 Ignacio Jofré-Fernández
Ignacio Jofré-Fernández Francisco Matus-Baeza
Francisco Matus-Baeza Carolina Merino-Guzmán
Carolina Merino-Guzmán- 1Scientific and Technological Bioresource Nucleus (BIOREN), Universidad de La Frontera, Temuco, Chile
- 2Laboratory of Geomicrobiology, Department of Chemical Sciences and Natural Resources, Universidad de La Frontera, Temuco, Chile
- 3Laboratory of Conservation and Dynamics of Volcanic Soils, Department of Chemical Sciences and Natural Resources, Universidad de La Frontera, Temuco, Chile
- 4Network for Extreme Environmental Research (NEXER), Universidad de La Frontera, Temuco, Chile
Soil organic matter (SOM) decomposition mechanisms in rainforest ecosystems are governed by biotic and abiotic procedures which depend on available oxygen in the soil. White-rot fungi (WRF) play an important role in the primary decomposition of SOM via enzymatic mechanisms (biotic mechanism), which are linked to abiotic oxidative reactions (e.g., Fenton reaction), where both processes are dependent on reactive oxygen species (ROS) and soil pH variation, which has yet been studied. In humid temperate forest soils, we hypothesize that soil pH is a determining factor that regulates the production and consumption of ROS during biotic and abiotic SOM decomposition. Three soils from different parent materials and WRF inoculum were considered for this study: granitic (Nahuelbuta, Schizophyllum commune), metamorphic (Alerce Costero, Stereum hirsutum), and volcanic-allophanic (Puyehue, Galerina patagonica). CO2 fluxes, lignin peroxidase, manganese peroxidase, and dye-decolorizing peroxidase levels were all determined. Likewise, the production of superoxide anion (O2•-), hydrogen peroxide (H2O2), and hydroxyl radicals (•OH) were assessed in soils microcosms after 36 days of anaerobic incubation with WRF inoculum and induced Fenton reaction under pH variations ranging from 2.5 to 5.1. ROS significantly increased biotic and abiotic CO2 emissions in all tested soils, according to the findings. The highest values (217.45 mg C kg−1) were found during the anaerobic incubation of sterilized and inoculated soils with WRF at a natural pH of 4.5. At pH 4.0, the lowest levels of C mineralization (82 mg C kg−1) were found in Nahuelbuta soil. Enzyme activities showed different trends as pH changed. The Fenton reaction consumed more H2O2 between pH 3 and 4, but less between pH 4.5 and 2.5. The mechanisms that oxidized SOM are extremely sensitive to variations in soil pH and the stability of oxidant radical and non-radical compounds, according to our findings.
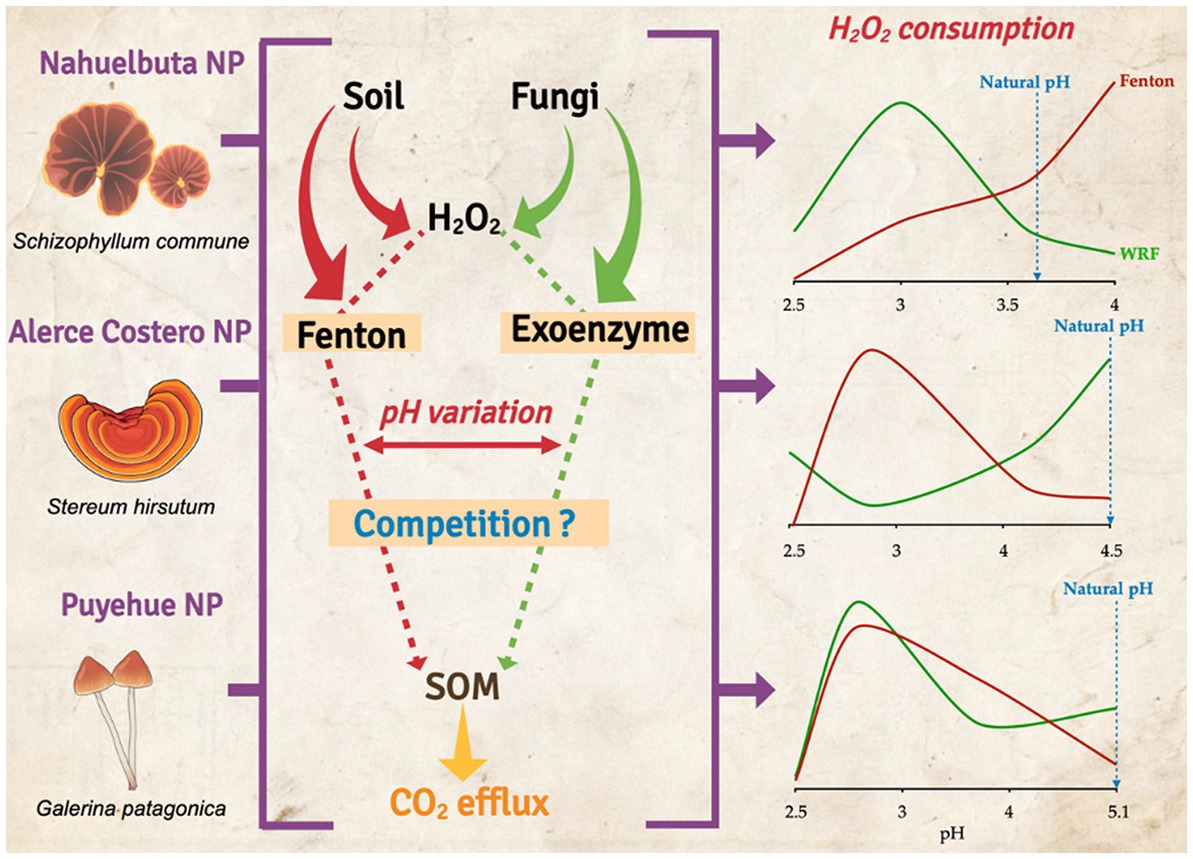
Graphical Abstract
1. Introduction
The rate of industrialization has accelerated changes in carbon dioxide (CO2), methane (CH4), and reactive nitrogen oxides (NxO) emissions from human activities over the last 150 years (Köster et al., 2017; Walsh et al., 2017). Furthermore, the geological and natural sources from which they are released frequently contribute to the current climate change problem. After photosynthesis, soils are the second largest source of CO2 fluxes in terrestrial ecosystems (Raich and Schlesinger, 1992; Dixon et al., 1994; Schlesinger and Andrews, 2000). The sources of emissions are well understood, such as rhizosphere basal microbial respiration and the decomposition of plant residues and soil organic matter (SOM) (Kuzyakov, 2006). As a result, microbial communities play an important role in GHG emissions (Xu et al., 2013). Fungi have a significant function in the forest soil biome because they recycle and decompose both labile and recalcitrant materials like wood lignin under aerobic and anaerobic conditions (Lin et al., 2021; Naylor et al., 2022). This allows fungi to deal with changes in soil redox state resulting from rainfall in the winter and drying in the summer. This means that factors like soluble O2, pH, and oxidation–reduction processes in iron-rich soils in temperate rain forests may have a significant impact on SOM stability under a warming scenario.
The most common type of fungi in these rainforest soils that can efficiently convert lignin into CO2 are white-rot fungi (WRF). This fungus produces a diverse family of hemoperoxidases, including lignin peroxidases (LiP), manganese peroxidase (MnP), and dye peroxidase (DyP), all of which catalyze the decomposition of recalcitrant SOM via a step-by-step reaction catalyzed by hydrogen peroxide (H2O2) as acceptor, and resulting in CO2 efflux (Lenhart et al., 2012). The primary abiotic source of H2O2 in the soil is rainfall deposition (Willey et al., 1996; Kakeshpour et al., 2022). This occurs as a result of a photochemical or electrochemical reaction, such as water photolysis or an electrical storm (electro-Fenton). Furthermore, volatile terpene compounds can produce reactive oxygen species (ROS) in forest ecosystems via indirect biotic processes, reacting with ozone to produce hydroxyl radicals (•OH) and hydrogen peroxide (H2O2), both of which are then deposited in soil by precipitation (Becker et al., 1990). Furthermore, some aerobic soil bacteria produce biotic sources of ROS as a byproduct of respiration by releasing superoxide anion (O2•-), which is rapidly converted into H2O2 by the activity of different rhizosphere isoforms of extracellular superoxide dismutase enzymes (Takahashi et al., 2003; Jofré et al., 2021). Furthermore, ROS stability has been measured over a wide pH range (i.e., 2–10), whereas the maximum rate for metal oxidation occurs only at pH 3 (Garrido-Ramírez et al., 2010; Du et al., 2019).
Although SOM decomposition stimulates GHG release (Merino et al., 2020, 2021a), it is unclear whether biotic (exoenzymes) and abiotic (e.g., Fenton reaction) ROS production can be correlated with soil carbon emission, and whether this mechanism is transversal to the forest with similar structure on a not local scale. This hypothesis appears to be dependent on optimal pH values for biotic GHG production, which is dependent on enzymatic activity and abiotic pathways (Merino et al., 2021a). The optimal soil pH for methanogenesis, according to some reports, is between 4 and 7, but higher CO2 emissions were recorded at circumneutral pH (Dalal and Allen, 2008). As a result, soil pH affects all chemical, physical, and biological soil properties, affecting C fluxes.
Under pH variation, factors such as the availability and consumption of oxidant and radical species (H2O2, •OH, and O2•-) in biotic and abiotic redox reactions should be considered to determine the potential of GHG emissions in temperate rainforest soils. According to changes in soil pH, we hypothesize that the stability and consumption of reactive oxygen species (ROS) from biotic and abiotic reactions can significantly drive the CO2 emission rate from temperate rainforest soils. We investigated the dynamics of ROS under oxygen-limiting conditions, with and without white-rot fungal inoculum, and the Fenton reaction in long-term incubations in this study.
2. Materials and methods
2.1. Study sites and sampling
Three temperate forest soil types with mean annual precipitation ranging from 1,500 to >5,000 mm were chosen. The first soil sampled was a loamy Inceptisol (Soil Survey Staff, 2014) which was derived from intrusive granodiorite rocks in Nahuelbuta National Park (37°47′S, 72°12′W) (Bernhard et al., 2018). This soil was formed by ancient Araucaria araucana and Nothofagus pumilio forests. The second soil sampled in Alerce Costero National Park in the Coastal range under Nothofagus spp. and Fitzroya cupressoides (40°22′S 73°38′W) was a loamy clay Ultisol derived from metamorphic mica-schist materials with illite-kaolinite as dominant clays (Luzio et al., 2003). The final soil was collected in the Andes Cordillera from a primary temperate rainforest of Nothofagus betuloides in Puyehue National Park (40°47′S, 72°12′W) as Andisol derived from recent volcanic ash and basaltic scoria deposits with a high content of allophane, imogolite, and ferrihydrite (Neculman et al., 2013).
Four composite soil samples were extracted from the top Ah mineral horizon (0–15 cm) at each site after the litter layer was removed. The samples were cleaned in the laboratory to remove coarse organic debris and separated into two parts: one was stored at 4°C for a microcosm experiment and microbial analyses, and the other was air-dried for further physico-chemical analyses.
2.2. Analytical soil procedures
The pH and electrical conductivity were measured in an aliquot of soil in a 1:2.5 suspension of soil:water. TOC-VCSH (Shimadzu, Kyoto, Japan) was used to determine soil organic C, and total N was determined using Kjeldahl distillation (VELP, Usmate, Italy). The aluminum, iron and manganese were extracted by oxalate; Alo, Feo and Mno respectively, using 0.2 M ammonium oxalate at pH 3 (Sadzawka et al., 2006). The aluminium, iron and Mn complexed with SOM, extracted by pyrophosphate, Alp, Fep and Mnp, respectively were obtained using a solution of 0.1 M sodium pyrophosphate (Van Reeuwijk, 2002). To identify exchangeable, crystalline, and complexed-SOM metals in soil samples, dithionite-citrate-bicarbonate (Fed) was used. Atomic absorption spectroscopy (Perkin Elmer 3110, Waltham, Massachusetts, United States) was used to determine Fe and Mn concentrations at 248.3 nm for Fe and 279.5 nm for Mn using a nitrous oxide acetylene flame. Cation exchange capacity (CEC) and nutrient characterization were carried out as Sadzawka et al. (2006) suggested. The total Fe concentration in 100 mg of dry soil was determined by adding 0.9 mL 0.28 M hydroxylamine hydrochloride and 1 mL 0.28 M HCl (Stookey, 1970). Approximately 100 μL of the extract was added to 4 mL of ferrozine color reagent (1 g ferrozine in 6.5 M ammonium acetate solution). Iron (II) concentration was determined in 100 mg of soil by adding 1 mL of 0.5 M HCl and vigorously shaking. Ferrozine reagent was added as previously described, and absorbance at 562 nm (ferrozine-Fe(II) complex standard) was measured in a UV spectrophotometer (Lovley and Phillips, 1987). The difference between total Fe and extractable Fe(II)-HCl forms was used to calculate the Fe(III) (oxyhydr) oxide content.
2.3. Culture conditions and fungal identification
In the same areas where soil samples were taken, white rot fungi were isolated from small fragments of decayed wood or fruiting body pieces. The fragments were placed in sterile tubes and stored at 4°C until they were analyzed. Small fragments of fungi fruiting bodies or decayed wood colonized by fungi were incubated on acidified glucose malt extract agar plates (15 g/L agar, 3.5 g/L malt extract, 10 g/L glucose, pH 5.5). Under aseptic conditions, pure mycelial cultures were obtained, and strains were identified using ITS sequencing. The DNA from each strain was extracted using the E.Z.N.A.® SP Fungal DNA Mini Kit D5524-01 (Omega, Bio-Tek, Norcross, GA, United States). The ITS1-5.8S—ITS2 rDNA was amplified using ITS1 and ITS4 primers (White et al., 1990). PCR was carried out in a total volume of 25 μL with 0.1 mM dNTPs, 0.1 mM of each primer, 5 U of Taq DNA polymerase, and the supplied reaction buffer (Promega Inc., Seoul, Korea). An ABI PRISM 3730 l DNA Analyzer System was used to sequence the PCR products (Macrogen, Seoul, Korea). The nucleotide sequences were compared in the GenBank database (Horisawa et al., 2013). Schizophyllum commune was identified in Nahuebuta soil, Stereum hirsutum in Alerce Costero soil, and Galerina patagonica in Puyehue soil (see Supplementary Table S1, for more information). In each microcosm, the isolates were inoculated at a final concentration of 3 × 108 CFU gr soil−1 in 100 μL of sterilized water.
2.4. Microcosm anaerobic experiments
Under anaerobic conditions, the contribution of and ROS production and consumption from soils at different pH levels was determined in a sterilized portion of soil at field capacity (80%) from each soil (oxygen-free by N2 purge). In a destructive sampling design, soil samples were incubated in 120 mL serum bottles for 36 h at 12°C to monitor CO2 evolution, ROS production, and enzymatic activity. To remove the microbial resistant structures, the soil samples were sterilized for 20 min at 121°C (in an autoclave) on four consecutive days. The soils were then fumigated with chloroform vapor for 24 h in a vacuum chamber (Trevors, 1996). Autoclaving was chosen because it has little effect on the structure of SOM, primarily causing changes in the carbohydrate and N-alkyl domains (Berns et al., 2008). Because it causes Fe reduction and oxidation, gamma radiation was avoided (Bank et al., 2008; Abedini et al., 2014; Sutherland et al., 2017). Gamma radiation also increased the bioavailability of Fe(III) (oxyhydr)oxide minerals, which helped in Fe(III) reduction (Brown et al., 2012). There were three treatments (WRF, Fenton reaction, and WRF + Fenton) and four replicates with four induced pH in each soil; 2.5, 3, 4, and the natural pH of each soil (3.6, 4.5, and 5.1 for Nahuelbuta, Alerce Costero, and Puyehue, respectively). A H2O2:Fe(II) ratio was also added to induce the Fenton reaction, as previously reported by Merino et al. (2020). This was achieved by adding 120–143 mL of H2O2 (0.1 M) and 1.29 g Fe(II) kg−1 soil as FeCl2 to the soil (Sigma Aldrich, United States). A total of 144 serum bottles with septums for gas sampling were used. CO2 was collected at 0.5, 4, 8, 12, 24, and 36 days of incubation. Approximately 10 mL of CO2 gas sample was extracted and injected into a gas chromatograph with Flame Ionization Detector (GC/FID) (Thermo Fisher ScientificTM, Austin, TX) with a 30 mm DB1-MS column and selected ion mode at each sampling time. At the end of the incubation period, 72 microcosm bottles were harvested, and soil was homogenized and quickly subsampled for ROS analysis and enzymatic activity at each sampling.
2.5. Reactive oxygen species (ROS) detection
Superoxide anion (O2•-) production was measured in 0.5 g of soil, which was extracted with 12 mL of potassium phosphate buffer (PPB, 65 mM, pH = 7.8) and centrifuged at 10,000×g for 15 min at 4°C. The supernatant (5 mL) was combined with 0.9 mL of PPB, 1 mL of sulfanilamide (17 mM), 1 mL of hydroxylamine hydrochloride (10 mM), and 1 mL of naphthylamine (7 mM) and incubated in the dark for 20 min at 25°C. At 530 nm, the absorbance of the samples was measured (Greenwald, 2018). Hydrogen peroxide (H2O2) was determined using the iodometric titration method (Velikova et al., 2000). The concentration of H2O2 in mg L−1 was determined using a calibration curve as the standard. Although iodometric titration is less precise than permanganate titration, it produces fewer interferences with SOM (Liang and He, 2018; Merino et al., 2020). Page et al. (2013) previously used terephthalic acid (TPA) in soil to measure •OH production. Approximately 200 l of soil water (1:2.5 soil:water suspension) was injected into 3 mL aliquots of deoxygenated water for the blank (in triplicate) and 3 mL of TPA for the test (in triplicate) (1.5 mM, in triplicate). TPA-treated soil suspension was incubated for 24 h in the dark to oxidize into hydroxyterephthalic acid (hTPA). Finally, the sample was filtered at 0.22 m and the fluorescence was measured in a microplate reader at λex = 310 nm, λem = 425 nm (Synergy HT, Biotek). The fluorescence intensity was compared to a standard curve with 2-hydroxyterephthalic acid (0, 2, 20, 40, and 80 nM) (hTPA, Sigma Aldrich).
2.6. Enzyme activity
Exoenzymatic activities were spectrophotometrically measured under pH variations after 36 h of incubation. The formation of Mn(III)-tartrate complex during the oxidation of 0.1 mM MnSO4 in 100 mM tartrate buffer at pH 5 was used to determine manganese peroxidase (MnP) activity. At 238 nm, manganese peroxidase was measured spectrophotometrically (Xu et al., 2018). The oxidation of 2,2′-azino-bis (3-ethylthiazoline-6-sulfonate) (ABTS) (2.5 mM) to its cation radical in 100 mM tartrate buffer at pH 5 was followed by dye-decolorizing peroxidase (DyP) activity. At 418 nm, dye-decolorizing peroxidase was measured spectrophotometrically (Salvachúa et al., 2013). Veratryl alcohol (2 mM) was used as a substrate for LiP during the oxidation of 0.21 mL of 50 mM sodium tartrate buffer at pH 2.5. At 310 nm, lignin peroxidase was measured. The enzymatic activity tests were all performed in the presence of 0.1 mM H2O2. All oxidative enzymatic activities were measured in units (U) per milliliter (i.e., one millimole of substrate oxidized per minute). The measured H2O2 was normalized in relation to the treatment values in terms of representation.
2.7. Statistical analysis
The normal data distribution and variance homogeneity were tested for each treatment and soil type. One-way ANOVA was used for cumulative gas sampling (CO2). Using a repeated-measures ANOVA test, the enzyme activity, ROS production, and Fenton contribution were tested over a 40-day incubation period. The last three variables measured were plotted as a six-sample time average. Duncan’s multiple range test was used for multiple comparison means because all ANOVA tests were significant at p 0.05. The RStudio software was used for all analyses (1.1.442).
3. Results
3.1. Soil properties
The soils studied are formed from a variety of parent materials, including Granitic (Nahuelbuta), Metamorphic (Alerce Costero), and Volcanic-allophanic (Puyehue); their textures range from sandy to clay. The pH ranged from 3.6 to 5.1, with organic C levels ranging from 9.2 ± 0.1 to 11.4% ± 0.3. Natural H2O2 levels in soil range between 25.6 ± 0.7 and 33.7 ± 0.5 μM g−1 soil (Nahuelbuta<Alerce Costero< Puyehue soils). Puyehue soil had the highest content of Al complexed with SOM (Alp) (11 g kg−1 soil ±1.5), while Nahuelbuta soil had the lowest (0.7 g kg−1 soil ±0.1). The Alo + 1/2 Feo > 2%, which indicates Andic properties <2, was found at higher levels of 3.8% in Puyehue soil. The Alo symbol represents Al-oxides associated with amorphous structures, such as Alp with SOM. Puyehue soil had the highest value of 3.1 ± 0.2 g kg−1 soil. The Puyehue soil had the lowest CEC (5.3 cmol(+) kg−1), while Alerce Costero soil had the highest (19.7 cmol(+) kg−1). The total Fe content was determined by combining the soluble ferrous iron Fe(II) and ferric iron Fe(III) forms, which ranged between 8.4 ± 0.5 and 11.8 ± 0.5 g kg−1. Alerce Costero soil had a higher total Fe content than other soils (Table 1).
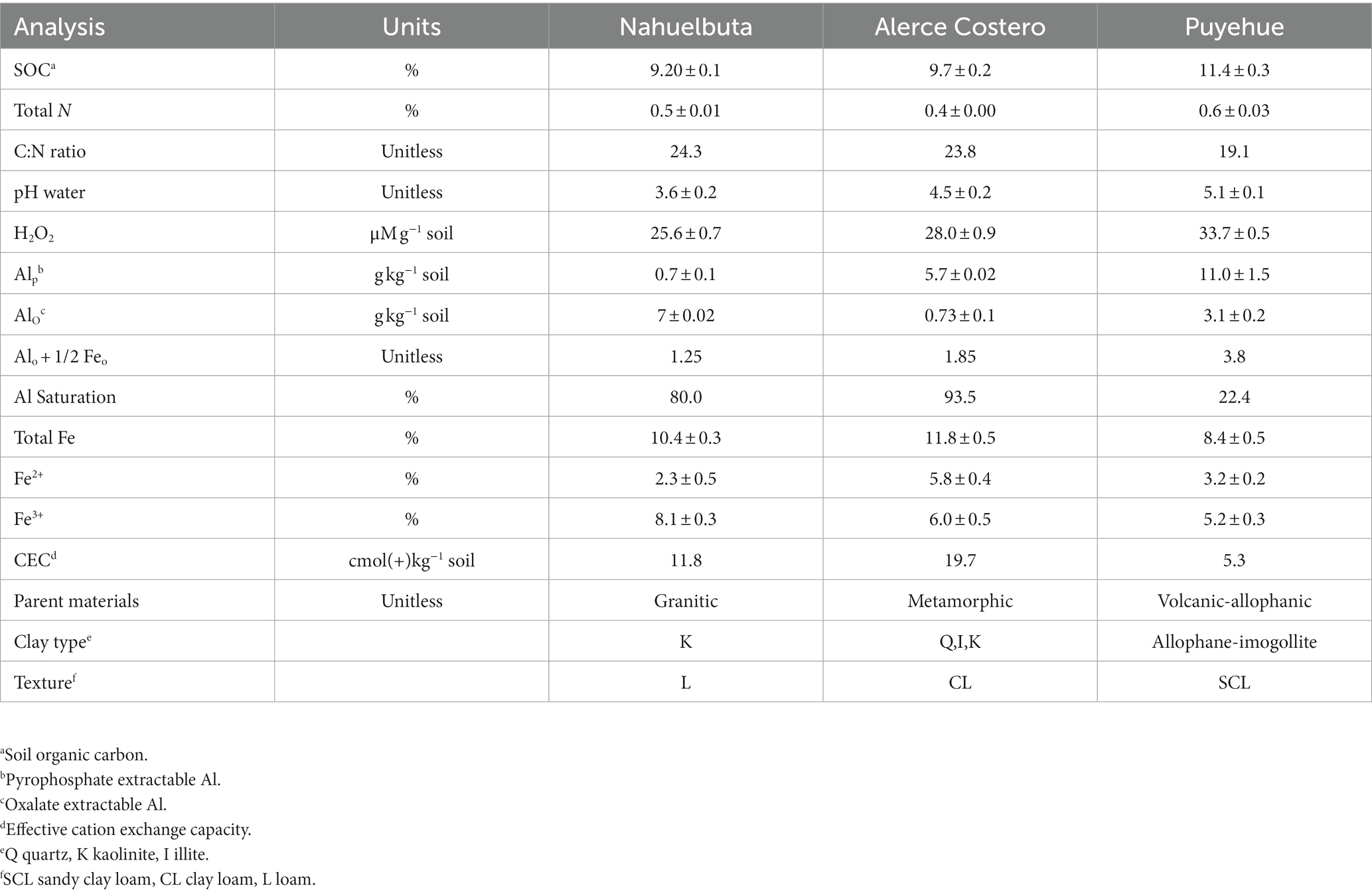
Table 1. Characteristics of soil used in the study.
3.2. CO2 evolution
Over the course of 36 days, increasing patterns of CO2 efflux were observed in all evaluated soils (Figures 1A–C). The highest values were found in Alerce Costero soil during anaerobic incubation of sterilized samples inoculated with WRF, at pH of 4.5 (217.45 mg C kg−1) (Figure 1B). The lowest levels of C mineralization (82 mg C kg−1) with the lowest pH 4 were found in Nahuelbuta soil (Figure 1A). Alerce Costero and Puyehue soils followed a similar pattern, with the highest CO2 release occurring at the highest pH levels, 4.5 and 5.1, and the lowest at induced pH 2.5 (Figures 1B,C). Figure 1A depicts the inverse trends for Nahuelbuta soil. Furthermore, all the soils treated with WRF without Fenton, and soils treated without inoculum, showed a lower average rate values than the combined treatments (WRF + Fenton). Similarly, the lowest average rate of mineralization was observed in sterile soils treated with Fenton, followed by WRF without Fenton induction (Figures 1A,C).
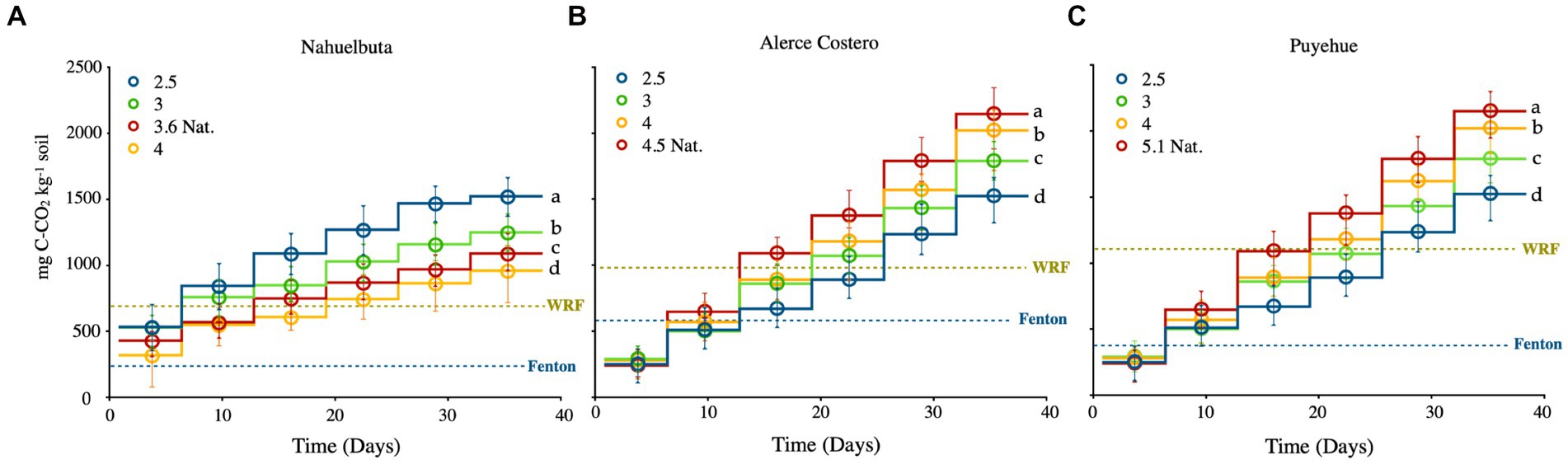
Figure 1. (A–C) CO2 evolution from anaerobic sterilized soils inoculated with WRF and H2O2/Fe for Fenton reaction, derived from three temperate rain forest sites with varying pH and incubated for 40 days at 12°C. The blue dotted lines represent the average rate of CO2 released by the induced Fenton reaction in sterilized soils at natural pH, while the green dotted lines represent the average rate of CO2 released by WRF at natural pH without the induction of the Fenton reaction. Significant differences (p < 0.05) are indicated by different letters in each panel. The ladder graph depicts the total amount of CO2 emitted between measurements.
3.3. Exo-enzyme activities
After 36 days of incubation, the activity of LiP, MnP, and DyP was measured. When compared to non-induced Fenton groups, all WRF plus induced-Fenton reactions increased proportionally to enzyme activity. The responses of enzyme activities to pH changes in the soils studied varied (Figure 2). Except for Puyehue soil, only the Nahuelbuta microcosm showed a decrease in activity as the pH increased, with the highest activity observed at pH 2.5 (Figures 2A,D,G). LiP, MnP, and DyP were one, two, and eight times higher, respectively, than at natural pH. The CO2 released was distributed uniformly across the soil pH gradient. With the exception of Alerce Costero with LiP and Puyehue, the highest pH showed the lowest enzymatic activity and CO2 release after 36 days of incubation. LiP activity in Alerce Costero soil displayed a distinct pattern of activity when compared to MnP and DyP. The activity of the LiP increased as the pH increased from acidic to moderate acidic, with higher activity observed at pH (4.5) (Figure 2B). MnP and DyP, on the other hand, significantly reduced their activity from pH 2.5 to 4.5, while CO2 increased at day 36 (Figures 2E,H). The pH variation in Puyehue soil was observed to have a non-linear distribution. Lip, MnP, and DyP activity were found to be lower in acidic conditions (pH 2.5), with the highest activity at pH 3 (Figures 2C,F,I), followed by a decrease in activity at natural pH (4.5).
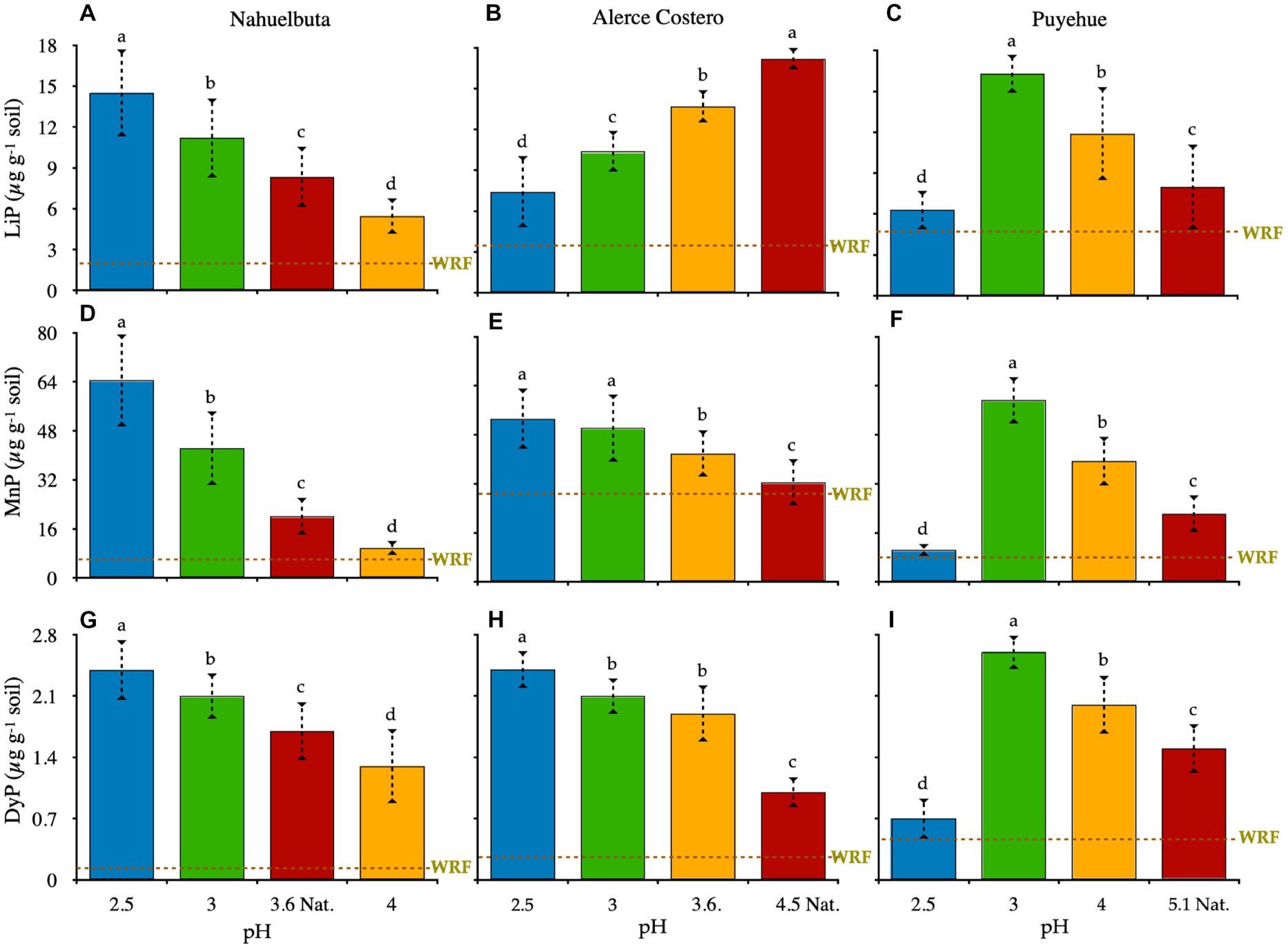
Figure 2. A–C: lignin peroxidase (LiP); D–F: manganese peroxidase (MnP); and G–I: dye-decolorizing peroxidase (DyP) in soils inoculated with WRF from three temperate forest soil sites at various pH levels. At natural pH, the green dotted lines represent the average rate of cumulative enzyme activity by WRF in the absence of the Fenton reaction. Within each panel, different letters indicate significant differences (p < 0.05).
3.4. Effect of pH on ROS generation
pH had an effect on O2•- production, and the variation was significant between treatments and soils, ranging from 0.5 to 1.3 μmol g−1 soil (Figures 3A,D,G). The highest values in Nahuelbuta soil were obtained at higher pH, in the Fenton + WRF group, and in the independent treatments (Figures 3A–C). This pattern was also observed in Alerce Costero soil, where pH 2.5 and 3 had the highest levels of O2•-, decreasing to 4 and 4.5 (Figure 3A). In comparison to the other soils, the values in Puyehue soil were inverse to the trend (Figure 3G). The lowest levels of O2•- were found in the most acidic values (2.5), but at pH 3, the maximum production was reached, and as the pH increased, the O2•- content decreased (Figure 3G). As the pH changed, the production of H2O2 varied without a clear trend (Figures 3B,C,H). The highest value of 3.4 μmol g−1 of soil was discovered in Alerce Costero at a natural pH of 4.5 (Figure 3E). Puyehue had the lowest H2O2 production of 1.3 μmol g−1 soil and the lowest pH of 2.5 (Figure 3H). These values take into account the subtraction of the added H2O2 to initiate the Fenton H2O2/Fe(II) reaction (10,1 ratio). When compared to the other reactive oxygen species, the production of hydroxyl radicals (•OH) was the highest (ROS) (Figures 3C,F,I). Similar to H2O2 production, there was no discernible trend in •OH production, with the highest value of 4.3 μmol g−1 soil being recorded at the lowest pH 2.5 in Nahuelbuta (Figure 3C). The almost undetectable •OH 0.7 μmol g−1 soil was recorded in Puyehue with the lowest pH of 2.5 (Figure 3I). The WRF + Fenton treatment produced the highest total ROS, followed by Fenton and the WRF treatments with lower contributions (Figure 3). Furthermore, the cumulative production of ROS (the sum of the values of each ROS in all soils) was found to be lowest for O2•-, followed by •OH, and H2O2 had the highest abundance.
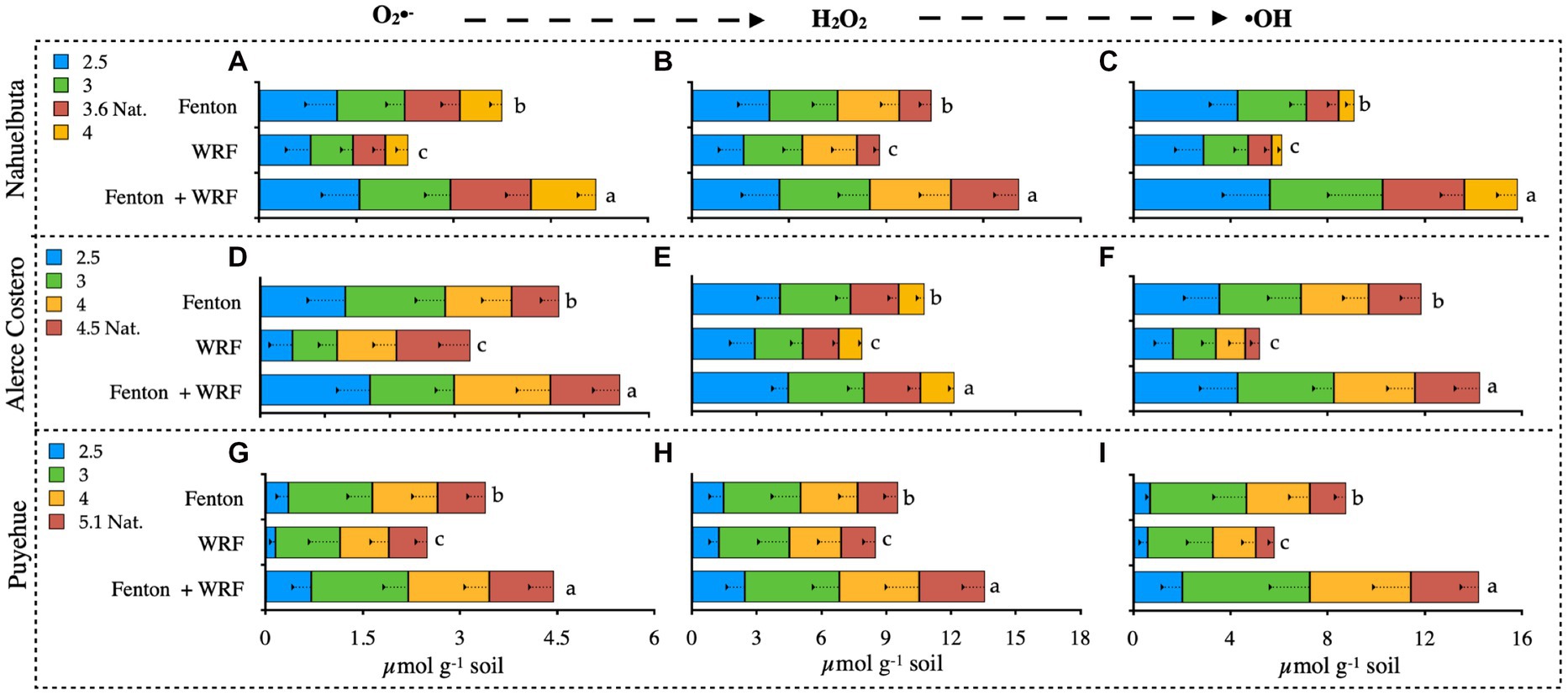
Figure 3. Detection of reactive oxygen species (ROS): superoxide anion (A,D,G); hydrogen peroxide (B,E,H); and hydroxyl radical (C,F,I) from soils inoculated with WRF from three temperate forest soil sites at different pHs. Significant differences (p < 0.05) are indicated by different letters in each panel.
3.5. H2O2 consumption
Figure 4 depicts the consumption of H2O2 by exoenzyme and Fenton reaction after 36 days of incubation due to their high oxidative content, allowing them to be used as a cofactor in catalysis and iron oxidation (Fenton). When WRF was added to Nahuelbuta soil, this increased H2O2 consumption between pH 2.5 and 3.6, but decreased at pH 4. The Fenton reaction, on the other hand, consumes more H2O2 at higher pH levels (pH 4) (Figure 4A). This was not the case with Alerce Costero soil. At moderate acidity pH 4.5 and acid condition (2.5), the exoenzyme consumes more H2O2, whereas consumption is similar but significantly reduced at pH 3 and 4. The Fenton reaction, on the other hand, exhibits higher consumption values between pH 3 and 4, but lower consumption values between pH 4.5 and 2.5 (Figure 4B). At pH 3.6, similar consumption values were observed in both exoenzyme and Fenton reactions, but not at the other pH tested. When the pH is raised, exoenzymes consume the least. At pH 2.5, the Fenton reaction produced the lowest values (Figure 4C).
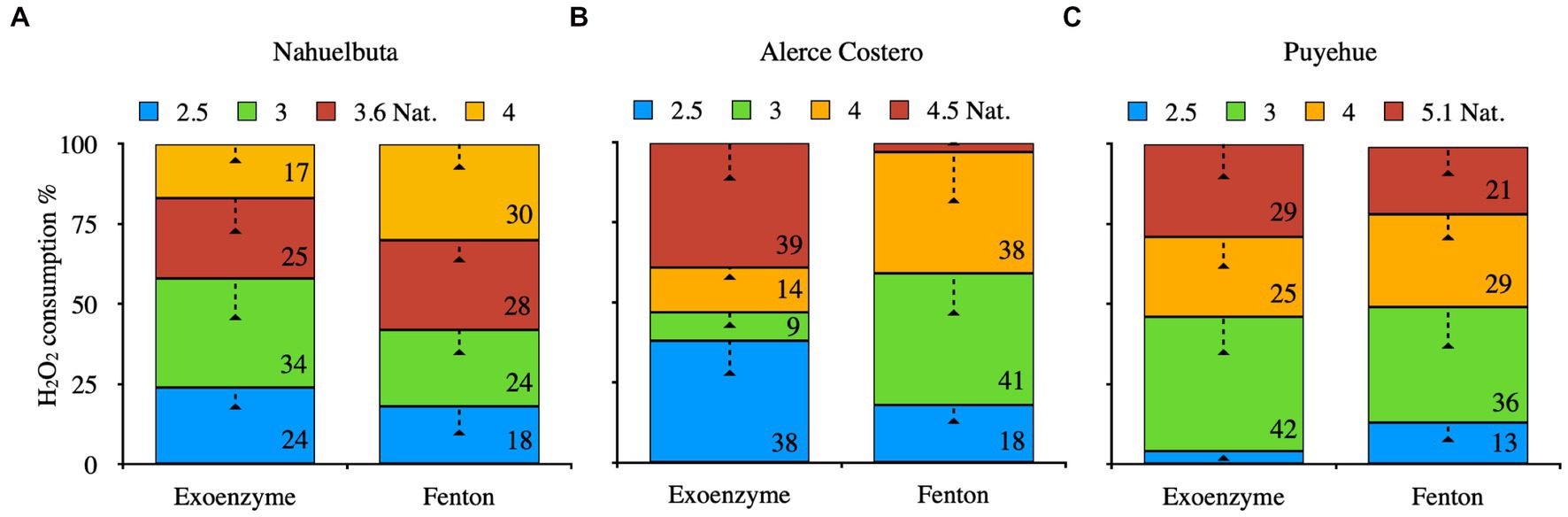
Figure 4. The average of exoenzyme activity or Fenton reaction from anaerobic sterilized soils inoculated with WRF from Nahuelbuta (A), Alerce Costero (B), and Puyehue (C) forest soil sites at different pH levels. Incubated for 36 days at 12°C. Significant differences (p < 0.05) are indicated by different letters in each panel.
3.6. Relationships between variables measured
The consumption of CO2 and H2O2 was found to have a positive and significant relationship (Figures 5A–C). CO2 production in WRF-treated soils was strongly related to peroxide consumption (p < 0.01, R2 > 0.85). Puyehue soil had a lower CO2 evolution trend associated with H2O2 consumption, but the tendency was lower in comparison to Alerce Costero soil, which had a higher CO2 production and H2O2 consumption at pH 3, with a steeper slope (Figures 5B,C).
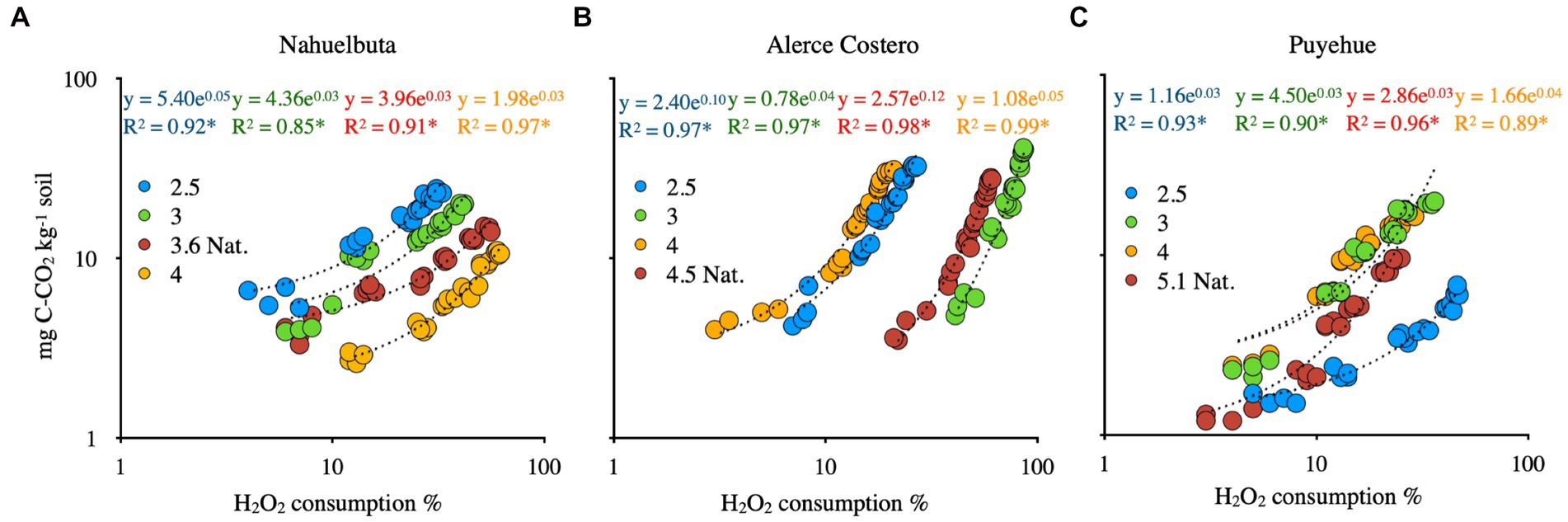
Figure 5. The relationships between CO2 and H2O2 consumption in anaerobic sterilized soils inoculated with WRF from Nahuelbuta (A), Alerce Costero (B) and Puyehue (C) forest soil sites at different pH levels. For 40 days, it was incubated at 12°C. All relationships are statistically significant at p < 0.01.
4. Discussion
The abiotic decomposition of SOM is driven by reactive oxygen species such as O2•-, •OH, and H2O2, which occurs naturally even in the absence of enzymatic activity (Piccolo et al., 2018). H2O2 can be incorporated into soils via wet and dry atmospheric deposition (Petigara et al., 2002), intense solar radiation (Georgiou et al., 2015), oxidation of Fe (II) or reduced dissolved organic carbon (DOC) by oxygen (Page et al., 2013; Trusiak et al., 2018), and biotic input via the release of various oxidative compounds by soil bacteria and fungi. The biotic or abiotic H2O2 input is critical in the decomposition of labile and recalcitrant SOM, increasing H2O2 concentration (Merino et al., 2020, 2021b). An excess of H2O2, on the other hand, causes competition for H2O2 between abiotic and biotic processes.
The WRF generates endogenous H2O2 that serves as a cofactor for many enzymes that control fungal biomass conversion and the activity of lignolytic enzymes (e.g., Mn-peroxidase, LiP-peroxidase, lytic polysaccharide monooxygenases), which can be driven in the absence of O2 (Wong, 2009; Suryadi et al., 2022). Brown-rot fungi, which lack lignin peroxidases, may use the released H2O2 to drive the Fenton reaction, which is unique to these fungi (Arantes et al., 2012). As a result, competition can have a significant impact on the efficiency with which SOM is mineralized. Excess H2O2 may cause parallel (additive or synergistic) biotic and abiotic reactions, whereas fungi native production will compensate for the oxidant’s substrate for the Fenton reaction if there is insufficient H2O2. This would have an antagonistic effect on SOM degradation efficiency.
Both biotic (Diaz et al., 2013; Shah et al., 2016) and abiotic (Garrido-Ramírez et al., 2010; Georgiou et al., 2015) factors influenced the level of H2O2 in soil. The pH of the soil influences both the biotic and abiotic response to oxidative processes, according to our findings. The biotic component is most sensitive to changes in pH, whereas Fenton-mediated oxidation increases H2O2 consumption and CO2 efflux. When the pH was close to 5.1, the Alerce Costero and Puyehue soils showed similar higher CO2 emission trends. The highest efflux was observed in Nahuelbuta soil at more extreme acidic pH levels (2.5). This could be attributed to the Schizophyllum commune strain, which grows well in vitro under acidic conditions, which could translate into increased enzymatic activity. Microbial activity regulates extracellular O2•- concentrations in the soil environment (Diaz et al., 2016), which may affect soil H2O2 levels. The effect of pH treatment on WRF activity in the three soils studied was comparable to previous studies (Leff et al., 2015; Ling et al., 2016). Soil CO2 emissions had a significant positive relationship with soil H2O2 consumption (Figure 5). Increased biodiversity may improve Fe mobilization and H2O2 decomposition in soils, according to these findings. This effect is closely related to soil functional selection, which includes nutritional and physical conditions that favor the establishment of specific microorganisms over others (Mimmo et al., 2014). Based on soil organic matter (SOM) availability and N content, previous research has shown that endemic microbiota can cause significant CO2 flux and reactive oxygen species (ROS) production (Merino et al., 2021a).
Reactive oxygen species are known to play a critical role in the generation of CO2 by WRF (Hammel et al., 2002), the soil CO2 released was investigated using ROS as a factor. Given that H2O2 has long been used as an oxidant in soil degradation and SOM assessment, our discovery of a strong and linear relationship between CO2 released and soil H2O2 concentrations was not surprising (Robinson, 1927). Previously, researchers discovered a strong positive linear correlation between CO2 production and H2O2 concentrations in deciduous forest soils (Jugold et al., 2012). It is possible that different types of soil precursors will react differently to H2O2. More research on the specific precursors of abiotic CH4 produced in soils should be conducted to determine how the precursors and ROS are likely to interact biochemically. However, we were unable to establish a clear relationship between H2O2 concentrations and the other ROS precursors, implying that the H2O2 levels used in this study were possibly too high and that other factors (e.g., soil Fe or Mn) had become limiting in the generation of ROS.
Our findings show that anaerobic soils produce a higher proportion of •OH in soils, and that Fe(II) is the primary electron donor allowing the synthesis of •OH and subsequent SOM decomposition, which results in CO2 release in soils. •OH measurements during anaerobic incubation were consistent with previous research (Page et al., 2013; Minella et al., 2015). •OH generation in anaerobic and low-O2 soil can also be supported by reducing conditions (i.e., high electron donating capacity and thus high concentrations of electron donors) present across a broad pH range (Page et al., 2013). The pH and oxygen levels in the studied soils were significantly lower (Lipson et al., 2012, 2013), and Fe(II) concentrations provided highly reducing conditions. •OH production from the oxidation of reduced components such as Fe(II) or reduced SOM can be significantly influenced by pH. •OH can also be produced by oxidizing Fe(II) at a pH close to neutral (Remucal and Sedlak, 2011). For example, oxidation of Fe(II) at pH 5 may result in the formation of ferryl iron (Fe(IV)) and •OH, reducing the ratio of •OH generated per mol oxidized Fe(II) (Vermilyea and Voelker, 2009; Remucal and Sedlak, 2011). Other studies have revealed that at low pH (5), Fe(II) oxidation produces more •OH than Fe(IV) (Remucal and Sedlak, 2011). In contrast to Fe(II) oxidation, the effect of pH on the production of •OH from reduced DOC oxidation has not been studied. Aeschbacher et al. (2012) found that the oxidation of DOC to produce •OH is more favorable at high pH than at low pH, implying that this process occurs more frequently at higher pH.
5. Conclusion
This research investigated the impact of reactive oxygen species (ROS) on CO2 efflux and enzymatic activity during long-term anaerobic incubation. Despite the fact that few studies have proven that H2O2 availability has an effect, our findings show that abiotic and biotic reactions using reactive oxygen species as a feed for both mechanisms have an antagonistic and synergistic effect on GHG emission. However, in all soils tested, ROS production significantly increased biotic and non-biotic CO2 emissions. This implies that microorganisms and soil structure have an additional influence on C efflux, which is pH dependent. When bacterial microbiota undergoes redox fluctuations, WRF contributes more than 70% of GHG emissions (N2O and CO2), which is consistent with previous research conducted in the same forest region. According to the findings of this study, the mechanisms of microbial SOM oxidation are highly dependent on the stability and abundance of oxidant radical and non-radical compounds, as well as changes in soil pH.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI—OQ341229, OQ339139 + OQ341421.
Author contributions
CM-G and IJ-F: conceptualization, resources, writing—original draft preparation, and funding acquisition. CM-G: methodology, formal analysis, and project administration. IJ-F: investigation. FM-B: writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by FONDECYT Postdoctorado, grant number 3200758, and FONDECYT regular, grant number 1220716. The APC was partially funded by Universidad de La Frontera, grant number DI21-1003.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1148750/full#supplementary-material
References
Abedini, A., Daud, A. R., Abdul Hamid, M. A., and Kamil Othman, N. (2014). Radiolytic formation of Fe3O4 nanoparticles: influence of radiation dose on structure and magnetic properties. PLoS One 9:e90055. doi: 10.1371/journal.pone.0090055
Aeschbacher, M., Graf, C., Schwarzenbach, R. P., and Sander, M. (2012). Antioxidant properties of humic substances. Environ. Sci. Technol. 46, 4916–4925. doi: 10.1021/es300039h
Arantes, V., Jellison, J., and Goodell, B. (2012). Peculiarities of brown-rot fungi and biochemical Fenton reaction with regard to their potential as a model for bioprocessing biomass. Appl. Microbiol. Biotechnol. 94, 323–338. doi: 10.1007/s00253-012-3954-y
Bank, T. L., Kukkadapu, R. K., Madden, A. S., Ginder-Vogel, M., Baldwin, M., and Jardine, P. (2008). Effects of gamma-sterilization on the physico-chemical properties of natural sediments. Chem. Geol. 251, 1–7. doi: 10.1016/j.chemgeo.2008.01.003
Becker, K. H., Brockmann, K. J., and Bechara, J. (1990). Production of hydrogen peroxide in forest air by reaction of ozone with terpenes. Nature 346, 256–258. doi: 10.1038/346256a0
Bernhard, N., Moskwa, L.-M., Schmidt, K., Oeser, R. A., Aburto, F., Bader, M. Y., et al. (2018). Pedogenic and microbial interrelations to regional climate and local topography: new insights from a climate gradient (arid to humid) along the coastal cordillera of Chile. Catena 170, 335–355. doi: 10.1016/j.catena.2018.06.018
Berns, A. E., Philipp, H., Narres, H.-D., Burauel, P., Vereecken, H., and Tappe, W. (2008). Effect of gamma-sterilization and autoclaving on soil organic matter structure as studied by solid state NMR, UV and fluorescence spectroscopy. Eur. J. Soil Sci. 59, 540–550. doi: 10.1111/j.1365-2389.2008.01016.x
Brown, M. E., Barros, T., and Chang, M. C. (2012). Identification and characterization of a multifunctional dye peroxidase from a lignin-reactive bacterium. ACS Chem. Biol. 7, 2074–2081. doi: 10.1021/cb300383y
Dalal, R. C., and Allen, D. E. (2008). Greenhouse gas fluxes from natural ecosystems. Aust. J. Bot. 56, 369–407. doi: 10.1071/BT07128
Diaz, J. M., Hansel, C. M., Apprill, A., Brighi, C., Zhang, T., Weber, L., et al. (2016). Species-specific control of external superoxide levels by the coral holobiont during a natural bleaching event. Nat. Commun. 7, 1–10. doi: 10.1038/ncomms13801
Diaz, J. M., Hansel, C. M., Voelker, B. M., Mendes, C. M., Andeer, P. F., and Zhang, T. (2013). Widespread production of extracellular superoxide by heterotrophic bacteria. Science 340, 1223–1226. doi: 10.1126/science.1237331
Dixon, R. K., Solomon, A., Brown, S., Houghton, R., Trexier, M., and Wisniewski, J. (1994). Carbon pools and flux of global forest ecosystems. Science 263, 185–190. doi: 10.1126/science.263.5144.185
Du, H.-Y., Yu, G.-H., Sun, F.-S., Usman, M., Goodman, B. A., Ran, W., et al. (2019). Iron minerals inhibit the growth of Pseudomonas brassicacearum J12 via a free-radical mechanism: implications for soil carbon storage. Biogeosciences 16, 1433–1445. doi: 10.5194/bg-16-1433-2019
Garrido-Ramírez, E. G., Theng, B. K., and Mora, M. L. (2010). Clays and oxide minerals as catalysts and nanocatalysts in Fenton-like reactions—a review. Appl. Clay Sci. 47, 182–192. doi: 10.1016/j.clay.2009.11.044
Georgiou, C. D., Sun, H. J., McKay, C. P., Grintzalis, K., Papapostolou, I., Zisimopoulos, D., et al. (2015). Evidence for photochemical production of reactive oxygen species in desert soils. Nat. Commun. 6:7100. doi: 10.1038/ncomms8100
Greenwald, R. A. (2018). Handbook Methods for Oxygen Radical Research CRC Press. doi: 10.1201/9781351072922
Hammel, K., Kapich, A., Jensen, K., and Ryan, Z. (2002). Reactive oxygen species as agents of wood decay by fungi. Enzym. Microb. Technol. 30, 445–453. doi: 10.1016/S0141-0229(02)00011-X
Horisawa, S., Sakuma, Y., and Doi, S. (2013). Identification and species-typing of wood rotting fungi using melting curve analysis. J. Wood Sci. 59, 432–441. doi: 10.1007/s10086-013-1349-z
Jofré, I., Matus, F., Mendoza, D., Nájera, F., and Merino, C. (2021). Manganese-oxidizing Antarctic Bacteria (Mn-Oxb) release reactive oxygen species (ROS) as secondary Mn (II) oxidation mechanisms to avoid toxicity. Biology 10:1004. doi: 10.3390/biology10101004
Jugold, A., Althoff, F., Hurkuck, M., Greule, M., Lelieveld, J., and Keppler, F. (2012). Non-microbial methane formation in oxic soils. Biogeosci. Discuss. 9, 5291–5301. doi: 10.5194/bgd-9-11961-2012
Kakeshpour, T., Metaferia, B., Zare, R. N., and Bax, A. (2022). Quantitative detection of hydrogen peroxide in rain, air, exhaled breath, and biological fluids by NMR spectroscopy. Proc. Natl. Acad. Sci. 119:e2121542119. doi: 10.1073/pnas.2121542119
Köster, K., Köster, E., Kulmala, L., Berninger, F., and Pumpanen, J. (2017). Are the climatic factors combined with reindeer grazing affecting the soil CO2 emissions in subarctic boreal pine forest? Catena 149, 616–622. doi: 10.1016/j.catena.2016.06.011
Kuzyakov, Y. (2006). Sources of CO2 efflux from soil and review of partitioning methods. Soil Biol. Biochem. 38, 425–448. doi: 10.1016/j.soilbio.2005.08.020
Leff, J. W., Jones, S. E., Prober, S. M., Barberán, A., Borer, E. T., Firn, J. L., et al. (2015). Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. 112, 10967–10972. doi: 10.1073/pnas.1508382112
Lenhart, K., Bunge, M., Ratering, S., Neu, T. R., Schüttmann, I., Greule, M., et al. (2012). Evidence for methane production by saprotrophic fungi. Nat. Commun. 3, 1–8. doi: 10.1038/ncomms2049
Liang, C., and He, B. (2018). A titration method for determining individual oxidant concentration in the dual sodium persulfate and hydrogen peroxide oxidation system. Chemosphere 198, 297–302. doi: 10.1016/j.chemosphere.2018.01.115
Lin, Y., Campbell, A. N., Bhattacharyya, A., DiDonato, N., Thompson, A. M., Tfaily, M. M., et al. (2021). Differential effects of redox conditions on the decomposition of litter and soil organic matter. Biogeochemistry 154, 1–15. doi: 10.1007/s10533-021-00790-y
Ling, N., Zhu, C., Xue, C., Chen, H., Duan, Y., Peng, C., et al. (2016). Insight into how organic amendments can shape the soil microbiome in long-term field experiments as revealed by network analysis. Soil Biol. Biochem. 99, 137–149. doi: 10.1016/j.soilbio.2016.05.005
Lipson, D. A., Raab, T. K., Goria, D., and Zlamal, J. (2013). The contribution of Fe (III) and humic acid reduction to ecosystem respiration in drained thaw lake basins of the Arctic coastal plain. Glob. Biogeochem. Cycles 27, 399–409. doi: 10.1002/gbc.20038
Lipson, D. A., Zona, D., Raab, T., Bozzolo, F., Mauritz, M., and Oechel, W. (2012). Water-table height and microtopography control biogeochemical cycling in an Arctic coastal tundra ecosystem. Biogeosciences 9, 577–591. doi: 10.5194/bg-9-577-2012
Lovley, D. R., and Phillips, E. J. (1987). Rapid assay for microbially reducible ferric iron in aquatic sediments. Appl. Environ. Microbiol. 53, 1536–1540. doi: 10.1128/aem.53.7.1536-1540.1987
Luzio, W., Sadzawka, A., Besoain, E., and Lara, P. (2003). Influence of volcanic materials on red clay soil genesis. R. C. Suelo Nutr. Veg. 3, 37–52.
Merino, C., Jofré, I., and Matus, F. (2021a). Soil redox controls CO2, CH4 and N2O efflux from White-rot Fungi in temperate Forest ecosystems. J. Fungi 7:621. doi: 10.3390/jof7080621
Merino, C., Kuzyakov, Y., Godoy, K., Cornejo, P., and Matus, F. (2020). Synergy effect of peroxidase enzymes and Fenton reactions greatly increase the anaerobic oxidation of soil organic matter. Sci. Rep. 10:11289. doi: 10.1038/s41598-020-67953-z
Merino, C., Kuzyakov, Y., Godoy, K., Jofré, I., Nájera, F., and Matus, F. (2021b). Iron-reducing bacteria decompose lignin by electron transfer from soil organic matter. Sci. Total Environ. 761:143194. doi: 10.1016/j.scitotenv.2020.143194
Mimmo, T., Del Buono, D., Terzano, R., Tomasi, N., Vigani, G., Crecchio, C., et al. (2014). Rhizospheric organic compounds in the soil–microorganism–plant system: their role in iron availability. Eur. J. Soil Sci. 65, 629–642. doi: 10.1111/ejss.12158
Minella, M., De Laurentiis, E., Maurino, V., Minero, C., and Vione, D. (2015). Dark production of hydroxyl radicals by aeration of anoxic lake water. Sci. Total Environ. 527-528, 322–327. doi: 10.1016/j.scitotenv.2015.04.123
Naylor, D., McClure, R., and Jansson, J. (2022). Trends in microbial community composition and function by soil depth. Microorganisms 10:540. doi: 10.3390/microorganisms10030540
Neculman, R., Rumpel, C., Matus, F., Godoy, R., Steffens, M., and de la Luz Mora, M. (2013). Organic matter stabilization in two Andisols of contrasting age under temperate rain forest. Biol. Fertil. Soils 49, 681–689. doi: 10.1007/s00374-012-0758-2
Page, S., Kling, G., Sander, M., Harrold, K., Logan, J., McNeill, K., et al. (2013). Dark formation of hydroxyl radical in Arctic soil and surface waters. Environ. Sci. Technol. 47, 12860–12867. doi: 10.1021/es4033265
Petigara, B. R., Blough, N. V., and Mignerey, A. C. (2002). Mechanisms of hydrogen peroxide decomposition in soils. Environ. Sci. Technol. 36, 639–645. doi: 10.1021/es001726y
Piccolo, A., Spaccini, R., Cozzolino, V., Nuzzo, A., Drosos, M., Zavattaro, L., et al. (2018). Effective carbon sequestration in Italian agricultural soils by in situ polymerization of soil organic matter under biomimetic photocatalysis. Land Degrad. Dev. 29, 485–494. doi: 10.1002/ldr.2877
Raich, J. W., and Schlesinger, W. H. (1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 44, 81–99. doi: 10.3402/tellusb.v44i2.15428
Remucal, C., and Sedlak, D. (2011). The role of iron coordination in the production of reactive oxidants from ferrous Iron oxidation by oxygen and hydrogen peroxide. ACS Symp. Ser. 1071, 177–197. doi: 10.1021/bk-2011-1071.ch009
Robinson, W. (1927). The determination of organic matter in soils by means of hydrogen peroxide. J. Agric. Res. 34, 339–356.
Sadzawka, A., Carrasco, M., Grez, R., Mora, M., Flores, H., and Neaman, A. (2006). Métodos de análisis de suelos recomendados para los suelos de Chile. Revision 2006. Instituto de Investigaciones Agropecuarias, Chile.
Salvachúa, D., Prieto, A., Martínez, Á. T., and Martínez, M. J. (2013). Characterization of a novel dye-decolorizing peroxidase (DyP)-type enzyme from Irpex lacteus and its application in enzymatic hydrolysis of wheat straw. Appl. Environ. Microbiol. 79, 4316–4324. doi: 10.1128/aem.00699-13
Schlesinger, W. H., and Andrews, J. A. (2000). Soil respiration and the global carbon cycle. Biogeochemistry 48, 7–20. doi: 10.1023/A:1006247623877
Shah, F., Nicolás, C., Bentzer, J., Ellström, M., Smits, M., Rineau, F., et al. (2016). Ectomycorrhizal fungi decompose soil organic matter using oxidative mechanisms adapted from saprotrophic ancestors. New Phytol. 209, 1705–1719. doi: 10.1111/nph.13722
Soil Survey Staff (2014). Keys to Soil Taxonomy, 12th Edn Washington, DC: Natural Resources Conservation Service, United States Department of Agriculture. [Google Scholar].
Stookey, L. L. (1970). Ferrozine---a new spectrophotometric reagent for iron. Anal. Chem. 42, 779–781. doi: 10.1021/ac60289a016
Suryadi, H., Judono, J. J., Putri, M. R., Eclessia, A. D., Ulhaq, J. M., Agustina, D. N., et al. (2022). Biodelignification of lignocellulose using ligninolytic enzymes from white-rot fungi. Heliyon 8:e08865. doi: 10.1016/j.heliyon.2022.e08865
Sutherland, T., Sparks, C., Joseph, J., Wang, Z., Whitaker, G., Sham, T., et al. (2017). Effect of ferrous ion concentration on the kinetics of radiation-induced iron-oxide nanoparticle formation and growth. Phys. Chem. Chem. Phys. 19, 695–708. doi: 10.1039/C6CP05456K
Takahashi, Y., Katoh, S., Shikura, N., Tomoda, H., and Omura, S. (2003). Superoxide dismutase produced by soil bacteria increases bacterial colony growth from soil samples. J. Gen. Appl. Microbiol. 49, 263–266. doi: 10.2323/jgam.49.263
Trevors, J. T. (1996). Sterilization and inhibition of microbial activity in soil. J. Microbiol. Methods 26, 53–59. doi: 10.1016/0167-7012(96)00843-3
Trusiak, A., Treibergs, L. A., Kling, G. W., and Cory, R. M. (2018). The role of iron and reactive oxygen species in the production of CO2 in arctic soil waters. Geochim. Cosmochim. Acta 224, 80–95. doi: 10.1016/j.gca.2017.12.022
Van Reeuwijk, L. (2002). Procedures for Soil Analysis. Wageningen, The Netherlands. doi: 10.4236/aim.2018.87040
Velikova, V., Yordanov, I., and Edreva, A. (2000). Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sci. 151, 59–66. doi: 10.1016/S0168-9452(99)00197-1
Vermilyea, A., and Voelker, B. (2009). Photo-Fenton reaction at near neutral pH. Environ. Sci. Technol. 43, 6927–6933. doi: 10.1021/es900721x
Walsh, J. E., Fetterer, F., Scott Stewart, J., and Chapman, W. L. (2017). A database for depicting Arctic Sea ice variations back to 1850. Geogr. Rev. 107, 89–107. doi: 10.1111/j.1931-0846.2016.12195.x
White, T., Bruns, T., Lee, S., Taylor, J., Innis, M., Gelfand, D., et al. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for Phylogenetics” in PCR Protocols: A Guide to Methods and Applications. eds. M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White (New York: Academic Press), 315–322.
Willey, J. D., Kieber, R. J., and Lancaster, R. D. (1996). Coastal rainwater hydrogen peroxide: concentration and deposition. J. Atmos. Chem. 25, 149–165. doi: 10.1007/BF00053789
Wong, D. W. (2009). Structure and action mechanism of ligninolytic enzymes. Appl. Biochem. Biotechnol. 157, 174–209. doi: 10.1007/s12010-008-8279-z
Xu, Z., Qin, L., Cai, M., Hua, W., and Jin, M. (2018). Biodegradation of Kraft lignin by newly isolated Klebsiella pneumoniae, pseudomonas putida, and Ochrobactrum tritici strains. Environ. Sci. Pollut. Res. 25, 14171–14181. doi: 10.1007/s11356-018-1633-y
Xu, S., Reuter, T., Gilroyed, B., Tymensen, L., Hao, Y., Hao, X., et al. (2013). Microbial communities and greenhouse gas emissions associated with the biodegradation of specified risk material in compost. Waste Manage. (New York) 33, 1372–1380. doi: 10.1016/j.wasman.2013.01.036
Glossary

Keywords: white-rot fungi, SOM decomposition, greenhouse gasses, reactive oxygen species, enzymatic activity
Citation: Jofré-Fernández I, Matus-Baeza F and Merino-Guzmán C (2023) White-rot fungi scavenge reactive oxygen species, which drives pH-dependent exo-enzymatic mechanisms and promotes CO2 efflux. Front. Microbiol. 14:1148750. doi: 10.3389/fmicb.2023.1148750
Edited by:
Mikhail Semenov, Russian Academy of Agricultural Sciences, RussiaReviewed by:
Ilya Yevdokimov, Russian Academy of Sciences, RussiaRakesh Kumar Sharma, Manipal University Jaipur, India
Copyright © 2023 Jofré-Fernández, Matus-Baeza and Merino-Guzmán. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carolina Merino-Guzmán, Y2Fyb2xpbmEubWVyaW5vQHVmcm9udGVyYS5jbA==