
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 23 November 2023
Sec. Microorganisms in Vertebrate Digestive Systems
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1228845
This article is part of the Research TopicWomen in Microorganisms in Vertebrate Digestive Systems: 2023View all 17 articles
 Anne Kauter1
Anne Kauter1 Julian Brombach2,3
Julian Brombach2,3 Antina Lübke-Becker2,3Dania Kannapin4
Antina Lübke-Becker2,3Dania Kannapin4 Corinna Bang5Sören Franzenburg5Sabita D. Stoeckle4
Corinna Bang5Sören Franzenburg5Sabita D. Stoeckle4 Alexander Mellmann6
Alexander Mellmann6 Natalie Scherff6Robin Köck6,7
Natalie Scherff6Robin Köck6,7 Sebastian Guenther8Lothar H. Wieler9
Sebastian Guenther8Lothar H. Wieler9 Heidrun Gehlen4
Heidrun Gehlen4 Torsten Semmler10
Torsten Semmler10 Silver A. Wolf10†
Silver A. Wolf10† Birgit Walther1,11*†
Birgit Walther1,11*†Introduction: Horse clinics are hotspots for the accumulation and spread of clinically relevant and zoonotic multidrug-resistant bacteria, including extended-spectrum β-lactamase producing (ESBL) Enterobacterales. Although median laparotomy in cases of acute equine colic is a frequently performed surgical intervention, knowledge about the effects of peri-operative antibiotic prophylaxis (PAP) based on a combination of penicillin and gentamicin on the gut microbiota is limited.
Methods: We collected fecal samples of horses from a non-hospitalized control group (CG) and from horses receiving either a pre-surgical single-shot (SSG) or a peri-operative 5-day (5DG) course of PAP. To assess differences between the two PAP regimens and the CG, all samples obtained at hospital admission (t0), on days three (t1) and 10 (t2) after surgery, were screened for ESBL-producing Enterobacterales and subjected to 16S rRNA V1–V2 gene sequencing.
Results: We included 48 samples in the SSG (n = 16 horses), 45 in the 5DG (n = 15), and 20 in the CG (for t0 and t1, n = 10). Two samples of equine patients receiving antibiotic prophylaxis (6.5%) were positive for ESBL-producing Enterobacterales at t0, while this rate increased to 67% at t1 and decreased only slightly at t2 (61%). Shannon diversity index (SDI) was used to evaluate alpha-diversity changes, revealing there was no significant difference between horses suffering from acute colic (5DG, SDImean of 5.90, SSG, SDImean of 6.17) when compared to the CG (SDImean of 6.53) at t0. Alpha-diversity decreased significantly in both PAP groups at t1, while at t2 the onset of microbiome recovery was noticed. Although we did not identify a significant SDImean difference with respect to PAP duration, the community structure (beta-diversity) was considerably restricted in samples of the 5DG at t1, most likely due to the ongoing administration of antibiotics. An increased abundance of Enterobacteriaceae, especially Escherichia, was noted for both study groups at t1.
Conclusion: Colic surgery and PAP drive the equine gut microbiome towards dysbiosis and reduced biodiversity that is accompanied by an increase of samples positive for ESBL-producing Enterobacterales. Further studies are needed to reveal important factors promoting the increase and residency of ESBL-producing Enterobacterales among hospitalized horses.
Compared with other companion animals, horses more often acquire gastro-intestinal tract (GIT) disorders that may lead to long-term suffering or even death (Traub-Dargatz et al., 2001). The complex pain caused by disorders of the GIT in horses is commonly referred to as “colic” (Traub-Dargatz et al., 2001; Stockle et al., 2018) and often requires surgical intervention (Archer, 2019). The composition of the bacterial community residing within the GIT has been regarded as beneficial and a prerequisite for the health and well-being of hindgut fermenters such as Equidaes (reviewed in Kauter et al., 2019). Previous reports indicated that colonization with the physiological endogenous microbiota shields the equine GIT against either direct or indirect pathogen-induced damage and that these protective effects are disrupted in various enteric diseases (Costa et al., 2012; Weese et al., 2015). In addition, it has been reported as a consequence of the administration of antibiotics such as penicillin (Baverud et al., 2003), enrofloxacin or ceftiofur (Liepman et al., 2022), and doxycycline (Davis et al., 2006) that the enteric microbial community enters a dysbiotic state (Costa et al., 2015). However, among other factors, the state of the microbiota at the time of the (antibiotic) perturbation (diet, species, functional diversity, and redundancy) and the characteristics of the perturbation (e.g., route of administration, antimicrobial spectrum, and duration of the antibiotic course) determine the extent of any eventual dysbiosis (Schwartz et al., 2020).
Peri-operative antibiotic prophylaxis (PAP) is recommended to prevent adverse postoperative events, such as surgical site infections, in horses subjected to abdominal surgery due to acute colic (Dallap Schaer et al., 2012). The most commonly used PAP regimen for horses requiring median laparotomy consists of a combination of penicillin and gentamicin (P/G) for a period of 3–10 days (Dallap Schaer et al., 2012; Wormstrand et al., 2014; Teschner et al., 2015; Ceriotti et al., 2021). In contrast, the standard regimen for similar surgical interventions lacking complicating circumstances in human and small animal medicine is a short-term PAP, which is provided as a single-shot therapy 30–60 min prior to incision (as described in Stöckle et al., 2021). To investigate the effects of prolonged administration beyond this immediate peri-operative timeframe (> 24 h after surgery), we conducted a pilot study focusing on the clinical outcome (Stöckle et al., 2021), the extended-spectrum β-lactamase (ESBL)-producing Escherichia coli (EC) carriage rates (Kauter et al., 2021), and the microbiome composition (present study). Beyond others, our results indicated non-inferiority of a “single-shot” vs. a 5-day course of P/G PAP with respect to the patients’ clinical outcomes (Stöckle et al., 2021). Regardless of the applied PAP-regimen, we noticed a worrisome increase of ESBL-EC carriage rates among horses during their hospital stay (Kauter et al., 2021).
Based on these previous observations, the current study aimed (i) to examine the extent of microbiome disturbance in hospitalized horses subjected to median laparotomy, (ii) to reveal alterations of the gut microbiota caused by a short and a long-term PAP regime, and (iii) to enable insights into changes of the microbiome that might play a role in the previously reported increased ESBL-EC carriage rates among equine patients.
Horses diagnosed with acute abdominal pain (colic) that required median laparotomy were included in this study and were randomized 1: 1 to the PAP regimens in an open-label study as the different antibiotic regimens prevented blinding (for a detailed description of the controlled and randomized study, see in Stöckle et al., 2021). Briefly, horses assigned to the SSG received short term P/G PAP, while the 5DG group received P/G PAP for 5 consecutive days. In both groups, PAP consisted of parenteral administration of sodium penicillin G (22,000 IU/kg four times daily) and gentamicin (6.6 mg/kg), as previously recommended for colic surgery (Dallap Schaer et al., 2012; Durward-Akhurst et al., 2013; Southwood, 2014). Specimens of 10 non-hospitalized farm horses that were free of clinical symptoms for any apparent illnesses served as a control group (CG). All animals in the CG were sampled within at a 3-day interval (t0 and t1), similarly to the hospitalized animals (see description of sample collection), in order to ensure the representativeness of the samples of this group (for a detailed description of the study cohort and their respective clinical results, see Stöckle et al., 2021). A graphical abstract of the study outline is provided in Figure 1.
Figure 1. Illustration of the sampling procedure. Horses subjected to colic surgery were (1) sampled directly at hospital admission (t0) and (2) allotted to either a single-shot perioperative antibiotic prophylaxis regimen or a 5-day-lasting protocol. Sampling was repeated for all horses on day 3 and 10 after surgery (3). All samples were subjected to DNA extraction and V1–V2 16S rRNA gene sequencing (4). Sequences were preprocessed and analyzed with respect to changes of microbiota composition, alpha and beta diversity (5).
Inclusion criteria for study participation were: study participants had to be free of clinical signs of infectious diseases prior to surgery. Additionally, since the juvenile microbiome is known for continuing changes during the foals’ gut maturation (Costa et al., 2016), all included horses were required to be 1 year or older. Equine patients were excluded from further consideration if their hospital stay had ended prematurely due to euthanasia and/or the antibiotic regimen they had originally been assigned to was not strictly followed, regardless of the particular reasons requiring these changes. Horses with incomplete sample sets (t0, t1, and t2, see sample collection) or resection (enhanced rates of surgical site infections, SSIs) were excluded from this study. For additional details on the clinical characteristics of colic horses, see Stöckle et al. (2021).
Fresh fecal samples were collected from each horse diagnosed with colic and undergoing midline celiotomy directly at hospital admission (t0), as described previously (Walther et al., 2018; Kauter et al., 2021). A second sample was collected 3 days (t1) after, and a third (final) sample, 10 days (t2, hospital discharge) after surgery. In order to gain insights into the gut microbiome associated with non-hospitalized horses lacking clinical signs of gastro-intestinal disorders, control samples were obtained from 10 horses residing in a barn. All specimens were stored directly at −80°C and shipped on dry ice.
Identification of ESBL-producing Enterobacterales was previously described (Kauter et al., 2021). Briefly, samples were cultured on Brilliance™ ESBL Agar plates (Thermo Scientific, Germany) overnight. Colonies showing characteristic growth signatures of ESBL-producing Enterobacterales on chromogenic screening plates were further investigated. In case of distinct phenotypic appearances of presumptive ESBL-producing colonies on the plates, all isolates were subjected to an ESBL confirmation test according to the Clinical Laboratory Standards Institute’s (CLSI) recommendations [Clinical and Laboratory Standards Institute (CLSI), 2020]. Species confirmation was achieved by matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometry (Bruker, Germany).
The sequencing of bacterial DNA was performed by the Institute of Clinical Molecular Biology (IKMB) at the Christian-Albrechts University of Kiel. DNA was extracted using the QIAamp Fast DNA stool mini kit (QIAGEN, Hilden, Germany) automated on the QIAcube (QIAGEN, Hilden, Germany). For this, approximately 200 mg feces were transferred to 0.70 mm Garnet Bead tubes filled with 1 mL InhibitEx buffer. Subsequently, bead beating was performed using a SpeedMill PLUS (QIAGEN, Hilden, Germany) for 45 s at 60 Hz. Samples were then heated to 95°C for 5 min and afterwards centrifuged for 1 min at 10,000 rpm. 200 μL of the resulting supernatant were transferred to a 1.5 mL microcentrifuge tube, which was placed in the QIAcube for follow-up automated DNA isolation according to the manufacturer’s protocol. DNA bound specifically to the QIAamp silica-gel membrane while contaminants passed through. PCR inhibitors were removed through means of InhibitEX (QIAGEN, Hilden, Germany), a unique adsorption resin, and an optimized buffer. DNA was diluted 1:10 prior to PCR, and 3 μL were used for further amplification. PCR-products were verified using agarose gel electrophoresis. PCR-products were then normalized using the SequalPrep Normalization Plate Kit (Thermo Fischer Scientific, Waltham, MA, United States), pooled equimolarily, and sequenced on an Illumina MiSeq v3 2 × 300 base pair (bp) platform (Illumina Inc., San Diego, CA, United States).
The V1–V2 region of the 16S rRNA gene was subsequently sequenced (Primerpair 27F-338R, dual MIDs inducing) on a MiSeq-platform (MiSeq Reagent Kit v2; Kozich et al., 2013). The resulting MiSeq raw fastq data were verified using an inhouse pipeline [Bcl2fastq Modul in CASAVA 1.8.2, Demultiplexing, FLASH software (v1.2) (Magoč and Salzberg, 2011), and fastx toolkit und UCHIME (v6.0) (Edgar et al., 2011)]. Demultiplexing was performed based on zero mismatches within the barcode sequences.
Sequence reads were preprocessed as described (Mach et al., 2020). In brief, paired-end reads were merged into continuous sequences using the “join_paired_ends.py” script of QIIME (v1.9.1) (Caporaso et al., 2010) in “fastq-join” mode (Aronesty, 2013). Defined parameters allowed a minimum overlap of 6 bp and a maximum difference within the overlap region of 8%. Reads which did not meet these criteria were removed from further analysis. Next, seqkit (v0.16.1) (Shen et al., 2016) was utilized to filter out reads which were too short (≤300 bp) or too long (≥470 bp). The remaining sequences were then quality filtered using the “split_libraries_fastq.py” script of QIIME (v1.9.1) (Edgar, 2010) by applying a PHRED quality threshold of 20. Reads were hereby required to have 50% of their bases to be consecutively of high quality. A maximum of three consecutive low-quality bases were allowed before truncating a read. Reads containing any ambiguous bases (“N”) were removed from further analysis.
Reads were clustered into operational taxonomic units (OTUs) using USEARCH (v11.0.667) (Edgar, 2010) and the Greengenes database (release 2013-08, gg_13_8_otus, 99_otus; DeSantis et al., 2006) with a 97% similarity cutoff. The “unnoise3” algorithm of USEARCH was utilized for additional filtering of chimeric OTUs. In order to improve taxonomic annotation, Greengenes OTUs as well as unmatched representative sequences were then mapped against the RDP database (v11.5) (Cole et al., 2014) using the SequenceMatch pipeline of rdptools (v2.0.3) (Cole et al., 2014), searching for the closest neighbor (k = 1) with a minimum sab score of 0.5. Counts were subsequently merged and the “filter_otus_from_otu_table.py” script of QIIME (v1.9.1) (Caporaso et al., 2010) was utilized to remove any singletons from the table (counts ≤3). Data were then exported using the biom (v 2.1.7) (McDonald et al., 2012) package and converted into the appropriate format for further data analysis. The resulting OTU table was then finalized by updating the available taxonomic labels with the recently released, new taxonomic names for bacterial phyla (Oren and Garrity, 2021).
Sample-size calculation was performed as recommended for microbiome studies by utilizing a permutation-based extension of the multivariate analysis of variance (PERMANOVA) on a matrix of pairwise distances (Kelly et al., 2015). Subsequent study power estimation was performed through the R package micropower (v0.4) (Kelly et al., 2015). Unfiltered OTU tables were hereby subjected to random rarefaction in order to assess key population parameters, including mean and standard deviation per OTU based on both the presence/absence of individual taxa as well as their abundance (Weighted Jaccard Distance) for a comparison of two groups of fixed size (n = 10). These parameters were then further utilized to simulate a range of distance matrices and effect sizes in order to estimate the statistical power for identifying an effect size given a specified sample size and respective OTU table. Permutation-based sample-size estimation, according to Kelly et al. (2015), revealed that a subset of samples from 10 horses per study group were sufficient to identify differences in taxonomic composition of effect size 0.020 with p = 0.05 and a power of 80% (90% to identify an effect size of 0.035).
The resulting OTU counts were randomly sub-sampled for each sample to a homogeneous level, defined by the counts of the lowest sample (12,178). OTU counts above 10,000 have been shown to provide adequate comparisons between differing sequencing depths for microbiome analyses (Mach et al., 2020). Rarefaction was performed by using the “rarefy_even_depth” function of phyloseq (v1.36.0) (McMurdie and Holmes, 2013) with rngseed = 1 in R (v4.1.1). Rarefied data were then utilized to assess the influence of antibiosis on the microbiome diversity among the equine patients and to visualize the distribution of taxa (OTUs) across the sample set. Within-sample diversity (alpha-diversity) was assessed through Shannon diversity indices (SDI) calculated using the R package microbiome (v1.14.0) (Lahti et al., 2017). Between-sample diversity (beta-diversity) was determined through the computation of Bray-Curtis distances by phyloseq (v1.36.0) (McMurdie and Holmes, 2013) on all OTUs of the rarefied table. The resulting diversity metrics were further visualized with ComplexHeatmap (v2.8.0) (Gu et al., 2016) and ggplot2 (v3.3.5) (Wickham et al., 2016) for subsequent comparisons between the study groups. Non-parametric Paired Wilcoxon Rank Sum tests were performed between groups of interest and results with a p < 0.05 were labeled as being significant. The Benjamini-Hochberg procedure was utilized for multiple-testing correction in order to limit the false discovery rate were applicable. OTUs were aggregated at selected taxonomic levels (including phlya) using the “aggregate_top_taxa” function of microbiome (v1.14.0) (Lahti et al., 2017). Differential taxa were identified between the PAP study groups (SSG/5DG) using the raw OTU table and the microbiomeExplorer R package (Reeder et al., 2021) with proportional normalization and the DESeq2 method (Love et al., 2014). Additional correlation analyses were also performed within the microbiome Explorer suite.
To explore the effects of both G/P PAP regimens and hospitalization on the gut microbiota of horses subjected to median laparotomy, microbiome sequencing and analysis were performed using sample sets obtained from a pilot study comparing the clinical outcomes of 67 patients that met the study’s inclusion criteria (Stöckle et al., 2021). Overall, 48 samples from 16 horses representing the SSG and 45 samples of 15 horses belonging to the 5DG were comparatively evaluated. As a non-hospitalized CG, we included additional 20 samples of 10 horses that lacked any apparent signs of clinical illness, resulting in 113 samples (Supplementary Table 1). At hospital admission, two samples (6.5%) were positive for ESBL-producing Enterobacterales (Supplementary Table 1), while, regardless of the study group, the overall rate increased to 67% (t1) and, only slightly decreased, at t2 (61%). There was no difference in carriage of ESBL-producing Enterobacterales between the SSG and 5DG (Fisher’s Exact Test, p = 1), while all CG samples were negative (Fisher’s Exact Test, p < 0.0001).
In total, 4,896,645 high quality OTU counts (ranging between 12,178–220,454 counts per sample, median = 32,730) were obtained and assigned to 17,035 different OTUs across 330 different taxonomic entities at genus level. Further taxonomic assignment of these OTUs revealed the top 10 bacterial taxa (phylum level) based on their total counts across the sample set: The phyla Bacteroidota (38%; previously Bacteroidetes; Oren and Garrity, 2021) and Bacillota (33%; previously: Firmicutes; Oren and Garrity, 2021) were predominant in the equine fecal samples at hospital admission (t0), followed by Verrucomicrobiota (11%), Pseudomonadota (9%; previously Proteobacteria; Oren and Garrity, 2021) and Spirochaetota (4%; Figure 2).

Figure 2. Taxonomic composition of microbiota in hospitalized horses. Stacked bar charts illustrating the relative abundances of the top 10 phyla for the single-shot group (SSG, n = 16) and the 5-day group (5DG, n = 15). t0, Hospital admission (SSG/5DG)/first sampling round (CG); t1 = 3 days after surgery (SSG/5DG)/3 days after first sampling (CG); t2 = 10 days after surgery (SSG/5DG).
In addition, each horse harbored an individual composition of bacterial fecal communities, as presented in Supplementary Figure 1.
The SDI is a measure of a sample’s diversity based on both, community richness and evenness. In the present study, SDI was selected to inspect the alpha-diversity within each sample and to compare these across study groups. Overall comparableness of the treatment groups was ensured by demonstrating the lack of significant difference in the mean SDI between both study groups at t0 (Wilcoxon test, p > 0.05). As expected, visualization of beta-diversity (“between sample diversity”) using ordination plots generated through principle component analysis (PCA) demonstrated an increased range of variance regarding the gut microbiota profiles of individual horses belonging to the SSG and the 5DG compared to the overall variance noticeable between the data points representing the CG samples (Figures 3A–C).
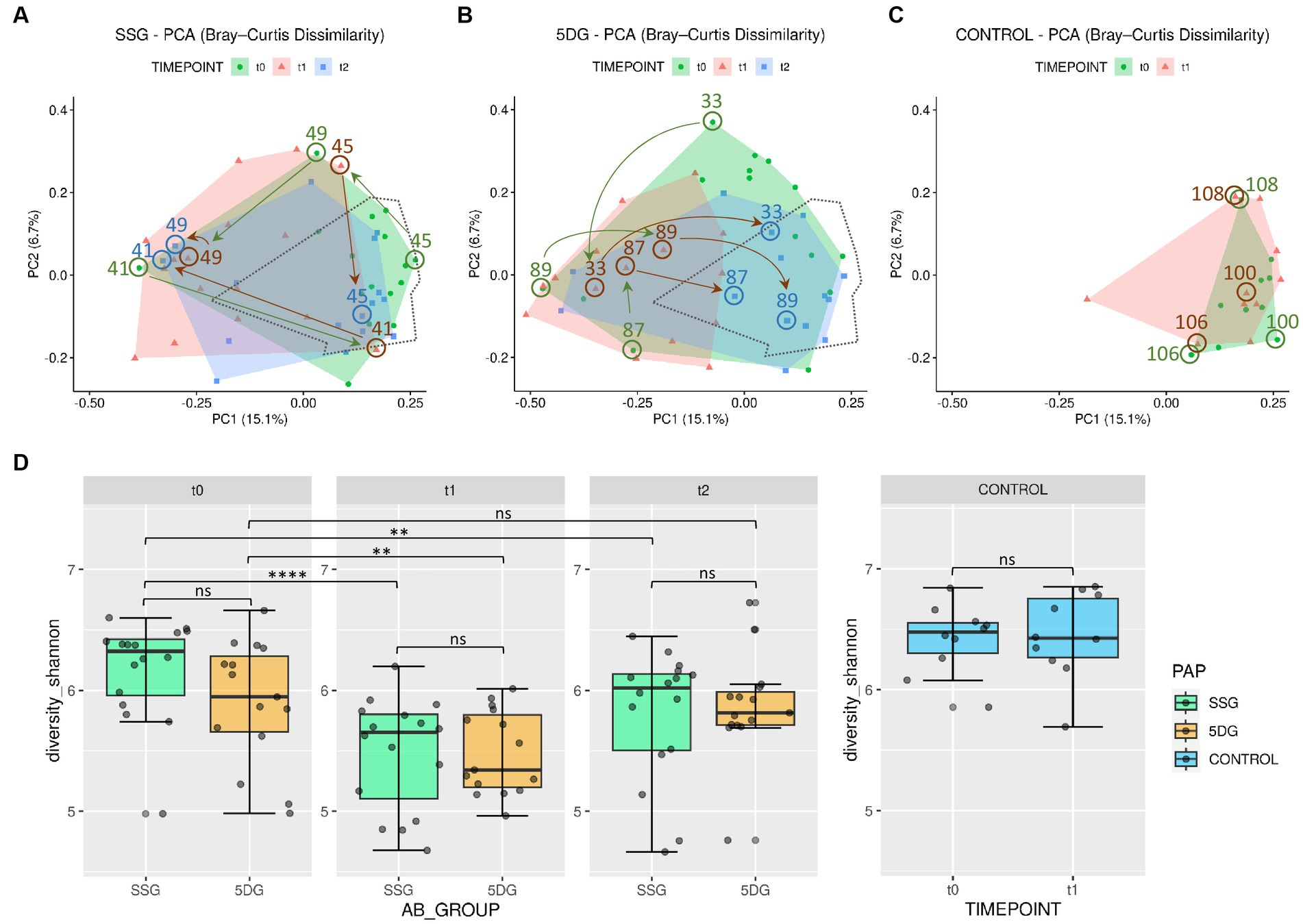
Figure 3. Diversity of the equine gut microbiome of hospitalized and non-hospitalized horses. (A–C) Principle component analysis (PCA) plots illustrating differences in gut microbiome composition (beta-diversity) based on unweighted Bray-Curtis dissimilarities across each group per sampling day. Axes represent the two dimensions that explain the largest proportion of variation across communities of each analysis. Distances between two data points reflect their similarity/dissimilarity with respect to sample composition. The colored areas represent the computed convex hull of data points from each sampling day, illustrating respective areas of minimal size. The area encompassing data points from the control group (CG) samples (C) is marked by a dotted line in (A,B) for direct comparison. Horses treated with peri-operative antibiotic prophylaxis (PAP) in the single-shot group (SSG) vs. the 5-day course group (5DG) show both large, intraindividual differences at t0 (A) and a strong shift away from the area defined by the CG samples at t1 (B). Samples of both study groups converge back to the CG area at t2. Numbered arrows indicate the trajectories of microbiota composition within selected horses. (D) Boxplots displaying the alpha-diversity indices (Shannon) for all three groups (SSG, n = 16; 5DG, n = 15; and control, n = 10; significance between groups: unpaired Wilcoxon Rank Sum tests, **p < 0.05, ****p < 0.0005, ns, not significant).
Of note, the mean SDI values of the CG samples were 6.42 and 6.44, respectively. At t0, most of the enteral microbiota profiles associated with SSG horses (11 of 16) clustered within the framework set by the CG samples (Figure 3A). The t0 microbiota profiles obtained for the 5DG, on the other hand, illustrated increased variance among the 5DG samples, and only four out of 15 samples clustered with the CG area, potentially indicating that more horses belonging to the 5DG had, in comparison to the SSG, gut microbiota perturbances at hospital admission (Figure 3B). This observation also seemed to correspond with the alpha-diversities, since the t0 samples representing the SSG were associated with an SDImean of 6.17, while the 5DG yielded an SDImean of 5.90, however the difference lacked statistical significance (Figure 3D).
At t1, most of the microbiota profiles obtained from fecal samples of the SSG (horses without antibiotics for at least 36 h) and the 5DG (horses at the third day of their 5-day course P/G PAP) visibly differed from those obtained at t0 by shifting away from the center of the CG area, indicating perturbations (Figure 3B). This shift was accompanied by a significant reduction in mean alpha-diversity across both groups [SSG: SDI from t0 = 6.17 to t1 = 5.48 (paired Wilcoxon Test, p = 0.000031); 5DG: SDI from t0 = 5.90 to t1 = 5.48 (paired Wilcoxon Test, p = 0.012); Supplementary Table 1; Figure 3D]. In addition, the overall distances between 5DG samples seemed considerably restricted compared to the situation at t0 (Figure 3B). Samples representing the SSG, on the other hand, were more widely scattered than samples representing the 5DG or even the CG, indicating an increased level of inter-individual differences among microbiome compositions for SSG horses at t1.
At t2, i.e., 10 (SSG) and 5 days (5DG) after the final PAP course was administered, the mean alpha-diversity of both study groups increased (SSG, SDImean = 5.80; 5DG, SDImean = 5.87), indicating the onset of microbiome recovery (Supplementary Table 1; Figure 3C).
To likewise illustrate common spatial shifts and individual deviation from common trajectories, data points representing samples of three individuals per group are highlighted in Figure 3.
At t0, the data points belonging to samples of equine patients 89, 33, and 87 clustered most distantly from the CG samples, indicating a considerable disturbance of the microbiome structure and composition at hospital admission. This finding is supported by the individual samples’ low SDI values (5.05, 5.75, and 5.29; Supplementary Table 1). At t1, samples 33 and 87 showed a different composition compared to the t0 situation, but both points still clustered distantly from the CG area. Only the t1 sample obtained from horse 89 showed some signs of movement toward the CG area (Figure 3B). At t2, data points representing the samples of horses 33, 87, and 89 clustered within the CG area, indicating the onset of microbiome recovery that was accompanied by a notable SDI increase (5.76, 6.02, and 5.76).
The initial data point representing the t0 sample of horse 45 (SDI 6.66) clustered near the area covered by the CG. While the corresponding data point at t1 indicated an increase of gut microbiome disturbance accompanied by an SDI decrease to 5.97, the t2 sample (SDI 6.15) clustered within the area framed by data points of the CG samples, once again indicating the onset of microbiome recovery.
However, some equine patients deviated from the aforementioned common temporal trajectory: The data points of horse 41 (SSG), for instance, indicated a considerable microbiome disturbance at t0 (SDI 5.05) followed by a brief relative recovery at t1 (SDI 5.99). Then, the data point of horse 41 shifted toward the opposite direction at t2 (Figure 3A), a reversal that is accompanied by an SDI decrease to 5.59.
Taken together, samples of the CG seemed to represent an overall beneficial structure and composition of equine gut microbiota, since all patients’ samples associated with an SDI > 5.8 clustered near or within the area covered by these samples, while samples associated with lower SDIs clustered elsewhere (Figure 3; Supplementary Table 1). Moreover, although a common temporal trajectory from hospital admission toward discharge was notable for the gut microbiota of most participants in both study groups, the temporal patterns of some horses deviated from these, emphasizing the individual nature of the GIT microbiome recovery process.
To gain insights into the putative effect of the different PAP regimens on the equine gut microbiota, log 2-fold changes (log2FC) were calculated based on OTU count changes on bacterial family level between sampling timepoints. Overall, the most prominent log2FC between t0 and t1 in the SSG were noticed for Bacteroidaceae (+5.16; p < 0.05), Enterobacteriaceae (+3.99; p < 0.05), and Pseudomonadaceae (+4.41; p < 0.05). Among the most prominent log2FC between t0 and t1 in samples of the 5DG were Bacteroidaceae (+2.96; p < 0.05) and Pseudomonadaceae (+3.33; p < 0.05).
In order to further investigate the relevance of the aforementioned observations and to better assess the influence of colic and midline laparotomy as well as antibiotic treatment on the abundances of specific bacterial families in the GIT of horses throughout this study, further log2FC values were determined based on the abundances calculated for the CG. Since the PCA confirmed the eligibility of the CG samples with respect to an overall favorable equine gut microbiota structure and composition, we determined the median relative abundance for bacteria (family level) among the CG samples. To enhance identification of relevant changes associated with microbiome disturbance among samples belonging to both study groups, the median for each bacterial family was calculated using the t0 CG samples as a baseline. Then, log2FC for each sample/timepoint/study group/ were determined to pin-point variation that might have been overlooked by simply investigating mean abundances. In order to compensate for individual differences between the equine patients (i.e., age, diet, medical history, severity and duration of the acute colic episode, housing and social contacts), an interval comprising at least 85% of the SSG and 5DG samples was defined (Figure 4; log2FC ±2.5).
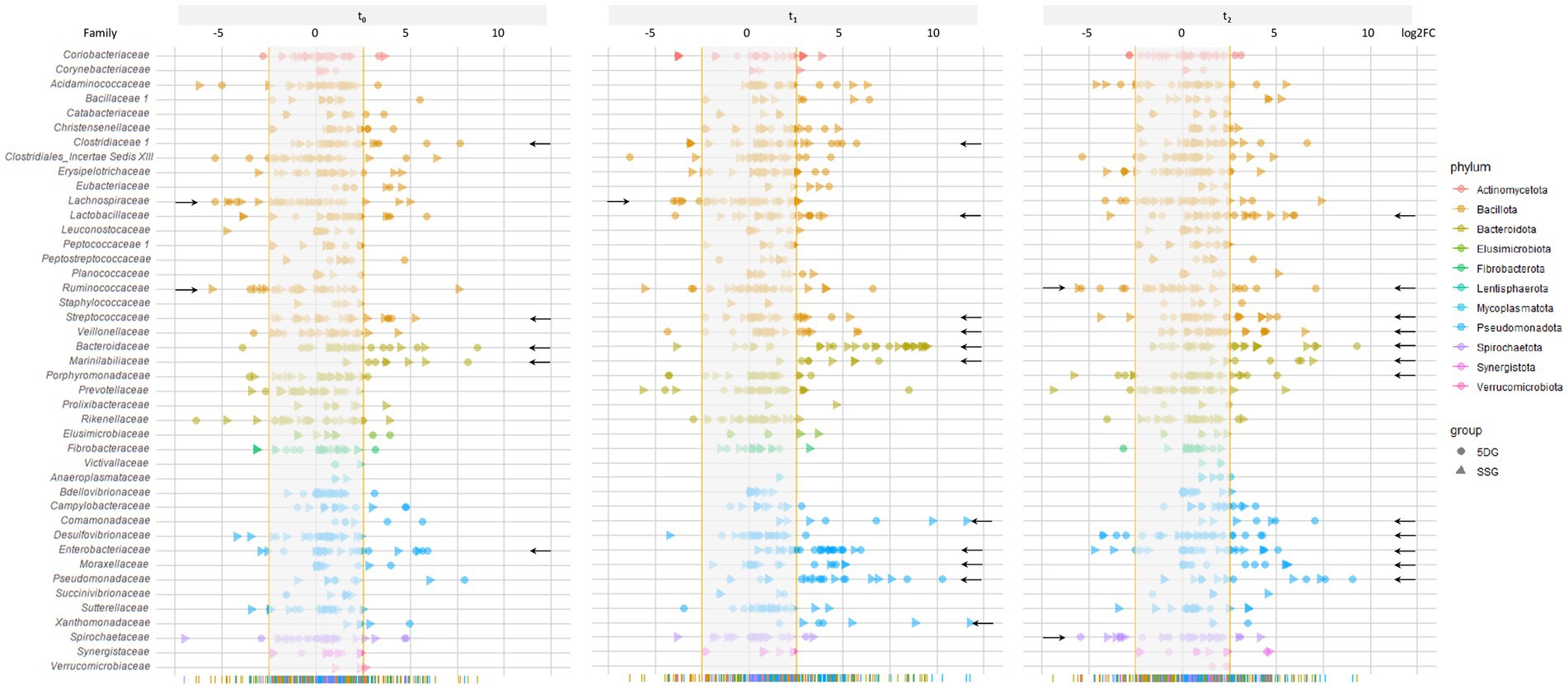
Figure 4. Changes of the fecal microbiome structure and composition during the study period. Illustration of log2FC abundance changes for bacterial families in each sample of both study groups with respect to a baseline calculated from the control group samples at t0. To acknowledge individual variation and to enhance the chances of overall trend detection, an interval comprising the majority of samples (85% at t0, log2FC of 2.5) was calculated, illustrated as the blurred area per timepoint. Individual values that were zero (i.e., no abundance difference compared to the control) were not shown for better clarification. Data points belonging to samples from the SSG are marked by a triangle and those from the 5DG by a dot, while deviation from the baseline within at least five samples are indicated by a black arrow.
To identify the most deviations with high potential relevancy, further in-depth explorative analysis was restricted to bacterial families associated with at least five samples (SSG and 5DG) that clustered beyond the 85% interval (Figure 4, black arrows indicate bacterial families fulfilling the restriction criteria).
Directly at hospital admission and before any antibiotics were administered (t0), six samples showed log2FC between 2.7 and 5.2 for Streptococcaceae, seven samples for Bacteroidaceae (log2FC between 2.9 and 8.6), seven samples for Marinilabiliaceae (log2FC between +2.8 and + 8.1), and six samples for Enterobacteriaceae (log2FC between +2.8 and + 5.9). Overall, lower values were noted for Lachnospiraceae (six samples, log2FC between −3.1 and − 5.4) and Ruminicoccaceae (five samples, log2FC between −2.7 and − 3.5; Figure 4, t0; indicated by black arrows).
At t1, increasing log2FC fulfilling the above mentioned criteria were noticed for Clostridiaceae, Lactobacillaceae, Streptococcaceae, Veillonellaceae, Marinilabiliaceae, and Xanthomonadaceae, but most prominently for Bacteriodaceae (22 samples, log2FC between +3.7 and + 9.5), Comamonadaceae (five samples, log2FC between +3.2 and + 11.6), Enterobacteriaceae (17 samples, log2FC between +2.7 and + 5.9), Moraxellaceae (six samples, log2FC between +3.5 and + 5.1), and Pseudomonadaceae (15 samples, log2FC between +2.8 and + 10.3; Figure 4). Lachnospiraceae, on the other hand, yielded log2FC between −2.7 and − 4.0 in five samples.
At t2, deviation from the baseline interval (log2FC ≥ ±2.5) defined for bacterial families were recognized for Bacteriodaceae (11 samples, log2FC between +2.8 and + 9.3), Enterobacteriaceae (eight samples, log2FC between +2.6 and + 5.1), and Lactobacillaceae (nine samples, log2FC between +2.8 and + 5.9) followed by Streptococcaceae, Veillonellaceae, Marinilabiliaceae, Comamonadaceae, Moraxellaceae, and Pseudomonadaceae (Figure 4). Overall, a shift back to the baseline was directly recognizable for most of the displayed bacterial families.
Taken together, OTU abundances of many bacterial families deviated considerably from those associated with samples of the CG (t0). In addition, overall deviation increased at t1 and decreased at t2, a result that is clearly in congruence with the SDI values and the trajectory pattern displayed by PCA (Figures 3A–D). Of note, clear differences between the study groups were not noticed in Figure 4. These results confirmed that although a common temporal trajectory pattern was recognizable, deviation of individuals contributed largely to the overall variances.
At t1, deviations of Bacteroidaceae and Enterobacteriaceae stood out considering the baseline defined by the CG samples (Figure 4). Although relative abundances of OTUs within an individual sample depended on each other, we noticed stunning trajectories of OTUs assigned to Bacteroides and Escherichia, the predominating genera of the above-mentioned families within our sample set (Figure 5).
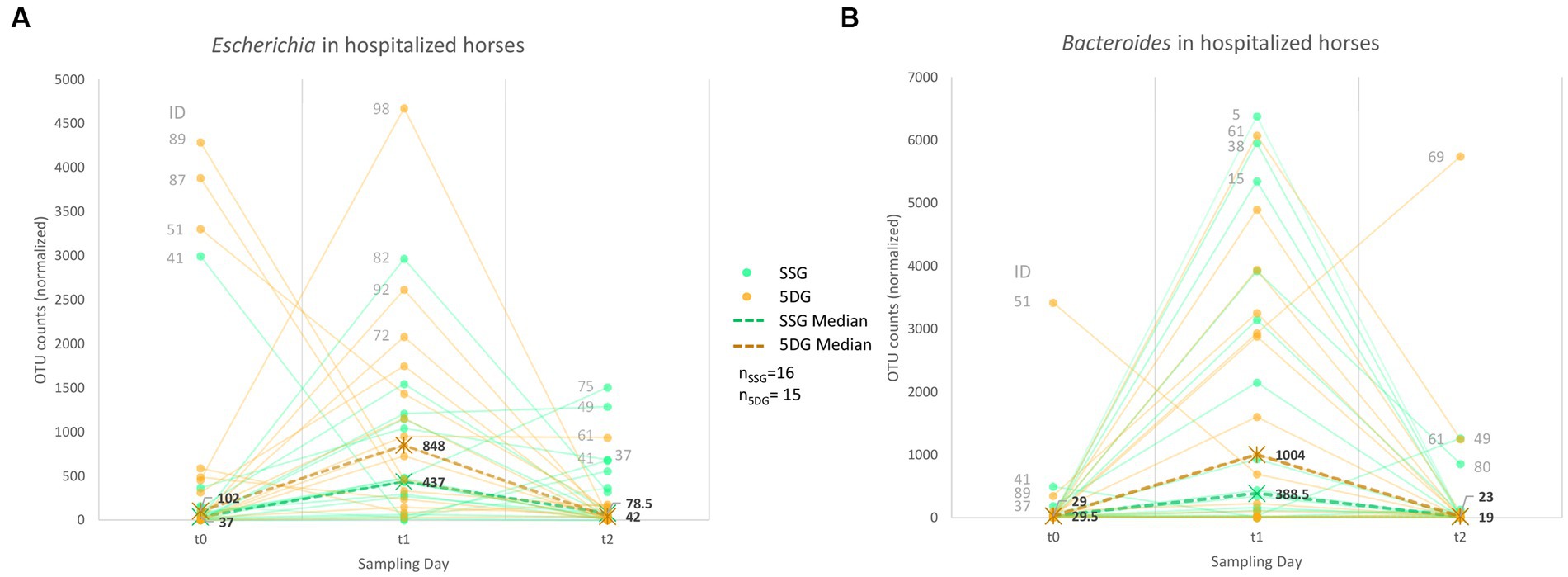
Figure 5. Abundance trajectories of OTUs assigned to the genera Escherichia (A) and Bacteroides (B). Charts showing a trend line based on normalized OTU counts per sample/study group and timepoint. A dashed line indicates the median per group and sampling day.
Apart from four horses (89, 87, 51, and 41), most study participants showed limited OTU counts (medianSSG = 37; median5DG = 102) classified as Escherichia (genus level) at t0, while the overall counts increased at t1 (Figure 5A), with a median OTU counts of 437 (SSG) and 848 (5DG) in samples of both groups, respectively (SSG vs. 5DG, t0: p = 0.14, t1: p = 0.086, t2: p = 0.95). This difference was significant between the SSG and 5DG from t1 to t2 (differences in counts, SSG vs. 5DG, t0 to t1: p = 1, t1 to t2: p = 0.0019). Of note, both study groups showed a decline to almost similar counts assigned to Escherichia at t2 (medianSSG = 79 OTU counts; median5DG = 42 OTU counts).
Interestingly, the hospitalized horses demonstrated a highly similar trajectory pattern for OTU counts assigned to the genus Bacteroides. Besides a single exception (horse 51, 5DG), both study groups had comparable counts for Bacteroides (medianSSG = 29.5 OTU counts, median5DG = 29 OTU counts) at t0 (SSG vs. 5DG, t0: p = 0.67, t1: p = 0.45, t2: p = 0.69). Changes in differences were not significant across time points (differences in counts, SSG vs. 5DG, t0 to t1: p = 0.89, t1 to t2: p = 0.92).
At t1, the OTU counts for Bacteroides doubled in abundance within the 5DG compared to the SSG (medianSSG = 389, median5DG = 1,004 OTU, p = 0.45). However, the Bacteroides count level decreased once more at the last day of sampling (t2) in both groups (medianSSG = 19; median5DG = 23, p = 0.69).
In summary, the abundance counts of both Escherichia and Bacteroides seemed to be associated with a more pronounced increase among samples representing the 5DG, i.e., horses that received the long-term PAP, compared to horses belonging to the SSG at t1. Correlation analysis revealed no obvious relationship between the two genera in the sample set investigated.
In the present study, we explored temporal alterations of the gut microbiota composition and the extent of its perturbation among hospitalized horses subjected to colic surgery that received either a short-term (SSG) or a 5-day course (5DG) of P/G PAP. Of note, comparison of fecal microbiota profiles of horses belonging to different cohorts or even between distinct individuals requires immense caution, since the intestinal microbiome is easily affected by external factors, including—but not limited to—exercise (Górniak et al., 2021), transport, fasting (Schoster et al., 2016) and diet (Al Jassim, 2006; Mshelia et al., 2018). In addition, Antwis et al. (2018) recently examined how spatial and social interactions affected the gut microbiome composition of semi-wild Welsh Ponies, revealing that up to 52.6% of the observed variation is attributable to individual variation.
At first, we determined the predominating microbial phyla in the sample set, revealing Bacteroidota (38%), Bacillota (33%) and Verrucomicrobiota (11%) as the top three main phyla at hospital admission (t0) and in the CG (Figure 2). This result is in line with previous reports defining the Bacteroidota and Bacillota as the major phyla of the core bacterial community in equines (Morrison et al., 2018; Edwards et al., 2020).
As summarized previously, most of the current clinical studies regarding the effects of antibiotics on the gut microbiome have been cross-sectional, while interventional or longitudinal approaches and comparisons to treatment-naive but diseased control groups are often missing (Zimmermann et al., 2021). As a result, it is difficult to differentiate between disease-mediated, drug-related hospitalization effects (Zimmermann et al., 2021), an aspect that clearly is a limitation with respect to the discussion of putative drug-related effects in the current study. Although the isolated and exact impact of the P/G regimen on the gut microbiota of horses participating in this study remains unknown due to the fact that our study was not an animal trial but a real-world scenario, recent research on the effect of parental administration of P/G on the developing infant gut microbiome clearly revealed an impact on Shannon diversity and overall gut microbiota composition (Reyman et al., 2022). However, at hospital admission and before antibiotic treatment and hospitalization (t0), the mean alpha-diversity measured by SDIs revealed strong signals of an already compromised bacterial biodiversity in samples representing the SSG and the 5DG compared with those of the CG (Figure 3D), although only the latter difference was found to be significant. A loss of species diversity seems to be among the prominent characteristic of a disturbed gut microbiota (Ramirez et al., 2020). At hospital admission, a decrease in bacterial richness and diversity accompanied with a greater inter-individual variability was reported for horses admitted due to colic compared with horses presented for elective surgical procedures or even healthy horses (Stewart et al., 2018; Park et al., 2021). A similar trend was detected in the current study, demonstrating a considerable range of variation between microbiota profiles and microbiome perturbations among horses suffering from colic compared to horses free of abdominal pain or other enteral disorders.
A previous study by Costa et al. (2015) examined the effects of intramuscular administration of procaine penicillin and ceftiofur sodium, and oral trimethoprim sulfadiazine on the fecal microbiome of healthy horses. Although the administration route and duration, the cohort under investigation and the combination of antibiotics (P/G) differed in the present study, general similarities should be mentioned. The strongest perturbation of the microbiome was recognized directly after the final course of antibiotics, including a strong effect on the microbial community membership (Costa et al., 2015).
The most prominent peak of microbiome disturbance is characterized by significant SDI decreases at t1 (i.e., 3 days after surgery/hospitalization; Figure 1). The considerable inter-individual distances between the microbiota profiles of 5DG horses at t0 have diminished immensely at t1, indicating a similar and consistent effect of the long-term PAP regimen on the gut microbiota composition that is specific for the long-term P/G PAP.
This observation is in line with the results of a recent study investigating the effects of commonly used antibiotics on the gut microbiome of healthy human volunteers before and after treatment, where the authors revealed drug-specific trajectories through the PCA space over time (Anthony et al., 2022). In strict contrast to the PCA of the 5DG in the present study, data points of the SSG showed increased inter-individual differences through the PCA space at t1, indicating the lack of a common selective factor. Therefore, other more distinguishing factors associated with the distinct individual such as appetite, stress and/or pain, environmental bacteria or even the onset of GIT function and/or microbiome recovery (reviewed in Kauter et al., 2019) might be mirrored here.
At t2, both study groups showed clear signs pointing toward the onset of microbiome recovery with respect to a rise of both SDImean, with most participants seeming to have already regained the baseline condition, since the difference between t0 and t2 lacked statistical differences (Figure 3). Interestingly, in a study of germ-free mice, the recovery of the gut microbiome after antibiotic treatment strongly depended on diet, community context and environmental reservoirs (Ng et al., 2019). The authors demonstrated that a reduction of environmental reservoirs impaired the process of microbiota recovery (Ng et al., 2019). This fact not only emphasizes the overall importance of the actual environmental bacteria in the immediate vicinity of hospitalized horses during microbiome recovery, but it also highlights the susceptibility of the equine gut microbiota to spatio-temporal local spreads of hospital-associated pathogens, explicitly including ESBL-EC, leading to worrisome carriage rates, as previously reported (Kauter et al., 2019).
Since studies on equine gut microbiomes are currently limited (Salem et al., 2019; Ang et al., 2022), it seems reasonable to assume that some general effects occur across different mammalian species: Exposure of the gut microbiota to antibiotics or their active metabolites reduces its overall diversity, which is either temporarily or permanently conserved after end of antibiotic treatment (Costa et al., 2015; Ramirez et al., 2020; Arnold et al., 2021). Moreover, the effect might not be limited to a reduced microbial diversity, since a recent study on healthy human volunteers showed a worrisome increase of virulence- and resistance associated-factors immediately after antibiotic treatment (Palleja et al., 2018). Further studies on the equine gut metagenome (Ang et al., 2022) after exposure to different antibiotics are required to gain more insights with respect to spatial trends of virulence- and resistance gene occurrences and abundances.
At first, we analyzed the significant log2FC of the SSG and the 5DG. Samples of horses belonging to both study groups were associated with a significant abundance increase for Bacteroidaceae, Enterobacteriaceae, and Pseudomonadaceae from t0 to t1. A previous study that examined changes in the equine fecal microbiome during hospitalization because of colic reported a similar increase for Bacteroides (Stewart et al., 2021). The increased abundances noticed for Enterobacteriaceae at t1 in both groups might enhance the risk of developing SSI for the equine patients, especially since multidrug-resistant Enterobacterales were often reported within horse clinics (Apostolakos et al., 2017; Walther et al., 2018; Shnaiderman-Torban et al., 2020), respectively in cases of SSI of horses subjected to laparotomy (Isgren et al., 2017; Dziubinski et al., 2020). Furthermore, significant changes were recognized regarding Pseudomonadaceae, a bacterial family that has been described as indicator of intestinal microbiome alterations in the human gut since an abundance increase seems to be associated with various gastrointestinal diseases (Alam et al., 2020; Chamorro et al., 2021).
Secondly, we explored the probable influence of the acute disease and other factors on the fecal microbiota. For this, we evaluated the abundance of different bacteria in fecal samples of both study groups compared to the control group at t0, revealing considerable deviation of OTUs belonging to 15 different bacterial families (Figure 4). Overall, a common temporal trajectory pattern was recognizable regarding Bacteroidaceae, Enterobacteriaceae, and Pseudomonadota, with a strikingly increased proportion in almost all t1 samples and decreasing, but still above-baseline level, OTU counts at t2. In addition, we observed a reduced frequency of Lachnospiraceae for the horses diagnosed with colic, compared to the horses belonging to the CG, which is in line with recent findings (Stewart et al., 2019). Moreover, a reduction of Lachnospiraceae was previously described for horses suffering from enteral disorders (Costa et al., 2012; Weese et al., 2015).
Although differences between the study groups were limited, we observed interesting differences with respect to the effect of the distinct P/G PAPs on the microbiota. Horses that received the 5-day course of PAP showed the highest loss of inter-individual diversity (Figure 3B, t1) which is clearly accompanied by the most prominent expansion of Escherichia (Figure 5). Further studies including metagenomic data are needed to reveal the particular dependences here.
Since penicillin and gentamicin were administered parenterally (Stöckle et al., 2021), the effect on the gut microbiome in general was expected to be lower than in cases when oral antibiotics where administered (Zhang et al., 2013). In the present study, however, the combined influences of (intestinal) illness, hospitalization, surgery and administration of P/G PAP showed a converged relative abundance trajectory of the genera Escherichia and Bacteroides over time, as demonstrated in the results (Figure 5). A recent review on the effects of distinct antibiotic classes on the GIT microbiome in humans highlighted an increase of Enterobacteriaceae and Bacteroidaceae after treatment with β-lactams in general (Patangia et al., 2022), which seems strikingly in line with the results presented here.
The obligatory anaerobic genus Bacteroides exclusively growing in the GIT of mammals is a major research focus of gut microbiology (reviewed in Wexler and Goodman, 2017). Since intrinsic resistance is reported for Bacteroides spp. [Pumbwe et al., 2006; Clinical and Laboratory Standards Institute (CLSI), 2020] at least an impact of a treatment-associated selective advantage can be assumed when considering the relative abundance increase among most of the fecal samples of both study group participants at t1 (Figure 5), in particular for the 5DG.
Although many Bacteroides species play a crucial role in degrading polysaccharides of a plant-based diet (Pereira et al., 2021; Cheng et al., 2022), the specific importance and role of distinct Bacteroides species in hindgut fermentation has not been investigated yet. More research on the subject including metabolomic profiles gained from metagenome sequencing projects is clearly required to shed more light on this subject.
Apart from outliers (four horses, Figure 5), most of the horses participating in the present study revealed a low relative abundance of Escherichia at hospital admission. This observation is in line with the previously reported increase of ESBL-EC carriage rates among patients of both study groups over time, i.e., from hospital admission to discharge (Supplementary Table 1), that indicated local spread of, for instance, distinct ESBL-EC clonal lineages, as reported previously (Kauter et al., 2021). We further speculate that during P/G PAP, ESBL-EC had a selective advantage in the GIT of horses and therefore proliferate, resulting not only in increased carriage rates (Kauter et al., 2021), but also in unavoidable environmental contamination via feces-contaminated litter. Since the immediate environment is among the main sources of GIT-associated bacteria during microbiome recovery (Ng et al., 2019), environmental sources seem to play an overwhelming role with respect to ESBL-EC carriage rates. Since the specific needs and surroundings of hospitalized horses (bedding, boxes, and floor surfaces, reviewed in Gehlen et al., 2020) differ immensely from those of small animal or even human patients, the environment of equine clinics seems to play an important role in ESBL-EC accumulation and spread, including strains known for their pathogenic potential in humans and animals, i.e., isolates belonging to sequence type (ST)10 and ST410 (Kauter et al., 2021).
Taken together, the spread of E. coli, especially ESBL-EC in horse clinics, seems to be promoted by (i) the selective advantage of these bacteria toward β-lactam antibiotics and (ii) the fact that the fecal microbiota structure is re-modeled by other factors occurring during the course of hospitalization, such as ingestion by food, contact to environmental sources or transmission via healthcare workers.
The colic patients originated from different stables and unknown feeding situations prior to hospitalization. Once hospitalized, they were provided a stable diet, comparable to horses of the control group. Further differences existed between the control and treated hospitalized groups because horses were kept in different environments (barn vs. clinic).
In the present study, we investigated the influence of two different PAP regimens (SSG vs. 5DG) in horses diagnosed with colic syndrome that were subjected to surgery with regard to changes of their gut microbiota composition. Colic surgery and PAP drive the equine gut microbiome toward dysbiosis and reduced biodiversity that is accompanied by a 10-fold increase of samples positive for ESBL-producing Enterobacterales (Kauter et al., 2021) and an abundance increase for Enterobacteriaceae. Further studies are needed to reveal the most important local sources of the resistant bacteria (i.e., environment, food, and contacts) and factors promoting the inclusion of ESBL-producing Enterobacterales in the equine gut microbiota.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, BioProject: PRJNA906950.
Ethical approval was not required for the studies involving animals in accordance with the local legislation and institutional requirements, as the comparison of perioperative antibiotic regimens does not require approval. Written informed consent was obtained from the owners for the participation of their animals in this study.
BW, AL-B, LW, and HG designed the project. AK, SS, HG, and BW conceived and designed the experiments. AK, DK, SS, JB, and AL-B performed laboratory analysis. CB and SF sequenced the samples. AK, SW, AL-B, BW, and TS analyzed the data. AK, SW, and BW wrote the first draft. AM, NS, RK, and SG helped to draft the manuscript and contributed to the discussion. All authors contributed to the article and approved the submitted version.
This work was funded by the German Federal Ministry of Education and Research (BMBF) for #1Health-PREVENT (grants 01KI2009A, 01KI2009D, and 01KI2009F) within the German Research Network of Zoonotic Diseases. The funding bodies did not influence data interpretation or writing the manuscript. This work was also supported by the DFG Research Infrastructure NGS_CC (project 407495230) as part of the Next Generation Sequencing Competence Network (project 423957469). Sequencing was carried out at the Competence Center for Genomic Analysis (Kiel).
We thank our colleagues from the Advanced Light and Electron Microscopy (ZBS 4) department of the Robert Koch Institute for their individual contribution and support. This manuscript was previously published as a preprint at: https://www.biorxiv.org/content/10.1101/2023.05.24.542119v1 (Kauter et al., 2023).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1228845/full#supplementary-material
5DG, 5-day course of peri-operative antibiotic prophylaxis group; Bp, Base pair; CG, Control group; EC, Escherichia coli; ESBL, Extended-spectrum β-Lactamase; G, Gentamicin; GIT, Gastrointestinal tract; i.e., id est; Log2FC, log2 fold change; MDR, Multi-drug resistance; OTU, Operational taxonomic units; P, Penicillin; PAP, Perioperative antibiotic prophylaxis; PCA, Principal component analysis; SDI, Shannon diversity index; SSG, Single-shot of peri-operative antibiotic prophylaxis group.
Al Jassim, R. A. M. (2006). Supplementary feeding of horses with processed sorghum grains and oats. Anim. Feed Sci. Technol. 125, 33–44. doi: 10.1016/j.anifeedsci.2005.05.019
Alam, M. T., Amos, G. C. A., Murphy, A. R. J., Murch, S., Wellington, E. M. H., and Arasaradnam, R. P. (2020). Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 12:1. doi: 10.1186/s13099-019-0341-6
Ang, L., Vinderola, G., Endo, A., Kantanen, J., Jingfeng, C., Binetti, A., et al. (2022). Gut microbiome characteristics in feral and domesticated horses from different geographic locations. Commun. Biol. 5:172. doi: 10.1038/s42003-022-03116-2
Anthony, W. E., Wang, B., Sukhum, K. V., D’souza, A. W., Hink, T., Cass, C., et al. (2022). Acute and persistent effects of commonly used antibiotics on the gut microbiome and resistome in healthy adults. Cell Rep. 39:110649. doi: 10.1016/j.celrep.2022.110649
Antwis, R. E., Lea, J. M. D., Unwin, B., and Shultz, S. (2018). Gut microbiome composition is associated with spatial structuring and social interactions in semi-feral Welsh Mountain ponies. Microbiome 6:207. doi: 10.1186/s40168-018-0593-2
Apostolakos, I., Franz, E., Van Hoek, A., Florijn, A., Veenman, C., Sloet-Van Oldruitenborgh-Oosterbaan, M. M., et al. (2017). Occurrence and molecular characteristics of ESBL/amp C-producing Escherichia coli in faecal samples from horses in an equine clinic. J. Antimicrob. Chemother. 72, 1915–1921. doi: 10.1093/jac/dkx072
Archer, D. C. (2019). Colic surgery: keeping it affordable for horse owners. Vet. Rec. 185, 505–507. doi: 10.1136/vr.l6062
Arnold, C. E., Pilla, R., Chaffin, M. K., Leatherwood, J. L., Wickersham, T. A., Callaway, T. R., et al. (2021). The effects of signalment, diet, geographic location, season, and colitis associated with antimicrobial use or Salmonella infection on the fecal microbiome of horses. J. Vet. Intern. Med. 35, 2437–2448. doi: 10.1111/jvim.16206
Aronesty, E. (2013). Comparison of sequencing utility programs. Open Bioinform. J. 7, 1–8. doi: 10.2174/1875036201307010001
Baverud, V., Gustafsson, A., Franklin, A., Aspan, A., and Gunnarsson, A. (2003). Clostridium difficile: prevalence in horses and environment, and antimicrobial susceptibility. Equine Vet. J. 35, 465–471. doi: 10.2746/042516403775600505
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Ceriotti, S., Westerfeld, R., Bonilla, A. G., and Pang, D. S. J. (2021). Use of clinical audits to evaluate timing of preoperative antimicrobials in equine surgery at a veterinary teaching hospital. Front. Vet. Sci. 8:630111. doi: 10.3389/fvets.2021.630111
Chamorro, N., Montero, D. A., Gallardo, P., Farfán, M., Contreras, M., De La Fuente, M., et al. (2021). Landscapes and bacterial signatures of mucosa-associated intestinal microbiota in Chilean and Spanish patients with inflammatory bowel disease. Microb. Cell 8, 223–238. doi: 10.15698/mic2021.09.760
Cheng, J., Hu, J., Geng, F., and Nie, S. (2022). Bacteroides utilization for dietary polysaccharides and their beneficial effects on gut health. Food Sci. Human Wellness 11, 1101–1110. doi: 10.1016/j.fshw.2022.04.002
Clinical and Laboratory Standards Institute (CLSI) (2020). Performance Standards for Antimicrobial Susceptibility Testing CLSI Supplement M100. Wayne, PA: Clinical and Laboratory Standards Institute
Cole, J. R., Wang, Q., Fish, J. A., Chai, B., Mcgarrell, D. M., Sun, Y., et al. (2014). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, D633–D642. doi: 10.1093/nar/gkt1244
Costa, M. C., Arroyo, L. G., Allen-Vercoe, E., Stämpfli, H. R., Kim, P. T., Sturgeon, A., et al. (2012). Comparison of the fecal microbiota of healthy horses and horses with colitis by high throughput sequencing of the V3-V5 region of the 16S rRNA gene. PLoS One 7:e41484. doi: 10.1371/journal.pone.0041484
Costa, M. C., Stampfli, H. R., Allen-Vercoe, E., and Weese, J. S. (2016). Development of the faecal microbiota in foals. Equine Vet. J. 48, 681–688. doi: 10.1111/evj.12532
Costa, M. C., Stampfli, H. R., Arroyo, L. G., Allen-Vercoe, E., Gomes, R. G., and Weese, J. S. (2015). Changes in the equine fecal microbiota associated with the use of systemic antimicrobial drugs. BMC Vet. Res. 11:19. doi: 10.1186/s12917-015-0335-7
Dallap Schaer, B. L., Linton, J. K., and Aceto, H. (2012). Antimicrobial use in horses undergoing colic surgery. J. Vet. Intern. Med. 26, 1449–1456. doi: 10.1111/j.1939-1676.2012.01024.x
Davis, J. L., Salmon, J. H., and Papich, M. G. (2006). Pharmacokinetics and tissue distribution of doxycycline after oral administration of single and multiple doses in horses. Am. J. Vet. Res. 67, 310–316. doi: 10.2460/ajvr.67.2.310
Desantis, T. Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E. L., Keller, K., et al. (2006). Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072. doi: 10.1128/AEM.03006-05
Durward-Akhurst, S. A., Mair, T. S., Boston, R., and Dunkel, B. (2013). Comparison of two antimicrobial regimens on the prevalence of incisional infections after colic surgery. Vet. Rec. 172:287. doi: 10.1136/vr.101186
Dziubinski, N., Mählmann, K., Lübke-Becker, A., and Lischer, C. (2020). Retrospective identification of bacterial isolates from emergency laparotomy surgical site infections in horses. J. Equine Vet. 87:102927. doi: 10.1016/j.jevs.2020.102927
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Edwards, J. E., Shetty, S. A., Van Den Berg, P., Burden, F., Van Doorn, D. A., Pellikaan, W. F., et al. (2020). Multi-kingdom characterization of the core equine fecal microbiota based on multiple equine (sub)species. Anim. Microb. 2:6. doi: 10.1186/s42523-020-0023-1
Gehlen, H., Simon, C., Reinhold-Fritzen, B., Lübke-Becker, A., Kauter, A., Walther, B., et al. (2020). Basis-Hygienemaßnahmen für den Pferdetierarzt in Praxis und Klinik. Berl. Munch. Tierarztl. Wochenschr. 133 doi: 10.2376/1439-0299-2020-3
Górniak, W., Cholewińska, P., Szeligowska, N., Wołoszyńska, M., Soroko, M., and Czyż, K. (2021). Effect of intense exercise on the level of Bacteroidetes and Firmicutes Phyla in the digestive system of thoroughbred racehorses. Animals 11:290. doi: 10.3390/ani11020290
Gu, Z., Eils, R., and Schlesner, M. (2016). Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 32, 2847–2849. doi: 10.1093/bioinformatics/btw313
Isgren, C. M., Salem, S. E., Archer, D. C., Worsman, F. C., and Townsend, N. B. (2017). Risk factors for surgical site infection following laparotomy: effect of season and perioperative variables and reporting of bacterial isolates in 287 horses. Equine Vet. J. 49, 39–44. doi: 10.1111/evj.12564
Kauter, A., Brombach, J., Luebke-Becker, A., Kannapin, D., Bang, C., Franzenburg, S., et al. (2023). Antibiotic prophylaxis and hospitalization of horses subjected to median laparotomy: gut microbiota trajectories and abundance increase of Escherichia. bioRxiv [Preprint]. doi: 10.1101/2023.05.24.542119v1
Kauter, A., Epping, L., Ghazisaeedi, F., Lübke-Becker, A., Wolf, S. A., Kannapin, D., et al. (2021). Frequency, local dynamics, and genomic characteristics of ESBL-producing Escherichia coli isolated from specimens of hospitalized horses. Front. Microbiol. 12:671676. doi: 10.3389/fmicb.2021.671676
Kauter, A., Epping, L., Semmler, T., Antao, E.-M., Kannapin, D., Stoeckle, S. D., et al. (2019). The gut microbiome of horses: current research on equine enteral microbiota and future perspectives. Anim. Microb. 1:14. doi: 10.1186/s42523-019-0013-3
Kelly, B. J., Gross, R., Bittinger, K., Sherrill-Mix, S., Lewis, J. D., Collman, R. G., et al. (2015). Power and sample-size estimation for microbiome studies using pairwise distances and PERMANOVA. Bioinformatics 31, 2461–2468. doi: 10.1093/bioinformatics/btv183
Kozich, J. J., Westcott, S. L., Baxter, N. T., Highlander, S. K., and Schloss, P. D. (2013). Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 79, 5112–5120. doi: 10.1128/AEM.01043-13
Lahti, L., Shetty, S., et al. (2017). Tools for microbiome analysis in R. Available at: https://microbiome.github.io/tutorials/ (Accessed September 20, 2022).
Liepman, R. S., Swink, J. M., Habing, G. G., Boyaka, P. N., Caddey, B., Costa, M., et al. (2022). Effects of intravenous antimicrobial drugs on the equine fecal microbiome. Animals 12:1013. doi: 10.3390/ani12081013
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Mach, N., Ruet, A., Clark, A., Bars-Cortina, D., Ramayo-Caldas, Y., Crisci, E., et al. (2020). Priming for welfare: gut microbiota is associated with equitation conditions and behavior in horse athletes. Sci. Rep. 10:8311. doi: 10.1038/s41598-020-65444-9
Magoč, T., and Salzberg, S. L. (2011). FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963. doi: 10.1093/bioinformatics/btr507
Mcdonald, D., Clemente, J. C., Kuczynski, J., Rideout, J. R., Stombaugh, J., Wendel, D., et al. (2012). The biological observation matrix (BIOM) format or: how I learned to stop worrying and love the ome-ome. GigaScience 1:7. doi: 10.1186/2047-217X-1-7
Mcmurdie, P. J., and Holmes, S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Morrison, P. K., Newbold, C. J., Jones, E., Worgan, H. J., Grove-White, D. H., Dugdale, A. H., et al. (2018). The equine gastrointestinal microbiome: impacts of age and obesity. Front. Microbiol. 9:3017. doi: 10.3389/fmicb.2018.03017
Mshelia, E. S., Adamu, L., Wakil, Y., Turaki, U. A., Gulani, I. A., and Musa, J. (2018). The association between gut microbiome, sex, age and body condition scores of horses in Maiduguri and its environs. Microb. Pathog. 118, 81–86. doi: 10.1016/j.micpath.2018.03.018
Ng, K. M., Aranda-Díaz, A., Tropini, C., Frankel, M. R., van Treuren, W., O’Loughlin, C. T., et al. (2019). Recovery of the gut microbiota after antibiotics depends on host diet, community context, and environmental reservoirs. Cell Host Microbe 26, 650–665.e4. doi: 10.1016/j.chom.2019.10.011
Oren, A., and Garrity, G. M. (2021). Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 71, 3491–3500. doi: 10.1099/ijsem.0.005056
Palleja, A., Mikkelsen, K. H., Forslund, S. K., Kashani, A., Allin, K. H., Nielsen, T., et al. (2018). Recovery of gut microbiota of healthy adults following antibiotic exposure. Nat. Microbiol. 3, 1255–1265. doi: 10.1038/s41564-018-0257-9
Park, T., Cheong, H., Yoon, J., Kim, A., Yun, Y., and Unno, T. (2021). Comparison of the fecal microbiota of horses with intestinal disease and their healthy counterparts. Vet. Sci. 8:113. doi: 10.3390/vetsci8060113
Patangia, D. V., Anthony Ryan, C., Dempsey, E., Paul Ross, R., and Stanton, C. (2022). Impact of antibiotics on the human microbiome and consequences for host health. MicrobiologyOpen 11:e1260. doi: 10.1002/mbo3.1260
Pereira, G. V., Abdel-Hamid, A. M., Dutta, S., D’alessandro-Gabazza, C. N., Wefers, D., Farris, J. A., et al. (2021). Degradation of complex arabinoxylans by human colonic Bacteroidetes. Nat. Commun. 12:459. doi: 10.1038/s41467-020-20737-5
Pumbwe, L., Ueda, O., Yoshimura, F., Chang, A., Smith, R. L., and Wexler, H. M. (2006). Bacteroides fragilis BmeABC efflux systems additively confer intrinsic antimicrobial resistance. J. Antimicrob. Chemother. 58, 37–46. doi: 10.1093/jac/dkl202
Ramirez, J., Guarner, F., Bustos Fernandez, L., Maruy, A., Sdepanian, V. L., and Cohen, H. (2020). Antibiotics as major disruptors of gut microbiota. Front. Cell. Infect. Microbiol. 10:572912. doi: 10.3389/fcimb.2020.572912
Reeder, J., Huang, M., Kaminker, J. S., and Paulson, J. N. (2021). MicrobiomeExplorer: an R package for the analysis and visualization of microbial communities. Bioinformatics 37, 1317–1318. doi: 10.1093/bioinformatics/btaa838
Reyman, M., Van Houten, M. A., Watson, R. L., Chu, M. L. J. N., Arp, K., De Waal, W. J., et al. (2022). Effects of early-life antibiotics on the developing infant gut microbiome and resistome: a randomized trial. Nat. Commun. 13:893. doi: 10.1038/s41467-022-28525-z
Salem, S. E., Maddox, T. W., Antczak, P., Ketley, J. M., Williams, N. J., and Archer, D. C. (2019). Acute changes in the colonic microbiota are associated with large intestinal forms of surgical colic. BMC Vet. Res. 15:468. doi: 10.1186/s12917-019-2205-1
Schoster, A., Mosing, M., Jalali, M., Staempfli, H. R., and Weese, J. S. (2016). Effects of transport, fasting and anaesthesia on the faecal microbiota of healthy adult horses. Equine Vet. J. 48, 595–602. doi: 10.1111/evj.12479
Schwartz, D. J., Langdon, A. E., and Dantas, G. (2020). Understanding the impact of antibiotic perturbation on the human microbiome. Genome Med. 12:82. doi: 10.1186/s13073-020-00782-x
Shen, W., Le, S., Li, Y., and Hu, F. (2016). Seq kit: a cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS One 11:e0163962. doi: 10.1371/journal.pone.0163962
Shnaiderman-Torban, A., Navon-Venezia, S., Dor, Z., Paitan, Y., Arielly, H., Ahmad, W. A., et al. (2020). Extended-Spectrum β-lactamase-producing Enterobacteriaceae shedding in farm horses versus hospitalized horses: prevalence and risk factors. Animals 10:282. doi: 10.3390/ani10020282
Southwood, L. L. (2014). Perioperative antimicrobials: should we be concerned about antimicrobial drug use in equine surgical patients? Equine Vet. J. 46, 267–269. doi: 10.1111/evj.12247
Stewart, H. L., Pitta, D., Indugu, N., Vecchiarelli, B., Engiles, J. B., and Southwood, L. L. (2018). Characterization of the fecal microbiota of healthy horses. Am. J. Vet. Res. 79, 811–819. doi: 10.2460/ajvr.79.8.811
Stewart, H. L., Pitta, D., Indugu, N., Vecchiarelli, B., Hennessy, M. L., Engiles, J. B., et al. (2021). Changes in the faecal bacterial microbiota during hospitalisation of horses with colic and the effect of different causes of colic. Equine Vet. J. 53, 1119–1131. doi: 10.1111/evj.13389
Stewart, H. L., Southwood, L. L., Indugu, N., Vecchiarelli, B., Engiles, J. B., and Pitta, D. (2019). Differences in the equine faecal microbiota between horses presenting to a tertiary referral hospital for colic compared with an elective surgical procedure. Equine Vet. J. 51, 336–342. doi: 10.1111/evj.13010
Stockle, S. D., Failing, K., Koene, M., and Fey, K. (2018). Postoperative complications in equine elective, clean orthopaedic surgery with/without antibiotic prophylaxis. Tierarztl. Prax. Ausg. G Grosst. Nutz. 46, 81–86. doi: 10.15653/TPG-170491
Stöckle, S. D., Kannapin, D. A., Kauter, A. M. L., Lübke-Becker, A., Walther, B., Merle, R., et al. (2021). A pilot randomised clinical trial comparing a short-term perioperative prophylaxis regimen to a long-term standard protocol in equine colic surgery. Antibiotics 10:10. doi: 10.3390/antibiotics10050587
Teschner, D., Barton, A. K., Klaus, C., and Gehlen, H. (2015). Antibiotikaeinsatz bei operierten Kolikpferden in Deutschland. Pferdeheilkunde 31, 235–240. doi: 10.21836/PEM20150305
Traub-Dargatz, J. L., Kopral, C. A., Seitzinger, A. H., Garber, L. P., Forde, K., and White, N. A. (2001). Estimate of the national incidence of and operation-level risk factors for colic among horses in the United States, spring 1998 to spring 1999. J. Am. Vet. Med. Assoc. 219, 67–71. doi: 10.2460/javma.2001.219.67
Walther, B., Klein, K.-S., Barton, A.-K., Semmler, T., Huber, C., Wolf, S. A., et al. (2018). Extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli and Acinetobacter baumannii among horses entering a veterinary teaching hospital: the contemporary "Trojan horse". PLoS One 13:e0191873. doi: 10.1371/journal.pone.0191873
Weese, J. S., Holcombe, S. J., Embertson, R. M., Kurtz, K. A., Roessner, H. A., Jalali, M., et al. (2015). Changes in the faecal microbiota of mares precede the development of post partum colic. Equine Vet. J. 47, 641–649. doi: 10.1111/evj.12361
Wexler, A. G., and Goodman, A. L. (2017). An insider's perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2:17026. doi: 10.1038/nmicrobiol.2017.26
Wickham, H., Chang, W., Henry, L., Pedersen, T.L., Takahashi, K., Wilke, C., et al. (2016). ggplot2: create elegant data visualisations using the grammar of graphics. R package version 2. Available at: https://ggplot2.tidyverse.org/reference/ggplot2-package.html (Accessed June 2022).
Wormstrand, B. H., Ihler, C. F., Diesen, R., and Krontveit, R. I. (2014). Surgical treatment of equine colic—a retrospective study of 297 surgeries in Norway 2005-2011. Acta Vet. Scand. 56:38. doi: 10.1186/1751-0147-56-38
Zhang, L., Huang, Y., Zhou, Y., Buckley, T., and Wang, H. H. (2013). Antibiotic administration routes significantly influence the levels of antibiotic resistance in gut microbiota. Antimicrob. Agents Chemother. 57, 3659–3666. doi: 10.1128/AAC.00670-13
Keywords: horse, microbiome, gastrointestinal tract, microbiota, 16S rRNA gene sequencing, hospitalization, colic, Escherichia
Citation: Kauter A, Brombach J, Lübke-Becker A, Kannapin D, Bang C, Franzenburg S, Stoeckle SD, Mellmann A, Scherff N, Köck R, Guenther S, Wieler LH, Gehlen H, Semmler T, Wolf SA and Walther B (2023) Antibiotic prophylaxis and hospitalization of horses subjected to median laparotomy: gut microbiota trajectories and abundance increase of Escherichia. Front. Microbiol. 14:1228845. doi: 10.3389/fmicb.2023.1228845
Edited by:
Rebeca Martín, INRAE Center Jouy-en-Josas, FranceReviewed by:
Sharanne Lee Raidal, Charles Sturt University, AustraliaCopyright © 2023 Kauter, Brombach, Lübke-Becker, Kannapin, Bang, Franzenburg, Stoeckle, Mellmann, Scherff, Köck, Guenther, Wieler, Gehlen, Semmler, Wolf and Walther. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Birgit Walther, YmlyZ2l0LndhbHRoZXJAdWJhLmRl
†These authors share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.