization of Anaephenes A–C, Alkylphenols from a filamentous cyanobacterium (Hormoscillasp., Oscillatoriales). J. Nat. Prod. 81, 2716–2721. doi: 10.1021/acs.jnatprod.8b00650
Bunbamrung, N., Intaraudom, C., Dramae, A., Komwijit, S., Laorob, T., Khamsaeng, S., et al. (2020). Antimicrobial, antimalarial and anticholinesterase substances from the marine-derived fungus aspergillus terreus BCC51799. Tetrahedron 76:131496. doi: 10.1016/j.tet.2020.131496
ClinicalTrials.gov. (2023a). Available at: https://clinicaltrials.gov/NCT00003677.
ClinicalTrials.gov. (2023b). AvailPable at: https://clinicaltrials.gov/NCT01669252.
ClinicalTrials.gov. (2023c). Avaialble at: https://clinicaltrials.gov/NCT00884845.
Carroll, A. R., Copp, B. R., Davis, R. A., and Keyzers, R. A. (2023). Prinsepf marine natural products. Nat. Prod. Rep. 40, 275–325. doi: 10.1039/D2NP00083K
Che, Q., Qiao, L., Han, X., Liu, Y., Wang, W., Gu, Q., et al. (2018). Anthranosides A–C, anthranilate derivatives from a sponge-DerivedStreptomycessp. CMN-62. Org. Lett. 20, 5466–5469. doi: 10.1021/acs.orglett.8b02382
Cho, E., Kwon, O.-S., Chung, B., Lee, J., Sun, J., Shin, J., et al. (2020). Antibacterial activity of Chromomycins from a marine-derived Streptomyces microflavus. Mar. Drugs 18:522. doi: 10.3390/md18100522
Chung, D., Nguyen, H., Yu, N. H., Yu, W.-J., Kwon, Y. M., Bae, S. S., et al. (2023). In vitro and in vivo antimicrobial activity of the fungal metabolite toluquinol against phytopathogenic bacteria. Front. Microbiol. 14:1221865. doi: 10.3389/fmicb.2023.1221865
Contreras-Castro, L., Martinez-Garcia, S., Cancino-Diaz, J. C., Maldonado, L. A., Hernandez-Guerrero, C. J., Martínez-Díaz, S. F., et al. (2020). Inhibits the growth of emerging bacterial pathogens and other multi-drug-resistant Bacteria. Pol. J. Microbiol. 69, 321–330. doi: 10.33073/pjm-2020-035
DrugBank Online. (2023a). AVailable at: http://www.drugbank.ca/DB05109.
DrugBank Online. (2023b). Available at: http://www.drugbank.ca/DB08871
DrugBank Online. (2023c). Available at: http://www.drugbank.ca/DB05158
da Silva, A. B., Pinto, F. C., Silveira, E. R., Costa-Lotufo, L. V., Costa, W. S., Ayala, A. P., et al. (2019). 4-Hydroxy-pyran-2-one and 3-hydroxy-N-methyl-2-oxindole derivatives of Salinispora arenicola from Brazilian marine sediments. Fitoterapia 138:104357. doi: 10.1016/j.fitote.2019.104357
Dadgostar, P. (2019). Antimicrobial resistance: implications and costs. Infect. Drug Resist. 12, 3903–3910. doi: 10.2147/IDR.S234610
Dang, H., and Lovell, C. R. (2016). Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 80, 91–138. doi: 10.1128/MMBR.00037-15
De Carvalho, C. C. C. R. (2018). Marine biofilms: a successful microbial strategy with economic implications. Front. Mar. Sci. 5:5. doi: 10.3389/fmars.2018.00126
de Sá, J. D. M., Kumla, D., Dethoup, T., and Kijjoa, A. (2022). Bioactive compounds from terrestrial and marine-derived Fungi of the genus Neosartorya. Molecules 27:2351. doi: 10.3390/molecules27072351
Deshmukh, S. K., Prakash, V., and Ranjan, N. (2018). Marine Fungi: a source of potential anticancer compounds. Front. Microbiol. 8:2536. doi: 10.3389/fmicb.2017.02536
Dobretsov, S., and Qian, P.-Y. (2006). Facilitation and inhibition of larval attachment of the bryozoan Bugula neritina in association with mono-species and multi-species biofilms. J. Exp. Mar. Biol. Ecol. 333, 263–274. doi: 10.1016/j.jembe.2006.01.019
Domínguez-Oliva, A., Hernández-Ávalos, I., Martínez-Burnes, J., Olmos-Hernández, A., Verduzco-Mendoza, A., and Mota-Rojas, D. (2023). The importance of animal models in biomedical research: current insights and applications. Animals 13:1223. doi: 10.3390/ani13071223
Dyshlovoy, S. A., and Honecker, F. (2018). Marine compounds and cancer: 2017 updates. Mar. Drugs 16:41. doi: 10.3390/md16020041
Elbandy, M. (2023). Anti-inflammatory effects of marine bioactive compounds and their potential as functional food ingredients in the prevention and treatment of Neuroinflammatory disorders. Molecules 28:2. doi: 10.3390/molecules28010002
El-Hossary, E. M., Cheng, C., Hamed, M. M., Hamed, A. N. E.-S., Ohlsen, K., Hentschel, U., et al. (2017). Antifungal potential of marine natural products. Eur. J. Med. Chem. 126, 631–651. doi: 10.1016/j.ejmech.2016.11.022
Encarnação, T., Pais, A. A., Campos, M. G., and Burrows, H. D. (2015). Cyanobacteria and microalgae: a renewable source of bioactive compounds and other chemicals. Sci. Prog. 98, 145–168. doi: 10.3184/003685015X14298590596266
Fahmy, N. M., and Abdel-Tawab, A. M. (2021). Isolation and characterization of marine sponge–associated Streptomyces sp. NMF6 strain producing secondary metabolite (s) possessing antimicrobial, antioxidant, anticancer, and antiviral activities. J. Genet. Eng. Biotechnol. 19, 1–14. doi: 10.1186/s43141-021-00203-5
Fijan, S., Kocbek, P., Steyer, A., Vodiˇcar, P. M., and Strauss, M. (2022). The antimicrobial effect of various single-strain and multi-strain probiotics, dietary supplements or other beneficial microbes against common clinical wound pathogens. Microorganisms 10:2518. doi: 10.3390/microorganisms10122518
Flemming, H. C., and Wingender, J. (2010). The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633. doi: 10.1038/nrmicro2415
Gao, C. H., Tian, X. P., Qi, S. H., Luo, X. M., Wang, P., and Zhang, S. (2010). Antibacterial and antilarval compounds from marine gorgonian-associated bacterium Bacillus amyloliquefaciens SCSIO 00856. J. Antibiot. 63, 191–193. doi: 10.1038/ja.2010.7
Gao, G., Wang, Y., Hua, H., Li, D., and Tang, C. (2021). Marine antitumor peptide Dolastatin 10: biological activity, structural modification and synthetic chemistry. Mar. Drugs 19:363. doi: 10.3390/md19070363
Giddings, L. A., and Newman, D. J. (2022). Extremophilic Fungi from marine environments: underexplored sources of antitumor, anti-infective and other biologically active agents. Mar. Drugs 20:62. doi: 10.3390/md20010062
Gladfelter, A. S., James, T. Y., and Amend, A. S. (2019). Marine fungi. Curr. Biol. 29, R191–R195. doi: 10.1016/j.cub.2019.02.009
Gui, C., Zhang, S., Zhu, X., Ding, W., Huang, H., Gu, Y.-C., et al. (2017). Antimicrobial spirotetronate metabolites from marine-derived Micromonospora harpali SCSIO GJ089. J. Nat. Prod. 80, 1594–1603. doi: 10.1021/acs.jnatprod.7b00176
Hafez Ghoran, S., Taktaz, F., Sousa, E., Fernandes, C., and Kijjoa, A. (2023). Peptides from marine-derived Fungi: chemistry and biological activities. Mar. Drugs 21:510. doi: 10.3390/md21100510
Hall-Stoodley, L., Costerton, J. W., and Stoodley, P. (2004). Bacterial biofilms: from the natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108. doi: 10.1038/nrmicro821
Hanif, N., Murni, A., Tanaka, C., and Tanaka, J. (2019). Marine natural products from Indonesian waters. Mar. Drugs 17:364. doi: 10.3390/md17060364
Hengzhuang, W., Wu, H., Ciofu, O., Song, Z., and Hoiby, N. (2011). Pharmacokinetics/pharmacodynamics of colistin and imipenem on mucoid and nonmucoid Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 55, 4469–4474. doi: 10.1128/AAC.00126-11
Huang, H. (2023). Bacteroidetes. Encyclopedia. Available at: https://encyclopedia.pub/entry/32508 (Accessed: 06 November 2023).
Islam, S., Farjana, M., Uddin, M. R., Akter, S., Jabin, A., Nafisa, H. T., et al. (2022). Molecular identification, characterization, and antagonistic activity profiling of Bacillus cereus LOCK 1002 along with the in-silico analysis of its presumptive bacteriocins. J. Adv. Vet. Anim. Res. 9, 663–675. doi: 10.5455/javar.2022.i635
Jahromi, S. T., and Barzkar, N. (2018a). Future direction in marine bacterial agarases for industrial applications. Appl. Microbiol. Biotechnol. 102, 6847–6863. doi: 10.1007/s00253-018-9156-5
Jahromi, S. T., and Barzkar, N. (2018b). Marine bacterial chitinase as sources of energy, eco-friendly agent, and industrial biocatalyst. Int. J. Biol. Macromol. 120, 2147–2154. doi: 10.1016/j.ijbiomac.2018.09.083
Jha, R. K., and Zi-rong, X. (2004). Biomedical compounds from marine organisms. Mar. Drugs 2, 123–146. doi: 10.3390/md203123
Ji, X., Guo, J., Liu, Y., Lu, A., Wang, Z., Li, Y., et al. (2018). Marine-natural-product development: first discovery of nortopsentin alkaloids as novel antiviral, anti-phytopathogenic-fungus, and insecticidal agents. J. Agric. Food Chem. 66, 4062–4072. doi: 10.1021/acs.jafc.8b00507
Jiménez, C. (2018). Marine natural products in medicinal chemistry. ACS Med. Chem. Lett. 9, 959–961. doi: 10.1021/acsmedchemlett.8b00368
Karim, M. R. U., Harunari, E., Oku, N., Akasaka, K., and Igarashi, Y. (2020). Bulbimidazoles A–C, antimicrobial and cytotoxic Alkanoyl Imidazoles from a marine Gammaproteobacterium MicrobulbiferSpecies. J. Nat. Prod. 83, 1295–1299. doi: 10.1021/acs.jnatprod.0c00082
Kariyazono, R., Ito, S., and Osanai, T. (2022). Carbon metabolism of great biotechnological interest: Metabolic engineering and synthetic biology of cyanobacteria. Cyanobact. Physiol., 189–200. doi: 10.1016/B978-0-323-96106-6.00003-4
Karthikeyan, A., Joseph, A., and Nair, B. G. (2022). Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 20:14. doi: 10.1186/s43141-021-00290-4
Khalifa, S. A., Elias, N., Farag, M. A., Chen, L., Saeed, A., Hegazy, M.-E. F., et al. (2019). Marine natural products: a source of novel anticancer drugs. Mar. Drugs 17:491. doi: 10.3390/md17090491
Khotimchenko, Yu. S., Biologically active substances from marine aquatic organisms - a source of new pharmaceutical substances and drugs. TMG. (2010). 2. Avaialble at: https://cyberleninka.ru/article/n/biologicheski-aktivnye-veschestva-iz-morskih-gidrobiontov-istochnik-novyh-farmatsevticheskih-substantsiy-i-lekarstv (Accessed: 21 November, 2023).
Kuo, J., Yang, Y.-T., Lu, M.-C., Wong, T.-Y., Sung, P.-J., and Huang, Y.-S. (2019). Antimicrobial activity and diversity of bacteria associated with Taiwanese marine sponge Theonella swinhoei. Ann. Microbiol. 69, 253–265. doi: 10.1007/s13213-018-1414-3
Lackner, G., Peters, E. E., Helfrich, E. J., and Piel, J. (2017). Insights into the lifestyle of uncultured bacterial natural product factories associated with marine sponges. Proc. Natl. Acad. Sci. U. S. A 114, E347–E356. doi: 10.1073/pnas.1616234114
Li, Z., Wang, Y., Li, X., Lin, Z., Lin, Y., Srinivasan, R., et al. (2019). The characteristics of antibiotic resistance and phenotypes in 29 outer-membrane protein mutant strains in Aeromonas hydrophila. Environ. Microbiol. 21, 4614–4628. doi: 10.1111/1462-2920.14761
Liu, K., Ding, H., Yu, Y., and Chen, B. (2019). A cold-adapted chitinase-producing bacterium from Antarctica and its potential in biocontrol of plant pathogenic fungi. Mar. Drugs 17:695. doi: 10.3390/md17120695
Long, L., Wang, R., Chiang, H. Y., Ding, W., Li, Y. X., Chen, F., et al. (2021). Discovery of Antibiofilm activity of Elasnin against marine biofilms and its application in the marine antifouling coatings. Mar. Drugs 19:19. doi: 10.3390/md19010019
Makarieva, T. N., Silchenko, A. S., Kicha, A. A., Lyakhova, E. G., Kolesnikova, S. A., Shubina, L. K., et al. (2014). Search and isolation of new natural compounds from marine invertebrates, studies of their structures and biological activities. Bull. Far East. Branch Russ. Acad. Sci. 1
Martyniuk, A. O., Skrotska, O. I., Shevchuk, T. A., and Pirog, T. P. (2020). Practically valuable metabolites of marine microorganisms. Biotechnol. Acta :3.
Maslin, M., Gaertner-Mazouni, N., Debitus, C., Joy, N., and Ho, R. (2021). Marine sponge aquaculture towards drug development: an ongoing history of technical, ecological, chemical considerations and challenges. Aquacult. Rep. 21:100813. doi: 10.1016/j.aqrep.2021.100813
Masschelein, J., Jenner, M., and Challis, G. L. (2017). Antibiotics from gram-negative bacteria: a comprehensive overview and selected biosynthetic highlights. Nat. Prod. Rep. 34, 712–783. doi: 10.1039/C7NP00010C
McCarthy, P. J., Roberts, B. F., Carbonell, A., Roberts, J., Wright, A. E., and Chakrabarti, D. (2019). Marine microbiome as a source of antimalarials. Trop. Med. Infect. Dis. 4:103. doi: 10.3390/tropicalmed4030103
Meyer, V., Basenko, E. Y., and Benz, J. P. (2020). Growing a circular economy with fungal biotechnology: a white paper. Biol. Biotechnol. 7:5. doi: 10.1186/s40694-020-00095-z
Miksch, S., Meiners, M., Meyerdierks, A., Probandt, D., Wegener, G., and Titschack, J. (2021). Bacterial communities in temperate and polar coastal sands are seasonally stable. ISME Commun. 1, 1–11. doi: 10.1038/s43705-021-00028-w
Mohie, T., El-dien, R., Mahmoud, B. K., and Abdelwahab, M. F. (2023). Paralemnalia thyrsoides-associated fungi: phylogenetic diversity, cytotoxic potential, metabolomic profiling and docking analysis. BMC Microbiol. 23:308. doi: 10.1186/s12866-023-03045-y
Mulani, M. S., Kamble, E. E., Kumkar, S. N., Tawre, M. S., and Pardesi, K. R. (2019). Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: a review. Front. Microbiol. 10:539. doi: 10.3389/fmicb.2019.00539
Naeem, A., Hu, P., Yang, M., Zhang, J., Liu, Y., Zhu, W., et al. (2022). Natural products as anticancer agents: current status and future perspectives. Molecules 27:8367. doi: 10.3390/molecules27238367
Newman, D. J., and Cragg, G. M. (2004). Advanced preclinical and clinical trials of natural products and related compounds from marine sources. Curr. Med. Chem. 11, 1693–1713. doi: 10.2174/0929867043364982
Newman, D. J., and Cragg, G. M. (2020). Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 83, 770–803. doi: 10.1021/acs.jnatprod.9b01285
Ng, Z. J., Zarin, M. A., Lee, C. K., and Tan, J. S. (2020). Application of bacteriocins in food preservation and infectious disease treatment for humans and livestock: a review. RSC Adv. 10, 38937–38964. doi: 10.1039/D0RA06161A
Ngamcharungchit, C., Chaimusik, N., Panbangred, W., Euanorasetr, J., and Intra, B. (2023). Bioactive metabolites from terrestrial and marine Actinomycetes. Molecules 28:5915. doi: 10.3390/molecules28155915
Nicoletti, R., Salvatore, M. M., and Andolfi, A. (2018). Secondary metabolites of mangrove-associated strains of Talaromyces. Mar. Drugs 16:12. doi: 10.3390/md16010012
Nielsen, J. C., Grijseels, S., Prigent, S., Ji, B., Dainat, J., Nielsen, K. F., et al. (2017). Global analysis of biosynthetic gene clusters reveals vast potential of secondary metabolite production in Penicillium species. Nat. Microbiol. 2, 1–9. doi: 10.1038/nmicrobiol.2017.44
Nobili, S., Lippi, D., Witort, E., Donnini, M., Bausi, L., Mini, E., et al. (2009). Natural compounds for cancer treatment and prevention. Pharmacol. Res. 59, 365–378. doi: 10.1016/j.phrs.2009.01.017
Nsanzabana, C. (2019). Resistance to artemisinin combination therapies (ACTs): do not forget the partner drug! Trop. Med. Infect. Dis. 4:26. doi: 10.3390/tropicalmed4010026
Nugraha, A. S., Firli, L. N., and Rani, D. M. (2023). Indonesian marine and its medicinal contribution. Nat. Prod. Rep. 13:38. doi: 10.1007/s13659-023-00403-1
O’Toole, G., Kaplan, H. B., and Kolter, R. (2000). Biofilm formation as microbial development. Annu. Rev. Microbiol. 54, 49–79. doi: 10.1146/annurev.micro.54.1.49
Obaid, H. M., Sale, S. S., and Boundenga, L. (2023). Pharmaceutical activity of a synthetic heterocyclic (C15H12N5OCl) compound on Entamoeba histolytica and Giardia lamblia. Russ. J. Infect. Immun. 13, 119–126. doi: 10.15789/2220-7619-PAO-2024
Ohno, H., Saheki, T., Awaya, J., Nakagawa, A., and Omura, S. (1978). Isolation and characterization of elasnin, a new human granulocyte elastase inhibitor produced by a strain of Streptomyces. J. Antibiot. 31, 1116–1123. doi: 10.7164/antibiotics.31.1116
Packiavathy, I. A. S. V., Kannappan, A., Thiyagarajan, S., Srinivasan, R., Jeyapragash, D., Paul, J. B. J., et al. (2021). AHL-lactonase producing Psychrobacter sp. from Palk Bay sediment mitigates quorum sensing-mediated virulence production in gram negative bacterial pathogens. Front. Microbiol. 12:634593. doi: 10.3389/fmicb.2021.634593
Pedrosa, R., Gaudêncio, P., and Vasconcelos, S. (2020). XVI international symposium on marine natural products| XI European conference on marine natural products. Mar. Drugs 18:40.
Perera, R. M. T. D., Herath, K. H. I. N. M., Sanjeewa, K. K. A., and Jayawardena, T. U. (2023). Recent reports on bioactive compounds from marine Cyanobacteria in relation to human health applications. Life (Basel, Switzerland) 13:1411. doi: 10.3390/life13061411
Pike, R. E., Haltli, B., and Kerr, R. G. (2013). Description of Endozoicomonas euniceicola sp. nov. and Endozoicomonas gorgoniicola sp. nov., bacteria isolated from the octocorals Eunicea fusca and Plexaura sp., and an emended description of the genus Endozoicomonas. Int. J. Syst. Evol. Microbiol. 63, 4294–4302. doi: 10.1099/ijs.0.051490-0
Plaza, G., and Achal, V. (2020). Biosurfactants: eco-friendly and innovative biocides against biocorrosion. Int. J. Mol. Sci. 21:2152. doi: 10.3390/ijms21062152
Poli, A., Finore, I., Romano, I., Gioiello, A., Lama, L., and Nicolaus, B. (2017). Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 5:25. doi: 10.3390/microorganisms5020025
Premarathna, A. D., Ahmed, T. A. E., Kulshreshtha, G., Humayun, S., Darko, C. N. S., Rjabovs, V., et al. (2024). Polysaccharides from red seaweeds: effect of extraction methods on physicochemical characteristics and antioxidant activities. Food Hydrocoll. 147:109307. doi: 10.1016/j.foodhyd.2023.109307
Puttaswamygowda, G. H., Olakkaran, S., Antony, A., and Purayil, A. K. (2019). Present status and future perspectives of marine Actinobacterial metabolites. Recent Dev. Appl. Microbiol. Biochem., 307–319. doi: 10.1016/B978-0-12-816328-3.00022-2
Qian, P. Y., Lau, S. C., Dahms, H. U., Dobretsov, S., and Harder, T. (2007). Marine biofilms as mediators of colonization by marine macroorganisms: implications for antifouling and aquaculture. Mar. Biotechnol. 9, 399–410. doi: 10.1007/s10126-007-9001-9
Ribeiro, I., Antunes, J. T., Alexandrino, D. A. M., Tomasino, M. P., Almeida, E., Hilário, A., et al. (2023). Actinobacteria from Arctic and Atlantic deep-sea sediments-biodiversity and bioactive potential. Front. Microbiol. 14:1158441. doi: 10.3389/fmicb.2023.1158441
Robles-Bañuelos, B., Durán-Riveroll, L. M., Rangel-López, E., Pérez-López, H. I., and González-Maya, L. (2022). Marine Cyanobacteria as sources of Lead anticancer compounds: a review of families of metabolites with cytotoxic, Antiproliferative, and antineoplastic effects. Molecules 27:4814. doi: 10.3390/molecules27154814
Rocha-Martin, J., Harrington, C., Dobson, A. D., and O’Gara, F. (2014). Emerging strategies and integrated systems microbiology technologies for biodiscovery of marine bioactive compounds. Mar. Drugs 12, 3516–3559. doi: 10.3390/md12063516
Rommasi, F. (2022). Bacterial-based methods for Cancer treatment: what we know and where we are. Oncol. Ther. 10, 23–54. doi: 10.1007/s40487-021-00177-x
Rua, C. P., Trindade-Silva, A. E., Appolinario, L. R., Venas, T. M., Garcia, G. D., Carvalho, L. S., et al. (2014). Diversity and antimicrobial potential of culturable heterotrophic bacteria associated with the endemic marine sponge Arenosclera brasiliensis. Peer J 2:e419. doi: 10.7717/peerj.419
Ruginescu, R., Enache, M., Popescu, O., Gomoiu, I., Cojoc, R., Microorgan Batrinescu-Moteau, C., et al. (2022). Characterization of some salt-tolerant bacterial hydrolases with potential utility in cultural heritage bio-cleaning. Nat. Prod. Rep. 10:644. doi: 10.3390/microorganisms10030644
Ruiz-Gil, T., Acuña, J. J., Fujiyoshi, S., Tanaka, D., Noda, J., Maruyama, F., et al. (2020). Airborne bacterial communities of outdoor environments and their associated influencing factors. Environ. Int. 145:106156. doi: 10.1016/j.envint.2020.106156
Rungprom, W., Siwu, E. R., Lambert, L. K., Dechsakulwatana, C., Barden, M. C., Kokpol, U., et al. (2008). Cyclic tetrapeptides from marine bacteria associated with the seaweed Diginea sp. and the sponge Halisarca ectofibrosa. Tetrahedron 64, 3147–3152. doi: 10.1016/j.tet.2008.01.089
Sannino, F., Parrilli, E., Apuzzo, G. A., de Pascale, D., Tedesco, P., Maida, I., et al. (2017). Pseudoalteromonas haloplanktis produces methylamine, a volatile compound active against Burkholderia cepacia complex strains. New Biotechnol. 35, 13–18. doi: 10.1016/j.nbt.2016.10.009
Saurav, K., Costantino, V., Venturi, V., and Steindler, L. (2017). Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 15:53. doi: 10.3390/md15030053
Schofield, M. M., Jain, S., Porat, D., Dick, G. J., and Sherman, D. H. (2015). Identification and analysis of the bacterial endosymbiont specialized for production of the chemotherapeutic natural product ET-743. Environ. Microbiol. 17, 3964–3975. doi: 10.1111/1462-2920.12908
Schultz, F., Anywar, G., Tang, H., Chassagne, F., Lyles, J. T., Garbe, L.-A., et al. (2020). Targeting ESKAPE pathogens with anti-infective medicinal plants from the greater Mpigi region in Uganda. Sci. Rep. 10, 1–19. doi: 10.1038/s41598-020-67572-8
Shafiekhani, M., Shekari, Z., and Boorboor, A. (2022). Bacterial and fungal co-infections with SARS-CoV-2 in solid organ recipients: a retrospective study. Virol. J. 19:35. doi: 10.1186/s12985-022-01763-9
Shahidi, F., and Santhiravel, S. (2022). Novel marine bioactives: application in functional foods, nutraceuticals, and pharmaceuticals. J. Food Bioact. 19:19. doi: 10.31665/JFB.2022.18316
Srilekha, V., Krishna, G., Seshasrinivas, V., and Charya, M. A. S. (2017). Antibacterial and anti-inflammatory activities of marine Brevibacterium sp. Res. Pharm. Sci. 12, 283–289. doi: 10.4103/1735-5362.212045
Srinivasan, R., Kannappan, A., Shi, C., and Lin, X. (2021). Marine bacterial secondary metabolites: a treasure house for structurally unique and effective antimicrobial compounds. Mar. Drugs 19:530. doi: 10.3390/md19100530
Stincone, P., and Brandelli, A. (2020). Marine bacteria as source of antimicrobial compounds. Crit. Rev. Biotechnol. 40, 306–319. doi: 10.1080/07388551.2019.1710457
Stonik, V. A., Makarieva, T. N., and Shubina, L. K. (2020). Antibiotics from marine Bacteria. Biochem. Mosc. 85, 1362–1373. doi: 10.1134/S0006297920110073
Sun, W., Wu, W., Liu, X., Zaleta-Pinet, D. A., and Clark, B. R. (2019). Bioactive compounds isolated from marine-derived microbes in China: 2009–2018. Mar. Drugs 17:339. doi: 10.3390/md17060339
Tajuddeen, N., and Van Heerden, F. R. (2019). Antiplasmodial natural products: an update. Malar. J. 18:404. doi: 10.1186/s12936-019-3026-1
Tan, L. T. (2007). Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 68, 954–979. doi: 10.1016/j.phytochem.2007.01.012
Tan, L. T. (2023). Impact of marine chemical ecology research on the discovery and development of new pharmaceuticals. Mar. Drugs 21:174. doi: 10.3390/md21030174
Uzair, B., Bano, A., Niazi, M. B. K., Khan, F., and Mujtaba, G. (2018). In vitro antifungal activity of 9, 10-dihydrophenanthrene-2-carboxylic acid isolated from a marine bacterium: Pseudomonas putida. Pak. J. Pharm. Sci. 31, 2733–2736.
Wang, W., Park, K. H., Lee, J., Oh, E., Park, C., Kang, E., et al. (2020). A new Thiopeptide antibiotic, Micrococcin P3, from a marine-derived strain of the bacterium Bacillus stratosphericus. Molecules. Basel, Switzerland 25:4383. doi: 10.3390/molecules25194383
Wefky, S., Hassan, M., and Antibacterial, (2018). Anticoagulant and anti-inflammatory activities of marine Bacillus cereus S1. J. Pure Appl. Microbiol. 10, 2593–2606. doi: 10.22207/JPAM.10.4.15
White, M. C., Holman, D. M., Boehm, J. E., Peipins, L. A., Grossman, M., and Henley, S. J. (2014). Age and cancer risk: a potentially modifiable relationship. Am. J. Prev. Med. 46, S7–S15. doi: 10.1016/j.amepre.2013.10.029
WHO (2020). World malaria report: 20 years of global progress and challenges. World malaria report 2020: 20 years of global progress and challenges 2020, 299
Xu, P., Ding, L., Wei, J., Li, Q., Gui, M., He, X., et al. (2020). A new aquatic pathogen inhibitor produced by the marine fungus aspergillus sp. LS116. Aquaculture 520:734670. doi: 10.1016/j.aquaculture.2019.734670
Xu, J.-L., Liu, H.-X., Chen, Y.-C., Tan, H.-B., Guo, H., Xu, L.-Q., et al. (2018). Highly substituted benzophenone aldehydes and Eremophilane derivatives from the Deep-Sea derived fungus Phomopsis lithocarpus FS508. Mar. Drugs 16:329. doi: 10.3390/md16090329
Yang, Y., Kessler, M. G. C., Marchán-Rivadeneira, M. R., and Han, Y. (2023). Combating antimicrobial resistance in the post-genomic era: rapid antibiotic discovery. Molecules 28:4183. doi: 10.3390/molecules28104183
Yasir, M. (2018). Analysis of bacterial communities and characterization of antimicrobial strains from cave microbiota. Braz. J. Microbiol. 49, 248–257. doi: 10.1016/j.bjm.2017.08.005
Ye, X., Anjum, K., Song, T., Wang, W., Liang, Y., Chen, M., et al. (2017). Antiproliferative cyclodepsipeptides from the marine actinomycete Streptomyces sp. P11-23B downregulating the tumor metabolic enzymes of glycolysis, glutaminolysis, and lipogenesis. Phytochemistry 135, 151–159. doi: 10.1016/j.phytochem.2016.12.010
Zhang, Z., Wang, P., Chen, M., Xie, L., Zhang, X., Shi, Y., et al. (2023). Antibacterial activity of two new Cassane Diterpenoids from Caesaplinia pulcherrima against Bacillus cereus by damage to cell membrane. Int. J. Mol. Sci. 24:4917.
Zhang, B., Wang, K. B., Wang, W., Bi, S. F., Mei, Y. N., Deng, X. Z., et al. (2018). Discovery, biosynthesis, and heterologous production of streptoseomycin, an anti-microaerophilic bacteria macrodilactone. Org. Lett. 20, 2967–2971. doi: 10.1021/acs.orglett.8b01006
Zhao, P., Xue, Y., and Li, J. (2019). Non-lipopeptide fungi-derived peptide antibiotics developed since 2000. Biotechnol. Lett. 41, 651–673. doi: 10.1007/s10529-019-02677-3
Zhou, W., Liang, H., Qin, X., Cao, D., Zhu, X., Ju, J., et al. (2020). The isolation of Pyrroloformamide congeners and characterization of their biosynthetic gene cluster. J. Nat. Prod. 83, 202–209. doi: 10.1021/acs.jnatprod.9b00321
Keywords: marine bacteria and fungi, metabolites for disease prevention and treatment, biological activity, bioactive compounds, human health
Citation: Barzkar N, Sukhikh S and Babich O (2024) Study of marine microorganism metabolites: new resources for bioactive natural products. Front. Microbiol. 14:1285902. doi: 10.3389/fmicb.2023.1285902
Edited by:
Danny Ionescu, Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), GermanyReviewed by:
Pau Loke Show, University of Nottingham Malaysia Campus, MalaysiaCopyright © 2024 Barzkar, Sukhikh and Babich. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Noora Barzkar, bm9vcmEuYmFyemthckBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.