 Marianne Quéméneur1*‡
Marianne Quéméneur1*‡ Nan Mei2‡
Nan Mei2‡ Christophe Monnin3
Christophe Monnin3 Anne Postec1Sophie Guasco1Julie Jeanpert4†Pierre Maurizot4
Anne Postec1Sophie Guasco1Julie Jeanpert4†Pierre Maurizot4 Bernard Pelletier5
Bernard Pelletier5 Gaël Erauso1
Gaël Erauso1- 1Aix Marseille Univ, Université de Toulon, CNRS, IRD, MIO, Marseille, France
- 2School of Chemistry, Chemical Engineering, and Life Science, Wuhan University of Technology, Wuhan, China
- 3Géosciences Environnement Toulouse, UMR 5563 (CNRS/UPS/IRD/CNES), Toulouse, France
- 4Direction de l’Industrie, des Mines et de l’Energie, Nouméa, New Caledonia
- 5Centre IRD de Nouméa, Nouméa, New Caledonia
The southeastern part of New Caledonia main island (Grande Terre) is the location of a large ophiolitic formation that hosts several hyperalkaline springs discharging high pH (∼11) and warm (<40°C) fluids enriched in methane (CH4) and hydrogen (H2). These waters are produced by the serpentinization of the ultrabasic rock formations. Molecular surveys had previously revealed the prokaryotic diversity of some of these New Caledonian springs, especially from the submarine chimneys of Prony Bay hydrothermal field. Here we investigate the microbial community of hyperalkaline waters from on-land springs and their relationships with elevated concentrations of dissolved H2 (21.1–721.3 μmol/L) and CH4 (153.0–376.6 μmol/L). 16S rRNA gene analyses (metabarcoding and qPCR) provided evidence of abundant and diverse prokaryotic communities inhabiting hyperalkaline fluids at all the collected springs. The abundance of prokaryotes was positively correlated to the H2/CH4 ratio. Prokaryotes consisted mainly of bacteria that use H2 as an energy source, such as microaerophilic Hydrogenophaga/Serpentinimonas (detected in all sources on land) or anaerobic sulfate-reducing Desulfonatronum, which were exclusively found in the most reducing (Eh ref H2 ∼ -700 mV) and the most H2-enriched waters discharging at the intertidal spring of the Bain des Japonais. The relative abundance of a specific group of uncultured Methanosarcinales that thrive in serpentinization-driven ecosystems emitting H2, considered potential H2-consuming methanogens, was positively correlated with CH4 concentrations, and negatively correlated to the relative abundance of methylotrophic Gammaproteobacteria. Firmicutes were also numerous in hyperalkaline waters, and their relative abundance (e.g., Gracilibacter or Dethiobacter) was proportional to the dissolved H2 concentrations, but their role in the H2 budget remains to be assessed. The prokaryotic communities thriving in New Caledonia hyperalkaline waters are similar to those found in other serpentinite-hosted high-pH waters worldwide, such as Lost City (North Atlantic) and The Cedars (California).
1. Introduction
Serpentinization is a natural process of mantle rock alteration that transforms olivine into serpentine, with the parallel formation of dihydrogen (H2) linked to water reduction and the oxidation of metals (principally iron) contained in the minerals (mainly olivine) (Sleep et al., 2004; Klein et al., 2013; Schrenk et al., 2013; Wang et al., 2014). This H2 can react with carbon-bearing species such as carbonic acid to form methane (CH4) as well as low-molecular-weight organic compounds such as acetate and formate (Schrenk et al., 2013; Wang et al., 2014; Konn et al., 2015). These organic compounds can be sources of energy (electron donors) and of carbon for microorganisms living in deep environments (McCollom and Seewald, 2013; Ménez, 2020). The environmental characteristics of serpentinizing environments are considered similar to those that have prevailed on the early Earth and on other rocky planets like Mars, thus providing a prebiotic chemistry that may have favored the emergence of life (Nealson et al., 2005; Russell et al., 2010). Thus, the study of these ecosystems allows to address fundamental questions such as the origin and limits of life on Earth, or more applied investigations such as the search of extremophiles (i.e., alkaliphiles) for biotechnological applications or the production of natural H2 (also called native H2) (Gaucher, 2020; Truche et al., 2020).
Several ecosystems found in such serpentinization-driven environments have been found in various areas around the world: (i) underwater, such as the shallow submarine Prony Bay hydrothermal field (PBHF) located at less than 50 meters below sea level (mbsl) in the southern lagoon of New Caledonia (South Pacific) (Monnin et al., 2014; Quéméneur et al., 2014; Postec et al., 2015), the deep submarine Lost City hydrothermal field (LCHF) near the Mid-Atlantic Ridge at 700–800 mbsl (Kelley et al., 2005), and the abyssal Old City hydrothermal field at 3,100 mbsl along the southwest Indian ridge (Lecoeuvre et al., 2021), or (ii) on-land, such as The Cedars spring system (CA, USA) (Suzuki et al., 2013), the Coast Range Ophiolite (CA, USA) (Twing et al., 2017), the Samail ophiolite (Oman) (Rempfert et al., 2017) and the Voltri ophiolitic springs (Italy) (Quéméneur et al., 2015). Phylogenetically and metabolically diverse microbial communities live inside the chimneys and concretions built by the venting of anoxic, high-pH fluids (Schrenk et al., 2013). Reactions involving H2, CH4, and sulfur compounds act as the energy source for these microbial communities, indicating that serpentinization-related fluids can sustain chemosynthesis rather than photosynthesis (McCollom and Seewald, 2013).
The southern part of the New Caledonia main island (Grande Terre) is covered by a large allochthonous sheet of oceanic lithosphere (ophiolite) thrusted over continental basement at the late Eocene (Avias, 1967; Cluzel et al., 2001; Pirard et al., 2013). There, a number of high-pH springs are located either on land, in the intertidal zone of the Prony Bay or at shallow depths in this bay (Cox et al., 1982; Monnin et al., 2014, 2021; Deville and Prinzhofer, 2016; Maurizot et al., 2020). In the Prony Bay, high-pH (up to 11.2) and warm (up to 42°C) fluids enriched in H2 (12–30% vol of dry gas) and CH4 (6–14% vol of dry gas) are currently venting into the lagoon (Monnin et al., 2014; Vacquand et al., 2018). Their mixing with seawater leads to the formation of brucite-carbonate chimneys and pinnacles, reaching heights up to tens of meters, as the 38-m high Aiguille de Prony (Launay and Fontes, 1985; Quéméneur et al., 2014). Previous molecular surveys have revealed the prokaryotic diversity of the coastal submarine PBHF, with diverse aerobic and anaerobic bacteria potentially involved in H2 consumption (e.g., Hydrogenophaga and Serpentinimonas) or H2 production (e.g., Clostridiales) (Quéméneur et al., 2014; Mei et al., 2016b). In addition, a low diversity of uncultured Methanosarcinales, potentially linked to CH4 production or oxidation, was observed in PBHF chimneys (Quéméneur et al., 2014; Postec et al., 2015; Frouin et al., 2018), and was also found in other serpentinization-associated submarine and terrestrial sites such as The Cedars or Lost City (Kelley et al., 2005; Suzuki et al., 2013; Quéméneur et al., 2015). However, the microbial communities from on-land springs of the New Caledonia ophiolite have been only studied from the geothermal spring of La Crouen (Quéméneur et al., 2021), where bacteria potentially involved in sulfur cycle (e.g., Candidatus Desulfobacillus, Thiofaba, Thiovirga) and H2 oxidation (e.g., Hydrogenophaga) dominated with Ca. Gracilibacteria in the waters depleted in H2. The concentrations of H2 and CH4 in gases and waters emitted at the high-pH springs of New Caledonia are quite variable (Monnin et al., 2014, 2021; Deville and Prinzhofer, 2016) and likely to play a major role on the abundance and diversity of microorganisms.
This study used 16S rRNA gene analyses (metabarcoding and qPCR) to investigate the abundance and composition of prokaryotic communities inhabiting the high-pH fluids discharging at several on-land and intertidal springs of New Caledonia. We evaluated the relationship between the main taxa and the physicochemical characteristics of the high-pH fluids, including their contents in H2 and CH4. We also compared the dominant microbial members with those of other serpentinite-hosted hyperalkaline springs worldwide to uncover specific taxonomic bioindicators of this type of ecosystem and proxies of natural H2 and CH4 emissions.
2. Materials and methods
2.1. Study site
The studied high-pH thermal springs are all located in the southeastern part of the main island (Figure 1). The water samples were collected in November–December 2014 at five different sites : (i) a shallow (10-cm deep) pool of “La Coulée” (CL) spring, close to the town of Nouméa, (ii) the source of “Rivière des Pirogues” (PG), located halfway between the cities of Nouméa and Yaté, (iii) a natural 1-m deep pool of “Montagne des Sources” (MDS), located in a natural reserve close to the city of Nouméa, (iv) a shallow (10-cm) natural pool (RKB) and the spring captured in a cemented pool 1-m deep (RKH) of “Rivière des Kaoris,” located in the Prony Bay, (v) the water venting at the “Bain des Japonais” (BJ), located in the intertidal zone of the Prony Bay (Figure 1). The spring locations have been previously described by Maurizot et al. (2020) and Monnin et al. (2021). All on-land springs are associated with carbonate deposits.
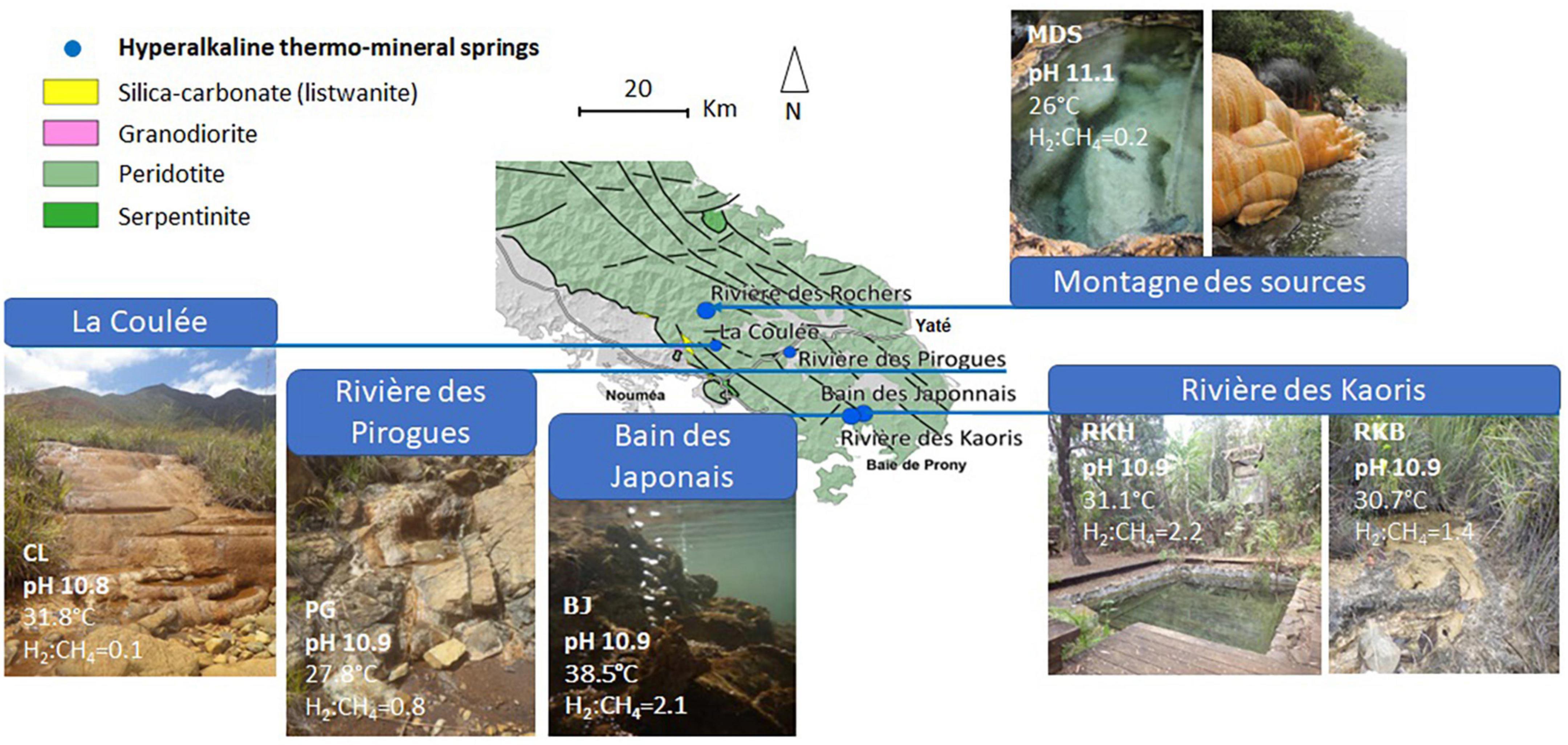
Figure 1. Location and photos of the high-pH springs of New Caledonia. CL corresponds to the highest pool of “La Coulée,” MDS is a pool of the site “Montagne des Sources,” PG indicates the source of the site “Rivière des Pirogues,” RKB and RKH correspond to a natural and an artificial pool of the area “Rivière des Kaoris,” respectively, and, BJ is a small intertidal vent of the site “Bain des Japonais” in the Prony Bay. The map is adapted from Maurizot et al. (2020). The H2:CH4 ratio is expressed in concentrations mol/mol.
2.2. Sample collection
Water samples were collected in cleaned 4-L plastic bottles using sterile and pre-rinsed syringes and stored in a portable icebox until arrival at the laboratory (about 2–3 h after sampling). Duplicate samples of two liters of water were filtered through 0.2 μm pore-size Isopore polycarbonate membrane filters 47-mm (Millipore). The filters were kept at -80°C before DNA extraction. Water samples dedicated to the chemical analyses were collected in duplicate in 120-ml glass bottles sealed with butyl-rubber stoppers. At springs where gas bubbles form, free gas samples were collected in 10-ml glass vials using the water displacement technique and sealed with butyl-rubber stoppers and aluminum caps in 10-ml glass vials these samples were kept at 4°C in the Nouméa (IRD) and Marseille (MIO) laboratories. The oxidation-reduction potential (Eh), dissolved oxygen (O2), pH, temperature, and conductivity were measured in situ using a WTW Multi 3420® Multimeter with adequate probes (Monnin et al., 2021).
2.3. Dissolved gas analysis
Dissolved gas analysis was performed using a headspace equilibration method adapted from Magen et al. (2014). Briefly, a headspace representing 10% of the vial volume was created in the collection bottle by water displacement with argon, then the bottle was manually shaken for 1 min and placed on a shaker for 1 h. The composition of the headspace gas was determined using a Shimadzu GC 8A gas chromatograph equipped with a thermal conductivity detector (GC/TCD) and a concentric column CTR1 (Alltech, USA), as described previously (Mei et al., 2014). Argon was used as carrier gas at a flow rate of 60 mL/min; the injector and detector temperatures were fixed at 150°C. The concentrations of the dissolved gases were calculated using Henry’s law (Sander, 2015).
2.4. DNA extraction
The filters were transferred to a sterile 2 mL tube containing a glass bead mixture (lysing matrix E from MP Biomedicals). The bacterial and archaeal cells of the filters were disrupted by a combination of mechanical (bead beating, according MP Biomedical recommendations) and chemical lyses by addition of 1 mL of sterile bacterial lysis buffer [100 mM NaCl, 100 mM Tris pH 8.0, 50 mM EDTA, 100 μL of 10 mg/mL lysozyme (Sigma-Aldrich, St. Louis, MO, USA), 20 μL of 10 μg/mL DNase-free RNase solutions] and incubation at 37°C for 15 min. Then, 100 μL of 10% SDS, 100 μL of 10% lauryl-sarkosyl and 50 μL of proteinase K (20 mg/mL were added to the mixture and incubated at 55°C for 1 h). DNA was extracted from the lysate with 1 volume of phenol:chloroform (1:1) mixture and then with 1 volume of chloroform. Total DNA was precipitated from the aqueous phase by adding 0.7 volume of isopropanol followed by centrifugation. The DNA pellet was washed in 75% ethanol and was again collected by centrifugation. The air-dried pellet was dissolved in 30 μL of TE buffer. The DNA concentrations were measured using a Qubit® fluorometer (Invitrogen).
2.5. Quantitative real-time PCR (qPCR)
The abundances of bacteria and archaea were determined by qPCR using, respectively, the primers set 341F/518R (Muyzer et al., 1993) and 344F/519R (Ovreås et al., 1997; Casamayor et al., 2002). The primer set used to quantify methanogens was ME3MF/ME2r’ targeting mcrA genes (Hales et al., 1996; Nunoura et al., 2008). The primer set used to quantify sulfate-reducers was DSRp2060f/DSR4R targeting dsrB genes (Wagner et al., 1998; Geets et al., 2006). The sequences of primers are given in Supplementary Table 1.
Each qPCR mixture (20 μL) contained 1X SsoAdvanced SYBR Green Supermix (Bio-Rad), 250 nM of each primer, 1 μl of DNA template (10-fold dilution series of standard PCR product or environmental DNA sample) or distilled water (negative control). All qPCR assays were performed in triplicate on a Bio-Rad CFX-96 real-time system (Bio-Rad). The qPCR cycling conditions were: 95°C for 2 min, followed by 40 cycles of a 2-step PCR protocol with a 15 sec denaturation phase at 95°C and a 15 sec annealing/elongation phase at 55°C. Fluorescence was measured at the end of each cycle. Following PCR, melt curves were generated between 65 and 95°C in 0.5°C increments, to verify PCR specificity.
qPCR standard curves were created from serial dilution of DNA standards of known concentration. For bacterial 16S rRNA and dsrB gene qPCR, standard DNA fragments were amplified from Desulfovibrio vulgarisT DSM 644 using the primer sets 27F/907R (Lane, 1991) and DSR1F/DSR4R (Wagner et al., 1998), respectively. For archaeal 16S rRNA and mcrA gene qPCR, standard DNA fragments were amplified from Methanosarcina barkeriT DSM 800 using the primer sets 109F/958R (Webster et al., 2006) and MLF/MLR (Luton et al., 2002), respectively. Triplicate PCR products were pooled and purified with the Nucleospin and PCR Clean-up kit (Macherey-Nagel), according to the manufacturer instructions. Purified PCR products were quantified using the BioSpec-nano Spectrophotometer (Shimadzu) and used as DNA standards. Copy number of DNA standards was calculated as described by Oldham and Duncan (2012). For each gene, the standard curve of CT (threshold cycle) versus the gene copy numbers was generated by using a 10-fold dilution series from 108 to 101 copies per ng of DNA. For all standard curves, the coefficients of determination (R2) were higher than 99.0%. The precision of the assay was measured by calculating the variation in Ct values across the three replicates. The abundance of targeted genes was reported as copy numbers per L of water.
2.6. 16S rDNA metabarcoding analyses of microbial communities
Bacterial and archaeal 16S rRNA gene V4 hypervariable regions were amplified by PCR using the 515F/806R universal primer set (Caporaso et al., 2011), with a barcode on the forward primer, as previously described by Dowd et al. (2008), and were sequenced by the MiSeq Illumina platform of the Molecular Research Laboratory (TX, USA). Sequence data were processed using the MR DNA analysis pipeline (MR DNA, Shallowater, TX, USA). In summary, sequences were joined (overlapping pairs) and grouped by samples following the barcodes before removing them. Short sequences (<150 bp) and sequences with ambiguous base calls were removed. Remaining sequences were denoised, operational taxonomic units (OTUs) were generated, and chimeras were checked using UCHIME and removed (Edgar et al., 2011). OTUs were clustered at 97% of similarity with USEARCH (Edgar, 2010) followed by removal of singleton sequences. Finally, OTUs were taxonomically classified using BLASTn against NCBI non-redundant (NR) reference database, and the top hit was taken as a taxonomic classification. The 16S rRNA gene sequences of the dominant OTUs have been deposited in the Genbank database under the accession numbers OQ551354-OQ551410. Raw sequence data were submitted to the NCBI SRA under BioProject PRJNA974011, BioSamples SAMN35158175–SAMN35158185.
2.7. Statistical analyses
All statistical analyses were performed using XLSTAT 2020.5.1 (Microsoft Excel add-in program; Addinsoft, Paris, France). The alpha diversity was calculated using the Shannon (1948) and Simpson (1949) indices from OTU abundance matrix. The beta diversity was based on Bray–Curtis’s dissimilarities and a principal coordinate analysis (PCoA) (from phyla/classes) or a dendrogram (from dominant OTUs) was generated to group samples into clusters. Spearman correlations and principal component analyses (PCA) were used to evaluate the relationship between the relative abundance of microbial taxa (classes/phyla and dominant OTUs) and the physico-chemical parameters of water. P-values < 0.05 are statistically significant. The abundance of the dominant OTUs in the water samples was also visualized by heatmap.
3. Results
3.1. Physico-chemical parameters and dissolved gases of water samples
The water samples collected from the six spring sites had high pH values ranging from 10.8 (CLW) to 11.1 (MDSW), and their temperatures varied from 26.0°C (MDSW) to 38.5 °C (BJW) (Table 1), as previously reported (Monnin et al., 2021). The lowest Eh value (-697 mV, ref. H2) was measured at the intertidal site of the “Bain des Japonais” (BJW). At this spring, very active gas bubbling is observed. The highest concentration of dissolved H2 (721.3 μmol/L) is also measured from the BJ waters (Table 1). The lowest dissolved O2 level (0.1 mg/L) was measured at the site of “Rivière des Kaoris” (RKB), also a site with intense gas bubbling (Monnin et al., 2014, 2021). There the concentration of dissolved CH4 (376.6 μmol/L) is the highest of all the studied springs, while the highest ratio of H2:CH4 was observed at the site RKH (Figure 2 and Table 1).
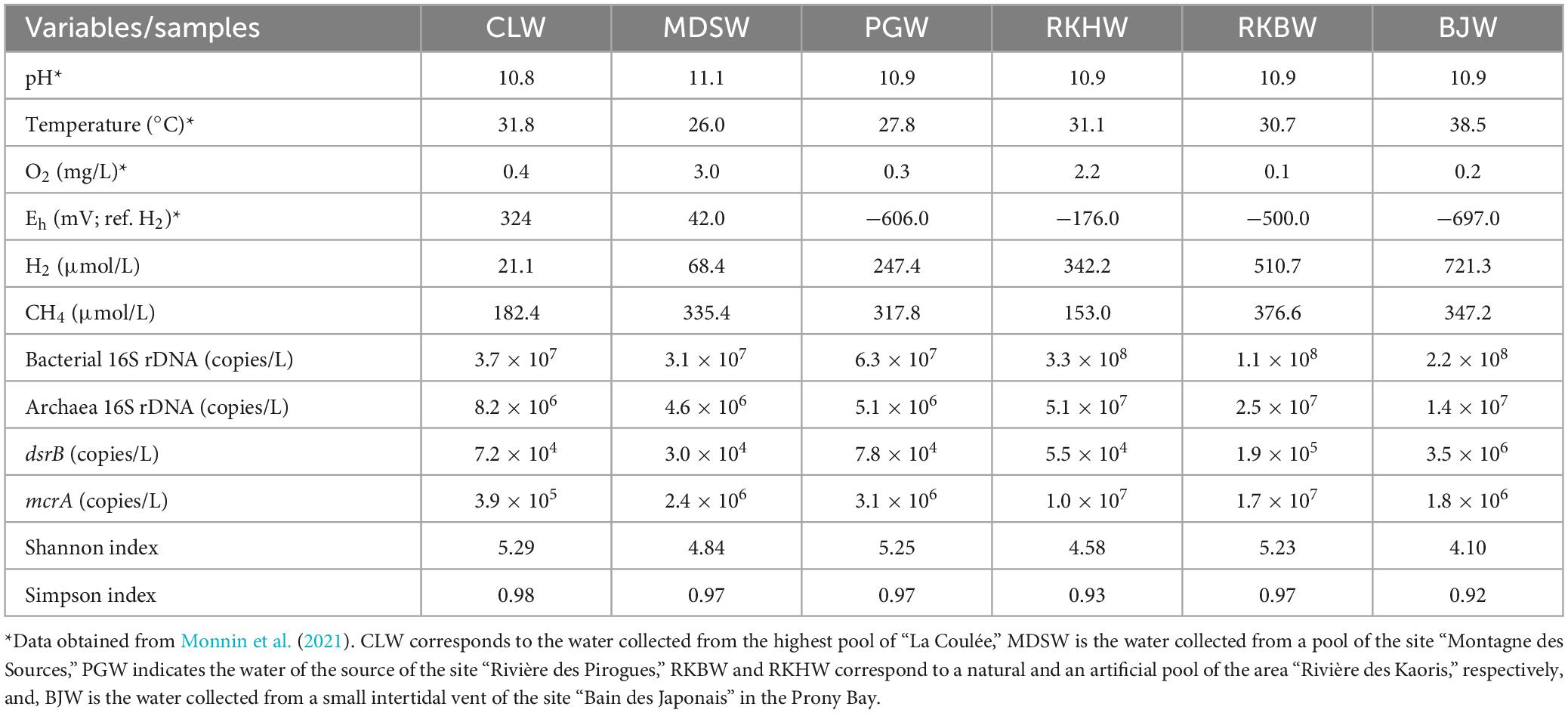
Table 1. In situ physicochemical parameters, diversity indices and gene abundances in water samples collected in the high-pH geothermal springs of New Caledonia.
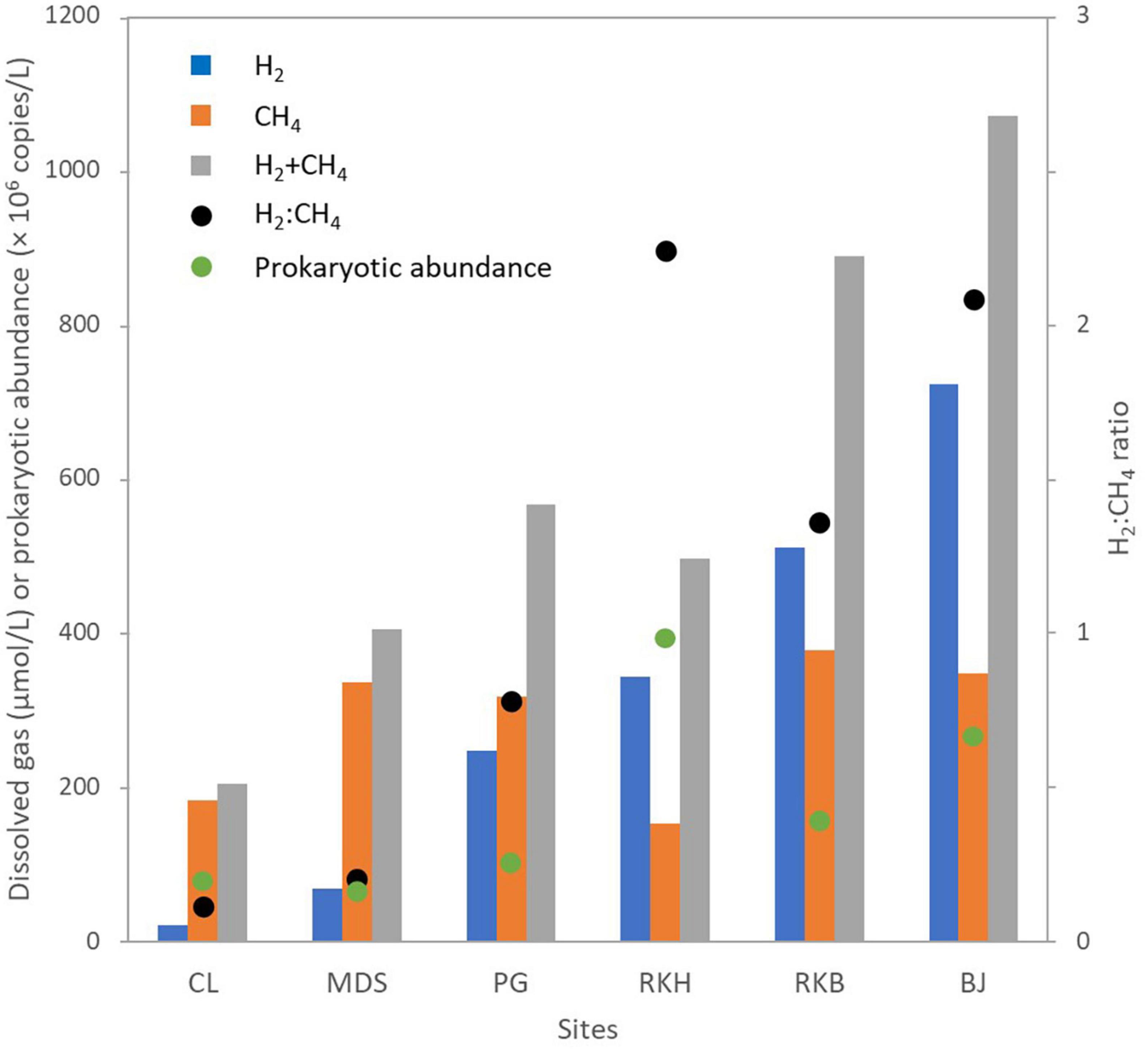
Figure 2. Dissolved gas (hydrogen and methane) and abundance of prokaryotes in water samples collected from the high-pH springs of New Caledonia. CL corresponds to the highest pool of “La Coulée,” MDS is a pool of the “Montagne des Sources,” PG indicates the source of the “Rivière des Pirogues,” RKB and RKH correspond to a natural and an artificial pool of the “Rivière des Kaoris,” respectively, and, BJ is a small intertidal vent of the “Bain des Japonais.” Sum and ratio of hydrogen and methane concentrations (H2:CH4 ratio) and sum of the abundance of archaea and Bacteria (total quantity of prokaryotes obtained by real-time PCR) were indicated.
3.2. Abundance and diversity indices of prokaryotic communities
Real-time qPCR assays and 16S rDNA metabarcoding analyses were performed on the water samples collected from the six high-pH springs. The qPCR experiments revealed that the bacterial 16S rRNA gene abundances ranged between 3.1 × 107 (MDSW) and 3.3 × 108 (RKHW) copies/L. The archaeal 16S rRNA gene abundances varied from 4.6 × 106 (MDSW) to 5.1 × 107 (RKHW) copies/L (Table 1). The abundance of bacterial 16S rRNA genes was more than one order of magnitude higher than archaeal 16S rRNA genes, as previously observed in PBHF chimneys (Quéméneur et al., 2014; Postec et al., 2015). The highest abundances of prokaryotes were measured in the waters of the PBHF sites, with maximum values in the RKH pool (displaying the highest H2 concentrations) (Figure 2). Prokaryotic abundances increased with increasing H2:CH4 ratio (Figure 2 and Supplementary Figure 1). The mcrA genes (markers of methanoarchaea) were detected in all samples but were more abundant in the waters of the Rivière des Kaoris (> 107 copies/L of RKHW and RKBW). The dsrB genes (markers of sulfate reducers) were also detected in all samples. They were more abundant in the BJ waters (BJW), where sulfate concentrations range between 10 and 100 μM/L (Monnin et al., 2014).
The Simpson diversity indices (1-D) ranged between 0.92 and 0.98, while Shannon indices (H) varied from 4.1 to 5.2 (Table 1). Based on both indices, the lowest microbial diversity was observed in the most reducing water sample BJW (displaying the highest H2 concentrations), while the highest diversity was found in CLW (displaying the highest Eh value and the lowest H2 concentrations).
3.3. Global prokaryotic community composition
Variation in microbial community composition was observed in the spring waters with high-pH, which were different from each other (Figure 3). This was also observed in a principal coordinates analysis (PCoA), where the replicated samples from the same spring were closely grouped (with the exception of MDS water samples), but were situated far from each other on the PCoA plot (Supplementary Figure 1).
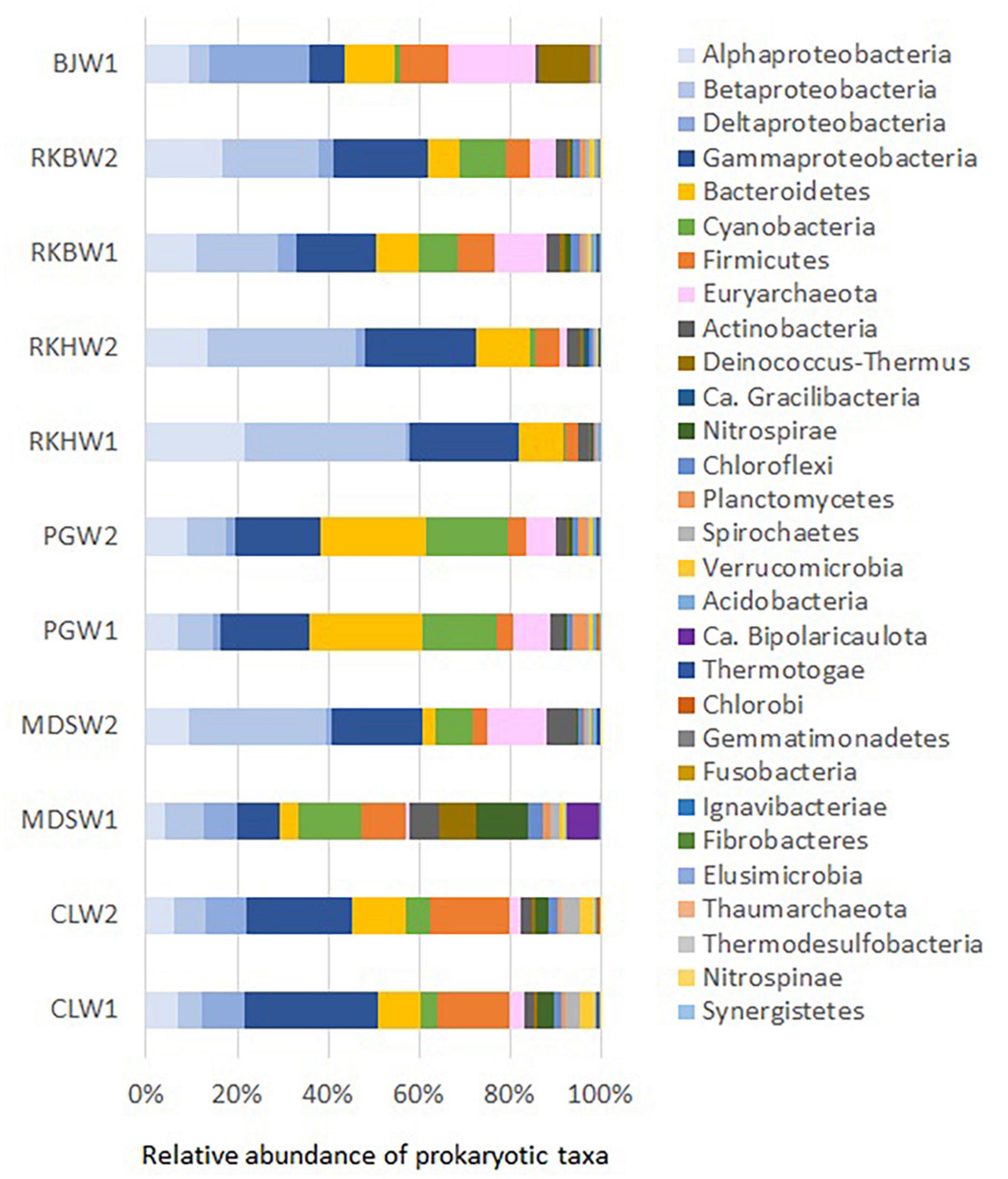
Figure 3. Prokaryotic phyla and proteobacterial classes (> 1% on average) in water samples collected from the high-pH springs of New Caledonia. CLW corresponds to the water of “La Coulée,” MDSW is the water of “Montagne des Sources,” PGW indicates the water of “Rivière des Pirogues,” RKBW and RKHW correspond to the waters of “Rivière des Kaoris” and BJW is the water of “Bain des Japonais.” Duplicated samples were indicated by numbers 1 or 2 (except for sample BJW).
Twenty-six different phyla were identified by 16S rRNA metabarcoding analyses of the water samples collected from the six high-pH springs (Figure 3). However, only half of these phyla (n = 12) were considered dominant (representing each more than 1% of prokaryotic sequences and together more than 97% of prokaryotic sequences). Proteobacteria was predominant in all samples (37.5 ± 10.0%), followed by Bacteroidetes (11.0 ± 6.9%), Firmicutes (7.9 ± 5.1%), Cyanobacteria (7.8 ± 6.1%) and Euryarchaeota (6.5 ± 5.9%; Figure 3). These five major phyla (each > 5% on average) accounted for 85% of all prokaryotic sequences. The other dominant phyla ranged between 1 and 5% of prokaryotic phyla (in average) and included Actinobacteria (3.1 ± 1.8%), Deinococcus-Thermus (2.2 ± 1.7%), Nitrospirae (2.0 ± 1.3%), Chloroflexi (1.3 ± 0.9%), Planctomycetes (1.1 ± 0.9%), Spirochaetes (1.3 ± 1.2%), Verrucomicrobia (1.1 ± 1.1%), followed by minor phyla (0.1-1%): Acidobacteria, Bipolaricaulota (formerly known as Acetothermia or OP1), Thermotogae and Gracilibacteria (formerly designated GN02/BD1-5).
The highest content of Euryarchaeota (mainly represented by Methanosarcinales) was observed in Bain des Japonais (BJ) waters (19.1%), which also displayed the highest content of Deltaproteobacteria (mainly represented by Desulfonatronum species). Both Deinococcus-Thermus (Meiothermus) and Firmicutes (Gracilibacter) were also detected in significant amounts in BJ waters (> 10% of prokaryotes) and in Montagne des Sources (MDS) waters (> 10%). Firmicutes were ubiquitous and abundant in all waters (> 1%) and reached a maximum in CL waters (16.7 ± 1.0%), where Dethiobacter species mainly represented them. The PG waters were primarily dominated by Bacteroidetes (24%) and Cyanobacteria (16%). The MDS waters displayed the highest occurrences of Bipolaricaulota.
3.4. Distribution and diversity of abundant prokaryotic OTUs
Marked variations in the prokaryotic community of high-pH waters are also illustrated on a heatmap showing the most abundant OTUs (each > 1% on average) (Figure 4). Altogether, these 57 dominant OTUs represented almost 2/3 of the total prokaryotic sequences (60% in average).
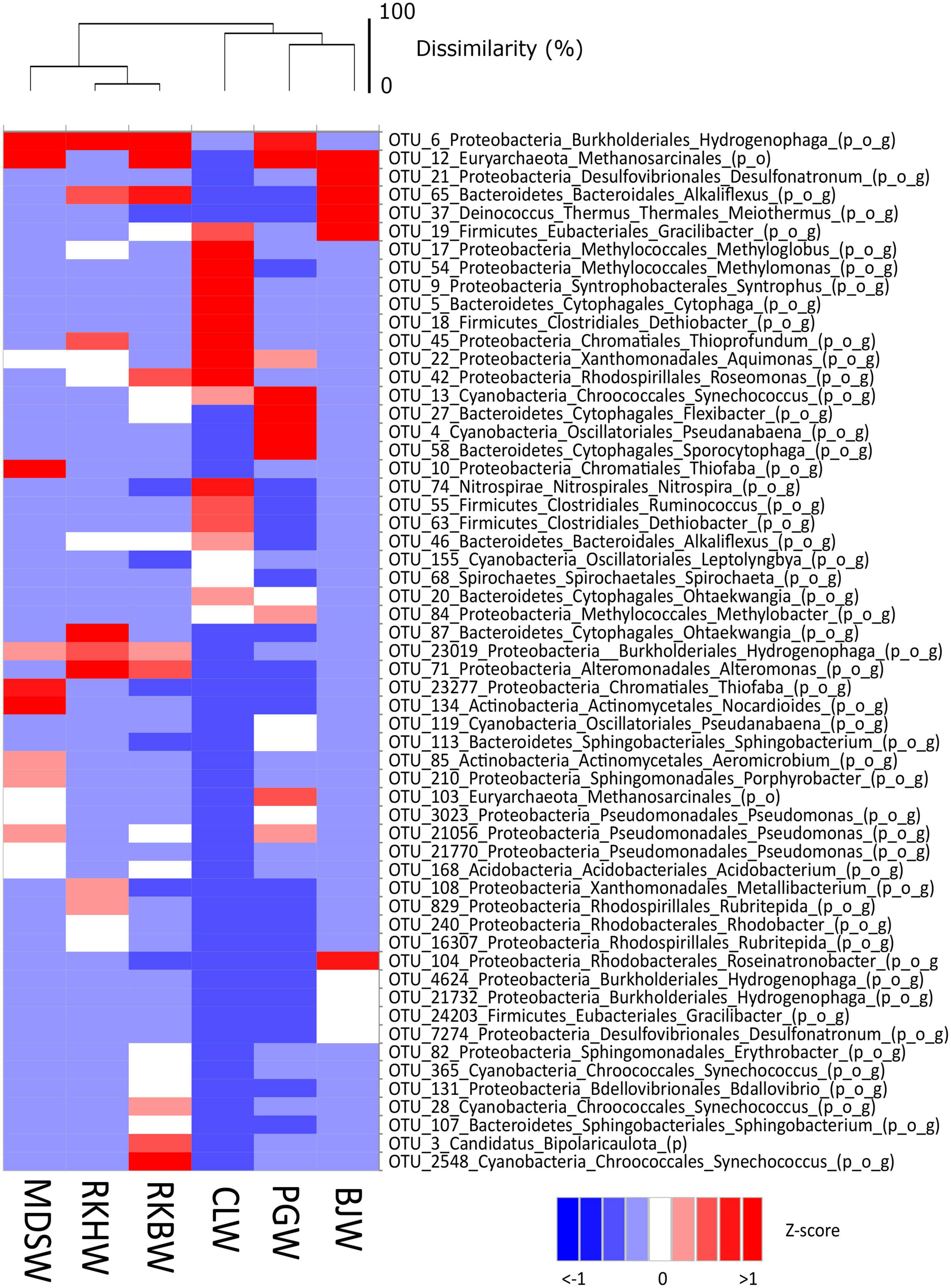
Figure 4. Heat map visualizing the Z-score distribution of the relative abundance of the dominant OTUs (> 1% on average) and their respective taxonomic affiliations in all water samples of the high-pH springs of New Caledonia. CLW corresponds to the water of “La Coulée,” MDSW is the water of “Montagne des Sources,” PGW indicates the water of “Rivière des Pirogues,” RKBW and RKHW correspond to the waters of “Rivière des Kaoris” and BJW is the water of “Bain des Japonais.” The average of replicates was calculated for each sampling site. The scale bar of the dendrogram represents the dissimilarity level (%) between microbial communities. The abbreviations of taxonomic ranks are p_ for phylum, o_ for order and g_ for genus.
In BJ waters (displaying the lowest Eh value and the highest H2 content), the prokaryotes were dominated by: (i) potential H2-oxidizing, sulfate-reducing Desulfonatronum OTUs (#21 and #7274, 19.5 and 1% of prokaryotes, respectively), closely related to Desulfonatronum cooperativum (96.7% identity; NR_043143), serpentinite-hosted PBHF clones (> 99% identity, KF886171; KT344938), and an uncultured deltaproteobacterium of deep groundwater (97.8%, LC055934), (ii) Methanosarcinales (OTU #12, 17.2% of prokaryotes) related to uncultured archaea from PBHF (KF886034 and KF886029; > 99.3% identity) and LCHF (SGYG644, SGYU755; 95.6% identity) classified as Lost City Methanosarcinales (LCMS); (iii) Meiothermus (OTU #37, 9.5%) closely related to Meiothermus hypogaeus (97.1% identity; NR_113226) and a PBHF clone (99.3% identity, KF886174), (iv) Bacteroidetes (OTU #65; 8.9%) related to uncultured bacteria from a deep subsurface gas storage aquifer (FJ168485, 90.2% identity) and deep groundwater from the Mizunami underground research laboratory (MIU) (LC055944, > 99% identity), (v) Firmicutes (OTUs #19 and #24203, 7.3 and 1.2%, respectively) affiliated with Gracilibacter thermotolerans (92–93%; NR_115692) and Firmicutes strains CE17 and CE8 (KX156793 and KX156784; > 95% identity) enriched from an in situ electrochemical experiment in a high-pH serpentinizing spring of The Cedars.
The RKH waters were dominated by: (i) H2-oxidizing Hydrogenophaga OTUs (#6 and #23019; 24.6 and 3.3% of prokaryotes) closely related to Hydrogenophaga aquatica (97.5% identity) and Serpentinimonas barnesii (NR_181590; 99.3 and 97.1% identity), (ii) Alteromonas OTUs (#71 and #7774; 4.8 and 1.2%), (iii) Bacteroidetes OTUs (#87 and #65, 4.6 and 2.6%) closely related to those of carbonate precipitates from the Voltri serpentinite-hosted hyperalkaline springs (99% identity; KP097469). The RKB waters, similarly to the RKH ones, were also dominated by the Hydrogenophaga OTUs #6 and #23019, but in smaller quantities (about half as much, 13.3 and 1.7% of prokaryotes). Both Alteromonas and Bacteroidetes OTUs were also less represented (∼2%), but Methanosarcinales OTUs #12 and #103 were more abundant in RKBW (6 and 0.5%), displaying higher CH4 content than in RKHW (∼0.5%). Bipolaricaulota OTU #3 related to Acetothermus autotrophicum (AP011801, 97% identity) accounted for 2.5% of RKBW prokaryotes. Cyanobacterial OTUs related to Synechococcus (#28, #365, and #2548) represented more than 5% of the prokaryotes.
In PG waters, the dominant OTUs were: (i) cyanobacterial Leptolyngbya (#4 and #13, 6.6 and 4.4% of prokaryotes) closely related to that of the mildly alkaline (pH 8.3–8.8) low-sulfur, low-carbonate Octopus hot spring (Yellowstone National Park, USA) (99% identity; KC236907, AY862014), (ii) Methanosarcinales (#12 and #103; 4.2 and 1.8%), (iii) Hydrogenophaga (#6; 2.7%), (iv) Methylobacter (#84; 1.3%).
In CL waters, the five dominant OTUs were affiliated with: (i) potential methylotrophic Methyloglobus (#17; 90% identity) and Methylomonas (#54), (ii) potential hydrocarbon-degrading Syntrophus gentianae (JQ346737; OTU #9; 95% identity), similar to uncultured deltaproteobacterium detected from deep groundwater (LC055948, > 98% identity), (iii) uncultured Firmicutes related to Dethiobacter alkaliphilus (OTUs #5, #18, #63; 94–96% identity), also detected from deep groundwater (LC055956) and from several serpentinite-hosted ecosystems (> 97% identity): PBHF (KJ149239, KF886127), Cabeço de Vide Aquifer (CVA) (AM777965), the hyperalkaline Allas Springs (Cyprus, 97.4% identity, JQ766804), the hyperalkaline spring GPS1 fed with deep groundwater at The Cedars (KC57503216S).
The MDS waters were dominated by six OTUs (representing more than 1/3 of the community) affiliated with (i) Hydrogenophaga/Serpentinimonas (#6 and #23019; 10.0 and 1.4% of prokaryotes), (ii) Methanosarcinales (#12 and #103; 10.0 and 1.1%), and (iii) sulfur-oxidizing Thiofaba (#10 and #23277, 8.2 and 2.4%).
3.5. Relationships between abundant taxa and environmental variables
Principal component analyses (PCA) was performed to identify the factors that affect the microbial community of New Caledonia high-pH spring waters (Figure 5). The first two principal components explained 63.6 and 76.1% of the data variability, for the phyla (Figure 5A) and major prokaryotic functional groups (Figure 5B), respectively. The first axis mostly separated the waters with the highest H2 contents (BJW, RKBW, and RKHW) from the others. The second axis separated the more oxygenated waters RKHW and MDSW from the others. Spearman’s rank correlation analyses examined the relationships between the microbial taxa, diversity indices and the environmental variables (Supplementary Tables 2–4).
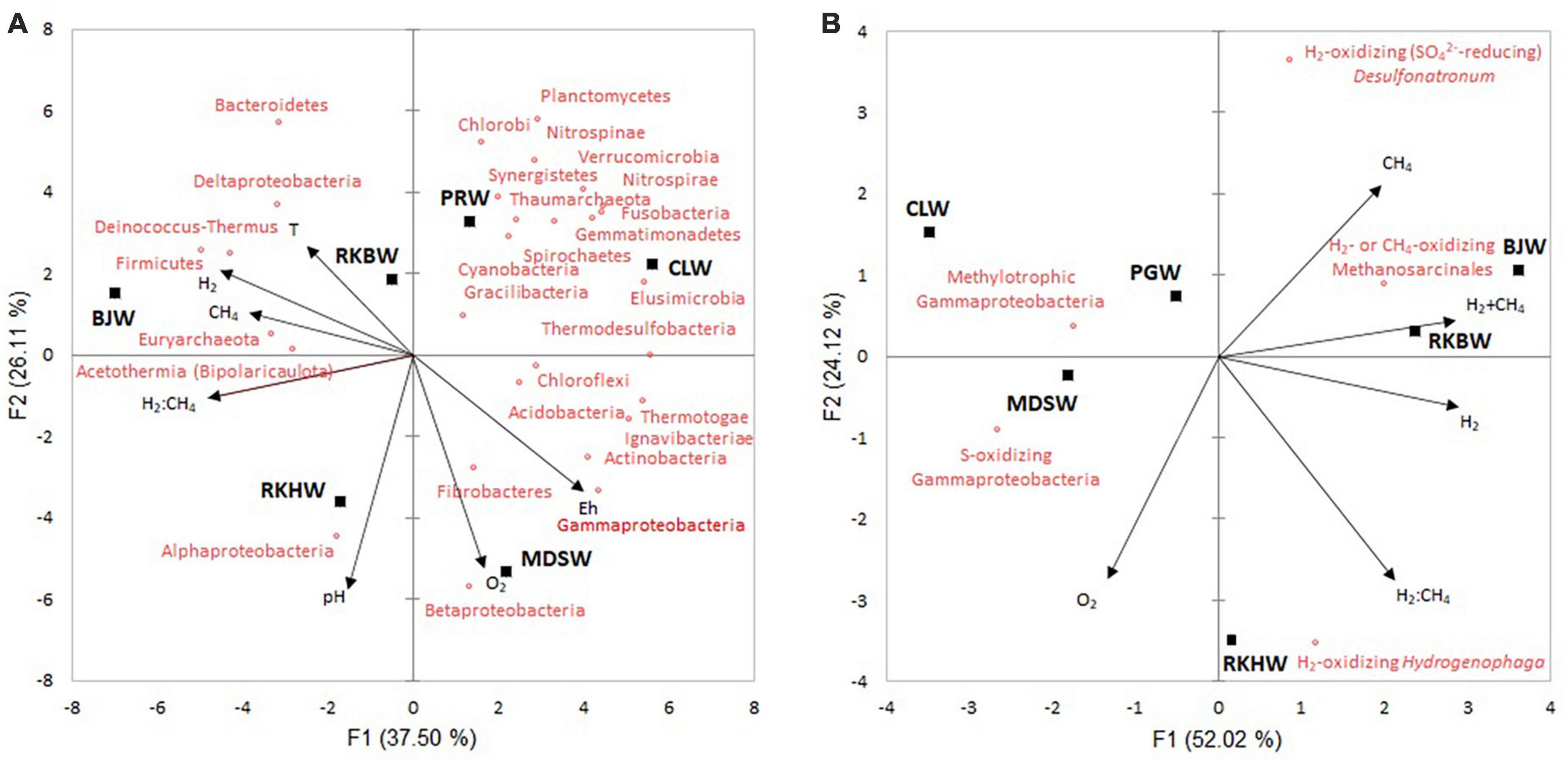
Figure 5. Principal component analysis (PCA) biplots show the variation among the high-pH water samples based on the physicochemical variables and the relative abundances of phyla (and proteobacterial classes) (A) and dominant prokaryotic functional groups (B). Black squares represent water samples of high-pH springs of New Caledonia. Arrows indicate the direction of each variable’s maximum increase and strength (through the length) to the overall distribution. CLW corresponds to the water of “La Coulée,” MDSW is the water of “Montagne des Sources,” PGW indicates the water of “Rivière des Pirogues,” RKBW and RKHW correspond to the waters of “Rivière des Kaoris” and BJW is the water of “Bain des Japonais.”
At the phylum/class level, Gammaproteobacteria was positively correlated to Eh (rS = 0.94, p < 0.05), and Deltaproteobacteria was positively correlated to temperature (rS = 0.83, p < 0.05). Firmicutes were positively correlated to H2 (rS = 0.96, p = 0.003), while the four phyla Elusimicrobia, Thermotogae, Ignavibacteria, and Thermodesulfovibrio were negatively correlated to H2 (rs = −0.89 to 0.99). Euryarchaeota (including methanogens) and Bipolaricaulota (Acetothermia) were positively correlated to CH4 (rs = 0.83 and 0.94, respectively; Supplementary Table 2).
Among the dominant OTUs, four OTUs (Gracilibacter #19, Meiothermus #37, Bacteroidetes #65, Roseinatronobacter #104) were positively correlated with H2. In comparison, five OTUs (Cyanobacteria #4, Synthrophus #9, Dethiobacter #63, Nitrospirae #74, Leptolyngbya #155) were negatively correlated with H2, and only one OTU (Methanosarcinales #12) was positively correlated with CH4 (Supplementary Table 3). Methanosarcinales (OTUs #12 and #103) were negatively correlated with methylotrophic gammaproteobacterial group (rS = −0.93, p < 0.02). The sulfur-oxidizing gammaproteobacterial group was negatively correlated with H2 (rS = −0.89, p < 0.05) (Supplementary Table 4 and Supplementary Figure 2). Prokaryotic abundances were positively correlated with H2:CH4 ratio (rS = 0.94, p < 0.02). No significant correlation was observed between diversity indices and the environmental variables measured (Supplementary Table 2).
4. Discussion
The prokaryotic communities of the New Caledonian high-pH waters show several taxa previously detected in other ecosystems sustained by serpentinization, such as The Cedars and Lost City (Schrenk et al., 2013; Suzuki et al., 2013). These microbial taxa are represented by H2-, CH4-, and S-cycling prokaryotes (Figure 6), supporting the importance of oxidation or reduction of these compounds for the growth of microbial communities in New Caledonian springs.
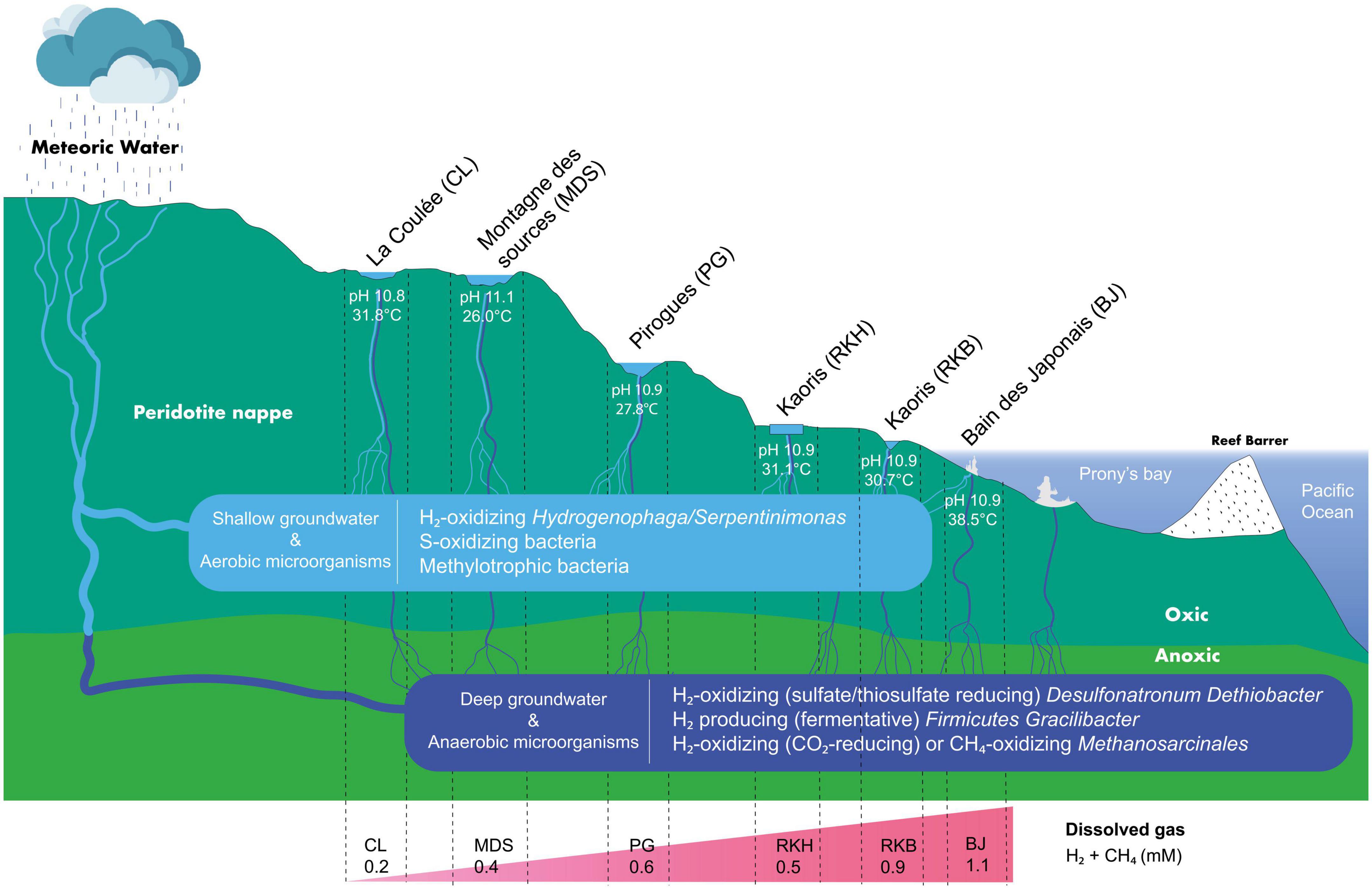
Figure 6. Schematic hydrothermal system diagram in southeastern New Caledonia’s high-pH springs (not to scale). The meteoric water moves underground into the subsurface, where the process of serpentinization produces hyperalkaline H2-rich fluids, before discharging at the geothermal springs of New Caledonia. Different microbial groups are observed depending on high-pH waters and their primary sources (shallow or deep).
Uncultured Methanosarcinales (OTUs #12 and #103) previously found in the submarine serpentinizing systems of PBHF (> 99% identity) (Quéméneur et al., 2014) and LCHF (> 95%) (Brazelton et al., 2010), could potentially anaerobically oxidize CH4 or produce it using H2 in an O2-deprived environment. Methanosarcinales phylotypes are also detected in on-land high-pH (∼11.5), H2- and CH4-rich springs of The Cedars (CA, USA) (Suzuki et al., 2013) and in the H2-depleted Voltri springs (Italy) (Quéméneur et al., 2015). Our study shows that the uncultured Methanosarcinales (related to LCMS phylotypes) are abundant in most New Caledonian springs but are found in small quantities (∼0.02% of the prokaryotes) in the CL waters, which have low CH4 and H2 contents. This was previously observed in the mildly alkaline (pH 9.3), CH4 and H2-poor waters of the La Crouen geothermal spring (Quéméneur et al., 2021). Moreover, the Methanobacterium phylotype was not detected in the hyperalkaline fluids of the studied on-land springs of New Caledonia. In contrast, this genus has been found in the alkaline geothermal spring of La Crouen (Quéméneur et al., 2021), where the novel species Methanobacterium alkalithermotolerans has been isolated (Mei et al., 2022), or in other on-land ecosystems sustained by serpentinization, such as the ophiolites of Samail (Oman) (Rempfert et al., 2017), Voltri (Northern Italy) (Quéméneur et al., 2015) and Zambales (Philippines) (Woycheese et al., 2015). On the other hand, in waters free from CH4 (e.g., La Crouen or CL), the concentrations of potential aerobic methanotrophic Gammaproteobacteria (e.g., Methylomonas, Methylobacter, Methylophaga) were higher (> 5%) than in the CH4-rich springs (<1%). These trends emphasize the importance of these potential methanogens and methanotrophs in the CH4 budget.
Bacterial members of the aerobic H2-oxidizing genera Hydrogenophaga (Lin et al., 2017) and Serpentinimonas (Bird et al., 2021) are also abundant in New Caledonian hyperalkaline springs, except in the BJ and CL waters (in which the H2 concentrations are contrasted, high and low, respectively). Both genera were frequently detected in waters and sediments/concretions of on-land serpentinite-hosted springs, where anoxic H2-rich subsurface fluids mix with oxygenated surface water (Suzuki et al., 2013; Quéméneur et al., 2015). They are present in the H2-rich hyperalkaline waters of The Cedars springs (Suzuki et al., 2013), as well as in the H2-depleted hyperalkaline waters of Voltri springs (Quéméneur et al., 2015), in the Cabeço de Vide Aquifer (Tiago and Veríssimo, 2013) and the H2-depleted alkaline waters of La Crouen (Quéméneur et al., 2021). The low level of dissolved H2 in these later spring waters could most likely be due to the high rate of H2 consumption by the hyperalkaliphilic H2-oxidizing Serpentinimonas (Marques et al., 2018).
BJ and CL waters displayed the lowest contents of Hydrogenophaga/Serpentinimonas, and the highest proportions of uncultured Firmicutes populations, previously identified from several serpentinizing systems. The dominant Firmicutes OTUs (#5 and #18) in CL waters were affiliated with Dethiobacter alkaliphilus (Sorokin et al., 2008), an anaerobic thiosulfate/polysulfide (not sulfate) reducer, polyextremophile, able to fix inorganic carbon through the Wood–Ljungdahl pathway using H2 as an electron donor (Melton et al., 2017). Dethiobacter phylotypes were abundant at Cabeço de Vide (Tiago and Veríssimo, 2013), The Cedars (Suzuki et al., 2013), the Coast Range Ophiolite (Twing et al., 2017), and the PBHF sites (Mei et al., 2016b). The dominant Firmicutes OTUs (#19 and #24203) in BJ waters are affiliated with Gracilibacter thermotolerans (Lee et al., 2006), a strict anaerobe able to produce H2 and acetate as a by-product of fermentation. Gracilibacter phylotypes were previously observed in the natural subsurface waters of the Coast Range Ophiolite (Twing et al., 2017) and the artificially enriched spring waters of The Cedars during an in situ electrochemical experiment (Rowe et al., 2017). Other potential acetogens and H2-producers, positively correlated with H2 and abundantly detected in MDS and RK waters, were affiliated with Bipolaricaulota (formerly known as Acetothermia and OP1). These microorganisms can transform low-molecular-weight organic compounds into H2 and acetate in the subsurface aquifer (Kadnikov et al., 2019).
The hydrogenotrophic, alkaliphilic, sulfate-reducing bacterium, D. cooperativum, has been isolated from a syntrophic culture growing on acetate and enriched from soda lake samples (Zhilina et al., 2005). It dominated the highly reduced waters discharged by the intertidal BJ spring (OTUs #21 and #7274), as found in a previous study (Mei et al., 2016b). These anaerobic bacteria can use sulfate, thiosulfate, elemental sulfur, or polysulfide as terminal electron acceptors. They were detected in low abundance in other on-land high-pH springs of New Caledonia (this work) and previously found in subsurface fluids of the Samail Ophiolite (Oman) (Rempfert et al., 2017). In New Caledonian high-pH waters, Desulfonatronum bacteria were inversely correlated with sulfur-oxidizing Gammaproteobacteria (e.g., Thiofaba). This points to a link between microbial activity and the sulfur chemistry, constrained by the redox conditions of the local environment. This was previously observed in borehole fluids of the Samail Ophiolite (Oman) (USA) and the California Coast Range Ophiolite (Glombitza et al., 2021).
5. Conclusion
The prokaryotic communities thriving in the hyperalkaline waters of several New Caledonia springs are mainly composed of microorganisms that use H2 as an energy source. They consist mainly of microaerophilic bacteria belonging to Hydrogenophaga/Serpentinimonas (in on-land spring waters) or anaerobic archaea belonging to a specific phylotype (designated LCMS) of uncultured Methanosarcinales (potentially able to produce CH4 using H2), which are previously detected in both submarine (i.e., Lost City, Prony Bay) (Schrenk et al., 2013; Quéméneur et al., 2014; Postec et al., 2015) and on-land serpentinizing systems (e.g., The Cedars, Voltri ophiolite) (Schrenk et al., 2013; Suzuki et al., 2013; Quéméneur et al., 2015). Thus, the relative abundance of these H2-consuming microorganisms could be used as signature-taxa (or taxonomic bioindicators) of serpentinite-hosted environments emitting natural H2.
The low abundance of Hydrogenophaga/Serpentinimonas in the water of the intertidal site BJ of the Prony Bay, where anaerobic sulfate-reducing Desulfonatronum proliferate together with other abundant anaerobic and thermophilic bacterial taxa (e.g., Bacteroidetes, Meiothermus, or Gracilibacter), suggests different water origins between terrestrial and marine springs, or subsurface seawater infiltrations through rock fractures (Monnin et al., 2014). Moreover, the co-existence of aerobic and anaerobic microorganisms in the hyperalkaline waters of other terrestrial hyperalkaline springs of the New Caledonia ophiolite could result in a mixing of deep (anoxic and warm) and surface (oxygenated and cold) waters before surface discharge at the various springs (Figure 6). This would corroborate a circulation pattern of the hydrothermal systems in ophiolites where waters flow through oxic and anoxic zones (Leong and Shock, 2020).
The relative abundance of uncultured Methanosarcinales-like sequences (designated LCMS), exclusively detected in serpentinite-hosted ecosystems (Frouin et al., 2018), was negatively correlated with aerobic methylobacteria, able to use CH4 as the sole source of carbon and energy in the waters of the hyperalkaline springs. This result suggests their implication in the consumption of CH4, also measured in the anoxic water generated by the serpentinization of the New Caledonia ophiolite.
Other anaerobic microbial taxa previously found in serpentinite-hosted environments were detected in New Caledonia hyperalkaline waters. It is the case of two anaerobic Firmicutes phylotypes affiliated with Dethiobacter and Gracilibacter genera and correlated with H2, which could also be considered as specific taxonomic bioindicators of H2 emissions in serpentinite-hosted environments. Although some Firmicutes bacteria have been isolated from the hyperalkaline concretions of the New Caledonian sites of the PBHF (including H2-producing bacteria) (Mei et al., 2014, 2016a,b; Postec et al., 2021), most remain uncultivated and their metabolisms unknown. Their role in the natural H2 budget remains to be assessed.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in this article/Supplementary material.
Author contributions
MQ, CM, and BP sampled water and gas during the H2NAT campaign. CM performed core parameter analyses. NM performed DNA extraction and PCR assays with the help of SG. MQ processed the MiSeq Illumina sequence data and wrote the first draft of the manuscript. All authors were involved in the critical revision and approval of the final version.
Funding
This project was financially supported by the French Institute of Research for Development (IRD), the INTERVIE CNRS INSU program, the H2NAT project funded by the Midi-Pyrénées Observatory (OMP, Toulouse), and the ANR MICROPRONY (N°19-CE02-0020-02).
Acknowledgments
We thank Jean Chatelier and the people at the IRD center and the DIMENC in Nouméa for their help on the field.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1196516/full#supplementary-material
References
Avias, J. (1967). Overthrust structure of the main ultrabasic New Caledonian massives. Tectonophysics 4, 531–541. doi: 10.1016/0040-1951(67)90017-0
Bird, L. J., Kuenen, J. G., Osburn, M. R., Tomioka, N., Ishii, S., Barr, C., et al. (2021). Serpentinimonas gen. nov., Serpentinimonas raichei sp. nov., Serpentinimonas barnesii sp. nov. and Serpentinimonas maccroryi sp. nov., hyperalkaliphilic and facultative autotrophic bacteria isolated from terrestrial serpentinizing springs. Int. J. Syst. Evol. Microbiol. 71:004945. doi: 10.1099/ijsem.0.004945
Brazelton, W. J., Ludwig, K. A., Sogin, M. L., Andreishcheva, E. N., Kelley, D. S., Shen, C. C., et al. (2010). Archaea and bacteria with surprising microdiversity show shifts in dominance over 1,000-year time scales in hydrothermal chimneys. Proc. Natl. Acad. Sci. U. S. A. 107, 1612–1617. doi: 10.1073/pnas.0905369107
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U. S. A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Casamayor, E. O., Massana, R., Benlloch, S., Øvreås, L., Díez, B., Goddard, V. J., et al. (2002). Changes in archaeal, bacterial and eukaryal assemblages along a salinity gradient by comparison of genetic fingerprinting methods in a multipond solar saltern. Environ. Microbiol. 4, 338–348. doi: 10.1046/j.1462-2920.2002.00297.x
Cluzel, D., Aitchison, J. C., and Picard, C. (2001). Tectonic accretion and underplating of mafic terranes in the late eocene intraoceanic fore-arc of New Caledonia (Southwest Pacific): Geodynamic implications. Tectonophysics 340, 23–59. doi: 10.1016/S0040-1951(01)00148-2
Cox, M. E., Launay, J., and Paris, J. (1982). “Geochemistry of low temperature geothermal systems in New Caledonia,” in Pacific geothermal conference and 4th NZ Geothermal Workshop, (Vienna: International Atomic Energy Agency), 453–459.
Deville, E., and Prinzhofer, A. (2016). The origin of N2-H2-CH4-rich natural gas seepages in ophiolitic context: A major and noble gases study of fluid seepages in New Caledonia. Chem. Geol. 440, 139–147. doi: 10.1016/j.chemgeo.2016.06.011
Dowd, S. E., Callaway, T. R., Wolcott, R. D., Sun, Y., McKeehan, T., Hagevoort, R. G., et al. (2008). Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 8:125. doi: 10.1186/1471-2180-8-125
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than blast. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C., and Knight, R. (2011). UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Frouin, E., Bes, M., Ollivier, B., Quéméneur, M., Postec, A., Debroas, D., et al. (2018). Diversity of rare and abundant prokaryotic phylotypes in the Prony Hydrothermal Field and comparison with other serpentinite-hosted ecosystems. Front. Microbiol. 9:102. doi: 10.3389/fmicb.2018.00102
Gaucher, E. C. (2020). New perspectives in the industrial exploration for native hydrogen. Elements 16, 8–9. doi: 10.2138/gselements.16.1.8
Geets, J., Borremans, B., Diels, L., Springael, D., Vangronsveld, J., van der Lelie, D., et al. (2006). DsrB gene-based DGGE for community and diversity surveys of sulfate-reducing bacteria. J. Microbiol. Methods 66, 194–205. doi: 10.1016/j.mimet.2005.11.002
Glombitza, C., Putman, L. I., Rempfert, K. R., Kubo, M. D., Schrenk, M. O., Templeton, A. S., et al. (2021). Active microbial sulfate reduction in fluids of serpentinizing peridotites of the continental subsurface. Commun. Earth Environ. 2:84. doi: 10.1038/s43247-021-00157-z
Hales, B. A., Edwards, C., Ritchie, D. A., Hall, G., Pickup, R. W., and Saunders, J. R. (1996). Isolation and identification of methanogen-specific DNA from blanket bog peat by PCR amplification and sequence analysis. Appl. Environ. Microbiol. 62, 668–675. doi: 10.1128/aem.62.2.668-675.1996
Kadnikov, V., Mardanov, A., Beletsky, A., Frank, Y., Karnachuk, O., and Ravin, N. (2019). Complete genome sequence of an uncultured bacterium of the candidate phylum bipolaricaulota. Microbiology 88, 461–468. doi: 10.1134/S0026261719040064
Kelley, D. S., Karson, J. A., Fruh-Green, G. L., Yoerger, D. R., Shank, T. M., Butterfield, D. A., et al. (2005). A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 307, 1428–1434. doi: 10.1126/science.1102556
Klein, F., Bach, W., and McCollom, T. M. (2013). Compositional controls on hydrogen generation during serpentinization of ultramafic rocks. Lithos 178, 55–69. doi: 10.1016/j.lithos.2013.03.008
Konn, C., Charlou, J. L., Holm, N. G., and Mousis, O. (2015). The production of methane, hydrogen, and organic compounds in ultramafic-hosted hydrothermal vents of the Mid-Atlantic Ridge. Astrobiology 15, 381–399. doi: 10.1089/ast.2014.1198
Lane, D. (1991). “16S/23S rRNA sequencing,” in Nucleic acid techniques in bacterial systematics, eds E. Stackebrandt and M. Goodfellow (New York, NY: Wiley).
Launay, J., and Fontes, J. C. (1985). Les sources thermales de Prony (Nouvelle Calédonie) et leurs précipités chimiques, exemple de formation de brucite primaire. Geol. France 1, 83–100.
Lecoeuvre, A., Ménez, B., Cannat, M., Chavagnac, V., and Gérard, E. (2021). Microbial ecology of the newly discovered serpentinite-hosted Old City hydrothermal field (southwest Indian ridge). ISME J. 15, 818–832. doi: 10.1038/s41396-020-00816-7
Lee, Y. J., Romanek, C. S., Mills, G. L., Davis, R. C., Whitman, W. B., and Wiegel, J. (2006). Gracilibacter thermotolerans gen. nov., sp. nov., an anaerobic, thermotolerant bacterium from a constructed wetland receiving acid sulfate water. Int. J. Syst. Evol. Microbiol. 56, 2089–2093. doi: 10.1099/ijs.0.64040-0
Leong, J. A. M., and Shock, E. L. (2020). Thermodynamic constraints on the geochemistry of low-temperature, continental, serpentinization-generated fluids. Am. J. Sci. 320, 185–235. doi: 10.2475/03.2020.01
Lin, S. Y., Hameed, A., Wen, C. Z., Hsu, Y. H., Liu, Y. C., Lai, W. A., et al. (2017). Hydrogenophaga aquatica sp. nov., isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 67, 3716–3721. doi: 10.1099/ijsem.0.002146
Luton, P. E., Wayne, J. M., Sharp, R. J., and Riley, P. W. (2002). The mcrA gene as an alternative to 16S rRNA in the phylogenetic analysis of methanogen populations in landfill. Microbiology 148, 3521–3530. doi: 10.1099/00221287-148-11-3521
Magen, C., Lapham, L. L., Pohlman, J. W., Marshall, K., Bosman, S., Casso, M., et al. (2014). A simple headspace equilibration method for measuring dissolved methane. Limnol. Oceanogr. Methods 12, 637–650. doi: 10.1093/chromsci/36.5.253
Marques, J. M., Etiope, G., Neves, M., Carreira, P., Rocha, C., Vance, S., et al. (2018). Linking serpentinization, hyperalkaline mineral waters and abiotic methane production in continental peridotites: An integrated hydrogeological-bio-geochemical model from the Cabeço de Vide CH4-rich aquifer (Portugal). Appl. Geochem. 96, 287–301. doi: 10.1016/j.apgeochem.2018.07.011
Maurizot, P., Sevin, B., Lesimple, S., Collot, J., Jeanpert, J., Bailly, L., et al. (2020). Mineral resources and prospectivity of non-ultramafic rocks of New Caledonia. Geol. Soc. Lond. Mem. 51, 215–245. doi: 10.1144/M51-2016-9
McCollom, T. M., and Seewald, J. S. (2013). Serpentinites, hydrogen, and life. Elements 9, 129–134. doi: 10.2113/gselements.9.2.129
Mei, N., Postec, A., Bartoli, M., Vandecasteele, C., Wils, L., Gil, L., et al. (2022). Methanobacterium alkalithermotolerans sp. nov., a novel alkaliphilic and hydrogen-utilizing methanogen isolated from an alkaline geothermal spring (La Crouen, New Caledonia). Int. J. Syst. Evol. Microbiol. 72:005554. doi: 10.1099/ijsem.0.005554
Mei, N., Postec, A., Erauso, G., Joseph, M., Pelletier, B., Payri, C., et al. (2016a). Serpentinicella alkaliphila gen. nov., sp. nov., a novel alkaliphilic anaerobic bacterium isolated from the serpentinite-hosted Prony hydrothermal field, New Caledonia. Int. J. Syst. Evol. Microbiol. 66, 4464–4470. doi: 10.1099/ijsem.0.001375
Mei, N., Postec, A., Monnin, C., Pelletier, B., Payri, C. E., Ménez, B., et al. (2016b). Metagenomic and PCR-based diversity surveys of [FeFe]-hydrogenases combined with isolation of alkaliphilic hydrogen-producing bacteria from the serpentinite-hosted Prony Hydrothermal Field, New Caledonia. Front. Microbiol. 7:1301. doi: 10.3389/fmicb.2016.01301
Mei, N., Zergane, N., Postec, A., Erauso, G., Ollier, A., Payri, C., et al. (2014). Fermentative hydrogen production by a new alkaliphilic Clostridium sp.(strain PROH2) isolated from a shallow submarine hydrothermal chimney in Prony Bay, New Caledonia. Int. J. Hydrogen Energy 39, 19465–19473. doi: 10.1016/j.ijhydene.2014.09.111
Melton, E. D., Sorokin, D. Y., Overmars, L., Lapidus, A. L., Pillay, M., Ivanova, N., et al. (2017). Draft genome sequence of Dethiobacter alkaliphilus strain AHT1 T, a gram-positive sulfidogenic polyextremophile. Standards Genomic Sci. 12, 1–9. doi: 10.1186/s40793-017-0268-9
Monnin, C., Chavagnac, V., Boulart, C., Ménez, B., Gérard, M., Gérard, E., et al. (2014). Fluid chemistry of the low temperature hyperalkaline hydrothermal system of Prony Bay (New Caledonia). Biogeosciences 11, 5687–5706.
Monnin, C., Quéméneur, M., Price, R., Jeanpert, J., Maurizot, P., Boulart, C., et al. (2021). The chemistry of hyperalkaline springs in serpentinizing environments: 1. the composition of free gases in New Caledonia compared to other springs worldwide. J. Geophys. Res. Biogeosci. 126:e2021JG006243.
Muyzer, G., De Waal, E. C., and Uitterlinden, A. (1993). Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 59, 695–700. doi: 10.1128/aem.59.3.695-700.1993
Nealson, K. H., Inagaki, F., and Takai, K. (2005). Hydrogen-driven subsurface lithoautotrophic microbial ecosystems (SLiMEs): Do they exist and why should we care? Trends Microbiol. 13, 405–410. doi: 10.1016/j.tim.2005.07.010
Nunoura, T., Oida, H., Miyazaki, J., Miyashita, A., Imachi, H., and Takai, K. (2008). Quantification of mcrA by fluorescent PCR in methanogenic and methanotrophic microbial communities. FEMS Microbiol. Ecol. 64, 240–247.
Oldham, A. L., and Duncan, K. E. (2012). Similar gene estimates from circular and linear standards in quantitative PCR analyses using the prokaryotic 16S rRNA gene as a model. PLoS One 7:e51931. doi: 10.1371/journal.pone.0051931
Ovreås, L., Forney, L., Daae, F. L., and Torsvik, V. (1997). Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 63, 3367–3373. doi: 10.1128/aem.63.9.3367-3373.1997
Pirard, C., Hermann, J., and O’Neill, H. S. C. (2013). Petrology and geochemistry of the crust–mantle boundary in a nascent arc, Massif du Sud ophiolite, New Caledonia, SW Pacific. J. Petrol. 54, 1759–1792.
Postec, A., Quéméneur, M., Bes, M., Mei, N., Benaïssa, F., Payri, C., et al. (2015). Microbial diversity in a submarine carbonate edifice from the serpentinizing hydrothermal system of the Prony Bay (New Caledonia) over a 6-year period. Front. Microbiol. 6:857. doi: 10.3389/fmicb.2015.00857
Postec, A., Quemeneur, M., Lecoeuvre, A., Chabert, N., Joseph, M., and Erauso, G. (2021). Alkaliphilus serpentinus sp. nov. and Alkaliphilus pronyensis sp. nov., two novel anaerobic alkaliphilic species isolated from the serpentinite-hosted Prony Bay Hydrothermal Field (New Caledonia). Syst. Appl. Microbiol. 44:126175. doi: 10.1016/j.syapm.2020.126175
Quéméneur, M., Bes, M., Postec, A., Mei, N., Hamelin, J., Monnin, C., et al. (2014). Spatial distribution of microbial communities in the shallow submarine alkaline hydrothermal field of the P rony Bay, New Caledonia. Environ. Microbiol. Rep. 6, 665–674. doi: 10.1111/1758-2229.12184
Quéméneur, M., Mei, N., Monnin, C., Postec, A., Wils, L., Bartoli, M., et al. (2021). Procaryotic Diversity and Hydrogenotrophic Methanogenesis in an Alkaline Spring (La Crouen, New Caledonia). Microorganisms 9:1360. doi: 10.3390/microorganisms9071360
Quéméneur, M., Palvadeau, A., Postec, A., Monnin, C., Chavagnac, V., Ollivier, B., et al. (2015). Endolithic microbial communities in carbonate precipitates from serpentinite-hosted hyperalkaline springs of the Voltri Massif (Ligurian Alps, Northern Italy). Environ. Sci. Pollut. Res. 22, 13613–13624. doi: 10.1007/s11356-015-4113-7
Rempfert, K. R., Miller, H. M., Bompard, N., Nothaft, D., Matter, J. M., Kelemen, P., et al. (2017). Geological and geochemical controls on subsurface microbial life in the Samail Ophiolite, Oman. Front. Microbiol. 8:56. doi: 10.3389/fmicb.2017.00056
Rowe, A. R., Yoshimura, M., LaRowe, D. E., Bird, L. J., Amend, J. P., Hashimoto, K., et al. (2017). In situ electrochemical enrichment and isolation of a magnetite-reducing bacterium from a high pH serpentinizing spring. Environ. Microbiol. 19, 2272–2285. doi: 10.1111/1462-2920.13723
Russell, M. J., Hall, A. J., and Martin, W. (2010). Serpentinization as a source of energy at the origin of life. Geobiology 8, 355–371.
Sander, R. (2015). Compilation of Henry’s law constants (version 4.0) for water as solvent. Atmosph. Chem. Phys. 15, 4399–4981.
Schrenk, M. O., Brazelton, W. J., and Lang, S. Q. (2013). Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 75, 575–606.
Sleep, N., Meibom, A., Fridriksson, T., Coleman, R., and Bird, D. (2004). H2-rich fluids from serpentinization: Geochemical and biotic implications. Proc. Natl. Acad. Sci. U. S. A. 101, 12818–12823. doi: 10.1073/pnas.0405289101
Sorokin, D. Y., Tourova, T., Mußmann, M., and Muyzer, G. (2008). Dethiobacter alkaliphilus gen. nov. sp. nov., and Desulfurivibrio alkaliphilus gen. nov. sp. nov.: Two novel representatives of reductive sulfur cycle from soda lakes. Extremophiles 12, 431–439. doi: 10.1007/s00792-008-0148-8
Suzuki, S., Ishii, S., Wu, A., Cheung, A., Tenney, A., Wanger, G., et al. (2013). Microbial diversity in The Cedars, an ultrabasic, ultrareducing, and low salinity serpentinizing ecosystem. Proc. Natl. Acad. Sci. U. S. A. 110, 15336–15341. doi: 10.1073/pnas.1302426110
Tiago, I., and Veríssimo, A. (2013). Microbial and functional diversity of a subterrestrial high pH groundwater associated to serpentinization. Environ. Microbiol. 15, 1687–1706. doi: 10.1111/1462-2920.12034
Truche, L., McCollom, T. M., and Martinez, I. (2020). Hydrogen and abiotic hydrocarbons: Molecules that change the world. Elements 16, 13–18.
Twing, K. I., Brazelton, W. J., Kubo, M. D., Hyer, A. J., Cardace, D., Hoehler, T. M., et al. (2017). Serpentinization-influenced groundwater harbors extremely low diversity microbial communities adapted to high pH. Front. Microbiol. 8:308. doi: 10.3389/fmicb.2017.00308
Vacquand, C., Deville, E., Beaumont, V., Guyot, F., Sissmann, O., Pillot, D., et al. (2018). Reduced gas seepages in ophiolitic complexes: Evidences for multiple origins of the H2-CH4-N2 gas mixtures. Geochim. Cosmochim. Acta 223, 437–461.
Wagner, M., Roger, A. J., Flax, J. L., Brusseau, G. A., and Stahl, D. A. (1998). Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration. J. Bacteriol. 180, 2975–2982. doi: 10.1128/JB.180.11.2975-2982.1998
Wang, X., Ouyang, Z., Zhuo, S., Zhang, M., Zheng, G., and Wang, Y. (2014). Serpentinization, abiogenic organic compounds, and deep life. Sci. China Earth Sci. 57, 878–887.
Webster, G., John Parkes, R., Cragg, B. A., Newberry, C. J., Weightman, A. J., and Fry, J. C. (2006). Prokaryotic community composition and biogeochemical processes in deep subseafloor sediments from the Peru Margin. FEMS Microbiol. Ecol. 58, 65–85.
Woycheese, K. M., Meyer-Dombard, D. A. R., Cardace, D., Argayosa, A. M., and Arcilla, C. A. (2015). Out of the dark: Transitional subsurface-to-surface microbial diversity in a terrestrial serpentinizing seep (Manleluag, Pangasinan, the Philippines). Front. Microbiol. 6:44. doi: 10.3389/fmicb.2015.00044
Zhilina, T. N., Zavarzina, D. G., Kuever, J., Lysenko, A. M., and Zavarzin, G. A. (2005). Desulfonatronum cooperativum sp. nov., a novel hydrogenotrophic, alkaliphilic, sulfate-reducing bacterium, from a syntrophic culture growing on acetate. Int. J. Syst. Evol. Microbiol. 55, 1001–1006. doi: 10.1099/ijs.0.63490-0
Keywords: bacteria, archaea, high-pH water, microbial diversity, natural hydrogen, methane, serpentinization, geothermal springs
Citation: Quéméneur M, Mei N, Monnin C, Postec A, Guasco S, Jeanpert J, Maurizot P, Pelletier B and Erauso G (2023) Microbial taxa related to natural hydrogen and methane emissions in serpentinite-hosted hyperalkaline springs of New Caledonia. Front. Microbiol. 14:1196516. doi: 10.3389/fmicb.2023.1196516
Received: 29 March 2023; Accepted: 20 June 2023;
Published: 06 July 2023.
Edited by:
Federico Lauro, Nanyang Technological University, SingaporeReviewed by:
Francesco Smedile, National Research Council (CNR), ItalyLauren M. Seyler, Stockton University, United States
Copyright © 2023 Quéméneur, Mei, Monnin, Postec, Guasco, Jeanpert, Maurizot, Pelletier and Erauso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marianne Quéméneur, bWFyaWFubmUucXVlbWVuZXVyQGlyZC5mcg==
†Present addresses: Julie Jeanpert, Agence de l’eau Rhône, Méditerranée Corse, Lyon, France
‡These authors have contributed equally to this work and share first authorship