 Yanan Li1,2
Yanan Li1,2 Shuxia Liu
Shuxia Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 19 May 2023
Sec. Terrestrial Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1170214
Long-term fertilization can result in the changes in carbon (C) cycle in the maize rhizosphere soil. However, there have been few reports on the impacts of microbial regulatory mechanisms on the C cycle in soil. In the study, we analyzed the response of functional genes that regulate the C fixation, decomposition and methane (CH4) metabolism in maize rhizosphere soil to different fertilization treatments using metagenomics analysis. As the dominant C fixation pathway in maize rhizosphere soil, the abundance of the functional genes regulating the reductive citrate cycle (rTCA cycle) including korA, korB, and IHD1 was higher under the chemical nitrogen (N) fertilizer treatments [nitrogen fertilizer (N), compound chemical fertilization (NPK), the combination of compound chemical fertilizer with maize straw (NPKS)] than maize straw return treatments [maize straw return (S), the combination of phosphorus and potassium fertilizer with maize straw (PKS)]. The NPK treatment decreased the abundance of functional genes involved in 3-hydroxypropionate bicycle (3-HP cycle; porA, porB, and porD), which was one of the major C fixation pathways in soil aside from dicarboxylate-hydroxybutyrate (DC/4-HB cycle) and Calvin cycle. The abundance of functional genes related to C degradation was higher in S, PKS and NPKS treatments than N and NPK treatments, and chemical N fertilizer application had a significant effect on C degradation. The dominant Methanaogenesis pathway in maize rhizosphere soil, used acetate as a substrate, and was significantly promoted under chemical N fertilizer application. The functional genes that were related to CH4 oxidation (i.e., pmoA and pmoB) were reduced under N and NPK treatments. Moreover, soil chemical properties had a significant impact on the functional genes related to C fixation and degradation, with SOC (r2 = 0.79) and NO3−-N (r2 = 0.63) being the main regulators. These results implied that N fertilization rather than maize straw return had a greater influence on the C cycle in maize rhizosphere soil.
In recent years, the essential role of soil microorganisms in regulating agricultural production and the carbon (C) cycle has been elucidated in numerous studies (Leadbeater et al., 2021; Ye et al., 2023). Yadav et al. (2021) proposed that soil microorganisms played significant roles in the ecosystem for nutrients cycling, and can be used for improving agro-environmental sustainability. Using microbiological systems to ameliorate agricultural production in a sustainable and eco-friendly way has been widely accepted as a future key-technology (Kumawat et al., 2022). Moreover, the structure and function of microbial community have an important impact on soil C emissions and are a major driver of the C cycle in rhizosphere micro-ecosystems, and the study of functional microbiomes has become a hot topic in recent years (Chen X. et al., 2023; Tang et al., 2023; Wu D. et al., 2023). Although the volume of rhizosphere soil is relatively small, due to the continuous secretion of various metabolites by plant roots and the discontinuous death and shedding of root surface tissues, the nutrients and microorganisms in rhizosphere soil are abundant, and their physical, chemical and biological properties are quite different from those in non-rhizosphere soil (Chen J.-Z. et al., 2023). And Cai et al. (2022) confirmed that the rhizosphere is the most active soil area, which plays an important role in soil biochemical cycling. Specially, the composition and function of the microorganisms in rhizosphere soil has drawn more attention due to the important role that the rhizosphere soil played in the uptake of mineral nutrients and water by plants (Castellano-Hinojosa et al., 2021). And soil functional microbiome is crucial to understand the ecological processes and functions such as nutrient cycles, conversion of C substances into carbon dioxide (Enebe and Babalola, 2022). The changes in the abundance of genes related to C metabolism and microbial community structure could lead to the differences in C metabolism functions, which would further affect the dynamics of soil “C source” and “C sink” in farmland ecosystem (Deng et al., 2019; Wang J. et al., 2021). Hence, the study of soil microorganisms could provide essential new insights into the regulatory mechanism of the C cycle in rhizosphere soils.
Long-term fertilization, which adds nutrients into agroecosystems to improve crop production to meet the growing food demands, is a common phenomenon in China at present (Li et al., 2009; Gao et al., 2023). However, long-term fertilization application, especially excessive fertilization could cause a series of environmental problems, such as soil acidification and greenhouse gas (GHG) emissions (Jin et al., 2022). Maize straw is abundant in organic matter, nitrogen (N), phosphorus, potassium and other nutrients, and could be used as renewable organic fertilizer (Cheng et al., 2018). Previous studies confirmed that long-term fertilization had a significant impact on the stability of soil C carbon and C emissions (Zhang et al., 2021). According to previous reports, soil physicochemical properties played important roles in controlling microbial community structure (Lauber et al., 2008; Val-Moraes et al., 2016). In addition, many bacterial communities are closely correlated with soil chemical properties and can be used as indicators of soil condition (Kuramae et al., 2015). Investigating the correlation between soil microbial communities and soil chemical properties can enable a better understanding of the mechanism of soil C cycle. However, the mechanism of soil C cycling under different fertilizer application is still unclear. Therefore, more attention should be paid to explore the association and underlying mechanism of long-term fertilization for regulating soil C cycle.
Fertilization has a significant effect on the dynamical balance of the soil C cycle (B. Liu et al., 2022). Hu et al. (2022) found that soil C fixation and degradation processes were significantly affected by manure application, but were not significantly changed under chemical fertilizer application. Fertilizer application promotes soil carbon dioxide (CO2) emissions by increasing the abundance of catabolic genes involved in carbon cycling (Enebe and Babalola, 2021). Maize straw is an important source of organic C in agroecosystem and can substantially increase the soil carbon stock (Chen et al., 2022). Furthermore, chemical N fertilization increased the abundance of methanogenic genes in croplands (Yuan et al., 2018). And the magnitude of the effect of fertilizer application on soil microbiome is depended on the soil properties such as soil N and C contents and fertilizer source (Castellano-Hinojosa et al., 2021). Recently, the studies investigating the soil functional microbial communities were mainly focused on the abundance of functional genes. However, the impacts of different long-term fertilization management on the microbial function genes relate to C cycling processes need to be determined.
In this study, the metagenomic sequencing technology was applied to quantify the C metabolic pathways (including metabolic modules and the functional genes related to C cycle) and analyze the potential metabolic response to long-term fertilizers application. We aimed to (i) explore the effects of long-term fertilizer application on soil C cycle processes at the level of metabolic modules and functional genes. (ii) To analyze the correlation between the soil properties and the abundance of functional genes related to C cycle in maize rhizosphere soil. This study hypothesized that the functional genes related to C cycle will be altered in maize rhizosphere under long-term fertilization.
The rhizosphere soil samples were collected from a long-term experimental site located in Jilin Agricultural University, Changchun City, Jilin Province, Northeast China, which has a semi-humid temperate continental climate (43°47′42″ N, 123°20′45″ E). The maize field experiment was established in 1984, with the maize seeds sown in April, and all maize residues were removed from the plot after harvest in October. The maize was rained without additional irrigation. We collected the rhizosphere soil in polyethylene bags by shaking the roots until the non-adhering soil fell off. Soil samples were collected after the harvest period, each plot was sampled at five points, and the samples were mixed into one sample for a total of 18 test samples. Six treatments, including no fertilization (CK), N fertilization (N, 150 kg/ha), compound chemical fertilization (NPK, N-P-K: 150–75-75 kg/ha), maize straw return (S, 5,000 kg/ha), the combination of phosphorus and potassium fertilizer with maize straw (PKS, P-K-S: 75-75-5,000 kg/ha), the combination of compound chemical fertilizer with maize straw (NPKS, N-P-K-S: 150-75-75-5,000 kg/ha), were implemented with three replicates per treatment for this study.
The methods adopted for the measurements of soil properties, i.e., soil pH, total N (TN), total phosphorus (TP), total potassium (TK), ammonium-N (NH4+-N), nitrate-N (NO3−-N), available phosphorus (AP) and available potassium (AK) were described in Li et al. (2020). The readily oxidizable organic carbon (ROC) was determined by potassium permanganate oxidation method, and dissolved organic carbon (DOC) was determined by TOC analyzer (Huang et al., 2021). The measured soil properties were listed in Supplementary Table S1.
The microbial DNA extraction and metagenomic sequencing was conducted according to previous research (Li et al., 2020). The non-redundant contigs set is obtained by using the MMseqs2 software. The (merged) contigs sequence set is de-redundant according to the similarity degree of 95% and the coverage degree of alignment area of 90%. Then the contigs sequence set were further filtered to remove nontarget fragment sequences and sequences with an actual depth of 0. Subsequently, contigs were used for the prediction of open reading frames (ORFs) using MetaGeneMark,1 then translated into amino acid sequences. The non-redundant gene catalog was searched against Kyoto Encyclopedia of Genes and Genomes2 for functional annotation. A total of 1,605,676,212 reads were obtained after sequencing, of which 51,418,797 contigs sequences were retained after filtering and removing chimeras (Supplementary Table S2).
We used analysis of variance (ANOVA) to analyze the differences among means, followed by least significant difference (LSD) test if the difference was significant. Redundancy analysis (RDA) was used to analyze the contribution of environmental factors (soil properties) to the changes in functional genes related to soil C cycle. Principal components analysis (PCA) was used to identify the effect of fertilizer application on the abundance of functional genes related to C cycle. The RDA and PCA analysis were performed by a free online platform for data analysis.3 We used Spearman’s rank correlation coefficients to examine the associations between the functional genes related to C cycle and soil properties, and illustrated heatmaps to show the results by an online tool of Majorbio Cloud Platform.4
There were a total of 164 genes participated in the soil C cycle according to the KEGG database in maize rhizosphere soil. The reductive citrate cycle (rTCA) was the dominant C metabolic pathway in maize rhizosphere soil (38 genes, 23.48–23.97%), and its relative abundance was changed significantly under different treatments (p < 0.05). The following was the dicarboxylate-hydroxybutyrate (DC/4-HB cycle, 20 genes, 15.37–15.95%), and N and NPK treatments significantly decreased its reductive abundance (p < 0.001). There were 21 genes participating in the Calvin cycle (12.24–12.51%) and 3-hydroxypropionate bicycle (3-HP cycle, 13.03–13.30%), and their relative abundance was not significantly different from each other under different fertilization treatments (p > 0.05). Only a small number of genes participated in the Crassulacean acid metabolism (CAM) cycle and C4-Dicarboxylic acid cycle, and the relative abundance of which increased significantly under N and NPK treatments (M00169, M00172, p < 0.001). Chemical fertilizer (N and NPK treatments) and NPKS treatments increased the abundance of glyceraldehyde-3P = > ribulose-5P (M00167). Furthermore, S and PKS treatments had no significant impact on C cycle at the module level compared with CK treatment except for M00375 (Figure 1; Supplementary Figure S1).
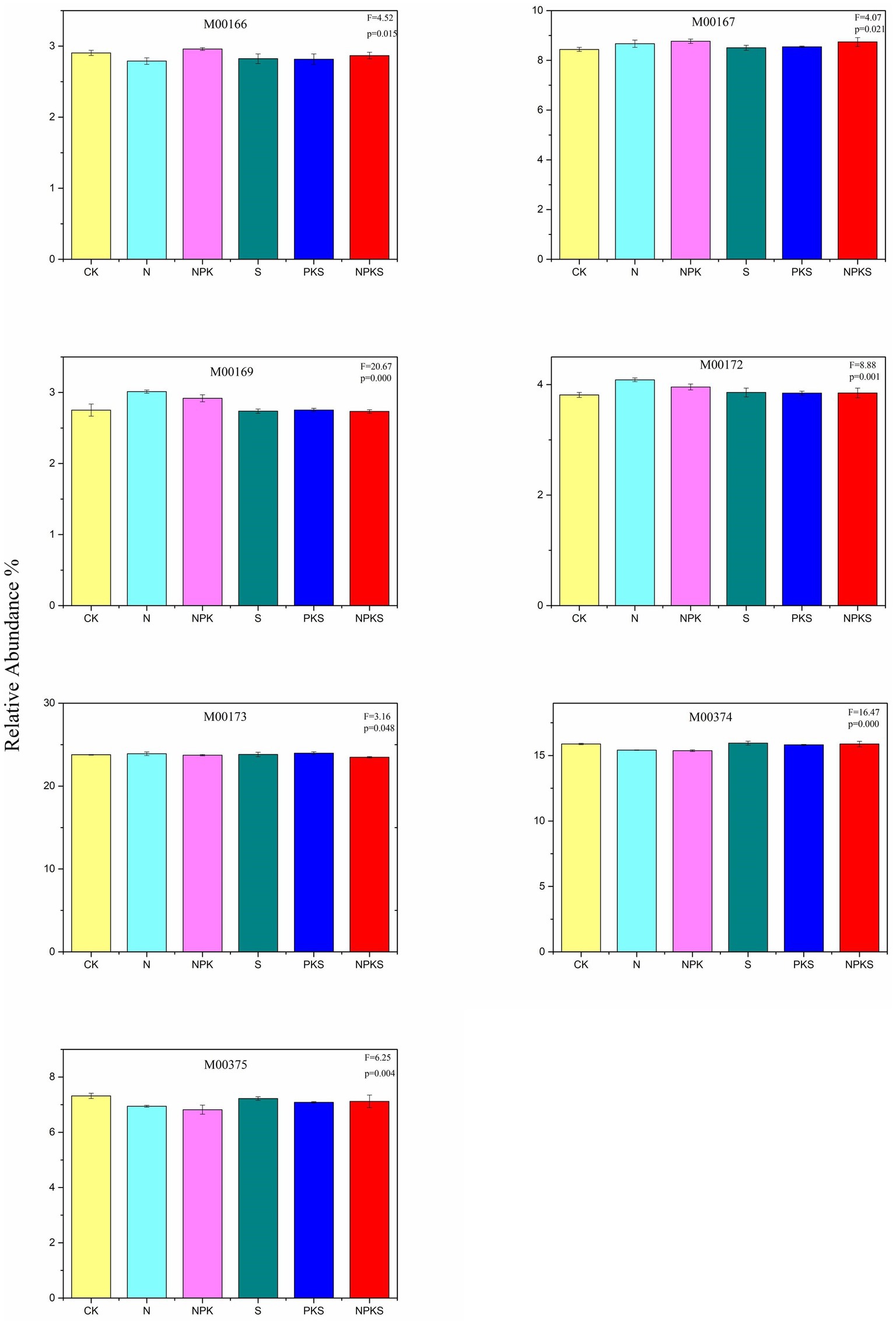
Figure 1. The relative abundance of CO2 fixation pathways (the significance test at p < 0.05 level) in maize rhizosphere soils under different fertilizer applications. Reductive pentose phosphate cycle (M00165, Calvin cycle; M00166, ribulose-5P = > glyceraldehyde-3P; M00167, glyceraldehyde-3P = > ribulose-5P), CAM cycle (M00168, dark; M00169, light), C4-Dicarboxylic acid cycle (M00170, M00171, M00172), rTCA cycle (M00173), DC/4-HB cycle (M00374), 3-HP/4-HB cycle (M00375), 3-HP cycle (M00376), and Wood-Ljungdahl pathway (M00377).
A total of 70 genes were involved in the methane (CH4) metabolic pathways. 11 genes participated in CH4 oxidation (M00174), and compared with CK treatment, the relative abundance of CH4 oxidation pathway had significantly changed under fertilization treatments except for N and PKS treatment. The relative abundance of CH4-producing metabolic pathways that used acetate as a substrate was the highest in maize rhizosphere soils (68.08–72.94%), and increased significantly under N fertilizer application (p < 0.001). In contrast, N application significantly decreased the relative abundance of CH4-producing metabolic pathways that used methylamine, dimethylamine and trimethylamine as substrates (p < 0.001). Furthermore, 19 genes participated in CH4-producing metabolic pathways that use CO2 as a substrate, and its relative abundance did not significantly change under different fertilizer application treatments (p > 0.05; Figure 2; Supplementary Figure S2).
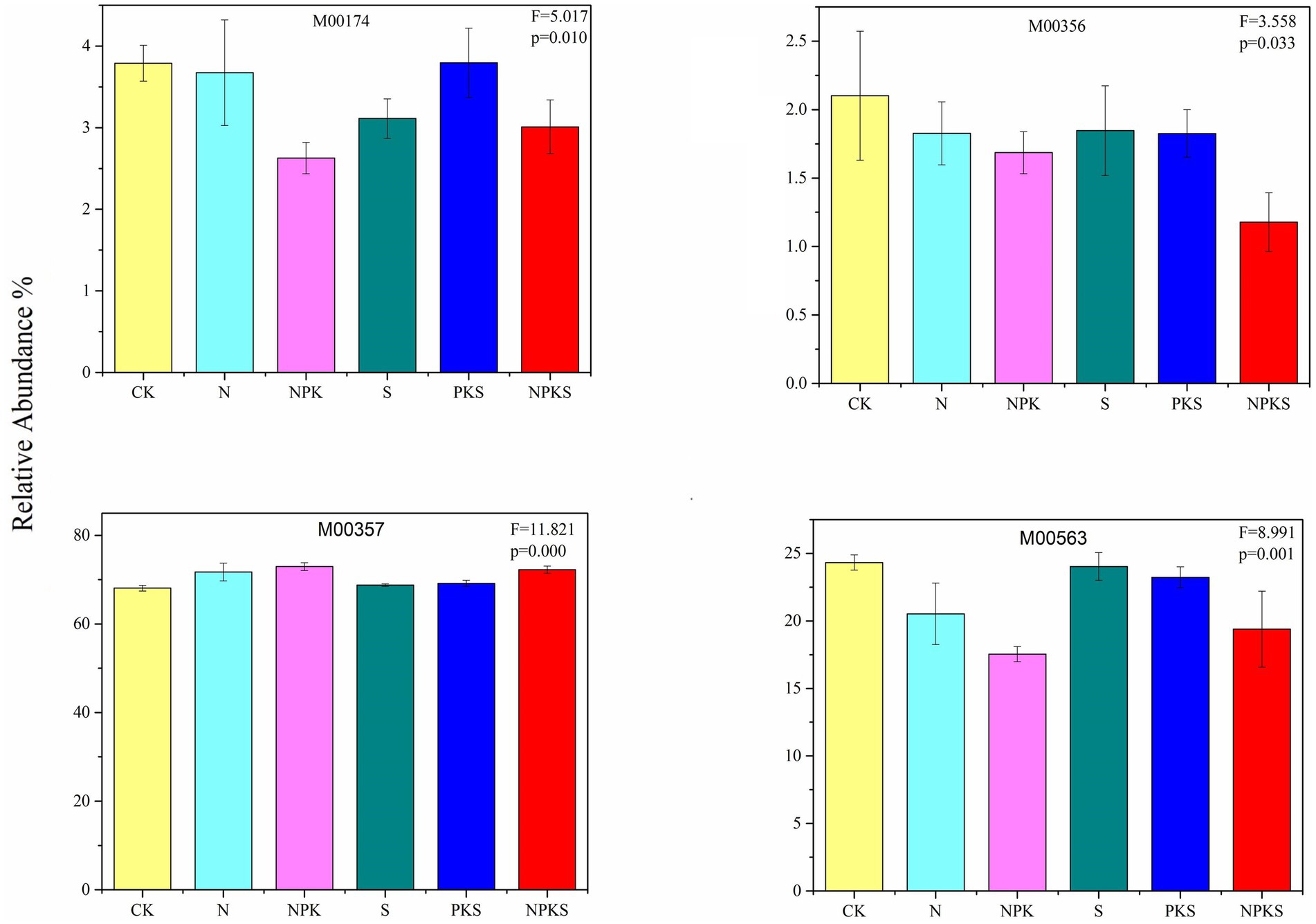
Figure 2. The relative abundance of methane metabolic pathways (the significance test at p < 0.05 level) in maize rhizosphere soils under different fertilizer applications. Methane oxidation: M00174, Methanaogenesis: methanol = > methane (M00356), acetate = > methane (M00357), methylamine/dimethylamine/trimethylamine = > methane (M00563).
A subset of 53 genes involved in C cycling was retrieved from the 18 metagenomes, all genes listed in Supplementary Table S3. They were involved in C fixation and degradation and CH4 metabolism (Wang S. et al., 2021). The functional genes related to C fixation exhibited the highest abundance, followed by the C degradation functional genes (Table 1). Within the C fixation group, the relative abundance of functional genes related to multiple systems (13.76–15.36%), C monoxide (CO) oxidation (18.34–21.15%), and the rTCA cycle (13.58–15.37%) was higher than that of functional genes participated in the other C fixation pathways. Within the C degradation group, functional genes related to hemicellulose degradation exhibited highest abundance (9.29–10.98%) across all treatments, followed by starch degradation (7.49–8.13%) and cellulose degradation (3.20–4.33%) genes.
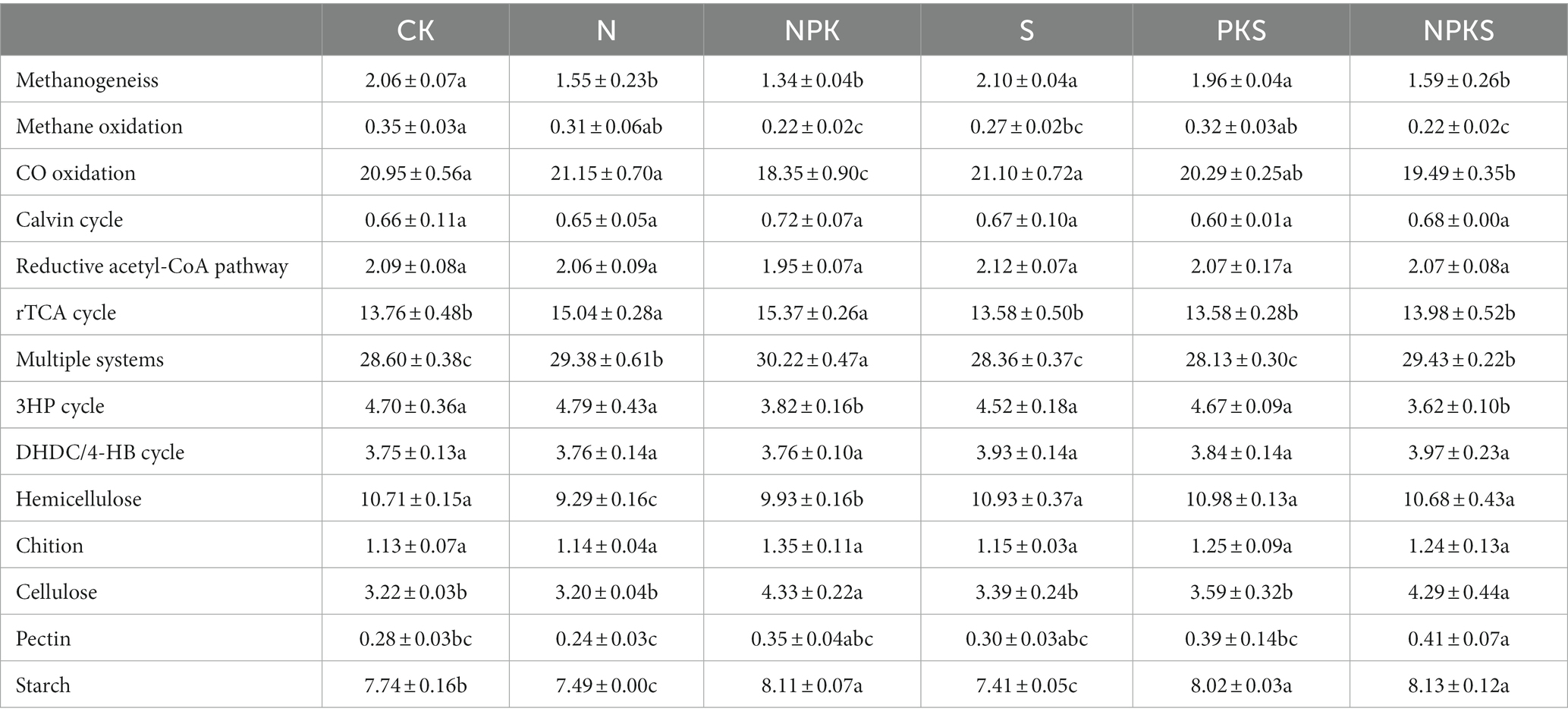
Table 1. The relative abundance of functional genes participated in carbon cycle in maize rhizosphere soils under different fertilizers application.
Functional genes participated in CO oxidation (coxS) were decreased significantly under N treatment (p < 0.05), similar with the effects of fertilizer application on cooC gene related to reductive acetyl-CoA pathway. In the rTCA cycle, the relative abundances of korA, korB and IHD1 were higher under chemical fertilization treatments than maize straw return treatments. The NPK treatment decreased the functional genes involved in 3-HP cycle (porA, porB, and porD). No significant changes were found in the functional genes related to Calvin cycle and DC/4-HB cycle under different fertilizer application treatments (p > 0.05). The relative abundance of PCCA, PCCB, accA, accC, and accD related to multiple system in C fixation was also markedly enhanced by the N, NPK and NPKS treatments (Figure 3).
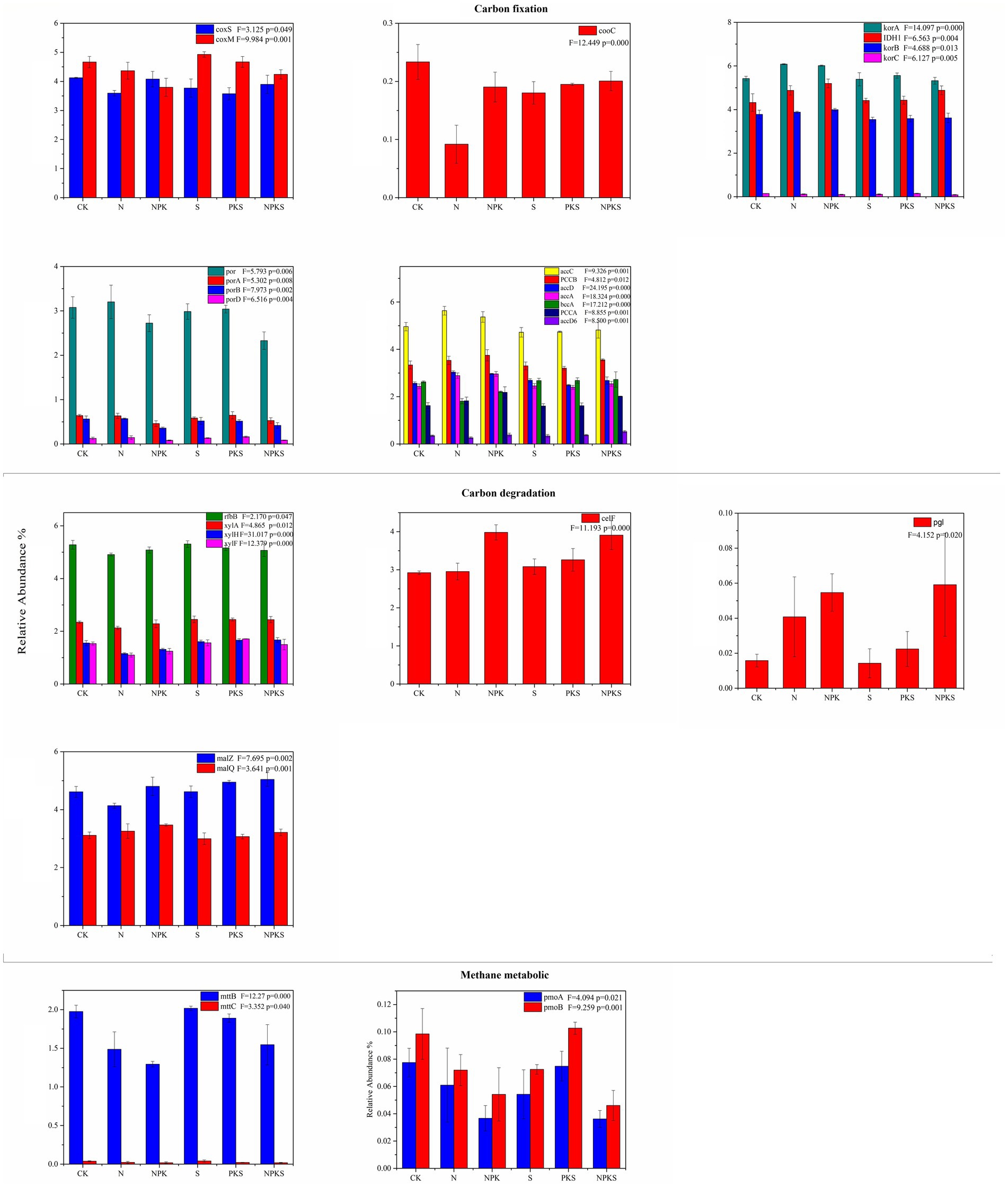
Figure 3. The relative abundance of functional genes participated in carbon cycle (the significance test at p < 0.05 level) in maize rhizosphere soils under different fertilizer applications. coxS and coxM involved in carbon-monoxide oxidation; cooC involved in reductive acetyl-CoA pathway; korA, korB, korC, and IDH1 involved in the rTCA cycle; porA, porB, porD, and por involved in 3HP cycle; bccA, PCCA, PCCB, accD6, accA, accC. and accD involved in multiple systems in carbon fixation. In carbon degradation xylA, xylF, xylH, and rfbB involved in hemicellulose; celF involved in cellulose; pgl involved in pectin; malZ and malQ involved in starch. In methane metabolic mttB and mttC involved in methanogenesis; pmoA, pmoB, involved in methane oxidation.
Compared with CK treatment, N and NPK treatment significantly decreased the relative abundance of functional genes (including xylA, xylH, xylF, and rfbB) related to hemicellulose degradation (p < 0.05). However, other treatments had no significant effects on hemicellulose degradation genes (p > 0.05). Besides, the relative abundance of the functional genes related to cellulose (celF) and pectin (pgl) for NPK and NPKS treatments were higher than that for other treatments. The abundance of starch degradation gene malZ declined under N fertilizer application, increased under NPKS fertilizer application (p < 0.05). The N, NPK, and NPKS treatments inhibited the process of methanogenesis. Compared with N fertilizer applications, the relative abundance of mttB, which participated in methanogenesis, was the higher under straw return application. Moreover, the N, NPK, and NPKS treatments reduced the abundance of CH4 oxidation genes in maize rhizosphere soil (Figure 3).
The results of PCA indicated that fertilizer application explained most of the variation in the C cycling genes across all samples (PC1 of PCA: 99.2%). The sample points from the CK-, S- and PKS-treated soils grouped together and were separated from those of other treatments (Figure 4A). The variations among these genes under different treatments were probably caused by the soil properties. The RDA showed that the different forms of soil C (soil organic carbon [SOC], dissolved organic carbon [DOC], readily oxidizable organic carbon [ROC]) and available nutrients (nitrate N [NO3−-N], active potassium [AK], active phosphorus [AP], total phosphorus [TP]) and pH had significant effects on C cycle (p < 0.05; Figure 4B). The soil properties had strong effects on the C fixation pathways including reductive acetyl-CoA pathway (cooC), rTCA cycle (korA, korB, and IDH1), 3HP (porB) and multiple systems (bccA, accA, accC, and accD). For C degradation, SOC, DOC, ROC, total nitrogen (TN), AP and AK were significantly positively correlated with the hemicellulose (xylF, xylH, and xylA) and cellulose (celF and cbhA) and pectin (pel) degradation. The NO3−-N was significantly negatively correlated with xylF, xylH, and xylA, which were relative with hemicellulose degradation, and was positively correlated with starch degradation gene malQ. However, the soil properties had no significant correlation with the functional genes related to the Calvin cycle. Moreover, the methanogenesis-relative genes mttB had positive correlation with pH and AK, and negative correlation with TN, ammonium nitrogen (NH4+-N) and NO3−-N. Functional gene pmoC that participated in CH4 oxidation had strong correlation with DOC, ROC, TN, AP, and TP (Figure 5; Supplementary Table S4).
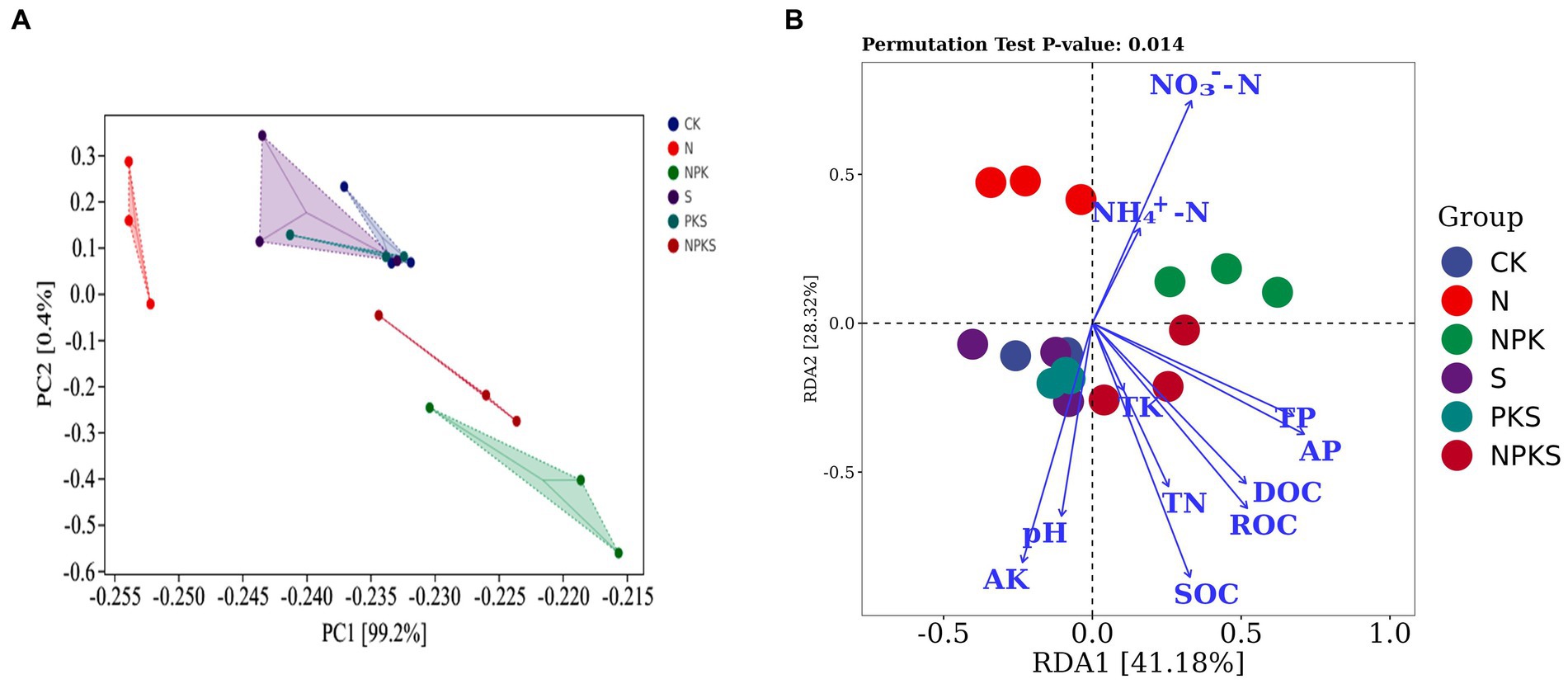
Figure 4. Effects of fertilizers application on functional genes in soil carbon cycles. Functional genes identified by principal components analysis (A). Redundancy analysis of microbial functional genes associated with soil properties (B). SOC, soil organic carbon; DOC, dissolved organic carbon; ROC, readily oxidizable organic carbon; TN, total nitrogen; TP, total phosphorus; TK, total potassium; NH4+-N, ammonium nitrogen; NO3−-N, nitrate nitrogen; AP, active phosphorus; AK, active potassium.
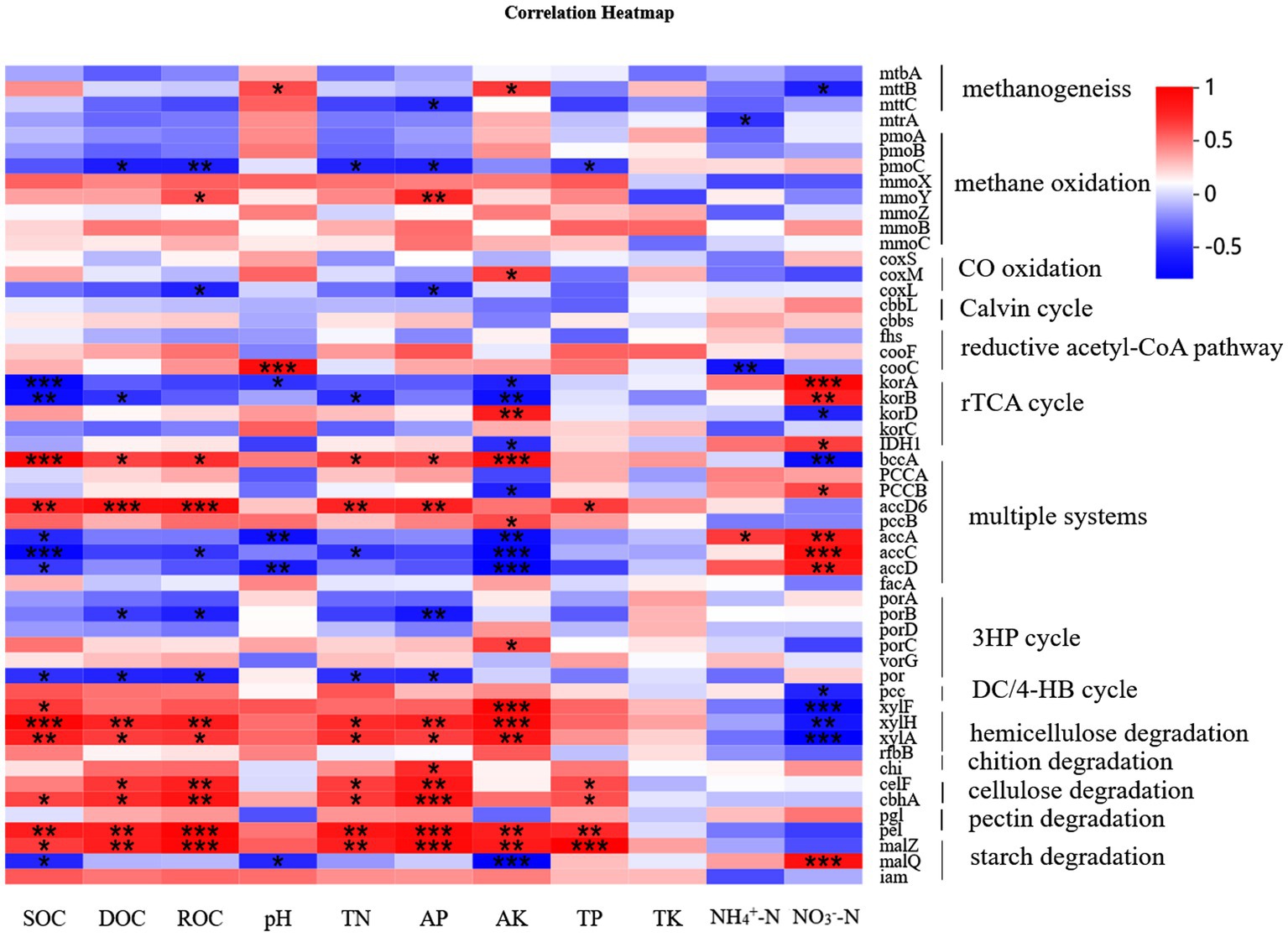
Figure 5. The heatmap of the correlation between microbial functional genes and soil properties.
Fertilizer application could influence soil C turnover (Gong et al., 2012). Similarly, in this study, chemical fertilizer application decreased the SOC content, while maize straw return and chemical fertilizer added with maize straw increased the SOC content compared to CK treatment. Active organic carbon is an essential component of soil organic carbon, the increase of its content can provide more available carbon sources for soil microorganisms, and then affect soil carbon cycle (Luo et al., 2018). Moreover, fertilizer application stimulated the transformation of stable SOC to labile forms such as DOC and ROC (Supplementary Table S1). The result confirmed that long-term fertilization can affect the C cycle by changing the content of different forms of soil carbon. Soil C transformation was found to be driven by microorganisms (Kramer and Gleixner, 2006) and previous studies have shown that soil microorganisms participate in the transformation of various forms of C in soil, and can mediate the CO2 fixation and C degradation (Beulig et al., 2016; Agarwal et al., 2017; Jacoby et al., 2017). This study assessed eight CO2 fixation pathways including the plant photosynthesis pathway (CAM cycle) in maize rhizosphere soil that existed in the KEGG database (Li et al., 2022). It might be because the maize rhizosphere could provide the substrates needed for microbial photosynthesis. As the dominant C fixation pathway, the rTCA cycle only needed two adenosine triphosphates (ATP) to complete this metabolic process (Claassens et al., 2016), and the combined application of chemical fertilizer and maize straw (NPKS) decreased the abundance of the rTCA (Figure 1). Compared to the rTCA cycle, the Calvin cycle, a major pathway for CO2 fixation that catalyzes the carboxylation of ribulose-1,5-bisphosphate into two molecules of 3-phosphoglycerate, requires more energy (Huang et al., 2022). Long-term fertilization could affect C cycle by altering C metabolism pathways. It was reported that long-term fertilizer application had no significant effects on the Calvin cycle (Hu et al., 2022), but the glyceraldehyde-3P = > ribulose-5P (M00167) metabolic pathway was strongly improved under chemical fertilization (N and NPK treatments) and the combined application of chemical fertilizer and maize straw (NPKS) in this study (Figure 1). Previous study concluded that soil water content and N fertilizer had significant effects on the CAM, and N fertilizer changed the net CO2 uptake and photosynthetic capacity of the CAM in soil (Ma et al., 2022). Similar results were observed in this study, that is, compared with CK treatment, chemical fertilization (N and NPK treatments) promoted the CAM metabolic pathway. In addition, chemical fertilization also increased the relative abundance of the C4-Dicarboxylic acid metabolic pathway. Moreover, compared with the CK treatment, maize straw return and the combined application of maize straw and PK fertilizer had no significant effects on C fixation metabolic pathways, indicating that the C fixation in soil was mainly affected by chemical N fertilizers.
Among the different agricultural production systems, paddy fields are a source of CH4 emissions whereas dryland is a weak sink for CH4 uptake (Snyder et al., 2009; Wu P. et al., 2023). Previous studies mainly focused on CH4 emissions, whereas the microbial process of CH4 metabolism in soil was less understood. Methane metabolism mainly consists of methanaogenesis and CH4 oxidation that occurred in maize rhizosphere soil. The dominant metabolic pathways of methanaogenesis used acetate as the substrate (Padhy et al., 2022), thus increasing significantly under chemical N fertilization and the combined application of chemical N fertilizer with maize straw. Oppositely, the chemical N fertilization inhibited the methanaogenesis pathway from methylamine, dimethylamine and trimethylamine. Methanosphaera could reduce the methyl group of methanol to CH4 (Ferry, 2010), the abundance of which decreased under chemical fertilizer application and combined application of chemical fertilizer and maize straw (Supplementary Figure S3). Moreover, chemical fertilization and maize straw return had no significant effect on CH4 metabolism that utilized CO2 as a substrate, probably because the Methanocella genera grow and produce CH4 from CO2 and H2, which was not significantly affected by N fertilization treatments (Yuan et al., 2018; Supplementary Figure S3).
The functional genes related to C fixation have been studied extensively in terrestrial ecosystems and most of the previous studies focused on the cbbL and cbbM genes driving the Calvin cycle (Liu et al., 2016; Li et al., 2017; Treseder et al., 2018). However, we found the relative abundance of cbbL and cbbM genes was lower than the functional genes participated in multiple system, CO oxidation and rTCA cycle. This result is similar to the pattern of soil C metabolism at module level. Chemical N addition can induce a shift in soil N availability, which influences soil C turnover (Wang et al., 2018). In this study, the abundances of the functional genes related to C fixation including the rTCA cycle, 3HP cycle, multiple system and CO oxidation responded distinctively to different fertilization treatments. This was similar with (Schleuss et al., 2019), but different from the results reported by Hu et al. (2022), which was probably because the samples were obtained from the maize rhizosphere soil in this study and surface soil in previous study, respectively. Three functional genes related with the CO oxidation process were obtained from the maize rhizosphere, among which the abundance of coxL was the highest and did not significantly vary among different treatments. However, the abundance of coxS and coxM decreased under chemical fertilizer application, and coxS was also influenced by maize straw return. It indicated that chemical N fertilizer rather than maize straw imposed significant effects on CO oxidation in soil. It is consistent with the dynamic pattern of functional genes related with the multiple systems and 3 HP cycle. The N and NPK treatments had higher functional gene abundance that was related to rTCA cycle (including korA, korB, and IDH1) than S and PKS treatments, and the relative abundance of PCCA, PCCB, accA, accC, and accD related to multiple systems in C fixation was also markedly enhanced by the N, NPK and NPKS treatments. These results indicated that chemical fertilizer contributed more to the variations in soil microbial C fixation than maize straw.
The functional genes related to C degradation mostly participated in hemicellulose, starch and cellulose degradation. The abundance of functional genes related to hemicellulose, cellulose and starch degradation were significantly different from each other under different fertilization treatments in this study. The amendment of maize straw could enhance the nutrient availability for microbial growth such as organic C and N in soil, and improve the activity soil hydrolytic enzymes, and provide more C source for soil microorganisms (Zhu et al., 2019; Wu et al., 2020). In addition, previous findings showed that maize straw addition increased the amount of macroaggregates in soil, thus improving the permeability of soil and promoting the growth and development of aerobic microorganisms (Zhang et al., 2022). Therefore, the abundance of functional genes related to C degradation was higher under S, PKS and NPKS treatments than N and NPK treatments. Moreover, chemical fertilizer application decreased the abundance of functional genes related to hemicellulose degradation (i.e., xylA, xylH, xylF, and rfbB). It was because that long-term application of chemical fertilizer resulted in the decrease of soil microbial diversity and the change in microbial community structure, thus reducing the capacity of microbial C decomposition in soil (Jin et al., 2022). Previous study concluded that the combined application of chemical fertilizer and maize straw improved the activity of soil enzymes that degraded cellulose and hemicellulose from straw into glucose (β-Cellobiohydrolase, βxylosidase and β-glucosidase; Liu et al., 2022). Consistently, in this study, the combined application of chemical fertilizer and maize straw increased the abundance of celF, a functional gene related to cellulose degradation.
The abundance of functional gene mttB which was the highest abundance gene for methanogenesis, and decreased by 0.49, 0.69, and 0.43% under N, NPK and NPKS treatments, respectively, which was consistent with the findings of Wu P. et al. (2023). Compared with S and PKS treatments (without chemical N fertilizer), N, NPK, and NPKS treatments produced lower CH4. Previous study concluded that the limiting factor of CH4 emission was the precursor for CH4 production instead of the mineral nitrogen in soil (Wang Xiaoqi et al., 2017). The decomposition of maize straw increases the content of SOC, which provides a higher level of C source for soil microorganisms (Liang et al., 2021; Ren et al., 2023). Consequently, the decomposition of organic C into monosaccharides and then acids by microorganisms was accelerated, which promotes the CH4 production (Wang Xiaoqi et al., 2017; Zhang and Zhang, 2020). In this study, long-term fertilizer application significantly affected the abundance of CH4 oxidation genes including pmoA and pmoB. In conclusion, chemical fertilizer application rather than maize straw addition exhibited significant influences on soil C fixation, degradation and CH4 metabolism. The above results indicated that long-term fertilization affect C cycle by alter functional genes related to C cycle.
Long-term chemical fertilization can greatly alter the microbial function by directly increasing available nutrients or indirectly changing the soil physical and chemical properties. The chemical properties including SOC, DOC, ROC, TN, NO3−-N, AK, AP, TP, and pH changed significantly under different fertilization treatments in the maize rhizosphere soil in this study (Supplementary Table S1), which resultantly influenced the C fixation and degradation (Figures 4B, 5). Carbon and N sources in soil such as SOC, ROC, DOC, and TN and NO3−-N provided C and N source for microbial growth (Xie et al., 2012). Labile SOC had negative effects on C fixation and positive effects on C degradation. However, the effects of NO3−-N on C fixation and degradation were opposite with that of SOC (Figure 5). Previous studies confirmed that soil P was the most influential driver of C cycle, and could efficiently maintain the microbial growth and enhance the microbial function for mediating soil C cycle (Yao et al., 2018; Hu et al., 2022). In this study, AP had significantly positive correlations with the abundance of bbcA, accD6, xylH, xylA, chi, celF, cbhA, pgl, malZ, and negative correlations with that of por, porB and coxL. Soil pH was also an important factor regulating the soil C cycle. Soil pH could alter the availability of soluble nutrients in soil, and directly affect the contents of dissolved CO2 and the use of C sources (CO2 and HCO3−) by soil microorganisms, thereby imposing a significant influence on the C cycle (Huang et al., 2022). The result also confirmed that long-term fertilization can affect soil functional genes and carbon cycle by changing soil physical and chemical properties.
In this study, there were eight C fixation pathways in maize rhizosphere soil, and the dominant one was the reductive citrate cycle (rTCA cycle), followed by the dicarboxylate-hydroxybutyrate (DC/4-HB cycle), 3-hydroxypropionate bicycle (3-HP cycle) and the Calvin cycle. Long-term fertilizer application had a significant effect on soil C fixation except for the Calvin and 3-HP cycles. Chemical fertilizers increased the abundance of the C4-Dicarboxylic acid cycle metabolic pathway and the CAM light metabolic pathway, and inhibited the DC/4-HB cycle. Compared with CK treatment, S and PKS treatments had no significant influences on the soil C fixation. The relative abundance of CH4-producing metabolic pathways that used acetate as a substrate was the highest in maize rhizosphere soils, and increased significantly under N fertilizer application relative to CK. However, long-term fertilizer application had no significant effect on CH4 metabolic pathway that used CO2 as a substrate. The functional genes related to C degradation mostly participated in hemicellulose, cellulose and starch degradation. The abundance of functional genes related to C degradation was higher under S, PKS and NPKS treatments than N and NPK treatments, indicating that chemical fertilizer application had a significant influence on C degradation. In addition, chemical fertilizer application decreased the abundance of methanogenesis genes (i.e., mttB and mttC) and CH4 oxidation genes (i.e., pmoA and pmoB). In conclusion, this study implied that N fertilizer rather than maize straw application had significant impacts on the C cycle in maize rhizosphere soil. Moreover, soil chemical properties had significant influences on the abundance of functional genes related to C fixation and degradation, with SOC and NO3−-N being the major drivers. Overall, our results highlighted the important of long-term fertilization in C cycle and the microorganism regulated C cycle in maize rhizosphere.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
SL and WH: conceptualization and funding acquisition. CW and HC: methodology. YL and YZ: software. SL: resources and project administration. YL: data curation, writing—original draft preparation, and visualization. YL and SL: writing—review and editing. CW: supervision. All authors contributed to the article and approved the submitted version.
This work is a contribution to Natural Science Foundation of Jilin Province of China (20210101100JC) and Special Project on Science and Technology Innovation Capacity Construction of Beijing Academy of Agriculture and Forestry Sciences (KJCX20220416).
We thanks to Junnan Wu for soil collecting and sample pretreatment, Daijia Fan, Beijing Academy of Agriculture and Forestry Sciences for polishing the paper to eliminate language problems.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1170214/full#supplementary-material
Agarwal, L., Dafale, N. A., and Purohit, H. J. (2017). “Microbial CO2 fixation bioprocesses and desert as future carbon sink” in Optimization and applicability of bioprocesses. eds. H. J. Purohit, V. C. Kalia, A. N. Vaidya, and A. A. Khardenavis (Singapore: Springer Singapore), 325–348. doi: 10.1007/978-981-10-6863-8_16
Beulig, F., Urich, T., Nowak, M., Trumbore, S. E., Gleixner, G., Gilfillan, G. D., et al. (2016). Altered carbon turnover processes and microbiomes in soils under long-term extremely high CO2 exposure. Nat. Microbiol. 1:15025. doi: 10.1038/nmicrobiol.2015.25
Cai, A., Tang, S., Waqas, M. A., Wang, B., Tian, D., Zhang, Y., et al. (2022). Magnitude, direction, and drivers of rhizosphere effect on soil nitrogen and phosphorus in global agroecosystem. Int. Soil Water Conserv. Res. doi: 10.1016/j.iswcr.2022.07.004
Claassens, N. J., Sousa, D. Z., Dos Santos, V. A. P. M., De Vos, W. M., and Van Der Oost, J. (2016). Harnessing the power of microbial autotrophy. Nat. Rev. Microbiol. 14, 692–706. doi: 10.1038/nrmicro.2016.130
Castellano-Hinojosa, A., Strauss, S. L., González-López, J., and Bedmar, E. J. (2021). Changes in the diversity and predicted functional composition of the bulk and rhizosphere soil bacterial microbiomes of tomato and common bean after inorganic N-fertilization. Rhizosphere 18:100362. doi: 10.1016/j.rhisph.2021.100362
Chen, X., Han, X.-Z., Wang, X.-H., Guo, Z.-X., Yan, J., Lu, X.-C., et al. (2022). Inversion tillage with straw incorporation affects the patterns of soil microbial co-occurrence and multi-nutrient cycling in a Hapli-Udic Cambisol. J. Integr. Agric. 22, 1546–1559. doi: 10.1016/j.jia.2022.12.011
Chen, J.-Z., Huang, X.-L., Sun, Q.-W., and Liu, J.-M. (2023). Bulk soil microbial reservoir or plant recruitment dominates rhizosphere microbial community assembly: evidence from the rare, endangered Lauraceae species Cinmaomum migao. Ecol. Indic. 148:110071. doi: 10.1016/j.ecolind.2023.110071
Chen, X., Xie, Y., Wang, J., Shi, Z., Zhang, J., Wei, H., et al. (2023). Presence of different microplastics promotes greenhouse gas emissions and alters the microbial community composition of farmland soil. Sci. Total Environ. 879:162967. doi: 10.1016/j.scitotenv.2023.162967
Cheng, C., Wang, J., Cheng, H., Luo, K., and Shang, Q. (2018). Effects of straw returning and tillage system on crop yield and soil fertility quality in paddy field under double-cropping-rice system. Acta Pedol. Sin. 55, 247–257. doi: 10.11766/trxb201707090275
Deng, J., Zhu, W., Zhou, Y., and You, Y. (2019). Soil organic carbon chemical functional groups under different revegetation types are coupled with changes in the microbial community composition and the functional genes. Forests 10:240. doi: 10.3390/f10030240
Enebe, M. C., and Babalola, O. O. (2021). Soil fertilization affects the abundance and distribution of carbon and nitrogen cycling genes in the maize rhizosphere. AMB Express 11:24. doi: 10.1186/s13568-021-01182-z
Enebe, M. C., and Babalola, O. O. (2022). Functional diversity of bacterial communities in the rhizosphere of maize grown on a soil under organic and inorganic fertilization. Sci. Afr. 16:e01212. doi: 10.1016/j.sciaf.2022.e01212
Ferry, J. G. (2010). The chemical biology of methanogenesis. Planet. Space Sci. 58, 1775–1783. doi: 10.1016/j.pss.2010.08.014
Gao, P., Zhang, T., Lei, X. Y., Cui, X. W., Lu, Y. X., Fan, P. F., et al. (2023). Improvement of soil fertility and rice yield after long-term application of cow manure combined with inorganic fertilizers. J. Integr. Agric. doi: 10.1016/j.jia.2023.02.037
Gong, W., Yan, X., and Wang, J. (2012). The effect of chemical fertilizer application on carbon input and export in soil—a pot experiment with wheat using natural 13C abundance method. Geoderma 189-190, 170–175. doi: 10.1016/j.geoderma.2012.05.007
Hu, X., Gu, H., Liu, J., Wei, D., Zhu, P., Cui, X. A., et al. (2022). Metagenomics reveals divergent functional profiles of soil carbon and nitrogen cycling under long-term addition of chemical and organic fertilizers in the black soil region. Geoderma 418:115846. doi: 10.1016/j.geoderma.2022.115846
Huang, Q., Huang, Y., Wang, B., Dippold, M. A., Li, H., Li, N., et al. (2022). Metabolic pathways of CO2 fixing microorganisms determined C-fixation rates in grassland soils along the precipitation gradient. Soil Biol. Biochem. 172:108764. doi: 10.1016/j.soilbio.2022.108764
Huang, R., Lan, T., Song, X., Li, J., Ling, J., Deng, O., et al. (2021). Soil labile organic carbon impacts C:N:P stoichiometry in urban park green spaces depending on vegetation types and time after planting. Appl. Soil Ecol. 163:103926. doi: 10.1016/j.apsoil.2021.103926
Jacoby, R., Peukert, M., Succurro, A., Koprivova, A., and Kopriva, S. (2017). The role of soil microorganisms in plant mineral nutrition-current knowledge and future directions. Front. Plant Sci. 8:1617. doi: 10.3389/fpls.2017.01617
Jin, B.-J., Bi, Q.-F., Li, K.-J., Yu, Q.-G., Ni, L., Lin, X.-Y., et al. (2022). Long-term combined application of chemical fertilizers and organic manure shapes the gut microbial diversity and functional community structures of earthworms. Appl. Soil Ecol. 170:104250. doi: 10.1016/j.apsoil.2021.104250
Kramer, C., and Gleixner, G. (2006). Variable use of plant- and soil-derived carbon by microorganisms in agricultural soils. Soil Biol. Biochem. 38, 3267–3278. doi: 10.1016/j.soilbio.2006.04.006
Kumawat, K. C., Razdan, N., and Saharan, K. (2022). Rhizospheric microbiome: bio-based emerging strategies for sustainable agriculture development and future perspectives. Microbiol. Res. 254:126901. doi: 10.1016/j.micres.2021.126901
Kuramae, E. E., Etienne, Y., Wong, L. C., Pijl, A. S., Van, V. J. A., and Kowalchuk, G. A. (2015). Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol. Ecol. 1, 12–24. doi: 10.1111/j.1574-6941.2011.01192.x
Lauber, C. L., Strickland, M. S., Bradford, M. A., and Fierer, N. (2008). The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 40, 2407–2415. doi: 10.1016/j.soilbio.2008.05.021
Leadbeater, D. R., Oates, N. C., Bennett, J. P., Li, Y., Dowle, A. A., Taylor, J. D., et al. (2021). Mechanistic strategies of microbial communities regulating lignocellulose deconstruction in a UK salt marsh. Microbiome 9:48. doi: 10.1186/s40168-020-00964-0
Li, W., Li, Y., Lv, J., He, X., Wang, J., Teng, D., et al. (2022). Rhizosphere effect alters the soil microbiome composition and C, N transformation in an arid ecosystem. Appl. Soil Ecol. 170:104296. doi: 10.1016/j.apsoil.2021.104296
Li, Y., Wang, C., Wang, T., Liu, Y., Jia, S., Gao, Y., et al. (2020). Effects of different fertilizer treatments on rhizosphere soil microbiome composition and functions. Land 9:329. doi: 10.3390/land9090329
Li, Z., Xu, M., Zhang, H., Zhang, W., and Gao, J. (2009). Grain yield trends of different food crops under long-term fertilization in China. Sci. Agric. Sin. 42, 2407–2424.
Li, H., Yang, S., Xu, Z., Yan, Q., Li, X., van Nostrand, J. D., et al. (2017). Responses of soil microbial functional genes to global changes are indirectly influenced by aboveground plant biomass variation. Soil Biol. Biochem. 104, 18–29. doi: 10.1016/j.soilbio.2016.10.009
Liang, Y., Al-Kaisi, M., Yuan, J., Liu, J., Zhang, H., Wang, L., et al. (2021). Effect of chemical fertilizer and straw-derived organic amendments on continuous maize yield, soil carbon sequestration and soil quality in a Chinese Mollisol. Agric. Ecosyst. Environ. 314:107403. doi: 10.1016/j.agee.2021.107403
Liu, J.-F., Mbadinga, S. M., Sun, X.-B., Yang, G.-C., Yang, S.-Z., Gu, J.-D., et al. (2016). Microbial communities responsible for fixation of CO2 revealed by using mcrA, cbbM, cbbL, fthfs, fefe-hydrogenase genes as molecular biomarkers in petroleum reservoirs of different temperatures. Int. Biodeterior. Biodegradation 114, 164–175. doi: 10.1016/j.ibiod.2016.06.019
Liu, B., Xia, H., Jiang, C., Riaz, M., Yang, L., Chen, Y., et al. (2022). 14 year applications of chemical fertilizers and crop straw effects on soil labile organic carbon fractions, enzyme activities and microbial community in rice-wheat rotation of middle China. Sci. Total Environ. 841:156608. doi: 10.1016/j.scitotenv.2022.156608
Luo, M., Tian, D., Gao, M., and Huang, R. (2018). Soil organic carbon of purple soil as affected by different application of biochar. Environ. Sci. 39, 4327–4337. doi: 10.13227/j.hjkx.201711067
Ma, H., Li, L., Liu, S., Shi, W., Wang, C., Zhao, Q., et al. (2022). Physiological response, phytohormone signaling, biomass production and water use efficiency of the CAM plant Ananas comosus under different water and nitrogen regimes. Agric. Water Manag. 266:107563. doi: 10.1016/j.agwat.2022.107563
Padhy, S. R., Bhattacharyya, P., Dash, P. K., Nayak, S. K., Parida, S. P., Baig, M. J., et al. (2022). Elucidation of dominant energy metabolic pathways of methane, Sulphur and nitrogen in respect to mangrove-degradation for climate change mitigation. J. Environ. Manag. 303:114151. doi: 10.1016/j.jenvman.2021.114151
Ren, G., Zhang, X., Xin, X., Yang, W., Zhu, A., Yang, J., et al. (2023). Soil organic carbon and nitrogen fractions as affected by straw and nitrogen management on the North China plain. Agric. Ecosyst. Environ. 342:108248. doi: 10.1016/j.agee.2022.108248
Schleuss, P.-M., Widdig, M., Heintz-Buschart, A., Guhr, A., Martin, S., Kirkman, K., et al. (2019). Stoichiometric controls of soil carbon and nitrogen cycling after long-term nitrogen and phosphorus addition in a Mesic grassland in South Africa. Soil Biol. Biochem. 135, 294–303. doi: 10.1016/j.soilbio.2019.05.018
Snyder, C. S., Bruulsema, T. W., Jensen, T. L., and Fixen, P. E. (2009). Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agric. Ecosyst. Environ. 133, 247–266. doi: 10.1016/j.agee.2009.04.021
Tang, S., Ma, Q., Marsden, K. A., Chadwick, D. R., Luo, Y., Kuzyakov, Y., et al. (2023). Microbial community succession in soil is mainly driven by carbon and nitrogen contents rather than phosphorus and Sulphur contents. Soil Biol. Biochem. 180:109019. doi: 10.1016/j.soilbio.2023.109019
Treseder, K. K., Berlemont, R., Allison, S. D., and Martiny, A. C. (2018). Nitrogen enrichment shifts functional genes related to nitrogen and carbon acquisition in the fungal community. Soil Biol. Biochem. 123, 87–96. doi: 10.1016/j.soilbio.2018.05.014
Val-Moraes, S. P., De Macedo, H. S., Kishi, L. T., Pereira, R. M., Navarrete, A. A., Mendes, L. W., et al. (2016). Liming in the sugarcane burnt system and the green harvest practice affect soil bacterial community in northeastern so Paulo, Brazil. Antonie Van Leeuwenhoek 109, 1643–1654. doi: 10.1007/s10482-016-0764-8
Wang, S., Ding, L., Liu, W., Wang, J., and Qian, Y. (2021). Effect of plastic mulching on soil carbon and nitrogen cycling-related bacterial community structure and function in a dryland spring maize field. Agriculture 11, 1–13. doi: 10.3390/agriculture11111040
Wang, J., Fu, X., Ghimire, R., Sainju, U. M., Jia, Y., and Zhao, F. (2021). Responses of soil bacterial community and enzyme activity to organic matter components under long-term fertilization on the loess plateau of China. Appl. Soil Ecol. 166:103992. doi: 10.1016/j.apsoil.2021.103992
Wang, H., Hu, G., Xu, W., Boutton, T. W., Zhuge, Y., and Bai, E. (2018). Effects of nitrogen addition on soil organic carbon mineralization after maize stalk addition. Eur. J. Soil Biol. 89, 33–38. doi: 10.1016/j.ejsobi.2018.10.002
Wang Xiaoqi, S. L., Yafeng, J., Wei, T., Qianqian, L., and Lei, M. (2017). Effect of addition of several types of straws on N2O and CH4 emissions from soil under flooding condition in Hainan Province. Acta Sci. Circumst. 37, 4004–4010.
Wu, Z., Li, H., Liu, Q., Ye, C., and Yu, F. (2020). Application of bio-organic fertilizer, not biochar, in degraded red soil improves soil nutrients and plant growth. Rhizosphere 16:100264. doi: 10.1016/j.rhisph.2020.100264
Wu, P., Liu, F., Zhao, Y., Bai, Y., Feng, B., Li, Y., et al. (2023). Diffusion and transformation of methane within the soil profile and surface uptake in dryland spring maize fields under different fertilizer application depths. Agric. Ecosyst. Environ. 344:108305. doi: 10.1016/j.agee.2022.108305
Wu, D., Ren, C., Ren, D., Tian, Y., Li, Y., Wu, C., et al. (2023). New insights into carbon mineralization in tropical paddy soil under land use conversion: coupled roles of soil microbial community, metabolism, and dissolved organic matter chemodiversity. Geoderma 432:116393. doi: 10.1016/j.geoderma.2023.116393
Xie, L., Chen, J., Wang, R., and Zhou, Q. (2012). Effect of carbon source and COD/NO3−–N ratio on anaerobic simultaneous denitrification and methanogenesis for high-strength wastewater treatment. J. Biosci. Bioeng. 113, 759–764. doi: 10.1016/j.jbiosc.2012.01.007
Yadav, A. N., Kour, D., Kaur, T., Devi, R., Yadav, A., Dikilitas, M., et al. (2021). Biodiversity, and biotechnological contribution of beneficial soil microbiomes for nutrient cycling, plant growth improvement and nutrient uptake. Biocatal. Agric. Biotechnol. 33:102009. doi: 10.1016/j.bcab.2021.102009
Yao, Q., Li, Z., Song, Y., Wright, S. J., Guo, X., Tringe, S. G., et al. (2018). Community proteogenomics reveals the systemic impact of phosphorus availability on microbial functions in tropical soil. Nat. Eco. Evol. 2, 499–509. doi: 10.1038/s41559-017-0463-5
Ye, L. F., Liu, H. Y., Dan Deng, H., Zheng, Y. P., Han, Y. W., Gao, X. T., et al. (2023). Effects of decadal nitrogen and phosphorus fertilization on microbial taxonomic and functional attributes associated with soil organic carbon decomposition and concentration in an alpine meadow. Ecol. Indic. 146:109790. doi: 10.1016/j.ecolind.2022.109790
Yuan, J., Yuan, Y., Zhu, Y., and Cao, L. (2018). Effects of different fertilizers on methane emissions and methanogenic community structures in paddy rhizosphere soil. Sci. Total Environ. 627, 770–781. doi: 10.1016/j.scitotenv.2018.01.233
Zhang, Z.-H., Nie, J., Liang, H., Wei, C.-L., Wang, Y., Liao, Y.-L., et al. (2022). The effects of co-utilizing green manure and rice straw on soil aggregates and soil carbon stability in a paddy soil in South China. J. Integr. Agric. doi: 10.1016/j.jia.2022.09.025
Zhang, Y. S., Wang, Y., Su, S., Bai, L., Wu, C., and Zeng, X. (2021). Long-term manure application enhances the stability of aggregates and aggregate-associated carbon by regulating soil physicochemical characteristics. Catena 203:105342. doi: 10.1016/j.catena.2021.105342
Zhang, W. K. H., and Zhang, R. (2020). Research progress of SOC function and transformation mechanisms. Sci. Agric. Sin. 53, 317–331.
Keywords: C fixation, C degradation, CH4 metabolism, long-term fertilization, metagenomics, maize rhizosphere soil
Citation: Li Y, Wang C, Chang H, Zhang Y, Liu S and He W (2023) Metagenomics reveals the effect of long-term fertilization on carbon cycle in the maize rhizosphere. Front. Microbiol. 14:1170214. doi: 10.3389/fmicb.2023.1170214
Edited by:
Upendra Kumar, National Rice Research Institute (ICAR), IndiaReviewed by:
Tin Mar Lynn, Ministry of Education, MyanmarCopyright © 2023 Li, Wang, Chang, Zhang, Liu and He. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shuxia Liu, bGl1c2h1eGlhNjlAMTYzLmNvbQ==; Wentian He, d2VudGlhbl9oZUBob3RtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.