 Karma L. Bhutia1*
Karma L. Bhutia1* Mahtab Ahmad1
Mahtab Ahmad1 Anima Kisku1
Anima Kisku1 R. A. Sudhan1Nangsol D. Bhutia2
R. A. Sudhan1Nangsol D. Bhutia2 V. K. Sharma1
V. K. Sharma1 Bishun Deo Prasad1
Bishun Deo Prasad1 Mahendar Thudi1Oliver Obročník3Viliam Bárek3Marian Brestic4,5Milan Skalicky5Ahmed Gaber6
Mahendar Thudi1Oliver Obročník3Viliam Bárek3Marian Brestic4,5Milan Skalicky5Ahmed Gaber6 Akbar Hossain7*
Akbar Hossain7*- 1Department of Agricultural Biotechnology and Molecular Biology, CBS&H, Dr. Rajendra Prasad Central Agricultural University, Pusa, Bihar, India
- 2College of Horticulture and Forestry, Central Agricultural University (Imphal), Pasighat, Arunachal Pradesh, India
- 3Department of Water Resources and Environmental Engineering, Faculty of Horticulture and Landscape Engineering, Slovak University of Agriculture, Nitra, Slovakia
- 4Institute of Plant and Environmental Sciences, Slovak University of Agriculture, Nitra, Slovakia
- 5Department of Botany and Plant Physiology, Faculty of Agrobiology, Food, and Natural Resources, Czech University of Life Sciences Prague, Prague, Czechia
- 6Department of Biology, College of Science, Taif University, Taif, Saudi Arabia
- 7Division of Soil Science, Bangladesh Wheat and Maize Research Institute, Dinajpur, Bangladesh
Introduction: The yield of chickpea is severely hampered by infection wilt caused by several races of Fusarium oxysporum f. sp. ciceris (Foc).
Methods: To understand the underlying molecular mechanisms of resistance against Foc4 Fusarium wilt, RNA sequencing-based shoot transcriptome data of two contrasting chickpea genotypes, namely KWR 108 (resistant) and GL 13001 (susceptible), were generated and analyzed.
Results and Discussion: The shoot transcriptome data showed 1,103 and 1,221 significant DEGs in chickpea genotypes KWR 108 and GL 13001, respectively. Among these, 495 and 608 genes were significantly down and up-regulated in genotypes KWR 108, and 427 and 794 genes were significantly down and up-regulated in genotype GL 13001. The gene ontology (GO) analysis of significant DEGs was performed and the GO of the top 50 DEGs in two contrasting chickpea genotypes showed the highest cellular components as membrane and nucleus, and molecular functions including nucleotide binding, metal ion binding, transferase, kinase, and oxidoreductase activity involved in biological processes such as phosphorylation, oxidation–reduction, cell redox homeostasis process, and DNA repair. Compared to the susceptible genotype which showed significant up-regulation of genes involved in processes like DNA repair, the significantly up-regulated DEGs of the resistant genotypes were involved in processes like energy metabolism and environmental adaptation, particularly host-pathogen interaction. This indicates an efficient utilization of environmental adaptation pathways, energy homeostasis, and stable DNA molecules as the strategy to cope with Fusarium wilt infection in chickpea. The findings of the study will be useful in targeting the genes in designing gene-based markers for association mapping with the traits of interest in chickpea under Fusarium wilt which could be efficiently utilized in marker-assisted breeding of chickpea, particularly against Foc4 Fusarium wilt.
1. Introduction
Chickpea (Cicer arietinum L) is a protein and nutrient-enriched pulse crops, cultivated in different parts of the world, with an area of about 15 million hectares (Mha) and an annual production of 16 million tons (MT; FAOSTAT, 2023). India contributes more than 75% of the world’s chickpea production (around 12 MT @ 1,192 kg ha−1) in an area of around 10 Mha (Indiastat, 2023). The demand for protein and other nutrient-dense foods like pulses is increasing to meet the food and nutritional security of the growing world population. However, the area and production of chickpea have only increased marginally in the past few decades. One of the reasons for the antagonistic production and productivity of chickpea is Fusarium wilt disease, caused by Fusarium oxysporum f. sp. ciceris (Foc). Fusarium wilt is a devastating biotic stress that causes up to 100% yield loss in chickpea (Jendoubi et al., 2017) if the proper disease management practices are not followed.
The Fusarium wilt disease is very difficult to manage by conventional methods like the application of fungicides, crop rotation, etc., as the nature of the pathogen is soil- and seed-borne and it can survive in the soil for up to 6 years without a host (Haware et al., 1996). However, most of the time, farmers’ preference is higher yield over disease resistance. Several high-yielding varieties that are preferred by farmers are susceptible to Fusarium wilt. Hence, the cultivation of wilt-resistant varieties of chickpea is the best way to overcome yield and production losses. Cultivation of one resistant variety over a long time in the same field may also lead to a breakdown of the resistance mechanism by pathogens, as many of these mechanisms are controlled by one major gene or oligogenes (Adugna, 2004). The different races of this pathogen cause wilt disease and other related symptoms in chickpea across chickpea-growing countries. The wilting symptom in chickpea is caused by specific races of this pathogen, namely 1A, 2, 3, 4, 5, and 6 (Jendoubi et al., 2017).
Therefore, understanding the molecular mechanisms behind the resistance to Fusarium wilt is a positive step toward the development of chickpea genotypes with better yield and quality, along with resistance to diseases. To understand the underlying molecular mechanisms in chickpea plants for resistance against Fusarium wilt, various approaches, including “omics” tools, are being utilized, such as cDNA-RAPD and cDNA-AFLP techniques (Gupta et al., 2009), small RNA sequencing to identify microRNAs (Kohli et al., 2014), genome-wide transcriptome profiling (Sharma et al., 2016), and a long SAGE transcriptome approach to understanding host-pathogen interaction (Upasani et al., 2017). Similarly, studies on root transcriptome, marker-trait association, marker-based DNA profiling, and race-specific marker development against several races of Foc were conducted (Yadava et al., 2023) and these reported several underlying genes and pathways involved in host-pathogen interaction and resistance mechanisms. However, most of these studies were conducted with races 1, 2, 3, and 5 of Foc and with different genotypic backgrounds of the chickpea.
Considering the above burning issues, the present study was conducted with two contrasting genotypes of chickpea (one highly resistant and one highly susceptible) against Foc4, aiming to reveal the important molecular pathways that are associated with Fusarium wilt resistance and the genes involved in it. The identified genes could be targeted to design candidate gene-based markers to be utilized in marker-trait association (MTA) studies. The MTA studies will enable us to understand how these genes are directly or indirectly imparting resistance against Fusarium wilt via different traits; that information will ultimately help to develop chickpea genotypes by using marker-assisted breeding, particularly against Foc4.
2. Materials and methods
2.1. Experimental details
Two chickpea genotypes, namely KWR 108 and GL 13001, were selected for the study as these two varieties were reported to be highly contrasting in response to Fusarium wilt (Kumar et al., 2019), particularly to Foc4. The experiment was conducted in the Fusarium wilt sick plot of TCA, Dholi farm (a designated plot for chickpea Fusarium wilt pathological trials from the All India Co-ordinated Research Projects on Chickpea located at TCA-Dholi under the Department of Agricultural Biotechnology and Molecular Biology, CBS&H, Dr. Rajendra Prasad Central Agricultural University, Pusa (Samastipur) 848,125, Bihar, India). F. oxysporum isolates were collected from the field of TCA, Dholi and identified as Foc4 (Sharma et al., 2014). Among these chickpea genotypes, KWR 108 was reported as highly resistant and GL 13001 was reported as a highly susceptible genotype against Fusarium wilt (Kumar et al., 2019). The seeds of KWR 108 and GL 13001 genotypes were procured from the ICAR-Indian Institute of Pulse Research, Kanpur, Uttar Pradesh, India. Seeds of both genotypes were sown in a grow bag filled with a mixture of autoclaved soil and compost at a ratio of 6:4. For each genotype, two seeds were sown in each grow bag. After germination, only one plant bag−1 was maintained for 25 days after sowing. Then plants of both genotypes were divided into two groups, namely control and treatment, which had three biological replicates. All the field experiments were carried out under the net house condition during rabi season 2021–2022, and the molecular works were carried out at the Functional Genomics Laboratory of the Department of Agricultural Biotechnology and Molecular Biology (25.98°N latitude and 85.67°E longitude and 52 m altitude from mean sea level), College of Basic Sciences and Humanities, Dr. Rajendra Prasad Central Agricultural University, Pusa 848,125, Bihar, India during the year 2021–2022.
2.2. Treatment of plants with Fusarium oxysporum f. sp. ciceris culture
For infecting the plants, broth culture was prepared from the pure culture plates of Foc4 (pure culture was derived from the wilt-infected chickpea plant grown at the wilt sick plot of TCA, Dholi). Small-size inoculums (0.5 cm2) from the pure culture plate were inoculated in broth culture media (Potato Dextrose Broth) with the help of a forcep. The broth culture was incubated at 26°C and 150 rpm in a rotary agitator for 7 days. For inducing wilt infection in the treatment group of plants, the broth culture was thoroughly mixed using a pipette, and 2 mL of the solution was added to each plant at the root zone with the help of a pipette. To further ensure the infection of plants in treatment groups, soil collected from the Fusarium wilt sick plot of TCA Dholi was filled on the top of the grow bags (treatment), followed by light irrigation. Symptoms appeared in the susceptible genotypes within 8–10 days of the treatment (Figures 1A,B). The plant leaf samples were collected after 10 days of treatment for transcriptome sequencing.
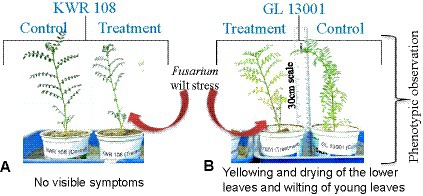
Figure 1. Phenotypic observations of (A) No visible symptoms in genotype KWR 108 and (B) Yellowing and drying of lower leaves and wilting of young leaves in genotype GL 13001 due to the infection of F. oxysporum as compared to control.
2.3. RNA isolation and preparation of library for transcriptome sequencing
The leaf samples were collected from control and treated plants of both the genotypes in three biological replicates and immediately dipped in the 2 mL micro-centrifuge tubes filled with RNA stabilizer solution (G-Biosciences) and outsourced for RNA isolation and transcriptome sequencing at Unigenome, Ahmedabad, Gujarat, India-380015. The leaf samples from three biological replicates were bulked together for RNA isolation. RNA was isolated by using the Alexgen Total RNA kit. RNA quality and quantity were analyzed using 1% agarose gel and Qubit® 4.0 fluorometer, respectively. The NEBNext® Ultra™ RNA Library Prep Kit for Illumina NEB #E 7770 was used for paired-end sequencing library preparation. The mRNA enrichment was performed as per the user manual and mRNA was first fragmented, followed by cDNA synthesis and repairing of ends and adenylation of 3′ end. Subsequently, the adapter was ligated at the fragmented ends and PCR amplification was performed for selective enrichment of adapter ligated DNA fragments. The adapter-based selectively enriched amplified fragments/libraries were analyzed on TapeStation 4,150 (Agilent using RNA ScreenTape® as per manufacturer’s instructions). The average size of libraries was observed to be 331 bp, 363 bp for GL 13001 (control and treatment) and 353 bp and 344 bp for KWR 108 (Control and Treatment), respectively. The final library was pooled with other samples, denatured, and loaded onto the flow cell. On the flow cell, cluster generation and sequencing were performed using the Illumina Novaseq 6,000 platform to generate 2 × 150 bp paired-end (PE) reads. Data were filtered to remove the adapter and low-quality reads. The raw transcriptome sequencing data obtained from both genotypes under control and F. oxysporum f. sp. ciceris-treated plants with two technical replicates were submitted to the SRA (Sequence Read Archive) database of NCBI (accessed on 20 October 2023).1
2.4. Reference genome-based transcript mapping, assembly, and further bioinformatics works
From the NCBI database (accessed on 20 October 2023),2 the reference genome of Cicer arietinum (GCA_000331145.1_ASM33114v1) and reference genome annotation GTF file were downloaded. Subsequently, by using STAR (v2.7.10a) aligner, reference genome-based assembly of transcripts was performed first by separately mapping high-quality (HQ) clean reads of control and treatment samples of GL 13001 and KWR 108 on the reference genome, respectively. The STAR Genome-Generate mode option was used to carry out indexing of the reference genome. Then the input reads, in FASTQ format, along with the indexed reference genome generated in the previous step, were given to the STAR aligner. To identify the positions of the origin of the reads, the STAR aligner created the alignment in BAM format for each sample through reference genome-guided mapping of the HQ reads followed by assembly of transcripts using StringTie (v2.2.1). After the assembly of transcripts by StringTie, the quantification of assembled transcripts was performed. The quantification of the assembled transcripts was done in order of most highly abundant transcripts to less abundant by using the network flow algorithm. The annotation of assembled transcripts was performed using the GFF annotation file of reference genome having “known” gene exon structures followed by quantification of the expression of known genes. Simultaneously, to merge the gene structure found in all four samples, the assembled transcripts of all four samples were passed to StringTie’s merge function to avoid assembly of only a partial version of any of the transcripts in the initial StringTie run.
2.5. Identification of novel transcripts and novel isoforms of transcripts
The reference GTF file and the string-tie merged GTF file were taken and the GFF compare utility was run to check or identify if there were any novel transcripts or novel isoforms of transcripts were present in the merge sample. Novel transcripts were then identified using a method described by Gleeson et al. (2022). The transcripts that did not have any available information in the annotation file of the reference genome were considered novel transcripts.
2.6. Differential gene expression analysis
Using the SringTie merge function, a consensus set of transcripts was derived by merging the structures of all the genes found in all four samples and simultaneously estimating the abundances of the merged transcripts in the four samples. The read count information that was obtained from the files generated by StringTie was extracted by using prepDE.py, a Python program (Gill and Dhillon, 2022). The edgeR package (Robinson et al., 2010) was then used to calculate differential gene expression (DEGs) by taking read count results of a Python program prepDE.py as input.
2.7. Gene ontology and KEGG enrichment pathway analysis
Gene ontology (GO) analysis was done to identify the distribution of significant differentially expressed genes (DEGs) into three major domains, i.e., cellular component, molecular function, and biological process. GO term was assigned to each significant differentially expressed transcript using Blas2GOcli 1.4. KAAS (KEGG automatic annotation server; Moriya et al., 2007) was used for ortholog identification and mapping of significant differentially expressed transcripts to the biological pathways. The criteria used to identify significantly down and up-regulated transcripts were log2FC > 0 and q value<0.05 (means significantly up-regulated), and log2FC < 0 and q value <0.05 (means significantly down-regulated). Using BLASTX with a threshold bit-score value of 60 (default), significant differentially expressed transcripts were compared against the KEGG database. A Heatmap of DEGs was generated for 25 highly significant down and up-regulated genes (a total top 50 transcripts) based on q value (i.e., least q value).
2.8. Identification of genes encoding differentially expressed transcripts
The reference of the genome-based transcript identification revealed that many of the transcripts showed differential expression under Fusarium wilt infections, which are the transcript fragments encoding the same proteins. Further analysis was conducted using the NCBI gene database. However, the protein IDs of the transcripts revealed that several proteins were the products/isoforms of the same genes. Using the sorting option available in MS Excel, the transcripts encoded by the same genes were sorted together to finally get the exact numbers of genes showing differential expression under Fusarium wilt stress in chickpea.
2.9. Primer design and real-time PCR analysis
From the RNA sequencing data, 15 genes that showed significant differential expression in the resistant genotype, i.e., KWR 108, were selected for real-time PCR verification of the RNA sequencing results. Primer Blast Program, available at NCBI database (accessed on 20 October 2023),3 was used for designing primers with CDS of 15 selected genes and CDS of Glucose-6-Phosphate Dehydrogenase (G6PD) gene as internal control (Reddy et al., 2016). RNA was extracted by using RNA isolation kit (Qiagen) from control and treatment plants of KWR 108 and cDNA was synthesized from RNA using the cDNA synthesis kit following the manufacturer’s protocol (Himedia, HiGenoMB). The gene expression analysis was done by using the cDNA template and 2× AB HS SYBR Green qPCR Mix (Thermo Scientific) in a qRT-PCR (Stratagene Mx3000P, Agilent). The details of real-time PCR primers specific to the ERF transcription factor family used in the experiment are listed in Table 1.
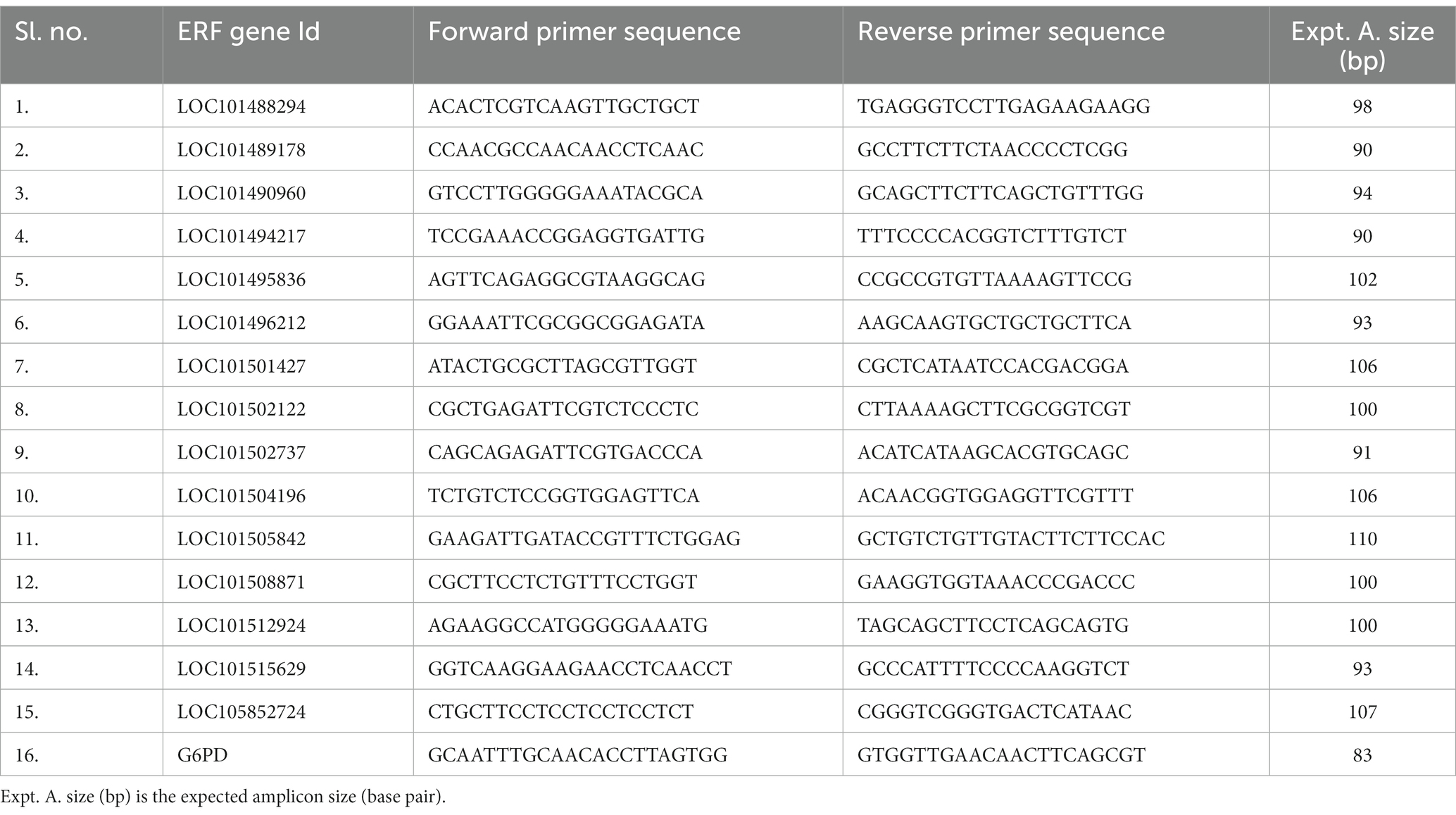
Table 1. List of qRT PCR primers specific to selected genes that showed significant up-regulation in KWR 108 in shoot transcriptome data for validation of RNA sequencing-based shoot transcriptome data.
2.10. Data analysis and graphing
Sample comparison was made for GL 13001 (control vs. treatment) and KWR 108 (control vs. treatment), respectively, for differential expression analysis. The analyses were performed in the edge R package to identify differentially expressed genes. Reads per million mapped reads or Counts per million mapped reads for both control and treatment samples, LogCPM (Log10 of CPM value), Log2Fold Change [the logarithm (base 2) of the fold change], pval (value of p for the statistical significance of this change), FDR [FDR adjusted value of p (q-value)], and the criteria used to identify significantly down and up-regulated transcripts were log2FC > 0 (means up-regulated), log2FC > 0 and q value<0.05 (means significantly up-regulated), log2FC < 0 (means down-regulated), log2FC < 0 and q value <0.05 (means significantly down-regulated). In real-time PCR analysis, the mean and standard deviation were calculated based on three pieces of biological replicated data and, using the Livak method (Schmittgen and Livak, 2008), the relative gene expression was calculated with G6PD as the internal reference gene. The rest of the data statistics and graphing were performed using MS Excel. The reliability of the RNAseq data was confirmed by comparing the expression data of selected genes using a linear regression model in Excel.
3. Results
3.1. Transcriptome data generation, transcript mapping, and assembly
In the present investigation, the total reads obtained from four samples, i.e., GL 13001 (control and treatment) and KWR 108 (control and treatment), ranged from 33.19 million reads in GL 13001-Control to 42.43 million reads in KWR 108-Treatment (Table 2).
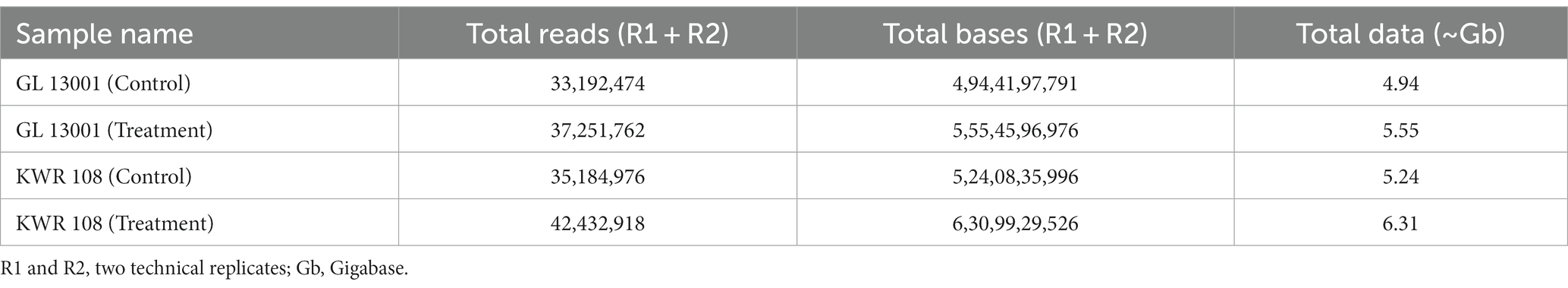
Table 2. The transcriptome data statistics (A) for clean data after removal of the adapter and low-quality bases.
The QC passed reads were then subjected to reference-based transcript assembly. The genome size for Cicer arietinum is 531 Mb. There are a total of 30,236 genes and 220,289 CDS as per the GFF file of the NCBI chickpea reference genome. The transcript assembly was performed by mapping clean reads of control and treatment samples of GL 13001 and KWR 108 on the reference genome using STAR (v 2.7.10a). The mapping statistics of HQ reads against the reference genome are presented in Table 3.
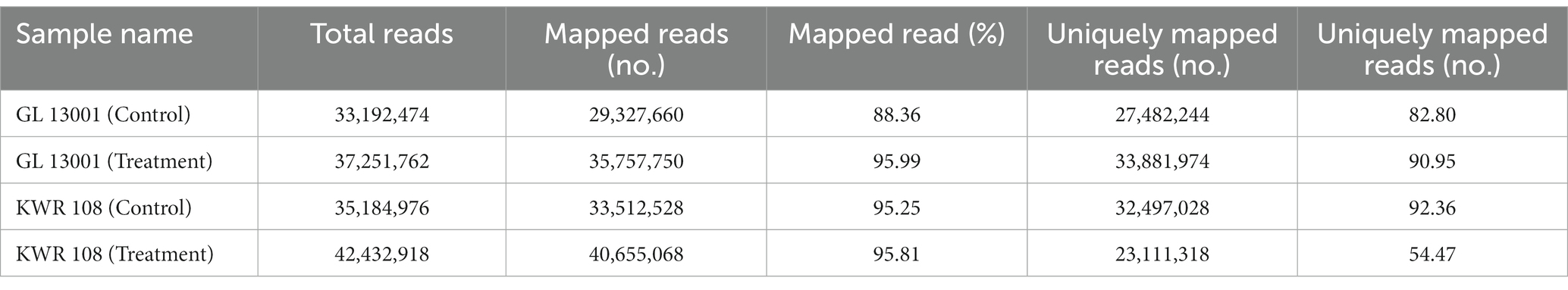
Table 3. Highlights of mapping statistics high-quality reads derived from shoot transcriptome mapped to chickpea reference genome using STAR aligner.
The number of assembled transcripts varied from 36,081 to 39,186 with an average transcript size of 2003.40 bp and 1992.60 bp in control and treated GL 1300, respectively. Similarly, the number of assembled transcripts ranged from 37,496 and 37,304 with an average transcript size of 2005.90 bp and 1984.60 bp in control and treated KWR 108 leaves samples, respectively. Simultaneously, StringTie’s merge resulted in several transcripts assembled as 51,353 with an average transcript size of 1898 bp in merge data (Table 4).
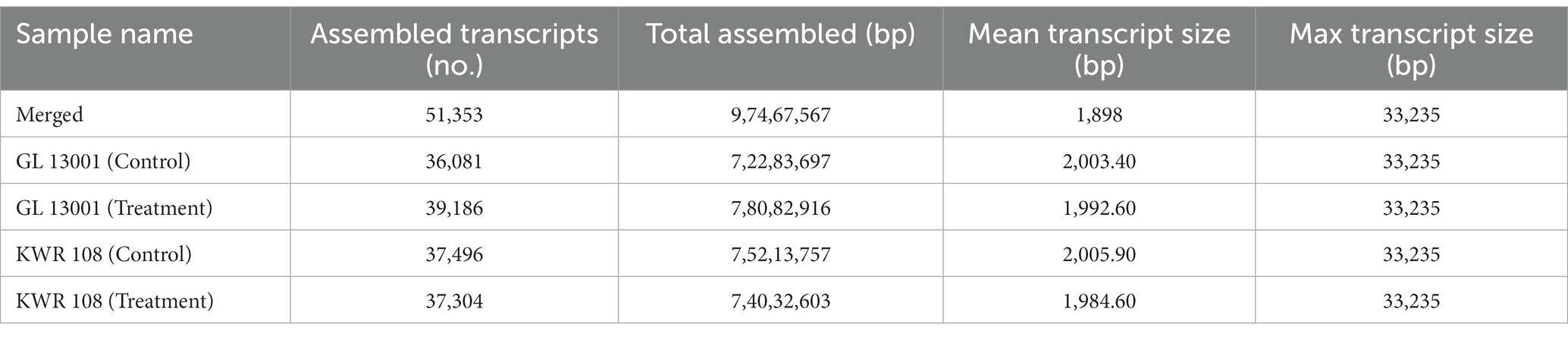
Table 4. The statistics of merged transcripts (generated by the combination of all four samples as well as reference genome) and individual transcript assembly using StringTie assembly and merge function.
StringTie’s merge function was used because individually in some of the samples, transcripts might only be partially covered by reads, leading to the assembly of only a partial version of those transcripts in the initial StringTie run. Thus, the set of transcripts that are consistent in all four samples was created to be compared in the subsequent steps using the merge step.
3.2. Novel transcripts and novel isoform transcripts identified
A total of 4,111 novel transcript isoforms corresponding to or encoded by 2,918 genes were identified as potential novel isoforms. Interestingly, 167 novel transcripts in KWR 108 and 170 novel transcripts in GL 13001, which were not available in the annotation file of the reference genome, were identified. The detailed list of novel transcripts and novel isoform transcripts identified are enlisted in Supplementary raw data file S1.
3.3. Identification of differentially expressed transcripts
Large numbers of transcripts were shown to be differentially expressed as revealed by differential expression analysis using edgeR software. Simultaneously, 7,504 and 6,698 transcripts were found to be significantly differentially expressed in GL 13001 and KWR 108, respectively (Table 5).
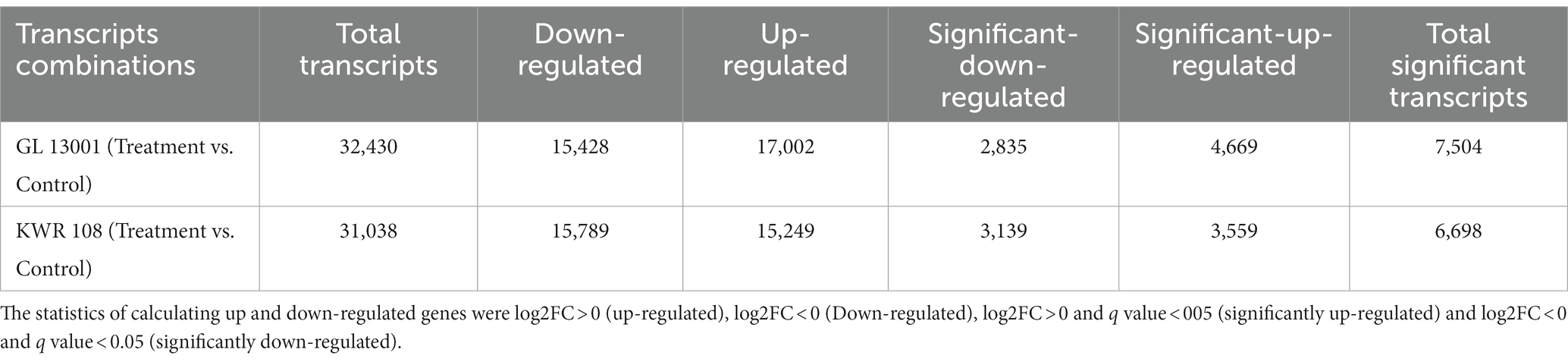
Table 5. The statistics of transcripts showing differential expression.
By transforming the data onto A (mean average) and M (log ratio) scales, the differences between measurements taken in control and treatment samples were visualized in MA plot. Simultaneously, a volcano plot that arranges differentially expressed transcripts along dimensions of statistical as well as biological significance was obtained from the edgeR software (Figures 2A–D). Similarly, the heatmap of 25 highly significant down and up-regulated genes (total 50 transcripts) based on q-value (i.e., least q value) and transcripts with proper annotations generated for both the genotypes are represented in Figures 2G,H.
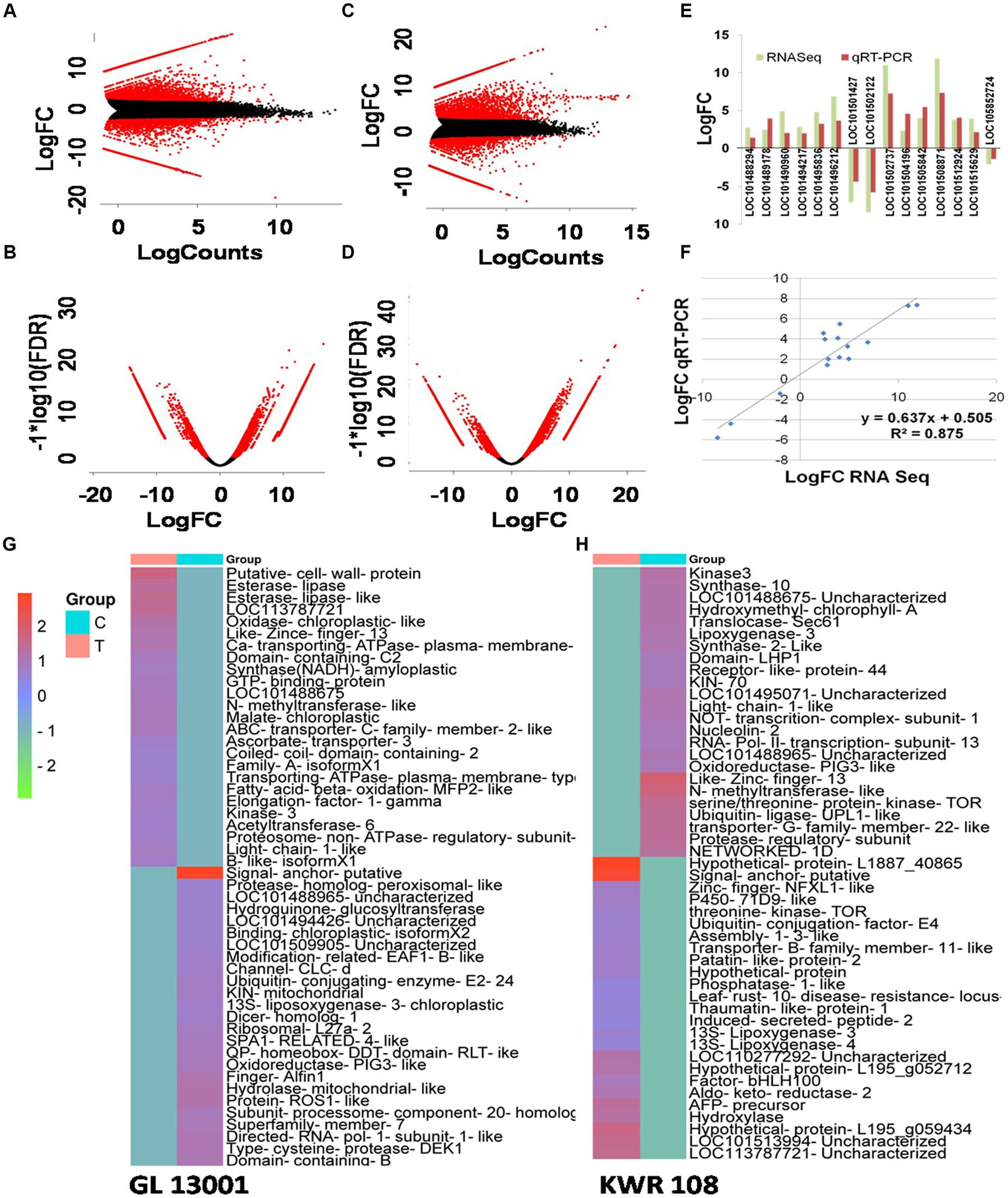
Figure 2. MA plot and volcano plot showing differentially expressed transcripts in treatment vs. control combination in GL 13001 (A,B) and KWR 108 (C,D). Red corresponds to transcripts with adjusted p value/q value <0.05. Validation of RNA-Seq data by qRT-PCR by using primers specific to 15 genes which in RNA-Seq result showed significant differential expression in KWR 108 (E,F). The heatmap of 25 highly significant down and up-regulated transcripts based on q value (i.e., least q value) in GL 13001 (G) and KWR 108 (H). The color coding ranges from red to green, where shades of red represent high transcript expression and shades of green represent low transcript expression. T means treatment sample and C means control sample.
3.4. Validation of RNAseq-based transcriptome data using a real-time PCR
The reliability of sequencing data was confirmed by real-time-based expression analysis of 15 selected genes that showed significant up-regulation in the resistant genotype under Fusarium wilt infection as revealed by RNAseq data. Similar to RNA sequencing results, the real-time PCR data of 15 selected genes showed significant differential expression of the genotype KWR 108 (Figure 2E). The linear regression analysis of RNAseq and real-time PCR-based expression data also showed significant similarity (R2 = 0.871), which confirmed that the RNAseq data were reliable for further analysis (Figure 2F).
3.5. Reference genome-based transcript identification
Identification of genes encoding differentially expressed transcripts revealed that, in GL 13001, 427 genes were significantly down-regulated and 794 were significantly up-regulated (Supplementary raw data file S2). Similarly, in KWR 108, 495 genes were significantly down-regulated and 608 genes were significantly up-regulated (Supplementary raw data file S2). When the genes that showed significant differential expression were compared between two genotypes, i.e., GL 13001 and KWR 108, it was revealed that 81 genes were down-regulated and 211 genes were up-regulated in both the genotypes. Ninety-one and 172 genes showed significant down-regulation and up-regulation only in GL 13001 and 93 and 97 genes showed significant down-regulation and up-regulation only in KWR 108, respectively. Similarly, 111 genes were significantly up-regulated in GL 13001 but significantly down-regulated in KWR 108 and 50 genes were significantly down-regulated in GL 13001 but significantly up-regulated in KWR 108.
3.6. Gene ontology enrichment analysis
The GO enrichment analysis of significant differentially expressed genes (DEGs) summarized the genes into three main GO categories, namely cellular component, molecular function, and biological process (Supplementary raw data file S2). The GO enrichment results of the top 50 significantly up-regulated and down-regulated genes in two genotypes are represented as pie charts (Figure 3).
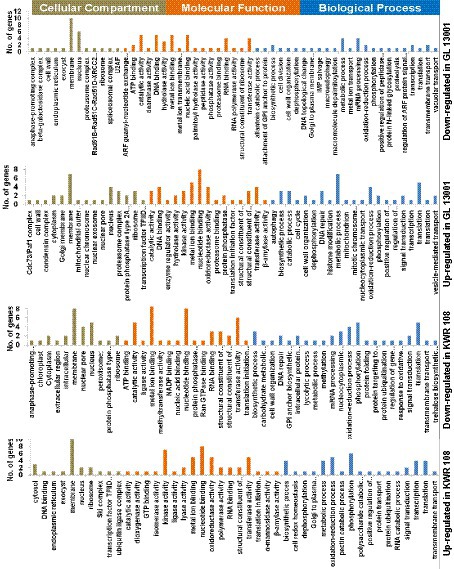
Figure 3. Gene ontology (GO) enrichment graphs of the top 50 up and 50 down-regulated genes in KWR 108 and GL 13001. Moss green color bar represents Cellular Compartments, Orange color bar represents Molecular functions, and Blue color bar represents biological processes in which the top 50 DEGs are involved.
The GO enrichment results of the top 50 significantly up and down-regulated genes revealed that the proteins encoded by most of these genes are the components of the membrane and nucleus. The products of most of the top 50 genes down-regulated in GL 13001 have nucleotide binding, catalytic, metal ion binding, transferase, and hydrolase activity involved in biological processes like metabolic processes, mRNA processing, DNA repair, oxidation–reduction process, and macroautophagy. Apart from membrane and nucleus, many of the proteins encoded by the top 50 genes up-regulated in GL 13001 are the components of proteasome and ribosome having nucleotide binding, catalytic, metal ion binding, oxidoreductase, and transferase activity involved in biological processes like metabolic processes, translation, DNA repair, and oxidation–reduction process.
Similar to the GO terms of the top 50 genes significantly up and down-regulated in GL 13001, the GO terms of the top 50 genes down-regulated in KWR 108 showed the highest cellular component as membrane and nucleus and molecular functions as nucleotide binding, metal ion binding, transferase, catalytic, and nuclear pore activities. However, the GO terms of the top 50 genes up-regulated in KWR 108 revealed that, apart from membrane and nucleus, the products of the genes are the components of ribosome and cytosol having molecular functions such as kinase activity, oxidoreductase activity, nucleotide binding, transferase, and metal ion binding activities involved in biological processes such as phosphorylation, oxidation–reduction process, metabolic and biosynthetic process, and cell redox homeostasis (Figure 3).
3.7. Pathways involving differentially expressed genes
KEGG enrichment analysis of the significant DEGs identified in both the genotypes of chickpea was performed to estimate the numbers of significant DEGs involved in different KEGG pathway levels. It revealed that, in the susceptible genotype, i.e., GL 13001, the number of significantly up-regulated genes involved in different pathways such as DNA replication and DNA repair, cell motility, lipid metabolism, carbohydrate metabolism, and DNA degradation is significantly higher than in KWR 108. Whereas the number of DEGs specifically involved in energy metabolism and environmental adaptation pathways are significantly higher in KWR 108 than in GL 13001 (Figure 4A). The same genes that were up-regulated in GL 13001 but down-regulated in KWR 108 were observed to be higher in most of the pathways; more specifically, the genes involved in cell motility and membrane transport were significantly up-regulated only in GL 13001 (Figure 4B). However, the same genes involved in carbohydrate metabolism were up-regulated in resistant genotype KWR 108 but down-regulated in susceptible genotype GL 13001 (Figure 4B).
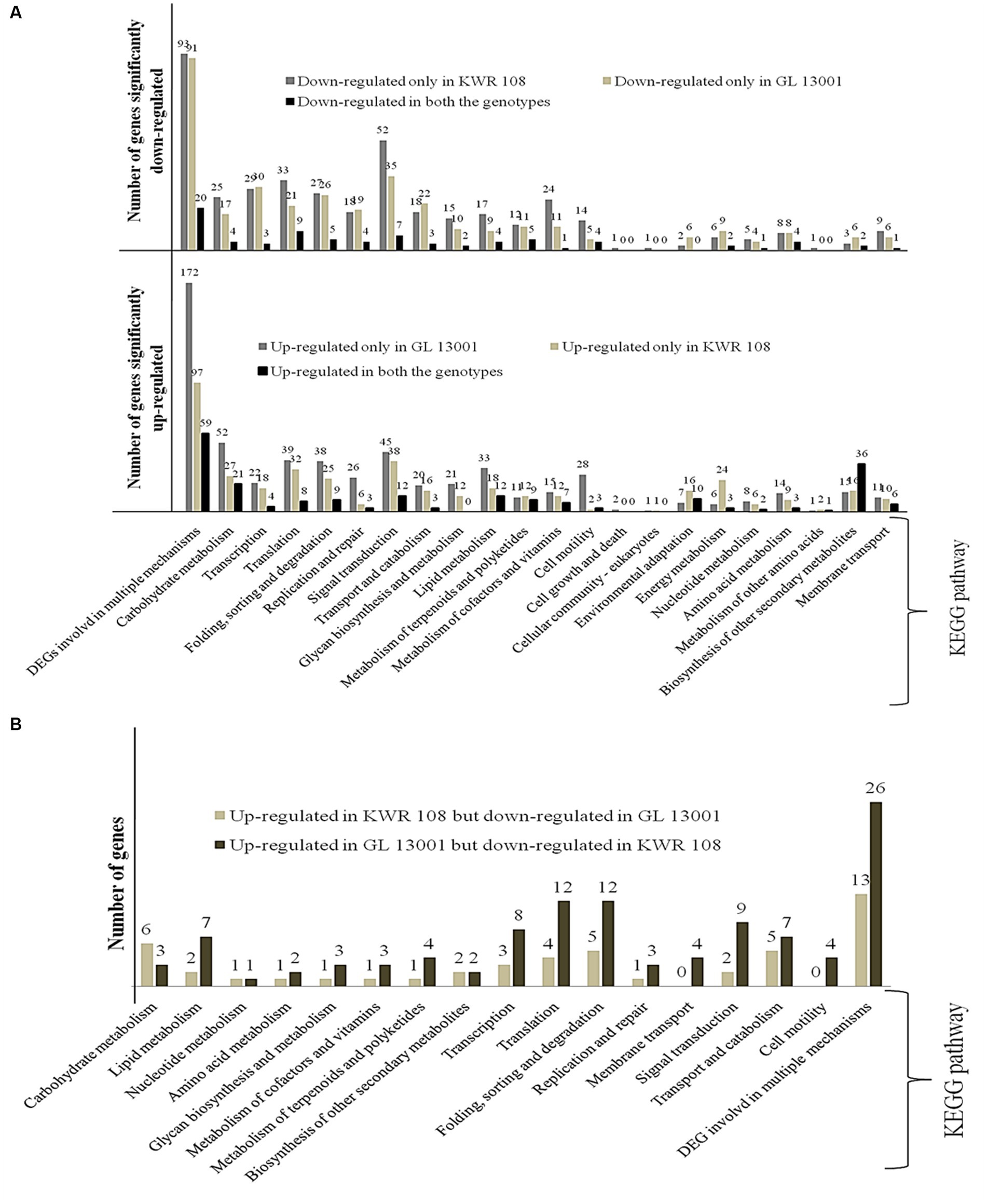
Figure 4. KEGG enrichment/Pathway analysis results (A) genes significantly up-regulated and down-regulated in two genotypes of chickpea, i.e., KWR 108 and GL 13001 and (B) genes significantly up-regulated in KWR 108 but down-regulated in GL 13001 and vice versa. ‘Y’ axis represents the number of significant DEGs.
4. Discussion
In the present study, transcriptome dissection was done to understand the probable molecular mechanisms imparting the ability to tolerate Fusarium wilt in the chickpea genotype, particularly in the genome background of KWR 108, a highly resistant variety screened at TCA-Dholi, Bihar, India (Kumar et al., 2019). The comparative analysis of the transcriptome data generated from susceptible genotypes GL 13001 and resistant variety KWR 108 was performed to identify the pathways and genes involved that are showing differential expression under Fusarium wilt stress. Compared to the susceptible genotypes, higher numbers of genes belonging to the pathways involved in environmental adaptation and energy metabolism showed highly significant up-regulation in resistant genotype KWR 108 and, along with genes involved in several other processes, more genes belonging to DNA repair mechanisms showed significant up-regulation in susceptible genotype GL13001 (Figure 4A).
4.1. Significant up-regulation of genes involved in DNA repair mechanism in susceptible genotype
In our present study, a comparison between the shoot transcriptome of KWR 108 and GL 13001 showed a significantly higher number of genes involved in DNA repair mechanisms were up-regulated in GL 13001, a susceptible genotype (Figure 4A). Although, under normal circumstances, DNA damage can be thought of as a frequent occurrence, it is likely to occur even more in response to several stress conditions (Nisa et al., 2019) either directly or indirectly due to increased level of reactive oxygen species (ROS; Szurman-Zubrzycka et al., 2023). The resistant or tolerant plants protect the DNA from damage by activating enzymatic and non-enzymatic antioxidant machinery (Moradbeygi et al., 2020). In the present study, the significantly up-regulated genes related to DNA repair mechanism in susceptible genotypes are crossover junction endonuclease EME1, DNA excision repair protein ERCC-1 & 4, DNA mismatch repair protein MLH3, DNA repair and recombination protein RAD54 and RAD54-like protein, DNA repair protein REV1, DNA-3-methyladenine glycosylase I, flap endonuclease-1, telomere length regulation protein, UV excision repair protein RAD23, and WD repeat-containing protein 48. This suggests that DNA replication and other similar processes of DNA synthesis are more stable in resistant genotypes as compared to susceptible genotypes under Fusarium wilt stress condition.
4.2. Significant up-regulation of environmental adaptations genes in the resistant genotype
The genes specifically belonging to environmental adaptation pathways are significantly up-regulated in the resistant genotype. The environmental adaptation genes basically encode for the proteins or enzymes involved in plant-pathogen interaction upon infection. In the present study, the significantly up-regulated environmental adaptation genes in KWR 108 were calcium-dependent protein kinase (CDPK), pathogenesis-related genes transcriptional activator (Pti5 & Pti6), calmodulin (CaM), calmodulin-like proteins (CML), cyclic nucleotide-gated channel (CNGC), mitochondrial translation elongation factor Tu (TUFM), chitin elicitor receptor kinase 1 (CERK1), enhanced disease susceptibility 1 protein (EDS1), disease resistance protein (RPM1/ RPS3), mitogen-activated protein kinase 3 (MPK3), mitogen-activated protein kinase 4 (MPK4), mitogen-activated protein kinase kinase 4/5 (MKK4_5), pathogenesis-related protein 1 (PR1), and WRKY transcription factor 33 (WRKY33). The important roles and involvement of environmental adaptation genes like CDPK (Asano et al., 2012; Baba et al., 2019; Wu et al., 2021), Pti5 & Pti6 (Yang et al., 2020; Wang Y. et al., 2021; Sun et al., 2022), CaM, and CML (Zeng et al., 2015; Ranty et al., 2016) in several biotic and abiotic stress responses have been extensively studied and reported. The plant accumulates Ca+ upon experiencing stress as a secondary messenger to modulate the cellular processes (Luan, 2009). CNGC, which showed significant up-regulation under Fusarium wilt in the present study, is mostly localized to the plasma membrane and acts as ligand-gated cation channels (Chin et al., 2009) for the accumulation of Ca+ in the cell during stress. Several reports suggested the potential role of CNGC in plant disease resistance through hypersensitive response (Clough et al., 2000; Ahn, 2007; Keisa et al., 2011; Ma and Berkowitz, 2011; Guo et al., 2018; Peng et al., 2019; Dietrich et al., 2020; Chakraborty et al., 2021; Zhao et al., 2021). Mitochondrial translation elongation factor Tu (TUFM) is a mitochondrial protein encoded by the nuclear genome that potently inhibits RLR (RIG-I-like receptors) signaling and promotes autophagy during a pathogen infection, particularly viruses (Lei, 2011; Wu et al., 2022). In the study, TUFM showed significant up-regulation in Fusarium wilt-resistant genotype KWR 108, suggesting its potential role in imparting resistance against fungal pathogens and other stresses as reported in crops like maize (Rao et al., 2004) and soybean (Yin et al., 2009). Chitin, which is an essential unit of fungal cell walls including chickpea wilt, causes F. oxysporum to act as an elicitor of plant immunity (Gong et al., 2020). Chitin elicitor receptor kinase 1 (CERK1), which showed significant up-regulation in resistant genotypes in the present study, is a heterotrimeric complex (Gigli-Bisceglia and Testerink, 2021) required for the perception of chitin and subsequent activation of the chitin-responsive cellular processes as well as for triggering the innate immunity of the plants against abiotic stress (Espinoza et al., 2017). CERK1 mutant (cerk1) plants showed increased susceptibility to fungal pathogens like Erisiphe cichoracearum, Alternaria brassicicola, and Blumeria graminis (Gigli-Bisceglia and Testerink, 2021), indicating its vital role in perceiving pathogen signals and subsequent induction of defense mechanism in resistant plants. The enhanced disease susceptibility 1 (EDS1) gene, which showed significant upregulation in KWR 108 in the present study, encodes a lipase-like protein that has been reported to be essential for resistance against biotrophic pathogens (Ochsenbein et al., 2006). In plants, it is basically required by Toll/Interleukin-1 receptor (TIR) domain of NLR (nucleotide-binding leucine-rich repeat) immune receptors (Johanndrees et al., 2023) and for inducing expression of PR (pathogen-related) proteins through the salicylic acid (SA) pathway against several pathogens (Yan et al., 2016; Bhandari et al., 2019; Neubauer et al., 2020). Likewise, RPM1/RPS3 (disease resistance protein), which in the present study showed significant up-regulation in the resistant genotype, is basically a CC-NBS-LRR protein that was first identified as a protein in Arabidopsis essential for resistance mechanism against P syringae pv. Maculicola. It was later reported to have roles in resistance against diseases caused by other pathogens like Puccinia striiformis f. sp. tritici through hypersensitive response and Ca+ concentration dynamics (Grant et al., 2000; Wang et al., 2020). In the present study, mitogen-activated protein kinases (MAPK) such as MPK3, MPK4, and MKK4_5 showed significant up-regulation in resistant genotypes that are basically involved in intracellular responses (Sinha et al., 2011). In plants, MAPKs are reported to induce stress tolerance (Lin et al., 2021) including biotic stresses (Zhang and Zhang, 2022) through JA (Jasmonic acid) and SA signaling pathways (Jagodzik et al., 2018). The SA signaling in plants under stress including fungal infection induces the expression of defense proteins like PR1 (pathogenesis-related protein 1; Almeida-Silva and Venancio, 2022; Anuradha et al., 2022). Antifungal and several other antimicrobial activities of PR1 have been well-reviewed and reported (Kattupalli et al., 2021; Almeida-Silva and Venancio, 2022; Anuradha et al., 2022; Pečenková et al., 2022). Similar to all these reports, in the present study also, PR1 genes showed significant up-regulation in chickpea under Fusarium wilt stress, confirming its role as a defense mechanism against Fusarium wilt disease. Likewise, in plant defense responses, an important oxidoreductase enzyme is peroxidase, which is involved in cell wall modifications and the generation of reactive oxygen species (ROS; Chang et al., 2021). Xue et al. (2017) reported that, in common bean, the peroxidase gene PvPOX1 enhanced resistance against Fusarium wilt. In the present study, a large number of peroxidase genes up-regulated in KWR 108, but none in GL 13001, suggesting its role in resistance against Fusarium wilt in chickpea also. Similarly, WRKY33, an important member of WRKY transcription factor family in plants, showed significant up-regulation in the resistant genotype. The role of WRKY transcription factor has been studied extensively and it plays a role in responses to several stimuli irrespective of physiological, biotic, or abiotic stress (Phukan et al., 2016; Cheng et al., 2021).
4.3. Significant up-regulation of energy metabolism genes in the resistant genotype
The important metabolites involved in redox homeostasis of plant cells are nicotinamide adenine dinucleotide (NAD) and its other derivatives, which are mostly nucleotide coenzymes (Kapoor et al., 2015). The redox state of NAD, i.e., NAD (P)+/NAD(P)H (ratio of oxidized to reduced form) in the cell plays an important role as a signal to ameliorate the bridge between cellular metabolic state and gene expression under various conditions, including biotic and abiotic stress condition (Kapoor et al., 2015), and are vital for homeostasis of cellular energy in plant cells under stress condition for optimum growth and development. In the present study, genes related to NAD (P)H, such as NADH-dependent glutamate synthase, NAD(P)H-quinone oxidoreductase subunit 4, NADH dehydrogenase (ubiquinone) Fe-S protein, NADH:quinone reductase, and NADH–ubiquinone oxidoreductase, showed significant up-regulation in resistant genotypes, confirming its role in energy homeostasis in plants under stress conditions. Other than NADH-related genes, shoot transcriptome data of resistant genotypes in the present study also showed significant up-regulation of other genes involved in energy metabolism, such as phosphoenolpyruvate carboxylase (Wang et al., 2016; Waseem and Ahmad, 2019), alcohol dehydrogenase (Su et al., 2020), alanine transaminase (Bashar et al., 2020), ATP citrate (pro-S)-lyase (Liu F. et al., 2022), ATPase (Li J. et al., 2022; Li Y. et al., 2022), carbonic anhydrase (Rudenko et al., 2021), cytochrome c, cytochrome c oxidase (Guerra-Castellano et al., 2018; Analin et al., 2020), fructose-bisphosphate aldolase (Lv et al., 2017; Cai et al., 2022), malate dehydrogenase (Song et al., 2022), 6-phosphofructokinase 1 (Wang H. et al., 2021), photosystem II P680 reaction center D1 protein (Landi and Guidi, 2022), phosphoserine aminotransferase (Wang et al., 2022), pyruvate-orthophosphate dikinase (Yadav et al., 2020), and serine O-acetyltransferase (Mulet et al., 2004; Liu D. et al., 2022), and their molecular mechanism and role in energy homeostasis in different crops is well reviewed. It indicates that the significant up-regulation of the genes involved in energy metabolism in KWR 108 might have helped it to cope with Fusarium wilt stress by maintaining energy homeostasis in cells under stress conditions.
The genes particularly involved in environmental adaptation, energy metabolism, and others which showed significant up-regulation in wilt-resistant genotypes will be a target for designing gene-based markers for marker-trait association studies. The marker-trait association study, especially for the traits attributing to wilt resistance and yield under wilt infection, will be our next target to check whether these up-regulated genes have any roles in imparting resistance against Fusarium wilt through some desirable phenotypic traits or whether there are no roles in phenotypic traits of chickpea under wilt. If these gene-based marker/s are found to be robustly associated with Fusarium wilt-resistant traits, then they could be utilized in marker-assisted breeding to develop a wilt-resistant high-yielding variety of chickpea.
5. Conclusion
The present study was conducted to gain insight into the molecular mechanism imparting resistance against Fusarium wilt infection in chickpea using an RNA sequencing-based shoot transcriptome approach. The shoot transcriptome data derived from two contrasting genotypes of chickpea, namely KWR 108 (wilt resistant variety) and GL 13001 (wilt susceptible variety), revealed that the resistant genotype efficiently maintains the energy homeostasis in the cell under stress conditions by up-regulating the large sets of genes responsible for maintaining the cellular energy homeostasis. The up-regulation of significant numbers of genes involved in environmental adaptation, particularly host-pathogen interaction in resistant genotypes, suggests that the resistant genotypes efficiently deploy the defense mechanism starting from perceiving and transduction of pathogen signal to activation and implementation of the defense-related mechanism. Similarly, the susceptible genotype showed significant up-regulation of genes involved in several mechanisms, including the DNA repair mechanism, thus it can be perceived that the DNA molecules of the susceptible genotype were more unstable and damaged than the DNA molecules of the resistant genotype, which showed optimum growth under Fusarium wilt stress. Therefore, it can be concluded that efficient energy metabolism, activation of environmental adaptation mechanisms, and DNA stability were key to resistance against Fusarium wilt infection in chickpea genotypes.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
KB: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft. MA: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – original draft. AK: Conceptualization, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – original draft. RS: Investigation, Methodology, Validation, Visualization, Writing – original draft. NB: Methodology, Software, Validation, Visualization, Writing – original draft. VS: Investigation, Methodology, Validation, Visualization, Writing – original draft. BP: Methodology, Software, Validation, Visualization, Writing – original draft. MT: Investigation, Methodology, Validation, Visualization, Writing – original draft. OO: Data curation, Formal analysis, Funding acquisition, Software, Writing - review & editing. VB: Data curation, Formal analysis, Funding acquisition, Software, Writing – review & editing. MB: Data curation, Formal analysis, Funding acquisition, Software, Writing – review & editing. MS: Data curation, Formal analysis, Funding acquisition, Software, Writing – review & editing. AG: Data curation, Formal analysis, Funding acquisition, Software, Writing – review & editing. AH: Data curation, Formal analysis, Funding acquisition, Software, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This experiment was funded by the Science and Engineering Research Board (SERB), DST-Government of India (Project Id No. EEQ/2020/000535). The study was also financially supported by the Slovak Research and Development Agency under contract no. APVV-15-0562, APVV-20-0071 and Science grant agency under contract no. VEGA-1/0300/22. The study received funding from the Deanship of Scientific Research, Taif University.
Acknowledgments
The authors are thankful to the Slovak Research and Development Agency for supporting the study. The researchers would also like to acknowledge the Deanship of Scientific Research, Taif University, Taif, Saudi Arabia for funding this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1265265/full#supplementary-material
Footnotes
1. ^https://submit.ncbi.nlm.nih.gov/subs/sra/SUB12177877/
References
Adugna, A. (2004). Alternate approaches in deploying genes for disease resistance in crop plants. Asian J. Plant Sci. 3, 618–623. doi: 10.3923/ajps.2004.618.623
Ahn, I. P. (2007). Disturbance of the Ca2+/calmodulin-dependent signalling pathway is responsible for the resistance of Arabidopsis dnd1 against Pectobacterium carotovorum infection. Mol. Plant Pathol. 8, 747–759. doi: 10.1111/j.1364-3703.2007.00428.x
Almeida-Silva, F., and Venancio, T. M. (2022). Pathogenesis-related protein 1 (PR-1) genes in soybean: genome-wide identification, structural analysis and expression profiling under multiple biotic and abiotic stresses. Gene 809:146013. doi: 10.1016/j.gene.2021.146013
Analin, B., Mohanan, A., Bakka, K., and Challabathula, D. (2020). Cytochrome oxidase and alternative oxidase pathways of mitochondrial electron transport chain are important for the photosynthetic performance of pea plants under salinity stress conditions. Plant Physiol. Biochem. 154, 248–259. doi: 10.1016/j.plaphy.2020.05.022
Anuradha, C., Chandrasekar, A., Backiyarani, S., Thangavelu, R., Giribabu, P., and Uma, S. (2022). Genome-wide analysis of pathogenesis-related protein 1 (PR-1) gene family from Musa spp. and its role in defense response during stresses. Gene 821:146334. doi: 10.1016/j.gene.2022.146334
Asano, T., Hayashi, N., Kikuchi, S., and Ohsugi, R. (2012). CDPK-mediated abiotic stress signaling. Plant Signal. Behav. 7, 817–821. doi: 10.4161/psb.20351
Baba, A. I., Rigó, G., Andrási, N., Tietz, O., Palme, K., Szabados, L., et al. (2019). “Striving towards abiotic stresses: role of the plant CDPK superfamily members” in International climate protection. eds. M. Palocz-Andresen, D. Szalay, A. Gosztom, L. Sípos, and T. Taligás (Cham: Springer), 99–105.
Bashar, K. K., Tareq, M. Z., and Islam, M. S. (2020). Unlocking the mystery of plants’ survival capability under waterlogging stress. Plant Sci. Today 7, 142–153. doi: 10.14719/pst.2020.7.2.663
Bhandari, D. D., Lapin, D., Kracher, B., von Born, P., Bautor, J., Niefind, K., et al. (2019). An EDS1 heterodimer signalling surface enforces timely reprogramming of immunity genes in Arabidopsis. Nat. Commun. 10:772. doi: 10.1038/s41467-019-08783-0
Cai, B., Ning, Y., Li, Q., Li, Q., and Ai, X. (2022). Effects of the chloroplast fructose-1, 6-bisphosphate aldolase gene on growth and low-temperature tolerance of tomato. Int. J. Mol. Sci. 23:728. doi: 10.3390/ijms23020728
Chakraborty, S., Toyota, M., Moeder, W., Chin, K., Fortuna, A., Champigny, M., et al. (2021). CYCLIC NUCLEOTIDE-GATED ION CHANNEL 2 modulates auxin homeostasis and signaling. Plant Physiol. 187, 1690–1703. doi: 10.1093/plphys/kiab332
Chang, Y., Sun, F., Sun, S., Wang, L., Wu, J., and Zhu, Z. (2021). Transcriptome analysis of resistance to Fusarium wilt in mung bean (Vigna radiata L.). Front. Plant Sci. 12:1213. doi: 10.3389/fpls.2021.679629
Cheng, Z., Luan, Y., Meng, J., Sun, J., Tao, J., and Zhao, D. (2021). WRKY transcription factor response to high-temperature stress. Plan. Theory 10:2211. doi: 10.3390/plants10102211
Chin, K., Moeder, W., and Yoshioka, K. (2009). Biological roles of cyclic-nucleotide-gated ion channels in plants: what we know and don’t know about this 20-member ion channel family. Botany 87, 668–677. doi: 10.1139/B08-147
Clough, S. J., Fengler, K. A., Yu, I. C., Lippok, B., Smith, R. K., and Bent, A. F. Jr. (2000). The Arabidopsis dnd1 “defense, no death” gene encodes a mutated cyclic nucleotide-gated ion channel. Proc. Natl. Acad. Sci. 97, 9323–9328. doi: 10.1073/pnas.15000569
Dietrich, P., Moeder, W., and Yoshioka, K. (2020). Plant cyclic nucleotide-gated channels: new insights on their functions and regulation. Plant Physiol. 184, 27–38. doi: 10.1104/pp.20.00425
Espinoza, C., Liang, Y., and Stacey, G. (2017). Chitin receptor CERK 1 links salt stress and chitin-triggered innate immunity in Arabidopsis. Plant J. 89, 984–995. doi: 10.1111/tpj.13437
FAOSTAT. (2023). World Chickpea production in 2020–21, UN FAO, Corporate Statistical Database. Available at: https://www.fao.org/faostat/en/#data/QCL/visualize. Retrieved on 5 January 2023.
Gigli-Bisceglia, N., and Testerink, C. (2021). Fighting salt or enemies: shared perception and signaling strategies. Curr. Opin. Plant Biol. 64:102120. doi: 10.1016/j.pbi.2021.102120
Gill, N., and Dhillon, B. (2022). “RNA-seq data analysis for differential expression” in Fusarium wilt. Methods in molecular biology. ed. J. Coleman, vol. 2391 (New York, NY: Humana)
Gleeson, J., Leger, A., Prawer, Y. D., Lane, T. A., Harrison, P. J., Haerty, W., et al. (2022). Accurate expression quantification from nanopore direct RNA sequencing with NanoCount. Nucleic Acids Res. 50:e19. doi: 10.1093/nar/gkab1129
Gong, B. Q., Wang, F. Z., and Li, J. F. (2020). Hide-and-seek: chitin-triggered plant immunity and fungal counterstrategies. Trends Plant Sci. 25, 805–816. doi: 10.1016/j.tplants.2020.03.006
Grant, M., Brown, I., Adams, S., Knight, M., Ainslie, A., and Mansfield, J. (2000). The RPM1 plant disease resistance gene facilitates a rapid and sustained increase in cytosolic calcium that is necessary for the oxidative burst and hypersensitive cell death. Plant J. 23, 441–450. doi: 10.1046/j.1365-313x.2000.00804.x
Guerra-Castellano, A., Díaz-Quintana, A., Pérez-Mejías, G., Elena-Real, C. A., González-Arzola, K., García-Mauriño, S. M., et al. (2018). Oxidative stress is tightly regulated by cytochrome c phosphorylation and respirasome factors in mitochondria. Proc. Natl. Acad. Sci. 115, 7955–7960. doi: 10.1073/pnas.1806833115
Guo, J., Islam, M. A., Lin, H., Ji, C., Duan, Y., Liu, P., et al. (2018). Genome-wide identification of cyclic nucleotide-gated ion channel gene family in wheat and functional analyses of TaCNGC14 and TaCNGC16. Front. Plant Sci. 9:18. doi: 10.3389/fpls.2018.00018
Gupta, S., Chakraborti, D., Rangi, R. K., Basu, D., and Das, S. (2009). A molecular insight into the early events of chickpea (Cicer arietinum) and Fusarium oxysporum f. sp. ciceri (race 1) interaction through cDNA-AFLP analysis. Phytopathology 99, 1245–1257. doi: 10.1094/PHYTO-99-11-1245
Haware, M. P., Nene, Y. L., and Natarajan, M. (1996). Survival of Fusarium oxysporum f. sp. ciceri in the soil in the absence of chickpea. Phytopathol. Mediterr. 35, 9–12.
Indiastat. (2023). India Chickpea production in 2020–21. Available at: https://www.indiastat.com/data/agriculture. Retrieved on 5 January 2023.
Jagodzik, P., Tajdel-Zielinska, M., Ciesla, A., Marczak, M., and Ludwikow, A. (2018). Mitogen-activated protein kinase cascades in plant hormone signaling. Front. Plant Sci. 9:1387. doi: 10.3389/fpls.2018.01387
Jendoubi, W., Bouhadida, M., Boukteb, A., Béji, M., and Kharrat, M. (2017). Fusarium wilt affecting chickpea crop. Agriculture 7:23. doi: 10.3390/agriculture7030023
Johanndrees, O., Baggs, E. L., Uhlmann, C., Locci, F., Läßle, H. L., Melkonian, K., et al. (2023). Variation in plant toll/Interleukin-1 receptor domain protein dependence on ENHANCED DISEASE SUSCEPTIBILITY 1. Plant Physiol. 191, 626–642. doi: 10.1093/plphys/kiac480
Kapoor, D., Sharma, R., Handa, N., Kaur, H., Rattan, A., Yadav, P., et al. (2015). Redox homeostasis in plants under abiotic stress: role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci. 3:13. doi: 10.3389/fenvs.2015.00013
Kattupalli, D., Srinivasan, A., and Soniya, E. V. (2021). A genome-wide analysis of pathogenesis-related Protein-1 (PR-1) genes from Piper nigrum reveals its critical role during Phytophthora capsici infection. Gene 12:1007. doi: 10.3390/genes12071007
Keisa, A., Kanberga-Silina, K., Nakurte, I., Kunga, L., and Rostoks, N. (2011). Differential disease resistance response in the barley necrotic mutant nec1. BMC Plant Biol. 11:66. doi: 10.1186/1471-2229-11-66
Kohli, D., Joshi, G., Deokar, A. A., Bhardwaj, A. R., Agarwal, M., Katiyar-Agarwal, S., et al. (2014). Identification and characterization of wilt and salt stress-responsive microRNAs in chickpea through high-throughput sequencing. PLoS One 9:e108851. doi: 10.1371/journal.pone.0108851
Kumar, S., Sahni, S., and Kumar, B. (2019). Screening of chickpea genotypes for resistance against Fusarium wilt. Current J. App. Sci. Technol. 38, 1–6. doi: 10.9734/CJAST/2019/v38i630409
Landi, M., and Guidi, L. (2022). Effects of abiotic stress on photosystem II proteins. Photosynthetica 61, 148–156. doi: 10.32615/ps.2022.043
Lei, Y. (2011). Biochemical and functional characterization of the mitochondrial immune Signaling protein complex. A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Curriculum of Oral Biology, School of Dentistry. doi: 10.17615/sr3j-8s02
Li, J., Guo, Y., and Yang, Y. (2022). The molecular mechanism of plasma membrane H+-ATPases in plant responses to abiotic stress. J. Genetics Genomics. 49, 715–725. doi: 10.1016/j.jgg.2022.05.007
Li, Y., Zeng, H., Xu, F., Yan, F., and Xu, W. (2022). H+-ATPases in plant growth and stress responses. Annu. Rev. Plant Biol. 73, 495–521. doi: 10.1146/annurev-arplant-102820-114551
Lin, L., Wu, J., Jiang, M., and Wang, Y. (2021). Plant mitogen-activated protein kinase cascades in environmental stresses. Int. J. Mol. Sci. 22:1543. doi: 10.3390/ijms22041543
Liu, D., Li, M., Guo, T., Lu, J., Xie, Y., Hao, Y., et al. (2022). Functional characterization of the serine acetyltransferase family genes uncovers the diversification and conservation of cysteine biosynthesis in tomato. Front. Plant Sci. 13:913856. doi: 10.3389/fpls.2022.913856
Liu, F., Ma, Z., Cai, S., Dai, L., Gao, J., and Zhou, B. (2022). ATP-citrate lyase B (ACLB) negatively affects cell death and resistance to verticillium wilt. BMC Plant Biol. 22:443. doi: 10.1186/s12870-022-03834-z
Luan, S. (2009). The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 14, 37–42. doi: 10.1016/j.tplants.2008.10.005
Lv, G. Y., Guo, X. G., Xie, L. P., Xie, C. G., Zhang, X. H., Yang, Y., et al. (2017). Molecular characterization, gene evolution, and expression analysis of the fructose-1, 6-bisphosphate aldolase (FBA) gene family in wheat (Triticum aestivum L.). Front. Plant Sci. 8:1030. doi: 10.3389/fpls.2017.01030
Ma, W., and Berkowitz, G. A. (2011). Ca2+ conduction by plant cyclic nucleotide gated channels and associated signaling components in pathogen defense signal transduction cascades. New Phytol. 190, 566–572. doi: 10.1111/j.1469-8137.2010.03577.x
Moradbeygi, H., Jamei, R., Heidari, R., and Darvishzadeh, R. (2020). Investigating the enzymatic and non-enzymatic antioxidant defense by applying iron oxide nanoparticles in Dracocephalum moldavica L. plant under salinity stress. Sci. Hortic. 272:109537. doi: 10.1016/j.scienta.2020.109537
Moriya, Y., Itoh, M., Okuda, S., Yoshizawa, A. C., and Kanehisa, M. (2007). KAAS: an automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 35, W182–W185. doi: 10.1093/nar/gkm321
Mulet, J. M., Alemany, B., Ros, R., Calvete, J. J., and Serrano, R. (2004). Expression of a plant serine O-acetyltransferase in Saccharomyces cerevisiae confers osmotic tolerance and creates an alternative pathway for cysteine biosynthesis. Yeast 21, 303–312. doi: 10.1002/yea.1076
Neubauer, M., Serrano, I., Rodibaugh, N., Bhandari, D. D., Bautor, J., Parker, J. E., et al. (2020). Arabidopsis EDR1 protein kinase regulates the association of EDS1 and PAD4 to inhibit cell death. Mol. Plant-Microbe Interact. 33, 693–703. doi: 10.1094/MPMI-12-19-0339-R
Nisa, M. U., Huang, Y., Benhamed, M., and Raynaud, C. (2019). The plant DNA damage response: signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 10:653. doi: 10.3389/fpls.2019.00653
Ochsenbein, C., Przybyla, D., Danon, A., Landgraf, F., Göbel, C., Imboden, A., et al. (2006). The role of EDS1 (enhanced disease susceptibility) during singlet oxygen-mediated stress responses of Arabidopsis. Plant J. 47, 445–456. doi: 10.1111/j.1365-313X.2006.02793.x
Pečenková, T., Pejchar, P., Moravec, T., Drs, M., Haluška, S., Šantrůček, J., et al. (2022). Immunity functions of Arabidopsis pathogenesis-related 1 are coupled but not confined to its C-terminus processing and trafficking. Mol. Plant Pathol. 23, 664–678. doi: 10.1111/mpp.13187
Peng, X., Zhang, X., Li, B., and Zhao, L. (2019). Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating nitric oxide production via cytosolic calcium ions. BMC Plant Biol. 19, 368–313. doi: 10.1186/s12870-019-1974-9
Phukan, U. J., Jeena, G. S., and Shukla, R. K. (2016). WRKY transcription factors: molecular regulation and stress responses in plants. Front. Plant Sci. 7:760. doi: 10.3389/fpls.2016.00760
Ranty, B., Aldon, D., Cotelle, V., Galaud, J. P., Thuleau, P., and Mazars, C. (2016). Calcium sensors as key hubs in plant responses to biotic and abiotic stresses. Front. Plant Sci. 7:327. doi: 10.3389/fpls.2016.00327
Rao, D., Momcilovic, I., Kobayashi, S., Callegari, E., and Ristic, Z. (2004). Chaperone activity of recombinant maize chloroplast protein synthesis elongation factor, EF-Tu. Eur. J. Biochem. 271, 3684–3692. doi: 10.1111/j.1432-1033.2004.04309.x
Reddy, D. S., Bhatnagar-Mathur, P., Reddy, P. S., Sri Cindhuri, K., Sivaji Ganesh, A., and Sharma, K. K. (2016). Identification and validation of reference genes and their impact on normalized gene expression studies across cultivated and wild cicer species. PLoS One 11:e0148451. doi: 10.1371/journal.pone.0148451
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rudenko, N. N., Borisova-Mubarakshina, M. M., Ignatova, L. K., Fedorchuk, T. P., Nadeeva-Zhurikova, E. M., and Ivanov, B. N. (2021). Role of plant carbonic anhydrases under stress conditions. Plant Stress Physiol. 4, 301–325. doi: 10.5772/intechopen.91971
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative C T method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Sharma, M., Nagavardhini, A., Thudi, M., Ghosh, R., Pande, S., and Varshney, R. K. (2014). Development of DArT markers and assessment of diversity in Fusarium oxysporum f. sp. ciceris, wilt pathogen of chickpea (Cicer arietinum L.). BMC Genomics 15, 454–414. doi: 10.1186/1471-2164-15-454
Sharma, M., Sengupta, A., Ghosh, R., Agarwal, G., Tarafdar, A., Nagavardhini, A., et al. (2016). Genome wide transcriptome profiling of Fusarium oxysporum f sp. ciceris conidial germination reveals new insights into infection-related genes. Sci. Rep. 6, 1–11. doi: 10.1038/srep37353
Sinha, A. K., Jaggi, M., Raghuram, B., and Tuteja, N. (2011). Mitogen-activated protein kinase signaling in plants under abiotic stress. Plant Signal. Behav. 6, 196–203. doi: 10.4161/psb.6.2.14701
Song, J., Zou, X., Liu, P., Cardoso, J. A., Schultze-Kraft, R., Liu, G., et al. (2022). Differential expressions and enzymatic properties of malate dehydrogenases in response to nutrient and metal stresses in Stylosanthes guianensis. Plant Physiol. Biochem. 170, 325–337. doi: 10.1016/j.plaphy.2021.12.012
Su, W., Ren, Y., Wang, D., Su, Y., Feng, J., Zhang, C., et al. (2020). The alcohol dehydrogenase gene family in sugarcane and its involvement in cold stress regulation. BMC Genomics 21, 521–517. doi: 10.1186/s12864-020-06929-9
Sun, M., Qiu, L., Liu, Y., Zhang, H., Zhang, Y., Qin, Y., et al. (2022). Pto interaction proteins: critical regulators in plant development and stress response. Front. Plant Sci. 13:774229. doi: 10.3389/fpls.2022.774229
Szurman-Zubrzycka, M., Jędrzejek, P., and Szarejko, I. (2023). How do plants cope with DNA damage? A concise review on the DDR pathway in plants. Int. J. Mol. Sci. 24:2404. doi: 10.3390/ijms24032404
Upasani, M. L., Limaye, B. M., Gurjar, G. S., Kasibhatla, S. M., Joshi, R. R., Kadoo, N. Y., et al. (2017). Chickpea-Fusarium oxysporum interaction transcriptome reveals differential modulation of plant defense strategies. Sci. Rep. 7, 1–12. doi: 10.1038/s41598-017-07114-x
Wang, Y., Feng, G., Zhang, Z., Liu, Y., Ma, Y., Wang, Y., et al. (2021). Overexpression of Pti4, Pti5, and Pti6 in tomato promote plant defense and fruit ripening. Plant Sci. 302:110702. doi: 10.1016/j.plantsci.2020.110702
Wang, L., Li, S., Sun, L., Tong, Y., Yang, L., Zhu, Y., et al. (2022). Over-expression of phosphoserine aminotransferase-encoding gene (AtPSAT1) prompts starch accumulation in L. turionifera under nitrogen starvation. Int. J. Mol. Sci. 23:11563. doi: 10.3390/ijms231911563
Wang, J., Tian, W., Tao, F., Wang, J., Shang, H., Chen, X., et al. (2020). TaRPM1 positively regulates wheat high-temperature seedling-plant resistance to Puccinia striiformis f. sp. tritici. Frontiers. Plant Sci. 10:1679. doi: 10.3389/fpls.2019.01679
Wang, H., Zhao, P., Shen, X., Xia, Z., Zhou, X., Chen, X., et al. (2021). Genome-wide survey of the phosphofructokinase family in cassava and functional characterization in response to oxygen-deficient stress. BMC Plant Biol. 21, 376–315. doi: 10.1186/s12870-021-03139-7
Wang, N., Zhong, X., Cong, Y., Wang, T., Yang, S., Li, Y., et al. (2016). Genome-wide analysis of phosphoenolpyruvate carboxylase gene family and their response to abiotic stresses in soybean. Sci. Rep. 6, 1–14. doi: 10.1038/srep38448
Waseem, M., and Ahmad, F. (2019). The phosphoenolpyruvate carboxylase gene family identification and expression analysis under abiotic and phytohormone stresses in Solanum lycopersicum L. Gene 690, 11–20. doi: 10.1016/j.gene.2018.12.033
Wu, W., Luo, X., and Ren, M. (2022). Clearance or hijack: universal interplay mechanisms between viruses and host autophagy from plants to animals. Front. Cell. Infect. Microbiol. 11:1345. doi: 10.3389/fcimb.2021.786348
Wu, Y., Zhang, L., Zhou, J., Zhang, X., Feng, Z., Wei, F., et al. (2021). Calcium-dependent protein kinase GhCDPK28 was identified and involved in verticillium wilt resistance in cotton. Front. Plant Sci. 12:772649. doi: 10.3389/fpls.2021.772649
Xue, R., Wu, X., Wang, Y., Zhuang, Y., Chen, J., Wu, J., et al. (2017). Hairy root transgene expression analysis of a secretory peroxidase (PvPOX1) from common bean infected by Fusarium wilt. Plant Sci. 260, 1–7. doi: 10.1016/j.plantsci.2017.03.011
Yadav, S., Rathore, M. S., and Mishra, A. (2020). The pyruvate-phosphate dikinase (C4-SmPPDK) gene from Suaeda monoica enhances photosynthesis, carbon assimilation, and abiotic stress tolerance in a C3 plant under elevated CO2 conditions. Front. Plant Sci. 11:345. doi: 10.3389/fpls.2020.00345
Yadava, Y. K., Chaudhary, P., Yadav, S., Rizvi, A. H., Kumar, T., Srivastava, R., et al. (2023). Genetic mapping of quantitative trait loci associated with drought tolerance in chickpea (Cicer arietinum L.). Sci. Rep. 13:17623. doi: 10.1038/s41598-023-44990-y
Yan, Z., Xingfen, W., Wei, R., Jun, Y., and Zhiying, M. (2016). Island cotton enhanced disease susceptibility 1 gene encoding a lipase-like protein plays a crucial role in response to verticillium dahliae by regulating the SA level and H2O2 accumulation. Front. Plant Sci. 7:1830. doi: 10.3389/fpls.2016.01830
Yang, X., Chen, L., Yang, Y., Guo, X., Chen, G., Xiong, X., et al. (2020). Transcriptome analysis reveals that exogenous ethylene activates immune and defense responses in a high late blight resistant potato genotype. Sci. Rep. 10, 21294–21314. doi: 10.1038/s41598-020-78027-5
Yin, G., Sun, H., Xin, X., Qin, G., Liang, Z., and Jing, X. (2009). Mitochondrial damage in the soybean seed axis during imbibition at chilling temperatures. Plant Cell Physiol. 50, 1305–1318. doi: 10.1093/pcp/pcp074
Zeng, H., Xu, L., Singh, A., Wang, H., Du, L., and Poovaiah, B. W. (2015). Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 6:600. doi: 10.3389/fpls.2015.00600
Zhang, M., and Zhang, S. (2022). Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 64, 301–341. doi: 10.1111/jipb.13215
Keywords: chickpea, Fusarium wilt, RNA sequencing, transcriptome, environmental adaptation, energy metabolism
Citation: Bhutia KL, Ahmad M, Kisku A, Sudhan RA, Bhutia ND, Sharma VK, Prasad BD, Thudi M, Obročník O, Bárek V, Brestic M, Skalicky M, Gaber A and Hossain A (2024) Shoot transcriptome revealed widespread differential expression and potential molecular mechanisms of chickpea (Cicer arietinum L.) against Fusarium wilt. Front. Microbiol. 14:1265265. doi: 10.3389/fmicb.2023.1265265
Edited by:
Mahyar Mirmajlessi, Ghent University, BelgiumReviewed by:
Arindam Ghatak, University of Vienna, AustriaArshi Jamil, Aligarh Muslim University, India
Copyright © 2024 Bhutia, Ahmad, Kisku, Sudhan, Bhutia, Sharma, Prasad, Thudi, Obročník, Bárek, Brestic, Skalicky, Gaber and Hossain. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karma L. Bhutia, klbhutia@rpcau.ac.in; Akbar Hossain, akbarhossainwrc@gmail.com