
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 28 April 2023
Sec. Food Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1150425
 Alexander Wenger1,2
Alexander Wenger1,2 Cornelia Bär1
Cornelia Bär1 Reto Portmann1
Reto Portmann1 Remo S. Schmidt1
Remo S. Schmidt1 Elisabeth Eugster3
Elisabeth Eugster3 Laure Weisskopf2
Laure Weisskopf2 Stefan Irmler1*
Stefan Irmler1*The function of the aminotransferase Aat (GenBank Protein WP_159211138) from Pediococcus acidilactici FAM 18098 was studied in vivo. For this purpose, the gene was replaced with an erythromycin resistance gene using the temperature-sensitive Escherichia coli-Pediococcus shuttle plasmid pSET4T_Δaat. The knockout was verified by PCR and genome sequencing. Subsequently, the differences between the metabolism of the knockout and of the wild-type strain were investigated by determining the free amino acids and organic acids in culture supernatants. It was found that the knockout mutant no longer synthesized 3-phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (HPLA). Additionally, the mutant strain no longer catabolized phenylalanine. Metabolic pathway analysis using the KEGG database indicate that P. acidilactici cannot synthesize α-ketoglutarate that is a predominant amino-group acceptor in many transamination reactions. To study the transfer of the amino group of phenylalanine, the wild-type strain was incubated with [15N] phenylalanine. Mass spectrometry showed that during fermentation, [15N] alanine was formed, indicating that pyruvic acid is an amino group acceptor in P. acidilactici. The present study shows that Aat plays a crucial role in PLA/HPLA biosynthesis and pyruvic acid is an amino acceptor in transamination reactions in P. acidilactici.
Fermentation has been used for hundreds of years to preserve food. Many fermentations rely on the metabolic activity of lactic acid bacteria. These bacteria form lactic acid from sugar and thus lower the pH of the food, resulting in a longer shelf life. Nowadays, when lactic acid bacteria are selected for fermentation, it is not only important that they perform a fast acidification but also that they possess specific properties. These include, for example, the formation of aroma substances and/or antimicrobial substances.
Starter cultures are used for controlled acidification during cheese manufacture. During cheese ripening, non-starter lactic acid bacteria then start to grow (Beresford and Williams, 2004). They are a significant component of the cheese microbiome at the end of ripening and can affect cheese quality both positively and negatively. Amongst others, Pediococcus acidilactici can be part of the non-starter lactic acid bacteria population.
We are interested in the amino acid metabolism of P. acidilactici since it exhibits properties that can influence cheese quality. For example, it has been reported that it can produce 3-phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (HPLA) from phenylalanine and tyrosine, respectively (Mu et al., 2012b). Both compounds, PLA and HPLA, exhibit antifungal activity (Lavermicocca et al., 2000). Additionally, it possesses the genes of the arginine deiminase pathway, which converts arginine to ornithine (Eugster et al., 2019; Wenger et al., 2020). This metabolic pathway releases carbon dioxide which could influence the eye formation in cheese. Finally, P. acidilactici can degrade serine and threonine and, in turn, form alanine and α-aminobutyric acid in medium and cheese (Eugster et al., 2019; Wenger et al., 2020). We assume that serine and threonine are converted to alanine and α-aminobutyric acid, respectively. Our hypothesis is that a dehydratase metabolizes serine and threonine to α-keto acids, which are substrates for both aminotransferases and dehydrogenases.
When we searched the genome sequence of P. acidilactici strain FAM 18098 for genes encoding aminotransferases, an aminotransferase class I/II-fold pyridoxal phosphate-dependent enzyme (GenBank Protein WP_159211138) was identified that may be involved in the formation of α-aminobutyric acid and alanine (Wenger et al., 2020). In order to analyze the enzymatic activity of this aminotransferase, the gene was cloned and overexpressed in Escherichia coli. We found that the purified aminotransferase exhibited activity toward various amino acids such as leucine, methionine, α-aminobutyric acid, alanine, cysteine, and phenylalanine. Because of the broad substrate specificity, we could not clearly deduce whether the aminotransferase plays a role in the formation α-aminobutyric acid or alanine in vivo.
The aim of this work was to study the role of aminotransferase Aat in vivo. For this purpose, the gene was inactivated in FAM 18098 by gene replacement. The composition of the organic acids and amino acid in the culture supernatants of the knockout strain was compared with those of the wild-type strain. When it became apparent that the mutant strain no longer synthesized PLA from phenylalanine, further studies were performed with [15N]-labeled phenylalanine to identify the in vivo amino group acceptors.
The bacterial strains used in this study are listed in Table 1. P. acidilactici strains were cultivated in MRS medium (de Man et al., 1960) at 30°C.
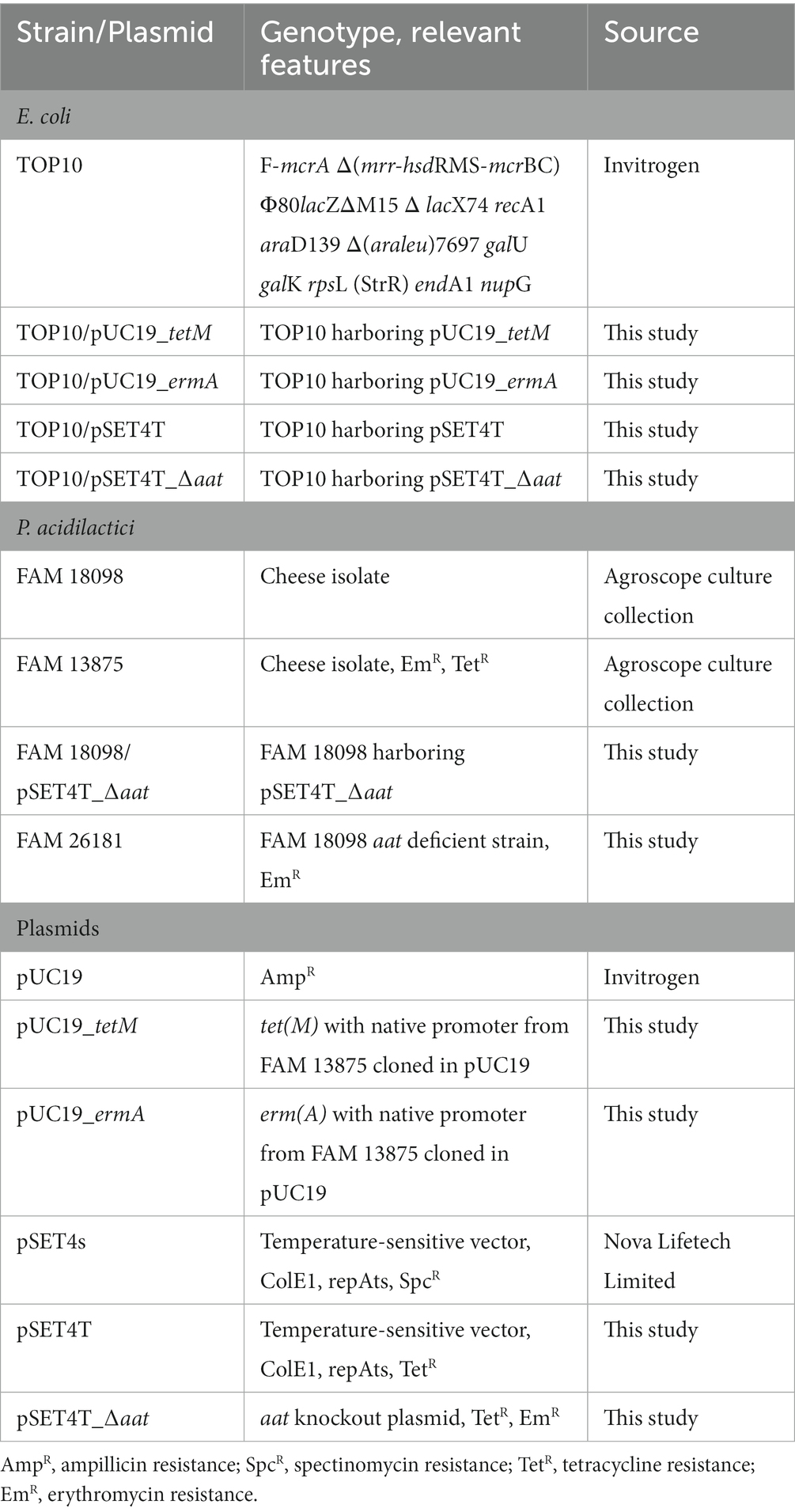
Table 1. Strains and plasmids used in this study.
To study amino acid metabolism, strains were grown at 30°C for 4 days in basal medium (pH 7.0 ± 0.2) that consisted of di-potassium hydrogen phosphate anhydrous (9 g L−1), yeast extract (5 g L−1), casein hydrolysate (2 g L−1), magnesium sulfate heptahydrate (0.2 g L−1), manganese dichloride tetrahydrate (0.2 g L−1), D-galactose (2 g L−1), 5 mM of L-serine, and 5 mM of L-threonine.
To study the transfer of the amino group of phenylalanine, P. acidilactici strain FAM 18098 was grown at 30°C for 3 days in MRS medium supplemented with 5 mM of [15N] L-phenylalanine (Cambridge Isotope Laboratories, Tewksbury, MA, United States).
Escherichia coli strains were grown in LB medium (Sambrook et al., 1989), with shaking (220 rpm) at 37°C. The LB medium was supplemented with ampicillin (100 μg mL−1) or tetracycline (50 μg mL−1), if necessary.
Genomic DNA (gDNA) was extracted as described by Berthoud et al. (2017), and DNA concentration was determined using the Qubit dsDNA BR Assay Kit (Thermo Fisher Scientific, Reinach, Switzerland). Plasmids were isolated using the FastGene Plasmid Mini Kit (Nippon Genetics Europe, Düren, Germany). Restriction and ligation were performed using the Invitrogen Anza Restriction Enzyme Cloning System (Thermo Fisher Scientific). One Shot TOP10 Chemically Competent E. coli (Thermo Fisher Scientific) were used for the transformation of E. coli.
The primers were designed using the Primer3Plus tool (Untergasser et al., 2012) and synthesized by Microsynth AG (Balgach, Switzerland). Their sequences are listed in Table 2. The polymerase chain reactions (PCR) were performed using the Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific) following the manufacturer’s instructions and used in all PCR reactions. The amplicons were purified using the FastGene Gel/PCR Extraction Kit (Nippon Genetics Europe). Their concentrations were determined using a NanoDrop Spectrophotometer (Thermo Fisher Scientific).
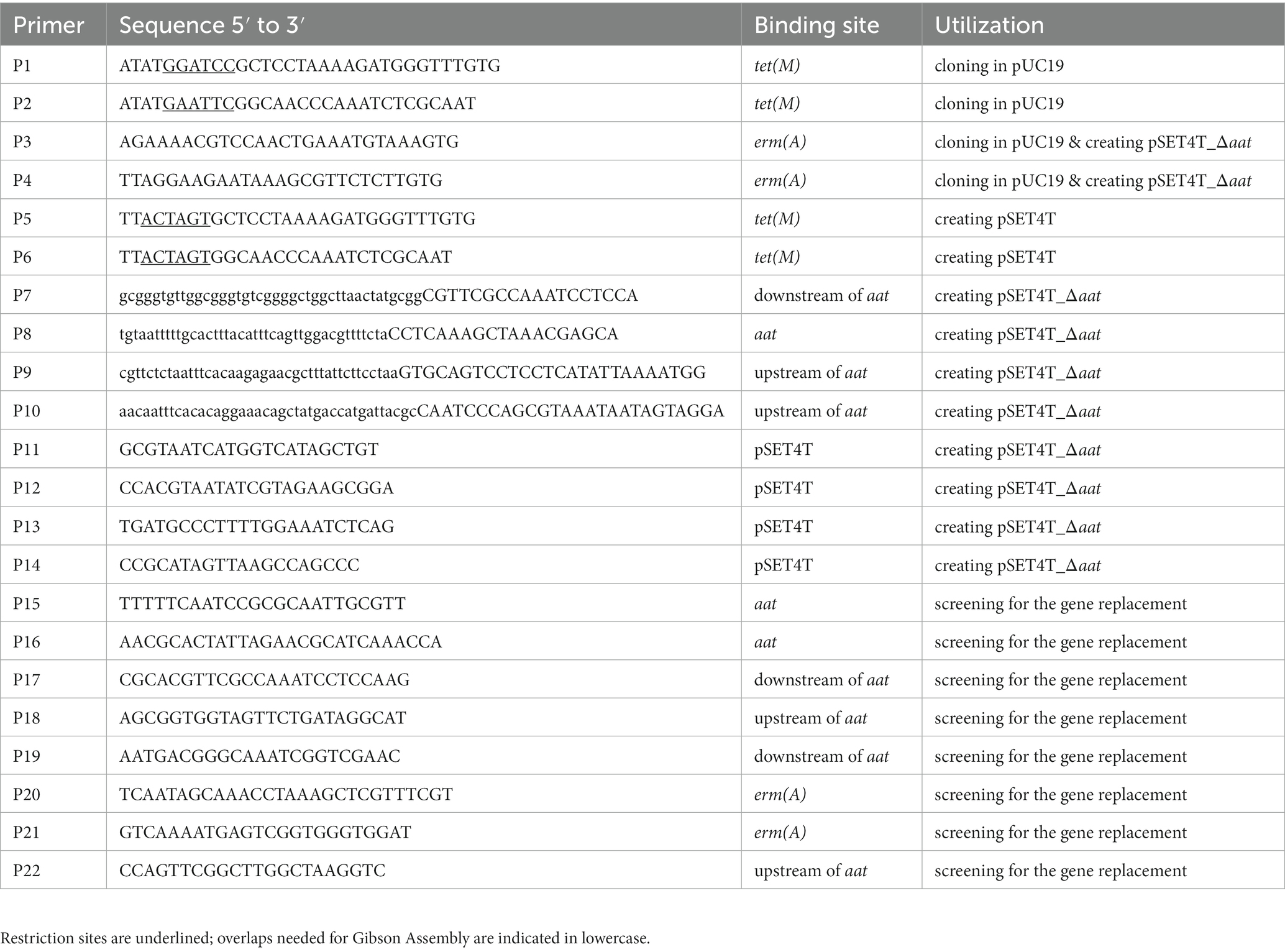
Table 2. Primers used in this study.
The workflow for the construction of pSET4T_∆aat is shown in Figure 1. The gene tet(M) (GenBank Nucleotide GBP49_00310) including the promoter region was amplified from the gDNA of P. acidilactici strain FAM 13875 using primers P1/P2. The amplicon was purified then digested with BamHI and EcoRI and ligated in the BamHI- and EcoRI-digested pUC19, resulting in pUC19_tetM. E. coli TOP10 were then transformed with the ligation reaction, and transformants were selected on LB agar containing tetracycline.
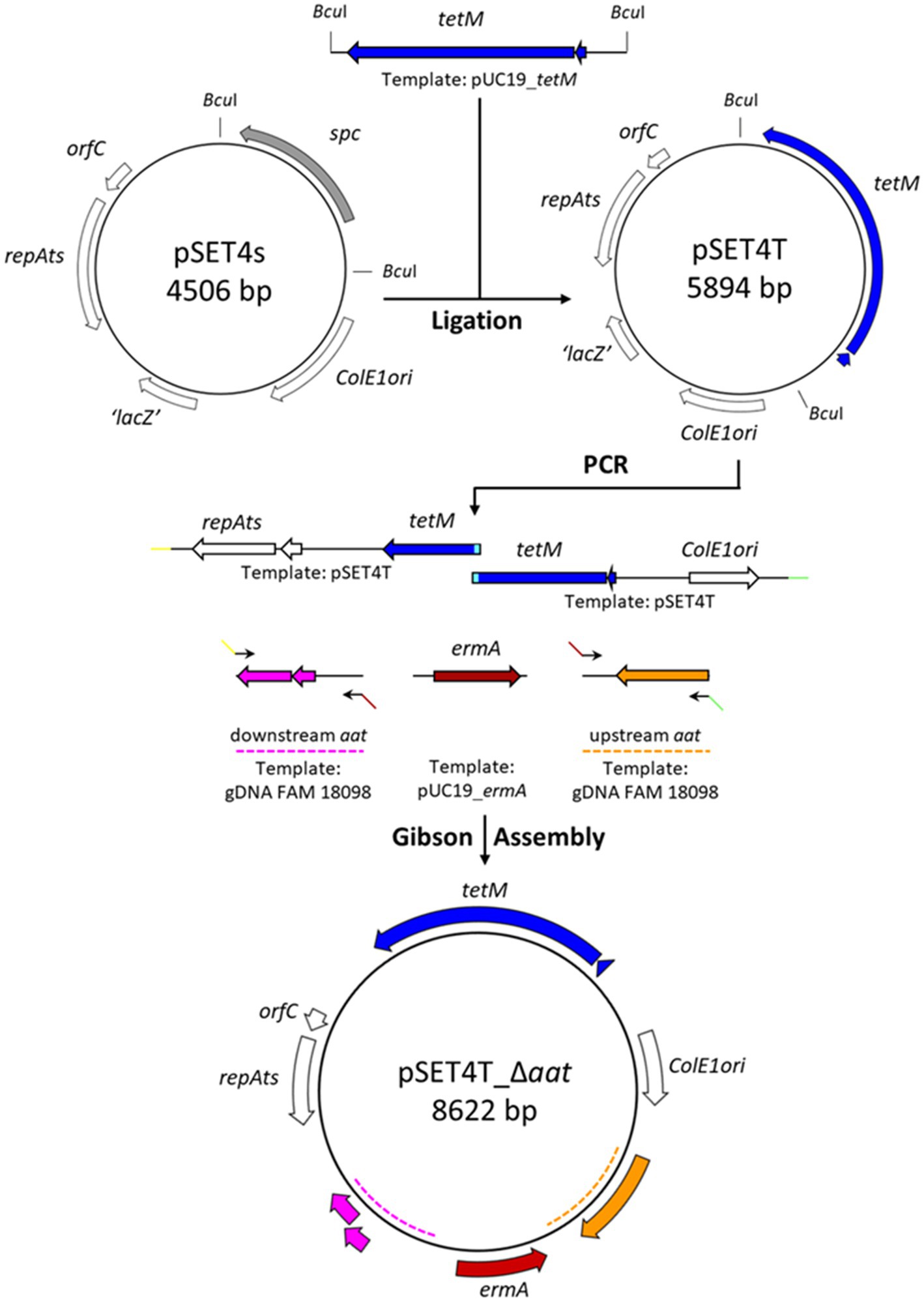
Figure 1. Schematic representation of the construction of the gene knockout plasmid pSET4T_Δaat. Tetracycline resistance gene tetM was amplified by PCR from pUC19_tetM vector and ligated into pSET4S using the two BcuI sites creating pSET4T. The gene replacement vector pSET4T_Δaat was constructed by PCR amplification of the plasmid backbone of pSET4T with two overlapping fragments (indicated in light blue). The regions 1-kbp upstream and 1-kbp downstream of aat were amplified from the genomic DNA of P. acidilactici strain FAM 18098 using overhang primers with homologous sequences of the other fragments (indicated by colored arrows). The erythromycin resistance gene ermA was amplified via PCR from pUC19_ermA. The five fragments were assembled using a Gibson Assembly resulting in pSET4T_Δaat. The wide arrows indicate open reading frames.
The gene erm(A) (GenBank Nucleotide GBP49_00290) including the promotor region was amplified from the gDNA of P. acidilactici strain FAM 13875 using primers P3/P4 and ligated to SmaI-digested pUC19. The ligation reaction was transformed into E. coli TOP10, which were then plated on LB agar supplemented with ampicillin and X-Gal. Transformants were selected by blue/white screening. Sanger sequencing was used to check the orientation of the insert.
Tetracycline resistance gene tet(M) was amplified from the pUC19_tetM plasmid using the primers P5/P6. The amplicon was digested by BcuI and then ligated into BcuI-digested pSET4S. After transformation into E. coli TOP10, transformants were selected on LB agar containing tetracycline. The resulting plasmid was named pSET4T.
The final gene replacement vector pSET4T_∆aat was created using a Gibson Assembly reaction. Therefore, the 1-kbp upstream and downstream regions of aat were amplified using the primers P7/P8 and P9/P10, respectively, from the gDNA of strain FAM 18098. The erm(A) gene was amplified with the primers P3/P4 from pUC19_ermA. Furthermore, two fragments of pSET4T were amplified using the primers P11/P12 and P13/P14. The five amplicons were purified and then assembled using the GeneArt™ Gibson Assembly® HiFi Cloning Kit (Thermo Fisher Scientific, Reinach, Switzerland) following the manufacturer’s instructions. The reaction mix was transformed into E. coli TOP10, and transformants were selected on LB agar containing tetracycline. The composition of the plasmids isolated from several transformants was analyzed by restriction analysis and PCR combined with Sanger sequencing. A correctly assembled plasmid was named pSET4T_∆aat and propagated and maintained in E. coli TOP10.
Using the protocol described by Caldwell et al. (1996), pSET4T_∆aat (220 ng) was transformed into P. acidilactici FAM 18098 by electroporation. Transformants were selected on MRS plates supplemented with erythromycin (5 μg mL−1) and grown at 30°C for 48 h. A transformant colony was then cultured in MRS medium containing erythromycin at 42°C to stop plasmid replication. After 24 h of growth, transformants were reinoculated into fresh MRS medium supplemented with erythromycin, creating a new culture. Colonies were obtained by diluting a sample of each culture by a factor of 10−6 and 10−7 with NA (0.8% sodium chloride, 0.1% peptone from casein, pancreatic digest), which was then plated on MRS agar containing erythromycin. Each time, 50 colonies were transferred with a sterile toothpick onto a new MRS agar plate containing erythromycin. After incubating for 24–48 h, colonies were replica plated onto an MRS agar plate containing tetracycline (100 μg mL−1). Colonies exhibiting resistance against erythromycin and sensitivity toward tetracycline were analyzed for the loss of pSET4T_∆aat and the replacement of aat using PCR with the primers P15–P22 (Table 2).
The gDNA of a mutant of interest (strain FAM 26181) was sequenced using Illumina Novaseq technology at the Next Generation Sequencing Platform of the University of Bern. The Illumina reads were trimmed using Trimmomatic (version 0.38, options: SLIDINGWINDOW:4:8 MINLEN:127) (Bolger et al., 2014). The trimmed reads were mapped to the genome sequence of Pediococcus acidilactici FAM 18098 (GenBank Assembly ASM980809v1) using bowtie2 (Langmead and Salzberg, 2012). Samtools (version 1.16.1) (Danecek et al., 2021) was then used to sort and index the reads and to identify regions with zero coverage. Furthermore, the trimmed reads were assembled using SPAdes (version 3.6.1, options: -careful -mismatch-correction -k 21,33,55,77,99,127). The assembly was deposited in the GenBank database under the accession number ASM2453963v1.
Free amino acids in culture supernatants were determined using high-performance liquid chromatography (HPLC) as previously described (Wenger et al., 2020).
Organic acids were determined by HPLC using an Aminex HPX-87H column (Bio-Rad, Cressier, Switzerland). Perchloric acid was added to the culture supernatant at a final concentration of 0.5 M, and the supernatant was then filtered (Nylon 0.45 μm, Millipore Millex-HN, Merck KGaA, Darmstadt, Germany). The chromatographic conditions were as follows: column temperature = 65°C, flow rate = 0.4 mL min−1, UV detection at 210 nm, injection volume = 2 μL. Organic acids, such as oxalic acid, citric acid, malic acid, succinic acid, formic acid, acetic acid, pyruvic acid, lactic acid, α-ketoglutaric acid, α-ketobutyric acid, phenyllactic acid, and 4-hydroxyphenyllactic acid, were used as standards.
Analyses were performed using a Vanquish liquid chromatography system coupled to a Q Exactive Plus Orbitrap mass spectrometer (Thermo Fisher Scientific) and controlled by the Thermo Xcalibur software (version 4.5.474.0, Thermo Fisher Scientific). Samples were filtered before analysis (nylon, 0.45 μm, Millipore Millex-HN, Merck KGaA, Darmstadt, Germany) and then separated on an Acquity UPLC HSS T3 column (150 × 2.1 mm i.d., 1.8 μm particle size; Waters Corporation, Milford, MA, United States) at a flow rate of 400 μL min−1 at 40°C. Mobile phase A consisted of H2O containing 0.1% formic acid, and mobile phase B consisted of acetonitrile containing 0.1% formic acid. Mass spectra were recorded at a resolution of 140,000 FWHM (full width at half maximum). Peak areas were determined using the Thermo Xcalibur Qual Browser software (version 4.1.31.9, Thermo Fisher Scientific).
For the determination of PLA and HPLA, 10 μL of the sample was injected. The gradient was as follows: 5% B for 2 min; 5–95% B for 17 min; 95% B for 4 min. Mass spectra were recorded over a mass range of 66.7–300 in negative ionization mode. The mass ranges from 165.045 to 165.065 m/z and from 181.045 to 181.065 m/z were used to analyze extracted ion chromatograms for PLA and HPLA, respectively. The results shown in this study reflect the mean and standard deviation of three independently performed biological experiments.
For the detection of [15N]-labeled amino acids, a standard solution containing the amino acids alanine, aspartic acid, glutamic acid, leucine, methionine, phenylalanine, valine, and 2-aminobutyric acid (each 1 pmol μL−1) was measured in triplicate. The measurements were used to determine retention times and to verify the accurate masses of the monoisotopic [14N]-peaks and the masses of the first naturally occurring [15N]-containing isotope of the amino acids. Filtered samples with or without incubation of Pediococcus acidilactici FAM 18098 were diluted hundredfold with water, of which 1 μL was injected and separated using the following gradient: 3% B for 3 min; 3–95% B for 7 min; 95% B for 5 min. Mass spectra of the samples were recorded over a mass range of 75–300 in positive ion mode. Extracted ion chromatograms of the monoisotopic [14N]- and the [15N]-peak with a tolerance of 2 ppm were generated to determine the peak areas for the substances of interest (Supplementary Table S1).
In order to better understand the function of aat, the gene was knocked out in this study using the thermosensitive vector pET4s, which was originally constructed for gene replacements in Streptococcus suis and carries the spc gene that confers spectinomycin resistance (Takamatsu et al., 2001). Because P. acidilactici FAM 18098 is resistant to spectinomycin, we selected tetracycline and erythromycin resistance genes as markers of selection. For this purpose, the tet(M) and erm(A) gene of P. acidilactici FAM 13875, a strain that showed resistance to tetracycline and erythromycin (Lüdin et al., 2018), were cloned. The final plasmid pSET4T_Δaat consisted of two origins of replication (ColE1ori and orfC/repAts), the tet(M) gene, and the nucleotide sequences needed for the exchange of the aat gene with erm(A) (Figure 1).
After transformation of strain FAM 18098 with the knockout plasmid, transformed cells were selected with erythromycin. Initially, the colonies also showed resistance to tetracycline. After three cultivation steps, a tetracycline-sensitive colony was found. PCR analysis confirmed that the colony had lost both the knockout plasmid and the aat gene (data not shown). The culture obtained from this colony was named FAM 26181.
In order to validate the disruption of the aat gene, the gDNA of this strain was sequenced using Illumina sequencing and compared with the already existing genome sequence of the wild-type strain FAM 18098. When the Illumina reads obtained from the knockout strain were mapped to the genome assembly of the wild-type, the region from position 606,059 to 607,168 of the wild-type’s chromosome containing the aat gene was not covered (data not shown). This confirmed the results of the PCR analysis and showed that the aat gene was no longer present in the knockout strain. The analysis of the de novo assembly showed that the erm(A) gene had been integrated at this site (Figure 2).
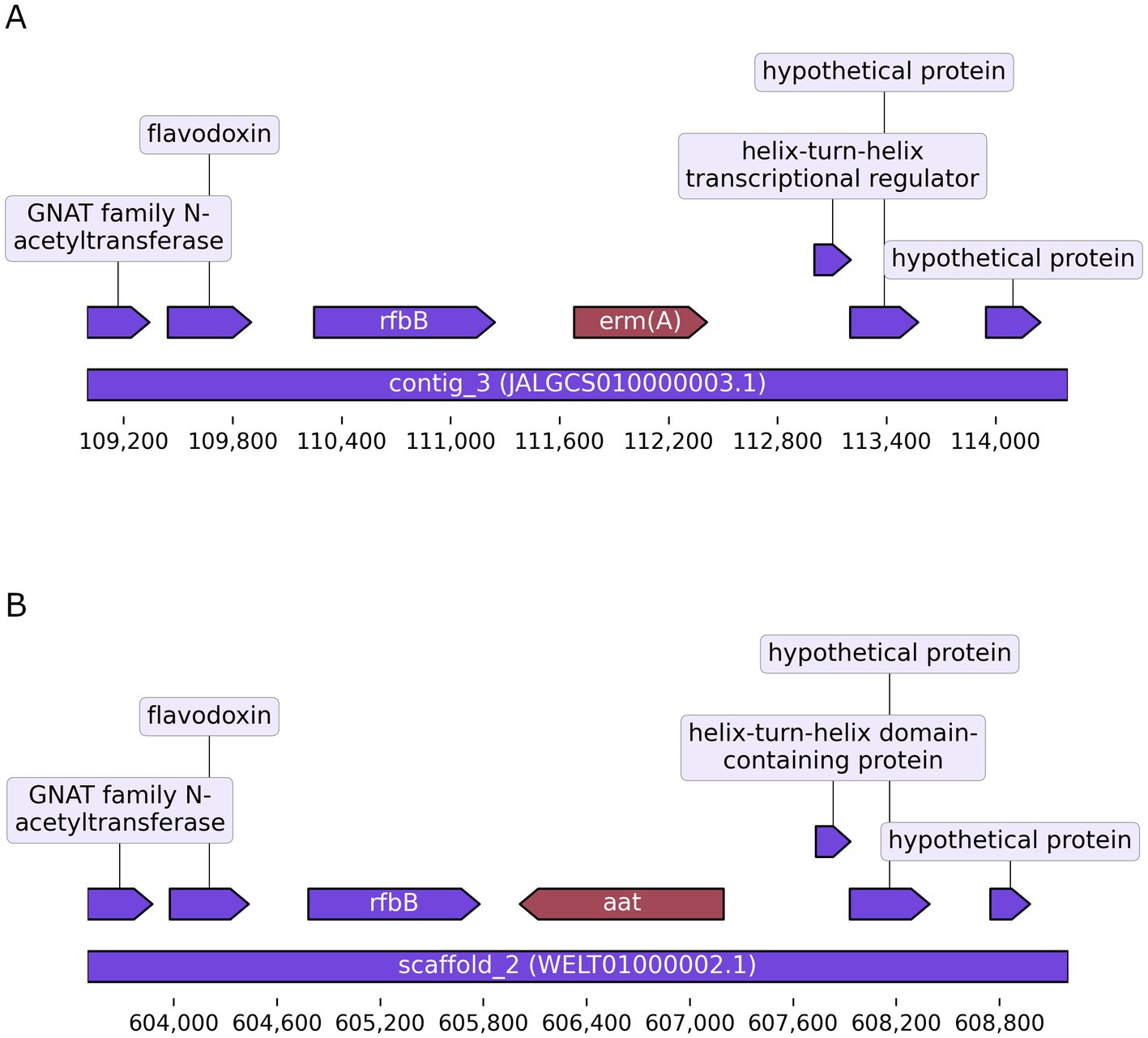
Figure 2. Gene replacement in Pediococcus acidilactici. The illustration shows the erm(A) gene that replaced the aat gene in the knockout strain (A). The corresponding region containing the aat gene in the wild-type strain is shown in the lower panel (B). Arrows represent protein-coding sequences. The rulers indicate the location in contig_3 (GenBank acc. no. JALGCS010000003.1) and chromosome (GenBank acc. no. WELT01000002.1).
Since the formation of α-aminobutyric acid by P. acidilactici can be observed in a so-called basal medium (Wenger et al., 2020), this medium was used to compare the amino acid metabolism of the aat knockout mutant with that of the wild-type strain.
When amino acid levels were determined in culture supernatants, the knockout strain was found to produce α-aminobutyric acid and alanine and to degrade serine and threonine (Figure 3). This was also true for the wild-type strain, except that the levels of α-aminobutyric acid and alanine were slightly higher than in the culture supernatant of the knockout strain. Furthermore, it was observed that both strains metabolized arginine and formed ornithine.
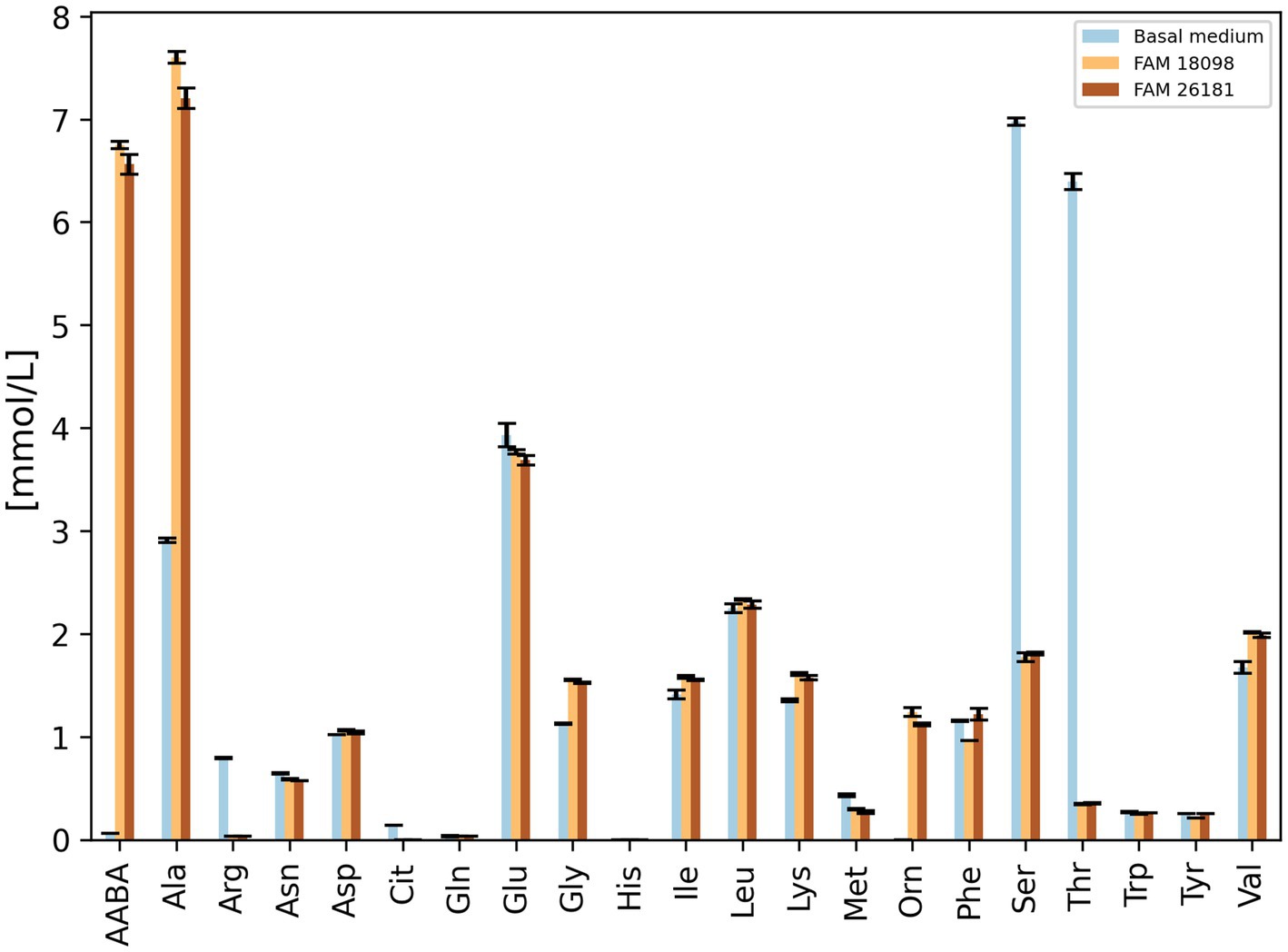
Figure 3. Amino acid concentrations in culture supernatants of wild-type strain FAM 18098 and aat knockout strain FAM 26181 after incubation for 4 days in the basal medium. Values represent the mean (±standard deviation) of two independent biological experiments. AABA: α-aminobutyric acid, Cit: citrulline, Orn: ornithine. All other amino acids are abbreviated according to IUPAC nomenclature.
It was noticeable that the phenylalanine content in the culture supernatants of the wild-type strain was considerably lower than in the basal medium and in the culture supernatants of the knockout strain (Figure 3). A similar, but less pronounced pattern was observed for tyrosine.
Since various lactic acid bacteria are able to form PLA in MRS medium (Mu et al., 2012c), this medium was used to compare the organic acid profile of the wild-type strain with that of the knockout strain.
Comparison of the organic acid chromatograms showed that two peaks were missing in the culture supernatant of the knockout strain (Figure 4). The first peak eluted with a retention time consistent with that of HPLA. The retention time of the second peak was consistent with that of PLA.
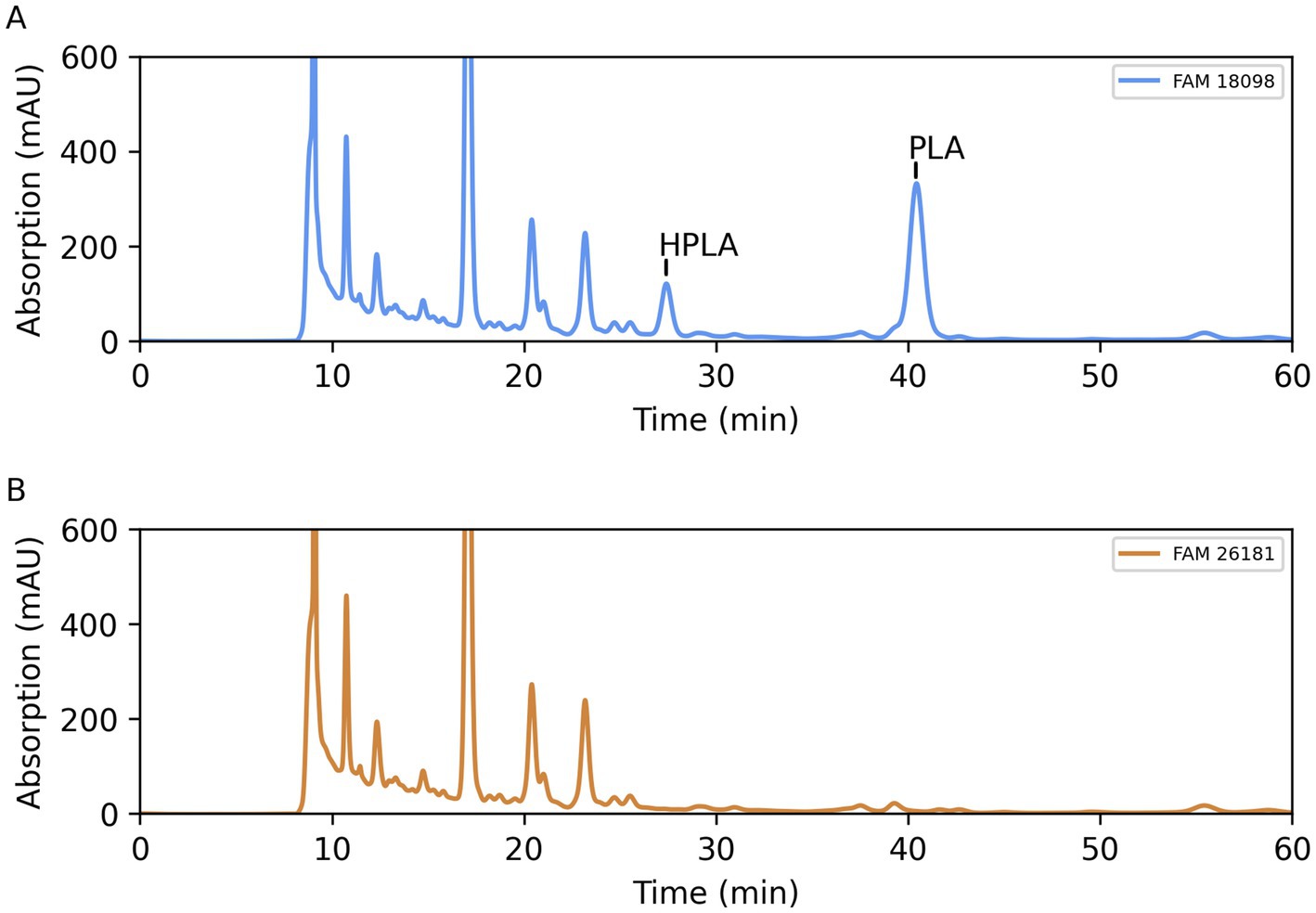
Figure 4. Organic acid profile of culture supernatants of wild-type strain FAM 18098 (A) and of aat knockout strain FAM 26181 (B) after incubation for 3 days in MRS medium. PLA: 3-phenyllactic acid, HPLA: 4-hydroxyphenyllactic acid.
In order to verify the identity of the two substances, a mass spectrometry method was established to specifically and quantitatively determine PLA and HPLA in the culture supernatants. Using standard curves generated with the pure substances, it was found that wild-type strain FAM 18098 produced 1.19 ± 0.11 mmol/L PLA and 0.38 ± 0.04 mmol/L HPLA in MRS medium, while neither substance was detected in the medium or in the culture supernatants of FAM 26181.
Since wild-type strain catabolizes phenylalanine, as described above, we investigated whether the amino group of phenylalanine is transferred to an α-keto acid. This would indicate that an aminotransferase is involved in the degradation of phenylalanine. For this purposse, [15N]-labeled phenylalanine was added to the medium and fermented with P. acidilactici FAM18098.
The following amino group acceptors were considered: pyruvic acid, α-ketobutyric acid, oxaloacetic acid, α-ketoglutaric acid, α-ketoisocaproic acid, α-ketomethylthiobutyric acid, and α-ketoisovaleric acid. If any of these acceptors were to receive the 15 N-labeled amino group of phenylalanine, [15N]-alanine, [15N]-α-aminobutyric acid, [15N]-aspartic acid, [15N]-glutamic acid, [15N]-leucine, [15N]-methionine, or [15N]-valine would be formed accordingly.
Of the amino acids mentioned, [15N]-α-aminobutyric acid was too low in concentration to be detected reliably in the medium or in the culture supernatant of FAM 18098. After fermentation with P. acidilactici FAM 18098, the levels of the unlabeled amino acids aspartic acid, methionine, and valine were found to be higher in the medium (Figure 5). No changes were observed for the unlabeled amino acids alanine, glutamic acid, leucine while phenylalanine was reduced. This pattern was also observed for the [15N]-labeled amino acids, except for alanine, which was clearly increased in the [15N]-labeled substance (Figure 5).
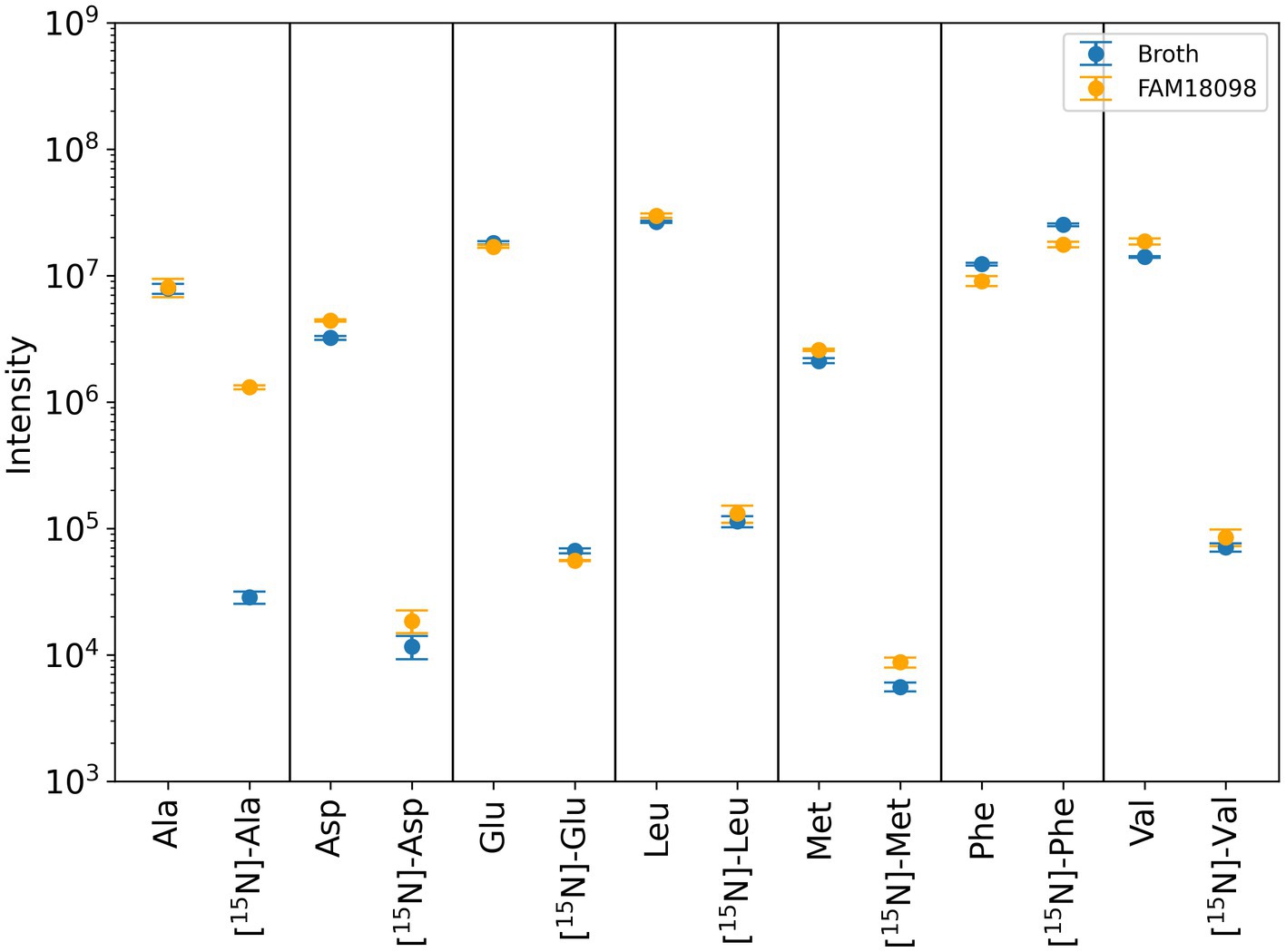
Figure 5. Determination of unlabeled and [15N]-labeled amino acids using mass spectrometry. Blue and orange dots represent the values determined in unfermented medium and medium fermented with P. acidilactici FAM 18098, respectively. Values represent the mean of three independent biological experiments (±standard deviation). Ala, alanine; Asp, aspartic acid; Glu, glutamic acid; Leu, leucine; Met, methionine; Phe, phenylalanine; Val, valine.
Various biochemical changes take place in cheese during ripening. Among others, amino acids are released from the caseins during proteolysis, which are then further metabolized by bacteria. Transamination is one of the major steps for amino acid conversion. For example, Rijnen et al. (2003) showed that the knockout of transaminases in Lactococcus lactis impacts flavor formation in cheese.
Pediococcus acidilactici can also be found in cheese. Interestingly, this species forms α-aminobutyric acid and alanine (Eugster et al., 2019; Wenger et al., 2020). The enzymes involved in the formation of these substances are not yet known, and it is not inconceivable that a transaminase plays a role here. The analysis of the genome of P. acidilactici FAM 18098 (GenBank Assembly ASM980809v1) showed that this strain has six genes encoding aminotransferases (Wenger et al., 2020). For one of these six—which encodes the protein WP_159211138 and is named aat in this study—no function could be predicted from sequence analysis. Characterization of the gene product in vitro showed that the enzyme exhibits a broad substrate specificity by converting glutamic acid, leucine, methionine, phenylalanine, alanine, and α-aminobutyric acid (Wenger et al., 2020).
The broad substrate specificity of Aat did not allow us to assign it to a metabolic pathway. Therefore, a major objective of the present study was to disrupt the gene and investigate whether the knockout has an effect on α-aminobutyric acid and alanine biosynthesis.
For the knockout, a plasmid was constructed that replicates in E. coli and P. acidilactici. The procedure presented in this study has the advantage that a knockout plasmid can be produced from five PCR fragments. The disadvantage of the methodology is that the knockout leads to an integration of an antibiotic resistance gene, so that the modified strains can only be used for laboratory purposes.
After confirming by PCR and whole genome sequencing that we had isolated a strain having lost the aat gene, the phenotypic characteristics of the wild-type strain were compared with the knockout strain. It was expected that the removal of an aminotransferase would affect amino acid and organic acid metabolism. Therefore, the composition of amino acids and organic acids was studied in the culture supernatants of the wild-type and knockout strains.
The comparison of the amino acid levels in the culture supernatants showed that the mutant and wild-type strains differed with respect to alanine, phenylalanine, and tyrosine. No differences were found for leucine, methionine, and α-aminobutyric acid —amino acids which Aat was able to convert in vitro (Wenger et al., 2020). It was concluded that Aat plays a role in vivo, mainly in the metabolism of alanine and the aromatic amino acids. This was confirmed when organic acid analysis revealed that the knockout mutant no longer synthesized PLA and HPLA (Figure 4).
Rajanikar et al. (2021) reviewed in detail two biosynthetic pathways for PLA known in lactic acid bacteria. In the first case, named “de novo pathway,” glucose is the precursor for the biosynthesis of PLA and does not require the activity of an aminotransferase. In the second case, called the “core pathway,” phenylalanine is the precursor. Phenylalanine is thereby converted into phenylpyruvic acid by the action of an aminotransferase. Phenylpyruvic acid is subsequently reduced to PLA by a dehydrogenase (Figure 6).
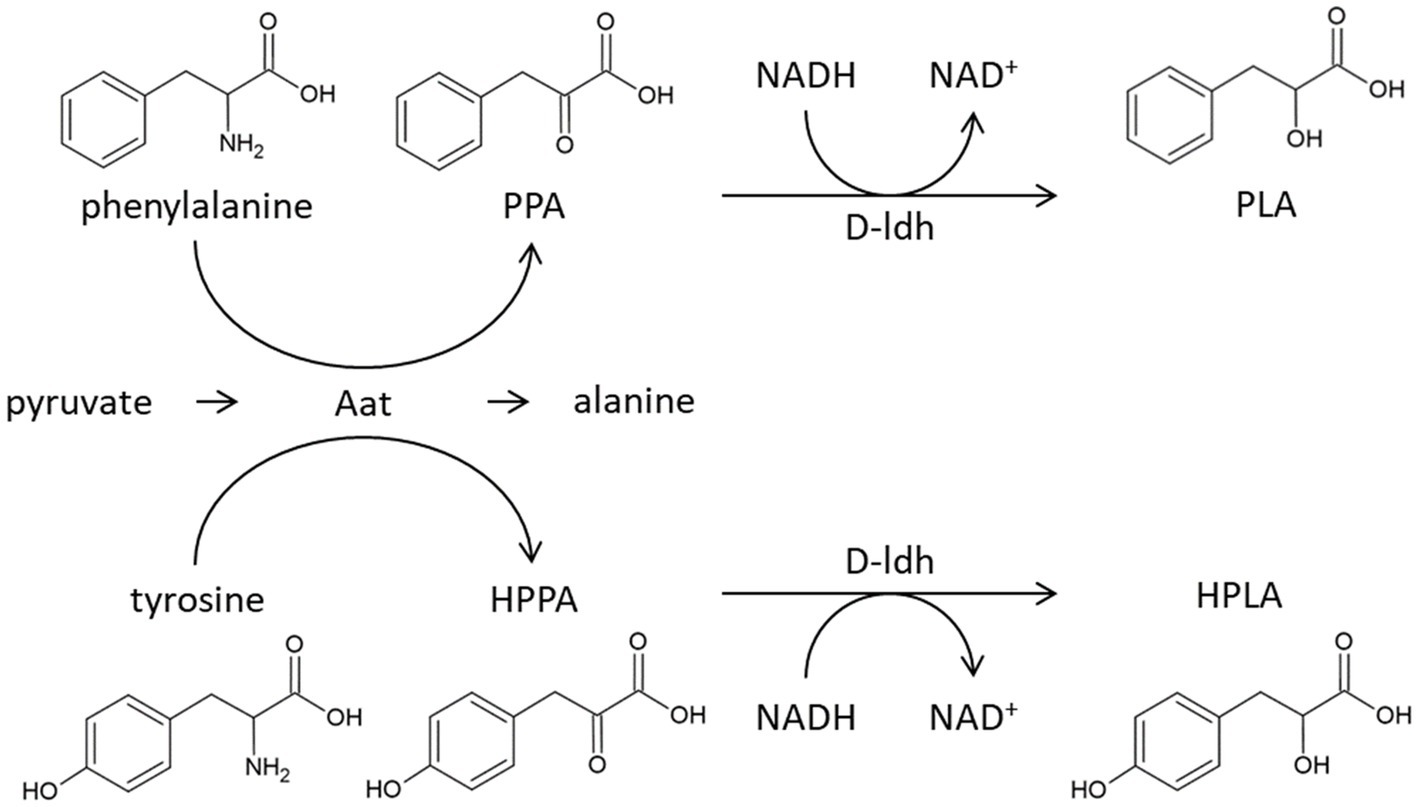
Figure 6. Proposed synthesis of 3-phenyllactic acid and 4-hydroxyphenyllactic acid in Pediococcus acidilactici. Firstly, phenylalanine and tyrosine are deaminated through the aminotransferase Aat (GenBank Protein WP_159211138), forming phenylpyruvic acid (PPA) and 4-hydroxyphenylpyruvic acid (HPPA), respectively. Then the keto acids are reduced by the D-lactate dehydrogenase (GenBank Protein WP_152689015) reaction, forming 3-phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (HPLA).
Since PLA and HPLA production was no longer detected in the knockout mutant, it can be concluded that the two substances are mainly formed via the “core pathway” and the aat gene catalyzes the first step (Figure 6). This conclusion is supported by the study of Mu et al. (2012b), which showed that PLA and HPLA biosynthesis in P. acidilactici can be increased by the addition of phenylalanine and tyrosine, respectively.
The second step, the conversion of phenylpyruvic acid to PLA, is linked to a lactate dehydrogenase coding gene (D-ldh). This is based on the findings of Mu et al. (2012a), who showed that the heterologously expressed D-ldh reduced pyruvic acid to PLA.
There is increasing evidence showing that the genes of proteins that interact functionally are often located close to each other in the genome (Bhatt et al., 2018). However, this does not apply to aat and D-ldh, which are separated from each other by about 215,000 base pairs in the genome of FAM 18098. In addition to this fact, both enzymes exhibit broad substrate specificity. This suggests that the two enzymes not only play a role in PLA/HPLA biosynthesis but probably have other functions as well. This could also explain why PLA is often a by-product of cellular metabolism and modifications of the core pathway genes do not necessarily lead to high levels of PLA in biotechnological applications (Rajanikar et al., 2021).
α-Ketoglutarate is the predominant amino group acceptor in many transamination reactions. The substance itself can be synthesized by glutamic acid dehydrogenase or isocitrate dehydrogenase. However, when the genome data of P. acidilactici FAM 18098 and other strains were analyzed, no genes encoding either of the two dehydrogenases were found (data not shown). This means that other amino group acceptors are involved in transamination reactions in P. acidilactici. When [15N] phenylalanine was used to analyze the transfer of nitrogen, the biosynthesis of [15N] alanine was observed, indicating that pyruvic acid is an amino group acceptor compound (Figure 5). Furthermore, it was observed that the [15N]-labeled phenylalanine was not completely catabolized. This is in line with findings from Mu et al. (2012b), who reported that P. acidilactici DSM 20284 does not convert all the available phenylalanine to PLA.
The present study shows that the production of knockout mutants is very helpful to elucidate metabolic pathways. Sequence analysis of Aat showed that it was annotated as a transaminase (EC 2.6.1.-), but depending on the search parameters and the database used, it was assigned to different EC numbers, such as EC 2.6.1.1 aspartate transaminase, EC 2.6.1.5 tyrosine transaminase, EC 2.6.1.9 histidinol-phosphate transaminase, or EC 2.6.1.57 aromatic-amino-acid transaminase. Analysis of the genetic context also did not help to assign the protein to a metabolic pathway (data not shown). When the enzyme was produced by heterologous expression, it exhibited broad substrate specificity (Wenger et al., 2020). With the knockout, it was then shown that aat plays a role in PLA/HPLA biosynthesis. As an outlook, comparative studies of cheese produced with the knockout and the wild-type strain could reveal whether other parameters, such as growth or flavor formation, are influenced by the newly characterized aminotransferase Aat.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
AW performed the experiments, analyzed the data, and wrote the first manuscript draft. RP assisted in the use and evaluation of the mass spectrometry analysis. AW, CB, and SI wrote the final manuscript. All authors provided intellectual input, reviewed the results, contributed to the discussion of the work and approved the final manuscript.
This work was supported by Open access funding by Agroscope.
We thank Eliane Binz and Tharmatha Bavan for their excellent technical support.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at:
https://www.frontiersin.org/articles/10.3389/fmicb.2023.1150425/full#supplementary-material
Beresford, T. P., and Williams, A. (2004). “The microbiology of cheese ripening” in Cheese: chemistry, physics & microbiology. eds. P. F. Fox, P. L. H. Mcsweeney, T. M. Cogan, and T. P. Guinee, vol. 1. Third edition ed (London: Elsevier Academic Press), 287–317.
Berthoud, H., Wüthrich, D., Bruggmann, R., Wechsler, D., Fröhlich-Wyder, M.-T., and Irmler, S. (2017). Development of new methods for the quantitative detection and typing of Lactobacillus parabuchneri in dairy products. Int. Dairy J. 70, 65–71. doi: 10.1016/j.idairyj.2016.10.005
Bhatt, V., Mohapatra, A., Anand, S., Kuntal, B. K., and Mande, S. S. (2018). FLIM-MAP: gene context based identification of functional modules in bacterial metabolic pathways. Front. Microbiol. 9:2183. doi: 10.3389/fmicb.2018.02183
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Caldwell, S. L., McMahon, D. J., Oberg, C. J., and Broadbent, J. R. (1996). Development and characterization of lactose-positive Pediococcus species for milk fermentation. Appl. Environ. Microbiol. 62, 936–941. doi: 10.1128/aem.62.3.936-941.1996
Danecek, P., Bonfield, J. K., Liddle, J., Marshall, J., Ohan, V., Pollard, M. O., et al. (2021). Twelve years of SAMtools and BCFtools. Gigascience 10:giab008. doi: 10.1093/gigascience/giab008
De Man, J. C., Rogosa, M., and Sharpe, M. E. (1960). A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135. doi: 10.1111/j.1365-2672.1960.tb00188.x
Eugster, E., Fuchsmann, P., Schlichtherle-Cerny, H., Bütikofer, U., and Irmler, S. (2019). Formation of alanine, α-aminobutyrate, acetate, and 2-butanol during cheese ripening by Pediococcus acidilactici FAM18098. Int. Dairy J. 96, 21–28. doi: 10.1016/j.idairyj.2019.04.001
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lavermicocca, P., Valerio, F., Evidente, A., Lazzaroni, S., Corsetti, A., and Gobbetti, M. (2000). Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 66, 4084–4090. doi: 10.1128/aem.66.9.4084-4090.2000
Lüdin, P., Roetschi, A., Wüthrich, D., Bruggmann, R., Berthoud, H., and Shani, N. (2018). Update on tetracycline susceptibility of Pediococcus acidilactici based on strains isolated from Swiss cheese and whey. J. Food Prot. 81, 1582–1589. doi: 10.4315/0362-028x.jfp-18-160
Mu, W., Yu, S., Jiang, B., and Li, X. (2012a). Characterization of D-lactate dehydrogenase from Pediococcus acidilactici that converts phenylpyruvic acid into phenyllactic acid. Biotechnol. Lett. 34, 907–911. doi: 10.1007/s10529-012-0847-1
Mu, W., Yu, S., Zhu, L., Jiang, B., and Zhang, T. (2012b). Production of 3-phenyllactic acid and 4-hydroxyphenyllactic acid by Pediococcus acidilactici DSM 20284 fermentation. Eur. Food Res. Technol. 235, 581–585. doi: 10.1007/s00217-012-1768-x
Mu, W., Yu, S., Zhu, L., Zhang, T., and Jiang, B. (2012c). Recent research on 3-phenyllactic acid, a broad-spectrum antimicrobial compound. Appl. Microbiol. Biotechnol. 95, 1155–1163. doi: 10.1007/s00253-012-4269-8
Rajanikar, R. V., Nataraj, B. H., Naithani, H., Ali, S. A., Panjagari, N. R., and Behare, P. V. (2021). Phenyllactic acid: a green compound for food biopreservation. Food Control 128:108184. doi: 10.1016/j.foodcont.2021.108184
Rijnen, L., Yvon, M., Van Kranenburg, R., Courtin, P., Verheul, A., Chambellon, E., et al. (2003). Lactococcal aminotransferases AraT and BcaT are key enzymes for the formation of aroma compounds from amino acids in cheese. Int. Dairy J. 13, 805–812. doi: 10.1016/S0958-6946(03)00102-X
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecular cloning: a laboratory manual. New York: Cold Spring Harbor Laboratory Press.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2001). Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46, 140–148. doi: 10.1006/plas.2001.1532
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3 - new capabilities and interfaces. Nucleic Acids Res. 40:e115. doi: 10.1093/nar/gks596
Keywords: Pediococcus acidilactici, aminotransferase, knockout, 3-phenyllactic acid, 4-hydroxyphenyllactic acid
Citation: Wenger A, Bär C, Portmann R, Schmidt RS, Eugster E, Weisskopf L and Irmler S (2023) The aminotransferase Aat initiates 3-phenyllactic acid biosynthesis in Pediococcus acidilactici. Front. Microbiol. 14:1150425. doi: 10.3389/fmicb.2023.1150425
Edited by:
Marc Stevens, University of Zürich, SwitzerlandReviewed by:
Michael Gänzle, University of Alberta, CanadaCopyright © 2023 Wenger, Bär, Portmann, Schmidt, Eugster, Weisskopf and Irmler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stefan Irmler, c3RlZmFuLmlybWxlckBhZ3Jvc2NvcGUuYWRtaW4uY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.